

Abstract
Amyotrophic lateral sclerosis (ALS) is characterized by the progressive degeneration of motor neurons. There is a pathological and genetic link between ALS and frontotemporal lobar degeneration (FTLD). Although FTLD is characterized by abnormal phosphorylated tau deposition, it is unknown whether tau is phosphorylated in ALS motor neurons. Therefore, this study assessed tau epitopes that are commonly phosphorylated in FTLD, including serine 396 (pS396), 214 (pS214), and 404 (pS404) in motor neurons from clinically pure sporadic ALS cases compared with controls. In ALS lower motor neurons, tau pS396 was observed in the nucleus or the nucleus and cytoplasm. In ALS upper motor neurons, tau pS396 was observed in the nucleus, cytoplasm, or both the nucleus and cytoplasm. Tau pS214 and pS404 was observed only in the cytoplasm of upper and lower motor neurons in ALS. The number of motor neurons (per mm2) positive for tau pS396 and pS214, but not pS404, was significantly increased in ALS. Furthermore, there was a significant loss of phosphorylated tau-negative motor neurons in ALS compared with controls. Together, our data identified a complex relationship between motor neurons positive for tau phosphorylated at specific residues and disease duration, suggesting that tau phosphorylation plays a role in ALS.
Keywords: Amyotrophic lateral sclerosis, Motor neurons, Motor neuron disease, Neurodegeneration, Neuropathology, Phosphorylation, Tau
INTRODUCTION
Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disease characterized by the progressive degeneration of motor neurons in the brain and spinal cord, leading to widespread muscle wasting and weakness (1). Approximately 40% of ALS patients show evidence of cognitive impairment and 13% fulfill criteria for a clinical diagnosis of frontotemporal dementia (FTD) (2), which is second only to Alzheimer disease (AD) as the most common cause of early-onset dementia. Frontotemporal lobar degeneration (FTLD) is the term used to describe the neuropathological forms of FTD (3). FTLD neuropathology is heterogeneous but may be broadly divided into 3 subgroups based on the main protein deposits found in the brains of patients (3). The 3 main subgroups are FTLD with tau pathology, tar DNA binding protein 43 (TDP-43) pathology or FET pathology (3). ALS and FTLD patients may show abnormal accumulation of the same proteins in neurons and glial cells (e.g. TDP-43 and fused in sarcoma), and familial cases may be due to mutations in the same genes (e.g. C90RF72) (4). Together, this suggests that ALS and FTLD are not single disease entities but are part of a disease spectrum (4).
Approximately 45% of FTLD patients show neuronal and glial inclusions composed of hyper-phosphorylated tau (4). Tau has an important role in modulating the assembly and stability of microtubules, cellular trafficking and maintaining neuronal morphology (5, 6). In FTLD with tau pathology and AD, tau becomes progressively hyper-phosphorylated causing it to detach from the microtubule, eventually forming inclusions including neurofibrillary tangles (NFTs) in neurons and glial cells (5). The phosphorylation of key epitopes on the tau molecule, including serine 214, 396, and 404, have been shown to be promoters of tau disassembly, aggregation, and mislocalization (7–10). Tau inclusions in FTLD and AD postmortem brain tissue and mouse models are positive for these same phosphorylated epitopes (8, 10–13).
There have been a number of studies examining tau in the brains of ALS patients. Increased protein levels and cytoplasmic inclusions positive for tau phosphorylated at various residues, most notably threonine 175, 217 and serine 208/210, have been identified throughout the brains of patients with ALS and cognitive dysfunction, and to a lesser extent in ALS (14–17); however, motor neurons were not examined. Tau phosphorylated at threonine 175 and 231 has also been identified in hippocampal and spinal cord neurons in ALS with chronic encephalopathy (18). Phosphorylated tau (AT8 clone, serine 202/threonine 205) has been identified in neurons and glia throughout the brain and spinal cord of familial ALS patients with a mutation in optineurin (19) and in the hippocampal neurons of patients with ALS/parkinsonism-dementia complex (20). In addition, in 2 ALS-FTD patients presenting with a unique distribution and combination of protein deposits, AT8 tau has also be identified in the hippocampus and entorhinal cortex neurons (21) and astrocytes (22). Although the accumulation of a neurotoxic tau45–230 fragment has been identified in ALS motor neurons, phosphorylated tau epitopes known to be involved in AD and FTLD were not examined (23). It was the aim of this study to assess the motor neurons of clinically pure sporadic ALS cases for accumulation of phosphorylated tau at serine residues 214, 396, and 404 by immunohistochemistry and fibrillar tau (NFTs) by silver staining. Together our data show that abnormal tau phosphorylation but not fibrillar tau is present in both the upper and lower motor neurons of clinically pure sporadic ALS cases.
MATERIALS AND METHODS
Case Selection
Cases with a pathological diagnosis of sporadic ALS with TDP-43 pathology (n = 8) and age-and-gender matched controls (n = 7) were selected from a neuropathological series collected by the NSW Brain Banks through regional donor programs in Sydney, Australia. The brain donor programs hold approval from the Human Research Ethics Committees of the University of New South Wales and University of Sydney, and comply with the statement on human experimentation issued by the National Health and Medical Research Council of Australia. Patients were diagnosed during life by specialized neurologists using standard diagnostic criteria following a medical interview, cognitive testing, and informant history (24–26). Any ALS case with documented clinical FTD symptoms, including cognitive impairment, was excluded. Any control case with evidence of neurological disease was excluded. Any ALS or control case with primary and secondary malignancies, head injuries, history of stroke, inflammatory insults, or family history of neurodegenerative diseases was excluded. Standardized neuropathological characterization was performed and all ALS cases demonstrated motor neuron loss and TDP-43 neuronal inclusions in surviving motor neurons. Demographic, postmortem delay (PMD) and standard neuropathological assessment details provided by the NSW Brain Banks, as per current guidelines (27–29) are provided for the individual cases in Table 1. A comparison of the control and ALS cohort demographics and PMD intervals are provided in Table 2. This research project was approved by the Human Research Ethics Committee of the University of Wollongong.
TABLE 1.
Demographic, PMD, and Neuropathological Details of Control and ALS Cases
| Gender | Age at Death (Years) | Age at Onset (Years) | Disease Duration (Years) | PMD (Hours) | A Score (Aβ/Amyloid Plaques) | B Score (NFTs) | C Score (NPs) | TDP-43 Stage | Braak LB Stage | |
|---|---|---|---|---|---|---|---|---|---|---|
| Control 1 | Female | 72 | N/A | N/A | 25 | 0 | 2 | 0 | – | – |
| Control 2 | Female | 85 | N/A | N/A | 10 | 2 | 1 | 0 | – | – |
| Control 3 | Male | 73 | N/A | N/A | 9 | 1 | 1 | 0 | – | – |
| Control 4 | Male | 67 | N/A | N/A | 29 | 3 | 1 | 2 | – | – |
| Control 5 | Female | 81 | N/A | N/A | 35 | 0 | 1 | 0 | – | – |
| Control 6 | Male | 66 | N/A | N/A | 20.5 | 0 | 1 | 0 | – | – |
| Control 7 | Male | 75 | N/A | N/A | 34 | 2 | 0 | 0 | – | – |
| ALS 1 | Female | 72 | 70 | 2 | 8 | 0 | 0 | 0 | 4 | 0 |
| ALS 2 | Female | 70 | 69 | 1 | 5 | 0 | 0 | 0 | 4 | 0 |
| ALS 3 | Male | 66 | 62 | 4 | 24 | 1 | 0 | 0 | 4 | 0 |
| ALS 4 | Female | 81 | 80 | 1 | 19 | 3 | 1 | 3 | 1 | 0 |
| ALS 5 | Female | 82 | 77 | 5 | 24 | 2 | 0 | 2 | 3 | 0 |
| ALS 6 | Male | 75 | 72 | 3 | 29 | 1 | 1 | 1 | 4 | 0 |
| ALS 7 | Male | 72 | 69 | 3 | 31 | 1 | 0 | 0 | 4 | 0 |
| ALS 8 | Male | 67 | 65 | 2 | 35 | 3 | 0 | 2 | 3 | 0 |
ALS, amyotrophic lateral sclerosis; N/A, not applicable; PMD, postmortem delay; NFTs, neurofibrillary tangles; NPs, neuritic plaques; LB, Lewy body.
TABLE 2.
Demographic and PMD Details of Control and ALS Cohorts
| Control | ALS | |
|---|---|---|
| n (% male) | 7 (57%) | 8 (50%)*,† |
| Age at death (year ± standard error) | 74.1 ± 2 | 73.1 ± 3*,‡ |
| Age at onset (year ± standard error) | N/A | 70.5 ± 2 |
| Disease duration (year ± standard error) | N/A | 2.6 ± 0.5 |
| PMD (hours ± standard error) | 23.2 ± 4 | 21.9 ± 4*,‡ |
ALS, amyotrophic lateral sclerosis; N/A, not applicable.
p > 0.05 compared with controls.
Chi square test.
Student t-test.
Immunohistochemistry
Fresh tissue was fixed in 15% neutral buffered formalin for 2 weeks and transferred to 10% neutral buffered formalin for long-term storage. Formalin was replaced every 2 years to maintain tissue integrity. Formalin-fixed, paraffin-embedded tissue sections from the cervical spinal cord (lower motor neurons) and motor cortex (upper motor neurons) were cut at a thickness of 10 μm and slide-mounted. This was performed by the NSW Brain Banks using standardized protocols. Routine immunoperoxidase staining was then performed. Briefly, tissue sections were pretreated with citrate buffer and incubated with primary antibodies against tau phosphorylated at serine 396 (pS396 1:500, ab109390, Abcam, Cambridge, UK), serine 214 (pS214, 1:500, ab170892, Abcam), and serine 404 (pS404, 1:100, ab92676, Abcam). Sections were then incubated with a biotinylated secondary antibody (Vector Laboratories, Burlingame, CA), followed by avidin-biotin tertiary complex (ABC Elite, Vector Laboratories). The reaction product was visualized using 3, 3′-diaminobenzidine (Sigma-Aldrich, St. Louis, MO) and sections lightly counterstained with hematoxylin (POCD Healthcare, Artarmon, Australia). For each phosphorylated tau antibody, both a positive control (inferior temporal cortex from an AD case) and a negative control (primary antibody omitted) was included. All cases (control, ALS, positive control, and negative control) were stained at the same time using the same in-date solutions for each phosphorylated tau epitope. Specificity of the pS396 antibody was confirmed using a blocking peptide (Abcam) (Supplementary DataFig. S1). A modified Bielschowsky silver stain was used to assess the tissue for protein fibrils, including NFTs, as previously described (30).
Quantitation of Tau Staining
Brain and spinal cord sections were assessed at ×200–×400 magnification using the Leica ICC50 HD microscope camera (Leica, Wetzlar, Germany), images were taken and motor neurons counted using the cell counter function on Fiji (V1.51n) (31). To be classified as a motor neuron, cells had to meet the following criteria: (i) localization within the ventral horn of the cervical spinal cord or layer V of the motor cortex; (ii) greater than or equal to 20 µm in diameter along their longest axis (32); and (iii) contain a nucleus identified by hematoxylin. Tau-positive nuclear staining was classified as “web-like” if the staining overlapped with the hematoxylin counterstain or “intense” if the staining engulfed the nucleus. The sections were also visualized using the UV wavelength on a Leica DMi8 microscope (Leica), supporting that the tau staining observed in the cytoplasm was not lipofuscin. The Color Atlas of the Brain and Spinal Cord by England and Wakely was used to identify motor neurons in the ventral horn of the cervical spinal cord (pages 181, 212, 214) and layer V in the motor cortex (pages 118, 288) (33). For each case, the area of the ventral horn in the spinal cord and layer V in the motor cortex was outlined using the freehand selection tool on Fiji (V1.51n) and the area determined using the measurement function on Fiji (V1.51n). The number of motor neurons per mm2 was then determined for each case. The mean number of motor neurons per mm2 within each group was converted to a percentage, whereby the total number of control motor neurons per mm2 was 100%. This provides a graphical representation of the difference in proportion of tau-negative to tau-positive motor neurons in controls compared with ALS cases and further demonstrates the loss of motor neurons in ALS compared with controls. Researchers were blinded to case details during analysis, with quantitation performed in 3 cases per region by the same investigator on different days and by 2 researchers blind to cases details, to give an intra and interrater variance of <5%.
Statistics
Statistical analyses were performed in GraphPad Prism (Version 6, San Diego, CA), with p < 0.05 considered significant. A D’Agostino-Pearson test was first used to test for normality, revealing that the data were not normally distributed. Therefore, nonparametric Mann Whitney U tests were used to determine differences in the number of motor neurons per mm2 between ALS and control cases, as well as differences in the number of tau-positive and tau-negative motor neurons per mm2 between the ALS and control cases. Differences in the cell localization of tau pS396 staining within the ALS cohort was determined using a Mann-Whitney U test for the lower motor neurons and a Kruskal-Wallis with Dunn’s multiple comparisons test for the upper motor neurons. Spearman rank correlations were performed to determine relationships between tau phosphorylation and disease duration, patient age at death, A score (Aβ/amyloid plaques), B score (NFTs), C score (neuritic plaques [NPs]), TDP-43 stage and Lewy body stage for ALS cases, and between tau phosphorylation and ABC scores for controls.
RESULTS
Tau pS396 Accumulation in the Nucleus and Cytoplasm of ALS Motor Neurons
The number of motor neurons per mm2 was reduced in ALS compared with control cases in both the spinal cord (lower motor neurons) and motor cortex (upper motor neurons). Four 10-μm-thick sections were analyzed, and on average a 53% loss of lower motor neurons was observed in ALS compared with controls (33.59 ± 3.95 × 10−12 mm2 vs 63.75 ± 6.94 × 10−12 mm2, respectively, p = 0.0012), and a 48% loss of upper motor neurons in ALS compared with controls (19.82 ± 2.63 × 10−12 mm2 vs 41.44 ± 6.39 × 10−12 mm2, respectively, p = 0.02). This is in line with previous studies (34, 35).
Tau pS396 was rarely observed in the control spinal cord (Fig. 1A) with only 1.57 ± 1.05% of total lower motor neurons (positive staining in 2/7 cases) showing weak cytoplasmic staining. In contrast, tau pS396 staining was identified in the nucleus (Fig. 1B, C), or both the nucleus and cytoplasm (Fig. 1D) of 92.9 ± 16.5% of ALS lower motor neurons (positive staining in 8/8 cases). Similarly, tau pS396 was absent from the majority of control upper motor neurons (Fig. 1E) with 15.55 ± 5.15% of upper motor neurons showing weak cytoplasmic staining. In contrast, tau pS396 staining was observed in the nucleus (Fig. 1F), cytoplasm (Fig. 1G), or both the nucleus and cytoplasm (Fig. 1H) of 91.55 ± 17.52% of upper motor neurons (positive staining in 8/8 cases). The number of tau pS396-positive ALS motor neurons per mm2 was significantly increased from controls in both the spinal cord (increased 35-fold, p = 0.0003; Fig. 1I) and motor cortex (increased 4-fold, p = 0.01; Fig. 1J). The number of tau pS396-negative motor neurons per mm2 was significantly reduced from controls in both the spinal cord (reduced 11-fold, p = 0.0003; Fig. 1I) and motor cortex (reduced 10-fold, p = 0.006; Fig. 1J).
FIGURE 1.

Tau pS396 immunoreactivity in motor neurons from control and ALS cases. Representative micrographs of lower motor neurons (LMNs) in the cervical spinal cord showing no immunoreactivity in controls (A) but nuclear (B, C) and both nuclear and cytoplasmic accumulation in ALS (D), and of upper motor neurons (UMNs) in the motor cortex showing no immunoreactivity in controls (E) but nuclear (F), cytoplasmic (G) and both nuclear and cytoplasmic accumulation in ALS (H). Comparison between the percentage of tau pS396-positive and negative motor neurons in control and ALS spinal cord (I) and motor cortex (J). Comparison between the cellular localization of tau pS396 in ALS motor neurons (K). Control: n = 7; ALS: n = 8. Scale bar in (H) is equivalent for A–H. Scale bar = 10 μm.
Within the ALS cohort, 2 tau pS396 nuclear morphologies were identified—the most prevalent was a web-like pattern that overlapped with the hematoxylin counterstain (Fig. 1B, F), however an intense stain that fully engulfed the nucleus was also observed (Fig. 1C) (now referred to as “web-like” vs “intense” nuclear pS396 tau; 94.9 ± 2.9% vs 5.1 ± 0.3% in the spinal cord; 93.9 ± 2.9% vs 6.0 ± 0.4% in the motor cortex). The majority of ALS motor neurons showed an enlarged cytoplasm, consistent with motor neuron swelling (36). There was a significant increase in the number of ALS lower motor neurons with nuclear compared with nuclear and cytoplasmic staining (90.0 ± 17.8% vs 10.0 ± 5.7%, p = 0.0005); however, there was no significant difference in pS396 cell localization in the upper motor neurons (nuclear = 39.3 ± 9.1%, cytoplasm = 16.9 ± 4.3%, nuclear + cytoplasm = 43.9 ± 10.7%; Fig. 1K).
Tau pS214 and pS404 Accumulation in the Cytoplasm of ALS Motor Neurons
Tau pS214 and pS404 staining was only observed in the cytoplasm of motor neurons. In the spinal cord, tau pS214 accumulation was minimal in controls affecting 15.6 ± 4.6% of lower motor neurons (positive staining in 5/7 cases; Fig. 2A) compared with 87.4 ± 13.5% of ALS lower motor neurons (positive staining in 8/8 cases; Fig. 2B). Similarly, in the motor cortex, only 7.2 ± 2.1% of control upper motor neurons showed tau pS214 staining (positive staining in 5/7 cases; Fig. 2C) compared with 88.4 ± 16.2% of ALS upper motor neurons (positive staining in 8/8 cases; Fig. 2D). The number of tau pS214-positive ALS motor neurons per mm2 was significantly increased from controls in both the spinal cord (increased 4-fold, p = 0.002; Fig. 2E) and motor cortex (increased 6-fold, p = 0.001; Fig. 2F). Similarly, the number of tau pS214-negative neurons per mm2 was significantly reduced from controls in both the spinal cord (reduced 11-fold, p = 0.0003; Fig. 2E) and motor cortex (reduced 15-fold, p = 0.0003; Fig. 2F).
FIGURE 2.
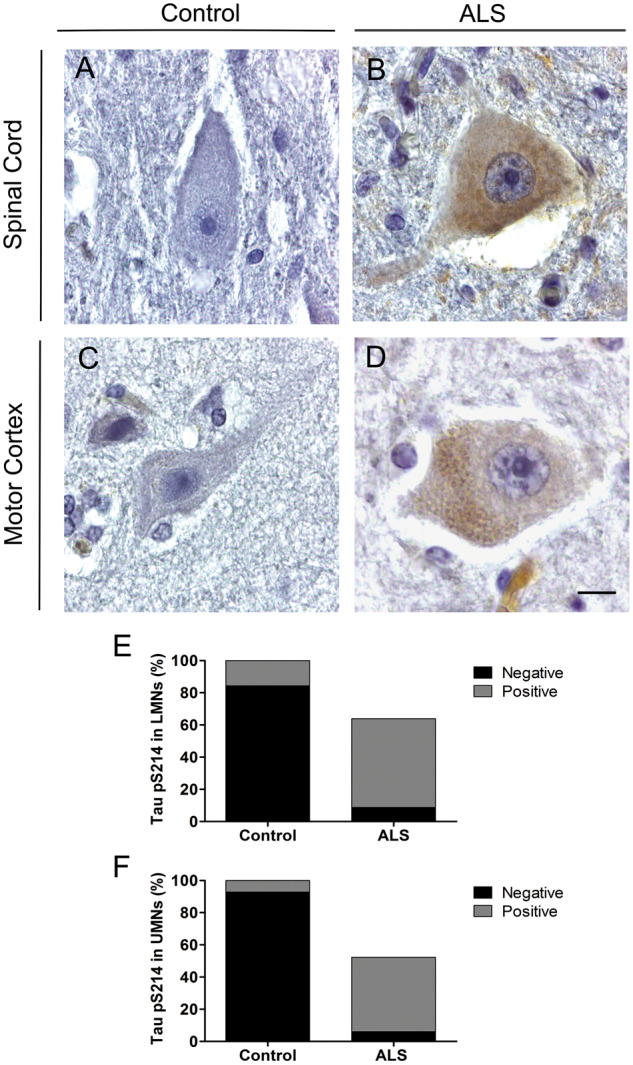
Tau pS214 immunoreactivity in motor neurons from control and ALS cases. Representative micrographs of lower motor neurons (LMNs) in the cervical spinal cord showing no immunoreactivity in controls (A) but cytoplasmic accumulation in ALS (B), and of upper motor neurons (UMNs) showing no immunoreactivity in controls (C) but cytoplasmic accumulation in ALS (D). Comparison between the percentage of tau pS214-positive and negative motor neurons in control and ALS spinal cord (E) and motor cortex (F). Control: n = 7; ALS: n = 8. Scale in (D) is equivalent for A–D. Scale bar = 10 μm.
In the spinal cord, tau pS404 staining was observed in 40.7 ± 11.0% of control lower motor neurons (positive staining in 7/7 cases; Fig. 3A) compared with 91.73 ± 29.8% of ALS lower motor neurons (positive staining in 8/8 cases; Fig. 3B). In the motor cortex, 18.5 ± 2.7% of control upper motor neurons showed tau pS404 staining (positive staining in 7/7 cases; Fig. 3C) compared with 96.5 ± 19.8% of ALS upper motor neurons (positive staining in 8/8 cases; Fig. 3D). Although the number of ALS motor neurons with tau pS404 staining per mm2 was not significantly different from controls, there was a significant reduction in the number of tau pS404-negative neurons per mm2 in both the spinal cord (reduced 13-fold, p = 0.008; Fig. 3E) and motor cortex (reduced 100-fold, p = 0.0003; Fig. 3F). In addition, small round deposits of tau pS404 were evident in some ALS lower motor neurons (Fig. 3C inset), which were not observed in controls.
FIGURE 3.
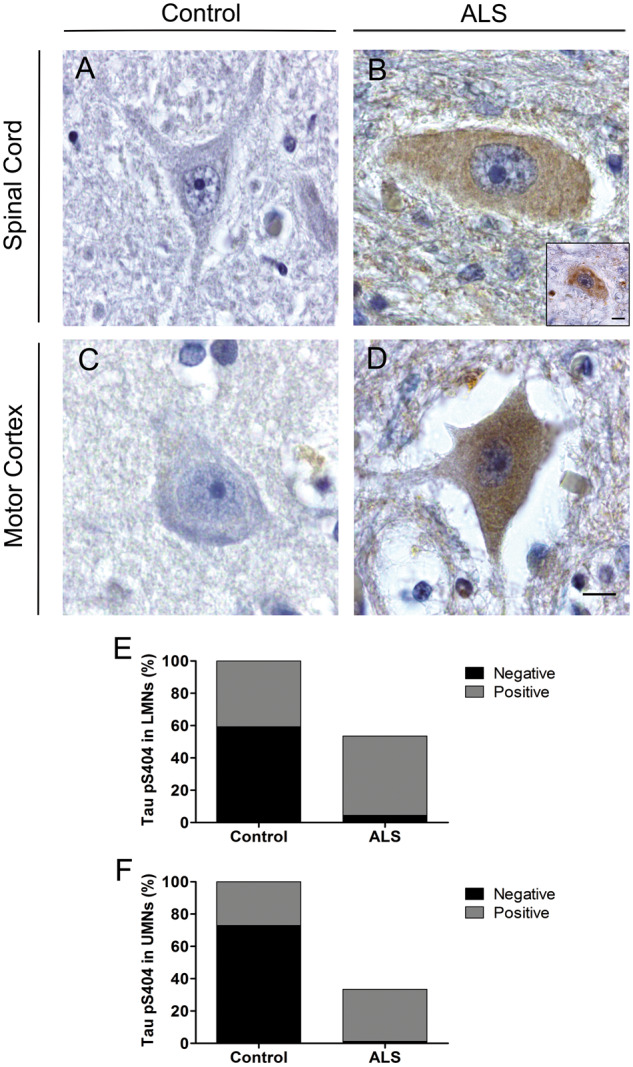
Tau pS404 immunoreactivity in motor neurons from control and ALS cases. Representative micrographs of lower motor neurons (LMNs) in the cervical spinal cord showing no immunoreactivity in controls (A), and both diffuse cytoplasmic accumulation (B) and small round deposits (B, inset) in ALS. Representative micrographs of upper motor neurons (UMNs) in the motor cortex showing no immunoreactivity in controls (C), and cytoplasmic accumulation in ALS (D). Comparison between the percentage of tau pS404-positive and negative motor neurons in control and ALS spinal cord (E) and motor cortex (F). Control: n = 7; ALS: n = 8. Scale in (D) is equivalent for A–D. Scale bars = 10 μm.
Absence of Fibrillar Tau in ALS Motor Neurons
Modified Bielschowsky silver staining was performed to examine ALS tissue for fibrillar tau. Despite the fact that abundant NFTs were observed in the positive control sections (inferior temporal cortex from an AD case), silver staining did not reveal any evidence of fibrils, including NFTs, in the spinal cord or motor cortex of ALS cases or controls (Supplementary DataFig. S2).
Correlation of Tau Phosphorylation With Case Demographics and Standard Neuropathological Assessment
The number of tau pS214-positive motor neurons per mm2 in the ALS spinal cord negatively correlated with disease duration (Rho = -0.812, p = 0.02; Fig. 4A). Conversely, there was a positive correlation between disease duration and the number of both tau pS214- (Rho = 0.812, p = 0.02; Fig. 4B) and pS404- (Rho = 0.934, p = 0.002; Fig. 4C) positive motor neurons per mm2 in the ALS motor cortex. A positive correlation was also observed between motor neurons with intense pS396 nuclear staining and disease duration in the motor cortex (Rho = 0.778, p = 0.018; Fig. 4D). There was no relationship between any other pS396 variable and disease duration or patient age at death, or between the number of tau pS214- and pS404-positive motor neurons per mm2 and patient age at death for either region.
FIGURE 4.

Linear relationship between tau pS214 immunoreactivity in the lower motor neurons (LMNs) and disease duration (A), and upper motor neurons (UMNs) and disease duration (B). Linear relationship between tau pS404 immunoreactivity in the UMNs and disease duration (C). Linear relationship between UMNs with intense nuclear tau pS396 staining and disease duration (D). ALS: n = 8.
The number of tau pS396 motor neurons per mm2 with nuclear and cytoplasmic staining in the ALS spinal cord negatively correlated with both the A (Rho = –0.785, p = 0.028; Fig. 5A) and C (Rho = –0.838, p = 0.015; Fig. 5B) scores. A negative correlation was also observed between the number of tau pS396-positive motor neurons per mm2 with cytoplasmic staining in the ALS motor cortex and the C score (Rho = –0.805, p = 0.022; Fig. 5C). In contrast, ALS motor neurons with cytoplasmic tau pS396 staining in the motor cortex positively correlated with the TDP-43 stage (Rho = 0.866, p = 0.012; Fig. 5D).
FIGURE 5.

Linear relationship between nuclear and cytoplasmic tau pS396 immunoreactivity in the lower motor neurons (LMNs) and the A score (Aβ/amyloid plaques) (A) and the C score (neuritic plaques) (B). Linear relationship between cytoplasmic tau pS396 immunoreactivity in the upper motor neurons (UMNs) and the C score (neuritic plaques) (C) and TDP-43 stage (D). ALS: n = 8.
There was no relationship between any other tau pS396 variable or the number of tau pS214- and pS404-positive motor neurons per mm2 with the A, B, or C scores, TDP-43 stage or Braak Lewy body stage in the ALS cases. There was no relationship between the number of tau pS396-, pS214-, or pS404-positive motor neurons per mm2 and the A, B, or C scores in the control cases.
DISCUSSION
In the present study, tau pS396, pS214, and pS404 immunoreactivity was assessed in the motor neurons from the cervical spinal cord and motor cortex of patients diagnosed with clinically pure ALS compared with age- and gender-matched healthy controls. For the first time, tau pS396 staining has been identified in the nucleus of ALS motor neurons. Increased numbers of motor neurons with cytoplasmic pS396 and pS214 staining were found in ALS cases compared with controls, and a reduction in tau-negative neurons was identified in ALS compared with controls for all phospho-tau epitopes.
While previous studies suggest that tau may play a role in ALS (14–16, 23, 37), phospho-tau epitopes have not been assessed in the motor neurons of ALS cases with no comorbidity, particularly FTD and cognitive symptoms. This is the first study to report accumulation of tau, specifically tau pS396 in the nucleus of ALS motor neurons, although PHF1 tau (S404/S396) has been identified in the nucleus of neurons throughout the brain of a patient with presenile dementia with motor neuron disease (38), supporting this finding. Previously, tau has been reported in the nucleus of both neuronal and nonneuronal cells where it is proposed to have a role in protecting DNA from cellular stress (39–41). During stress, nonphosphorylated tau may also be found in the nucleolus where it has been shown to be important in heterochromatin stability and ribosomal DNA transcriptional repression (39, 42). Furthermore, stress induced by glutamate, formaldehyde and Aβ42 has been shown to result in increased nuclear tau phosphorylated at Thr231 (42), Thr181 and S396 (43), and PHF1 (pS404/S396) (44), respectively. This indicates the existence of different tau species in the nucleus, and suggests that they may play different roles depending on the type and duration of stress (42).
Recently, a study assessed human brain for the presence of AT100 tau (pThr212/Ser214) finding increased nuclear tau with aging, but a significant reduction in cancer and with progression of AD (45, 46). AT100 nuclear tau was shown to colocalize with DAPI-positive chromatin in neurons and was suggested to be important for the cell to maintain control of genomic DNA (45). Interestingly, AT100 tau showed a mottled appearance in the nucleus of aged pyramidal neurons (40–60 years) and increased in intensity in senile pyramidal neurons (>60 years), which also showed light cytoplasmic staining (45). This is similar to the findings in the present study of web-like nuclear pS396 tau staining that overlapped with hematoxylin (stains nuclear chromatin) and the intense nuclear pS396 tau staining that fully engulfed the nucleus. In addition, the present study also observed motor neurons with both nuclear and cytoplasmic staining. This may indicate a progression of pS396 tau in ALS motor neurons, supported by the correlation between increasing intense nuclear tau and disease duration in the motor cortex. Interestingly, degenerating AD neurons with cytoplasmic NFTs have been shown to contain almost no nuclear tau (45, 46), in contrast to the present findings of phosphorylated nuclear tau, coupled with an absence of NFTs in ALS. Similarly, Gallyas silver staining has previously revealed intra and extraneuronal tau aggregates in ALS with dementia (15, 47) that were absent from clinically pure ALS cases (15). Western blotting of ALS and AD brain samples for tau has also revealed different banding patterns in the insoluble fraction suggesting the presence of different tau isoforms (16). Comparing the RNA and protein profiles of motor neurons from clinically pure ALS cases compared with ALS-FTLD, FTLD with tau pathology and AD may shed light on the mechanisms involved in fibrillar tau formation. Furthermore, the negative correlation observed between nuclear and cytoplasmic tau pS396 in the ALS spinal cord and the A (Aβ/amyloid plaques) and C (neuritic plaques) scores, and between cytoplasmic tau pS396 in the ALS motor cortex and the C score (neuritic plaques), may suggest a protective role for pS396 tau against Aβ neuropathology. Our findings therefore indicate that the role of phosphorylation at specific residues of tau should be the subject of future research in ALS. Additional methods including the use of recently developed fluorescent dyes and probes may also be considered to further assess ALS tissue for evidence of tau aggregation (48, 49). The positive correlation observed between cytoplasmic tau pS396 in the ALS motor cortex and TDP-43 stage is also of interest in light of recent work in a mouse model of chronic traumatic encephalopathy that found a reduction in TDP-43 pathology after elimination of phosphorylated tau (50), and the subsequent suggestion that that there may be a synergistic toxic relationship between the 2 proteins (51). Taken together, this suggests that different mechanisms may be responsible for tau accumulation in ALS compared with AD and ALS with dementia/cognitive impairment.
A change in tau phosphorylation is suggested to be responsible for alterations in nuclear tau; however, the exact mechanism of entry into the nucleus is still not known (39). Studies have shown that disrupting the balance between tau phosphorylation and O-linked glycosylation changes tau nuclear localization (52), and that glycosylation has epitope-specific effects on levels of tau phosphorylation (53). Reduced O-glycosylation has been reported in spinal cord motor neurons from a SOD1 transgenic mouse model of ALS (54), however, given the differences observed in tau accumulation between human ALS tissue and the SOD1 mouse (23), assessing glycosylation levels in ALS patient tissue would be valuable.
Similar to FTLD tau and AD, phosphorylated tau was observed in the cytoplasm of ALS motor neurons. However, the staining was diffusely spread throughout the cytoplasm and didn’t form specific structures, with the exception of round pS404 tau-positive deposits seen in some ALS lower motor neurons. Cellular stress is known to cause increased tau phosphorylation, reducing its binding affinity to microtubules, eventually resulting in its abnormal accumulation in the cytoplasm (55, 56). In this context, the phosphorylation of tau at S396, S214 and S404 has been directly linked to the disassociation of tau from the microtubule complex (7, 57, 58). Combined with the evidence of cellular stress observed in the current ALS cohort (swollen motor neurons indicative of chromatolysis), the cytoplasmic accumulation of phosphorylated tau in ALS is consistent with the hypothesis of a stress-induced mislocalization of tau.
Cytoplasmic accumulation of tau pS214 and pS404 in the motor cortex of ALS patients positively correlated with disease duration, suggesting an increase in the number of neurons affected over the disease course. Conversely, however, a negative correlation was observed between cytoplasmic accumulation of tau pS214 and disease duration in the spinal cord. A positive correlation between intense nuclear tau pS396 staining and disease duration was also observed in the motor cortex but not in the spinal cord. These region-specific changes may be due to differences in the severity of motor neuron loss between the spinal cord and motor cortex, but are also interesting in light of recent studies that have highlighted differences between the spinal cord and motor cortex in ALS. Positron emission tomography studies using 18F-fluorodeoxyglucose to assess metabolic activity observed higher tracer uptake in the spinal cord of ALS patients compared with controls, but reduced uptake in the cortex (59). Similarly the molecule semaphorin 3A, which has been implicated in the death of motor neurons, is increased in the motor cortex but reduced in the spinal cord of ALS patients, and has been shown to enhance survival of spinal cord motor neurons but reduce survival of cortical neurons (60). Our data thus support this different temporal progression of the mechanisms underlying disease progression in the upper versus lower motor neurons.
Conclusion
This study has identified increased tau phosphorylation in the nucleus and cytoplasm of upper and lower motor neurons from clinically pure sporadic ALS patients. Tau phosphorylated at S396 was identified in the nucleus and cytoplasm of ALS motor neurons, while S214 and S404 were identified only in the cytoplasm of ALS motor neurons. There were region-specific differences in the relationships between phosphorylated tau-positive motor neurons and disease duration, supporting a different temporal progression of tau phosphorylation in the motor neurons from the spinal cord versus motor cortex. Overall, this study contributes to the growing body of evidence suggesting that phosphorylated tau is involved in ALS; however, it remains to be determined whether it is neurodegenerative or neuroprotective.
Supplementary Material
ACKNOWLEDGMENTS
Tissues were received from the New South Wales Brain Tissue Resource Centre at the University of Sydney and the Sydney Brain Bank at Neuroscience Research Australia which are supported by the University of New South Wales, Neuroscience Research Australia and Schizophrenia Research Institute. Research reported in this publication was supported by the National Institute On Alcohol Abuse And Alcoholism of the National Institutes of Health under Award Number R28AA012725. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Lezanne Ooi is supported by a National Health and Medical Research Council (NHMRC) of Australia Boosting Dementia Research Leadership Fellowship (APP1135720). Glenda M. Halliday is supported by a NHMRC Senior Principal Research Fellow (APP1079679).
The authors have no duality or conflicts of interest to declare.
Supplementary Data can be found at academic.oup.com/jnen.
REFERENCES
- 1. Cleveland DW, Rothstein JD. From Charcot to Lou Gehrig: Deciphering selective motor neuron death in ALS. Nat Rev Neurosci 2001;2:806–19 [DOI] [PubMed] [Google Scholar]
- 2. Phukan J, Elamin M, Bede P, et al. The syndrome of cognitive impairment in amyotrophic lateral sclerosis: A population-based study. J Neurol Neurosurg Psychiatry 2012;83:102–8 [DOI] [PubMed] [Google Scholar]
- 3. Mackenzie IR, Neumann M. Molecular neuropathology of frontotemporal dementia: Insights into disease mechanisms from postmortem studies. J Neurochem 2016;138(Suppl. 1):54–70 [DOI] [PubMed] [Google Scholar]
- 4. Tan RH, Ke YD, Ittner LM, et al. ALS/FTLD: Experimental models and reality. Acta Neuropathol 2017;133:177–96 [DOI] [PubMed] [Google Scholar]
- 5. Spillantini MG, Goedert M. Tau pathology and neurodegeneration. Lancet Neurol 2013;12:609–22 [DOI] [PubMed] [Google Scholar]
- 6. Medina M, Hernandez F, Avila J. New features about tau function and dysfunction. Biomolecules 2016;6:21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Evans DB, Rank KB, Bhattacharya K, et al. Tau phosphorylation at serine 396 and serine 404 by human recombinant tau protein kinase II inhibits tau's ability to promote microtubule assembly. J Biol Chem 2000;275:24977–83 [DOI] [PubMed] [Google Scholar]
- 8. Augustinack JC, Schneider A, Mandelkow E-M, et al. Specific tau phosphorylation sites correlate with severity of neuronal cytopathology in Alzheimer's disease. Acta Neuropathol 2002;103:26–35 [DOI] [PubMed] [Google Scholar]
- 9. Haase C, Stieler JT, Arendt T, et al. Pseudophosphorylation of tau protein alters its ability for self-aggregation. J Neurochem 2004;88:1509–20 [DOI] [PubMed] [Google Scholar]
- 10. Zheng-Fischhofer Q, Biernat J, Mandelkow EM, et al. Sequential phosphorylation of Tau by glycogen synthase kinase-3beta and protein kinase A at Thr212 and Ser214 generates the Alzheimer-specific epitope of antibody AT100 and requires a paired-helical-filament-like conformation. Eur J Biochem 1998;252:542–52 [DOI] [PubMed] [Google Scholar]
- 11. Gotz J, Chen F, van Dorpe J, et al. Formation of neurofibrillary tangles in P301l tau transgenic mice induced by Abeta 42 fibrils. Science 2001;293:1491–5 [DOI] [PubMed] [Google Scholar]
- 12. Flores-Rodriguez P, Ontiveros-Torres MA, Cardenas-Aguayo MC, et al. The relationship between truncation and phosphorylation at the C-terminus of tau protein in the paired helical filaments of Alzheimer's disease. Front Neurosci 2015;9:33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. van Eersel J, Stevens CH, Przybyla M, et al. Early-onset axonal pathology in a novel P301S-Tau transgenic mouse model of frontotemporal lobar degeneration. Neuropathol Appl Neurobiol 2015;41:906–25 [DOI] [PubMed] [Google Scholar]
- 14. Yang W, Strong MJ. Widespread neuronal and glial hyperphosphorylated tau deposition in ALS with cognitive impairment. Amyotroph Lateral Scler 2012;13:178–93 [DOI] [PubMed] [Google Scholar]
- 15. Yang W, Sopper MM, Leystra-Lantz C, et al. Microtubule-associated tau protein positive neuronal and glial inclusions in ALS. Neurology 2003;61:1766–73 [DOI] [PubMed] [Google Scholar]
- 16. Strong M, Yang W, Strong W, et al. Tau protein hyperphosphorylation in sporadic ALS with cognitive impairment. Neurology 2006;66:1770–1 [DOI] [PubMed] [Google Scholar]
- 17. Behrouzi R, Liu X, Wu D, et al. Pathological tau deposition in Motor Neurone Disease and frontotemporal lobar degeneration associated with TDP-43 proteinopathy. Acta Neuropathol Commun 2016;4:33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Moszczynski AJ, Strong W, Xu K, et al. Pathologic Thr175 tau phosphorylation in CTE and CTE with ALS. Neurology 2018;90:e380–e7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Ayaki T, Ito H, Komure O, et al. Multiple proteinopathies in familial ALS cases with optineurin mutations. J Neuropathol Exp Neurol 2018;77:128–38 [DOI] [PubMed] [Google Scholar]
- 20. Hata Y, Ma N, Yoneda M, et al. Nitrative stress and tau accumulation in amyotrophic lateral sclerosis/parkinsonism-dementia complex (ALS/PDC) in the Kii peninsula, Japan. Front Neurosci 2017;11:751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Guedes ACB, Santin R, Costa ASR, et al. Distinct Phospho-TDP-43 brain distribution in two cases of FTD, one associated with ALS. Dement Neuropsychol 2017;11:249–54 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Takeuchi R, Toyoshima Y, Tada M, et al. Globular glial mixed four repeat tau and TDP-43 proteinopathy with motor neuron disease and frontotemporal dementia. Brain Pathol 2016;26:82–94 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Vintilescu CR, Afreen S, Rubino AE, et al. The neurotoxic Tau45-230 fragment accumulates in upper and lower motor neurons in amyotrophic lateral sclerosis subjects. Mol Med 2016;22:477–86 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Al-Chalabi A, Hardiman O, Kiernan MC, et al. Amyotrophic lateral sclerosis: Moving towards a new classification system. Lancet Neurol 2016;15:1182–94 [DOI] [PubMed] [Google Scholar]
- 25. Hardiman O, van den Berg LH, Kiernan MC. Clinical diagnosis and management of amyotrophic lateral sclerosis. Nat Rev Neurol 2011;7:639–49 [DOI] [PubMed] [Google Scholar]
- 26. Kiernan MC, Vucic S, Cheah BC, et al. Amyotrophic lateral sclerosis. Lancet 2011;377:942–55 [DOI] [PubMed] [Google Scholar]
- 27. Hyman BT, Phelps CH, Beach TG, et al. National Institute on Aging-Alzheimer's Association guidelines for the neuropathologic assessment of Alzheimer's disease. Alzheimers Dement 2012;8:1–13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Montine TJ, Phelps CH, Beach TG, et al. National Institute on Aging-Alzheimer's Association guidelines for the neuropathologic assessment of Alzheimer's disease: A practical approach. Acta Neuropathol 2012;123:1–11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Brettschneider J, Del Tredici K, Toledo JB, et al. Stages of pTDP-43 pathology in amyotrophic lateral sclerosis. Ann Neurol 2013;74:20–38 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Halliday G, Flowers D, Baum L. Analysis of staining methods for different cortical plaques in Alzheimer's disease. Acta Neuropathol 1994;87:174–86 [DOI] [PubMed] [Google Scholar]
- 31. Schindelin J, Arganda-Carreras I, Frise E, et al. Fiji: An open-source platform for biological-image analysis. Nat Methods 2012;9:676–82 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Wakayama I. Morphometry of spinal motor neurons in amyotrophic lateral sclerosis with special reference to chromatolysis and intracytoplasmic inclusion bodies. Brain Res 1992;586:12–8 [DOI] [PubMed] [Google Scholar]
- 33.England MA, Wakely J. Color Atlas of the Brain and Spinal Cord. Second Edition. Philadelphia, PA: Mosby Elsevier; 2006 [Google Scholar]
- 34. Saberi S, Stauffer JE, Schulte DJ, et al. Neuropathology of amyotrophic lateral sclerosis and its variants. Neurol Clin 2015;33:855–76 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Coan G, Mitchell CS. An assessment of possible neuropathology and clinical relationships in 46 sporadic amyotrophic lateral sclerosis patient autopsies. Neurodegener Dis 2015;15:301–12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Cragg BG. What is the signal for chromatolysis? Brain Res 1970;23:1–21 [DOI] [PubMed] [Google Scholar]
- 37. Moszczynski AJ, Yang W, Hammond R, et al. Threonine175, a novel pathological phosphorylation site on tau protein linked to multiple tauopathies. Acta Neuropathol Commun 2017;5:1–14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Papasozomenos SC. Nuclear tau-immunoreactivity in presenile-dementia with motor-neuron disease – a case-report. Clin Neuropathol 1995;14:100–4 [PubMed] [Google Scholar]
- 39. Sultan A, Nesslany F, Violet M, et al. Nuclear tau, a key player in neuronal DNA protection. J Biol Chem 2011;286:4566–75 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Violet M, Delattre L, Tardivel M, et al. A major role for Tau in neuronal DNA and RNA protection in vivo under physiological and hyperthermic conditions. Front Cell Neurosci 2014;8:84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Maina MB, Al-Hilaly YK, Serpell LC. Nuclear tau and its potential role in Alzheimer's disease. Biomolecules 2016;6:9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Maina MB, Bailey LJ, Wagih S, et al. The involvement of tau in nucleolar transcription and the stress response. Acta Neuropathol Commun 2018;6:70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Lu J, Miao JY, Su T, et al. Formaldehyde induces hyperphosphorylation and polymerization of Tau protein both in vitro and in vivo. Biochim Biophys Acta 2013;1830:4102–16 [DOI] [PubMed] [Google Scholar]
- 44. Noel A, Barrier L, Ingrand S. The Tyr216 phosphorylated form of GSK3 beta contributes to tau phosphorylation at PHF-1 epitope in response to A beta in the nucleus of SH-SY5Y cells. Life Sci 2016;158:14–21 [DOI] [PubMed] [Google Scholar]
- 45. Gil L, Federico C, Pinedo F, et al. Aging dependent effect of nuclear tau. Brain Res 2017;1677:129–37 [DOI] [PubMed] [Google Scholar]
- 46. Hernandez-Ortega K, Garcia-Esparcia P, Gil L, et al. Altered machinery of protein synthesis in Alzheimer's: From the nucleolus to the ribosome. Brain Pathol 2016;26:593–605 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Noda K, Katayama S, Watanabe C, et al. Gallyas- and tau-positive glial structures in motor neuron disease with dementia. Clin Neuropathol 1999;18:218–25 [PubMed] [Google Scholar]
- 48. Verwilst P, Kim HS, Kim S, et al. Shedding light on tau protein aggregation: The progress in developing highly selective fluorophores. Chem Soc Rev 2018;47:2249–65 [DOI] [PubMed] [Google Scholar]
- 49. Lai HM, Ng WL, Gentleman SM, et al. Chemical probes for visualizing intact animal and human brain tissue. Cell Chem Biol 2017;24:659–72 [DOI] [PubMed] [Google Scholar]
- 50. Albayram O, Kondo A, Mannix R, et al. Cis P-tau is induced in clinical and preclinical brain injury and contributes to post-injury sequelae. Nat Commun 2017;8:1000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Moszczynski AJ, Hintermayer MA, Strong MJ. Phosphorylation of threonine 175 tau in the induction of tau pathology in amyotrophic lateral sclerosis-frontotemporal spectrum disorder (ALS-FTSD). A review. Front Neurosci 2018;12:259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Lefebvre T, Ferreira S, Dupont-Wallois L, et al. Evidence of a balance between phosphorylation and O-GlcNAc glycosylation of Tau proteins—a role in nuclear localization. Biochim Biophys Acta 2003;1619:167–76 [DOI] [PubMed] [Google Scholar]
- 53. Yu Y, Zhang L, Li X, et al. Differential effects of an O-GlcNAcase inhibitor on tau phosphorylation. PLoS One 2012;7:e35277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Shan XY, Vocadlo DJ, Krieger C. Reduced protein O-glycosylation in the nervous system of the mutant SOD1 transgenic mouse model of amyotrophic lateral sclerosis. Neurosci Lett 2012;516:296–301 [DOI] [PubMed] [Google Scholar]
- 55. Naini SMA, Soussi-Yanicostas N. Tau hyperphosphorylation and oxidative stress, a critical vicious circle in neurodegenerative tauopathies? Oxid Med Cell Longev 2015;2015:151979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Mietelska-Porowska A, Wasik U, Goras M, et al. Tau protein modifications and interactions: Their role in function and dysfunction. Int J Mol Sci 2014;15:4671–713 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Illenberger S, Zheng-Fischhöfer Q, Preuss U, et al. The endogenous and cell cycle-dependent phosphorylation of tau protein in living cells: Implications for Alzheimer’s disease. Mol Biol Cell 1998;9:1495–512 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Cho JH, Johnson G. Glycogen synthase kinase 3 beta phosphorylates tau at both primed and unprimed sites – differential impact on microtubule binding. J Biol Chem 2003;278:187–93 [DOI] [PubMed] [Google Scholar]
- 59. Marini C, Morbelli S, Cistaro A, et al. Interplay between spinal cord and cerebral cortex metabolism in amyotrophic lateral sclerosis. Brain 2018;141:2272–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Birger A, Ottolenghi M, Perez L, et al. ALS-related human cortical and motor neurons survival is differentially affected by Sema3A. Cell Death Dis 2018;9:1–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.