

Abstract
Purpose
Papillary thyroid cancer (PTC) has a high incidence of BRAFV600E mutation. The purpose of this study was to evaluate the potential relationship between thyroiditis and BRAFV600E mutation status in patients with PTC. We investigated how a selective inhibitor of BRAFV600E PLX4032 affects the proliferation and inflammatory cytokine levels of thyroid cancer.
Methods
Two thyroid cancer cell lines TPC1 and 8505C were treated with PLX4032, an analysis was done on cell growth, cell cycle, the degree of apoptosis, and levels of inflammatory cytokines. To identify the functional links of BRAF, we used the STRING database.
Results
Docking results illustrated PLX4032 blocked the kinase activity by exclusively binding on the serine/threonine kinase domain. STRING results indicated BRAF is functionally linked to mitogen-activated protein kinase. Both cell lines showed a dose-dependent reduction in growth rate but had a different half maximal inhibitory concentration value for PLX4032. The reaction to PLX4032 was more sensitive in the 8505C cells than in the TPC1 cells. PLX4032 induced a G2/M phase arrest in the TPC1 cells and G0/G1 in the 8505C cells. PLX4032 induced apoptosis only in the 8505C cells. With PLX4032, the TPC1 cells showed decreased levels of vascular endothelial growth factor, granulocyte-macrophage colony-stimulating factor, chemokine (C-C motif) ligand 2/monocyte chemoattractant protein 1, whereas the 8505C cells showed significantly decreased levels of IL-8, serpin E1/plasminogen activator inhibitor-1, and matrix metalloproteinase (MMP)-3.
Conclusion
PLX4032 was cytotoxic in both TPC1 and 8505C cells and induced apoptosis. In the 8505C cells, inflammatory cytokines such as IL-8 and MMP-3 were down-regulated. These findings suggest the possibility that the BRAFV600E mutation needs to target inflammatory signaling pathways in the treatment of thyroid cancer.
Keywords: BRAFV600E, Cytokines, Thyroid neoplasms, Vemurafenib
INTRODUCTION
Korea and other parts of the world have seen an increase in the incidence of thyroid carcinoma [1,2]. It is also known that thyroid cancer has a close link with thyroiditis [3,4,5]. An epidemiological association between Hashimoto's thyroiditis and thyroid cancer has been identified. The progression of thyroid cancer is likely to be accelerated with concomitant inflammation. IL-6 and IL-8 are especially significant in systemic and chronic inflammatory conditions [4]. IL-6 plays a major role in thyroid cancer development. In papillary thyroid cancer (PTC) patients, the IL-6 level in cancer tissue correlates with the aggressiveness of the cancer [6,7]. The cytokine levels of IFN-alpha and IL-8 showed a strong positive step association in thyroid cancer patients [7]. Inflammatory status boosts cytokine secretion such as interleukins and TNF-alpha [7]. These cytokines facilitate the proliferation and metastasis of thyroid cancer. Due to the prognosis of thyroid cancer, 2 cytokines, IL-6 and IL-8, are considered to be associated with thyroid cancer [7]. It is also recognized that the matrix metalloproteinase (MMP) molecule participates in cancer metastasis [8]. It is involved in the pathogenesis of many diseases because of its complex regulatory activity [8]. Some studies have shown that MMPs can be induced by BRAFV600E and a positive association between BRAFV600E and MMP-9 and MMP-2 expressions has also been documented, both associated with tumor extrathyroidal extension [8].
BRAFV600E mutation is much higher (73%–86%) in Korea than in Western countries (29%–69%), and is associated with larger tumor size, lymphovascular invasion or metastases, and mortality in different human cancers [1,9]. The valine-to-glutamic acid substitution at amino acid 600 (BRAFV600E) strongly enhances BRAF kinase activity. The phosphorylation activity on ERK1/2 is 480-fold higher than the wild type BRAF or other BRAF mutants [10].
BRAFV600E mutation is also the most common genetic lesion in PTC, which is seen in 40% of cases [10]. The mutation in BRAFV600E has a strong correlation with PTC. By adding a negatively charged residue adjacent to the phosphorylation site at T598 and mimicking phosphorylation at Thr598, and mimicking phosphorylation at Thr598 and Ser601 residues with the effect of increasing ERK1/2 photography, the V600E mutation significantly improves BRAF kinase operation [10].
In the past 2 years, with the approval of new treatments, including pathway inhibitor drugs and a monoclonal antibody, the clinical landscape has been changed. Approved by the U.S. Food and Drug Administration in 2011, vemurafenib (PLX4032) is a selective kinase inhibitor for the most common mutant form of BRAF kinase (BRAFV600E) to prevent melanoma transformation and proliferation. It is also a highly selective inhibitor of BRAF kinase activity and has a half maximal inhibitory concentration (IC50) value in the nM range (60–450 nM) against V600E-mutant BRAF [11].
In this study, we observed how the BRAF mutation affects the progression of thyroid cancer, we focused on selective BRAFV600E kinase inhibitor and its responses in the progression and cytokine levels of thyroid cancer.
METHODS
Molecular docking
The renowned software Autodock tools (ADT) 1.5.6 (Scripps Research Institute, La Jolla, CA, USA) was used for docking analysis. Protein structures for BRAF wild type (PDB: 4H58) and BRAFV600E (PDB: 4FK3) were used from a protein data bank (www.rcsb.org), and crystallized inhibitors were cleared from receptor molecules and energy minimized for reuse. The PLX4032 structure was obtained from PubChem with ID:42611257 and prepared for docking. For molecular docking, default parameters were used and generated 100 confirmations for robustness. We then clustered the structures based on root mean square deviation of the confirmations, and the largest group was selected as the best fit model.
Functional analysis using STRING
The search tool for the retrieval of interacting genes/proteins (STRING, https://string-db.org) is a web-based tool that provides a critical assessment and integration of protein–protein interactions. In this study, STRING was applied to determine the functional protein partners of each BRAF member. Gene fusion, and coexpression were taken into consideration as active protein–protein interaction sources. The protein–protein network was constructed using both known and predicted interactions with no network clustering.
Cell culture
We chose 8505C mutant cell line derived from an anaplastic thyroid cancer; and, for BRAF, wild type TPC1 cell line was used, derived from a PTC with a wild type BRAF [12,13]. The study was approved by Institutional Review Board at Konkuk University Medical Center, Seoul, Korea (No. 2020-12-062). Unless otherwise specified, all reagents were purchased from Sigma (St. Louis, MO, USA). The 8505C and TPC1 cells were cultured in Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS). For the culture, 5 × 105 cells were cultured at 37℃ in a humidified atmosphere containing 5% CO2.
Assessment of cell growth following PLX4032 treatment
For each cell line, 5 × 105 cells were seeded in DMEM with 10% FBS. After 24 hours, the cells were washed with phosphate-buffered saline (PBS) and cultured in fresh media. PLX4032 was added at 0.1, 1, 5, or 10 µM for 72 hours. The cells were then counted in a Neubauer chamber after a period of incubation with 0.4% trypan blue dye.
Cell cycle analysis
After being treated with 10 µM of PLX4032 for 72 hours, the cells were harvested, fixed in 70% ethanol for 1 hour, and washed with PBS. The cells were then treated with 100 µg/mL of RNase A for 1 hour at 37℃ and stained with 10 µg/mL of propidium iodide. Flow cytometry was performed in triplicate for each experiment with a FACS caliber system (BD Biosciences, San Jose, CA, USA).
Apoptosis analysis with annexin V staining
Annexin V staining was performed in accordance with the manufacturer's protocol (BD PharMingen, San Diego, CA, USA). Trypsin-EDTA was used to obtain single-cell suspensions. The cells were subjected to centrifugation, washed twice with cold PBS, and resuspended in a binding buffer (10-mM HEPES, pH 7.4; 150-mM NaCl; 5-mM KCl; 1-mM MgCl2; and 1.8-mM CaCl2) at a concentration of 1 × 106 cells/mL. An aliquot (100 µL) of the solution containing 1 × 105 cells was transferred to a 5 mL culture tube and supplemented with 5 µL of annexin V-fluorescein isothiocyanate (FITC) and 5 µL of propidium iodide. The cells were vortexed and then incubated for 15 minutes at room temperature (25℃) in the dark. A 400-µL sample of 1× binding buffer was added to each tube. Finally, flow cytometry was performed with a FACS caliber system within 1 hour.
Secreted protein (growth factor, cytokine, matrix metalloproteinase) analysis
A sample of 5 × 105 TPC1 cells, which were seeded in DMEM with 10% FBS and 10 µM of PLX4032, was added and the cells were incubated for a further 72 hours. The 8505C cells were cultured under the same conditions with 5 µM of PLX4032. The media were collected and centrifuged, and only the upper layer of the media without any cell pellets was harvested. The secreted protein was analyzed with the commercially available Human Multianalyte Profiling Assay Base Kit A (catalog No. LMP000, R&D Systems Inc., Minneapolis, MN, USA). The levels of secreted protein were measured with a Bio-Plex 200 system (Bio-Rad, Hercules, CA, USA).
Statistical analysis
All data were compiled from a minimum of 3 replicate experiments. The data for the statistical analysis were expressed as the mean plus or minus the standard error. The results from a Student t-test of the treated cells were compared with those of the control cells using Excel software (Microsoft, Redmond, WA, USA). A P-value of less than 0.05 was considered statistically significant.
RESULTS
Subsection
Binding patterns of PLX4032 on BRAF
To illustrate the differential binding effects of PLX4032 between wild type BRAF and mutated BRAF-V600E, we employed the molecular docking approach. For comparative analysis, we have docked the PLX4032 into BRAF wild type and BRAFV600E using the ADT 1.5.6 docking software. Interestingly, observed that PLX4032 showed more promiscuity toward mutated BRAF than wild type. When explored further, the ligand deeply penetrated the hydrophobic core of the mutated BRAF and precisely blocked the ATP binding site (483K), which is important for phosphorylation activity; it is not observed in wild type complex. Furthermore, V600E and PLX4032 complex possessed more hydrogen bonds (T529, C532, D594, F595, and G596) compared to wild type (N580, I526, and W530). PLX4032 showed similarity in not binding to active site residue D576 of both variants but showed efficacy toward mutated types by binding on the ATP binding site (Fig. 1).
Fig. 1. Docking for PLX4032 in wild type BRAF and V600E mutant binding patterns of PLX4032 on BRAF. Molecular docking of PLX4032 with BRAF wild type and V600E. (A) The first panel illustrates PLX4032 docked into the hydrophobic core of wild type BRAF showing hydrogen, hydrophobic, and cation interactions. (B) The second panel indicates the best-docked confirmation of PLX4032 into BRAFV600E along with binding interactions representation.

BRAF functional interaction
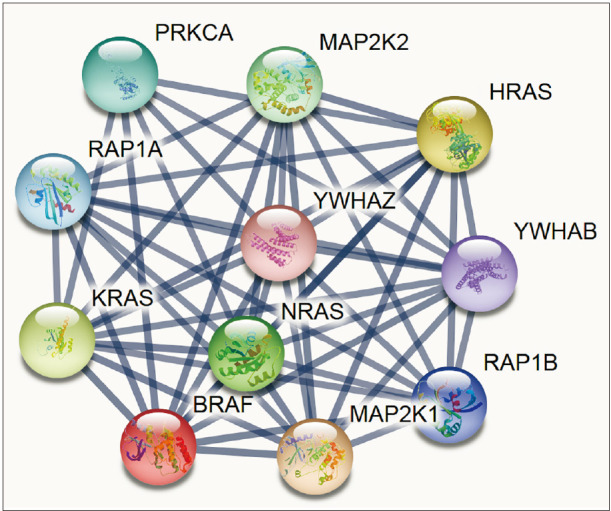
To identify the functional interaction of BRAF, we used STRING, an online biological database and web-based tool, to analyze known and predicted protein–protein interactions. Here, we tried to gain some insights at the system-level of the functional interactions of BRAF with other closely related proteins via a protein–protein interaction network. The database was individually queried for BRAF with Homo sapiens as organism, where the respective functional link was obtained as the output. Whereas each node in functional network represents all the proteins produced by a single, protein-coding gene locus, an edge represents protein association; however, does not necessarily represent a physical binding between the pair of genes. Rather, associated proteins jointly contribute to a shared function. We selected knowledge-based functional protein partners of BRAF based on curated databases. The top 10 predicted proteins of BRAF (with the corresponding gene names) included PRKCA, MP2K2, HRAS, YWHAB, RAP1B, MAP2K1, NRAS, KRAS, RAP1A, and YWHAZ. These interacting genes are basically involved in kinase activity and activated by the extracellular cytokine and growth factor binding thereby entering the mitogen-activated protein kinase (MAPK) signal transduction pathway through RAS and RAF activation, especially MAP2K1, MP2K2 key roles in the MAPK/ERK cascade function—a crucial complex for major biological activities like cell growth, survival, and differentiation [14]. YWHAB and YWHAZ are the tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation proteins and housekeeping genes linked with protein kinase activity; and, in phosphorylated form, interacts with Akt-signaling and MAPK pathway [15]. Altogether, all the BRAF interacting genes are activated by extracellular cytokine activity and shown broadly in various cancers (Fig. 2).
Fig. 2. Predicting the interaction network of BRAF using STRING (https://string-db.org). Interacting nodes are displayed in circles, obtained using STRING. Predicted functional partners of BRAF are shown, taking into consideration functional links, coexpression, colocalization, genetic interactions, pathway, physical interactions, predicted, and shared protein domains.
PLX4032-induced inhibition of the growth of thyroid cancer in a dose-dependent manner
PLX4032 is known as an anticancer drug that targets wild and mutant BRAF. We selected the TPC1 and 8505C cell lines so that we could see how PLX4032 affects thyroid cancer. Seventy-two hours of treatment with PLX4032 inhibits the growth of both cell lines in a dose-dependent manner (Fig. 3). The results reveal different IC50 doses for PLX4032 in the 2 cell lines. The TPC1 cells show an IC50 value of 10 µM for PLX4032, whereas the 8505C cells show IC50 values of 5 µM for PLX4032. These IC50 doses were used in growth assays to observe the cell cycle and apoptosis of both cell lines.
Fig. 3. Growth rate of cells after PLX4032 treatment. TPC1 and 8505C cells cultured in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum. (A) TPC1 cells were treated for 72 hours with 100 nM, 1 µM, and 10 µM of DMSO (CTL) or PLX4032. (B) 8505C cells were treated for 72 hours with 100 nM, 1 µM, 5 µM, and 10 µM of DMSO (CTL) or PLX4032. Relative cell survival rate is shown as percentage of survival vs. control cells after treatment with PLX4032. Values presented here are mean values and standard errors from at least 3 independent experiments. Square box indicates the IC50 value of the TPC1 and 8505C cells. DMSO, dimethyl sulfoxide; CTL, cytotoxic T lymphocyte; IC50, half maximal inhibitory concentration. *P < 0.05, **P < 0.01 vs. control.

PLX4032-induced cell cycle arrest
Previous studies have reported that PLX4032 leads to different responses in melanoma and thyroid carcinoma cells through apoptosis and cell cycle arrest [11,16,17]. We investigated the cell cycle after treatment with these drugs. The cell cycle analysis showed that the TPC1 cells responded in a different manner to the 8505C cells. In the TPC1 cells, there was a 6% increase in the G2/M phase with 10 µM of PLX4032. In the 8505C cells, there was a 14% increase in the G0/G1 phase with 5 µM of PLX4032 (Fig. 4).
Fig. 4. Cell cycle analysis after PLX4032 treatment. (A) TPC1 cells were treated for 72 hours with 10 µM of DMSO (CTL) or PLX4032. (B) 8505C cells were treated for 72 hours with 5 µM of DMSO (CTL) or PLX4032. Cell cycle was analyzed by flow cytometry after DNA staining with propidium iodide. Data represent percentage of cells at each stage of cell cycle. Data are from representative experiment (out of a total of 3 experiments). DMSO, dimethyl sulfoxide; CTL, cytotoxic T lymphocyte.

PLX4032-induced apoptosis
We observed apoptosis through propidium iodide and annexin V staining to determine how the growth inhibition affects the top of the cell cycle arrest after treatment with PLX4032. The apoptotic responses to each drug were different. The 1-µM sample of PLX4032 induced apoptosis in 7% of the 8505C cells; however, the same sample induced no apoptosis in the TPC1 cells (Fig. 5). The propidium iodide and annexin V staining showed that PLX4720 failed to induce any apoptosis in either of the cell lines (Fig. 3).
Fig. 5. Apoptosisanalysisafter PLX4032 treatment. (A) TPC1 cells were treated for 72 hours with 10 µM of DMSO (CTL) or PLX4032. (B) 8505C cells were treated for 72 hours with at 5 µM of DMSO (CTL) or PLX4032. The cell cycle was analyzed by flow cytometry after DNA staining with propidium iodide. All cells were stained with FITC-conjugated annexin V in buffer containing propidium iodide and analyzed by flow cytometry. For each treatment, percentage of viable cells is shown in lower left quadrant, which indicates low levels of annexin V and propidium iodide. Data are from representative experiment (out of a total of 3 experiments). DMSO, dimethyl sulfoxide; CTL, cytotoxic T lymphocyte; FITC, fluorescein isothiocyanate.

Analysis of secretory proteins after treatment with PLX4032
We investigated how PLX4032 affects the growth factors, cytokines, lipid metabolism factors, and MMPs. Specifically, we compared the basal levels with the stimulated levels of these factors in the TPC1 cells and 8505C cells after treatment with PLX4032. Some of these factors were selected if the cell expression level exceeded 100 pg/mL. The TPC1 cells expressed high levels of IL-6 (Fig. 6A), vascular endothelial growth factor (VEGF) (Fig. 7A), granulocyte-macrophage colony-stimulating factor (GM-CSF) (Fig. 7A), serpin E1/plasminogen activator inhibitor 1 (PAI-1) (Fig. 8A), chemokine (C-C motif) ligand 2 (CCL2)/monocyte chemoattractant protein 1 (MCP-1) (Fig. 8A), and MMP-2 (Fig. 9A). In contrast, the 8505C cells expressed high levels of IL-8 (Fig. 6B), MMP-1 (Fig. 9B), and MMP-3 (Fig. 9B). The levels of these selected factors are indicative of the effects of PLX4032. After treatment with PLX4032, the levels of IL-8, VEGF, GM-CSF, CCL2/MCP-1, MMP-2, and MMP-3 were reduced in the TPC1 cells (Figs. 6A, 7A, 8A, and 9A); and the levels of IL-8, serpin E1/PAI-1, and MMP-3 were reduced in the 8505C cells (Figs. 6A, 7A, 8A, and 9B). The CCL2/MCP-1 level was the only factor that showed reduction in the 8505C cells after the PLX4032 treatment.
Fig. 6. Expression of IL-6 and IL-8 after PLX4032 treatment. (A) TPC1 cells were treated for 72 hours with 10 µM of DMSO (CTL) or PLX4032. (B) 8505C cells were treated for 72 hours with 5 µM of DMSO (CTL) or PLX4032. Density levels of IL-6 and IL-8 were measured with Bio-Plex 200 system (Bio-Rad, Hercules, CA, USA). DMSO, dimethyl sulfoxide; CTL, cytotoxic T lymphocyte. *P < 0.05, **P < 0.01 vs. control.

Fig. 7. Expression of VEGF and GM-CSF after PLX4032 treatment. (A) TPC1 cells were treated for 72 hours with 10 µM of DMSO (CTL) or PLX4032. (B) 8505C cells were treated for 72 hours with 5 µM of DMSO (CTL) or PLX4032. Density levels of VEGF and GM-CSF were measured with Bio-Plex 200 system (Bio-Rad, Hercules, CA, USA). VEGF, vascular endothelial growth factor; GM-CSF, granulocyte-macrophage colony-stimulating factor; DMSO, dimethyl sulfoxide; CTL, cytotoxic T lymphocyte. *P < 0.05, **P < 0.01 vs. control.

Fig. 8. Expression of CCL2/MCP-1 and serpin E1/PAI-1 after PLX4032 treatment. (A) TPC1 cells were treated for 72 hours with 10 µM of DMSO (CTL) or PLX4032. (B) 8505C cells were treated for 72 hours with 5 µM of DMSO (CTL) or PLX4032. Density levels of CCL2/MCP-1 and serpin E1/PAI-1 were measured with Bio-Plex 200 system (Bio-Rad, Hercules, CA, USA). CCL2, chemokine (C-C motif) ligand 2; MCP-1, monocyte chemoattractant protein 1; PAI-1, plasminogen activator inhibitor 1; DMSO, dimethyl sulfoxide; CTL, cytotoxic T lymphocyte. *P < 0.05, **P < 0.01 vs. control.

Fig. 9. Expression of MMP-1, 2, and 3 after PLX4032 treatment. (A) TPC1 cells were treated for 72 hours with 10 µM of DMSO (CTL) or PLX4032 and PLX4720. (B) 8505C cells were treated for 72 hours with 5 µM of DMSO (CTL) or PLX4032. Density levels of MMP-1, -2, and -3 were measured with Bio-Plex 200 system (Bio-Rad, Hercules, CA, USA). MMP, matrix metalloproteinase; DMSO, dimethyl sulfoxide; CTL, cytotoxic T lymphocyte. *P < 0.05, **P < 0.01 vs. control.

DISCUSSION
Approval of vemurafenib (PLX4032) was a milestone in thyroid cancer therapy which targets the altered and constantly activated BRAFV600E. Our results showed that a BRAF is functionally linked to the MAPK pathway which encodes genes like MMP and IL-8. A kinase in the MAPK pathway is genetically modified in approximately 37%–59% of all melanomas [7,8]. Here, most substantial differences in the sensitivity of wild type and mutant populations were detected for acute PLX4032 treatment. We compared 2 thyroid cancer cell lines (8505C, TPC1) that differed in their oncogene profile (BRAFV600E and BRAF wild type). With these results, we confirmed that PLX4032 significantly affects the inhibition of growth, the cell cycle arrest, and the downregulation of secreted proteins in thyroid cancer.
As PLX4032 targets BRAF mutation and effectively inhibits cellular growth in various cancer cells with the BRAF mutation [11,16,18], our results showed similar results. PLX4032 induced growth inhibition in thyroid cancer cell lines but had different effects on the cell cycle and apoptosis. PLX4032 induced a G2/M phase arrest but showed no apoptotic effects in TPC1 cells. However, PLX4032 induced a G0/G1 phase arrest and apoptosis in 8505C cells. These results are thought to be attributed to the presence of the BRAF mutation, which is found only in the 8505C cells. These findings are very interesting, and the presence of a BRAF mutation might be considered the target of an anti-apoptotic pathway.
Several lines of evidence have recently indicated that inflammation and carcinogenesis may be strongly associated with the progression of a tumor [19,20]. An estimated 20% of all tumors arise from inflammatory conditions such as chronic infections or autoimmune diseases [20,21,22]. Whether the inflammation can influence the progression and aggressiveness of advanced thyroid cancer is still open to debate [3,23].
Our evaluation suggests that PLX4032 can target specific functions of secretory proteins related to inflammatory, angiogenic, and metastatic processes of advanced thyroid carcinomas. The important secretory proteins for tumor progression and cell-to-cell interactions of various cancers are inflammatory cytokines [24]. Previous research indicating that resistance of dacarbazine-conditioned BRAFV600E cells to PLX4032 is mainly due to the re-activation of MAPK pathway by autocrine IL-8 cytokine stimulation [10].
In our experiments, PLX4032 significantly reduced IL-8 and serpin E1/PAI-1 in the BRAF mutant cell line 8505C. Following exposure to BRAF inhibitors, mice with BRAF mutant melanoma xenografts show a significant reduction in the plasma concentration of IL-8 [23,25]. Advances in proteomics suggest that IL-8 has a diagnostic value between benign and malignant thyroid nodules [26]. An orthotopic model of anaplastic thyroid carcinoma in athymic nude mice showed an increase in IL-8 expression in immunohistochemical staining [27]. IL-8 also increases the population of cancer stem cells and strengthens drug resistance in breast cancer [28,29].
Many cancers could be easily treated if they did not metastasize to other organs or distant organs. MMPs play a key role in the metastasis of tumor cells [30]. We observed that PLX4032 reduced MMP-3. These results indicate that PLX4032 may have an inhibitory effect on the metastasis of thyroid cancer.
The resistance to BRAF inhibitors can occur by various genetic and nongenetic alterations [10,18]. Common genetic alterations that lead to BRAF inhibitor resistance include NRAS mutation, MAP2K1 and MAP2K2 amplification, and mutation in the PI3K pathway [18].
Whether selective BRAF inhibitors will eventually find a role in thyroid cancer therapy is difficult to predict [18]. An appealing feature of these findings is the striking selectivity for the mutant form of BRAF [18]. Increasingly, a patient's tumor genotype will come to influence therapeutic decision making in advanced thyroid cancer [18]. Lessons learned from the highly selective clinical activity of drugs such as trastuzumab (Herceptin, Genentech, Inc., South San Francisco, CA, USA) in patients with Her2/Neu amplified breast cancer, and erlotinib (Tarceva, Genentech, Inc.) in epidermal growth factor receptor mutated lung cancer will help to inform clinical trial design in thyroid cancer [18].
Taken together, the BRAFV600E inhibitor PLX4032 starts to inhibit proliferation, reduces the levels of secretory proteins such as IL-8 and MMP-3 in thyroid cancer cell lines, and induces apoptosis in thyroid cancer cell lines. The impressive results in cell lines further suggest that this therapeutic treatment achieves great results for patients with thyroid cancer, and may further benefit from additional combination with emerging immunotherapies.
Going forward, it will be important to determine the effect of this treatment on metastasized thyroid cancer, and, over an extended period, to elucidate the genetic or epigenetic contributions leading to variable responses to treatment and to further characterize the immune response to treatment. These findings now show high potential for clinical impact on patients with BRAFV600E-positive thyroid cancer, who currently lack an effective treatment. Further study is needed to determine how PLX4032 is related to secretory proteins in thyroid cancer. Future in vivo studies should be done to evaluate the therapeutic positive results by the inhibiting of cytokine secretion in thyroid tumor microenvironments; this can be an alternative therapeutic strategy.
Footnotes
Fund/Grant Support: This paper was supported by Konkuk University in 2018.
Conflict of Interest: No potential conflict of interest relevant to this article was reported.
- Conceptualization: KSP.
- Formal Analysis: MS, EYY.
- Investigation: TYJ, HRR, AH, WSP, SYP.
- Methodology: SEN, YBY, JHY, JBK.
- Project Administration: KSP, YKY.
- Writing — Original Draft: MS, KSP.
- Writing — Review & Editing: YKY, WSP, SYP, EYY, TYJ, HRR, AH, SEN, YBY, JHY, JBK.
References
- 1.Won YJ, Sung J, Jung KW, Kong HJ, Park S, Shin HR, et al. Nationwide cancer incidence in Korea, 2003–2005. Cancer Res Treat. 2009;41:122–131. doi: 10.4143/crt.2009.41.3.122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Jemal A, Siegel R, Ward E, Murray T, Xu J, Thun MJ. Cancer statistics, 2007. CA Cancer J Clin. 2007;57:43–66. doi: 10.3322/canjclin.57.1.43. [DOI] [PubMed] [Google Scholar]
- 3.Larson SD, Jackson LN, Riall TS, Uchida T, Thomas RP, Qiu S, et al. Increased incidence of well-differentiated thyroid cancer associated with Hashimoto thyroiditis and the role of the PI3k/Akt pathway. J Am Coll Surg. 2007;204:764–773. doi: 10.1016/j.jamcollsurg.2006.12.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Zhang L, Li H, Ji QH, Zhu YX, Wang ZY, Wang Y, et al. The clinical features of papillary thyroid cancer in Hashimoto's thyroiditis patients from an area with a high prevalence of Hashimoto's disease. BMC Cancer. 2012;12:610. doi: 10.1186/1471-2407-12-610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Teng W, Shan Z, Teng X, Guan H, Li Y, Teng D, et al. Effect of iodine intake on thyroid diseases in China. N Engl J Med. 2006;354:2783–2793. doi: 10.1056/NEJMoa054022. [DOI] [PubMed] [Google Scholar]
- 6.Kobawala TP, Trivedi TI, Gajjar KK, Patel DH, Patel GH, Ghosh NR. Significance of interleukin-6 in papillary thyroid carcinoma. J Thyroid Res. 2016;2016:6178921. doi: 10.1155/2016/6178921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kobawala TP, Patel GH, Gajjar DR, Patel KN, Thakor PB, Parekh UB, et al. Clinical utility of serum interleukin-8 and interferon-alpha in thyroid diseases. J Thyroid Res. 2011;2011:270149. doi: 10.4061/2011/270149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Stetler-Stevenson WG. The role of matrix metalloproteinases in tumor invasion, metastasis, and angiogenesis. Surg Oncol Clin N Am. 2001;10:383–392. [PubMed] [Google Scholar]
- 9.Kebebew E, Weng J, Bauer J, Ranvier G, Clark OH, Duh QY, et al. The prevalence and prognostic value of BRAF mutation in thyroid cancer. Ann Surg. 2007;246:466–470. doi: 10.1097/SLA.0b013e318148563d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nucera C, Lawler J, Parangi S. BRAF(V600E) and microenvironment in thyroid cancer: a functional link to drive cancer progression. Cancer Res. 2011;71:2417–2422. doi: 10.1158/0008-5472.CAN-10-3844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sala E, Mologni L, Truffa S, Gaetano C, Bollag GE, Gambacorti-Passerini C. BRAF silencing by short hairpin RNA or chemical blockade by PLX4032 leads to different responses in melanoma and thyroid carcinoma cells. Mol Cancer Res. 2008;6:751–759. doi: 10.1158/1541-7786.MCR-07-2001. [DOI] [PubMed] [Google Scholar]
- 12.Meireles AM, Preto A, Rocha AS, Rebocho AP, Máximo V, Pereira-Castro I, et al. Molecular and genotypic characterization of human thyroid follicular cell carcinoma-derived cell lines. Thyroid. 2007;17:707–715. doi: 10.1089/thy.2007.0097. [DOI] [PubMed] [Google Scholar]
- 13.Koh CS, Ku JL, Park SY, Kim KH, Choi JS, Kim IJ, et al. Establishment and characterization of cell lines from three human thyroid carcinomas: responses to all-trans-retinoic acid and mutations in the BRAF gene. Mol Cell Endocrinol. 2007;264:118–127. doi: 10.1016/j.mce.2006.10.017. [DOI] [PubMed] [Google Scholar]
- 14.Nadeau V, Guillemette S, Bélanger LF, Jacob O, Roy S, Charron J. Map2k1 and Map2k2 genes contribute to the normal development of syncytiotrophoblasts during placentation. Development. 2009;136:1363–1374. doi: 10.1242/dev.031872. [DOI] [PubMed] [Google Scholar]
- 15.Dubois T, Rommel C, Howell S, Steinhussen U, Soneji Y, Morrice N, et al. 14-3-3 is phosphorylated by casein kinase I on residue 233. Phosphorylation at this site in vivo regulates Raf/14-3-3 interaction. J Biol Chem. 1997;272:28882–28888. doi: 10.1074/jbc.272.46.28882. [DOI] [PubMed] [Google Scholar]
- 16.Halaban R, Zhang W, Bacchiocchi A, Cheng E, Parisi F, Ariyan S, et al. PLX4032, a selective BRAF(V600E) kinase inhibitor, activates the ERK pathway and enhances cell migration and proliferation of BRAF melanoma cells. Pigment Cell Melanoma Res. 2010;23:190–200. doi: 10.1111/j.1755-148X.2010.00685.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Emery CM, Vijayendran KG, Zipser MC, Sawyer AM, Niu L, Kim JJ, et al. MEK1 mutations confer resistance to MEK and B-RAF inhibition. Proc Natl Acad Sci U S A. 2009;106:20411–20416. doi: 10.1073/pnas.0905833106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Salerno P, De Falco V, Tamburrino A, Nappi TC, Vecchio G, Schweppe RE, et al. Cytostatic activity of adenosine triphosphate-competitive kinase inhibitors in BRAF mutant thyroid carcinoma cells. J Clin Endocrinol Metab. 2010;95:450–455. doi: 10.1210/jc.2009-0373. [DOI] [PubMed] [Google Scholar]
- 19.Wang L, Yi T, Zhang W, Pardoll DM, Yu H. IL-17 enhances tumor development in carcinogen-induced skin cancer. Cancer Res. 2010;70:10112–10120. doi: 10.1158/0008-5472.CAN-10-0775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation. Nature. 2008;454:436–444. doi: 10.1038/nature07205. [DOI] [PubMed] [Google Scholar]
- 21.Pikarsky E, Porat RM, Stein I, Abramovitch R, Amit S, Kasem S, et al. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature. 2004;431:461–466. doi: 10.1038/nature02924. [DOI] [PubMed] [Google Scholar]
- 22.Muzza M, Degl'Innocenti D, Colombo C, Perrino M, Ravasi E, Rossi S, et al. The tight relationship between papillary thyroid cancer, autoimmunity and inflammation: clinical and molecular studies. Clin Endocrinol (Oxf) 2010;72:702–708. doi: 10.1111/j.1365-2265.2009.03699.x. [DOI] [PubMed] [Google Scholar]
- 23.Lumachi F, Basso SM, Orlando R. Cytokines, thyroid diseases and thyroid cancer. Cytokine. 2010;50:229–233. doi: 10.1016/j.cyto.2010.03.005. [DOI] [PubMed] [Google Scholar]
- 24.Reynolds JV, Donohoe CL, Doyle SL. Diet, obesity and cancer. Ir J Med Sci. 2011;180:521–527. doi: 10.1007/s11845-010-0653-5. [DOI] [PubMed] [Google Scholar]
- 25.Crawford S, Belajic D, Wei J, Riley JP, Dunford PJ, Bembenek S, et al. A novel B-RAF inhibitor blocks interleukin-8 (IL-8) synthesis in human melanoma xenografts, revealing IL-8 as a potential pharmacodynamic biomarker. Mol Cancer Ther. 2008;7:492–499. doi: 10.1158/1535-7163.MCT-07-0307. [DOI] [PubMed] [Google Scholar]
- 26.Linkov F, Ferris RL, Yurkovetsky Z, Marrangoni A, Velikokhatnaya L, Gooding W, et al. Multiplex analysis of cytokines as biomarkers that differentiate benign and malignant thyroid diseases. Proteomics Clin Appl. 2008;2:1575–1585. doi: 10.1002/prca.200780095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kim S, Park YW, Schiff BA, Doan DD, Yazici Y, Jasser SA, et al. An orthotopic model of anaplastic thyroid carcinoma in athymic nude mice. Clin Cancer Res. 2005;11:1713–1721. doi: 10.1158/1078-0432.CCR-04-1908. [DOI] [PubMed] [Google Scholar]
- 28.Charafe-Jauffret E, Ginestier C, Iovino F, Wicinski J, Cervera N, Finetti P, et al. Breast cancer cell lines contain functional cancer stem cells with metastatic capacity and a distinct molecular signature. Cancer Res. 2009;69:1302–1313. doi: 10.1158/0008-5472.CAN-08-2741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ginestier C, Liu S, Diebel ME, Korkaya H, Luo M, Brown M, et al. CXCR1 blockade selectively targets human breast cancer stem cells in vitro and in xenografts. J Clin Invest. 2010;120:485–497. doi: 10.1172/JCI39397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Stamenkovic I. Matrix metalloproteinases in tumor invasion and metastasis. Semin Cancer Biol. 2000;10:415–433. doi: 10.1006/scbi.2000.0379. [DOI] [PubMed] [Google Scholar]