

Abstract
Objective
Necrotizing enterocolitis (NEC) is the most frequent life-threatening gastrointestinal disease experienced by premature infants in neonatal intensive care units all over the world. The objective of the present study was to take advantage of RNA-Seq data from the analysis of intestinal specimens of preterm infants diagnosed with NEC. Function enrichments with Gene Ontology and the Kyoto Encyclopedia of Genes and Genomes were used to analyse previous data in order to identify biological and functional processes, which could provide more insight into the pathogenesis of NEC in infants.
Results
Gene set enrichment analysis indicated that the most significant biological pathways over-represented in NEC neonates were closely associated with innate immune functions. One of the striking observations was the highly modulated expression of inflammatory genes related to the IL-17 pathway including such as pro-inflammatory cytokines (CXCL8), chemokines (CXCL5 and CXCL10) and antimicrobials (DEF5A, DEF6A, LCN2, NOS2) in the intestine of neonates diagnosed with NEC. Interestingly, the increase in IL-17 expression appeared to be under the IL-17F form, as reported in Crohn's disease, another inflammatory bowel disease. Further investigation is thus still needed to determine the precise role of IL-17F and its downstream targets in NEC.
Keywords: Human intestine, Preterm newborn, Transcriptomics, Gene expression, Interleukin-17
Introduction
Necrotizing enterocolitis (NEC) is the most common life-threatening condition of premature infants occurring in neonatal intensive care units [1–3]. This gastrointestinal disease is characterized by severe intestinal inflammation, intestinal necrosis and high morbidity [4]. The infants who survive NEC are at a higher risk for developing short bowel syndrome, impaired growth and neurodevelopmental outcomes [5, 6]. Several epidemiological investigations have suggested that preterm birth, enteral feeding and abnormal bacterial colonization are likely to play major roles in the pathogenesis of NEC [7, 8]. However, the precise mechanisms leading to the development of NEC remain to be elucidated.
One of the greatest challenges for neonatologists is to identify early reliable clinical signs and symptoms of NEC [1, 2]. In this context, the ahead of time clinical manifestations of NEC are relatively nonspecific and at diagnosis NEC is often already at an advanced stage due first to the initially insidious condition and then to the fierce progression of the disease [9, 10]. One strategy to prevent or treat NEC would be to develop an early diagnostic tool allowing the identification of preterm infants either at risk of developing NEC or at the onset of symptoms to aid in the diagnostic dilemma and treatment.
Recently, we used the high-throughput sequencing of RNA transcripts (RNA-Seq) approach to determine the complete gene expression profiles of ileal specimens, which were resected from preterm infants diagnosed with NEC vs non-NEC conditions [11]. This recently important work has led to the identification of potential biomarkers for the prediction of the disease. The objective of the present study was to provide new insights into the molecular mechanisms of NEC pathophysiology by conducting function enrichment analyses with Gene Ontology (GO) and the Kyoto Encyclopedia of Genes and Genomes (KEGG). Our analysis revealed that immune and inflammatory responses were strongly modulated in the small intestine of neonates with NEC, supporting the finding that NEC development is related to the immaturity of the intestinal mucosa challenged by microbial dysbiosis [12, 13]. Interestingly, we also observed that several interleukin-17 (IL-17) target genes, including chemokines and antimicrobial proteins, were strongly modulated in the small intestine of neonates with NEC. Pro-inflammatory IL-17 has been linked to the pathogenesis of diverse autoimmune and chronic inflammatory diseases while being essential for host defense against microbial colonization [14, 15]. Our results suggest a role for IL-17 signaling, specifically IL-17F, in the development of NEC.
Main text
Materials and methods
Study population and informed consent
As detailed in the previous report [11], the present collaborative study recruited premature infants from neonatal intensive care units at the Centre Hospitalier Universitaire de Sherbrooke (Sherbrooke, QC, Canada), Erasmus MC-Sophia Children’s Hospital (Rotterdam, The Netherlands), Children’s Hospital of Eastern Ontario (Ottawa, ON, Canada) and Hôpital Pierre Zobda-Quitman (Fort-de-France, Martinique), between October 2008 and May 2013. After the approval of the Institutional Review Committee for the use of human material in each center, written informed consent from parents or guardians was obtained for each patient.
Premature infants who had undergone bowel resection were eligible for the study. The diagnosis and staging of NEC were based according to the Bell's criteria [9]. Freshly resected intestinal specimens taken from the ileum were conserved in RNAlater (Ambion) before RNA extraction. Preterm patients who had undergone bowel resection for stage III acute NEC were referred to as NEC, and preterm patients who had undergone resection for diseases other than NEC were included as control patients (CTRL) as detailed in [11].
Sample preparation and RNA sequencing
The procedures for sample preparation, library preparation and sequencing, and data analysis were performed as previously described by Tremblay et al. [11].
Functional pathway enrichment analysis
The bioinformatics software DAVID 6.8 (https://david.ncifcrf.gov/) [16] was used to perform Gene Ontology (GO) analysis and KEGG pathways enrichment analysis of differentially expressed genes (DEGs) identified in the pathogenesis of NEC.
Data validation by quantitative polymerase chain reaction (qPCR)
All qPCR reactions were performed as previously described [11]. The genes investigated were chemokine (C-X-C motif) ligand 8 (CXCL8) and 10 (CXCL10), alpha-defensin 5 (DEFA5) and 6 (DEFA6), interleukin-17A (IL-17A) and IL1-7F (IL-17F), interleukin 6 (IL6), lipocalin 2 (LCN2), nuclear factor kappa B subunit 1 (NFKB1), regenerating islet-derived 3 alpha (REG3A), ribosomal protein lateral stalk subunit P0 (RPLP0), ribosomal protein 23 (RPS23), and tumor necrosis factor (TNF). Primers (listed in Additional file 1: Table S1) were generated using the primer formation software Primer3 (http://bioinfo.ut.ee/primer3). Differences in gene expression were evaluated by comparing the expression of control and NEC samples using the equation R = (Etarget)∆Ct target/(Ereference)∆Ct reference) [17]. Samples were normalized to a set of 2 reference genes, RPLP0 and RPS23 [18, 19].
Results
RNA-Seq analysis and identification of DEGs
The data for RNA-Seq analysis on ileum of preterm infants with NEC vs without NEC (CTRL) have been deposited at the National Center for Biotechnology Information's Gene Expression Omnibus and are accessible through GEO Series accession number GSE64801 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=gse64801). In total, 804 DEGs were identified (p < 0.05), 383 upregulated and 421 downregulated [11].
GO and KEGG pathway analyses in NEC samples
In the previous study, IPA analysis revealed an impaired immune response in the intestine of NEC neonates where several related pathways and biological functions were found to be altered in the samples of NEC infants [11]. To gain more insight in the function pattern of the DEGs obtained previously, GO and KEGG pathway analyses were conducted by DAVID. Among the 151 GO "Biological Process" terms enriched in DEGs, the top 8 terms, which were selected based on their highest p values, were highly associated with innate and adaptive immune as well as inflammatory responses (Fig. 1a). The same relationship with the immune responses was also noted with KEGG analyses (Fig. 1b). Intriguingly, a further detailed analysis of the DEGs revealed that many of those genes were highly associated with the signature target genes of IL-17 (Fig. 2), suggesting a role for this cytokine in the pathogenesis of NEC. Among these IL-17 target genes, changes in expression of pro-inflammatory cytokines, chemokines, antimicrobials and matrix metalloproteinase were identified.
Fig. 1.

Functional pathways in NEC samples. The negative logarithm of p-values (Fisher's test) calculated for the 8 most significant pathways overrepresented in small intestinal NEC samples compared to control non-NEC samples for GO Biological process (a) and KEGG (b) pathways. Dotted line: -Log 0.05 corresponds to 1.3
Fig. 2.

Association of DEG identified in NEC samples and IL-17 target genes. IL-17 receptor (IL-17R) can bind to IL-17A, IL-17F or IL-17A/F to activate various signaling pathways leading to the regulation of IL-17 targets genes involved in tissue remodeling and anti-microbial activity as well as mediators of inflammation as adapted from [15, 20]. Genes identified as being significantly modulated in NEC samples compared with control non-NEC samples are labeled in green and red while those for which no significant differences were notes are labeled in grey
Validation of the gene expression profile in NEC samples
To further investigate the gene expression profile identified in RNA-Seq analyses, we used qPCR to test representative DEGs known to be involved in IL-17 signaling in NEC samples as depicted in Fig. 2. As shown in Fig. 3a, qPCR analyses confirmed the upregulation of many of the IL-17 downstream genes in NEC samples. Incidentally, transcript expression of the two main IL-17 cytokines, IL-17A and IL-17F was also investigated and revealed a significant induction of IL-17F in NEC samples (Fig. 3b) confirming alterations in the IL-17 signature gene profile in the intestinal samples of patients with NEC.
Fig. 3.
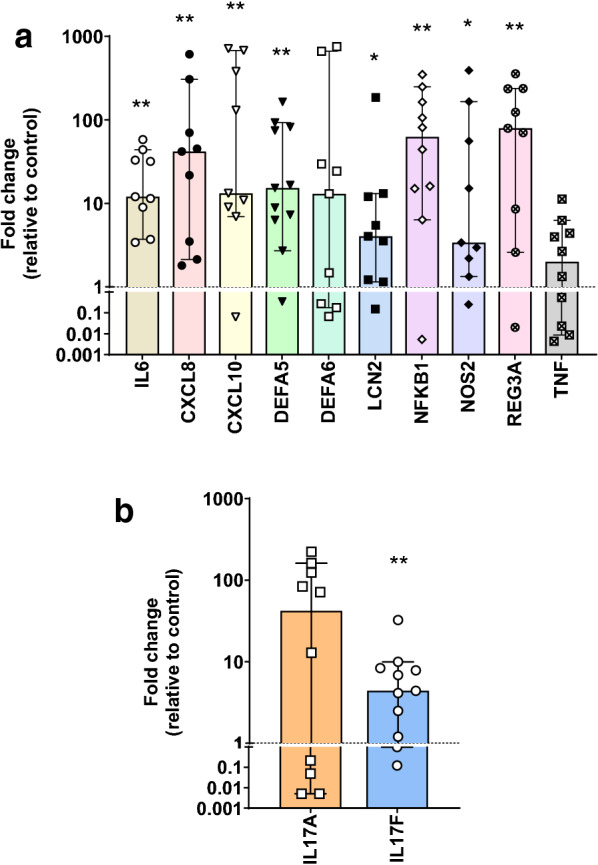
Differential expression of IL-17-related genes assessed in intestinal NEC and non-NEC control samples by real-time qPCR. Transcript levels of IL-17 target genes (a) and IL17A and IL17F transcripts (b) in NEC relative to control. Results are expressed as a scatter dot plot showing individual data and as median with 95% confidence interval. * and ** are p < 0.05 and p < 0.01, respectively, using the one sample Wilcoxon test
Discussion
NEC is a complex and a potentially fatal disease that remains a major cause of morbidity in neonates. Although significant effort has been made on the further understanding of this disease, no effective treatment or further diagnostic tools have been discovered [3]. In the present study, we performed GO and KEGG pathway analyses on RNA-seq data that was previously done with the aim to further deepen our knowledge of the molecular mechanisms underlying NEC pathogenesis. GO and KEGG pathway analyses showed strong enrichment in genes involved in innate and adaptive immune responses, consistent with a disproportionate inflammatory response following an exposure to luminal microorganisms [11]. Indeed, excessive inflammatory response and overexpression of proinflammatory cytokines are the main pathophysiological characteristics of NEC that contribute to damage the intestinal epithelium [13]. Moreover, by reviewing in detail all enriched genes in these pathways, we highlighted several target genes of the IL-17 pathway [20], which were modulated in NEC, suggesting a role for IL-17 signaling in the pathophysiology of this severe inflammatory disease.
The IL-17 family of cytokines consists of six structurally related proteins, IL-17A through IL-17F [15]. IL-17A and IL-17F are the most related members and they are usually secreted as homodimers or heterodimers by various innate immune cells [21] including those residing in the gut [22]. One of the major functions of IL-17 is to provide a protective inflammatory response to the host against pathogens [23–25]. However, despite the protective effects of IL-17 during infection, there is emerging evidence that dysregulated IL-17 expression could worsen the gravity of some diseases [26, 27], namely NEC [28, 29]. It is noteworthy that IL-17 exerts only modest activity by its own, its immunomodulatory role arising from synergistic action with other cytokines [24]. In preterm infants, IL-17 level was found to be increased after red blood cell transfusion that could lead to a higher risk of developing NEC [30]. Moreover, an increased IL-17 level has been related to impairment of enterocyte tight junctions and increased epithelial cell apoptosis through Toll-like receptor 4-dependent Th17 polarization in mouse and human NEC models [31] consistent with its key role in host defence [24]. Indeed, as reviewed recently, although evidence for the involvement of adaptative immunity in NEC initiation and perpetuation is still insufficient, the potential for a pathogenic role of Th17 responses is pointed out [25]. In our study, we showed that the transcript expression of IL-17F, but not IL-17A, is upregulated in the intestine of premature infants diagnosed with NEC. Distinct roles between IL-17A and IL-17F have been previously demonstrated in IL-17F-deficient mice [32, 33]. Upregulated IL-17F expression was also reported in intestinal samples obtained from patients with active Crohn's disease [34]. In the experimental colitis model induced by DSS, IL-17F−/− mice were protected, whereas IL-17-knockout mice showed a more severe condition [32]. In addition, a recent study showed that a potential genetic variant of IL-17F, but not IL-17A, was associated with an increased severity of NEC [35]. Taken together, these observations support a potential involvement for the upregulated expression of IL-17F in the development of NEC. Based on the similarity between NEC and Crohn's disease [11], one may speculate that as proposed for Crohn's disease [36], the development of specific inhibitors for IL-17F could become a potential option for treating NEC although it is important to consider that other organs such as the lung or the brain could also be affected by IL-17 blockade [37, 38].
Upregulation of IL-17 controls the expression of inflammatory genes in the majority of non-hematopoietic cells [39]. The trademark of IL-17 signaling is induction of pro-inflammatory cytokines, chemokines, antimicrobial proteins and inflammatory effectors, to maintain mucosal barrier integrity against infections [20, 40]. Among the IL-17 signature genes, we have found the transcription factor NFKB1, chemokines such as CXCL8, CXCL10 as well as a plethora of antimicrobial molecules or regulators (DEFA5, LCN2, NOS2, REG3A) to be upregulated in NEC intestinal samples as compared to control. These observations suggest that IL-17 could be involved in a protective mucosal reaction towards pathogenic microorganisms in the intestine of NEC infants. Unexpectedly, we observed that expression levels of IL-17-related pro-inflammatory cytokines (CSF2, CSF3, and TNF) were either downregulated or unchanged in NEC tissues. In the early phase of infection, these cytokines have been reported to be involved in neutrophil recruitment for an effective innate immunity and rapid control against pathogens [41, 42]. One possible explanation of this discrepancy could be the time lapse between early induction of these cytokines and the resection of tissue that occurred at the late phase of the disease.
In conclusion, this study has led to the identification of a characteristic IL-17 core gene signature in the pro-inflammatory intestinal microenvironment of the premature infant with NEC. Furthermore, it appears that it is IL-17F, rather than IL-17A, which is consistently involved in the pro-inflammatory cascade in neonates affected with NEC. However, further investigations need to be undertaken to validate the role of IL-17F in the development of NEC at the functional level.
Limitations
Intestinal samples from premature infants diagnosed with NEC are difficult to obtain [43]. It is noteworthy that the small cohort of control and NEC patients represents a limitation in interpreting the results. A larger cohort which could also include NEC patients at earlier stages would help to better document the role of IL-17 in NEC initiation as well as increase the statistical power to better detect differentially expressed genes.
Supplementary Information
Additional file1: Table S1. Primers used for qPCR.
Acknowledgements
The authors thank Elizabeth Herring for reviewing the manuscript. We also thank the other members of the Research Consortium on Child Intestinal Inflammation who contributed to the initial work: Valérie Bertelle and Sandeep Mayer, Université de Sherbrooke; Marcos Bettolli and David Grynspan, University of Ottawa; Jean-François Colombani, CHU de Martinique, Olivier Mouterde, CHU de Rouen, France; and Ingrid B. Renes, Erasmus MC-Sophia Children’s Hospital, The Netherlands.
Abbreviations
- CTRL
Control
- DEG
Differentially expressed genes
- GO
Gene ontology
- IL-17
Interleukin-17
- KEGG
Kyoto Encyclopedia of Genes and Genomes
- NEC
Necrotizing enterocolitis
- qPCR
Quantitative polymerase chain reaction
- RNA-Seq
High-throughput sequencing of RNA transcripts
Authors’ contributions
ET, EF, CB, KMB, EL and JFB were collectively involved in the study concept and the design of the approach; ET and EF acquired the data; ET, EF and JFB analyzed and interpreted the data; ET and JFB wrote the manuscript; EF, CB, KMB and EL reviewed the manuscript. All authors read and approved the final manuscript.
Funding
This work was funded by a Grant from the Canadian Institutes of Health Research # MOP-136991.
Availability of data and materials
The datasets used during the current study are available through GEO Series accession number GSE64801. The datasets analyzed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
This research was approved by the Institutional Review Committee for the use of human material from the Centre Hospitalier Universitaire de Sherbrooke while the initial research was approved Institutional Review Committee for the use of human material for each of the collaborating centers and written informed consent from parents or guardians was obtained for each patient.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Éric Tremblay, Email: eric.tremblay@usherbrooke.ca.
Emanuela Ferretti, Email: eferretti@ton.on.ca.
Corentin Babakissa, Email: corentin.babakissa@usherbrooke.ca.
Karolina Maria Burghardt, Email: karolina.burghardt@gmail.com.
Emile Levy, Email: emile.levy@recherche-ste-justine.qc.ca.
Jean-François Beaulieu, Email: jean-francois.beaulieu@usherbrooke.ca.
Supplementary Information
The online version contains supplementary material available at 10.1186/s13104-021-05489-9.
References
- 1.Henry MC, Moss RL. Necrotizing enterocolitis. Annu Rev Med. 2009;60:111–124. doi: 10.1146/annurev.med.60.050207.092824. [DOI] [PubMed] [Google Scholar]
- 2.Neu J, Walker WA. Necrotizing enterocolitis. N Engl J Med. 2011;364(3):255–264. doi: 10.1056/NEJMra1005408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Neu J. Necrotizing enterocolitis: the future. Neonatology. 2020;117(2):240–244. doi: 10.1159/000506866. [DOI] [PubMed] [Google Scholar]
- 4.Berman L, Moss RL. Necrotizing enterocolitis: an update. Semin Fetal Neonatal Med. 2011;16(3):145–150. doi: 10.1016/j.siny.2011.02.002. [DOI] [PubMed] [Google Scholar]
- 5.Huda S, Chaudhery S, Ibrahim H, Pramanik A. Neonatal necrotizing enterocolitis: clinical challenges, pathophysiology and management. Pathophysiology. 2014;21(1):3–12. doi: 10.1016/j.pathophys.2013.11.009. [DOI] [PubMed] [Google Scholar]
- 6.Bazacliu C, Neu J. Necrotizing enterocolitis: long term complications. Curr Pediatr Rev. 2019;15(2):115–124. doi: 10.2174/1573396315666190312093119. [DOI] [PubMed] [Google Scholar]
- 7.Afrazi A, Sodhi CP, Richardson W, Neal M, Good M, Siggers R, et al. New insights into the pathogenesis and treatment of necrotizing enterocolitis: toll-like receptors and beyond. Pediatr Res. 2011;69(3):183–188. doi: 10.1203/PDR.0b013e3182093280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Grave GD, Nelson SA, Walker WA, Moss RL, Dvorak B, Hamilton FA, et al. New therapies and preventive approaches for necrotizing enterocolitis: report of a research planning workshop. Pediatr Res. 2007;62(4):510–514. doi: 10.1203/PDR.0b013e318142580a. [DOI] [PubMed] [Google Scholar]
- 9.Bell MJ, Ternberg JL, Feigin RD, Keating JP, Marshall R, Barton L, et al. Neonatal necrotizing enterocolitis. Therapeutic decisions based upon clinical staging. Ann Surg. 1978;187(1):1–7. doi: 10.1097/00000658-197801000-00001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pietz J, Achanti B, Lilien L, Stepka EC, Mehta SK. Prevention of necrotizing enterocolitis in preterm infants: a 20-year experience. Pediatrics. 2007;119(1):e164–e170. doi: 10.1542/peds.2006-0521. [DOI] [PubMed] [Google Scholar]
- 11.Tremblay E, Thibault MP, Ferretti E, Babakissa C, Bertelle V, Bettolli M, et al. Gene expression profiling in necrotizing enterocolitis reveals pathways common to those reported in Crohn's disease. BMC Med Genomics. 2016;9:6. doi: 10.1186/s12920-016-0166-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Berrington JE, Stewart CJ, Cummings SP, Embleton ND. The neonatal bowel microbiome in health and infection. Curr Opin Infect Dis. 2014;27(3):236–243. doi: 10.1097/QCO.0000000000000061. [DOI] [PubMed] [Google Scholar]
- 13.Nanthakumar N, Meng D, Goldstein AM, Zhu W, Lu L, Uauy R, et al. The mechanism of excessive intestinal inflammation in necrotizing enterocolitis: an immature innate immune response. PLoS ONE. 2011;6(3):e17776. doi: 10.1371/journal.pone.0017776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Amatya N, Garg AV, Gaffen SL. IL-17 Signaling: The Yin and the Yang. Trends Immunol. 2017;38(5):310–322. doi: 10.1016/j.it.2017.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.McGeachy MJ, Cua DJ, Gaffen SL. The IL-17 family of cytokines in health and disease. Immunity. 2019;50(4):892–906. doi: 10.1016/j.immuni.2019.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.da Huang W, Sherman BT, Lempicki RA. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009;37(1):1–13. doi: 10.1093/nar/gkn923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001;29(9):e45. doi: 10.1093/nar/29.9.e45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Thibault MP, Tremblay E, Wallace JL, Beaulieu JF. Effect of Ketoprofen and ATB-352 on the Immature Human Intestine: Identification of Responders and Non-responders. J Pediatr Gastroenterol Nutr. 2019;68(5):623–629. doi: 10.1097/MPG.0000000000002308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Dydensborg AB, Herring E, Auclair J, Tremblay E, Beaulieu J-F. Normalizing genes for quantitative RT-PCR in differentiating human intestinal epithelial cells and adenocarcinomas of the colon. Am J Physiol Gastrointest Liver Physiol. 2006;290(5):G1067–G1074. doi: 10.1152/ajpgi.00234.2005. [DOI] [PubMed] [Google Scholar]
- 20.Onishi RM, Gaffen SL. Interleukin-17 and its target genes: mechanisms of interleukin-17 function in disease. Immunology. 2010;129(3):311–321. doi: 10.1111/j.1365-2567.2009.03240.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Reynolds JM, Angkasekwinai P, Dong C. IL-17 family member cytokines: regulation and function in innate immunity. Cytokine Growth Factor Rev. 2010;21(6):413–423. doi: 10.1016/j.cytogfr.2010.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cua DJ, Tato CM. Innate IL-17-producing cells: the sentinels of the immune system. Nat Rev Immunol. 2010;10(7):479–489. doi: 10.1038/nri2800. [DOI] [PubMed] [Google Scholar]
- 23.Chamoun MN, Blumenthal A, Sullivan MJ, Schembri MA, Ulett GC. Bacterial pathogenesis and interleukin-17: interconnecting mechanisms of immune regulation, host genetics, and microbial virulence that influence severity of infection. Crit Rev Microbiol. 2018;44(4):465–486. doi: 10.1080/1040841X.2018.1426556. [DOI] [PubMed] [Google Scholar]
- 24.Veldhoen M. Interleukin 17 is a chief orchestrator of immunity. Nat Immunol. 2017;18(6):612–621. doi: 10.1038/ni.3742. [DOI] [PubMed] [Google Scholar]
- 25.Cho SX, Berger PJ, Nold-Petry CA, Nold MF. The immunological landscape in necrotising enterocolitis. Expert Rev Mol Med. 2016;18:e12. doi: 10.1017/erm.2016.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bai H, Cheng J, Gao X, Joyee AG, Fan Y, Wang S, et al. IL-17/Th17 promotes type 1 T cell immunity against pulmonary intracellular bacterial infection through modulating dendritic cell function. J Immunol. 2009;183(9):5886–5895. doi: 10.4049/jimmunol.0901584. [DOI] [PubMed] [Google Scholar]
- 27.Feinen B, Jerse AE, Gaffen SL, Russell MW. Critical role of Th17 responses in a murine model of Neisseria gonorrhoeae genital infection. Mucosal Immunol. 2010;3(3):312–321. doi: 10.1038/mi.2009.139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ma F, Li S, Gao X, Zhou J, Zhu X, Wang D, et al. Interleukin-6-mediated CCR9(+) interleukin-17-producing regulatory T cells polarization increases the severity of necrotizing enterocolitis. EBioMedicine. 2019;44:71–85. doi: 10.1016/j.ebiom.2019.05.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pang Y, Du X, Xu X, Wang M, Li Z. Monocyte activation and inflammation can exacerbate Treg/Th17 imbalance in infants with neonatal necrotizing enterocolitis. Int Immunopharmacol. 2018;59:354–360. doi: 10.1016/j.intimp.2018.04.026. [DOI] [PubMed] [Google Scholar]
- 30.Dani C, Poggi C, Gozzini E, Leonardi V, Sereni A, Abbate R, et al. Red blood cell transfusions can induce proinflammatory cytokines in preterm infants. Transfusion. 2017;57(5):1304–1310. doi: 10.1111/trf.14080. [DOI] [PubMed] [Google Scholar]
- 31.Egan CE, Sodhi CP, Good M, Lin J, Jia H, Yamaguchi Y, et al. Toll-like receptor 4-mediated lymphocyte influx induces neonatal necrotizing enterocolitis. J Clin Invest. 2016;126(2):495–508. doi: 10.1172/JCI83356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yang XO, Chang SH, Park H, Nurieva R, Shah B, Acero L, et al. Regulation of inflammatory responses by IL-17F. J Exp Med. 2008;205(5):1063–1075. doi: 10.1084/jem.20071978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ogawa A, Andoh A, Araki Y, Bamba T, Fujiyama Y. Neutralization of interleukin-17 aggravates dextran sulfate sodium-induced colitis in mice. Clin Immunol. 2004;110(1):55–62. doi: 10.1016/j.clim.2003.09.013. [DOI] [PubMed] [Google Scholar]
- 34.Seiderer J, Elben I, Diegelmann J, Glas J, Stallhofer J, Tillack C, et al. Role of the novel Th17 cytokine IL-17F in inflammatory bowel disease (IBD): upregulated colonic IL-17F expression in active Crohn's disease and analysis of the IL17F p.His161Arg polymorphism in IBD. Inflamm Bowel Dis. 2008;14(4):437–445. doi: 10.1002/ibd.20339. [DOI] [PubMed] [Google Scholar]
- 35.Tian J, Liu Y, Jiang Y, Zhou H, Zhu T, Zhao X, et al. Association of single nucleotide polymorphisms of IL23R and IL17 with necrotizing enterocolitis in premature infants. Mol Cell Biochem. 2017;430(1–2):201–209. doi: 10.1007/s11010-017-2972-6. [DOI] [PubMed] [Google Scholar]
- 36.Zwicky P, Unger S, Becher B. Targeting interleukin-17 in chronic inflammatory disease: a clinical perspective. J Exp Med. 2020;217:1. doi: 10.1084/jem.20191123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ye B, Tao T, Zhao A, Wen L, He X, Liu Y, et al. Blockade of IL-17A/IL-17R Pathway Protected Mice from Sepsis-Associated Encephalopathy by Inhibition of Microglia Activation. Mediators Inflamm. 2019;2019:8461725. doi: 10.1155/2019/8461725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chenuet P, Fauconnier L, Madouri F, Marchiol T, Rouxel N, Ledru A, et al. Neutralization of either IL-17A or IL-17F is sufficient to inhibit house dust mite induced allergic asthma in mice. Clin Sci (Lond) 2017;131(20):2533–2548. doi: 10.1042/CS20171034. [DOI] [PubMed] [Google Scholar]
- 39.Iwakura Y, Ishigame H, Saijo S, Nakae S. Functional specialization of interleukin-17 family members. Immunity. 2011;34(2):149–162. doi: 10.1016/j.immuni.2011.02.012. [DOI] [PubMed] [Google Scholar]
- 40.Chang SH, Dong C. Signaling of interleukin-17 family cytokines in immunity and inflammation. Cell Signal. 2011;23(7):1069–1075. doi: 10.1016/j.cellsig.2010.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Curtis MM, Way SS. Interleukin-17 in host defence against bacterial, mycobacterial and fungal pathogens. Immunology. 2009;126(2):177–185. doi: 10.1111/j.1365-2567.2008.03017.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Iwakura Y, Nakae S, Saijo S, Ishigame H. The roles of IL-17A in inflammatory immune responses and host defense against pathogens. Immunol Rev. 2008;226:57–79. doi: 10.1111/j.1600-065X.2008.00699.x. [DOI] [PubMed] [Google Scholar]
- 43.Allin BSR, Long AM, Gupta A, Lakhoo K, Knight M. British Association of Paediatric Surgeons Congenital Anomalies Surveillance System Necrotising Enterocolitis C. One-year outcomes following surgery for necrotising enterocolitis: a UK-wide cohort study. Arch Dis Child Fetal Neonatal Ed. 2018;103(5):F461–F466. doi: 10.1136/archdischild-2017-313113. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file1: Table S1. Primers used for qPCR.
Data Availability Statement
The datasets used during the current study are available through GEO Series accession number GSE64801. The datasets analyzed during the current study are available from the corresponding author on reasonable request.