

Abstract
Breast cancer is the most common type of cancer among women worldwide. The Hippo signaling pathway is strongly associated with cell proliferation, migration, invasion, metastasis and resistance to breast cancer treatment. The upstream factors involved in the Hippo signaling pathway, including mammalian Ste20 kinases 1/2, large tumor suppressor kinases 1/2 and transcription coactivator Yes-associated protein (YAP)/transcriptional coactivator with PDZ-binding motif (TAZ), have been extensively studied as they are considered therapeutic targets for breast cancer. Recently, it has been suggested that the transcriptional enhancer factor domain (TEAD) family of transcription factors, particularly TEAD4, plays an important role in breast cancer. TEADs interact with YAP/TAZ to act as transcription factors. Notably, recent studies have demonstrated that TEAD4 may also function in a YAP/TAZ-independent manner and serve as a prognostic marker for breast cancer. The present review summarizes the current research on the effect of the aberrant activation of the Hippo signaling pathway on breast cancer progression. Furthermore, the latest advances on the role of the TEAD family in breast cancer are highlighted, and the role of TEAD4 as a potential target for therapeutic intervention in breast cancer is discussed.
Keywords: Hippo signaling pathway, breast cancer, transcriptional enhancer factor domain 4
1. Introduction
In women, breast cancer is the most diagnosed cancer (24.5% of total cases), followed by colorectal (9.4%) and lung (8.4%) cancers (1). Breast cancer is also the leading cause of cancer-associated mortality (15.5% of total cancer-associated mortalities) among females, followed by lung (13.7%) and colorectal (9.5%) cancers (1). Currently, chemotherapy and surgery are the main treatment approaches for breast cancer. However, the recurrence and chemoresistance of breast cancer are obstacles to its treatment, and the molecular mechanism underlying these phenomena remain unclear (2). Using a single biomarker is insufficient in adequately assessing tumor growth, thus, identifying the molecular mechanisms underlying tumor growth and cell proliferation, through multiple signaling pathways consisting of a set of core regulatory elements, may be more effective.
The Hippo signaling pathway, a novel and evolutionarily conserved tumor suppression pathway first discovered in Drosophila, regulates tissue growth and organ size by attenuating cell proliferation and simultaneously promoting cell death (3). In humans, the core Hippo pathway consists of the scaffold protein merlin, encoded by the neurofibromatosis type 2 (NF2) gene, KIBRA protein, WW and the C2 domain containing 1 (WWC1) gene and a cascade of kinases, including the upstream sterile 20-like kinase 1 and 2 (MST1/2), the cofactor Salvador homologue 1 (SAV1), MOB kinase activator 1A and B (MOB1A/B), and the downstream large tumor suppressors 1 and 2 (LATS1/2). In response to high cell density, NF2 and WWC1 activate the Hippo signaling pathway by binding to LATS1/2 to facilitate their activation by MST1/2 (4,5). Subsequently, MST1/2 binds to SAV1 to form an enzymatic complex that phosphorylates and activates LATS1/2 kinases and their MOB1A/B regulatory subunits. LATS1/2 phosphorylates the transcriptionalcoactivator Yes-associated protein (YAP) and WW domain-containing transcription regulator protein 1, also known as transcriptional coactivator with PDZ-binding motif (TAZ), which in turn translocate in the cytoplasm and are inactivated (6). In cancer, the Hippo signaling pathway is abnormally inactivated, thus, LATS1/2 is not activated. Following nuclear translocation, the YAP/TAZ complex binds to the transcriptional enhancer factor domain (TEAD)/TEF to activate the transcription of its target genes, including connective tissue growth factor (CTGF), cysteine-rich angiogenic inducer 61 (CYR61), glioma-associated oncogene family zinc finger 2 (Gli2) and amphiregulin (AREG), which in turn promote tumor cell proliferation and growth (5,7) (Fig. 1). Previous studies have demonstrated that the YAP/TAZ complex is often abnormally expressed in breast cancer, and its overactivation eventually leads to several tumor-promoting processes, such as epithelial-to-mesenchymal transition(EMT), cancer stem cell generation and chemotherapy resistance (7,8).
Figure 1.
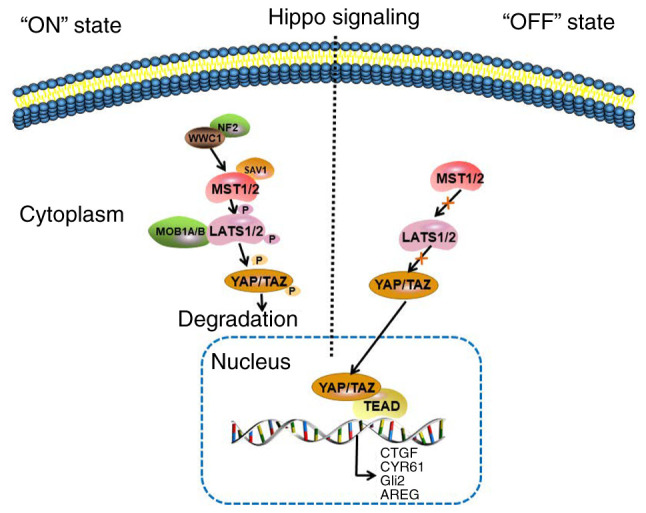
Hippo signaling pathway. In response to high cell density, NF2 and WWC1 activate the Hippo signaling pathway by binding to LATS1/2 to facilitate their activation by MST1/2. In turn, MST1/2 bind to the cofactor, SAV1, to form an enzymatic complex that phosphorylates and activates the LATS1/2 kinases, and the MOB1A/B regulatory subunits of LAST1/2. Subsequently, LATS1/2 phosphorylate the transcriptional coactivators, YAP and TAZ, which in turn translocate in the cytoplasm and get inactivated. In cancer, the Hippo signaling is abnormally inactivated and LATS1/2 are not activated by MST1/2. The YAP/TAZ complex translocates to the nucleus and binds with TEAD to activate the transcription of its target genes, including CTGF, CYR61, Gli2 and AREG. NF2, neurofibromatosis type 2; WWC1, WW And C2 domain containing 1 gene; LATS1/2, large tumor suppressors 1 and 2; MST1/2, upstream sterile 20-like kinase 1 and 2; SAV1, Salvador homologue 1; MOB1A/B, MOB kinase activator 1A and B; YAP, Yes-associated protein; TAZ, transcriptional coactivator with PDZ-binding motif; TEAD, transcriptional enhancer factor domain; CTGF, connective tissue growth factor; CYR61, cysteine-rich angiogenic inducer 61; Gli2, glioma-associated oncogene family zinc finger 2; AREG, amphiregulin.
The present review summarizes the current research on the key targets of the Hippo signaling pathway in the treatment of breast cancer. Furthermore, the latest advances on the role of the TEAD family in breast cancer are highlighted, and the role of TEAD4 as a potential target for therapeutic intervention in breast cancer is discussed.
2. Aberrant expression of various components of the Hippo signaling pathway in breast cancer
Increasing evidence suggests that the expression of the upstream kinases involved in the Hippo signaling pathway is consistent in different subtypes of breast cancer. For example, MST1 and MST2 expression levels are downregulated in human breast cancer tissues compared with normal breast tissues (9). In addition, upregulated MST1 expression is associated with high overall survival and disease-free survival rates, suggesting that MST1 functions as a tumor suppressor in breast cancer (10). Phosphorylation of MST1/2 (p-MST1/2) promotes the proliferation and migration of MDA-MB-231 cells (11), while the nuclear localization of p-MST1/2 exhibits tumor suppressive effects in human epidermal growth factor receptor 2-positive and triple-negative breast cancer (TNBC) (12). The loss of LATS2 is associated with embryonic lethality in mouse embryos, and it was hypothesized that the embryonic lethality was caused by abnormal cell proliferation, mitotic defects and accumulated genomic instability; however, LATS1-null mice survive. LATS1 and LATS2 are considered key regulators of cell fate as they modulate the functions of tumor suppressive effectors, which exert a highly conserved function during evolution (13).
In addition to the close association between the kinase chain of the Hippo signaling pathway and breast cancer progression, the transcription coactivators, YAP and TAZ, serve important roles in the occurrence and development of breast cancer (8,14,15). The activity of the YAP/TAZ complex is associated with high histological grade and breast cancer stem cell traits, and can induce multiple processes involved in tumor progression and metastasis of breast cancer (14–16). In luminal type A breast cancer, low YAP1 expression is associated with a low recurrence-free survival rate (14); however, this expression trend is reversed in TNBC (15). In addition, the protein expression levels and activity of TAZ are elevated in prospective breast cancer stem cells and in poorly differentiated human tumors (16). Consistently, overexpression of TAZ in differentiated breast cancer cells can induce cell transformation and confer tumorigenicity and migratory ability (17). Nuclear TAZ expression is strongly associated with the TNBC subtype, which is further strengthened in the basal-like subtype. Furthermore, 90% of breast carcinomas with morphological EMT features are TAZ-positive (18). The downstream effectors of TEAD transcription factors include the oncogenes, CYR61, CTGF, MYC, AREG and Gli2 (19–23).
3. Upstream kinases as therapeutic targets for breast cancer
Gene and drug therapies can inhibit the development of breast cancer by affecting the expression of the MST1/2 downstream kinases. It has been reported that Ras association domain-containing protein 6 (RASSF6), discs large homolog 5 (DLG5), microtubule affinity regulating kinase 4 (MARK4) and meningioma-associated protein (MAC30)act on the Hippo signaling pathway MST1/2 upstream kinases (24–26). Previous studies have demonstrated that RASSF6 (24) and DLG5 (25) can promote the phosphorylation of MST1/2 and LATS1, and inhibit YAP expression. Conversely, MARK4 can bind and phosphorylate MST and SAV, and attenuate the formation of the MST-SAV and LATS complexes in MDA-MB-231 cells (11). In addition, MAC30 inhibits MST1/2 and LATS1/2 expression levels in MDA-MB-157 and MDA-MB-231 cells (26). It has also been demonstrated that neoadjuvant therapy activates the nuclear localization of p-MST1/2, resulting in tumor suppressive effects (12).
LATS1/2 are considered important kinases involved in the Hippo signaling pathway, thus, drug and gene therapies targeting LATS1/2 have been extensively studied (27–29). Phospholipase D (PLD), phosphatidic acid (PA) and vascular endothelial growth factor (VEGF) can regulate the activity of LATS1/2. PLD activates PA, which in turn directly interacts with LATS to disrupt the formation of the LATS-MOB1 complex, thereby promoting the carcinogenic activity of YAP (27). Another study demonstrated that VEGF-NRP2 signaling can activate GTPase Rac1, which inhibits the Hippo kinase, LATS, and activates TAZ and stem-like traits in MDA-MB-231 cells (28). Conversely, resveratrol phosphorylates and inactivates YAP by activating LATS1 in TNBC cells (29).
Kinases are regulated and activated by various genes, and are also degraded in cells. Protein kinases are degraded by E3 ubiquitin ligase and are deubiquitinated by deubiquitinating enzymes to maintain protein stability (30–34). A study revealed that breast cancer type 1 susceptibility protein could turn ‘off’ the Hippo signaling pathway via ubiquitination of NF2 (30). Furthermore, NF2 can inhibit the interaction between the E3 ubiquitin ligase, CRL4DCAF1 and LATS1; however, when derepressed, CRL4DCAF1 can ubiquitylate and block LATS1/2 in the cell nucleus (31). Similarly, it has been reported that Itch, a HECT class E3 ubiquitin ligase, can form a complex with LATS1 in MDA-MB-231 cells through the PPxY motifs of LATS1 and the WW domains of Itch (32). In addition, a study demonstrated that E3 ubiquitin-protein ligase seven in absentia homolog 2 destabilizes LATS2 in response to hypoxia (33). It has also been reported that deubiquitinase ubiquitin specific peptidase 9 X-linked strongly interacts with LATS kinase and enhances its stability (34).
4. Transcriptional coactivators YAP/TAZ as therapeutic targets for breast cancer
Several studies have focused on the phosphorylation of YAP/TAZ, and the interaction between YAP/TAZ and TEAD to activate transcription of their target genes. It has been demonstrated that leukemia inhibitory factor receptor can promote YAP phosphorylation and its cytoplasmic retention, eventually resulting in YAP inactivation (35). Another study reported that the loss of FAT atypical cadherin 1 promotes the resistance of estrogen receptor (ER)-positive breast cancer to cyclin-dependent kinase (CDK) 4/6 inhibitors through the accumulation of YAP/TAZ transcription factors on the CDK6 promoter (36). Similarly, the loss of Ca2+-ATPase isoform 2 (37) and extracellular regulated protein kinase 1 (38) can activate YAP/TAZ expression. Τhe concept that regular exercise can prevent breast cancer is based on scientific evidence suggesting that the serum levels of catecholamines, including epinephrine and norepinephrine, increase following exercise, thus resulting in YAP1 phosphorylation and its cytoplasmic retention, eventually promoting its tumor suppressive effects (39).
The ubiquitination and deubiquitination of YAP/TAZ can also affect breast cancer development (40,41). A study demonstrated that YAP1 is polyubiquitinated and degraded by the E3 ubiquitin-protein ligase, RNF187, thus acquiring tumor suppressive effects in TNBC (40). In addition to ubiquitination and deubiquitination, SUMOylation is also considered a common modified degradation method. The ubiquitin thioesterase protein, OTUB2, was demonstrated to be poly-SUMOlated on K233, thus allowing its binding with the YAP/TAZ complex, resulting in the deubiquitination and activation of the complex (41) (Table I).
Table I.
Gene and drug therapies targeting the Hippo signaling pathway.
| Gene and drug therapies | Hippo target | Mechanisms | (Refs.) |
|---|---|---|---|
| RASSF6, DLG5 | MST1/2 and LATS1 | Promotes the phosphorylation of MST1/2 and LATS1 | (24,25) |
| MARK4 | MST and SAV | Weakens the formation of the MST-SAV and LATS complex | (11) |
| MAC30 | MST1/2 and LATS1/2 | Attenuates MST1/2 and LATS1/2 levels | (26) |
| Neoadjuvant therapy | MST1/2 | Activates the nuclear localization of pMST1/2 | (12) |
| VEGF | LATS1/2 | Activates GTPase Rac1, thereby inhibiting LATS | (28) |
| Resveratrol | LATS1 | Activates LATS1 | (29) |
| BRCA1 | NF2 | Contributes to the inactivation of the Hippo signaling pathway through NF2 ubiquitination | (30) |
| CRL4DCAF1, Itch | LATS1/2 | Ubiquitylates and inhibits LATS1/2 | (31) |
| SIAH2 | LATS1/2 | Inactivates LATS1/2 via proteases | (33) |
| USP9X | LATS | Interacts with LATS kinase and enhances the stability of LATS | (34) |
| LIFR | YAP | Promotes YAP phosphorylation and cytoplasmic retention | (35) |
| FAT1 | YAP/TAZ | The loss of FAT1 promotes YAP/TAZ expression | (36) |
| SPCA2, ERK1 | YAP/TAZ | Activates YAP/TAZ expression | (37,38) |
| EPI and NE | YAP1 | Through YAP1 phosphorylation and cytoplasmic retention | (39) |
| RNF187 | YAP1 | Polyubiquitinates and degrades YAP1 | (40) |
| OTUB2 | YAP/TAZ | Deubiquitinates and activates YAP/TAZ | (41) |
RASSF6, Ras association domain-containing protein 6; DLG5, discs large homolog 5; MARK4, microtubule affinity regulating Kinase 4; MAC30, meningioma-associated protein; VEGF, vascular endothelial growth factor; BRCA1, breast cancer type 1 susceptibility protein; LIFR, leukemia inhibitory factor receptor; FAT1, FAT atypical cadherin 1; SPCA2, Ca2+-ATPase isoform 2; ERK1, extracellular regulated protein kinase 1; EPI, epinephrine; NE, norepinephrine; MST1/2, upstream sterile 20-like kinase 1 and 2; LATS1/2, large tumor suppressors 1 and 2; YAP, Yes-associated protein; TAZ, transcriptional coactivator with PDZ-binding motif; SAV, Salvador homologue; NF2, neurofibromatosis type 2.
5. TEAD4 as a promising target for breast cancer treatment
Although several studies have focused on investigating the therapeutic potential of the regulation of YAP phosphorylation and its nuclear localization (35,39), the regulation and function of the TEAD family have not been extensively studied. TEAD is an important transcription factor that binds to YAP/TAZ in the nucleus. The mammalian encoding TEAD family consists of four homologous genes, namely TEAD1-4. Following binding of YAP to TEAD, it depends on the DNA binding domain of TEAD to initiate transcription of the downstream target genes (5,7,8). It has been demonstrated that the oncogenes, interleukin 6 (42), angiomotin (43) and activator protein-1 (44) can promote the formation of the YAP/TAZ-TEAD complex, and facilitate the proliferation and invasion of breast cancer cells. Furthermore, the YAP/TAZ-TEAD complex increases programmed cell death 1 ligand 1 promoter activity to enhance the cell invasive ability and immune evasion in breast cancer (45). Notably, it has been reported that Ski activates the non-classical Hippo signaling pathway by binding to the TAZ-TEAD complex and recruiting the transcriptional repressor nuclear receptor corepressor 1 to the TAZ-TEAD complex to inhibit TAZ transcription (46).
Previous studies have demonstrated that the TEAD family, particularly TEAD4, is upregulated in various tumor tissues, including gastric, colon, breast and prostate cancers, and is associated with poor prognosis (47–49). TEAD1 and TEAD4 play crucial roles in cancer by promoting tumor growth (50–53). A study revealed that TEAD4 expression is upregulated in breast cancer, thus serving as a prognostic marker for breast cancer (50). In MCF-7 cells, the YAP1-TEAD4 complex binds to the enhancer region of ERα to activate the expression of the enhancer RNA transcription markers by recruiting the mediator complex subunit 1, a component of the enhancer activation mechanism. ERα is subsequently activated by binding to the estrogen ligand 17b-estradiol (E2), thereby regulating CTGF expression (51). Notably, complementation experiments using wild-type TEAD4 and YAP-binding mutant TEAD4, Y429H, demonstrated that both factors can reverse EMT, increase the expression levels of the mesenchymal marker vimentin, and enhance the cell invasive and migratory abilities in a YAP-independent manner (52,53). Thus, nuclear TEAD4 expression is considered a biomarker for colon cancer progression and poor prognosis (52,53). It has also been reported that the activation of glucocorticoids promotes the interaction between glucocorticoid receptor and TEAD4, eventually resulting in the formation of a complex. This complex is subsequently recruited to the TEAD4 promoter to enhance its own expression and promote breast cancer stem cell-like properties (54). Thus, TEAD4 knockdown can inhibit glucocorticoid-induced chemotherapy resistance in breast cancer, suggesting new properties of TEAD4 in breast cancer treatment (54). Analysis of the Hippo signaling pathway-related differentially expressed genes between breast cancer tissues and normal tissues revealed that only TEAD4 is overexpressed among all members of the TEAD family (Fig. 2A). Notably, TEAD4 expression is upregulated in TNBC tissues compared with the normal breast tissues (Fig. 2Β). Taken together, these results suggest that the use of agents targeting TEAD4 and inhibiting the formation of the YAP/TAZ-TEADs complex, exert promising results in in vitro experiments and animal models of different types of cancer. Thus, TEAD4 may be used as a potential target for the therapeutic intervention of tumors.
Figure 2.

Volcano maps of the Hippo signaling pathway-related differentially expressed genes in (A) Breast cancer tissues vs. normal tissues (B) TNBC tissues vs. normal tissues. Red dots represent upregulatedgenes, while blue dots represent downregulated genes in breast cancer or TNBC. Gray dots indicate fold-change <2 or no statistically significant difference. TNBC, triple-negative breast cancer; TEAD, transcriptional enhancer factor domain.
6. Conclusions
Breast cancer is the most common type of cancer and the leading cause of cancer-associated mortality among women worldwide. The treatment of breast cancer has gained significant global attention. The Hippo signaling pathway includes a cascade of signals that control tissue growth and organ size by attenuating cell proliferation and promoting cell death. When the Hippo signaling pathway is activated, the transcriptional activity of its target genes, which exert a pivotal effect on carcinogenesis, is repressed. Conversely, when the Hippo signaling pathway is aberrantly inactivated, the transcriptional activity of its target genes is enhanced, resulting in uncontrolled cell proliferation.
With regards to the upstream kinase cascade in the Hippo signaling pathway, the expression of oncogenes, tumor suppressor genes, as well as the ubiquitinating and deubiquitinating modifications can disrupt the function of these cascades, eventually affecting the human body. Increasing evidence suggest that the transcriptional coactivators, YAP and TAZ, serve important role in the Hippo signaling pathway. Studies on the regulation of the phosphorylation and nuclear localization of YAP/TAZ confirm that these coactivators play a key role in the development of breast cancer.
The TEAD family is the most important family of transcription factors for the transcription of YAP/TAZ coactivators. It is generally accepted that following YAP/TAZ activation, the transcriptional activity of TEADs increases. Notably, it has been reported that the function of TEADs is independent of YAP/TAZ. TCGA database analysis revealed that among the members of the TEAD family, TEAD4 expression is significantly upregulated in breast cancer, particularly in TNBC tissues compared with normal breast tissues. In conclusion, TEAD4 may act as a potential target for the treatment of breast cancer by targeting the Hippo signaling pathway.
Acknowledgements
Not applicable.
Funding Statement
The present review was supported by the Natural Science Foundation of China (grant nos. 81873052 and 81503301).
Funding
The present review was supported by the Natural Science Foundation of China (grant nos. 81873052 and 81503301).
Availability of data and materials
The datasets generated and analyzed during the current study are available in the TCGA repository https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga.
Authors' contributions
YW drafted and revised the initial manuscript. CH conceived and modified the manuscript. YW, ML and JL analyzed the data from TCGA database. YW and CH confirmed the authenticity of all the raw data. All authors have read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021 doi: 10.3322/caac.21660. (Epub ahead of print) [DOI] [PubMed] [Google Scholar]
- 2.Gadag S, Sinha S, Nayak Y, Garg S, Nayak UY. Combination therapy and nanoparticulate systems: Smart approaches for the effective treatment of breast cancer. Pharmaceutics. 2020;12:524. doi: 10.3390/pharmaceutics12060524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zanconato F, Cordenonsi M, Piccolo S. YAP/TAZ at the roots of cancer. Cancer Cell. 2016;29:783–803. doi: 10.1016/j.ccell.2016.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Yin F, Yu J, Zheng Y, Chen Q, Zhang N, Pan D. Spatial organization of Hippo signaling at the plasma membrane mediated by the tumor suppressor Merlin/NF2. Cell. 2013;154:1342–1355. doi: 10.1016/j.cell.2013.08.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Maugeri-Sacca M, De Maria R. The Hippo pathway in normal development and cancer. PharmacolTher. 2018;186:60–72. doi: 10.1016/j.pharmthera.2017.12.011. [DOI] [PubMed] [Google Scholar]
- 6.Moroishi T, Hansen CG, Guan KL. The emerging roles of YAP and TAZ in cancer. Nat Rev Cancer. 2015;15:73–79. doi: 10.1038/nrc3876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Maugeri-Sacca M, Barba M, Pizzuti L, Vici P, Di Lauro L, Dattilo R, Vitale I, Bartucci M, Mottolese M, De Maria R. The Hippo transducers TAZ and YAP in breast cancer: Oncogenic activities and clinical implications. Expert Rev Mol Med. 2015;17:e14. doi: 10.1017/erm.2015.12. [DOI] [PubMed] [Google Scholar]
- 8.Lamar JM, Stern P, Liu H, Schindler JW, Jiang ZG, Hynes RO. The Hippo pathway target, YAP, promotes metastasis through its TEAD-interaction domain. Proc Natl Acad Sci USA. 2012;109:E2441–E2450. doi: 10.1073/pnas.1212021109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wang Y, Li J, Gao Y, Luo Y, Luo H, Wang L, Yi Y, Yuan Z, Jim Xiao ZX. Hippo kinases regulate cell junctions to inhibit tumor metastasis in response to oxidative stress. Redox Biol. 2019;26:101233. doi: 10.1016/j.redox.2019.101233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lin XY, Cai FF, Wang MH, Pan X, Wang F, Cai L, Cui RR, Chen S, Biskup E. Mammalian sterile 20-like kinase 1 expression and its prognostic significance in patients with breast cancer. Oncol Lett. 2017;14:5457–5463. doi: 10.3892/ol.2017.6852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Heidary Arash E, Shiban A, Song S, Attisano L. MARK4 inhibits Hippo signaling to promote proliferation and migration of breast cancer cells. EMBO Rep. 2017;18:420–436. doi: 10.15252/embr.201642455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ercolani C, Di Benedetto A, Terrenato I, Pizzuti L, Di Lauro L, Sergi D, Sperati F, Buglioni S, Ramieri MT, Mentuccia L, et al. Expression of phosphorylated Hippo pathway kinases (MST1/2 and LATS1/2) in HER2-positive and triple-negative breast cancer patients treated with neoadjuvant therapy. Cancer Biol Ther. 2017;18:339–346. doi: 10.1080/15384047.2017.1312230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Furth N, Aylon Y. The LATS1 and LATS2 tumor suppressors: Beyond the Hippo pathway. Cell Death Differ. 2017;24:1488–1501. doi: 10.1038/cdd.2017.99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lehn S, Tobin NP, Sims AH, Stal O, Jirstrom K, Axelson H, Landberg G. Decreased expression of Yes-associated protein is associated with outcome in the luminal a breast cancer subgroup and with an impaired tamoxifen response. BMC Cancer. 2014;14:119. doi: 10.1186/1471-2407-14-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Maugeri-Sacca M, De Maria R. Hippo pathway and breast cancer stem cells. Crit Rev Oncol Hematol. 2016;99:115–122. doi: 10.1016/j.critrevonc.2015.12.004. [DOI] [PubMed] [Google Scholar]
- 16.Cordenonsi M, Zanconato F, Azzolin L, Forcato M, Rosato A, Frasson C, Inui M, Montagner M, Parenti AR, Poletti A, et al. The Hippo transducer TAZ confers cancer stem cell-related traits on breast cancer cells. Cell. 2011;147:759–772. doi: 10.1016/j.cell.2011.09.048. [DOI] [PubMed] [Google Scholar]
- 17.Bartucci M, Dattilo R, Moriconi C, Pagliuca A, Mottolese M, Federici G, Benedetto AD, Todaro M, Stassi G, Sperati F, et al. TAZ is required for metastatic activity and chemoresistance of breast cancer stem cells. Oncogene. 2015;34:681–690. doi: 10.1038/onc.2014.5. [DOI] [PubMed] [Google Scholar]
- 18.Diaz-Martin J, Lopez-Garcia MA, Romero-Perez L, Atienza-Amores MR, Pecero ML, Castilla MA, Biscuola M, Santon A, Palacios J. Nuclear TAZ expression associates with the triple-negative phenotype in breast cancer. EndocrRelat Cancer. 2015;22:443–454. doi: 10.1530/ERC-14-0456. [DOI] [PubMed] [Google Scholar]
- 19.Huang YT, Lan Q, Lorusso G, Duffey N, Ruegg C. The matricellular protein CYR61 promotes breast cancer lung metastasis by facilitating tumor cell extravasation and suppressing anoikis. Oncotarget. 2017;8:9200–9215. doi: 10.18632/oncotarget.13677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Di Benedetto A, Mottolese M, Sperati F, Ercolani C, Di Lauro L, Pizzuti L, Vici P, Terrenato I, Sperduti I, Shaaban AM, et al. The Hippo transducers TAZ/YAP and their target CTGF in male breast cancer. Oncotarget. 2016;7:43188–43198. doi: 10.18632/oncotarget.9668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Elster D, Jaenicke LA, Eilers M, von Eyss B. TEAD activity is restrained by MYC and stratifies human breast cancer subtypes. Cell Cycle. 2016;15:2551–2556. doi: 10.1080/15384101.2016.1207837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Xiang G, Liu F, Liu J, Meng Q, Li N, Niu Y. Prognostic role of Amphiregulin and the correlation with androgen receptor in invasive breast cancer. Pathol Res Pract. 2019;215:152414. doi: 10.1016/j.prp.2019.04.006. [DOI] [PubMed] [Google Scholar]
- 23.Niu J, Ma J, Guan X, Zhao X, Li P, Zhang M. Correlation between doppler ultrasound blood flow parameters and angiogenesis and proliferation activity in breast cancer. Med Sci Monit. 2019;25:7035–7041. doi: 10.12659/MSM.914395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.He Z, Zhao TT, Jin F, Li JG, Xu YY, Dong HT, Liu Q, Xing P, Zhu GL, Xu H, Miao ZF. Downregulation of RASSF6 promotes breast cancer growth and chemoresistance through regulation of Hippo signaling. BiochemBiophys Res Commun. 2018;503:2340–2347. doi: 10.1016/j.bbrc.2018.06.159. [DOI] [PubMed] [Google Scholar]
- 25.Liu J, Li J, Li P, Wang Y, Liang Z, Jiang Y, Li J, Feng C, Wang R, Chen H, et al. Loss of DLG5 promotes breast cancer malignancy by inhibiting the Hippo signaling pathway. Sci Rep. 2017;7:42125. doi: 10.1038/srep42125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Song GQ, Zhao Y. MAC30 knockdown involved in the activation of the Hippo signaling pathway in breast cancer cells. Biol Chem. 2018;399:1305–1311. doi: 10.1515/hsz-2018-0250. [DOI] [PubMed] [Google Scholar]
- 27.Han H, Qi R, Zhou JJ, Ta AP, Yang B, Nakaoka HJ, Seo G, Guan KL, Luo R, Wang W. Regulation of the Hippo pathway by phosphatidic Acid-Mediated Lipid-Protein Interaction. Mol Cell. 2018;72:328–340.e8. doi: 10.1016/j.molcel.2018.08.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Elaimy AL, Guru S, Chang C, Ou J, Amante JJ, Zhu LJ, Goel HL, Mercurio AM. VEGF-neuropilin-2 signaling promotes stem-like traits in breast cancer cells by TAZ-mediated repression of the Rac GAP β2-chimaerin. Sci Signal. 2018;11:eaao6897. doi: 10.1126/scisignal.aao6897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kim YN, Choe SR, Cho KH, Cho DY, Kang J, Park CG, Lee HY. Resveratrol suppresses breast cancer cell invasion by inactivating a RhoA/YAP signaling axis. Exp Mol Med. 2017;49:e296. doi: 10.1038/emm.2016.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Verma S, Yeddula N, Soda Y, Zhu Q, Pao G, Moresco J, Diedrich JK, Hong A, Plouffe S, Moroishi T, et al. BRCA1/BARD1-dependent ubiquitination of NF2 regulates Hippo-YAP1 signaling. Proc Natl Acad Sci USA. 2019;116:7363–7370. doi: 10.1073/pnas.1822155116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Li W, Cooper J, Zhou L, Yang C, Erdjument-Bromage H, Zagzag D, Snuderl M, Ladanyi M, Hanemann CO, Zhou P, et al. Merlin/NF2 loss-driven tumorigenesis linked to CRL4(DCAF1)-mediated inhibition of the hippo pathway kinases Lats1 and 2 in the nucleus. Cancer Cell. 2014;26:48–60. doi: 10.1016/j.ccr.2014.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ho KC, Zhou Z, She YM, Chun A, Cyr TD, Yang X. Itch E3 ubiquitin ligase regulates large tumor suppressor 1 stability [corrected] Proc Natl Acad Sci USA. 2011;108:4870–4875. doi: 10.1073/pnas.1101273108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ma B, Chen Y, Chen L, Cheng H, Mu C, Li J, Gao R, Zhou C, Cao L, Liu J, et al. Hypoxia regulates Hippo signalling through the SIAH2 ubiquitin E3 ligase. Nat Cell Biol. 2015;17:95–103. doi: 10.1038/ncb3073. [DOI] [PubMed] [Google Scholar]
- 34.Toloczko A, Guo F, Yuen HF, Wen Q, Wood SA, Ong YS, Chan PY, Shaik AA, Gunaratne J, Dunne MJ, et al. Deubiquitinating enzyme USP9X suppresses tumor growth via LATS kinase and core components of the Hippo pathway. Cancer Res. 2017;77:4921–4933. doi: 10.1158/0008-5472.CAN-16-3413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chen D, Sun Y, Wei Y, Zhang P, Rezaeian AH, Teruya-Feldstein J, Gupta S, Liang H, Lin HK, Hung MC, Ma L. LIFR is a breast cancer metastasis suppressor upstream of the Hippo-YAP pathway and a prognostic marker. Nat Med. 2012;18:1511–1517. doi: 10.1038/nm.2940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Li Z, Razavi P, Li Q, Toy W, Liu B, Ping C, Hsieh W, Sanchez-Vega F, Brown DN, Da Cruz Paula AF, et al. Loss of the FAT1 tumor suppressor promotes resistance to CDK4/6 inhibitors via the hippo pathway. Cancer Cell. 2018;34:893–905.e8. doi: 10.1016/j.ccell.2018.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Dang DK, Makena MR, Llongueras JP, Prasad H, Ko M, Bandral M, Rao R. A Ca2+-ATPase regulates E-cadherin biogenesis and epithelial-mesenchymal transition in breast cancer cells. Mol Cancer Res. 2019;17:1735–1747. doi: 10.1158/1541-7786.MCR-19-0070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yu S, Zhang M, Huang L, Ma Z, Gong X, Liu W, Zhang J, Chen L, Yu Z, Zhao W, et al. ERK1 indicates good prognosis and inhibits breast cancer progression by suppressing YAP1 signaling. Aging (Albany NY) 2019;11:12295–12314. doi: 10.18632/aging.102572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Dethlefsen C, Hansen LS, Lillelund C, Andersen C, Gehl J, Christensen JF, Pedersen BK, Hojman P. Exercise-induced catecholamines activate the Hippo tumor suppressor pathway to reduce risks of breast cancer development. Cancer Res. 2017;77:4894–4904. doi: 10.1158/0008-5472.CAN-16-3125. [DOI] [PubMed] [Google Scholar]
- 40.Wang Z, Kong Q, Su P, Duan M, Xue M, Li X, Tang J, Gao Z, Wang B, Li Z, et al. Regulation of Hippo signaling and triple negative breast cancer progression by an ubiquitin ligase RNF187. Oncogenesis. 2020;9:36. doi: 10.1038/s41389-020-0220-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhang Z, Du J, Wang S, Shao L, Jin K, Li F, Wei B, Ding W, Fu P, van Dam H, et al. OTUB2 promotes cancer metastasis via Hippo-independent activation of YAP and TAZ. Mol Cell. 2019;73:7–21.e7. doi: 10.1016/j.molcel.2018.10.030. [DOI] [PubMed] [Google Scholar]
- 42.Hou L, Xie S, Li G, Xiong B, Gao Y, Zhao X, Hu J, Deng S, Jiang J. IL-6 triggers the migration and invasion of oestrogen receptor-negative breast cancer cells via regulation of Hippo pathways. Basic Clin PharmacolToxicol. 2018;123:549–557. doi: 10.1111/bcpt.13042. [DOI] [PubMed] [Google Scholar]
- 43.Lv M, Lv M, Chen L, Qin T, Zhang X, Liu P, Yang J. Angiomotin promotes breast cancer cell proliferation and invasion. Oncol Rep. 2015;33:1938–1946. doi: 10.3892/or.2015.3780. [DOI] [PubMed] [Google Scholar]
- 44.Zanconato F, Forcato M, Battilana G, Azzolin L, Quaranta E, Bodega B, Rosato A, Bicciato S, Cordenonsi M, Piccolo S. Genome-wide association between YAP/TAZ/TEAD and AP-1 at enhancers drives oncogenic growth. Nat Cell Biol. 2015;17:1218–1227. doi: 10.1038/ncb3216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Janse van Rensburg HJ, Azad T, Ling M, Hao Y, Snetsinger B, Khanal P, Minassian LM, Graham CH, Rauh MJ, Yang X. The Hippo pathway component TAZ promotes immune evasion in human cancer through PD-L1. Cancer Res. 2018;78:1457–1470. doi: 10.1158/0008-5472.CAN-17-3139. [DOI] [PubMed] [Google Scholar]
- 46.Rashidian J, Le Scolan E, Ji X, Zhu Q, Mulvihill MM, Nomura D, Luo K. Ski regulates Hippo and TAZ signaling to suppress breast cancer progression. Sci Signal. 2015;8:ra14. doi: 10.1126/scisignal.2005735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Holden JK, Cunningham CN. Targeting the Hippo pathway and cancer through the TEAD family of transcription factors. Cancers (Basel) 2018;10:81. doi: 10.3390/cancers10030081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Qi Y, Yu J, Han W, Fan X, Qian H, Wei H, Tsai YH, Zhao J, Zhang W, Liu Q, et al. A splicing isoform of TEAD4 attenuates the Hippo-YAP signalling to inhibit tumour proliferation. Nat Commun. 2016;7:ncommss11840. doi: 10.1038/ncomms11840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gibault F, Sturbaut M, Bailly F, Melnyk P, Cotelle P. Targeting transcriptional enhanced associate domains (TEADs) J Med Chem. 2018;61:5057–5072. doi: 10.1021/acs.jmedchem.7b00879. [DOI] [PubMed] [Google Scholar]
- 50.Zhou Y, Huang T, Cheng AS, Yu J, Kang W, To KF. The TEAD family and its oncogenic role in promoting tumorigenesis. Int J Mol Sci. 2016;17:138. doi: 10.3390/ijms17010138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhu C, Li L, Zhang Z, Bi M, Wang H, Su W, Hernandez K, Liu P, Chen J, Chen M, et al. A non-canonical role of YAP/TEAD is required for activation of estrogen-regulated enhancers in breast cancer. Mol Cell. 2019;75:791–806.e8. doi: 10.1016/j.molcel.2019.06.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Liu Y, Wang G, Yang Y, Mei Z, Liang Z, Cui A, Wu T, Liu CY, Cui L. Increased TEAD4 expression and nuclear localization in colorectal cancer promote epithelial-mesenchymal transition and metastasis in a YAP-independent manner. Oncogene. 2016;35:2789–2800. doi: 10.1038/onc.2015.342. [DOI] [PubMed] [Google Scholar]
- 53.Yu MH, Zhang W. TEAD1 enhances proliferation via activating SP1 in colorectal cancer. Biomed Pharmacother. 2016;83:496–501. doi: 10.1016/j.biopha.2016.06.058. [DOI] [PubMed] [Google Scholar]
- 54.He L, Yuan L, Sun Y, Wang P, Zhang H, Feng X, Wang Z, Zhang W, Yang C, Zeng YA, et al. Glucocorticoid receptor signaling activates TEAD4 to promote breast cancer progression. Cancer Res. 2019;79:4399–4411. doi: 10.1158/0008-5472.CAN-19-0012. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets generated and analyzed during the current study are available in the TCGA repository https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga.