

Abstract
Allowing dairy cattle to access pasture can promote natural behaviour and improve their health. However, the psychological benefits are poorly understood. We compared a cognitive indicator of emotion in cattle either with or without pasture access. In a crossover experiment, 29 Holstein–Friesian dairy cows had 18 days of overnight pasture access and 18 days of full-time indoor housing. To assess emotional wellbeing, we tested cows on a spatial judgement bias task. Subjects learnt to approach a rewarded bucket location, but not approach another, unrewarded bucket location. We then presented cows with three “probe” buckets intermediate between the trained locations. Approaching the probes reflected an expectation of reward under ambiguity—an “optimistic” judgement bias, suggesting positive emotional states. We analysed the data using linear mixed-effects models. There were no treatment differences in latency to approach the probe buckets, but cows approached the known rewarded bucket slower when they had pasture access than when they were indoors full-time. Our results indicate that, compared to cattle housed indoors, cattle with pasture access display less anticipatory behaviour towards a known reward. This reduced reward anticipation suggests that pasture is a more rewarding environment, which may induce more positive emotional states than full-time housing.
Subject terms: Psychology, Zoology
Introduction
Picture the scene: Holstein–Friesian cows grazing lush pasture on a summer afternoon. Consumers overwhelmingly support such pasture-based systems and oppose housing cattle indoors full-time. Ninety-five percent of the British public believe pasture access benefits dairy cows1. That figure is 88% in Germany2, 84% in the United States and Canada3, and 81% in Brazil4. Industry stakeholders also value pasture access, noting its importance for cattle welfare3,5. However, housing cows indoors full-time facilitates feeding high-energy diets and increasing herd size on the same geographical area6. As a result, farmers across Europe and North America increasingly keep cattle indoors all year round6,7. In Denmark, Greece, and Poland, less than a quarter of dairy cows went out to pasture in 20197. In the United States, only 20% of lactating cows and 34% of dry cows accessed pasture in 20138. Some countries buck this trend. Over 95% of British and Irish dairy cows went out to pasture in 20197, whilst Finland, Norway, and Sweden have banned full-time housing7,9. Worldwide, though, most milk now comes from dairy cows without any pasture access10.
Dairy cattle themselves also value pasture access. Given the choice between pasture and housing, cattle usually spend longer at pasture, particularly at night11–15. When pasture access requires walking long distances12,16 or pushing weighted doors17, cows appear to value pasture as highly as fresh food. Lying behaviour is strongly motivated and crucial for cow comfort18. Cows at pasture have longer lying bouts and are less restless19,20. Softer lying and walking surfaces also reduce the risk of injuries21, lameness20,22, and mastitis23,24. Cubicle barns have limited space and localised resources, so herds at pasture are less aggressive25 and more synchronous19,25. Full-time housing has some welfare benefits, such as protection from extreme weather26, lower risk of gastrointestinal parasites27, and greater control over nutrition28. Despite these advantages, several recent reviews have concluded that pasture access improves dairy cattle welfare29–32.
However, pasture’s importance for cows’ psychological wellbeing is poorly understood. Wellbeing requires a favourable balance of positive and negative emotions33–35. Two dimensions characterise emotion: arousal, the intensity of activation, and valence, whether the state is pleasant or unpleasant36–40. In humans, for instance, both elation and terror are high-arousal states, but elation is positively valenced, whereas terror is negatively valenced. Measures of cognition, physiology, and behaviour can reveal where animals fall along the two axes39,41. Based on these indicators, does pasture access induce positive emotions in dairy cows?
Emotions influence cognition, including attention, judgement, and memory42–44. For humans, positive emotions cause more optimistic judgements about ambiguous stimuli (“judgement bias”)45–49. Optimism also indicates psychological wellbeing in animals50, from primates to insects (reviews and meta-analyses:51–55). When presented with ambiguous stimuli, animals in positive-valence states expect more positive outcomes than animals in negative-valence states. To measure this judgement bias, researchers train subjects to respond differently to two unidimensional stimuli (e.g. spatial locations56). One stimulus (P) signals a relatively positive outcome, whereas the other stimulus (N) signals a relatively negative outcome. After training, subjects are exposed to ambiguous intermediate stimuli (probes). P-like responses to the probes indicate that the animal expects a positive outcome (i.e. optimism), whereas N-like responses indicate that the animal expects a negative outcome (i.e. pessimism). In a meta-analysis of 71 studies on 22 species, Lagisz et al.52 linked better housing and husbandry to more optimistic judgements about ambiguous stimuli.
Judgement biases are a popular indicator of livestock emotions and welfare57. For example, Neave et al.58 trained dairy calves to respond differently to red and white screens. “Go” responses (nose-touching) to one colour (P) yielded a milk reward, whilst “No-go” responses to the other colour (N) averted a one-minute time-out. When subsequently tested on ambiguous probe colours (pink screens), calves made more Go (i.e. optimistic) responses before hot-iron disbudding than after (see also59). In other calf studies, maternal separation induced pessimism60, and pair-housing induced optimism61. Pasture access also led to optimistic judgement biases in horses62,63. To our knowledge, judgement bias has not been investigated in adult cattle, but this method could reveal whether pasture access influences cows’ psychological wellbeing29.
The present crossover study measured emotional wellbeing in cows, which were given 18 days of overnight pasture access (PAS treatment) and 18 days of full-time housing (PEN treatment). We trained subjects on a spatial Go/No-go task, where a bucket at one location (P) contained food and a bucket at another location (N) was empty. Go responses and short response latencies to three intermediate probe locations indicated optimistic judgement biases. We hypothesised that cows in the PAS treatment would make more Go responses and have shorter response latencies to the probes than cows in the PEN treatment, indicating greater emotional wellbeing. We also predicted that likelihood to respond to the probe buckets would decrease—and latency would increase—with day number, as subjects learnt that the probes were unreinforced65. This was, to our knowledge, the first judgement bias study on cows64.
Methods
Ethical approval
This research was approved by Queen’s University Belfast’s Animal Research Ethics Committee, School of Biological Sciences (approval number: QUB-BS-AREC-18-05). The experiment was below the threshold of procedures detailed within the Animals (Scientific Procedures) Act 1986, and animal welfare was prioritised throughout. We confirm that all methods were carried out in accordance with relevant guidelines and regulations.
Subjects and housing
We performed this study from May to August 2018 at the Agri-Food and Biosciences Institute (AFBI), Hillsborough, County Down, Northern Ireland (54° 5′ N; 6° 1′ W). We recruited 29 lactating Holstein–Friesian dairy cows (2.7–8.7 years old, mean: 4.3 years; 209–273 days calved, mean: 241 days). Subjects were managed alongside three non-study individuals, which maintained a 1:1 cow/cubicle ratio, giving a total herd size of 32 animals. Every cow had been at pasture that spring but was cubicle-housed for 8 weeks before the experiment began. During this time, the herd was managed as one group. Cows were kept in two interconnected pens (pen dimensions: 113.1 m2; 13.3 × 8.5 m), each containing 16 cantilever-type cubicles in three rows. Two of the rows, arranged “head-to-head”, had five cubicles each (cubicle dimensions: 2.2 m × 1.2 m); the third row, facing the pens’ back wall, had six cubicles (cubicle dimensions: 2.5 m × 1.2 m). Every cubicle contained a Pasture Mat rubber mat (Promat, Woodstock, Canada). The pens also had concrete standing and walking areas, which were scrape-cleaned six times daily. Subjects were given ad libitum access to fresh water and ad libitum grass silage delivered at approximately 09.00. Every day, milking occurred at 06.30 and 15.00 in a Boumatic Daytona RMS-X Exterior 50-Point Rotary Parlour.
Timeline and treatments
We carried out judgement bias training from 24th May to 18th June 2018, judgement bias inclusion testing from 19th to 22nd June 2018, and the experiment from 23rd June to 3rd August 2018. The day before the experiment, we separated the pens and pseudorandomly divided subjects into two herds. Plywood visual barriers were installed to limit emotional contagion between treatments66. We balanced the groups for mobility scores and P side (left/right) in the judgement bias task. To account for mobility, a veterinary graduate (H.K.) assessed each animal’s gait 4 days before the experiment began (following67). Both groups had equal numbers of non-lame (scores: 0 or 1) and lame animals (scores: 2 or 3; see19 for details).
This experiment used a two-period crossover design with repeated measures. The first period was from 25th June to 13th July 2018 (18 days); the second period was from 16th July to 3rd August 2018 (18 days). There were two parallel treatments: the PAS treatment had daily pasture access from 16.00 to 10.00, whilst the PEN treatment lived in cubicle housing 24 h per day (except during milking). PAS cows were managed in a rotational grazing system, with grazing area 1370–3950 m2 (depending on herbage mass available) and distance to parlour 190–295 m. During each experimental period, we cut grass samples three times (six total). Herbage was high quality: mean oven dry matter (DM) content was 239 (SD 9) g/kg in the first period and 215 (SD 9) g/kg in the second period; mean crude protein content was 226 (SD 12) g/kg DM in the first period and 207 (SD 12) g/kg DM in the second period; and mean metabolizable energy content was 12.0 MJ/kg DM in the first period and 10.9 MJ/kg DM in the second period.
We collected judgement bias data during the daytime (10.00–15.00), when both groups were kept indoors with equivalent silage provision. The group in the PAS treatment during the first experimental period (PAS-first) contained 14 study animals (2.7–8.7 years old, mean: 4.5 years; 219–260 days calved, mean: 240 days). The group in the PEN treatment during the first experimental period (PAS-second) contained 15 study animals (2.7–7.8 years old, mean: 4.2 years; 209–273 days calved, mean 242 days). With the three non-study animals, both groups contained 16 individuals. The second experimental period was identical to the first, except the groups swapped treatments.
Judgement bias
Judgement bias testing involved two pens adjacent to the home pens: the holding area, where subjects were kept before sessions and during inter-trial intervals, and the testing area (13.3 × 3.1 m), where the task was carried out. Subjects in the holding area could not see the testing area. Once per weekday, experimenter 1 (A.C.) individually moved each cow into the holding area (subject order randomised each day). We used a spatial Go/No-go judgement bias task, with a bucket at one of five locations56,68,69 (Fig. 1). The P and N stimuli were buckets at the right and left locations (location counterbalanced between subjects). Rewarded P buckets contained 130 g of grain-based concentrate feed, which cattle find very desirable70. N buckets were not reinforced. The ambiguous probe stimuli were buckets at three intermediate spatial locations; these were also not reinforced. We ended trials if subjects did not make a Go response within 20 s. If the subject made a Go response and the bucket was rewarded, we allowed an additional 30 s to feed. We pseudorandomised trial order—subjects never had more than two consecutive buckets at the same location. Between trials, experimenter 1 moved the cow back into the holding area, and experimenter 2 (K.J.) re-set the bucket. Experimenter 2 filmed sessions on a tripod-mounted Sony HDR-CX450 1080p Camcorder.
Figure 1.
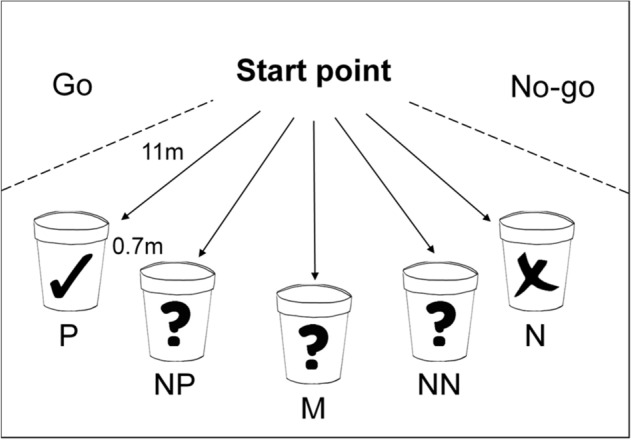
Diagram of the experimental setup, illustrating the five bucket locations (positive, P; near-positive, NP; middle, M; near-negative, NN; negative, N) and trained responses (Go, No-go).
Before the experiment began, cows underwent 18 training days. We divided these into six blocks of 3 days (Table 1), with one session of consecutive trials per subject per day. By the final 3-day training block, each cow received two rewarded P trials (P-Rew), one unrewarded P trial (P-Unr), and three unrewarded N trials (N-Unr) per day across a six-trial session. P-Unr trials introduced a one-third variable reinforcement ratio. Variable reinforcement reduces extinction learning towards unrewarded probes, which can look like increased pessimism without any change in emotional state65. To maintain task motivation, subjects never received more than two consecutive unreinforced trials (either P-Unr or N-Unr), and the last P trial was always P-Rew.
Table 1.
Training timeline, with the number of rewarded P trials (P-Rew), unrewarded P trials (P-Unr), and unrewarded N trials (N-Unr) per cow in each consecutive three-day block.
| Days | P-Rew trials | P-Unr trials | N-Unr trials | Total trials |
|---|---|---|---|---|
| 1–3 | 1 | 0 | 0 | 1 |
| 4–6 | 2 | 0 | 0 | 2 |
| 7–9 | 2 | 0 | 1 | 3 |
| 10–12 | 2 | 0 | 2 | 4 |
| 13–15 | 2 | 0 | 3 | 5 |
| 16–18 | 2 | 1 | 3 | 6 |
After the training phase, we carried out 3 days of inclusion testing to confirm that subjects had learnt the spatial discrimination task. We recorded responses in six inclusion trials per day (18 trials total). Three trials per day involved the P location (2 × P-Rew; 1 × P-Unr) and three trials per day involved the N location (3 × N-Unr). For each subject, we extracted the latency for all 18 trials, with No-go responses given a ceiling latency of 20 s. Our inclusion criteria are outlined in the “Statistical analyses” section.
During both experimental phases, we carried out judgement bias testing every Monday, Wednesday, and Friday (8 × testing sessions per individual per phase; 16 × testing sessions per individual total). Half of testing sessions included three P trials (2 × Rew; 1 × Unr) and two N trials, whilst the other half included two P trials (2 × Rew) and three N trials. The remaining trial was a probe bucket at one of three equidistant intermediate locations: near-positive (NP; 0.7 m from P), middle (M; 1.4 m from both P and N), and near-negative (NN; 0.7 m from N). The probe trial randomly replaced either P-Unr or N trials. Experimenter 3 (H.K.) extracted data for the P, N, and probe buckets from video footage. If the subject’s muzzle touched or entered the bucket, a Go response was recorded. Otherwise, a No-go response was recorded. Latency was also measured, from one hoof crossing a standardised start line to the Go response (distance: 11 m). Experimenter 3 was blind to both treatment (PAS/PEN) and which bucket location was rewarded (P/N). Throughout the experiment, we continued training sessions on Tuesdays and Thursdays. This increased the P/N:probe ratio, further reducing extinction learning towards the probes51.
Statistical analysis
We analysed the data in R (R Core Team, Cran-r-project, Vienna, Austria, version 3.6.2). We checked data and model assumptions using histograms and qqplots, applying transformations where appropriate. We used the package “lme4”71 to run mixed-effects models and dropped interactions when this reduced the model’s Akaike Information Criterion value by > 5. We then extracted p values using type III Wald’s tests. Where factors had multiple levels or interactions involved multiple comparisons, we performed a Tukey’s post-hoc test (“lsmeans” package72) to quantify differences between levels or comparisons. Unless otherwise stated, we present data as means ± standard error of the mean.
For the training data, we used a statistical inclusion criterion. We ran a Wilcoxon test on the latency data from each cow’s inclusion trials (n = 18; 9 × P, 9 × N). To proceed, subjects needed shorter response latencies to the P location than the N location (p < 0.05). We also ran a general linear mixed-effects model on the inclusion data to establish that subjects learnt the left/right association, rather than using olfactory cues to approach the rewarded location. Latency was included in the model as the response variable; location/reward category (P-Rew, P-Unr, N-Unr) was included as a fixed effect; and cow ID was included as a random effect. We identified differences between categories with a Tukey’s post-hoc test. Subjects not using olfactory cues would show no difference between P-Rew and P-Unr, but a difference between P and N; subjects using olfactory cues would show no difference between P-Unr and N-Unr, but a difference between Rew and Unr.
For the judgement bias data, we ran models with both binary Go/No-go responses and response latency as the dependent variables52. We fitted a generalised linear mixed-effects model for the Go/No-go data (binomial distribution, logit link). We ran a general linear mixed-effects model for the latency data, which we transformed by taking the natural logarithm of the value + 1. We excluded No-go responses from this model. In both models, the fixed effects were housing treatment (PAS, PEN), treatment order (PAS-first, PAS-second), bucket location (P, NP, M, NN, N), and day number (1–16). We also included treatment × treatment order, treatment × bucket location, and bucket location × day number interactions as fixed effects. Cow ID was included as a random effect. We also ran a separate model on latency (log-transformed + 1) to the P location only. Fixed and random effects were the same as for the previous model, except that we removed bucket location. To account for food motivation, we also included body condition score and time of day as fixed effects, as well as body condition score × treatment and time of day × treatment interactions.
Results
Judgement bias training
During inclusion testing, all 29 cows approached the P location faster than the N location (all subjects: p < 0.001) and advanced to the experimental phase. Investigating the effect of bucket location and food reward presence/absence, we found a difference in latency between P-Rew trials (median latency ± SD: 5.75 ± 0.92 s), P-Unr trials (5.75 ± 0.93 s), and N-Unr trials (20 ± 4.36 s; χ22 = 2248, p < 0.001). Post-hoc tests revealed no difference between P-Rew and P-Unr (z = − 0.14, p = 0.989). However, subjects were faster to both P-Rew (z = − 42.34, p < 0.001) and P-Unr (z = − 33.62, p < 0.001), compared to N-Unr.
Judgement bias testing
We collected data from 2741 judgement bias trials. Excluding the 1342 No-go trials, latency data were available from 1399 Go trials. Latency from start line to bucket ranged from 2.75 to 18.91 s (mean: 7.03 s).
Cows in the PAS treatment were less likely to approach the buckets (PAS: 47.75% trials; PEN: 53.24% trials; χ21 = 9.90, p < 0.001) and took longer to do so (PAS: 7.12 ± 0.07 s; PEN: 6.42 ± 0.05 s; χ21 = 26.91, p < 0.001). Treatment order did not affect approach likelihood (χ21 = 2.35, p = 0.13) or latency (χ21 = 0.38, p = 0.54). There was a treatment × treatment order interaction for both likelihood to approach (χ21 = 14.99, p < 0.001) and latency to approach (χ21 = 6.08, p = 0.01; Fig. 2). During the PEN treatment, the PAS-first group had a smaller increase in approach likelihood and reduction in approach latency than the PAS-second group.
Figure 2.
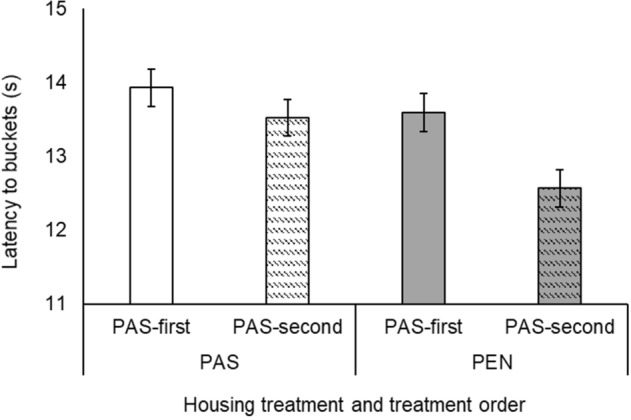
Interaction between housing treatment (pasture access: PAS; cubicle housing: PEN) and treatment order (PAS-first, PAS-second) in response latency to all five bucket locations. Error bars represent standard error of the mean.
There was an effect of bucket location on both the number of Go responses (χ24 = 816.31, p < 0.001) and approach latency (χ24 = 1089.89, p < 0.001; Fig. 3). Post-hoc tests revealed that all five bucket locations differed from one another in terms of both approach likelihood and latency (Table 2). There was no treatment × bucket location interaction for approach likelihood (χ24 = 2.11, p = 0.72), but there was an interaction for approach latency (χ24 = 15.87, p < 0.005). This showed that the main effect of treatment on latency was localised to the P location: cows were slower to approach P when they were in the PAS treatment than the PEN treatment (PAS: 6.38 ± 0.04 s; PEN: 6.28 ± 0.05 s; t1,386 = 6.39, p < 0.001; Fig. 4). There was no treatment difference in latency to any other location (NP: t1,385 = 0.42, p > 0.99; M: t1,386 = − 0.66, p > 0.99; NN: t1,387 = − 1.60, p = 0.85; N: t1,387 = 0.45, p > 0.99).
Figure 3.

Response latency to the five bucket locations throughout the experiment (negative: N; near-negative: NN; middle: M; near-positive: NP; positive: P). Error bars represent standard error of the mean.
Table 2.
Pairwise comparisons of the likelihood and latency to approach each bucket location, and for the bucket location × day number interaction.
| Comparison | Bucket location approach | Bucket location × day number | ||||||
|---|---|---|---|---|---|---|---|---|
| Likelihood | Latency | Likelihood | Latency | |||||
| z | p | t | p | z | p | t | p | |
| P – NP | − 5.98 | < 0.001 | 9.40 | < 0.001 | − 2.86 | 0.03 | 3.88 | < 0.005 |
| P – M | − 11.00 | < 0.001 | 13.14 | < 0.001 | − 4.11 | < 0.001 | 2.91 | 0.03 |
| P – NN | − 15.52 | < 0.001 | 14.70 | < 0.001 | − 2.59 | 0.07 | − 0.22 | < 1.00 |
| P – N | − 28.23 | < 0.001 | 26.29 | < 0.001 | − 2.39 | 0.12 | − 1.34 | 0.67 |
| NP – M | − 3.84 | < 0.005 | 4.48 | < 0.001 | − 0.93 | 0.88 | − 0.20 | < 1.00 |
| NP – NN | − 7.73 | < 0.001 | 7.26 | < 0.001 | 0.42 | 0.99 | − 2.46 | 0.10 |
| NP – N | − 16.78 | < 0.001 | 15.52 | < 0.001 | 1.21 | 0.75 | − 3.81 | < 0.005 |
| M – NN | − 4.17 | < 0.001 | 3.09 | 0.02 | 1.42 | 0.61 | − 2.11 | 0.22 |
| M – N | − 13.80 | < 0.001 | 10.06 | < 0.001 | 2.42 | 0.11 | − 3.20 | 0.01 |
| NN – N | − 9.53 | < 0.001 | 6.01 | < 0.001 | 0.79 | 0.93 | − 0.64 | 0.97 |
Bold p values are significant.
Figure 4.
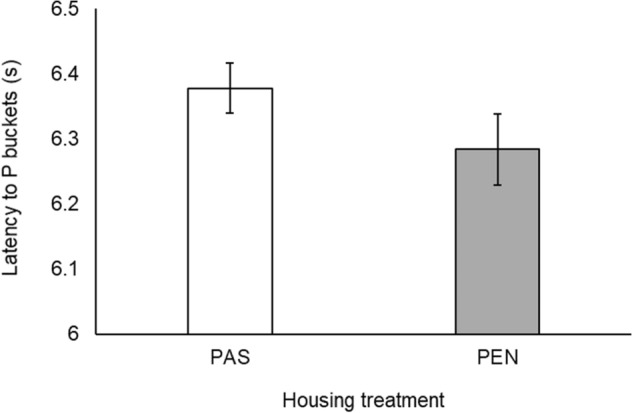
Response latency to the positive (P) bucket location in each housing treatment (pasture access: PAS; cubicle housing: PEN). Error bars represent standard error of the mean.
As the study progressed (i.e. day number increased), likelihood to approach the buckets decreased (χ21 = 27.62, p < 0.001; Fig. 5a) and approach latency increased (χ21 = 19.28, p < 0.001; Fig. 5b). There was also a bucket location × day number interaction for approach likelihood (χ24 = 21.72, p < 0.001) and latency (χ24 = 25.83, p < 0.001; Table 2).
Figure 5.
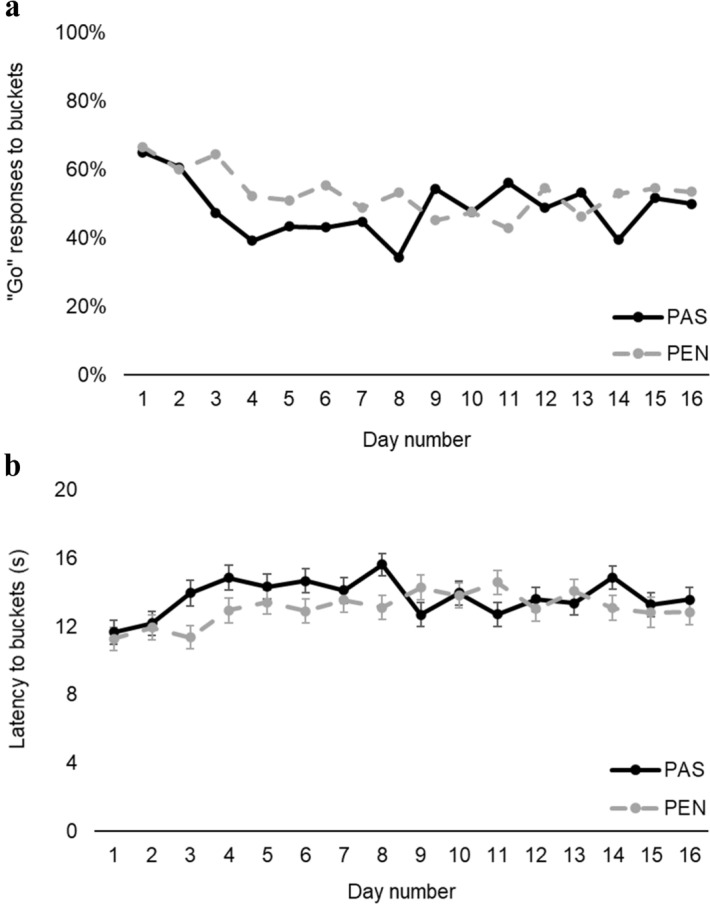
(a) Percentage of “Go” responses and (b) response latency to all buckets in each treatment (pasture access: PAS; cubicle housing: PEN) throughout the experiment (days 1–16). Error bars represent standard error of the mean.
When we modelled latency to the P location, there was no effect of either time of day (χ21 = 0.66, p = 0.42) or body condition score (χ21 = 0.00, p = 0.96). There was no treatment × time of day (χ21 = 0.53, p = 0.47) or treatment × body condition score interactions (χ21 = 0.20, p = 0.66).
Discussion
We investigated whether pasture access impacts judgement bias in dairy cows, to assess whether pasture improves emotional wellbeing. Cattle learnt a spatial judgement bias task, and optimistic responses increased with proximity to the rewarded location. However, there was no treatment difference in judgement bias. Subjects with pasture access were neither likelier nor faster to approach buckets when the reward contingency was ambiguous. Cows with pasture access nonetheless approached known rewarded buckets slower than cows kept indoors full-time.
We did not predict that cows would be slower to the P location in the PAS treatment than the PEN treatment. A core assumption of judgement bias tasks is that emotions bias decision-making when outcomes are uncertain39,73,74. As a result, treatment effects in judgement bias are expected towards the probes—not the trained P and N stimuli39,73. Most studies meet this assumption52 (for exceptions, see50,75,76). Moreover, we expected pasture access to reduce approach latency, representing a higher expectation of reward and an optimistic judgement bias. One explanation for our surprising result is that cows could have been less food motivated in the PAS treatment than the PEN treatment (e.g. see77–79). At pasture, cattle begin grazing at dawn (around 06.00)80, whereas subjects in our PEN treatment were only offered fresh silage at 09.00. PAS cows may, therefore, have fed for longer than PEN cows before testing. However, if food motivation were responsible, the effect would be strongest earliest in the day and decrease as all subjects spent longer with equivalent rations. Time of day did not affect latency to the P location. Additionally, we scored every cows’ body condition during both experimental phases. Higher scores reflect better nutrition67, so body condition score is inversely correlated with food motivation. We found no relationship between P latency and body condition score, suggesting that food motivation did not influence P latency. These converging lines of evidence indicate that treatment differences in nutrition were not responsible.
We propose that reduced reward anticipation, linked to positive emotional states, explains why the PAS treatment were slower to the P location than the PEN treatment. Spruijt et al35 hypothesised that animals exposed to fewer, lower-quality rewards value each reward more (Fig. 6; reviewed by81,82; see83 for a critique). As an example, dairy calves in basic housing responded to a reward-predicting cue with more behavioural transitions and shorter response latencies than calves in enriched housing84 (see also85). This effect means that, in a judgement bias task, we predict opposite welfare-based differences in response patterns towards the P stimulus and the probes. If animals have received more, higher-quality rewards, the P stimulus will elicit less anticipatory behaviour, whereas the probes will elicit more82. Latency to a rewarded bucket meets Spruijt et al.’s definition of anticipatory behaviour: “responses elicited by rewarding stimuli that lead to and facilitate consummatory behavior” (p. 160). We, therefore, suggest that PAS cows’ longer P latencies reflected lower reward anticipation, indicating that they had more rewarding lives and better welfare, rather than pessimistic judgement biases, which would indicate less rewarding lives and worse welfare. Our earlier findings support this interpretation: compared to cows in the PEN treatment, cows in the PAS treatment had longer total lying times; fewer, longer lying bouts; and more synchronous lying behaviour19. These results indicate that cows at pasture were more comfortable and less restless. The inverse relationship between reward frequency and reward anticipation does not apply to chronically stressed animals, which display reduced reward valuation (anhedonia86). Our results indicate that PEN cows were not anhedonic but had less rewarding lives than PAS cows.
Figure 6.
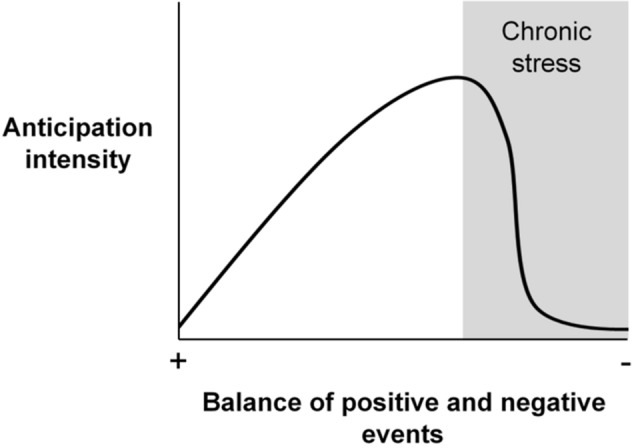
Relationship between the balance of positive and negative events in an animal’s life and anticipation intensity towards individual rewards (adapted from96).
Our reward anticipation findings highlight how judgement bias tasks may quantify effects besides judgement bias51,53. However, few studies have compared judgement bias and reward anticipation. Optimistic judgement biases were linked to reduced anticipatory behaviour in dolphins87, whereas enriched housing did not affect chickens’ responses to either a judgement bias or reward anticipation task88. Other studies suggest treatment differences in anticipatory behaviour influenced responses to a judgement bias task. For example, disrupting reward-related behaviours in chicks reduced latencies towards the P stimulus76. The antidepressant reboxetine likewise reduced P responses in rats75, and deep-litter enrichment reduced P responses in quail89 (exp. 4), although neither finding was consistent across experiments. Like our results, these P response patterns can be attributed to increased reward anticipation among subjects in more negative emotional states. Moreover, because judgement bias and reward anticipation predict opposite responses, it is possible that they cancel each other out. In a meta-analysis of judgement bias studies using pharmacological manipulations, effect sizes were smaller for the P stimulus than either the probes or N stimulus54. To differentiate the effects of judgement bias and reward anticipation, we suggest that judgement biases are more plausible when treatment differences only occur towards the probes, whereas treatment differences localised around the P stimulus imply reward anticipation.
Despite P responses indicating that the PAS treatment induced more positive emotional states, pasture access did not influence judgement bias. This is surprising, as aversive events lead to pessimism in dairy calves58–61, and pasture access leads to optimism in horses62,63. However, studies on pigs90, chickens88, and quail89 have found no difference in judgement bias between housing conditions. There are two possible reasons for our null results. First, pasture access may not influence emotional state in dairy cows. This explanation might seem implausible, given pasture’s potential behavioural and health benefits, and cows’ preference and motivation for pasture29,30. Subjects’ behaviour during our experiment—with fewer, longer lying bouts during the PAS treatment—also indicate that PAS cows were more comfortable19. However, we tested judgement bias during the daytime, when both treatments were kept indoors. Pasture may only improve emotional wellbeing whilst cows are at pasture, without persisting after they go inside. Ruet et al91 found that, when confined indoors again, horses given pasture access rapidly returned to previous poor welfare states. Conversely, Anderson and Adolphs92 identified persistence as a defining feature of emotions. Their characterisation is consistent with our reward anticipation findings, which indicate that positive emotional states from overnight pasture access carried over into daytime indoor housing.
The second potential explanation for our null judgement bias results is that treatment differences in emotional state existed, but our task did not detect them. In their meta-analysis, Lagisz et al.52 identified four methodological factors that may be responsible for our findings. (1) Sex: males exhibit larger effects than females, and our population was female. (2) Stimuli: sound and tactile stimuli lead to larger effects than spatial stimuli, which we used. (3) Responses: Go/Go tasks (where both P and N require active responses) produce larger effects than Go/No-go tasks; we tested the latter. (4) Reinforcement: methods with rewarded/punished stimuli or differentially rewarded stimuli generate larger effects than rewarded/unreinforced stimuli, which we used. In addition, cognitive tasks can be inherently rewarding93–95. Thus, performing the judgement bias task may have itself influenced cows’ emotional state, especially in the unstimulating PEN treatment. Another potential confound in spatial judgement bias tasks is olfactory cues, which can signal reward presence. However, our cows were no faster to P-Rew buckets than P-Unr buckets, suggesting that olfaction did not play a role. Additionally, repeated judgement bias testing can increase response latencies as subjects learn the probes are unreinforced65. We observed this effect, but our statistical models accounted for repeated measures. Some combination of these methodological factors potentially overrode treatment differences in judgement bias.
In conclusion, giving dairy cattle pasture access appears to induce more positive emotional states than cubicle housing. We previously showed that cows are more comfortable at pasture: they exhibit longer lying times, less restlessness, and greater herd synchrony. These behaviour data are partially consistent with the present findings, collected during the same experiment. We found no difference in judgement bias between cows with and without pasture access. In our judgement bias task, however, the pasture treatment was slower to approach a known reward. This effect implies reduced reward anticipation, suggesting that cows in the pasture-based system had more rewarding lives. Collectively, our results indicate that pasture access improves emotional wellbeing in dairy cows.
Acknowledgements
We thank the Agri-food and Biosciences Institute and its staff, especially Mike Davies, Deborah McConnell, and Gillian Scoley.
Author contributions
A.C., E.J.B., C.P.F., and G.A. designed the study; A.C., K.J., and H.K. carried out the experiments; A.C. & J.W. analysed the data; E.J.B., C.P.F., and G.A. supervised the project; A.C. drafted the manuscript; all authors reviewed the manuscript and accept responsibility for its contents.
Funding
A.C. was supported by a postgraduate studentship from Northern Ireland’s Department for the Economy. This research is part of a project that has received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme, Grant Number 851145.
Data availability
The datasets generated and analysed in this study are available from the corresponding author on reasonable request.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Ellis KA, Billington K, McNeil B, McKeegan DEF. Public opinion on UK milk marketing and dairy cow welfare. Anim. Welf. 2009;18(3):267–282. [Google Scholar]
- 2.Weinrich R, Kühl S, Zühlsdorf A, Spiller A. Consumer attitudes in Germany towards different dairy housing systems and their implications for the marketing of pasture raised milk. Int. Food Agribus. Manag. Rev. 2014;17(20):205–222. [Google Scholar]
- 3.Schuppli CA, von Keyserlingk MAG, Weary DM. Access to pasture for dairy cows: Responses from an online engagement. J. Anim. Sci. 2014;92(11):5185–5192. doi: 10.2527/jas.2014-7725. [DOI] [PubMed] [Google Scholar]
- 4.Hötzel MJ, Cardoso CS, Roslindo A, von Keyserlingk MAG. Citizens’ views on the practices of zero-grazing and cow-calf separation in the dairy industry: Does providing information increase acceptability? J. Dairy Sci. 2017;100(5):4150–4160. doi: 10.3168/jds.2016-11933. [DOI] [PubMed] [Google Scholar]
- 5.Cardoso CS, Hötzel MJ, Weary DM, Robbins JA, von Keyserlingk MAG. Imagining the ideal dairy farm. J. Dairy Sci. 2016;99(2):1663–1671. doi: 10.3168/jds.2015-9925. [DOI] [PubMed] [Google Scholar]
- 6.Robbins JA, von Keyserlingk MAG, Fraser D, Weary DM. Invited review: Farm size and animal welfare. J. Anim. Sci. 2016;94(12):5439–5455. doi: 10.2527/jas.2016-0805. [DOI] [PubMed] [Google Scholar]
- 7.Van den Pol-van Dasselaar A, Hennessy D, Isselstein J. Grazing of dairy cows in Europe—an in-depth analysis based on the perception of grassland experts. Sustainability. 2020;12(3):1098. doi: 10.3390/su12031098. [DOI] [Google Scholar]
- 8.USDA. Dairy Cattle Management Practices in the United States, 2014. www.aphis.usda.gov/animal_health/nahms/dairy/downloads/dairy14/Dairy14_dr_PartI.pdf (2016).
- 9.Jordbruksverket. Statens Jordbruksverks Författningssamling Saknr L., 106. http://www.jordbruksverket.se/download/18.229ea55815233ba0390e8c59/1452694447806/Landsbygdsprogrammet$+$2014-2020.pdf (2017).
- 10.EFSA Scientific opinion on welfare of dairy cows in relation to behaviour, fear and pain based on a risk assessment with special reference to the impact of housing, feeding, management and genetic selection. EFSA J. 2009;7:1139. doi: 10.2903/j.efsa.2009.1139. [DOI] [Google Scholar]
- 11.Charlton GL, Rutter SM, East M, Sinclair LA. Effects of providing total mixed rations indoors and on pasture on the behavior of lactating dairy cattle and their preference to be indoors or on pasture. J. Dairy Sci. 2011;94(8):3875–3884. doi: 10.3168/jds.2011-4172. [DOI] [PubMed] [Google Scholar]
- 12.Charlton GL, Rutter SM, East M, Sinclair LA. The motivation of dairy cows for access to pasture. J. Dairy Sci. 2013;96(7):4387–4396. doi: 10.3168/jds.2012-6421. [DOI] [PubMed] [Google Scholar]
- 13.Falk AC, Weary DM, Winckler C, Von Keyserlingk MAG. Preference for pasture versus freestall housing by dairy cattle when stall availability indoors is reduced. J. Dairy Sci. 2012;95(11):6409–6415. doi: 10.3168/jds.2011-5208. [DOI] [PubMed] [Google Scholar]
- 14.Legrand AL, Von Keyserlingk MAG, Weary DM. Preference and usage of pasture versus free-stall housing by lactating dairy cattle. J. Dairy Sci. 2009;92(8):3651–3658. doi: 10.3168/jds.2008-1733. [DOI] [PubMed] [Google Scholar]
- 15.Shepley E, Bergeron R, Vasseur E. Daytime summer access to pasture vs free-stall barn in dairy cows with year-long outdoor experience: A case study. Appl. Anim. Behav. Sci. 2017;192:10–14. doi: 10.1016/j.applanim.2016.11.003. [DOI] [Google Scholar]
- 16.Motupalli PR, Sinclair LA, Charlton GL, Bleach EC, Rutter SM. Preference and behavior of lactating dairy cows given free access to pasture at two herbage masses and two distances. J. Anim. Sci. 2014;92(11):5175–5184. doi: 10.2527/jas.2014-8046. [DOI] [PubMed] [Google Scholar]
- 17.Von Keyserlingk MA, Cestari AA, Franks B, Fregonesi JA, Weary DM. Dairy cows value access to pasture as highly as fresh feed. Sci. Rep. 2017;7:44953. doi: 10.1038/srep44953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tucker CB, Jensen MB, de Passillé AM, Hänninen L, Rushen J. Invited review: Lying time and the welfare of dairy cows. J. Dairy Sci. 2021;104(1):20–46. doi: 10.3168/jds.2019-18074. [DOI] [PubMed] [Google Scholar]
- 19.Crump A, Jenkins K, Bethell EJ, Ferris CP, Arnott G. Pasture access affects behavioral indicators of wellbeing in dairy cows. Animals. 2019;9(11):902. doi: 10.3390/ani9110902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Olmos, et al. Hoof disorders, locomotion ability and lying times of cubicle-housed compared to pasture-based dairy cows. Livest. Sci. 2009;125(2–3):199–207. doi: 10.1016/j.livsci.2009.04.009. [DOI] [Google Scholar]
- 21.der Tol V, et al. Frictional forces required for unrestrained locomotion in dairy cattle. J. Dairy Sci. 2005;88(2):615–624. doi: 10.3168/jds.S0022-0302(05)72725-9. [DOI] [PubMed] [Google Scholar]
- 22.Haskell MJ, Rennie LJ, Bowell VA, Bell MJ, Lawrence AB. Housing system, milk production, and zero-grazing effects on lameness and leg injury in dairy cows. J. Dairy Sci. 2006;89(11):4259–4266. doi: 10.3168/jds.S0022-0302(06)72472-9. [DOI] [PubMed] [Google Scholar]
- 23.Goldberg, et al. The influence of intensively managed rotational grazing, traditional continuous grazing, and confinement housing on bulk tank milk quality and udder health. J. Dairy Sci. 1992;75(1):96–104. doi: 10.3168/jds.S0022-0302(92)77743-1. [DOI] [PubMed] [Google Scholar]
- 24.Washburn SP, White SL, Green JT, Jr, Benson GA. Reproduction, mastitis, and body condition of seasonally calved Holstein and Jersey cows in confinement or pasture systems. J. Dairy Sci. 2002;85(1):105–111. doi: 10.3168/jds.S0022-0302(02)74058-7. [DOI] [PubMed] [Google Scholar]
- 25.Miller K, Wood-Gush DGM. Some effects of housing on the social behaviour of dairy cows. Anim. Sci. 1991;53(3):271–278. doi: 10.1017/S0003356100020262. [DOI] [Google Scholar]
- 26.Van Iaer E, Moons CPH, Sonck B, Tuyttens FAM. Importance of outdoor shelter for cattle in temperate climates. Livest. Sci. 2014;159:87–101. doi: 10.1016/j.livsci.2013.11.003. [DOI] [Google Scholar]
- 27.Charlier J, Claerebout E, De Mûelenaere E, Vercruysse J. Associations between dairy herd management factors and bulk tank milk antibody levels against Ostertagia ostertagi. Vet. Parasitol. 2005;133(1):91–100. doi: 10.1016/j.vetpar.2005.05.030. [DOI] [PubMed] [Google Scholar]
- 28.Kolver ES, Muller LD. Performance and nutrient intake of high producing Holstein cows consuming pasture or a total mixed ration. J. Dairy Sci. 1998;81(5):1403–1411. doi: 10.3168/jds.S0022-0302(98)75704-2. [DOI] [PubMed] [Google Scholar]
- 29.Arnott G, Ferris CP, O’Connell NE. Welfare of dairy cows in continuously housed and pasture-based production systems. Animal. 2017;11(2):261–273. doi: 10.1017/S1751731116001336. [DOI] [PubMed] [Google Scholar]
- 30.Charlton GL, Rutter SM. The behaviour of housed dairy cattle with and without pasture access: A review. Appl. Anim. Behav. Sci. 2017;192:2–9. doi: 10.1016/j.applanim.2017.05.015. [DOI] [Google Scholar]
- 31.Mee JF, Boyle LA. Assessing whether dairy cow welfare is “better” in pasture-based than in confinement-based management systems. N. Z. Vet. J. 2020;68(3):168–177. doi: 10.1080/00480169.2020.1721034. [DOI] [PubMed] [Google Scholar]
- 32.Smid AMC, Weary DM, von Keyserlingk MAG. The influence of different types of outdoor access on dairy cattle behavior. Front. Vet. Sci. 2020;7:257. doi: 10.3389/fvets.2020.00257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Boissy, et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007;92(3):375–397. doi: 10.1016/j.physbeh.2007.02.003. [DOI] [PubMed] [Google Scholar]
- 34.Fraser D, Duncan IJ. ‘Pleasures’, ‘pains’ and animal welfare: Toward a natural history of affect. Anim. Welf. 1998;7:383–396. [Google Scholar]
- 35.Spruijt BM, van den Bos R, Pijlman FT. A concept of welfare based on reward evaluating mechanisms in the brain: Anticipatory behaviour as an indicator for the state of reward systems. Appl. Anim. Behav. Sci. 2001;72(2):145–171. doi: 10.1016/S0168-1591(00)00204-5. [DOI] [PubMed] [Google Scholar]
- 36.Crump, et al. Emotion in animal contests. Proc. R. Soc. B. 2020;287(1939):20201715. doi: 10.1098/rspb.2020.1715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ede T, Lecorps B, von Keyserlingk MAG, Weary DM. Symposium review: Scientific assessment of affective states in dairy cattle. J. Dairy Sci. 2019;102(11):10677–10694. doi: 10.3168/jds.2019-16325. [DOI] [PubMed] [Google Scholar]
- 38.Kremer L, Holkenborg SK, Reimert IS, Bolhuis JE, Webb LE. The nuts and bolts of animal emotion. Neurosci. Biobehav. Rev. 2020;113:273–286. doi: 10.1016/j.neubiorev.2020.01.028. [DOI] [PubMed] [Google Scholar]
- 39.Mendl M, Burman OH, Paul ES. An integrative and functional framework for the study of animal emotion and mood. Proc. R. Soc. B. 2010;277(1696):2895–2904. doi: 10.1098/rspb.2010.0303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Russell JA. A circumplex model of affect. J. Pers. Soc. Psychol. 1980;39(6):1161. doi: 10.1037/h0077714. [DOI] [Google Scholar]
- 41.Désiré L, Boissy A, Veissier I. Emotions in farm animals: A new approach to animal welfare in applied ethology. Behav. Processes. 2002;60(2):165–180. doi: 10.1016/S0376-6357(02)00081-5. [DOI] [PubMed] [Google Scholar]
- 42.Crump A, Arnott G, Bethell EJ. Affect-driven attention biases as animal welfare indicators: Review and methods. Animals. 2018;8(8):136. doi: 10.3390/ani8080136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mendl M, Paul ES. Animal affect and decision-making. Neurosci. Biobehav. Rev. 2020;112:144–163. doi: 10.1016/j.neubiorev.2020.01.025. [DOI] [PubMed] [Google Scholar]
- 44.Paul ES, Harding EJ, Mendl M. Measuring emotional processes in animals: The utility of a cognitive approach. Neurosci. Biobehav. Rev. 2005;29(3):469–491. doi: 10.1016/j.neubiorev.2005.01.002. [DOI] [PubMed] [Google Scholar]
- 45.Blanchette I, Richards A. The influence of affect on higher level cognition: A review of research on interpretation, judgement, decision making and reasoning. Cogn. Emot. 2010;24(4):561–595. doi: 10.1080/02699930903132496. [DOI] [Google Scholar]
- 46.Everaert J, Podina IR, Koster EH. A comprehensive meta-analysis of interpretation biases in depression. Clin. Psychol. Rev. 2017;58:33–48. doi: 10.1016/j.cpr.2017.09.005. [DOI] [PubMed] [Google Scholar]
- 47.Hirsch CR, Meeten F, Krahé C, Reeder C. Resolving ambiguity in emotional disorders: The nature and role of interpretation biases. Annu. Rev. Clin. Psychol. 2016;12:281–305. doi: 10.1146/annurev-clinpsy-021815-093436. [DOI] [PubMed] [Google Scholar]
- 48.Schoth DE, Liossi C. A systematic review of experimental paradigms for exploring biased interpretation of ambiguous information with emotional and neutral associations. Front. Psychol. 2017;8:171. doi: 10.3389/fpsyg.2017.00171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Stuijfzand S, Creswell C, Field AP, Pearcey S, Dodd H. Research Review: Is anxiety associated with negative interpretations of ambiguity in children and adolescents? A systematic review and meta-analysis. J. Child Psychol. Psychiatry. 2018;59(11):1127–1142. doi: 10.1111/jcpp.12822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Harding EJ, Paul ES, Mendl M. Animal behaviour: Cognitive bias and affective state. Nature. 2004;427(6972):312. doi: 10.1038/427312a. [DOI] [PubMed] [Google Scholar]
- 51.Bethell EJ. A “how-to” guide for designing judgment bias studies to assess captive animal welfare. J. Appl. Anim. Welf. Sci. 2015;18:S18–S42. doi: 10.1080/10888705.2015.1075833. [DOI] [PubMed] [Google Scholar]
- 52.Lagisz, et al. Optimism, pessimism and judgement bias in animals: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2020;118:3–17. doi: 10.1016/j.neubiorev.2020.07.012. [DOI] [PubMed] [Google Scholar]
- 53.Mendl M, Burman OH, Parker RM, Paul ES. Cognitive bias as an indicator of animal emotion and welfare: Emerging evidence and underlying mechanisms. Appl. Anim. Behav. Sci. 2009;118(3):161–181. doi: 10.1016/j.applanim.2009.02.023. [DOI] [Google Scholar]
- 54.Neville, et al. Pharmacological manipulations of judgement bias: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2020;108:269–286. doi: 10.1016/j.neubiorev.2019.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Roelofs S, Boleij H, Nordquist RE, van der Staay FJ. Making decisions under ambiguity: Judgment bias tasks for assessing emotional state in animals. Front. Behav. Neurosci. 2016;10:119. doi: 10.3389/fnbeh.2016.00119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Burman OHP, Parker R, Paul ES, Mendl M. A spatial judgement task to determine background emotional state in laboratory rats, Rattus norvegicus. Anim. Behav. 2008;76:801–809. doi: 10.1016/j.anbehav.2008.02.014. [DOI] [Google Scholar]
- 57.Baciadonna L, McElligott AG. The use of judgement bias to assess welfare in farm livestock. Anim. Welf. 2015;24:81–91. doi: 10.7120/09627286.24.1.081. [DOI] [Google Scholar]
- 58.Neave HW, Daros RR, Costa JH, von Keyserlingk MAG, Weary DM. Pain and pessimism: Dairy calves exhibit negative judgement bias following hot-iron disbudding. PLoS One. 2013;8(12):e80556. doi: 10.1371/journal.pone.0080556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Lecorps B, Ludwig BR, von Keyserlingk MAG, Weary DM. Pain-induced pessimism and anhedonia: Evidence from a novel probability-based judgment bias test. Front. Behav. Neurosci. 2019;13:20. doi: 10.3389/fnbeh.2019.00054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Daros RR, Costa JH, von Keyserlingk MAG, Hötzel MJ, Weary DM. Separation from the dam causes negative judgement bias in dairy calves. PLoS One. 2014;9(5):e98429. doi: 10.1371/journal.pone.0098429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Bučková K, Špinka M, Hintze S. Pair housing makes calves more optimistic. Sci. Rep. 2019;9(1):1–9. doi: 10.1038/s41598-019-56798-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Henry S, Fureix C, Rowberry R, Bateson M, Hausberger M. Do horses with poor welfare show ‘pessimistic’ cognitive biases? Sci. Nat. 2017;104(1–2):8. doi: 10.1007/s00114-016-1429-1. [DOI] [PubMed] [Google Scholar]
- 63.Löckener S, Reese S, Erhard M, Wöhr AC. Pasturing in herds after housing in horseboxes induces a positive cognitive bias in horses. J. Vet. Behav. 2016;11:50–55. doi: 10.1016/j.jveb.2015.11.005. [DOI] [Google Scholar]
- 64.Crump, et al. Is the grass half-full? Investigating optimism as a welfare indicator for dairy cows with and without pasture-access. Pharmacol. Rep. 2019;71(6):1308. doi: 10.1016/j.pharep.2019.09.003. [DOI] [Google Scholar]
- 65.Doyle, et al. The effect of repeated testing on judgement biases in sheep. Behav. Processes. 2010;83(3):349–352. doi: 10.1016/j.beproc.2010.01.019. [DOI] [PubMed] [Google Scholar]
- 66.Špinka M. Social dimension of emotions and its implication for animal welfare. Appl. Anim. Behav. Sci. 2012;138(3–4):170–181. doi: 10.1016/j.applanim.2012.02.005. [DOI] [Google Scholar]
- 67.AHDB. Agriculture and Horticulture Development Board. https://dairy.ahdb.org.uk/resourceslibrary/technical-information/health-welfare/mobility-score-instructions/#.W2w3w9JKjIU (2020).
- 68.Hintze, et al. A cross-species judgement bias task: Integrating active trial initiation into a spatial Go/No-go task. Sci. Rep. 2018;8(1):5104. doi: 10.1038/s41598-018-23459-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Lecorps B, Weary DM, Keyserlingk MAG. Pessimism and fearfulness in dairy calves. Sci. Rep. 2018;8(1):1421. doi: 10.1038/s41598-017-17214-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Webb, et al. What do calves choose to eat and how do preferences affect behaviour? Appl. Anim. Behav. Sci. 2014;161:7–19. doi: 10.1016/j.applanim.2014.09.016. [DOI] [Google Scholar]
- 71.Bates DM, Mächler M, Bolker B, Walker S. Fitting linear mixed-effects models using lme4”. J. Stat. Softw. 2015;67(1):1–48. doi: 10.18637/jss.v067.i01. [DOI] [Google Scholar]
- 72.Lenth R, Lenth MR. Package ‘lsmeans’. Am. Stat. 2018;34(4):216–221. [Google Scholar]
- 73.Gygax L. The A to Z of statistics for testing cognitive judgement bias. Anim. Behav. 2014;95:59–69. doi: 10.1016/j.anbehav.2014.06.013. [DOI] [Google Scholar]
- 74.Trimmer PC, Paul ES, Mendl MT, McNamara JM, Houston AI. On the evolution and optimality of mood states. Behav. Sci. 2013;3(3):501–521. doi: 10.3390/bs3030501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Anderson MH, Munafò MR, Robinson ES. Investigating the psychopharmacology of cognitive affective bias in rats using an affective tone discrimination task. Psychopharmacology. 2013;226(3):601–613. doi: 10.1007/s00213-012-2932-5. [DOI] [PubMed] [Google Scholar]
- 76.Seehuus B, Mendl M, Keeling LJ, Blokhuis H. Disrupting motivational sequences in chicks: Are there affective consequences? Appl. Anim. Behav. Sci. 2013;148(1–2):85–92. doi: 10.1016/j.applanim.2013.07.008. [DOI] [Google Scholar]
- 77.Burman, et al. Using judgement bias to measure positive affective state in dogs. Appl. Anim. Behav. Sci. 2011;132(3–4):160–168. doi: 10.1016/j.applanim.2011.04.001. [DOI] [Google Scholar]
- 78.Freymond, et al. Behaviour of horses in a judgment bias test associated with positive or negative reinforcement. Appl. Anim. Behav. Sci. 2014;158:34–45. doi: 10.1016/j.applanim.2014.06.006. [DOI] [Google Scholar]
- 79.Verbeek E, Ferguson D, Lee C. Are hungry sheep more pessimistic? The effects of food restriction on cognitive bias and the involvement of ghrelin in its regulation. Physiol. Behav. 2014;123:67–75. doi: 10.1016/j.physbeh.2013.09.017. [DOI] [PubMed] [Google Scholar]
- 80.Kilgour RJ. In pursuit of “normal”: A review of the behaviour of cattle at pasture. Appl. Anim. Behav. Sci. 2012;138(1–2):1–11. doi: 10.1016/j.applanim.2011.12.002. [DOI] [Google Scholar]
- 81.Van der Harst JE, Spruijt BM. Tools to measure and improve animal welfare: Reward-related behaviour. Anim. Welf. 2007;16:67–73. [Google Scholar]
- 82.Watters JV, Krebs BL. Assessing and enhancing the welfare of animals with equivocal and reliable cues. Animals. 2019;9(9):680. doi: 10.3390/ani9090680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Anderson C, von Keyserlingk MAG, Lidfors LM, Weary DM. Anticipatory behaviour in animals: A critical review. Anim. Welf. 2020;29(3):231–238. doi: 10.7120/09627286.29.3.231. [DOI] [Google Scholar]
- 84.Neave HW, Webster JR, Zobel G. Anticipatory behaviour as an indicator of the welfare of dairy calves in different housing environments. PLoS One. 2021;16(1):e0245742. doi: 10.1371/journal.pone.0245742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Van der Harst JE, Baars AM, Spruijt BM. Standard housed rats are more sensitive to rewards than enriched housed rats as reflected by their anticipatory behaviour. Behav. Brain Res. 2003;142(1–2):151–156. doi: 10.1016/S0166-4328(02)00403-5. [DOI] [PubMed] [Google Scholar]
- 86.Treadway MT, Zald DH. Reconsidering anhedonia in depression: Lessons from translational neuroscience. Neurosci. Biobehav. Rev. 2011;35(3):537–555. doi: 10.1016/j.neubiorev.2010.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Clegg IL, Delfour F. Cognitive judgement bias is associated with frequency of anticipatory behavior in bottlenose dolphins. Zoo Biol. 2018;37(2):67–73. doi: 10.1002/zoo.21400. [DOI] [PubMed] [Google Scholar]
- 88.Wichman A, Keeling LJ, Forkman B. Cognitive bias and anticipatory behaviour of laying hens housed in basic and enriched pens. Appl. Anim. Behav. Sci. 2012;140(1–2):62–69. doi: 10.1016/j.applanim.2012.05.006. [DOI] [Google Scholar]
- 89.Horváth M, Pichová K, Košťál Ľ. The effects of housing conditions on judgement bias in Japanese quail. Appl. Anim. Behav. Sci. 2016;185:121–130. doi: 10.1016/j.applanim.2016.09.007. [DOI] [Google Scholar]
- 90.Carreras, et al. Housing conditions do not alter cognitive bias but affect serum cortisol, qualitative behaviour assessment and wounds on the carcass in pigs. Appl. Anim. Behav. Sci. 2016;185:39–44. doi: 10.1016/j.applanim.2016.09.006. [DOI] [Google Scholar]
- 91.Ruet, et al. Effects of a temporary period on pasture on the welfare state of horses housed in individual boxes. Appl. Anim. Behav. Sci. 2020;105027:20. [Google Scholar]
- 92.Anderson DJ, Adolphs R. A framework for studying emotions across species. Cell. 2014;157(1):187–200. doi: 10.1016/j.cell.2014.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Hagen K, Broom DM. Emotional reactions to learning in cattle. Appl. Anim. Behav. Sci. 2004;85(3–4):203–213. doi: 10.1016/j.applanim.2003.11.007. [DOI] [Google Scholar]
- 94.Manteuffel G, Langbein J, Puppe B. From operant learning to cognitive enrichment in farm animal housing: Bases and applicability. Anim. Welf. 2009;18(1):87–95. [Google Scholar]
- 95.Meagher RK, Strazhnik E, von Keyserlingk MAG, Weary DM. Assessing the motivation to learn in cattle. Sci. Rep. 2020;10(1):1–7. doi: 10.1038/s41598-020-63848-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Watters JV. Searching for behavioral indicators of welfare in zoos: Uncovering anticipatory behavior. Zoo Biol. 2014;33(4):251–256. doi: 10.1002/zoo.21144. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets generated and analysed in this study are available from the corresponding author on reasonable request.