

ABSTRACT
The emergence of New Delhi Metallo-β-lactamase (NDM)-producing Klebsiella pneumoniae has aroused critical concern worldwide. Herein, we reported the first emergence of NDM-5-producing K. pneumoniae isolates in a 68-year-old lung transplant recipient, who died of septic shock 13 days after surgery. The K. pneumoniae strain KP22937 isolated from the bloodstream of the patient was analyzed for phenotypes and genotypes. KP22937 belonged to sequence type (ST) 65 and capsule serotype K2, contained iucABCDiutA and iroBCDN virulence clusters, showed high virulence to mice, and was therefore considered a hypervirulent K. pneumoniae. The blaNDM-5 gene was located on a genomic island region of the IncX3-type plasmid pNDM22937, which was successfully transferred to Escherichia coli EC600 with insignificant fitness costs. The transconjugant demonstrated similar antimicrobial susceptibility and growth kinetics to the recipient E. coli EC600. The plasmid pNDM22937 was almost identical to the blaNDM-5-carrying IncX3 plasmids previously reported in K. pneumoniae strains with different ST types and in other species. Our findings raise concerns about the horizontal spread of blaNDM-5 gene mediated by IncX3 plasmid, where hypervirulent K. pneumoniae strains are also involved. Stricter control measures are needed to prevent the dissemination of the novel clone in hospital settings.
KEYWORDS: ST65, NDM-5, Klebsiella pneumoniae, IncX3, hypervirulence
New Delhi Metallo-β-lactamase (NDM) producing Klebsiella pneumoniae strains are capable of hydrolyzing all β-lactams except monobactam. The blaNDM-5 gene was first identified in 2011 from an Escherichia coli strain EC045 from a patient with a hospitalization history in India [1]. Subsequently, this carbapenemase gene has been detected in K. pneumoniae, and has caused sporadic outbreaks worldwide [2]. It has been identified in various ST types of K. pneumoniae, among which ST29 K. pneumoniae strain SCNJ1 and ST35 K. pneumoniae strain RJY9645 have been confirmed to be carbapenem-resistant hypervirulent K. pneumoniae (CR-hvKp) [3,4]. The treatment options are limited for severe infection caused by CR-hvKp [4]. Herein, we reported an NDM-5-producing hypervirulent K. pneumoniae ST65 strain in a post-transplant patient, which is a serious public health concern.
In June 2019, a 68-year-old lung transplantation recipient experienced a severe postoperative infection. Multiple specimens were sent for pathogen testing on postoperative day 2, 5, 6 and 7, and empirical antibacterial treatment was administered according to the patient's symptoms until the pathogen results were returned (Figure 1A). Aztreonam was immediately administered after receiving the antimicrobial susceptibility test (AST) result, and the patient's temperature and circulation gradually improved. However, the patient developed chest emphysema on postoperative day 10, and bronchoscopy revealed a large amount of pus moss. His condition dramatically deteriorated, and he died of septic shock.
Figure 1.

Clinical data of the lung transplantation recipient and genetic features of the two plasmids (pNDM22937 and pVir22937) of K. pneumoniae strain KP22937. (A) Time and site of isolation of K. pneumoniae from the patient and antimicrobial therapy. TZP, piperacillin/tazobactam; VA, vancomycin; CBF, caspofungin; AMB, amphotericin B; TGC, tigecycline; MEM, meropenem; ATM, aztreonam; VRC, voriconazole; PLB, polymyxin B. (B) Circular plasmid maps of the two plasmids (red colour) and their respective reference plasmids (blue and orange colours). Purple colour region is the genomic island in the plasmid pNDM22937. Red text on the plasmid maps indicates the main virulence determinants and resistance gene of strain KP22937. (C) Genetic contents of blaNDM-5 gene. ORFs are indicated by arrows. Sequences of shared homology between two plasmids are marked by grey shading.
A total of eight K. pneumoniae isolates were recovered from different specimens of the patient (Figure 1A), including blood (n = 3), bronchoalveolar lavage fluid (BALF, n = 3), sputum (n = 1), and drainage fluid (n = 1). AST using Vitek-2 system and microdilution broth method showed that these isolates were highly resistant to most antibiotics including carbapenems, cephalosporins, and β-lactam/β-lactamase inhibitors, but were susceptible to aztreonam, aminoglycosides, quinolones, tigecycline and colistin (Table S1). We suspected that these isolates produce Metallo-β-lactamase. Modified carbapenem inactivation method (mCIM) and EDTA-modified carbapenem inactivation method (eCIM) confirmed our suspicion.
We selected the first strain from four different specimens (strain KP22937 from blood; strain KP22866 from BALF; strain KP22877 from sputum; strain KP23025 from drainage fluid; GenBank accession number SAMN17245924-SAMN17245927) for genome sequencing using Illumina HiSeq 2500 sequencing platform, and then strain KP22937 was sequenced using nanopore sequencing method on MinION flow cells to obtain complete plasmid sequences. De novo assembly was conducted using SPAdes Genome Assembler v3.13.1 [5] and Unicycler [6]. Gene prediction was performed using Prokka 1.12 [7]. Genomic islands were predicted using IslandViewer 4 [8]. Antimicrobial resistance genes and plasmid replicon analyses were performed using ResFinder and PlasmidFinder tools via the CGE server (https://cge.cbs.dtu.dk/services/). Virulence genes were identified using the BIGSdb Klebsiella genome database (http://bigsdb.Pasteur.fr/klebsiella/klebsiella.html). Multilocus sequence types (MLST) and K type were determined using Kleborate 0.3.0 (https://github.com/katholt/Kleborate) and Kaptive tool [9], respectively.
The analysis showed that strain KP22937 belonged to ST65 and capsule serotype K2, and carried several hypervirulent determinants such as iucABCDiutA and iroBCDN, as well as a hypermucoviscous phenotype regulator gene rmpA. Mouse lethality assay was performed for assessing the in vivo virulence of strain KP22937. The result showed that all eight pathogen-free female BALB/c mice (6–8 weeks and 18–20 g) injected with KP22937 at a concentration of 105 CFU/ml died within 12 h, while mice injected with low-virulence clinical K. pneumoniae strain KPZ03 (GenBank accession number SAMN17245928) and PBS survived for over a week (Figure S1). The above results indicated that the ST65 K. pneumoniae strain KP22937 that caused infection in the lung transplant recipient was hvKp, which was consistent with a previous report [10].
Two plasmids were identified in K. pneumoniae strain KP22937. The plasmid pNDM22937, 45048 bp in size, belongs to the IncX3 group and did not harbour any resistance genes other than blaNDM-5. BLASTN search for GenBank showed that pNDM22937 was nearly identical to blaNDM-5-carrying plasmids previously reported in K. pneumonia of different ST types, and other species. Specifically, pNDM22937 showed 100% identity and coverage with plasmid pNDM5_020026 (CP034957) from E. coli SCEC020026, and 100% identity and 97% coverage with the plasmid pNDM_MGR194 (KF220657), which was a typical blaNDM-5-carrying plasmid recovered from a K. pneumoniae isolate in India (Figure 1B) [11]. The genetic environment of blaNDM-5 gene in pNDM22937 (ΔTn2-ΔIS3000-IS5-ΔISAba125-blaNDM-5-ble-trpF-dsbC-IS26-ΔumuD) was almost identical to pNDM_MGR194 and pNDM5_020026, except that part of the downstream region of IS3000 and upstream region of ISAba125 were deleted from pKP22937 (Figure 1C). Moreover, we found a genomic island region with 15 open reading frames (ORFs) in the plasmid pNDM22937 (Figure 1B). The first ORF shared 44.6% identity with an integrase from Serratia sp, and the last ORF in the genomic island was a twin-arginine translocation (TAT) pathway signal sequence domain protein. The blaNDM-5 gene and its flanking contents were also present in this region.
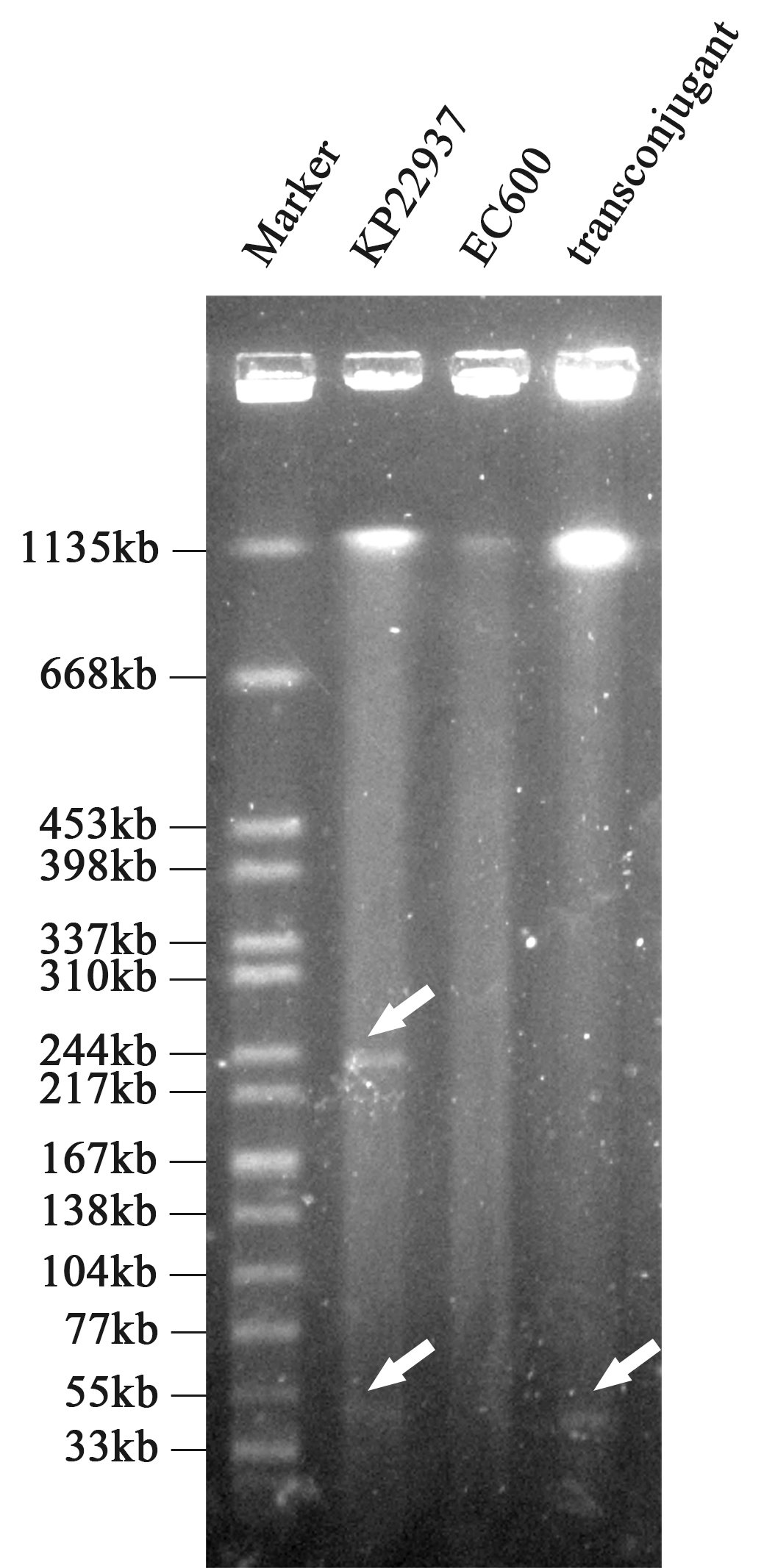
Several conjugal transfer genes were also identified in pNDM22937, such as virB4, virB8, virB9 and virD4 (Figure 1B). Conjugation experiment showed that the strain KP22937 transferred the plasmid carrying blaNDM-5 to the recipient E. coli EC600 at a frequency of 10−5 (transconjugant/recipient), PCR amplification and sequencing of the transconjugants and S1-PFGE experiment (Figure S2) confirmed successful transfer to the recipient, suggesting that blaNDM-5 was located on a self-transmissible plasmid, which could mediate dissemination of antibiotic resistance. The recipient E. coli EC600 was susceptible to all antibiotics, whereas the transconjugant displayed a similar antibiotic resistance phenotype to the strain KP22937 after obtaining the blaNDM-5-harbouring plasmid (Table S1). The fitness cost of the IncX3 plasmid harbouring blaNDM-5 was evaluated through growth kinetics assays. The results showed that the growth of the transconjugant was almost indistinguishable from that of E. coli EC600, revealing that the acquisition of IncX3 plasmid did not confer a fitness cost to the host. Accordingly, we hypothesized that the strain that caused postoperative infection in lung transplant patients in this study was produced by obtaining a self-transmissible IncX3 plasmid carrying blaNDM-5 gene by a hypervirulent ST65 K. pneumoniae strain.
In the absence of antibiotic pressure, the presence of resistance genes in plasmids will impose fitness costs on their host [12]. However, a recent in vitro study has shown that up to 75.9% (22/29) Enterobacteriaceae strains did not produce fitness costs after obtaining the IncX3 plasmid through conjugation test [13], which may facilitate the dissemination of the plasmid. Genomic islands are clusters of genes of probable horizontal origin in bacterial or archaeal genomes. A previous study revealed that genomic islands are a major driver of genome evolution, and they can enhance the fitness of bacteria within a niche [14]. In this study, the blaNDM-5 gene and its flanking content were located in the genomic island region of plasmid pNDM22937, which may contribute to the lower fitness cost of K. pneumoniae strain KP22937. On all accounts, the lower fitness cost may partly explain the rapid dissemination of blaNDM-5 among Enterobacteriaceae strains.
Another plasmid of K. pneumoniae KP22937, pVir22937, which was 234457 bp in length, showed 99–100% identity and 91–92% coverage with two classical hypervirulent plasmids pLVPK (NC_005249) and pK2044 (NC_006625) (Figure 1B). Virulence gene clusters iucABCDiutA and iroBCDN, and rmpA gene were located on this plasmid. This plasmid was not self-transmissible, and had no conjugal transfer gene.
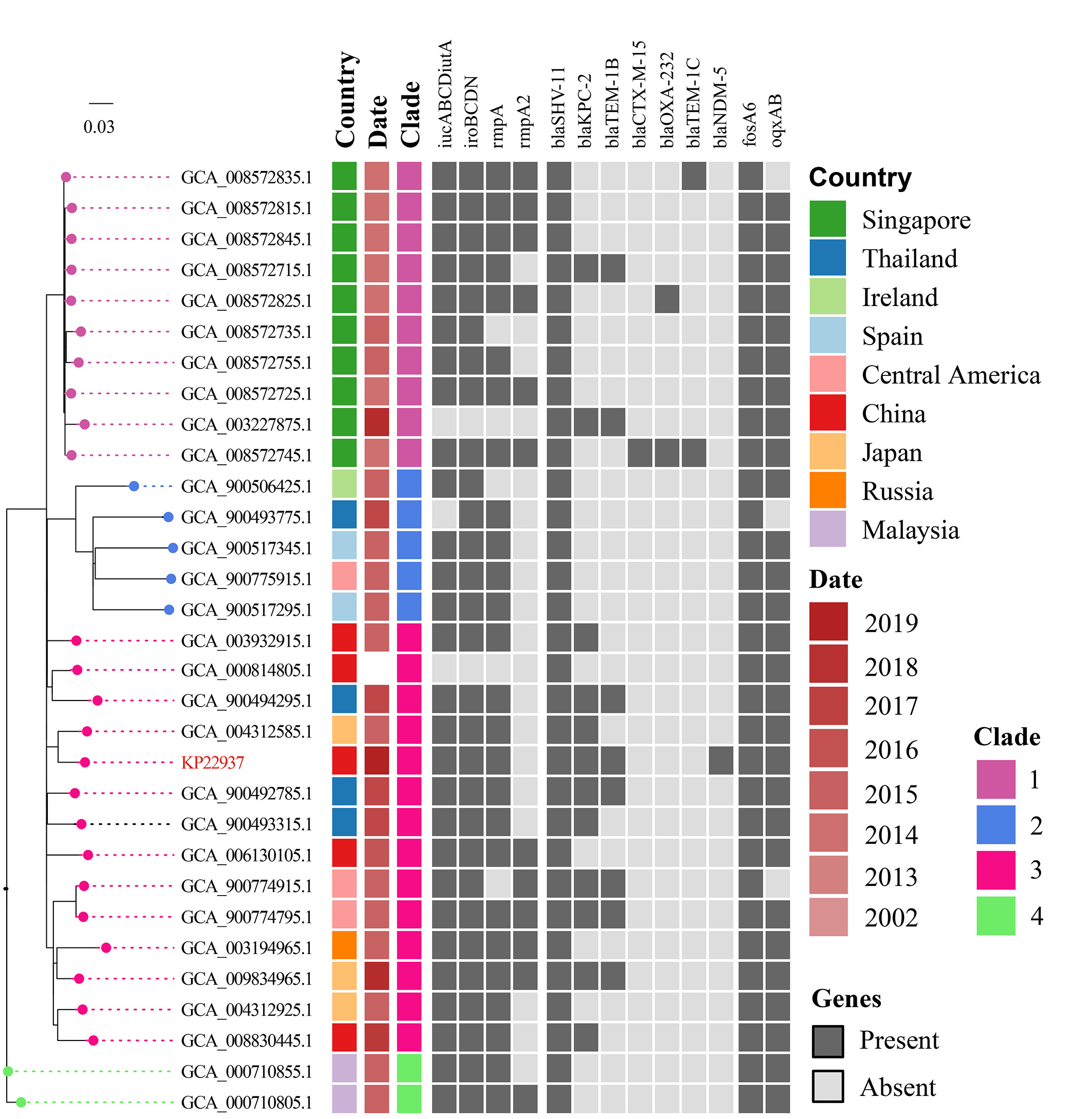
In the present study, we reported an NDM-5-producing hypervirulent K. pneumoniae ST65 strain. To investigate whether this clone has been previous reported, we retrieved 9157 genomic sequences of K. pneumoniae with different assembly levels from the NCBI genome database on July 1, 2020. MLST analysis of these isolates was performed to include only ST65 clone. Eventually, 30 genomic sequences of ST65 K. pneumoniae were involved. Analysis of resistance and virulence determinants showed that the blaKPC-2 gene was present in many ST65 K. pneumoniae strains, but the occurrence of blaNDM-5 gene in ST65 K. pneumoniae strain was comparatively rare. This study was the first report that found the blaNDM-5 gene in a clinical ST65 K. pneumoniae strain (Figure S3).
In summary, this study reported a fatal infection in a post-transplant patient caused by an NDM-5-producing hypervirulent K. pneumoniae ST65 clone. The blaNDM-5 gene was located on a self-transmissible IncX3 plasmid, which could be transferred to E. coli without significant fitness costs. The current finding raises concerns about horizontal spread of blaNDM-5 gene mediated by IncX3 plasmid, where hypervirulent K. pneumonia strains are also involved.
Supplementary Material
{kind=link}
{kind=link}
{kind=link}
Acknowledgements
We thank Chunlei Wang, Yudi Xia, Yue Zhu, Yufei Ye, Haibo Li, Xiaohui Zou and Lingxiao Sun for their assistance.
Funding Statement
This work was supported by the National Key Research and Development Program of China [grant number 2017YFC1309300, 2017YFC1309301, 2018YFC1200100 and 2018YFC1200102]; the Scientific Research Project of China-Japan Friendship Hospital [grant number 2018-2-QN-23] and the CAMS Innovation Fund for Medical Sciences [grant number CIFMS 2018-I2M-1-003].
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- 1.Hornsey M, Phee L, Wareham DW.. A novel variant, NDM-5, of the New Delhi metallo-beta-lactamase in a multidrug-resistant Escherichia coli ST648 isolate recovered from a patient in the United Kingdom. Antimicrob Agents Chemother. 2011 Dec;55(12):5952–5954. doi: 10.1128/AAC.05108-11. PubMed PMID: 21930874; PubMed Central PMCID: PMCPMC3232805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rojas LJ, Hujer AM, Rudin SD, et al. . NDM-5 and OXA-181 beta-lactamases, a significant threat continues to spread in the americas. Antimicrob Agents Chemother. 2017 Jul;61(7):e00454–17. doi: 10.1128/AAC.00454-17. PubMed PMID: 28461314; PubMed Central PMCID: PMCPMC5487671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Yuan Y, Li Y, Wang G, et al. . Bla NDM-5 carried by a hypervirulent Klebsiella pneumoniae with sequence type 29. Antimicrob Resist Infect Control. 2019;8:140. doi: 10.1186/s13756-019-0596-1. PubMed PMID: 31452874; PubMed Central PMCID: PMCPMC6701021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Shen Z, Gao Q, Qin J, et al. . Emergence of an NDM-5-producing hypervirulent Klebsiella pneumoniae sequence type 35 strain with chromosomal integration of an integrative and conjugative element, ICEKp1. Antimicrob Agents Chemother. 2020;64(1):e01675–19. doi: 10.1128/AAC.01675-19. PubMed PMID: 31611359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bankevich A, Nurk S, Antipov D, et al. . SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012 May;19(5):455–477. doi: 10.1089/cmb.2012.0021. PubMed PMID: 22506599; PubMed Central PMCID: PMCPMC3342519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wick RR, Judd LM, Gorrie CL, et al. . Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput Biol. 2017;13(6):e1005595. doi: 10.1371/journal.pcbi.1005595. PubMed PMID: 28594827; PubMed Central PMCID: PMCPMC5481147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014 Jul 15;30(14):2068–2069. doi: 10.1093/bioinformatics/btu153. PubMed PMID: 24642063. [DOI] [PubMed] [Google Scholar]
- 8.Bertelli C, Laird MR, Williams KP, et al. . Islandviewer 4: expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017 Jul 3;45(W1):W30–W35. doi: 10.1093/nar/gkx343. PubMed PMID: 28472413; PubMed Central PMCID: PMCPMC5570257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wyres KL, Wick RR, Gorrie C, et al. . Identification of Klebsiella capsule synthesis loci from whole genome data. Microb Genom. 2016 Dec;2(12):e000102. doi: 10.1099/mgen.0.000102. PubMed PMID: 28348840; PubMed Central PMCID: PMCPMC5359410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lev AI, Astashkin EI, Kislichkina AA, et al. . Comparative analysis of Klebsiella pneumoniae strains isolated in 2012–2016 that differ by antibiotic resistance genes and virulence genes profiles. Pathog Glob Health. 2018;112(3):142–151. doi: 10.1080/20477724.2018.1460949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Krishnaraju M, Kamatchi C, Jha AK, et al. . Complete sequencing of an IncX3 plasmid carrying blaNDM-5 allele reveals an early stage in the dissemination of the blaNDM gene. Indian J Med Microbiol. 2015 Jan-Mar;33(1):30–38. doi: 10.4103/0255-0857.148373. PubMed PMID: 25559999. [DOI] [PubMed] [Google Scholar]
- 12.Andersson DI, Hughes D.. Antibiotic resistance and its cost: is it possible to reverse resistance? Nat Rev Microbiol. 2010 Apr;8(4):260–271. doi: 10.1038/nrmicro2319. PubMed PMID: 20208551. [DOI] [PubMed] [Google Scholar]
- 13.Ma T, Fu J, Xie N, et al. . Fitness cost of blaNDM-5-carrying p3R-IncX3 plasmids in wild-type NDM-free enterobacteriaceae. Microorganisms. 2020 Mar 7;8(3):377. doi: 10.3390/microorganisms8030377. PubMed PMID: 32156014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dobrindt U, Hochhut B, Hentschel U, et al. . Genomic islands in pathogenic and environmental microorganisms. Nat Rev Microbiol. 2004 May;2(5):414–424. doi: 10.1038/nrmicro884. PubMed PMID: 15100694. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.