

Abstract
We have synthesized several conformationally constrained dipeptide analogues as possible substrates for incorporation into proteins. These have included three cyclic dipeptides formed from Boc derivatives of 2,4-diaminobutyric acid, ornithine and lysine, having 5-, 6-, and 7-membered lactam rings, respectively. These dipeptides were used to activate a suppressor tRNA transcript, the latter of which had been prepared by in vitro transcription. Using modified E. coli ribosomes described previously, these activated suppressor tRNAs enabled the incorporation of the three cyclic dipeptides into dihydrofolate reductase (DHFR) at positions 18 and 49. The suppression yields increased with increasing lactam ring size and were found to proceed in suppression yields ranging from 3.4 to 8.9% at two different protein sites for the 5-, 6- and 7-membered lactam dipeptides. The greater facility of incorporation of the 7-membered lactam prompted us to prepare two 7-membered cyclic acylhydrazides (4 and 5) by 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide (EDCI)-mediated cyclization of amino acids having selectively protected hydrazine functional groups in their side chains. In common with the lactam dipeptides, acylhydrazide dipeptides 4 and 5 could be used to activate the same suppressor tRNA transcript and to incorporate the cyclic dipeptides into DHFR. They were incorporated into the same two DHFR sites in suppression yields ranging from 8.3 to 11.2%.
Keywords: Conformational constraint, Lactam and acyl hydrazide cassettes, Non-proteinogenic dipeptides, Protein modification
1. Introduction
Nature has provided numerous examples of peptide hormones and other peptide-based mediators of biological processes.1–5 In nature, it is not unusual for these to be employed under precise temporal and spatial control. In contrast, the use of such compounds to control biological processes experimentally, including during therapeutic intervention, requires consideration of the chemical and metabolic stability of the peptide analogues employed. Frequently, it also requires a consideration of peptide stability and conformation to optimize the desired properties of the peptides as agonists and antagonists.6–11
Peptide stability has long been studied, e.g. by the replacement of proteinogenic amino acids in peptides with amino acid and peptide surrogates resistant to natural peptidases, or which can help to stabilize the native conformation.12–14 Some of these structural surrogates can also confer useful conformational constraints, for example at peptide sites known to be conformationally labile or important for receptor recognition.15 In recent years, however, it has become increasingly common to control peptide conformation by creating peptides in which much of the structure has more limited conformational freedom, e.g. by the use of cyclic peptides. These can be made to restrain linear amino acid sequences, such as in cyclic peptides which mimic the RGD domain of fibronectin.16–18 Modified ribosomal systems are currently also employed in chemoenzymatic protocols to elaborate libraries of modified cyclic peptides containing non-proteinogenic amino acids.19–21
In comparison, the modification of proteins with non-proteinogenic amino acids to confer protein stability to cellular proteases is less common, although a number of examples may be cited.22,23 Likewise, while many modifications undoubtedly alter protein conformation, the purposeful construction of structural cassettes designed to constrain specific regions of proteins to a subset of the conformations usually accessible has not been pursued as actively.
Recently, we have studied the use of modified bacterial ribosomes24 to introduce dipeptides and dipeptidomimetic species into proteins.25–28 Presently, we describe the preparation of several cyclic dipeptides that are constrained conformationally, but which can be incorporated into proteins from an activated suppressor tRNA using modified bacterial ribosomes. In particular, E. coli ribosomal clone 010328R225 containing a modified 23S rRNA was shown to incorporate five conformationally constrained cyclic dipeptides (1 – 5) (Figure 1) into two different positions of dihydrofolate reductase.
Figure 1.
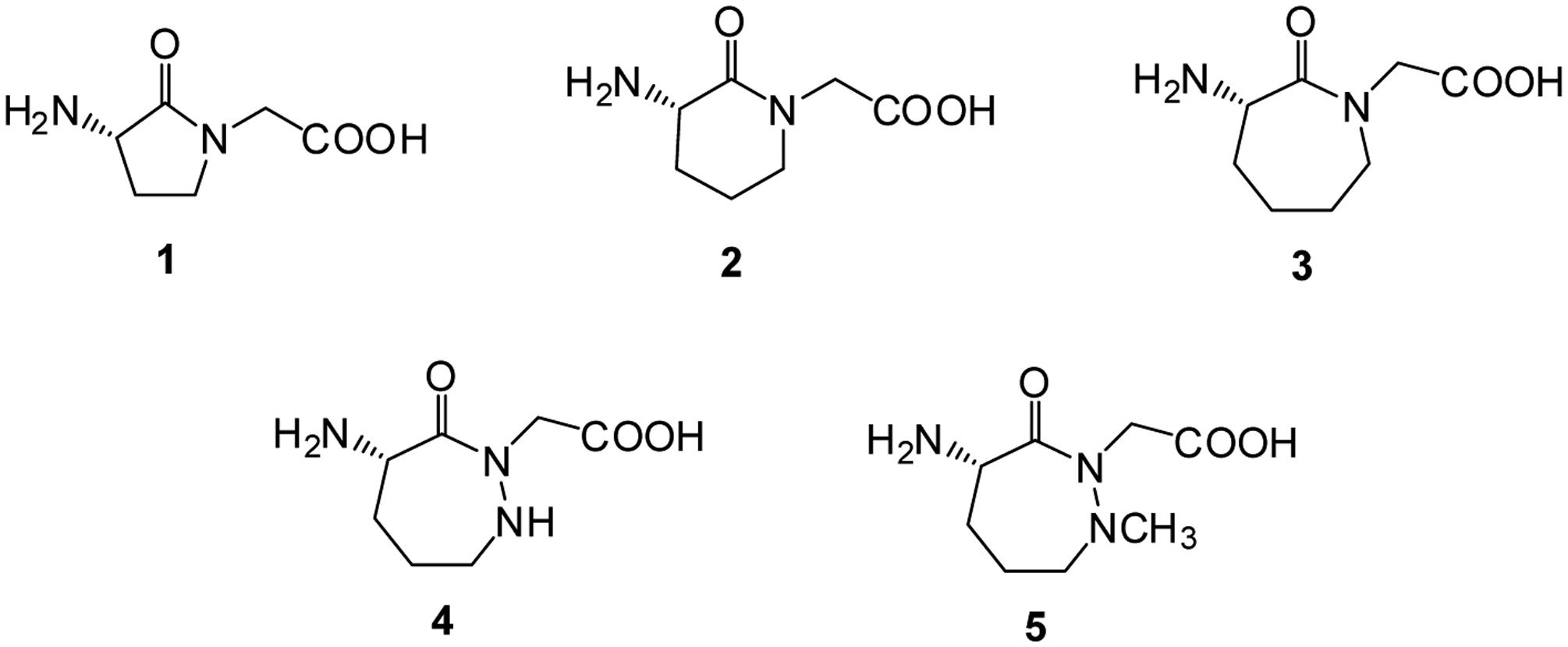
Structures of five conformationally constrained cyclic dipeptides synthesized and incorporated into E. coli dihydrofolate reductase at positions 18 and 49.
2. Results and discussion
Compounds 1 – 3 were prepared for potential use to induce conformational constraints in peptides in a report by Freidinger et al.29 Amblard et al.30 subsequently incorporated them in lieu of Pro7-Phe8 in bradykinin and found that the resulting bradykinin analogues bound to the human bradykinin B2 receptor. The preparation of dipeptide analogue 1, and the procedure used for its activation of a suppressor tRNACUA transcript obtained by in vitro transcription, is shown in Scheme 1. The routes used here for the preparation of compounds 1 – 3 were different from those reported by Freidinger et al.,29 and there were no identical compounds due to differences in protecting groups and the order of steps, so no comparison of properties can be made.
Scheme 1.
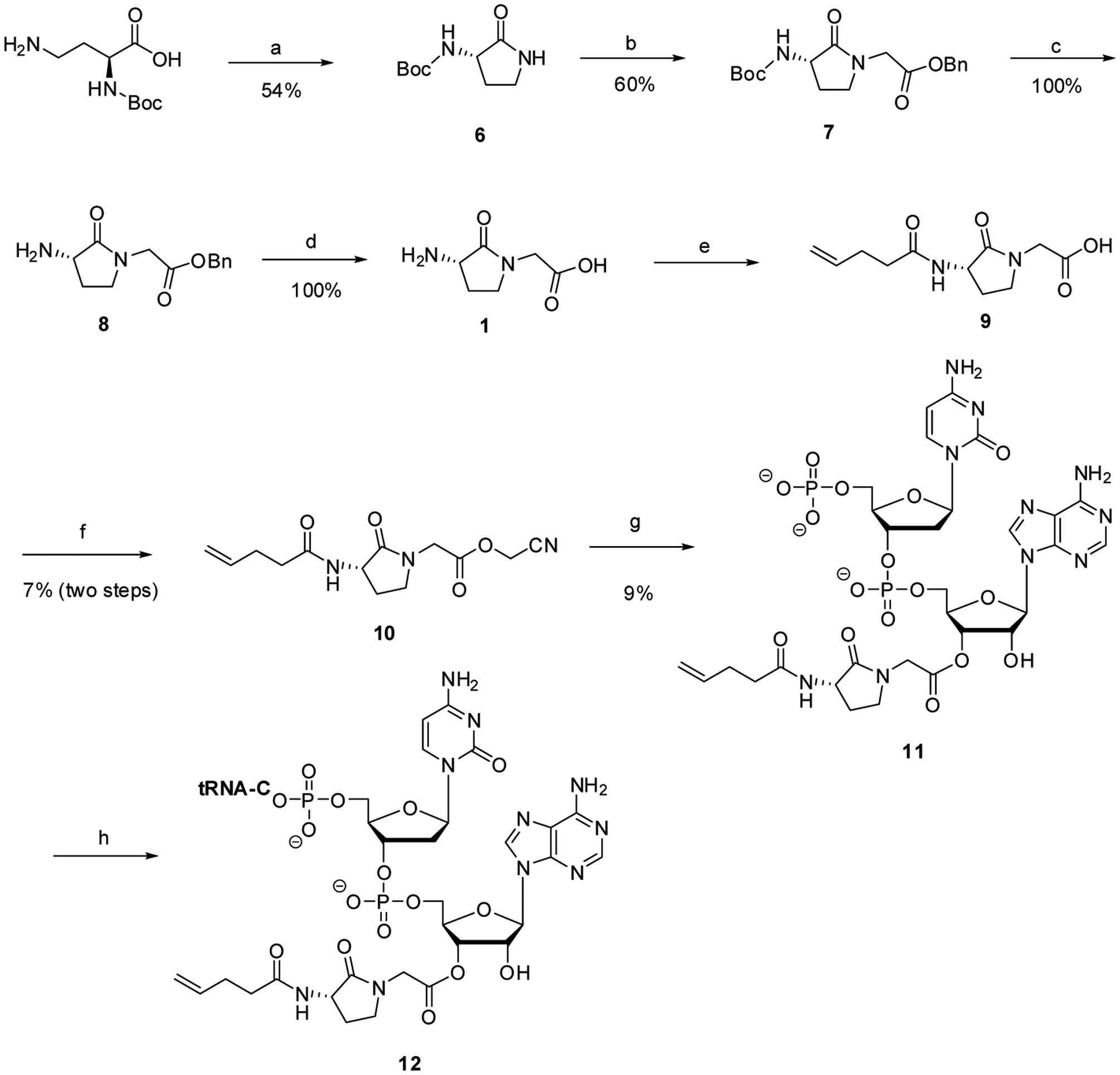
General procedures for the synthesis of a suppressor tRNACUA transcript activated with five-membered lactam-constrained dipeptide analogue 1. Reagents and conditions: (a) BOP reagent, NaHCO3,DMF, rt, 12 h; (b) NaH, benzyl bromoacetate, anh. THF, rt, 5 h; (c) 20% CF3COOH, anh. CH Cl o2 2, 0 C, 1 h; (d) 10% Pd/C, H2, 95% EtOH, rt, 1h; (e) 4-pentenoyloxysuccinimide, diisopropylethylamine, anh. DMF, rt, overnight; (f) chloroacetonitrile, Et3N, anh. DMF, rt, overnight; (g) pdCpA TBA salt, 9:1 anh. DMF-Et3N, sonication, rt, 12 h; (h) T4 RNA ligase, tRNA-COH, ATP, 37 °C, 2 h.
The synthesis of dipeptide 1 began with Nα-Boc (2S)-2,4-diaminobutyric acid, which was converted to lactam 6 in 54% yield by treatment with the BOP reagent ((benzotriazol-1-yloxy)tris(dimethylamino)phosphonium hexafluorophosphate). Treatment of lactam 6 with sodium hydride and benzyl bromoacetate in THF at room temperature afforded fully protected cyclic dipeptide 7 as a colorless oil in 60% yield after purification by silica gel chromatography. Following deprotection in quantitative yield by successive treatments with CF3COOH and hydrogenolysis over 10% palladium on carbon, free dipeptide 1 was isolated as its colorless trifluoroacetic salt. The dipeptide was reprotected on Nα of the pyrrolidine moiety as its N-pentenoyl derivative 9, and then activated as cyanomethyl ester 10 in low overall yield (7%) following purification by column chromatography on silica gel.
In order to enable tRNA activation, activated ester 10 was combined with the tetrabutylammonium salt of the dinucleotide pdCpA, which is a sequence common to the 3′-end of all cytoplasmic tRNAs. The resulting pdCpA ester of 10 (compound 11) was formed in low yield, and purified by C18 reversed phase HPLC, as were all of the pdCpA derivatives in this study. As shown in Scheme 1, this activated dinucleotide was attached to a suppressor tRNA in vitro transcript by enzyme mediated ligation with T4 RNA ligase.24 The N-pentenoyl protecting group attached to Nα of the pyrrolidine moiety of 12 could be removed prior to the use of the activated dipeptidyl tRNA in protein synthesis by brief treatment with dilute aqueous iodine.31,32
The synthesis of dipeptide 2 was carried out in a fashion closely analogous to the preparation of 1, except starting with Nα-Boc-protected ornithine rather than Nα-Boc 2,4-diaminobutyric acid (Schemes 1 and S1). The yields of the intermediates leading to the individual dipeptides 1 and 2 were also quite similar. Interestingly, while the two-step procedure leading to cyanomethyl ester 10 was very low, the analogous preparation of cyanomethyl ester 17 proceeded in 23% yield. Likewise, while pdCpA derivative 11 was formed in 9% yield (Scheme 1), the yield of 18 was 18% (Scheme S1). The synthesis of dipeptide 3 was also carried out as for 1 and 2, but starting with S-lysine. The yields of 3 were also quite similar to those realized in the steps leading to 1 and 2. For dipeptide 3, conversion to the fully protected cyanomethyl ester 24 proceeded in 21% yield and the pdCpA derivative was obtained in 38% yield (Scheme S2).
The synthesis of dipeptide 4 is outlined in Scheme 2. Ethyl (2S)-2-[(benzoxycarbonyl)(tert-butoxycarbonyl)amino]-5-oxopentanoate33 was converted to hydrazone 27 in 68% yield by treatment with tert-butyl carbazate, and then reduced to the corresponding hydrazine derivative 28 by treatment with NaBH3CN. Suitable positioning of protecting groups, and subsequent cyclization afforded a 1,2-diazepane with CBz protecting groups on N-1 and the 4-NH2 substituent (31). Treatment with ethyl bromoacetate then introduced the C-atoms of glycine at the 2-position of the diazepane. Following removal of the CBz protecting groups, the 4-NH2 substituent was protected by alkylation with 4-pentenoyloxysuccinimide, giving 34 in a yield of 62% for the two steps. Conversion of ethyl ester 34 to activated cyanomethyl ester 35 provided an intermediate capable of effecting esterification of dinucleotide pdCpA with the modified 1,2- diazepine, albeit in low yield. Dinucleotide 36 was ligated to the abbreviated tRNACUA-COH transcript, forming activated suppressor tRNA 37.
Scheme 2.
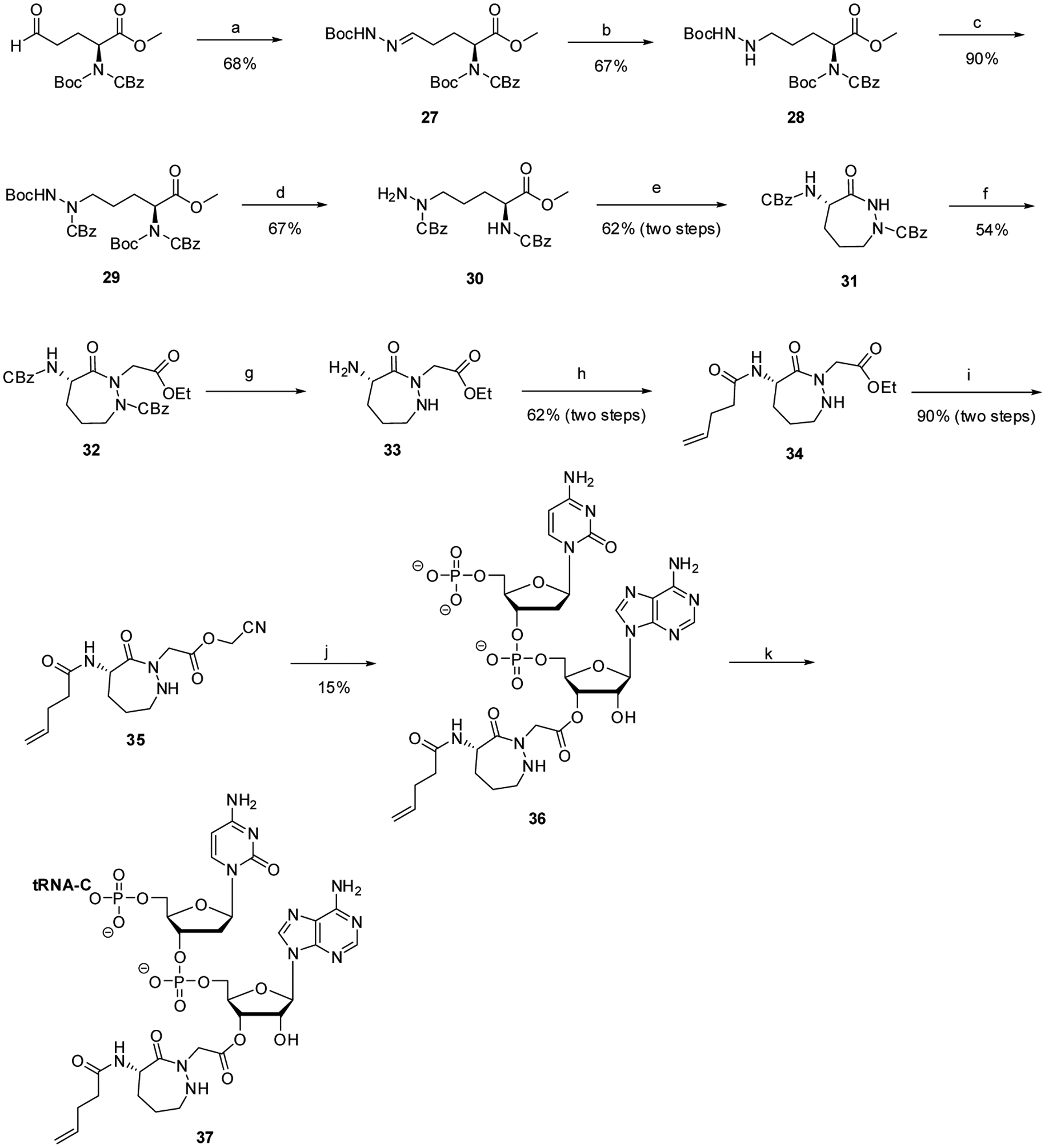
General procedures for the synthesis of a suppressor tRNACUA transcript activated with seven-membered cyclic acylhydrazide-constrained dipeptide analogue 4. Reagents and conditions: (a) tert-butyl carbazate, anh. THF, 0 °C, 18 h; (b) NaBH3CN, HOAc, EtOH, 0 °C, 17 h; (c) benzyl chloroformate, diisopropylethylamine, anh. CH2Cl2, rt, overnight; (d) 37.5% CF COOH, anh. CH o 3 2Cl2, 0 C, 3 h; (e) (1) 3:1 THF–MeOH, LiOH, 0 °C, 3 h; (2) HOBt, EDCI, diisopropylethylamine, CH3CN, rt, 18 h; (f) NaH, ethyl bromoacetate, anh. THF, rt, overnight; (g) 10% Pd/C, H2, 95% EtOH, rt, anh. MeOH, overnight; (h) 4-pentenoyl chloride, triethylamine, anh. CH2Cl2, rt, overnight; (i) (1) 3:1 THF–MeOH, LiOH, rt, 3 h; (2) chloroacetonitrile, Na2CO3, anh. DMF, rt, overnight; (j) pdCpA TBA salt, 9:1 anh. DMF–Et3N, rt, 15 h; (k) T4 RNA ligase, tRNA-COH, ATP, 37 °C, 2 h.
Methyl (2S)-2-[(enzoxycarbonyl)(tert-butoxycarbonyl)amino]-5-(tert butoxycarbonylhydrazinyl)pentanoate (28), an intermediate in the synthesis of dipeptide 4, was also used for the preparation of dipeptide 5 (Scheme 3). Following methylation of the hydrazine moiety in 76% yield, compound 38 was treated with CF3COOH to remove the Boc protecting groups and LiOH to hydrolyze the methyl ester. Cyclization by treatment with HOBt, diisopropylethylamine, and EDCI then effected cyclization, affording (4S)-4-(benzoxycarbonyl)amino-1-methyl--3-oxo-1,2-diazepane (40). Following treatment with ethyl bromoacetate to effect substitution at position 2 of the 1,2-diazepane in 92% yield, the CBz group at position 4 was replaced with an N-pentenoyl protecting group. Replacement of the ethyl ester at ring position 2 with a cyanomethyl ester then enabled esterification of the dinucleotide pdCpA with protected dipeptide 5.
Scheme 3.
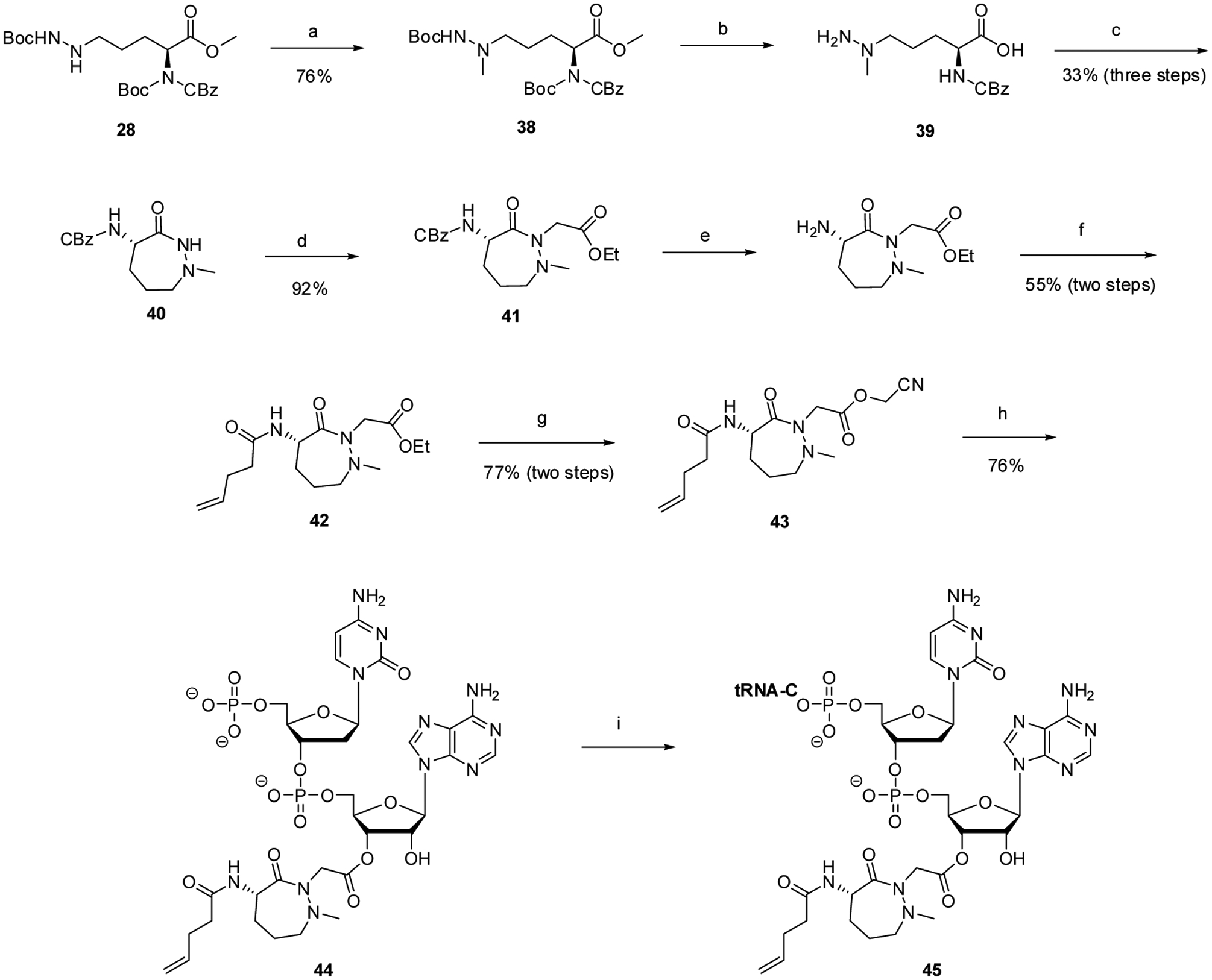
General procedures for the synthesis of a suppressor tRNACUA transcript activated with seven-membered cyclic acylhydrazide-constrained dipeptide analogue 5. Reagents and conditions: (a) CH3I, diisopropylethylamine, anh. DMF, rt, overnight; (b) (1) 50% CF COOH, anh. CH o3 2Cl2, 0 C, 3 h; (2) LiOH, 16:5:1 THF–MeOH–H2O, rt, 1 h; (c) 1:1 CH3CN–DMF, HOBt, diisopropylethylamine, EDCI, rt, 18 h, (d) NaH, anh. THF, ethyl bromoacetate, rt, overnight; (e) 10% Pd/C, anh. MeOH, rt, overnight; (f) anh. CH2Cl2, diisopropylethylamine, 4-pentenoyl chloride, rt, overnight; (g)(1) LiOH, 3:1 THF–MeOH;(2) anh. DMF, chloroacetonitrile, Na2CO3, rt, overnight; (h) pdCpA TBA salt, 9:1 anh. DMF–Et3N, rt, 15 h; (i) T4 RNA ligase, tRNA-COH, ATP, 37 °C, 2 h.
The cyclic peptidyl-pdCpA derivatives (compounds 11, 18, 25, 36 and 44) were all ligated to an abbreviated suppressor tRNACUA-COH prepared by in vitro transcription to afford the full length activated tRNAs by the use of T4 RNA ligase, as illustrated in Scheme 1 for the conversion of 11 → 12.31,32 The ligation of the cyclic dipeptidyl-pdCpAs to the abbreviated tRNAs was verified by analysis on 8% polyacrylamine–7 M urea gels at pH 5.0 (Figure 2).34 As noted in the Schemes, these tRNAs were protected with N-pentenoyl protecting groups, and were deprotected prior to their use in a protein synthesizing system by treatment with aqueous iodine at room temperature.31,32
Figure 2.
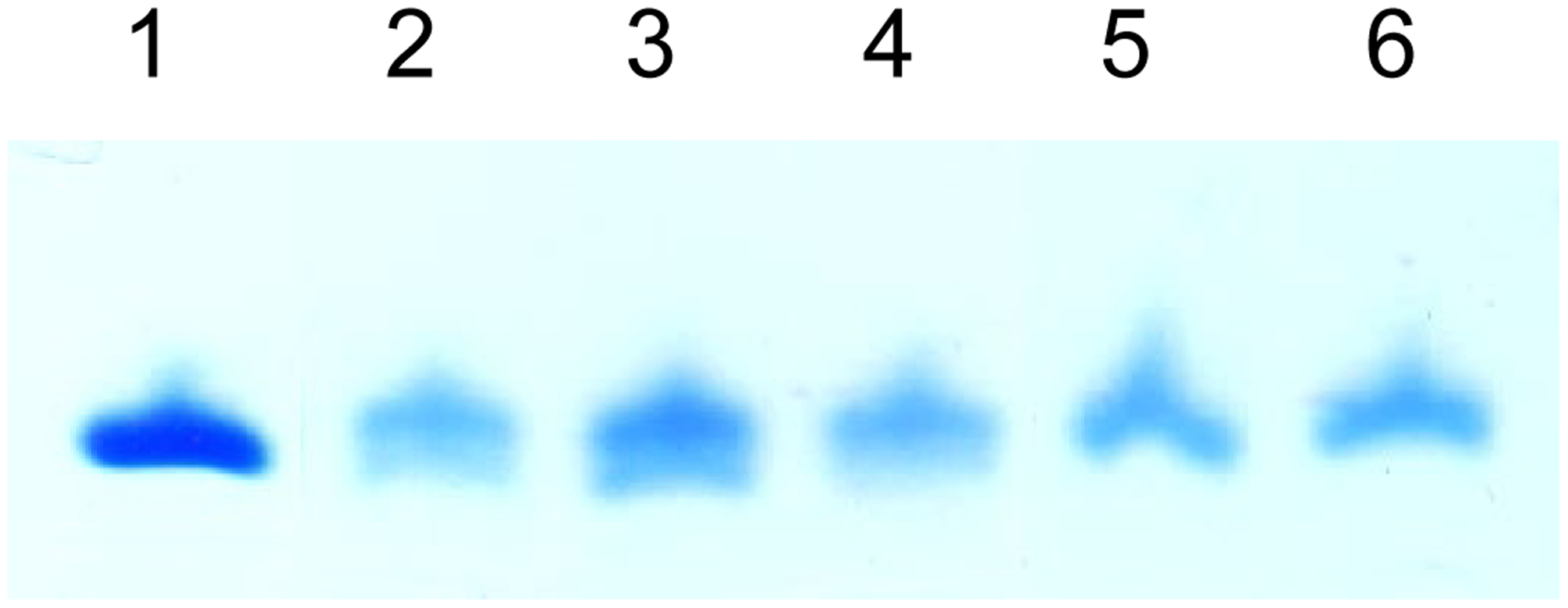
Analysis of the activation of a suppressor tRNA transcript with cyclic dipeptides by polyacrylamide gel electrophoresis under acidic conditions. Lane 1, abbreviated tRNACUA-COH (74 nucleotides). Lanes 2 – 6, ligation products of tRNACUA-COH with pdCpA derivatives 11, 18, 25, 36 and 44, respectively (76 nucleotides). The lower bands in lanes 2, 3 and 4 comigrated with the starting material (tRNA-COH).
Because E. coli ribosomes normally incorporate only α-L-amino acids, we have devised a strategy to alter the 23S ribosomal RNA at key positions, and to select modified ribosomes capable of recognizing activated tRNAs containing non-proteinogenic amino acids.24 These have included D-amino acids, β-amino acids, phosphorylated amino acids, dipeptides and dipeptidomimetics.35 In the present case, we utilized ribosomal clone 010328R2, originally selected to recognize a puromycin derivative that contained the dipeptide phenylalanylglycine.25 Because this clone was found to recognize several other dipeptides and dipeptidomimetic species, we used it in the present study to determine whether it would incorporate conformationally constrained dipeptides 1 – 5 into the model protein E. coli dihydrofolate reductase (DHFR).
Protein synthesis was carried out in triplicate independent experiments using an S-30 preparation with ribosomes from clone 010328R2. As shown in Figure 3, the five dipeptides were all incorporated, although the amounts varied from one analogue to another, and seemed to be incorporated somewhat more efficiently at position 18 than at position 49. The yields are summarized in Table 1. As shown in the Table, at both positions of DHFR, dipeptide analogue 3 (7-membered ring) was incorporated more efficiently than analogue 2 (six-membered ring), which was incorporated more efficiently than analogue 1 (five-membered ring). The suppression yields for 7-membered ring dipeptides 4 and 5 at position 18 (11.2 ± 0.9% and 10.2 ± 0.3%, respectively) were greater than for 7-membered ring 3 (8.9 ± 0.7%). The same relative efficiencies of incorporation were noted at position 49, although the differences between them was smaller (Table 1). Importantly, no significant incorporation of 1 – 5 was noted when an S-30 preparation made from wild-type ribosomes was employed (not shown). The choice of positions 18 and 49 in DHFR for replacement with non-proteinogenic amino acids was made to facilitate characterization as described.36 Mass spectrometric data supporting the incorporation of 4 and 5 into DHFR at position 49 is illustrated in Table S1.
Figure 3.
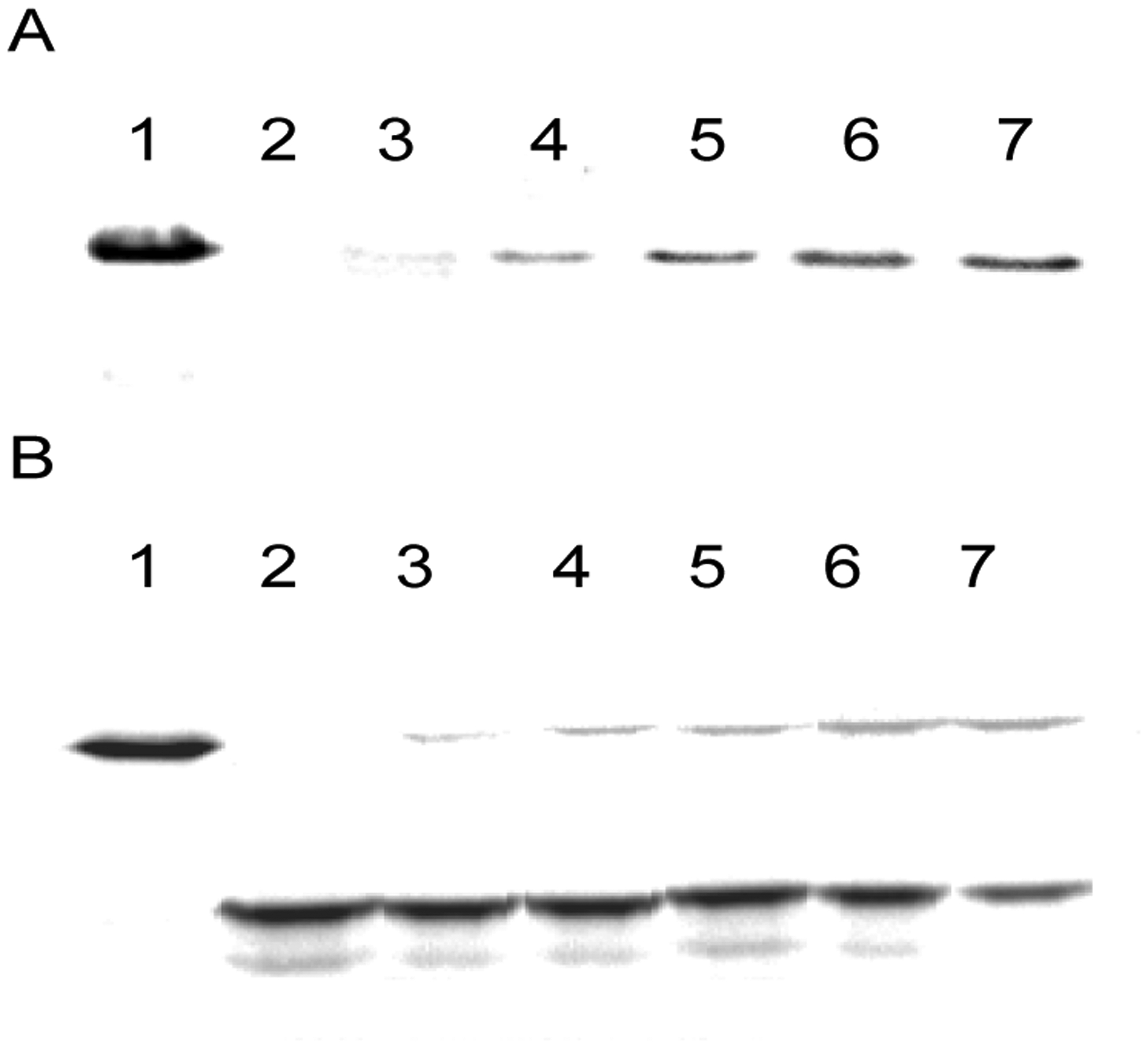
SDS‒polyacrylamide gel electrophoretic analysis of the synthesis of proteins containing cyclic peptides 1 – 5 at DHFR positions 18 (panel A) or 49 (panel B). (A) Comparison of wild-type DHFR (lane 1) and proteins synthesized using a mRNA with a UAG codon at position 18 in the absence of any suppressor tRNA (lane 2) or in the presence of tRNACUA activated with dipeptide 1 (lane 3), dipeptide 2 (lane 4), dipeptide 3 (lane 5), dipeptide 4 (lane 6) and dipeptide 5 (lane 7). (B) Comparison of wild-type DHFR (lane 1) and proteins synthesized using a mRNA with a UAG codon at position 49 in the absence of any suppressor tRNA (lane 2) or in the presence of tRNACUA activated with dipeptide 1 (lane 3), dipeptide 2 (lane 4), dipeptide 3 (lane 5), dipeptide 4 (lane 6) and dipeptide 5 (lane 7). In panel B, the stronger lower bands are DHFR truncated at position 49 (no suppression). In panel A, the shorter truncated protein ran off the gel.
Table 1.
Characterization of yield of full length DHFR during in vitro translation from modified genes in the presence of different cyclic dipeptidyl-tRNACUAs
| Cyclic dipeptides | Amount of full length proteins from modified genes, % wt | |
|---|---|---|
| TAG in position 18 | TAG in position 49 | |
| - | 0.6 ± 0.1 | 0.8 ± 0.2 |
| 1 | 4.2 ± 1.2 | 3.4 ± 0.8 |
| 2 | 6.3 ± 1.1 | 4.8 ± 0.7 |
| 3 | 8.9 ± 0.7 | 7.9 ± 2.5 |
| 4 | 11.2 ± 0.9 | 8.7 ± 0.8 |
| 5 | 10.2 ± 0.3 | 8.3 ± 1.2 |
The suppression yields in these experiments were less than what is sometimes observed for in vitro suppression using wild-type ribosomes. Nonetheless, earlier studies from our laboratory using DHFR37–39 have explored a range of substitutions enabling several biochemical studies to be carried out, and the yields reported here will clearly allow such cyclic dipeptides to be used in support of mechanistically focused mutagenesis experiments. One further issue that needs to be addressed in future experiments is whether conformational constraint, which has been exceptionally useful in producing peptides with improved properties, can also be exploited productively for protein modifications. In this regard, it may be noted that nature already employs proline to control conformation. This is accomplished both through judicious placement of proline residues at protein sites that can benefit from conformational constraint,40 and from cis-trans isomerization of the polypeptide backbone to effect reversible control of protein conformation.41 It is anticipated that the conformationally constrained dipeptides described here may find utility though the constraint of protein structure.
3. Conclusions
Five cyclic dipeptides have been prepared to enable the study of conformational constraint in proteins, a type of modification that has proven very useful in peptides. The modifications have included lactam-derived dipeptides having five-, six and seven-membered rings (1 – 3), and seven-membered acylhydrazides (diazepanes) 4 and 5. Each of these synthetic dipeptides was activated as a pdCpA ester and used to prepare a suppressor tRNA by the use of T4 RNA ligase. The activated cyclic dipeptidyl-tRNAs were used to introduce the five dipeptides into positions 18 and 49 of E. coli dihydrofolate reductase. The suppression yields obtained in replicate experiments were entirely sufficient to provide the amounts of protein required for routine biochemical experiments.
4. Experimental section
All chemical and biochemical methods are described in detail in the Supplementary Material sections.
Supplementary Material
Funding
This study was supported by National Institute of General Medical Sciences Grant GM12367, National Institutes of Health.
Footnotes
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- 1.Wu Q, Brow MR. Signaling and function of insulin-like peptides in insects. Annu Rev Entomol. 2006;51:1–24. [DOI] [PubMed] [Google Scholar]
- 2.Mack C, Wilson J, Athanacio J, Reynolds J, Langero K, Guss s, Vu C, Roth J, Parkes D. Pharmacological actions of the peptide hormone amylin in the long-term regulation of food intake, food preference, and body weight. Am J Physiol Regul Integr Comp Physiol. 2007; 293:R1855–R1863. [DOI] [PubMed] [Google Scholar]
- 3.Holst JJ. The physiology of glucagon-like peptide 1. Physiol Rev. 2007;87;1409–1439. [DOI] [PubMed] [Google Scholar]
- 4.Drucker DJ. The role of gut hormones in glucose homeostasis. J Clin Invest. 2007;117:24–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Clapp C, Thebault S, Jeziorski MC, Martinez de la Escalera G. Peptide hormone regulation of angiogenesis. Physiol Rev. 2009;80:1177–1215. [DOI] [PubMed] [Google Scholar]
- 6.Stymiest JL, Mitchell BF, Wong S, Vederas JC. Synthesis of biologically active dicarba analogues of the peptide hormone oxytocin using ring-closing metathesis Org Lett. 2003;5:47–49. [DOI] [PubMed] [Google Scholar]
- 7.Holz GG, Chepurny OG. Glucagon-Like Peptide-1 Synthetic analogs: new therapeutic agents for use in the treatment of diabetes mellitus Current Med Chem. 2003;10:2471–2483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Goodwin D, Simerska P, Toth L. Peptides as therapeutics with enhanced bioactivity Current Med Chem. 2003;19:4451–4461. [DOI] [PubMed] [Google Scholar]
- 9.Smith III AB, Wang W, Sprengler PA, Hirschmann R. Design, synthesis and solution structure of a pyrrolinone-based b-turn peptidomimetic. J Am Chem Soc. 2000;122:11037–11038. [Google Scholar]
- 10.Hirschmann RF, Nicolaou KC, Angeles AR, Chen JS, Smith AB III. The β-D-glucose scaffold as a β-turn mimetic. Acc Chem Res. 2009;42:1511–1520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Balaev AN, Osipov VN, Khachatryan DS. Synthetic peptide analogs of somatostatin: trends in the synthesis of and prospects in the search for new anticancer drugs. Pharmaceut Chem J. 2015;49:345–351. [Google Scholar]
- 12.Henchley LK, Jochim AL, Arora PS. Contemporary strategies for the stabilization of Peptides in the α-helical conformation. Current Opin Chem Biol. 2008;12:692–697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Estieu-Gionnet K, Guichard G. Stabilized helical peptides: overview of the technologies and therapeutic promises. Exp Opin Drug Discovery. 2011;6:937–963. [DOI] [PubMed] [Google Scholar]
- 14.Diderich P, Bertoldo D, Dessen P, Khan MM, Pizzitola I, Held W, Huelsken J, Heinis C. Phage selection of chemically stabilized α-helical peptide ligands. ACS Chem Biol. 2016;11:1422–1427. [DOI] [PubMed] [Google Scholar]
- 15.Haney EF, Hunter HN, Matsuzaki K, Vogel HJ. Solution NMR studies of amphibian antimicrobial peptides:linking structure to function? Biochim Biophys Acta. 2009;1788:1639–1655. [DOI] [PubMed] [Google Scholar]
- 16.Pierschbacher MD, Ruoslahti E. Cell attachment activity of fibronection can be duplicated by small synthetic fragments of the molecule. Nature. 1984;309:30–33. [DOI] [PubMed] [Google Scholar]
- 17.Dechantsreiter MA, Planker E, Matha B, Lohof E, Hölzemann G, Jonczyk A, Goodman SL, Kessler H. N-Methylated cyclic RGD peptides as highly active and selective αvβ3 integrin antagonists. J Med Chem. 1999;42:3033–3040. [DOI] [PubMed] [Google Scholar]
- 18.Belvisi L, Bernardi A, Checchia A, Manzoni L, Potenza D, Scolastico C, Castorina M, Cupelli A, Giannini G, Carminati P, Pisano C. Potent integrin antagonists from a small library of RGD-including cyclic pseudopeptides. Org Lett. 2001;3:1001.–. [PubMed] [Google Scholar]
- 19.Roberts RW, Szostak JW. RNA-peptide fusions for the in vitro selection of peptides and proteins Proc Natl Acad Sci USA. 1997;94:12297–12302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ishizawa T, Kawakami T, Reid PC, Murakami H. J Am Chem Soc. 2013;135:5433.–. [DOI] [PubMed] [Google Scholar]
- 21.Maini R, Kimura H, Takatsuji R, Katoh T, Goto Y, Suga H. Ribosomal formation of thioamide bonds in polypeptide synthesis. J Am Chem Soc. 2019;141:20004.–. [DOI] [PubMed] [Google Scholar]
- 22.Kazlauskas R Engineering more stable proteins. Chem Soc Rev. 2018;47:9026–9045. [DOI] [PubMed] [Google Scholar]
- 23.Li JC, Liu T, Wang Y, Mehta AP, Schultz PG. Enhancing protein stability with genetically encoded noncanonical amino acids. J Am Chem Soc. 2018;140:15997–16000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dedkova LM, Fahmi NE, Paul R, del Rosario M, Zhang L, Chen S, Feder G, Hecht SM. β-Puromycin selection of modified ribosomes for in vitro incorporation of β-amino Acids. Biochemistry. 2012;51:401–415. [DOI] [PubMed] [Google Scholar]
- 25.Maini R, Dedkova LM, Paul R, Madathil MM, Roy Chowdhury S, Chen S, Hecht SM. Ribosome-mediated incorporation of dipeptides and dipeptide analogues into proteins in vitro. J Am Chem Soc. 2015;137:11206–11209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Roy Chowdhury S, Maini R, Dedkova LM, Hecht SM. Synthesis of fluorescent dipeptidomimetics and their ribosomal incorporation into green fluorescent protein, Bioorg Med Chem Lett. 2015;25:4715–4718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Roy Chowdhury S, Chauhan PS, Dedkova LM, Bai X, Chen S, Talukder P, Hecht SM.Synthesis and evaluation of a library of fluorescent dipeptidomimetic analogues as substrates for modified bacterial ribosomes. Biochemistry. 2016;55:2427–2440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chen S, Ji X, Gao M, Dedkova LM, Hecht SM. In cellulo synthesis of proteins containing a fluorescent oxazole amino acid. J Am Chem Soc. 2019;141:5597–5601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Freidinger RM, Perlow DS, Veber DF. Protected lactam-bridged dipeptides for use as conformational constraints in peptides. J Org Chem. 1982;47:104–109. [Google Scholar]
- 30.Amblard M, Daffix I, Berge G, Calmes M, Dodey P, Pruneau D, Paquet J, Luccarini J, Belichard P, Martinez J. Synthesis and characterization of bradykinin B2 receptor agonists containing constrained dipeptide mimics. J Med Chem. 1999;42:4193–4201. [DOI] [PubMed] [Google Scholar]
- 31.Lodder M, Golovine S, Hecht SM. Chemical deprotection strategy for the elaboration of misacylated transfer RNA’s. J. Org. Chem 1997;62:778–779. [Google Scholar]
- 32.Lodder M, Golovine S, Laikhter AL, Karginov VA, Hecht SM. Misacylated transfer RNAs having a chemically removable protecting group. J. Org. Chem 1998;63:794–803 [DOI] [PubMed] [Google Scholar]
- 33.Nasopoulou M, Georgiadis D, Matziari M, Dive V, Yiotakis A. A versatile annulation protocol toward novel constrained phosphinic peptidomimetics. J Org Chem. 2007;72:7222–7228. [DOI] [PubMed] [Google Scholar]
- 34.Varshney U, Lee CP, RajBhandary UL. Direct analysis of aminoacylation levels of tRNAs in vivo. Application to studying recognition of Escherichia coli initiator tRNA mutants by glutaminyl-tRNA synthetase. J Biol Chem. 1991;266:24712–24718. [PubMed] [Google Scholar]
- 35.Dedkova LM, Hecht SM. Expanding the scope of protein synthesis using modified ribosomes. J Am Chem Soc. 2019;141:6430–6447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Maini R, Nguyen DT, Chen S, Dedkova LM, Roy Chowdhury S, Alcala-Torano R, Hecht SM. Incorporation of β-amino acids into dihydrofolate reductase by ribosomes having modifications in the peptidyltransferase center. Bioorg Med Chem. 2013;21:1088–1096. [DOI] [PubMed] [Google Scholar]
- 37.Karginov VA, Mamaev SV, An H, Van Cleve MD, Hecht SM, Komatsoulis GA, Abelson JN. Probing the role of an active site aspartic acid in dihydrofolate reductase. J Am Chem Soc. 1997;119:8166–8176. [Google Scholar]
- 38.Eisenhauer BM, Hecht SM. Site-specific incorporation of (aminoxy)acetic acid into proteins. Biochemistry 2002;41:11472–11478. [DOI] [PubMed] [Google Scholar]
- 39.Fahmi NE, Dedkova L, Wang B, Golovine S, Hecht, SM. Site specific incorporation of glycosylated serine and tyrosine derivatives into proteins. J Am Chem Soc. 2007;129:3586–3597. [DOI] [PubMed] [Google Scholar]
- 40.Vanhoof G, Goossens F, De Meester I, Scharpé S. Proline motifs in peptides and their biological processing. FASEB J. 1995;9:736–744. [PubMed] [Google Scholar]
- 41.Taler-Vercic A, Hasanbasic S, Berbic S, Stoka V, Turk D, Zerovnik E. Proline residues as switches in conformational changes leading to amyloid fibril formation. Int J Mol Sci. 2017;18:549. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.