

Abstract
Severe Acute Respiratory Syndrome-Coronavirus-2 (SARS-CoV-2), the etiological agent of Coronavirus Diseases 2019 (COVID-19), triggers an adaptive immunity in the infected host that results in the production of virus-specific antibodies and T cells. Although kinetic and quantitative aspects of antibodies have been analyzed in large patient cohorts, similar information about SARS-CoV-2-specific T cells are scarce. We summarize the available knowledge of quantitative and temporal features of the SARS-CoV-2 T-cell response in this review. Currently, most of the data are derived only from the analysis of the circulatory compartment. Despite this limitation, early appearance, multi-specificity and functionality of SARS-CoV-2-specific T cells are associated with accelerated viral clearance and with protection from severe COVID-19.
Keywords: COVID-19, T cells, virus-specific T cells
T and B cells act together to resolve viral infections but are performing nonredundant functions: B cells produce antibodies that recognize directly viral proteins and can prevent virus infection of the targeted cells. T cells instead recognize viral proteins not directly but only in association with Major Histocompatibility Complex (MHC)-classes I and II molecules displayed on the surface of the cells. MHC-class I molecules are displayed in variable quantities at the surface of all nucleated cells of the body, and they present viral proteins that are synthesized within the cells. Consequently, T cells recognizing MHC-class I molecules complexed with viral peptides (CD8+ cytotoxic T cells) target specifically the cells where viruses are replicating. Through direct lysis of virus-infected cells or through the secretion of antiviral cytokines (mainly IFN-γ), CD8 T cells are involved in the direct suppression of viral production and in the containment of infection. The T cells recognizing viral proteins associated with MHC-class II (CD4+ helper T cells) perform a different task. MHC-class II is expressed principally only by professional antigen presenting cells (dendritic cells, monocytes, macrophages), which are not infected but present viral antigen that has been taken up from the surrounding environment. T-helper cells (or CD4 T cells), by recognizing viral antigen presented by the professional antigen-presenting cells, produce diverse cytokines (IL-2, IL-21, Interferon (IFN)-γ, Tumor Necrosis Factor (TNF)-alpha) essentially supporting the expansion and the maturation of CD8 T and B cells.
Despite the importance of such coordinated involvement of T and B cells in prevention and clearance of viral infections, we know that different viruses can preferentially require diverse contributions of the distinct components of adaptive immunity. For example, protection from highly direct cytopathic viruses requires a particularly efficient antibody response that limits infection of target cells. In contrast, low or nondirectly cytopathic viruses are better controlled by cytotoxic T cells [1].
Such distinction is still not established in human coronavirus infections but data in animal models support the role of T cells in viral protection. For example, mice are protected from mouse adapted SARS-CoV infection by CD4 T cells and by production of IFN-γ [2,3]. In a similar model, it was also shown that clearance of SARS-CoV infection was dependent on the expansion of an early and robust virus-specific CD8 T-cell response [4]. More importantly, protective studies performed in rhesus macaques challenged with SARS-CoV-2 demonstrated that CD8 T cells play an essential role in the control of infection in animals that have sub-optimal levels of neutralizing antibodies [5]. Furthermore, prospective studies in humans are starting to reveal the importance of SARS-CoV-2 T cells in disease protection [6].
In this review, we will discuss, based on the currently available knowledge, quantitative and kinetic parameters of SARS-CoV-2 T cells. Other recent reviews and commentaries have already discussed aspects of SARS-CoV-2 T cells related to protection, phenotype and knowledge gap [7–9]. We hope that a summary of the quantitative and kinetic aspects of the SARS-CoV-2-specific cellular immune response might provide a solid reference for future study design to better define the role of SARS-CoV-2 T cells in human infection and vaccination.
CHARACTERIZATION OF SARS-COV-2 T CELLS IN HUMANS: THE LIMITATIONS
The first studies of immune cellular parameters in patients with COVID-19 reported marked lymphopenia [10, 11], signs of T-cell activation and cytokine production impairment on total T cells [12–14]. These initial studies investigating the activation levels and functional profile of total T cells were then followed by the characterization of the real players, T cells specific for the different proteins of SARS-CoV-2. Within a few months of the start of the pandemic, several groups showed that individuals infected by SARS-CoV-2 do not only produce antibodies but possess SARS-CoV-2-specific CD4 and CD8 T cells. The first studies were done in COVID-19 convalescents [15–23] immediately followed by characterization of individuals after asymptomatic infection [24]. Importantly, most of these studies also reported the presence of SARS-CoV-2-specific T cells in variable frequencies in unexposed healthy individuals [15–18, 25].
Before discussing the kinetic and quantitative aspects of SARS-CoV-2-specific T cells, we need to point out some inherent limitations of these studies in humans. The first one is that SARS-CoV-2 T cells in humans have so far only been analyzed in peripheral blood but studies in animal models showed that virus-specific T cells are preferentially recruited into the respiratory tract [2, 4].
Circulating T cells detected in the blood therefore are the fraction of the total pool of SARS-CoV-2 T cells present in an infected patient. The numerical value of such fraction is unknown, it is unlikely to be a fixed ratio but it will be dependent on the degree of inflammatory processes present in the respiratory tract. As the surface area of the respiratory tract is large (∼145 m2), it is likely that a very large quantity of T cells is trapped there during infection, particularly in patients with severe disease. This could also explain the marked general lymphopenia detected in their peripheral blood [10, 23]. Of note, in severe influenza a specific enrichment of influenza-specific T cells in the lung (∼45 times more than blood) was reported [26]. Furthermore, functional differences between the virus-specific T cells resident in different anatomical sites were also reported in animal studies [2]. CD4 T cells present in the airway produce more cytokines than the other parenchymal or vascular T cells and their ability to produce concomitantly IFN-γ and IL-10 appears to be required for optimal protection [2].
Limitations are also introduced by the methods used for T cell analysis.
T-cell analysis with MHC-class I or II multimers is precise and do not require the functionality of the T cells, but a more limited T-cell repertoire is detectable in comparison to the use of an unbiased peptide library. For example, SARS-CoV-2-specific T-cell analysis performed with a large library of MHC-class I multimers detected some SARS-CoV-2-specific CD8 T cells (27a), but robust frequencies were detected only after in vitro enrichment [27b, 28]. Direct visualization of T cells with MHC-tetramers also allows in studying their possible ‘functional phenotype’ like the expression of exhaustion/inhibitory markers (PD-1, TIM-3, CTLA-4). However, the significance of the expression of these markers during acute viral infection correlates more with the activation status than with functional exhaustion [29].
The number of epitopes visualized in individual patients by MHC-tetramers complexed with peptides selected by epitope predictive algorithms is low (2–3 epitopes) [27b] in comparison to what was detected with assays based on the use of peptide libraries (10–15 different pools positive) [15, 21] or by minigenes expressing viral antigens [30] (∼3 epitopes for each MHC-class I). However, for their detection, T cells must be functionally efficient and produce cytokines or proliferate or express activation markers and there is an accumulating evidence showing a degree of reduced production of cytokines (particularly IFN-γ) in SARS-CoV-2 T cells of symptomatic COVID-19 patients [20, 31]. The data are still preliminary, and a full functional T cell exhaustion of SARS-CoV-2 T cells has not been demonstrated [29]. Nevertheless, T cells of symptomatic patients secrete lower levels of IFN-γ than the ones of individuals with asymptomatic infection [31]. This could be one of the reasons why analysis of T cells with ‘activation induced markers (AIM)’, namely, CD40L, OX40 and CD69 detect more SARS-CoV-2 T cells than the methods based on IFN-γ production (intracellular cytokine staining and ELISpot). One the other hand, it is important to mention that detection of SARS-CoV-2-specific T cells utilizing AIM expression obtained after 24 h of peptide stimulation has the potential to induce bystander T-cell activation which might inflate the number of T cells detected.
Which specific functional assays might provide T-cell quantifications that correlate more with their potential protective ability is still unknown, but certainly worth future evaluation. In HIV infection, for example, it is only the number of T cells producing high levels of cytokines that correlate with viral protection [32].
In addition, performing a comprehensive analysis of the full repertoire of T cells against the whole SARS-CoV-2 proteome is challenging. The full proteome of SARS-CoV-2 is large, about 10 000 amino acids. Antibodies have protective ability when targeting the antigen located on the surface of the virus and particularly the region binding to the ACE2 receptors [33, 34]. In contrast, understanding the protective ability of different T cells is complex and not necessarily correlated with their quantity. T cells can be specific for all the distinct viral proteins produced within an infected cell. The endogenous synthesized viral proteins are processed and presented as epitopes which can elicit protective T cells irrespective to their quantitative expression [35, 36]. Furthermore, only few studies with a limited number of COVID-19-recovered patients have performed a comprehensive analysis of virus-specific T cells to the full SARS-CoV-2 proteome with peptide libraries [15, 21] or with minigenes expressing antigens [30].
A more robust set of data is instead available for T cells recognizing structural proteins, particularly the viral antigens spike, membrane and nucleoprotein. These proteins represent less than a third of the whole SARS-CoV-2 proteome [37, 38]. Even though they might not always contain the immunodominant epitope [30], they are immunogenic for both CD4 and CD8 T cells, and appear to elicit a strong T-cell response both in symptomatic and asymptomatic individuals [15, 20, 21, 24, 31, 39]. There are, however, also criticisms of investigating T cells utilizing only synthetic peptides. T cells activated by peptides presented by antigen-presenting cells expressing high quantity of MHC-class I or II molecules might just be ‘peptide specific’ or low affinity T cells and not really able to recognize virus infected targets. This might overestimate the number of T cells able to recognize efficiently the virus-infected cells and this criticism has been raised particularly for the significance of SARS-CoV-2 T cells present in healthy unexposed individuals [40].
KINETICS OF SARS-COV-2 T CELLS: INDUCTION AND EXPANSION
Despite all the different caveats, the results produced by many different groups are starting to define some clear pattern of induction, expansion and contraction of the humoral and cellular immune response during and following SARS-CoV-2 infection.
The kinetics profile of the antibody response after coronavirus infection is well defined. Several studies have shown that antibodies are detectable in the sera of the majority of SARS-CoV-2-infected symptomatic individuals within 4–5 days after the onset of symptoms with levels that rise for at least 2 weeks and that are higher in cases with more severe disease [41–44].
The knowledge of virus-specific T-cell kinetics and its association with disease severity is instead still limited as fewer works have examined the kinetics of SARS-CoV-2-specfic T cells during the acute phase of infection.
We know that SARS-CoV-2 T cells are detected, like antibodies [45], in almost the totality of infected individuals after recovery. Virtually, all SARS-CoV-2-infected symptomatic individuals positive for antibodies against Nucleoprotein (NP) or Spike possess a broad repertoire of T cells recognizing different structural proteins of SARS-CoV-2 (i.e. NP, M, Spike, ORF3a) [15, 20, 21].
Importantly, there is however also a discordance between virus-specific antibody levels and T-cell responses. For example, in some individuals, particularly the ones with mild or asymptomatic infection, SARS-CoV-2 multi-specific T cells are detectable despite the absence of a concomitant antibody response [24, 31, 46]; furthermore, different levels of neutralizing antibodies are detected in convalescent COVID-19 patients with similar quantity of SARS-CoV-2 T cells [47].
Like antibodies, SARS-CoV-2 T cells are detected within a week from onset of symptoms (likely ∼7–10 days from infection—time of infection cannot be precisely determined in human). This has been clearly shown in symptomatic COVID-19 patients [20, 48] and such T cells can be detected remarkably early. For example, Schulien et al. [28] visualized SARS-CoV-2 CD8+ T cells directly with MHC-multimers at Day 1 after symptoms, and showed that these virus-specific CD8 T cells were already activated. In other studies, where T cells were detected with functional assays (ELISpot for IFN-γ producing T cells or AIM), functionally responsive SARS-CoV-2 T cells were detected at 3–5 days after onset of symptoms (Figure 1) [20, 48]. This accelerated induction of SARS-CoV-2 T cells indicates that in many infected individuals, a perfect rapid and coordinated activation of innate and adaptive immunity are occurring. Note that, for example, infections with viruses, which evade or suppress innate immune recognition, like HBV or HCV, trigger an induction of virus-specific T cells only after 4–6 weeks from infection [49]. Indeed, the rapid expansion of the adaptive immune response is a proxy for the efficiency of the innate immunity triggering.
Figure 1:
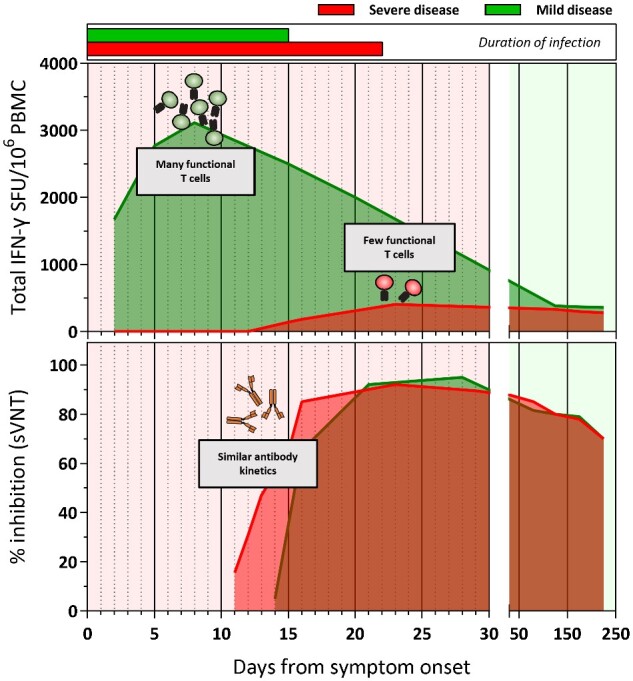
Kinetics of SARS-CoV-2-specific T cell and antibody response in COVID-19 patients with mild and severe disease.
The early induction of virus-specific antibodies and T cells after SARS-CoV-2 infections are however associated with different outcome.
Although antibodies against Spike and NP can be detected early in all the patients irrespective of clinical outcome, the early detection (<10 days from symptoms onset) of SARS-CoV-2 T cells in COVID-19 patients is associated with milder disease course and accelerated viral clearance [20, 48]. Delayed appearance (>15 days from symptoms onset) and weak induction of SARS-CoV-2 T cells were observed only in patients with severe COVID-19 [15, 19, 47], further supporting the concept that SARS-CoV-2 T cells are indispensable for viral control and that antibodies alone cannot clear an established infection. In severe COVID-19, the defective induction of a fully functional virus-specific T-cell response is then associated with what has been defined as ‘immunology misfiring’ [14]: robust and persistent activity of different components of innate immunity with a lack of a clear induction of a Th1-like immune response.
An open question about the SARS-CoV-2 T-cell kinetics is whether a rapid virus-specific T-cell induction also occurs in asymptomatic infection. The lack of symptoms conceals the possibility to recruit asymptomatic individuals at the beginning of infection and the lower level of SARS-CoV-2 antibodies detected in these individuals [43] have suggested that virus-specific B- and T-cell levels are directly proportional to symptom levels. Initial data derived from asymptomatic SARS-CoV-2 exposed individuals’ months after recovery supported this interpretation [21, 46]. However, a robust induction and functionally efficient SARS-CoV-2 T-cell response to levels even superior to what is detectable in symptomatic COVID-19 patients have been found in recently infected asymptomatic individuals [24, 31]. We think it is plausible to hypothesize that asymptomatic infection is characterized by a very rapid and efficient induction of a virus-specific cellular immune response.
One other question related to SARS-CoV-2 T-cell induction is whether T cells specific for the different SARS-CoV-2 viral proteins are induced with different kinetics and whether a preferential induction of a specific T-cell determinant is associated with better or worse viral control. For antibody responses, preferential induction of spike-specific antibodies is associated with faster SARS-CoV-2 clearance and mild disease, while an augmented anti-NP-specific antibody response is associated with worse diseases outcome [50]. However, if such findings are explained by the different protective efficiency of the antibodies against NP or Spike, a hypothetical parallel for T cells cannot be derived as we do not have any indications if T cells specific for different proteins have distinct protective values.
At the moment, most of the data indicate that the first detectable SARS-CoV-2-specific T-cell response is already multi-specific, with T cells simultaneously recognizing different epitopes in different proteins already at early stages of infection [16, 20, 51]. However, in a small study of patients analyzed longitudinally during the early phase of infection we noted a dynamic modulation of the hierarchy of SARS-CoV-2-specific T cells. T cells specific for ORF7 and ORF8 regions of SARS-CoV-2 were induced early and were more robustly detected in the early phases of infection than in convalescence [48]. These data need to be confirmed in larger studies, but the possibility that some viral antigens can be more efficient than the others in triggering a specific T-cell response might occur and it is compatible with different possibilities.
T cells specific for nonstructural proteins, like, for example, ORF-1-coded proteins, might be induced earlier as a consequence of the complex replication strategy of coronaviruses that requires the formation of a viral replicase–transcriptase complex essential for the subsequent transcription of the viral genome [52]. The early produced nonstructural proteins might trigger T cells ahead of T cells specific for structural viral proteins. Indications that this might occur are starting to appear in the literature, with demonstration of dominant epitopes detected in the ORF-1 region [30]. One other possible explanation for the differential kinetics of induction of T cells with different specificities can be derived from the pre-existence of SARS-CoV-2 cross-reactive T cells prior to the infection induced by other coronaviruses, vaccinations or other pathogens. As we briefly mentioned earlier, the existence of T cells specific for SARS-CoV-2 peptides in unexposed individuals has been reported in multiple studies [15–18, 25] and their impact in the protection and pathogenesis of SARS-CoV-2 infection is highly controversial [40, 53, 54].
Nevertheless, the possibility that SARS-CoV-2 T cell induction can be augmented by pre-existent memory T cells induced by other coronaviruses has been recently reported [55].
Furthermore, the first SARS-CoV-2 vaccines are designed to induce spike-specific immunity only, as spike antibodies have protective ability. Spike-specific T cells should theoretically have identical protective values than the T cells specific for other viral antigens. It will be however interesting to evaluate the impact that changes in the T cell immunodominance induced by vaccination will exert on the protective ability of the multi-specific T-cell response induced physiologically by the natural infection.
KINETICS OF SARS-COV-2 T CELLS: CONTRACTION PHASE
After the expansion phase that usually lasts for 10–20 days after symptom onset, T cells slowly decline but remain still detectable in the majority of individuals tested at least within 6–8 months after infection [31, 46, 55–59].
The kinetics of contraction of CD8 and CD4 T cells appears to slightly differ; both SARS-CoV-2-specific CD4 and CD8 T cells are present within the first 2 weeks after onset of symptoms, but while CD8 T cells show signs of progressive reduction after viral clearance (>1 month after infection) [20, 30], the SARS-CoV-2 CD4 T-cell frequency is more stable and appears higher in individuals tested in the initial recovery phase (1–2 months after infection) than immediately after infection [20]. These differential kinetics of expansion and contraction can explain the preferential detection of CD4 helper T cells in studies that have been investigated SARS-CoV-2 convalescent individuals [15, 17, 21, 55–59] and it mirrors the recent evidence of an identical persistence and progressive increase of SARS-CoV-2-specific B cells during the convalescent phase [57, 60]. The causes of such phenomena have not been demonstrated, but a sound hypothesis is that the viral antigens can persist as an antigenic depot in dendritic cells within lymph nodes [60].
Indeed, initial reports detecting a decline of SARS-CoV-2 antibodies immediately after the acute phase of infection [41, 61] have suggested that SARS-CoV-2-specific T cells were able to persist longer than the corresponding antibody response. However, more extended cross-sectional data showed that the virus-specific humoral and cellular immunity are, after an initial contraction phase [41, 48, 61], remarkably stable for at least 6–8 months after infection [31, 57, 58]. A recent cross-sectional study of COVID-19 recovered individuals analyzed >6 months after infection has calculated t1/2 of ∼3.5 months for CD4 and CD8 T cells detected with activation induced markers [57]. One other study that analyzed T cells with ELISpots has shown that 6 months after recovery, all COVID-19 convalescents have detectable T cells at variable frequency with a mean of 250 spots × million cells for T cells specific for NP, membrane and Spike [56].
One other unknown related to the contraction phase of SARS-CoV-2 T cells is whether this is faster in SARS-CoV-2-infected individuals in relation to the severity of disease. A recent comparative analysis of the SARS-CoV-2-specific T cells in asymptomatic versus symptomatic groups detected a very similar magnitude of SARS-CoV-2 T-cell responses within 1–3 months after infection [31]. However, a faster decline of the SARS-CoV-2 T-cell quantities was observed in asymptomatic individuals studied 6 months after infection compared with symptomatic COVID-19 patients at the same timepoint [56]. More studies will be needed to define whether cellular immunity decline is associated with the degree of symptom severity.
In conclusion, in these initial paragraphs, we summarized most of the presently available data related to the kinetics of appearance and disappearance of SARS-CoV-2 T cells. Experimental evidence indicates that the kinetics of induction of a multi-specific T-cell response is an important parameter of antiviral efficacy and that such multi-specific T cells can persist after viral clearance. Whether the quantity of T cells persisting as memory cells months after recovery will be able to mediate protection is still unknown as it is unknown whether those kinetic parameters of T cells are associated with age, sex and concomitant pathologies, variables that play important roles in the severity of SARS-CoV-2 infection.
QUANTITY AND IMMUNODOMINANCE OF THE SARS-COV-2 T CELL RESPONSE
What is the overall magnitude of virus-specific T cells present in SARS-CoV-2-infected patients? The quantity of virus-specific T cells in other viral diseases varies: some viruses (like HCMV, EBV, HIV) elicit high quantities of T cells which can often reach frequencies of 5–10% of total circulating T cells [32, 62, 63], while other virus-specific T cells (HBV, HCV) are seldom detected in blood at frequencies >0.1% [64]. These differences are not due to a single cause, but the fact that different viruses target different cell types and organs are the important factor. T cells are not evenly distributed among the body. As we have already mentioned, T cells in SARS-CoV-2-infected individuals have been so far analyzed only in peripheral blood but studies in animal models [2, 4] or other human respiratory diseases [26] show that virus-specific T cells can be preferentially recruited in the respiratory tract.
We try to summarize in Tables 1 and 2 the known frequencies of total circulating SARS-CoV-2-specific T cells detected in SARS-CoV-2-infected individuals published in different manuscripts. Note that all T-cell responses that have been so far reported in SARS-CoV-2 infection are Th1/Th0-type and not Th2-type. We also compared the frequency of Spike-specific T cells induced by natural infection with the one induced by the SARS-CoV-2 vaccines that have terminated Phase II clinical trials at the time of writing and in which Spike-specific T cell quantities were reported [65–72]. Frequency of T cells found within 1–3 months from infection or 2–4 weeks after vaccination is displayed (Tables 1 and 2).
Table 1:
Summary of SARS-CoV-2-specific T-cell responses induced by natural infection
| References | Severity | Disease state | Sampling date (post disease onset) | Type of T-cell assay | Specificity | Median SFU/mil PBMCs | |
|---|---|---|---|---|---|---|---|
| Sekine et al. [24] | Mild | Convalescent | 1–3 months | IFN-γ ELISPOT | S | ∼110 | |
| Mild | Convalescent | 1–3 months | IFN-γ ELISPOT | S, M, N | ∼310 | ||
| Severe | Convalescent | 1–3 months | IFN-γ ELISPOT | S | ∼110 | ||
| Severe | Convalescent | 1–3 months | IFN-γ ELISPOT | S, M, N | ∼530 | ||
| Peng et al. [21] | Mild | Convalescent | 1–3 months | IFN-γ ELISPOT | S | ∼250 | |
| Mild | Convalescent | 1–3 months | IFN-γ ELISPOT | S, M, N | ∼500 | ||
| Severe | Convalescent | 1–3 months | IFN-γ ELISPOT | S | ∼500 | ||
| Severe | Convalescent | 1–3 months | IFN-γ ELISPOT | S, M, N | ∼1050 | ||
| Le Bert et al. [31] | 75% mild/25% severe | Convalescent | 1–3 months | IFN-γ ELISPOT | S | ∼80 | |
| 75% mild/25% severe | Convalescent | 1–3 months | IFN-γ ELISPOT | S, M, N | ∼350 | ||
| 75% mild/25% severe | Asymptomatic | NA | IFN-γ ELISPOT | S | ∼60 | ||
| 75% mild/25% severe | Asymptomatic | NA | IFN-γ ELISPOT | S, M, N | ∼250 | ||
|
| |||||||
| References | Severity | Disease state | Sampling date (post disease onset) | Type of T-cell assay | Specificity | Median +/CD4 (%) | Median +/CD8 |
|
| |||||||
| Grifoni et al. [15] | ALL | Convalescent | 1–3 months | AIM | S | ∼0.2 | ∼0.1 |
| ALL | Convalescent | 1–3 months | AIM | S, M, N | ∼0.5 | ∼0.25 | |
| Rydyznski Moderbacher et al. [20] | ALL | Convalescent | 1–3 months | AIM | S | ∼0.3 | ∼0.03 |
| ALL | Convalescent | 1–3 months | AIM | S, M, N | ∼0.65 | ∼0.1 | |
| ALL | Acute | <1 month | AIM | S | ∼0.08 | ∼0.02 | |
| ALL | Acute | <1 month | AIM | S, M, N | ∼0.16 | ∼0.07 | |
| Weiskopf et al. [16] | Severe | Acute | <1 month | AIM | S | ∼0.6 | ∼1 |
| Breton et al. [58] | 80% mild/20% severe | Convalescent | 1–6 months | IFN-γ ICS | S, M, N, ORF3a | ∼0.25 | ∼0.1 |
| Dan et al. [57] | 90% mild/10% severe | Acute/ Convalescent | Up to 8 months | AIM | S, M, N, ORF3a, NSP3 | ∼1.1–0.4 | ∼1–0.3 |
Table 2:
summary of SARS-CoV-2-specific T-cell response induced by different vaccines
| Study | Age group | Type of T-cell assay | When? | Max achievable median response (SFU per mil PBMCs; SFU per mil CD4/CD8 T cells; IFN-γ pg/ml) |
|---|---|---|---|---|
| Pfizer—mRNA | 18–55 | IFN-γ ELISPOT | 7-day post boost/28-day post prime | ∼2000/2000 |
| 18–55 | IFN-γ ELISPOT | 28-days post vaccine/no boost | ∼50/50 | |
| Astrazeneca—Adenoviral | 18–55 | IFN-γ ELISPOT | 14-days post vaccine/no boost | ∼800 |
| 18–55 | IFN-γ ELISPOT | 28-days post vaccine/no boost | ∼550 | |
| CanSino—Adenoviral | 18–55 (10% >55) | IFN-γ ELISPOT | 28-days post vaccine/no boost | ∼100 |
| Gamaleya—Adenoviral | 18–60 | IFN-γ secretion | 28-days post vaccine/no boost | ∼30 pg/ml |
| Sinovac—Inactivated | 18–59 | IFN-γ ELISPOT | 14-days post boost/28-days post prime | ∼50 |
| Sinopharm—Inactivated | 18–59; >60 (1:1) | Not done | NA | NA |
|
| ||||
| Study | Age group | Type of T-cell assay | When? |
Max achievable median response (%IFN-γ+ out of CD4/CD8 T cells) |
|
| ||||
| Moderna—mRNA | 18–55 | IFN-γ ICS | 14-day post boost/42-day post prime | ≤0.2/≤0.1% |
| >56 | IFN-γ ICS | 14-day post boost/42-day post prime | ∼0.3/≤0.1% | |
| Janssen J&J—Adenoviral | 18–55 | IFN-γ ICS | 14-day post single dose | ∼0.1/0.09% |
| >65 | IFN-γ ICS | 14-day post single dose | ∼0.3/0.05% | |
Overall, the results so far accumulated show that SARS-CoV-2 T-cell quantity is, like antibodies, highly heterogeneous among different patients. On average T cells specific for the different structural proteins (NP, M and Spike) within 2–3 months from COVID-19 recovery are present at 0.1–1% of total CD4 or CD8 T cells with methods that detect T-cell AIM. ELISpot results show quantities of ∼100–1000 spots × million peripheral blood mononuclear cells (PBMC) against different individual proteins. The overall quantity does not differ between mild symptomatic and asymptomatic SARS-CoV-2 T-exposed individuals, but the functionality of SARS-CoV-2 T cells in asymptomatic seems superior [31]. These data are also in accordance with results obtained in patients with severe COVID-19 who display lower quantities and lower functionality of SARS-CoV-2-specific T cells than the patients with mild disease [20, 39].
The frequency of Spike-specific T cells induced by different preparations of vaccines is also reported. The different vaccine preparations, perhaps, with the exception of the inactivated Sinovac vaccine [69], trigger variable quantities of Spike-specific CD4 and CD8 T cells. Note, however, that a quantitative comparison between Spike-specific T cells induced by natural infection and vaccination is often incongruous as Spike-specific T cells were measured latest at 28 days after vaccination [71].
It remains to be analyzed whether vaccinations can induce a long-lasting memory T-cell response to levels similar to what can be achieved by natural infection and whether the induced Spike-specific T cells play a role in the vaccine efficacy.
There are two other important questions related to the magnitude of SARS-CoV-2 T cells: one is its relation with antibody responses and the other is related to the immunodominance of different epitopes.
The relation between the magnitude of SARS-CoV-2 T cells and antibodies is ‘intricate’. The quantity of neutralizing antibodies and total SARS-CoV-2-specific CD4 and CD8 T cells correlates in convalescent mild COVID-19 patients [20, 56]. Also, the frequency of Spike-specific CD4+CXCR5+ (a phenotype marker that defines circulating follicular helper T cells) was positively correlated with high level of Spike antibody and neutralizing activity [22], despite most of the T-cell response is targeting the Spike protein outside the Receptor Binding Domain (RBD) region [51]. However, neutralizing antibody levels were inversely correlated with the quantity of Spike-specific CD4+ CCR6+ CXCR3− T cells able to produce IL-17 [21] and no correlations were also detected between NP-specific T cell responses (Th1-like) and the magnitude and titers of neutralizing or NP-specific antibodies [17].
Furthermore, even though a coordinated expansion of both cellular and humoral arms of adaptive immunity is often found in mild but not severe COVID-19 patients [20], a robust SARS-CoV-2 T cell response in the absence of antibodies was found in some patients with mild COVID-19 who successfully control the infection [20, 24, 31, 46, 55]. Interestingly, the scientific literature does not report any evidence of COVID-19 patients who controlled the infection only by eliciting an antibody response in the absence of detectable SARS-CoV-2 T cells [20, 48], while high titers of antibodies (even neutralizing antibodies) are present in severe disease [73].
The discrepancy between the presence of virus-specific T cells and the absence of specific antibodies is detected also in some asymptomatic cases [24, 31] and in other coronaviruses infections like Middle East Respiratory Syndrome- Coronavirus (MERS-CoV), in whom subjects who were likely in contact with the viruses present MERS-CoV-specific T cells but no antibodies [74, 75]. Similarly, SARS-CoV-specific T cells (CD4 and CD8 T cells) can be detected at 11 and 17 years after SARS-CoV infection [17, 76] in the absence of detectable antibodies.
The cause of this detection discrepancy is still unknown. Some individuals can mount initially exclusively a cellular or a humoral response after exposure. Demonstration of extrafollicular B-cell maturation particularly in severe COVID-19 patients [77] might partially explain the presence of strong antibody responses in the absence of T cells [78, 79] but the immunological mechanisms that lead to the exclusive induction of cellular immunity are not known. An alternative interpretation of these results is that humoral and cellular responses might decay in some people with different kinetics. The data collected within the first 6 months after infection do not support such interpretation since waning of antibody titres and T cells appear similar [56], but it is still possible, as seen in SARS-CoV infection [80], that at later time points the rate of reduction of antibodies and T cells could diverge.
Quantitative features of the antiviral T-cell response are not only related to the total numbers of SARS-CoV-2 T cells present but also on their diversity. CD8 and CD4 T cells can recognize different epitopes located in the same and/or different proteins. The SARS-CoV-2 T cell response in COVID-19 recovered individuals is highly multi-specific with T cells recognizing not only multiple proteins [15, 21, 24], but also multiple regions within a single protein [17, 27, 30, 51]. This diffuse repertoire of T cells recognizing several epitopes displays a very heterogeneous T cell receptor (TCR) repertoire with the presence of some public TCR usage shared by some individuals [81].
Whether multi-specificity is the key of protection is still uncertain. A correlation between multi-specificity and mild disease has been reported [27]. Hypothetically, the ability to mount a broad T-cell response against several epitopes located in different viral proteins might be advantageous and might avoid the selection of mutated viruses able to escape CD8 T-cell recognition. In other viral infections (i.e. HBV), the multi-specificity of the T-cell response appears to be an important determinant of viral clearance [82]. Nevertheless, the data at the moment are only showing associations and not causality. It might be possible that despite the broad T-cell multi-specificity observed in SARS-CoV-2-recovered individuals, robust T cells specific for a single or limited number of epitopes present in a single protein can be equally protective. Detailed analysis of different epitopes and their element of restrictions is starting to appear in the scientific literature [27, 30, 51] and such information will be indispensable to understand whether single T-cell determinants are more or less important for viral clearance during natural infection. This information will have also to be translated into the analysis of T cells induced by vaccines that, currently, include only the single Spike protein or only its RBD region. Such vaccines can certainly not induce the broad T-cell repertoire as seen by the natural infection, and this Spike-focused T-cell induction might not be ideal. Nevertheless, the protective response induced by a vaccine is likely to be mediated by the ability to obtain a coordinated induction of both Spike-specific antibodies and T cells. Note that a very efficacious prophylactic vaccine, the one against HBV, also elicits a combined antibody and T cell response focused only against the S antigen of the virus (the envelope protein of HBV) [83] despite HBV control after natural infection is correlating with induction of a multi-specific T-cell response [82]. The two viruses display different viral replication kinetics (peak viral replication for SARS-CoV-2 within 1 to 2 weeks from infection [73], 4–6 weeks after infection for HBV [49]), but this comparative hypothesis is in line with the initial evidence of the protective efficacy of Spike-based vaccines recently reported.
CONCLUDING REMARKS
The SARS-CoV-2 pandemic has changed our life. It has not only caused the loss of many people but it has also compromised and changed social and economic habits. The response to this crisis has also transformed the scientific world. The pace of new scientific information related to SARS-CoV-2 has been growing exponentially. Scientific hypotheses are confirmed or become obsolete not after years, but after few months. When we accepted to write this review on virus-specific T cells in SARS-CoV-2 infection, we did not fully realize that this time we were asked to summarize a scientific argument that is far from being established but is in dynamic development. The risk to describe phenomena that could soon be considered obsolete or insignificant is high. Nevertheless, we report here kinetics and quantitative features of virus-specific T cells that in our opinion have fundamental importance to define their role either in the protection or in the pathogenesis of the disease caused by SARS-CoV-2. We think that the most of the data support a protective role of the quantity and diversity of virus-specific T cells in SARS-CoV-2 infection. T cells are indispensable in mouse models of coronavirus infection, are critical in protective studies in rhesus monkeys and their early presence, functionality and multi-specificity are correlated with protection and rapid viral control in infected individuals. Nevertheless, more data are needed to fully understand the role of virus-specific T cells in different groups of patients with different disease severity, the relation of SARS-CoV-2 T cell quantity and function with age, sex and other pathologies affecting infected individuals, or the role of T cells in protective vaccination and in the pathogenesis of long-term COVID-19, in which a possible pathogenetic role of T cells can certainly not be excluded.
FUNDING
This study is supported by the Singapore Ministry of Health’s National Medical Research Council under its COVID-19 Research Fund (COVID19RF3-0060), the Singapore Ministry of Health’s National Medical Research Council MOH-000019 (MOH-StaR17Nov-0001) and National Research Foundation, Singapore (NRF-CRP17-2017-06).
CONFLICT OF INTEREST STATEMENT
A.Bertoletti, A.T.Tan and N.Le Bert declare that they submitted a patent for a method to monitor SARS-CoV-2 specific T cells in biological samples. A. Bertoletti reported personal fees for consultances from Oxford Immunotech and Qiagen in relation to SARS-CoV-2 immunity outside this work.
DATA AVAILABILITY STATEMENT
Data analyzed in this study were re-analysis of existing data, which are openly available at locations cited in the reference section.
REFERENCES
- 1. Hangartner L, Zinkernagel RM, Hengartner H.. Antiviral antibody responses: the two extremes of a wide spectrum. Nat Rev Immunol 2006;6:231–43. [DOI] [PubMed] [Google Scholar]
- 2. Zhao J, Zhao J, Mangalam AK. et al. Airway memory CD4 + T cells mediate protective immunity against emerging respiratory coronaviruses. Immunity 2016;44:1379–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. McMahan K, Yu J, Mercado NB. et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature 2021; 590: 630–634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Zhao J, Zhao J, Perlman S.. T cell responses are required for protection from clinical disease and for virus clearance in severe acute respiratory syndrome coronavirus-infected mice. J Virol 2010;84:9318–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. McMahan K, Yu J, Mercado NB. et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature 2021:590:630–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Wyllie D, Mulchandani R, Jones HE. et al. SARS-CoV-2 responsive T cell numbers are associated with protection from COVID-19: A prospective cohort study in keyworkers. medRxiv 2020;3:e2010182–24. [Google Scholar]
- 7. Altmann DM, Boyton RJ.. SARS-CoV-2 T cell immunity: Specificity, function, durability, and role in protection. Sci Immunol 2020;5:eabd6160. [DOI] [PubMed] [Google Scholar]
- 8. Lipsitch M, Grad YH, Sette A. et al. Cross-reactive memory T cells and herd immunity to SARS-CoV-2. Nat Rev Immunol 2020;20:709–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Karlsson AC, Humbert M, Buggert M.. The known unknowns of T cell immunity to COVID-19. Sci Immunol 2020;5:eabe8063–9. [DOI] [PubMed] [Google Scholar]
- 10. Chen G, Wu D, Guo W. et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J Clin Invest 2020;130:2620–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Liu J, Li S, Liu J. et al. Longitudinal characteristics of lymphocyte responses and cytokine profiles in the peripheral blood of SARS-CoV-2 infected patients. EBioMedicine 2020;55:102763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. De Biasi S, Meschiari M, Gibellini L. et al. Marked T cell activation, senescence, exhaustion and skewing towards TH17 in patients with COVID-19 pneumonia. Nat Commun 2020;11:1199–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Mazzoni A, Salvati L, Maggi L. et al. Impaired immune cell cytotoxicity in severe COVID-19 is IL-6 dependent. J Clin Invest 2020;130:4694–703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Lucas C, Wong P, Klein J. et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 2020;584:463–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Grifoni A, Weiskopf D, Ramirez SI. et al. Targets of T cell responses to SARS-CoV-2 coronavirus in humans with COVID-19 disease and unexposed individuals. Cell 2020;181:1489–1501.e15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Weiskopf D, Schmitz KS, Raadsen MP. et al. Phenotype and kinetics of SARS-CoV-2-specific T cells in COVID-19 patients with acute respiratory distress syndrome. Sci Immunol 2020;5:eabd2071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Le Bert N, Tan AT, Kunasegaran K. et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020;584:457–62. [DOI] [PubMed] [Google Scholar]
- 18. Braun J, Loyal L, Frentsch M. et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature 2020;587:270–4. [DOI] [PubMed] [Google Scholar]
- 19. Zhou R, To KK-W, Wong YC. et al. Acute SARS-CoV-2 infection impairs dendritic cell and T cell responses. Immunity 2020;53:864–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Rydyznski Moderbacher C, Ramirez SI, Dan JM. et al. Antigen-specific adaptive immunity to SARS-CoV-2 in acute COVID-19 and associations with age and disease severity. Cell 2020;183:996–1012.e19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Peng Y, Mentzer AJ, Liu G. et al. Broad and strong memory CD4+ and CD8+ T cells induced by SARS-CoV-2 in UK convalescent individuals following COVID-19. Nat Immunol 2020;21:1336–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Juno JA, Tan H-X, Lee WS. et al. Humoral and circulating follicular helper T cell responses in recovered patients with COVID-19. Nat Med 2020:1–22. [DOI] [PubMed] [Google Scholar]
- 23. Schub D, Klemis V, Schneitler S. et al. High levels of SARS-CoV-2-specific T cells with restricted functionality in severe courses of COVID-19. JCI Insight 2020;5:1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Sekine T, Perez-Potti A, Rivera-Ballesteros O. et al. Robust T cell immunity in convalescent individuals with asymptomatic or mild COVID-19. Cell 2020;183:158–168.e14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Mateus J, Grifoni A, Tarke A, et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science. 2020. 370 (6512) 89–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Zhao Y, Zhang Y-H, Denney L. et al. High levels of virus-specific CD4+ T cells predict severe pandemic influenza A virus infection. Am J Respir Crit Care Med 2012;186:1292–7. [DOI] [PubMed] [Google Scholar]
- 27a. Kared H, Redd AD, Bloch EM. et al. SARS-CoV-2-specific CD8+ T cell responses in convalescent COVID-19 individuals. J Clin Invest 2021;131, doi:10.1172/JCI145476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27b. Nelde A, Bilich T, Heitmann JS. et al. SARS-CoV-2-derived peptides define heterologous and COVID-19-induced T cell recognition. Nat Immunol 2020;8:247–12. [DOI] [PubMed] [Google Scholar]
- 28. Schulien I, Kemming J, Oberhardt V. et al. Characterization of pre-existing and induced SARS-CoV-2-specific CD8+ T cells. Nat Med 2020;181:1489–8. [DOI] [PubMed] [Google Scholar]
- 29. Rha M-S, Jeong HW, Ko J-H. et al. PD-1-Expressing SARS-CoV-2-specific CD8+ T cells are not exhausted, but functional in patients with COVID-19. Immunity 2021;54:44–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Ferretti AP, Kula T, Wang Y. et al. Unbiased screens show CD8+ T cells of COVID-19 patients recognize shared epitopes in SARS-CoV-2 that largely reside outside the spike protein. Immunity 2020;53:1095–1107.e3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Le Bert N, Clapham HE, Tan AT. et al. Highly functional virus-specific cellular immune response in asymptomatic SARS-CoV-2 infection. J Exp Med 2021;218, doi:10.1084/jem20202617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Betts MR. HIV nonprogressors preferentially maintain highly functional HIV-specific CD8+ T cells. Blood 2006;107:4781–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Schmidt F, Weisblum Y, Muecksch F. et al. Measuring SARS-CoV-2 neutralizing antibody activity using pseudotyped and chimeric viruses. J Exp Med 2020;217:284–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Schäfer A, Muecksch F, Lorenzi JCC. et al. Antibody potency, effector function, and combinations in protection and therapy for SARS-CoV-2 infection in vivo. J Exp Med 2020;218:828–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Yewdell JW. Confronting complexity: real-world immunodominance in antiviral CD8+ T cell responses. Immunity 2006;25:533–43. [DOI] [PubMed] [Google Scholar]
- 36. Gallimore A, Dumrese T, Hengartner H. et al. Protective immunity does not correlate with the hierarchy of virus-specific cytotoxic T cell responses to naturally processed peptides. J Exp Med 1998;187:1647–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Irigoyen N, Firth AE, Jones JD. et al. High-resolution analysis of coronavirus gene expression by RNA sequencing and ribosome profiling. PLoS Pathog 2016;12:e1005473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. de Wit E, van Doremalen N, Falzarano D. et al. SARS and MERS: recent insights into emerging coronaviruses. Nat Rev Microbiol 2016;14:523–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Sattler A, Angermair S, Stockmann H. et al. SARS–CoV-2–specific T cell responses and correlations with COVID-19 patient predisposition. J Clin Invest 2020;130:6477–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Bacher P, Rosati E, Esser D. et al. Low avidity CD4+ T cell responses to SARS-CoV-2 in unexposed individuals and humans with severe COVID-19. Immunity 2020;53:1258–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Long Q-X, Liu B-Z, Deng H-J. et al. Antibody responses to SARS-CoV-2 in patients with COVID-19. Nat Med 2020;26:845–8. [DOI] [PubMed] [Google Scholar]
- 42. Henss L, Scholz T, Rhein von C. et al. Analysis of humoral immune responses in SARS-CoV-2 infected patients. J Infect Dis 2020, doi: 10.1093/infdis/jiaa680. [DOI] [PubMed] [Google Scholar]
- 43. Long Q-X, Tang X-J, Shi Q-L. et al. Clinical and immunological assessment of asymptomatic SARS-CoV-2 infections. Nat Med 2020;63:706–12. [DOI] [PubMed] [Google Scholar]
- 44. Robbiani DF, Gaebler C, Muecksch F. et al. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature 2020;584:437–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Wajnberg A, Amanat F, Firpo A. et al. Robust neutralizing antibodies to SARS-CoV-2 infection persist for months. Science 2020;370:1227–30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Reynolds CJ, Swadling L, Gibbons JM. et al. Discordant neutralizing antibody and T cell responses in asymptomatic and mild SARS-CoV-2 infection. Sci Immunol 2020. doi:10.1126/sciimmunol.abf3698 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Chia WN, Zhu F, Ong WX. et al. Multifaceted dynamics of SARS-CoV-2 neutralising antibody responses predicts a wide range of immunity longevity from days to decades. Lancet Microbe 2021;9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Tan AT, Linster M, Tan CW. et al. Early induction of functional SARS-CoV-2-specific T cells associates with rapid viral clearance and mild disease in COVID-19 patients. Cell Rep 2021;53:108728–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Wieland SF, Chisari FV.. Stealth and cunning: hepatitis B and hepatitis C viruses. J Virol 2005;79:9369–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Atyeo C, Fischinger S, Zohar T. et al. Distinct early serological signatures track with SARS-CoV-2 survival. Immunity 2020;53:524–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Tarke A, Sidney J, Kidd CK. et al. Comprehensive analysis of T cell immunodominance and immunoprevalence of SARS-CoV-2 epitopes in COVID-19 cases. Cell Rep Med 2021;181:100204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Perlman S, Netland J.. Coronaviruses post-SARS: update on replication and pathogenesis. Nat Rev Microbiol 2009;7:439–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Sagar M, Reifler K, Rossi M. et al. Recent endemic coronavirus infection is associated with less severe COVID-19. J Clin Invest 2021;131. doi:10.1172/JCI143380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Welsh RM, Selin LK.. No one is naive: the significance of heterologous T-cell immunity. Nat Rev Immunol 2002;2:417–26. [DOI] [PubMed] [Google Scholar]
- 55. Bonifacius A, Tischer-Zimmermann S, Dragon AC. et al. COVID-19 immune signatures reveal stable antiviral T cell function despite declining humoral responses. Immunity 2021;54:340–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Zuo J, Dowell A, Pearce H. et al. Robust SARS-CoV-2-specific T-cell immunity is maintained at 6 months following primary infection. bioRxiv 2020, doi org./10.1101/2020.11.01.362319v1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Dan JM, Mateus J, Kato Y. et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021;371. doi:10.1126/science.abf4063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Breton G, Mendoza P, Hägglöf T. et al. Persistent cellular immunity to SARS-CoV-2 infection. J Exp Med 2021;218:4781–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Rodda LB, Netland J, Shehata L. et al. Functional SARS-CoV-2-specific immune memory persists after mild COVID-19. Cell 2021;184:169–183.e17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Gaebler C, Wang Z, Lorenzi JCC. et al. Evolution of antibody immunity to SARS-CoV-2. Nature 2021. Doi:10.1038/s41586-021-03207-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Seow J, Graham C, Merrick B. et al. Longitudinal observation and decline of neutralizing antibody responses in the three months following SARS-CoV-2 infection in humans. Nature Microbiol 2020;5:1598–607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Lübke M, Spalt S, Kowalewski DJ. et al. Identification of HCMV-derived T cell epitopes in seropositive individuals through viral deletion models. J Exp Med 2019;217:613–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Long HM, Meckiff BJ, Taylor GS.. The T-cell response to Epstein-Barr virus-new tricks from an old dog. Front Immunol 2019;10:2193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Tan AT, Loggi E, Boni C. et al. Host ethnicity and virus genotype shape the hepatitis B virus-specific T-cell repertoire. J Virol 2008;82:10986–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Zhu F-C, Guan X-H, Li Y-H. et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: a randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2020;396:479–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Ramasamy MN, Minassian AM, Ewer KJ. et al. Safety and immunogenicity of ChAdOx1 nCoV-19 vaccine administered in a prime-boost regimen in young and old adults (COV002): a single-blind, randomised, controlled, phase 2/3 trial. Lancet 2020;396:1979–93.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Walsh EE, Frenck RW Jr., Falsey AR. et al. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. N Engl J Med 2020;383:2439–450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Sahin U, Muik A, Derhovanessian E. et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature 2020;586:594–9. [DOI] [PubMed] [Google Scholar]
- 69. Logunov DY, Dolzhikova IV, Zubkova OV. et al. Safety and immunogenicity of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine in two formulations: two open, non-randomised phase 1/2 studies from Russia. Lancet 2020;396:887–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Sadoff J, Le Gars M, Shukarev G. et al. Safety and immunogenicity of the Ad26.COV2.S COVID-19 vaccine candidate: interim results of a phase 1/2a, double-blind, randomized, placebo-controlled trial. medRxiv 2020, doi: 10.1101/2020.09.23.20199604. [Google Scholar]
- 71. Zhang Y, Zeng G, Pan H. et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18–59 years: a randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect Dis 2020;21:181–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Jackson LA, Anderson EJ, Rouphael NG. et al. An mRNA vaccine against SARS-CoV-2—preliminary report. N Engl J Med 2020;383:1920–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Wang Y, Zhang L, Sang L. et al. Kinetics of viral load and antibody response in relation to COVID-19 severity. J Clin Invest 2020;130:5235–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Zhao J, Alshukairi AN, Baharoon SA. et al. Recovery from the Middle East respiratory syndrome is associated with antibody and T-cell responses. Sci Immunol 2017;2:eaan5393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Perlman S, Zumla A.. MERS-CoV in Africa—an enigma with relevance to COVID-19. Lancet Infect Dis 2020;21:303–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Ng O-W, Chia A, Tan AT. et al. Memory T cell responses targeting the SARS coronavirus persist up to 11 years post-infection. Vaccine 2016;34:2008–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Woodruff MC, Ramonell RP, Nguyen DC. et al. Extrafollicular B cell responses correlate with neutralizing antibodies and morbidity in COVID-19. Nat Immunol 2020;21:1506–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Fagarasan S, Honjo T.. T-Independent immune response: new aspects of B cell biology. Science 2000;290:89–92. [DOI] [PubMed] [Google Scholar]
- 79. Sweet RA, Ols ML, Cullen JL. et al. Facultative role for T cells in extrafollicular Toll-like receptor-dependent autoreactive B-cell responses in vivo. Proc Natl Acad Sci 2011;108:7932–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Cao W-C, Liu W, Zhang P-H. et al. Disappearance of antibodies to SARS-associated coronavirus after recovery. N Engl J Med 2007;357:1162–3. [DOI] [PubMed] [Google Scholar]
- 81. Shomuradova AS, Vagida MS, Sheetikov SA. et al. SARS-CoV-2 epitopes are recognized by a public and diverse repertoire of human T cell receptors. Immunity 2020:1–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Bertoletti A, Ferrari C.. Adaptive immunity in HBV infection. J.Hepatol 2016;64:S71–83. [DOI] [PubMed] [Google Scholar]
- 83. Shouval D. Hepatitis B vaccines. J.Hepatol 2003;39 (Suppl 1):S70–6. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data analyzed in this study were re-analysis of existing data, which are openly available at locations cited in the reference section.