

Abstract
The human acetyltransferase NAT10 has recently been shown to catalyze formation of N4-acetylcytidine (ac4C), a minor nucleobase known to alter RNA structure and function. In order to better understand the role of RNA acetyltransferases in biology and disease, here we report the development and application of chemical methods to study ac4C. First, we demonstrate that ac4C can be conjugated to carrier proteins using optimized protocols. Next, we describe methods to access ac4C-containing RNAs, enabling the screening of anti-ac4C antibodies. Finally, we validate the specificity of an optimized ac4C affinity reagent in the context of cellular RNA by demonstrating its ability to accurately report on chemical deacetylation of ac4C. Overall, these studies provide a powerful new tool for studying ac4C in biological contexts, as well as new insights into the stability and half-life of this highly conserved RNA modification. More broadly, they demonstrate how chemical reactivity may be exploited to aid the development and validation of nucleobase-targeting affinity reagents designed to target the emerging epitranscriptome.
Graphical Abstract
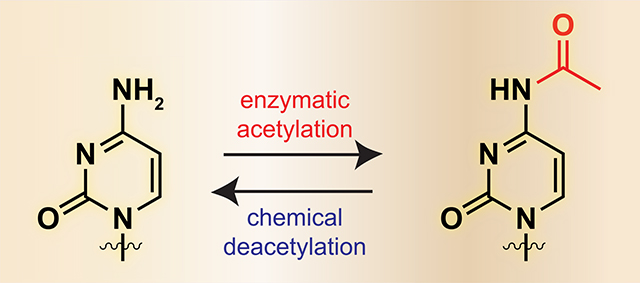
N-acetyltransferase 10 (NAT10) is a human acetyltransferase enzyme that has been implicated in the pathogenesis of many diseases, including a subtype of ovarian cancer with poor prognosis,1 highly malignant colorectal cancer,2 and premature aging syndromes.3 While NAT10 was initially characterized as a lysine acetyltransferase (KAT),4–6 it is unique among enzymes of the KAT superfamily in that it contains transfer RNA (tRNA) recognition and helicase modules, most commonly found in RNA-binding proteins (Figure 1a). The purpose for this unique domain architecture was recently revealed when several groups reported that NAT10 harbors intrinsic RNA acetyltransferase activity. Specifically, NAT10 and its yeast homologue were found to catalyze formation of the minor nucleobase N4-acetylcytidine (ac4C) in 18S ribosomal RNA (rRNA), as well as in leucine and serine tRNAs (Figure 1b,c).7–10 This finding extends the repertoire of enzyme-catalyzed cytidine acetylation, which had previously been studied in the context of bacterial tRNAs,11–13 to eukaryotes. Furthermore, it suggests that establishment of ac4C may play a functional role in pathological NAT10 activity, or serve as a biomarker thereof.
Figure 1.

Enzyme-catalyzed RNA acetylation. (a) Domain architecture of NAT10. (b) Catalytic activity of NAT10. (c) Known targets of RNA acetylation in human cells.
One challenge in studying NAT10 and related RNA acetyltransferase enzymes stems from the limited methods for their analysis. Radiolabeled 14C-acetyl-CoA has been used to validate NAT10’s biochemical activity9 but is relatively insensitive and challenging to utilize in kinetic studies. Chemoproteomic probes can be used to profile cellular NAT10 and study its interaction with small molecules14,15 but do not specifically report on its RNA acetyltransferase activity. LC-MS/MS can be used to study cellular ac4C7–10 but requires multistep digestion and specialized mass spectrometry equipment for analysis. Furthermore, standard LC-MS methods report only global levels of ac4C and are limited in their potential to interrogate the dynamics of specific sites, which may be influenced by adapter molecules.8,16,17 These considerations led us to explore the development of affinity and reactivity-based methods to study NAT10-catalyzed ac4C.
Inspired by the utility of m6A antibodies,18,19 which have provided powerful tools for its detection and location analysis in RNA, we set out to develop affinity reagents for profiling ac4C. Literature studies suggested the feasibility of this approach, as a mouse antibody to ac4C has been previously reported.20 We immobilized ac4C to two separate carrier proteins (ovalbumin and keyhole limpet hemocyanin) using a traditional periodate oxidation/reductive amination method.21 SDS-PAGE analysis of ovalbumin observed a mass shift consistent with ac4C-conjugation using either NaBH4 or NaCNBH3 as the reducing agent in the conjugation procedure (Figure 2). However, in model studies we found NaBH4 rapidly consumes ac4C, while NaCNBH3 was inert (Figure S1).20,22 This led us to produce the final immunogens using the latter reducing agent. These ac4C-conjugated carrier proteins were then injected into rabbits and applied to produce antisera, hybridomas, and ultimately monoclonal antibodies using previously reported methods.23
Figure 2.

Development of an ac4C antibody. (a) Scheme for ac4C carrier protein conjugation and antibody development. (b) SDS-PAGE gels indicating conjugation of ac4C to ovalbumin (OVA).
To screen these immunological reagents, we next required methods to generate ac4C-containing RNA probes. Unlike m6A and other RNA modifications, ac4C-containing RNAs cannot be readily accessed synthetically. This is because N4-acylation serves as a protecting group in conventional phosphoramidite RNA synthesis protocols and is cleaved during the final base-catalyzed resin cleavage step.24 This factor led us to examine whether ac4C-containing RNAs could be alternatively produced using a chemoenzymatic approach, specifically via in vitro transcription (IVT). To facilitate these studies, we first synthesized ac4CTP by direct phosphitylation of ac4C, using an adaptation of a known procedure (Scheme S1). Next, we performed IVT reactions using T7 Polymerase, a DNA template (consisting of a T7 promoter upstream of a gene sequence), and nucleoside triphosphates (NTPs). To establish the scope of this approach we tested three IVT templates of varying cytidine content and secondary structure, two of which encode biological RNAs known to contain ac4C (18S rRNA helix 45, E. coli tRNA Met) and a third that is predicted to be largely unstructured (beta-globin; Figure S2). Gel-based analysis found efficient transcription to occur both in the control reactions, which contained the four canonical NTPs, as well as when CTP was substituted with ac4CTP (Figure 3a). Furthermore, digestion of an IVT-produced RNA probe, followed by analysis of nucleoside composition by HPLC, verified incorporation of ac4C by T7 Polymerase (Figure 3b). Precedent exists for the ability of T7 Polymerase to use CTP analogues modified at the N4 position,25 although we noticed that, in general, ac4CTP reactions were less efficient (10–50% reduced yields) relative to those utilizing CTP. This approach provides access to synthetic ac4C-containing RNA probes for antibody screening. Using these probes, we systematically screened immunological reagents (serum, hybridomas, clones) to produce an optimized rabbit monoclonal ac4C antibody. Key to this antibody’s in vitro recognition properties, we found it was able to detect RNA probes synthesized in the presence of ac4CTP, but not CTP (Figure 3c). IVT ac4C RNAs are detected down to an abundance of 0.1 ng, which compares well with the sensitivity of other antibodies that have been used to detect modified cytidines within RNA.26 These studies define the selectivity and in vitro recognition of RNA by an anti-ac4C antibody.
Figure 3.

Preparation of ac4C-containing RNA probes. (a) ac4CTP is efficiently used as a substrate in T7 Polymerase-catalyzed in vitro transcription and can be incorporated into multiple templates. Full gel image is provided in the Supporting Information. (b) HPLC analysis verifies incorporation of ac4C into an in vitro transcribed RNA probe. (c) Dot blot analysis demonstrates concentration-dependent detection of in vitro transcribed ac4C-containing RNAs using a monoclonal anti-ac4C antibody.
Considering the reactivity of N4-acylation during resin-supported RNA synthesis, we next sought to examine whether it may also be possible to chemically deacetylate ac4C in aqueous buffers. We reasoned such an analysis of ac4C reactivity could fulfill two purposes. First, studying ac4C reactivity may yield insights into the enzymatic and nonenzymatic factors that regulate the dynamics of this modification in cells. Of note, while cytidine acetyltransferases have been characterized in bacteria, yeast, and humans,7–10,12,13 no cytidine deacetylase enzymes are yet known. Second, several studies have found that antibodies which show good specificity in vitro often display unanticipated cross-reactivity when presented with more complex biological epitopes.27 Selective deacetylation of ac4C could enable us to better validate our antibody, by allowing assessment of its specificity against the heterogeneous background of human RNA which contains >50 modified nucleobases.28
To facilitate our studies of reactivity, we first developed an assay to assess ac4C stability in model reactions. We found that straightforward UV spectroscopy analysis could be used to assess the deacetylation of the free ac4C nucleoside by simultaneously monitoring the disappearance of ac4C (300 nm) and appearance of cytidine (270 nm; Figure 4).11 Using this method, we first assessed the baseline stability of ac4C in aqueous buffers. We observed pH-dependent cleavage of ac4C, consistent with a base-catalyzed hydrolysis mechanism. At pH 7, ~25% of ac4C was cleaved over 18 h (Figure S3a). In contrast, at higher pH, ac4C exhibited much more rapid hydrolysis, which allowed calculation of half-lives (t1/2 = 8.2 h at pH 9; t1/2 = 1.8 h at pH 10). Next, we screened a small panel of nucleophiles for their ability to accelerate this process and found that hydroxylamine rapidly cleaves the electron-deficient acetamide of ac4C (Figure S3b, Figure 4b). Consistent with its ability to deacetylate the free nucleoside, hydroxylamine treatment also significantly reduced the detection of ac4C in IVT RNA probes, indicating its ability to cleave the electrophilic acetamide of ac4C in the context of the phosphodiester backbone (Figure 4c). Hydroxylamine did not cause detectable degradation of RNA in vitro, further establishing the relatively mild nature of these conditions (Figure S4). Moving to apply this method in the context of cellular RNA, we considered potential side reactions of hydroxylamine with canonical RNA nucleobases. For example, hydroxylamine is known to be capable of adding to the 5,6-double bond of cytidine and uridine. However, these reactions typically require elevated concentrations (1–10 M) and extreme pH,29 suggesting that a window for selective cleavage of ac4C may exist. Consistent with this hypothesis, treating total RNA isolated from HeLa cells with hydroxylamine under mild conditions (50 mM, pH 7.0) had relatively small effects on levels of canonical nucleosides (C and U, Figure 4d) but led to a substantial (~70%) reduction in ac4C levels (Figure 4e). Having validated our methods for chemical deacetylation of ac4C, we next tested the ability of our antibody to report on this change. Analysis of total RNA by anti-ac4C dot blot found a significantly decreased signal in hydroxylamine-treated samples compared to untreated samples (Figure 4f). It is important to note that these results do not rule out the possibility that hydroxylamine’s effects on ac4C detection may reflect a coordinate change in a cross-reactive nucleobase. However, by providing a measure of specificity that is complementary to genetic methods, chemical ablation of antibody recognition increases the confidence with which ac4C-targeting affinity reagents may be used.
Figure 4.

Profiling ac4C using its specific reactivity. (a) Scheme for chemical deacetylation of ac4C. (b) Effect of hydroxylamine on ac4C free nucleoside monitored by UV spectroscopy. Reaction conditions: 0.25 mM ac4C [free nucleoside], 50 mM hydroxylamine, Tris at pH 7.0. (c) Effect of hydroxylamine on ac4C in RNA monitored by anti-ac4C dot blot. Reaction conditions: 50 mM hydroxylamine, Tris at pH 8.0, 65 °C, 1 h. (d) Effect of hydroxylamine (50 mM, pH 7.0) on overall cytidine and uridine base content in total RNA analyzed by HPLC. (e) Effect of hydroxylamine (50 mM, pH 7.0, 65 °C, 1 h) on ac4C in total cellular RNA analyzed by LC-MS/MS. (f) Anti-ac4C antibody can detect chemical ablation of ac4C in total cellular RNA.
Here, we have described the development of chemical methods for the affinity and reactivity-based analysis of RNA acetylation. Our efforts were inspired by the recent observation that NAT10 regulates cytidine acetylation in rRNA and tRNA and led us to develop an antibody for the facile detection and analysis of ac4C. This reagent should prove valuable for the biochemical and biological profiling of NAT10, as well as related cytidine acetyltransferases of fungal and bacterial origin.10,12,13 In addition, we demonstrated the utility of IVT as a strategy for the universal incorporation of ac4C into synthetic RNA probes. This represents the first time synthetic ac4C-containing RNAs have been accessed and raises the possibility of using ligation or polymerase walking-based methods30 in future studies to enable the preparation of site-specifically modified ac4C probes for biophysical or functional analyses. Finally, we characterize the reactivity of ac4C and apply this knowledge to provide an additional validation of antibody specificity in the context of cellular RNA. Notably, this approach should not be limited to ac4C, as many modified nucleobases have site-specific chemistries associated with them. Such strategies may be especially useful for the validation of antibodies directed toward nucleobase modifications that are established by multiple functionally redundant writers, and which are thus challenging to genetically ablate.31,32 One caveat of ablative chemical methods is their potential to cause unanticipated changes in cellular RNA (hydroxylamine, for example, is a well-known mutagen and may cause RNA hydrolysis in other sequence contexts), necessitating that control samples be analyzed when using these methods in next-generation sequencing experiments. Another important finding was that chemical deacetylation of cytidine occurs more slowly in the context of RNA oligonucleotides than with free ac4C nucleoside (Figure 4b,e), suggesting the half-lives of ac4C observed in our model studies likely underestimate ac4C’s stability. This observation implies ac4C may persist even in RNAs with relatively long half-lives and sets the stage for site-specific analyses of ac4C dynamics in settings where pathogenic NAT10 activity has been identified, as well as the exploration of whether new targets of this modification exist. Such applications are underway and will be reported in due course.
METHODS
Preparation of ac4C-Conjugated Carrier Proteins.
Conjugation of ac4C to keyhole limpet hemocyanin (KLH) and ovalbumin (OVA) was performed according to the procedure of Erlanger and Beiser.18,21 Briefly, 5 mg of ac4C was oxidized with aqueous 0.1 M NaIO4 for 20 min. Excess periodate was removed using 25 μL of ethylene glycol. A total of 200 μL of aqueous solution of KLH or OVA (2 mg) was added, and the pH of the solution was adjusted to 9 using 5% potassium carbonate and incubated for 45 min. Reductive amination of carrier-bound ac4C was achieved via addition of NaBH3CN (200 μL, 0.6 M) on ice for 2 h. Excess NaBH3CN was quenched by formic acid, and the [pH] of the solution was adjusted to 8 using ammonium hydroxide prior to desalting using a Zeba size-exclusion column (7K MWCO). KLH and OVA conjugates were eluted in water and concentrated by lyophilization. Protein content of the resulting ac4C–protein conjugates was determined with a Qubit Protein Assay Kit.
In Vitro Transcription and Analysis of ac4C-Containing RNA Probes.
In vitro transcription was performed with the Megascript T7 Transcription Kit (ThermoFisher), according to the manufacturer’s instructions using a DNA template containing a T7 promoter upstream of a gene sequence encoding 18S rRNA helix 45, E. coli tRNA Met, or a mouse beta-globin gene sequence (Figure S2). For ac4C-containing transcripts, CTP was replaced in the reaction mixture with ac4CTP (7.5 mM; synthesis detailed in the Supporting Information). In vitro transcription reactions were analyzed by denaturing polyacrylamide gel electrophoresis on 10% TBE-urea gels and visualized using SYBR Gold staining. For a comparison of IVT using different CTP analogues (Figure 3a) an equal volume of each reaction was loaded onto the gel. Incorporation of ac4C into RNA probes was verified by subjecting in vitro transcribed products to digest with nuclease P1 (1U/10 μg RNA, #N8630, Sigma) in 50 μL of buffer containing 100 mM ammonium acetate [pH 5.5] for 16 h at 37 °C. Samples were heated at 60 °C for 5 min and returned to 37 °C for an additional hour, and 5 μL of 1 M ammonium bicarbonate [pH 8.3] and Bacterial Alkaline Phosphatase (0.5U/10 μg RNA, #18011–015, ThermoFisher) were added. Samples were further incubated at 37 °C for 2 h, adjusted to 150 μL with RNase-free water, and spin-filtered to remove enzymatic constituents (Amicon Ultra 3K, #UFC500396). Following lyophilization, samples were reconstituted in 10 μL of RNase-free water and injected into an Agilent Technologies 1260 Infinity HPLC equipped with a UV detector (Agilent Technologies). Nucleosides were separated on a Kinetex 2.6u C18 100A 100 × 2.1 mm column at a flow rate of 0.25 mL/min (chromatography gradient conditions below). The UV detector was set at 254 nm with a bandwidth of 4 nm. Buffer A: 0.01% formic acid. Buffer B: 50% acetonitrile, 0.01% formic acid [pH 3.5] with the gradient as follows, 0–1 min, 100% A; 1–2.4 min, 99.8% A; 2.4–3.8 min, 99.2% A; 3.8–5.2 min, 98.2% A; 5.2–6.6 min, 96.8% A; 6.6–10 min, 95% A; 10–12.5 min, 92% A; 12.5–18 min, 70% A; 18–18.5 min, 0% A; 18.5–20 min, 0% A; 20–21 min, 100% A; 21–30 min, 100% A.
Dot Blot Analyses.
Dot blot analyses were used to test the specificity of the ac4C antibody in vitro and in the context of cellular ac4C. For the former, synthetic RNA probes containing C or ac4C were serially diluted, denatured at 65 °C for 5 min, spotted onto Amersham Hybond-N+ membranes (GE Healthcare), and cross-linked twice with 150 mJ/cm2 in the UV254 nm Stratalinker 2400 (Stratagene, San Diego, CA). Membranes were blocked with 5% nonfat milk in 0.1% Tween-20 PBS (PBST) at RT for 30 min, and probed with anti-ac4C antibody (1:2000) in 5% nonfat milk at RT for 1 h. Membranes were washed three times with 0.1% PBST and incubated with HRP-conjugated secondary antirabbit IgG (1:10000 dilution, Cell Signaling). Membranes were washed four times with 0.1% PBST and developed with the SuperSignal ELISA Femto Maximum Sensitivity Substrate (Pierce, ThermoScientific, Waltham, MA). To detect ac4C from cellular RNA, total RNA was first purified from cultured cells using Trizol (ThermoFisher). A total of 1–10 μg of total RNA was denatured at 75 °C for 5 min, immediately placed on ice for 1 min, and loaded onto Hybond-N+ membranes. Dot blots were performed as described above.
Hydrolysis and Nucleophilic Cleavage of ac4C.
Model reactions to assess chemical deacetylation ac4C were performed using the free N4-acetylcytidine nucleoside. Reactions consisted of N4-acetylcytidine (250 μM) and reaction buffer (500 mM Tris-Cl [pH 7.5]) in Greiner-UV Star 96-well microplates (half area #655801). Reaction volumes were adjusted to 90 μL using RNase-free water and initiated by the addition of neutral hydroxylamine (0–50 mM). Chemical deacetylation was analyzed on a Biotek Synergy plate reader by monitoring the absorbance of N4-acetylcytidine (λmax = 300 nm) and cytidine (λmax = 270 nm) over 2–18 h. For chemical ablation of ac4C in cellular samples, RNA was treated with 50 mM hydroxylamine for 1 h at 65 °C prior to analysis by LC-MS/MS or anti-ac4C dot blot.
Supplementary Material
A ACKNOWLEDGMENTS
The anti-ac4C antibody was produced in partnership with Abcam. This work is supported by the Intramural Research Program of NIH, the National Cancer Institute, The Center for Cancer Research (ZIA BC011488-04), and the CCR FLEX Program. This project has been funded in whole or in part with Federal funds from the National Cancer Institute, National Institutes of Health, under contract number HHSN261200800001E
Footnotes
The authors declare no competing financial interest.
A ASSOCIATED CONTENT
Supporting Information
The Supporting Information is available free of charge on the ACS Publications website at DOI: 10.1021/acschem-bio.7b00734.
Figures S1–S4 and supporting schemes, materials, and methods (PDF)
R REFERENCES
- (1).Tan TZ, Miow QH, Huang RY, Wong MK, Ye J, Lau JA, Wu MC, Bin Abdul Hadi LH, Soong R, Choolani M, Davidson B, Nesland JM, Wang LZ, Matsumura N, Mandai M, Konishi I, Goh BC, Chang JT, Thiery JP, and Mori S (2013) Functional genomics identifies five distinct molecular subtypes with clinical relevance and pathways for growth control in epithelial ovarian cancer. EMBO Mol. Med 5, 1051–1066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (2).Zhang H, Hou W, Wang HL, Liu HJ, Jia XY, Zheng XZ, Zou YX, Li X, Hou L, McNutt MA, and Zhang B (2014) GSK-3beta-regulated N-acetyltransferase 10 is involved in colorectal cancer invasion. Clin. Cancer Res 20, 4717–4729. [DOI] [PubMed] [Google Scholar]
- (3).Larrieu D, Britton S, Demir M, Rodriguez R, and Jackson SP (2014) Chemical inhibition of NAT10 corrects defects of laminopathic cells. Science 344, 527–532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (4).Lv J, Liu H, Wang Q, Tang Z, Hou L, and Zhang B (2003) Molecular cloning of a novel human gene encoding histone acetyltransferase-like protein involved in transcriptional activation of hTERT. Biochem. Biophys. Res. Commun 311, 506–513. [DOI] [PubMed] [Google Scholar]
- (5).Shen Q, Zheng X, McNutt MA, Guang L, Sun Y, Wang J, Gong Y, Hou L, and Zhang B (2009) NAT10, a nucleolar protein, localizes to the midbody and regulates cytokinesis and acetylation of microtubules. Exp. Cell Res 315, 1653–1667. [DOI] [PubMed] [Google Scholar]
- (6).Liu X, Tan Y, Zhang C, Zhang Y, Zhang L, Ren P, Deng H, Luo J, Ke Y, and Du X (2016) NAT10 regulates p53 activation through acetylating p53 at K120 and ubiquitinating Mdm2. EMBO Rep 17, 349–366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (7).Ito S, Akamatsu Y, Noma A, Kimura S, Miyauchi K, Ikeuchi Y, Suzuki T, and Suzuki T (2014) A single acetylation of 18 S rRNA is essential for biogenesis of the small ribosomal subunit in Saccharomyces cerevisiae. J. Biol. Chem 289, 26201–26212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (8).Sharma S, Langhendries JL, Watzinger P, Kotter P, Entian KD, and Lafontaine DL (2015) Yeast Kre33 and human NAT10 are conserved 18S rRNA cytosine acetyltransferases that modify tRNAs assisted by the adaptor Tan1/THUMPD1. Nucleic Acids Res 43, 2242–2258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (9).Ito S, Horikawa S, Suzuki T, Kawauchi H, Tanaka Y, Suzuki T, and Suzuki T (2014) Human NAT10 is an ATP-dependent RNA acetyltransferase responsible for N4-acetylcytidine formation in 18 S ribosomal RNA (rRNA). J. Biol. Chem 289, 35724–35730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (10).Taoka M, Ishikawa D, Nobe Y, Ishikawa H, Yamauchi Y, Terukina G, Nakayama H, Hirota K, Takahashi N, and Isobe T (2014) RNA cytidine acetyltransferase of small-subunit ribosomal RNA: identification of acetylation sites and the responsible acetyltransferase in fission yeast, Schizosaccharomyces pombe. PLoS One 9, e112156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (11).Stern L, and Schulman LH (1978) The role of the minor base N4-acetylcytidine in the function of the Escherichia coli noninitiator methionine transfer RNA. J. Biol. Chem 253, 6132–6139. [PubMed] [Google Scholar]
- (12).Ikeuchi Y, Kitahara K, and Suzuki T (2008) The RNA acetyltransferase driven by ATP hydrolysis synthesizes N4-acetylcytidine of tRNA anticodon. EMBO J 27, 2194–2203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (13).Chimnaronk S, Suzuki T, Manita T, Ikeuchi Y, Yao M, Suzuki T, and Tanaka I (2009) RNA helicase module in an acetyltransferase that modifies a specific tRNA anticodon. EMBO J 28, 1362–1373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (14).Montgomery DC, Sorum AW, and Meier JL (2014) Chemoproteomic profiling of lysine acetyltransferases highlights an expanded landscape of catalytic acetylation. J. Am. Chem. Soc 136, 8669–8676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (15).Montgomery DC, Garlick JM, Kulkarni RA, Kennedy S, Allali-Hassani A, Kuo YM, Andrews AJ, Wu H, Vedadi M, and Meier JL (2016) Global Profiling of Acetyltransferase Feedback Regulation. J. Am. Chem. Soc 138, 6388–6391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (16).Sharma S., Yang J., van Nues R., Watzinger P., Kotter P., Lafontaine DLJ., Granneman S., and Entian KD. (2017) Specialized box C/D snoRNPs act as antisense guides to target RNA base acetylation. PLoS Genet. 13, e1006804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (17).Zhang X, Jiang G, Sun M, Zhou H, Miao Y, Liang M, Wang E, and Zhang Y (2017) Cytosolic THUMPD1 promotes breast cancer cells invasion and metastasis via the AKT-GSK3-Snail pathway. Oncotarget 8, 13357–13366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (18).Munns TW, Liszewski MK, and Sims HF (1977) Characterization of antibodies specific for N6-methyladenosine and for 7-methylguanosine. Biochemistry 16, 2163–2168. [DOI] [PubMed] [Google Scholar]
- (19).Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, and Jaffrey SR (2012) Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons. Cell 149, 1635–1646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (20).Reynaud C, Bruno C, Boullanger P, Grange J, Barbesti S, and Niveleau A (1992) Monitoring of urinary excretion of modified nucleosides in cancer patients using a set of six monoclonal antibodies. Cancer Lett 61, 255–262. [DOI] [PubMed] [Google Scholar]
- (21).Erlanger BF, and Beiser SM (1964) Antibodies Specific for Ribonucleosides and Ribonucleotides and Their Reaction with DNA. Proc. Natl. Acad. Sci. U. S. A 52, 68–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (22).Cerutti P, and Miller N (1967) Selective reduction of yeast transfer ribonucleic acid with sodium borohydride. J. Mol. Biol 26, 55–66. [DOI] [PubMed] [Google Scholar]
- (23).Huang Y, Gu B, Wu R, Zhang J, Li Y, and Zhang M (2007) Development of a rabbit monoclonal antibody group against Smads and immunocytochemical study of human and mouse embryonic stem cells. Hybridoma 26, 387–391. [DOI] [PubMed] [Google Scholar]
- (24).Scaringe SA (2000) Advanced 5′-silyl-2′-orthoester approach to RNA oligonucleotide synthesis. Methods Enzymol 317, 3–18. [DOI] [PubMed] [Google Scholar]
- (25).Stengel G, Urban M, Purse BW, and Kuchta RD (2010) Incorporation of the fluorescent ribonucleotide analogue tCTP by T7 RNA polymerase. Anal. Chem 82, 1082–1089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (26).Delatte B, Wang F, Ngoc LV, Collignon E, Bonvin E, Deplus R, Calonne E, Hassabi B, Putmans P, Awe S, Wetzel C, Kreher J, Soin R, Creppe C, Limbach PA, Gueydan C, Kruys V, Brehm A, Minakhina S, Defrance M, Steward R, and Fuks F (2016) RNA biochemistry. Transcriptome-wide distribution and function of RNA hydroxymethylcytosine. Science 351, 282–285. [DOI] [PubMed] [Google Scholar]
- (27).Fuchs SM, Krajewski K, Baker RW, Miller VL, and Strahl BD (2011) Influence of combinatorial histone modifications on antibody and effector protein recognition. Curr. Biol 21, 53–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (28).Machnicka MA, Milanowska K, Osman Oglou O, Purta E, Kurkowska M, Olchowik A, Januszewski W, Kalinowski S, Dunin-Horkawicz S, Rother KM, Helm M, Bujnicki JM, and Grosjean H (2013) MODOMICS: a database of RNA modification pathways–2013 update. Nucleic Acids Res 41, D262–267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (29).Rhodes D (1975) Accessible and inaccessible bases in yeast phenylalanine transfer RNA as studied by chemical modification. J. Mol. Biol 94, 449–460. [DOI] [PubMed] [Google Scholar]
- (30).Liu Y, Holmstrom E, Zhang J, Yu P, Wang J, Dyba MA, Chen D, Ying J, Lockett S, Nesbitt DJ, Ferre-D’Amare AR, Sousa R, Stagno JR, and Wang YX (2015) Synthesis and applications of RNAs with position-selective labelling and mosaic composition. Nature 522, 368–372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (31).Liu J, Yue Y, Han D, Wang X, Fu Y, Zhang L, Jia G, Yu M, Lu Z, Deng X, Dai Q, Chen W, and He C (2013) A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol 10, 93–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (32).Safra M, Nir R, Farouq D, Vainberg Slutskin I, and Schwartz S (2017) TRUB1 is the predominant pseudouridine synthase acting on mammalian mRNA via a predictable and conserved code. Genome Res 27, 393–406. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.