

Abstract
Drug resistance of Pseudomonas aeruginosa is a leading problem in hospital infections. The aim of this study was to determine the best molecular genetic discrimination method for Pseudomonas spp. isolates among 94 outpatients and inpatients and see their grouping by phenotype characteristics (biofilm formation, frequency of serotypes, pigmentation, production of different class of beta-lactamases, and susceptibility to different antibiotic classes) and genotype. The most common serotypes were P1, P6, and P11, while co-productions of pyoverdine and pyocyanin were observed in 70 % of isolates. A total of 77.66 % isolates were mostly weak and moderate biofilm producers. Isolates were susceptible to colistin (100 %), aztreonam (97.87 %), imipenem (91.49 %), doripenem (90.43 %), and meropenem (84.04 %). MICs values confirmed susceptibility to ceftazidime and cefepime and singled out meripenem as the most effective inhibitor. Most isolates were resistant to aminoglycosides and fluoroquinolones. Only two isolates produced ESBL, eight were carbapenemase producers, and five isolates produced MBLs. Twenty-nine isolates were multidrug-resistant; 82.8 % of which produced both pigments, 58.3 % were non-typeable, while the P6 and P11 serotypes were equally distributed (16.7 %). Thirteen MDR isolates were strong enzyme producers. RAPD PCR analysis using primer 272 proved the best at discriminatory fingerprinting for Pseudomonas isolates, as it allocated 12 clusters. A correlation between DNA patterns and antibiotic resistance, production of pigments, serotypes distribution, and biofilm formation was not observed, and only confirmed higher genetic heterogeneity among P. aeruginosa isolates, which suggests that other molecular methods are needed to reveal potential relations between genotypic patterns and phenotypic characteristics.
Keywords: clinical isolates, multidrug resistance, phenotypic characteristics, RAPD PCR profiling
Abstract
Antibiotska rezistencija Pseudomonas aeruginosa vodeći je problem u bolničkim infekcijama. Cilj ovoga istraživanja bio je utvrditi najbolju diskriminatorno molekularno-genetičku metodu među 94 ambulantna i bolnička Pseudomonas spp. izolata kako bi se uvidjelo njihovo grupiranje u smislu različitih fenotipskih obilježja (stvaranje biofilma, učestalost serotipova, pigmentacija, proizvodnja različitih klasa beta-laktamaza i osjetljivost na različite skupine antibiotika) u skladu s genotipom. Najčešći serotipovi bili su P1, P6 i P11, a proizvodnja i pioverdina i piocijanina primijećena je kod 70 % izolata. Ukupno 77,66 % izolata uglavnom je iskazalo slabu i umjerenu proizvodnju biofilma. Izolati su bili osjetljivi na kolistin (100 %), aztreonam (97,87 %), imipenem (91,49 %), doripenem (90,43 %) i meropenem (84,04 %). Vrijednosti MIC-ova potvrdile su podložnost izolata ceftazidimu i cefepimu, a izdvojile su meropenem kao najučinkovitiji inhibitor. Većina izolata bila je otporna na aminoglikozid i fluorokinolon. Samo dva izolata proizvela su ESBL, osam izolata sintetiziralo je karbapenemaze, a pet izolata imalo je sposobnost proizvodnje MBL-a. Dvadeset devet izolata bilo je višestruko rezistentno na antibiotike, od kojih je 82,8 % proizvodilo oba pigmenta, 58,3 % bili su netipabilni, a serotipovi P6 i P11 bili su podjednako zastupljeni među njima (16,7 %). Trinaest MDR izolata bili su snažni proizvođači enzima. RAPD PCR analiza korištenjem 272 početnica pokazala se kao najbolja diskriminatorna metoda otiskom prsta (fingerprinting) za Pseudomonas izolate, izdvajajući čak 12 različitih klastera. U ovom istraživanju nije zabilježena povezanost između DNA obrazaca i otpornosti na antibiotike, proizvodnje pigmenata, distribucije serotipova i stvaranja biofilma, što potvrđuje puno veću genetičku heterogenost unutar samih izolata P. aeruginosa, pod čim se podrazumijeva uključivanje drugih molekularnih metoda u otkrivanju potencijalnih odnosa između genetičkih obrazaca i fenotipskih obilježja.
Ključne riječi: fenotipska obilježja, klinički izolati, RAPD PCR profiliranje, višestruka otpornost
Pseudomonas aeruginosa has cosmopolitan distribution and can easily survive in moist environments. It is used in the production of various pigments such as pyocyanin (blue and green), pyoverdine (yellow-green fluorescent pigment), pyorubin (dark red), and pyomelanin (brown). It also makes normal part of the human microbiota of the skin (0–2 %), nasal mucosa (0–3.3 %), larynx (0–6.6 %), and colon (10 %), while in faecal samples it can range from 2.6 to 24 % (1). It can be found in 5 % of saliva and 80 % of burns and wounds in hospitalised patients (2) and is one of the leading causes of hospital infections, as it easily colonises the skin and mucous membrane and most often causes infection in immunocompromised patients. In the world it causes between 11 and 13.8 % of all hospital infections (3), and Serbia with its 13.3 % is no different (4) and has high mortality rates due to weakened host immune response, high virulence, and high toxicity of its enzymes and toxins.
In addition, it has developed high resistance to many antibiotic classes (4, 5) through its defence mechanisms, which involve reducing antibiotic concentration in the cell, changing the target site, and antibiotic inactivation by its own or acquired enzymes. Multidrug resistance (MDR) of P. aeruginosa is defined as non-susceptibility to at least one type of antibiotic from three or more classes of antipseudomonal antibiotics (5). This MDR to different groups or even all antibiotics is owed to a large number of mutations, horizontal gene transfers, and increased efflux, which limits treatment choices to aminoglycosides and imipenem as intravenous antibiotics (6, 7). Treatment options may further be limited to colistin (aka polymyxin E) alone (8), if the cell membrane loses porins (OprD) and is less permeable to aminoglycosides or if P. aeruginosa produces metallo-beta-lactamases (MBLs), which hydrolyse all beta-lactams except aztreonam, and certain extended-spectrum beta-lactamase (ESBLs), which modify aminoglycosides (7, 9). Six such MBL enzyme types (VIM, IMP, SPM, GIM, AIM, and, NDM), have been identified in clinical P. aeruginosa isolates in Serbia (6).
Biofilm formation is a complex process that begins with the binding of bacteria to the surface, after which they multiply, aggregate and form multilayer deposits, which could serve as a reservoir for the spreading of living cells and cause the development of chronic infections. P. aeruginosa has a natural tendency to bind to wet surfaces and form a biofilm from exopolysaccharides (EPS), nucleic acids, lipids, and ions (10), and – as such bacteria dominate the wound infections – postpone or prevent wound healing and diminish antibiotic activity (11). Prevention of biofilm formation in infections is crucial, because resistance to antibiotic increases 10–1,000 times, and P. aeruginosa in biofilm is more pathogenic than in the planktonic form (12).
Since P. aeruginosa is a major cause of hospital infections, identifying its antigen serotypes and biofilm formation ability is highly important for its epidemiological subtyping (13). However, methods such as protein profiling and pulsed-field gel electrophoresis (PFGE), which are used as gold standards for molecular discrimination of isolates, are often limited in their separation ability of Pseudomonas spp. genotypes and require additional expensive and time-consuming species-specific profiling methods (14). Consequently, they are replaced by various polymerase chain reaction (PCR) techniques, such as random amplification of polymorphic DNA (RAPD) and repetitive element sequence-based PCR (rep-PCR). These ensure reliable grouping of the same bacterial strains in corresponding clusters and ultimately simplify isolate selection for further identification (15). DNA fingerprinting is simple, inexpensive, quickly discriminates bacterial isolates, and is reliable for classification and typing of a wide range of Gram-negative bacteria (15).
The aim of our study was to determine the frequency of multidrug-resistant P. aeruginosa subtypes among outpatient and inpatient isolates from the south of Serbia and see how their phenotypic characteristics (susceptibility to different antibiotic classes, the expression of different class of beta-lactamases, frequency of serotypes, production of pigments, and biofilm formation) relate to their genotypic patterns. We also wanted to find the best molecular genetic discrimination method.
Materials and methods
Bacterial strains
Between 2013 and 2015, we collected 94 bacterial isolates, initially characterised as Pseudomonas spp., from the urine, sputum, and wound, throat, tongue, ear, and vaginal swabs of in and outpatients treated at the Clinical Hospital Centre Aleksinac.
The obtained samples were inoculated within a few hours on the following substrates: Columbia blood agar base, MacConkey agar, endo agar, tryptone soy agar, Mueller-Hinton agar (MHA), glucose, and thioglycollate broths (all purchased from Oxoid Ltd., Basingstoke, UK). Luria-Bertani (LB) medium and cetrimide agar (Lab M Limited, Bury, UK) were used for further testing. All isolates were grown overnight at 37 °C, stocked in 30 % glycerol, and stored at -80 °C until further use.
Ethical approval
The use of clinical strains was approved by the hospital’s ethics committee (decision No. 3242). Patient-identifiable information was coded and hidden from us and we had no contact with patients who gave the samples, so no informed consent was necessary for this study. None of the clinical data previously obtained were associated with the isolates of this study.
Serotyping
Pseudomonas spp. isolates were serotyped with the slide agglutination test according to the manufacturer’s instructions (Bio-Rad, Marnes-la-Coquette, France). The test set contained four polyvalent (PMA, PMF, PME, and PMC) and 16 monovalent serotypes (P1, P3, P4, and P6 from the PMA group; P7, P8, P11, and P12 from the PMF group; P2, P5, P15, and P16 from the PME group; and P9, P10, P13, and P14 from the PMC group). Agglutination was positive only if clear within two minutes. The isolates were scored as polyagglutinable, monoagglutinable, or non-typeable.
Pigmentation
Pigment production of Pseudomonas spp. isolates was tested on specialised media: pyocyanin (blue pigment) and/or pyomelanin (brown pigment) on the King agar A and pyoverdine (yellow-green pigment) on Pseudomonas fluorescein agar (all from HiMedia, Mumbai, India).
Biofilm formation
Biofilm formation was quantified with a modified version of the method described by Stepanović et al. (16). Each isolate was tested in triplicate (in three wells of 96-well microtiter plate), and all tests were carried in three separate experiments during three consecutive days. The wells of a 96-well flat-bottomed microplate were filled with a final volume of 200 μL [180 μL of Mueller-Hinton broth (MHB) and 20 μL of bacteria (5×105 CFU/mL)]. Negative control wells contained MHB only. After incubation at 35 °C for 24 h, the plates were decanted and the wells washed three times with 300 μL of phosphate buffer (1×PBS, pH 7.2, 25 °C). The remaining attached bacteria were fixed with 150 μL of methanol per well, and the plates emptied after 20 min and left to air dry. The final step was staining with 150 μL of 2 % Crystal Violet (Lach-Ner, Neratovice, Czech Republic) per well for 15 min. Excessive stain was rinsed off with running tap water. After air drying, the remaining stain was solved with 150 μL of 96 % ethanol per well. Forty-eight hours later, we measured biofilm optical density (OD) of each strain with an automated Multiskan FC reader (Flow Titertek Multiskan Plus, Flow Laboratories Co., Turku, Finland) at 570 nm. The obtained readings served to set the OD cut-off values (ODc) (three standard deviations above the average mean OD of negative control) as described elsewhere (16). Each strain was classified into one of the following categories: not a biofilm producer (OD≤ODc, category 0 or -); weak biofilm producer (OD≤ODc≤2×ODc, category 1 or +), moderate biofilm producer (2×ODc≤OD≤4×ODc, category 2 or ++), or strong biofilm producer (4×ODc≤OD, category 3 or +++).
Disk diffusion method
Antimicrobial susceptibility of the isolates was tested with the disk diffusion method following the protocol described by the Clinical and Laboratory Standards Institute (17). For this purpose we used standardised single antibiotic discs (Rosco Diagnostica, Taastrup, Denmark) at the following concentrations (μg per disc): 5 μg for ofloxacin, ciprofloxacin, and levofloxacin; 10 μg for piperacillin/tazobactam, imipenem, doripenem, meropenem, colistin, aztreonam, gentamicin, and tobramycin; and 30 μg for ceftazidime, cefepime, netilmicin, and amikacin. Readings were taken after incubation at 37 °C for 24 h and expressed in mm. The isolates were classified as susceptible (S), intermediate (I) or resistant (R).
Minimum inhibitory concentration assay
The sensitivity of our Pseudomonas spp. isolates to antibiotics was determined with a broth microdilution test in 96-well microtitre plates according to the protocol of the European Committee for Antimicrobial Susceptibility Testing (18) and the ISO 20776-1:2006 (19) standard. Clinical isolates were suspended in a saline solution and adjusted to turbidity of 0.5 per McFarland standard (Bio-Merieux, France), which corresponds to 1.0×108 CFU/mL. Antibiotics ceftazidime, cefepime, and meropenem (Sigma-Aldrich, St. Louis, MO, USA) were first dissolved in purified sterile water at stock concentration of 1000 μg/mL and then serially diluted in MHB in the following concentrations: 1–32 μg/mL (for ceftazidime and cefepime) and 1–16 μg/mL (for meropenem). Besides negative control, sterility control was also tested. All dilutions of antibiotics were done in triplicate, and the experiment was repeated twice over two consecutive days. Each well, except for the sterility control, was inoculated with 20 μL of bacterial culture (5×105 CFU/mL), reaching the final volume of 200 μL. Ten μL of 2,3,5-triphenyltetrazolium chloride (TTC, Sigma-Aldrich) was added to MHB. TTC is a colourless redox indicator that is enzymatically reduced with cellular dehydrogenase to a red metabolite 2,3,5-triphenyltetrazolium formazan if the cells are live, and metabolically active. The plates were incubated aerobically at 37 °C for 20 h. The lowest antibiotic concentration which yielded no change of colour was defined as the minimum inhibitory concentration (MIC). The results are expressed in μg/mL.
Identification of beta-lactamase-producing isolates
Extended spectrum beta-lactamase (ESBL)
The production of ESBLs was detected with the double-disk synergy test (DDST) and confirmed with the combination disk test (CDT) (20). For DDST, the discs of amoxicillin/clavulanic acid (AMC, 20/10 μg), cephalexin (CL, 30 μg), and cefotaxime (CTX, 30 μg) were placed on plates inoculated with Pseudomonas spp. isolates at the optimal distance of 17 mm. After an overnight aerobic incubation at 37 °C, the isolate was considered ESBL-positive if zones of inhibition spread toward the disc with beta-lactamase inhibitor (AMC) or any additional inhibition zone was observed. If the strain did not produce beta-lactamase, however, such characteristic changes on the plate were not observed. In case of a positive test, a confirmative CDT test was carried out by placing ceftazidime/clavulanic acid (30/10 μg) and ceftazidime (30 μg) discs. ESBL production was confirmed if the difference in the zones of inhibition around the discs with and without ESBL inhibitor were 5 mm or more.
Carbapenemases
Carbapenemase production was tested with a modified Hodge test (21). An overnight culture of E. coli ATCC 25922 (turbidity adjusted to 0.5 per McFarland standard) was inoculated on the surface of an MHA plate. A meropenem disk (10 μg) was placed at the centre of the plate, and isolates of Pseudomonas spp. were placed around the disc in radial stripes (extending from the edge of the disc to the periphery of the plate). The plates were incubated at 37 °C overnight. The test was positive if a clover leaf-like indentation was formed in the meropenem inhibition along the streaks of the tested isolate.
Metallo-beta-lactamase (MBL)
The isolates were tested for MBL production with the EDTA synergy test as described elsewhere (22). Briefly, an overnight liquid culture of the tested isolate (turbidity adjusted to 0.5 per McFarland standard) was spread on the surface of the MHA plate. Two imipenem discs (10 μg) were placed on the agar 15 mm apart and 10 μL of 0.5 EDTA was pipetted on one of the imipenem discs. After an overnight incubation at 37 °C, the expanded inhibition zone between the two discs or expansion of more than 6 mm in the imipenem/EDTA disc were interpreted as positive for MBL production.
PCR-based DNA amplifications
Genomic DNA (gDNA) was isolated from the Pseudomonas spp. following the method described by Janakiev et al. (23). The BOX (CTACGGCAAGGCGACG CTGACG), ERIC-1R (CACTTAGGGGTCCTGAATGTA), ERIC-2 (AAGTAAGTGACTGGGGTGAGCG), 272 (AGCGGGCCAA), and 208 (ACGGCCGACC) primers were rep-PCR and RAPD PCR fingerprinted (24, 25) and amplified (each in the final concentration of 10 μmol/L) in a 25 μL reaction mixture containing 1 μL of template DNA, 1.5 μL of 25 mmol/L MgCl2 (KAPA Biosystems, Wilmington, MA, USA), 2.5 μL of 10 mmol/L 10×KAPA Taq buffer, 0.5 μL of 10 mmol/L dNTP (KAPA Biosystems), and 0.1 μL of KAPA Taq polymerase (KAPA Biosystems).
The ERIC and BOX PCR reactions were done in the following steps: initial denaturation at 95 °C for 7 min, 30 cycles of denaturation at 94 °C for 1 min, primer annealing 46 °C for 1 min, and polymerisation at 65 °C for 8 min. For the 272 primer the steps were as follows: initial denaturation at 94 °C for 2 min, 35 cycles of denaturation at 94 °C for 30 s; primer annealing at 35 °C for 30 s, and polymerisation at a 72 °C for 2 min. The steps for the 208 primer were: four cycles of auto-extension (each consisting of initial denaturation at 94 °C for 5 min, primer annealing at 36 °C for 5 min, and polymerisation at 72 °C for 5 min), 30 cycles of denaturation at 94 °C for 1 min; primer annealing at 36 °C for 1 min, and polymerisation at 72 °C for 2 min. The final extension step for BOX and ERIC was done at 65 °C for 16 min and for 208 and 272 primers at 72 °C for 10 min.
From the obtained DNA fingerprints we built a dendrogram using the PyElph 1.4 software (26) and then determined clustering patterns using the unweighted pair group method with arithmetic mean (UPGMA) algorithm with a bootstrap value of 100. The position of pattern strips was checked manually.
The total DNA of selected isolates was used for 16S rRNA PCR amplification and isolate identification with universal primers UN116sF (GAGAGTTTGATCCTGGC) and UN116sR (AGGAGGTGATCCAGCCG). The reaction mixture was prepared as described above, and the conditions were the usual ones for primer annealing at 50 ºC for 1 min. The amplicons were purified on a QIAquick Gel Extraction KIT/250 column (QIAGEN GmbH, Hilden, Germany) and sequenced commercially (Macrogen, Amsterdam, Netherlands). The obtained sequences were searched for homology at the National Center for Biotechnology Information using the Basic Logical Alignment Search Tool, aligned with the ClustalW multiple sequence alignment in program BioEdit 7.1.3 (Tom Hall, North Carolina State University, Raleigh, NC, USA) and checked manually. The phylogenetic tree was constructed with the MEGA 7.0 software (Pennsylvania State University, Philadelphia, PA, USA) using the neighbour-joining method based on a pairwise distance matrix obtained with the Kimura two-parameter nucleotide substitution model. The topology of the trees was evaluated with the bootstrap resampling method with 1000 replicates.
Statistical analysis
The data obtained in this study were analysed with descriptive statistics using IBM SPSS Statistics for Windows, version 25.0 (IBM Corp., Armonk, NY, USA).
Results and discussion
Isolate origins
Of the 94 Pseudomonas spp. isolates 44 were identified in outpatients and 50 in inpatients. The predominant number of isolates came from wound swabs (46.8 %) of inpatients only. The majority of other isolates originated from urine samples (21.3 %) and throat swabs (13.8 %), mostly from outpatients. Isolates from sputum samples (9.6 %), ear (4.3 %), tongue (2.1 %), and vaginal swabs (2.1 %) were rare. We already reported about their high prevalence in urine (36.79 %) and wounds (29 %) nearly ten years ago (27).
Isolate serotypes, pigmentation, and biofilm formation
Table 1 shows our findings in terms of isolate serotypes, pigmentation, and biofilm formation. Twenty-eight isolates were non-typeable. All of the remaining identified serotypes were from polyvalent groups (PMA, PME, PMC, and PMF). Most belonged to the PMA and PMF groups, which included the P1 and P6 serotypes. The PMC group included the P9 and P10, while the PME and PMF groups P5 and P11 serotypes, respectively. The most commonly identified monovalent serotypes were P1 (17.02 %), P6 (22.34 %), and P11 (15.96 %). Co-production of pyoverdine and pyocyanin was observed in 70 % of clinical isolates, while the production of only pyoverdine in 22.3 % and of pyocyanin in 7.4 % of the isolates. Most isolates (77.66 %) expressed some biofilm formation ability, mostly weak (53.42 %, most of them originating from inpatient wounds and outpatient urine cultures) and moderate (39.73 %, most of them originating from inpatient wounds), while only five isolates were strong biofilm producers.
Table 1.
Serological identification of polyvalent and monovalent serotypes, pigmentation, and biofilm formation of Pseudomonas spp. clinical isolate of different origin
| Polyvalent and monovalent serotypes | Pigments type | Biofilm formation | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Isolate origin | PMA | PME | PMC | PMF | NT | Pyov+ Pyoc | Pyov | Pyoc | Cat. 0(-) | Cat. 1(+) | Cat. 2(++) | Cat. 3(+++) | ||
| P1 | P6 | P5 | P9 | P10 | P11 | |||||||||
| Throat | 4 | 4 | - | - | 1 | 1 | 3 | 9 | 3 | 1 | 2 | 6 | 5 | - |
| Tongue | - | 1 | - | - | 1 | - | - | 1 | 1 | - | 1 | - | 1 | - |
| Wound | 7 | 7 | 3 | 6 | - | 8 | 13 | 30 | 12 | 2 | 12 | 17 | 14 | 1 |
| Cer-vagmal canal | - | - | - | - | - | - | 2 | 1 | 1 | - | 1 | - | 1 | - |
| Ear | - | 2 | - | - | - | 1 | 1 | 2 | 1 | 1 | - | - | 4 | - |
| Sputum | 1 | 4 | - | 1 | - | 1 | 2 | 7 | 1 | 1 | 2 | 4 | 1 | 2 |
| Urine culture | 4 | 3 | - | 2 | - | 4 | 7 | 16 | 2 | 2 | 3 | 12 | 3 | 2 |
| Σtotal isloates | 16 | 21 | 3 | 9 | 2 | 15 | 28 | 66 | 21 | 7 | 21 | 39 | 29 | 5 |
NT – non-typeable; Pyov – pyoverdine; Pyoc – pyocyanin; Cat. 0 (-) – not a biofilm producer; Cat. 1 (+) – weak biofilm producer; Cat. 2 (++) – moderate biofilm producer; Cat. 3 (+++) – strong biofilm producer
These findings are consistent with earlier reports showing dominance of the P3, P6, and P11 serotypes (28, 29, 30) and pigmentation where the synthesis of pyoverdine (79.75 %) was more pronounced than the synthesis of pyocyanin (44.14 %) (27). The pigments of the clinical Pseudomonas spp. isolates participate in the development and maintenance of tissue lesions under inflammation in different ways. Therefore, the production of pigment is not only a phenotype characteristic but also an indicator of the level of pathogenicity of individual strains (27). However, a similar study in Serbia reported that only 2.45 % of 163 clinical P. aeruginosa isolates did not form a biofilm. 34.36 % were moderate, and most (39.26 %) were strong biofilm producers (31). Usually, biofilm forming is strong in invasive infections associated with the application of medical devices (such as urinary and central venous catheters or tracheostomy and endotracheal tubes) or with chronic pulmonary diseases like cystic fibrosis and chronic obstructive bronchitis, where bacteria firmly adhere to the plastic surface or viscous mucus. In our study, however, the isolates were collected from less invasive and/or superficial infections, which may explain weak to moderate biofilm formation. Similarly weak to moderate biofilm formation was reported by da Costa Lima et al. (32), whereas Nasirmoghadas et al. (33) evidenced that isolates may significantly differ in their ability to form biofilm even in the same type of infection. They reported that between multidrug-resistant and extended drug-resistant strains of P. aeruginosa isolated from patient burns, most of the isolates were weak (67 %) or moderate (22 %) biofilm producers.
Antibiotic susceptibility and enzyme production
Susceptibility to 15 antibiotics from different classes such as aminoglycosides (amikacin, gentamicin, netilmicin and tobramycin), fluoroquinolones (ofloxacin, ciprofloxacin and levofloxacin), polypeptides (colistin), penicillin combination (piperacillin/tazobactam), monobactams (aztreonam), cephalosporins (ceftazidime and cefepime), and carbapenems (doripenem, imipenem and meropenem) was tested in all isolates. Figure 1 shows that all isolates were sensitive to colistin, and most to aztreonam (97.87 %), imipenem, and doripenem (91.49 % and 90.43 %, respectively), while 84.04 % of the isolates were susceptible to meropenem and the piperacillin/tazobactam combination. These findings single out colistin as antibiotic of choice, but only as a last resort due to its neurotoxicity (34).
Figure 1.
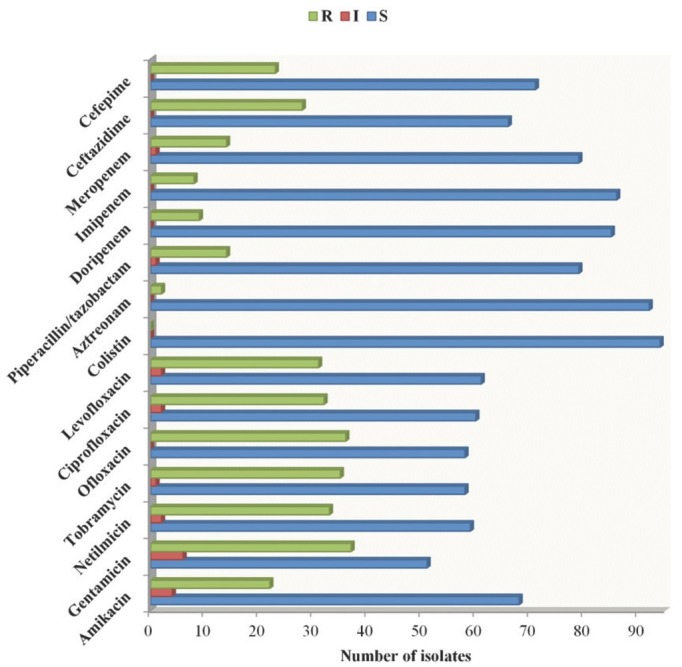
Susceptibility of clinical isolates of Pseudomonas spp. on different antibiotic classes. The isolates were classified as resistant (R), intermediate (I), or susceptible (S)
Regarding ceftazidime and cefepime susceptibility, 70.21 % and 75.53 % of the isolates were sensitive to 3rd and 4th generation cephalosporins, respectively. The highest number of resistant isolates was observed in testing with aminoglycoside [gentamicin (39.36 %), netilmicin (35.10 %) and tobramycin (37.23 %)], and fluoroquinolone [ofloxacin (38.30 %), ciprofloxacin (34.04 %) and levofloxacin (32.98 %)] classes. Twenty-three of inpatient isolates were resistant to fluoroquinolone ofloxacin (46 %) and seventeen of outpatient isolates to aminoglycoside gentamicin (38.64 %). Cefepime best inhibited Pseudomonas inpatient isolates, and ceftazidime outpatient isolates (Table 2).
Table 2.
Antibiotic susceptibility of inpatient and outpatient Pseudomonas spp. isolates
| Inpatients | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Isolate coded name | Specimen | Aminoglycosides | Fluoroquinolones | Polyp | Monob | Penicillin comb. | Carbapenems | Cephalosporins | ||||||||
| Amik | Gent | Net | Tobr | Oflox | Cipr | Levofl | Col | Aztr | Pip/taz | Dor | Imp | Mer | Ceft | Cef | ||
| 5660 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 5661 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 1183 | Urine | S | R | R | R | R | R | R | S | S | S | S | S | R | R | S |
| 2638 | Urine | R | R | R | R | R | R | R | S | S | R | R | R | R | R | R |
| 2844 | Urine | S | R | S | R | R | R | R | S | S | S | S | S | S | S | R |
| 3853 | Urine | S | R | R | R | R | R | R | S | S | S | S | S | S | R | S |
| 611 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 821 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | R | S | S |
| 971 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | R | R |
| 1087 | Wound | R | R | R | R | R | R | R | S | S | R | R | R | R | R | R |
| 1416 | Wound | S | S | S | S | R | S | S | S | S | S | S | S | S | S | S |
| 2064 | Wound | S | S | R | S | S | S | S | S | S | S | S | S | S | R | S |
| 2073 | Wound | R | R | R | R | R | R | R | S | S | S | R | R | R | R | R |
| 2581 | Wound | R | R | R | R | R | R | R | S | S | R | S | S | S | R | S |
| 2645 | Wound | R | R | R | R | R | R | R | S | S | R | R | R | R | R | R |
| 2900 | Wound | R | R | R | R | R | R | R | S | S | S | R | R | R | R | R |
| 3122 | Wound | S | S | R | S | R | S | S | S | S | S | S | S | S | S | S |
| 3322 | Wound | R | R | R | R | R | R | R | S | S | R | S | S | R | S | S |
| 3451 | Wound | S | R | R | R | R | R | R | S | S | R | S | S | S | R | S |
| 3563 | Wound | R | R | R | R | R | R | R | S | S | S | S | S | S | S | S |
| 3595 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 3658 | Wound | S | R | R | R | R | R | R | S | S | S | R | S | S | S | S |
| 3714 | Wound | S | S | S | R | S | S | S | S | R | S | S | R | R | R | R |
| 3883 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 4071 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 4082 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 4211 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | R | S |
| 4212 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 4312 | Wound | R | R | R | S | R | R | S | S | S | S | S | S | S | R | S |
| 4314 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 4354 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 4473 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 4541 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 5348 | Wound | R | R | R | R | R | R | R | S | S | R | S | S | S | S | S |
| 5774 | Wound | R | R | R | R | R | R | R | S | S | S | S | S | I | S | R |
| 5797 | Wound | R | R | R | R | R | R | R | S | S | S | S | S | S | R | S |
| 6412 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 6982 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 7543 | Wound | R | R | R | R | R | R | R | S | S | R | S | S | S | R | R |
| 7698 | Wound | R | R | I | R | S | I | S | S | S | S | S | S | S | S | S |
| 7881 | Wound | S | S | R | S | S | S | S | S | S | S | S | S | S | S | S |
| 8283 | Wound | I | I | R | R | R | R | R | S | S | S | S | S | S | S | S |
| 8702 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 8913 | Wound | R | R | R | R | R | R | R | S | S | S | S | S | S | S | S |
| 10195 | Wound | S | S | S | S | R | I | I | S | S | S | S | S | S | R | R |
| 10336 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 10708 | Wound | R | I | I | S | S | S | S | S | S | S | S | S | S | S | S |
| 10800 | Wound | S | I | S | S | S | S | S | S | S | S | R | R | S | R | S |
| 11947 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 19677 | Wound | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Outpatients | ||||||||||||||||
| Isolate coded name | Specimen | Aminoglycosides | Fluoroquinolones | Polyp | Monob | Penicillin comb. | Carbapenems | Cephalosporins | ||||||||
| Amik | Gent | Net | Tobr | Oflox | Cipr | Levofl | Col | Aztr | Pip/taz | Dor | Imip | Mer | Ceft | Cef | ||
| 2609 | Ear | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S |
| 4278 | Ear | S | I | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 5518 | Ear | S | S | S | S | S | S | S | S | S | S | S | S | R | R | R |
| 7546 | Ear | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 294 | Sputum | S | R | S | I | S | S | S | S | S | S | S | S | S | S | S |
| 2124 | Sputum | S | S | S | R | R | R | R | S | S | R | S | S | S | R | R |
| 2941 | Sputum | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 2966 | Sputum | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 3496 | Sputum | S | S | S | S | S | S | S | S | S | S | S | S | S | R | R |
| 3919 | Sputum | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 8142 | Sputum | S | R | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 9921 | Sputum | R | R | R | R | R | R | R | S | S | S | S | S | S | R | R |
| 11838 | Sputum | S | R | S | S | R | S | S | S | S | S | S | S | S | S | S |
| 1863 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 2005 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 2047 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 3238 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 3477 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 3540 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 3864 | Throat | S | I | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 4087 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 4646 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 9642 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 10412 | Throat | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 2383 | Tongue | S | R | S | S | S | S | S | S | S | R | S | S | S | S | S |
| 2967 | Tongue | I | R | R | R | S | S | S | S | S | R | S | S | S | R | R |
| 1408 | Urine | R | R | S | R | R | R | R | S | S | S | S | S | S | S | S |
| 1790 | Urine | I | R | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 2285 | Urine | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 2588 | Urine | R | R | R | R | R | R | I | S | S | S | S | S | R | S | S |
| 2597 | Urine | S | R | R | R | R | R | R | S | R | S | S | S | S | S | R |
| 2711 | Urine | R | I | R | R | S | S | R | S | S | S | S | R | S | S | R |
| 2875 | Urine | R | R | S | R | R | R | R | S | S | S | S | S | S | S | S |
| 3199 | Urine | S | R | R | R | R | R | R | S | S | S | S | S | S | S | S |
| 3214 | Urine | S | R | R | R | R | R | R | S | S | R | S | S | S | R | R |
| 3777 | Urine | S | R | R | R | R | R | R | S | S | S | R | S | R | S | R |
| 4188 | Urine | I | R | R | R | R | R | R | S | S | S | R | S | R | S | R |
| 4362 | Urine | S | S | S | S | S | S | S | S | S | S | S | S | S | R | R |
| 5586 | Urine | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 8599 | Urine | S | R | R | R | R | R | R | S | S | S | S | S | R | R | S |
| 10019 | Urine | R | R | R | R | R | R | R | S | S | I | S | S | S | S | R |
| 10600 | Urine | S | S | S | S | S | S | S | S | S | R | S | S | S | R | S |
| 2671 | Vag. Swab | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| 2689 | Vag. Swab | S | S | S | S | S | S | S | S | S | S | S | S | S | R | S |
Amik – amikacin; Gent – gentamicin; Net – netilmicin; Tobr – tobramycin; Oflox – ofloxacin; Cipr – ciprofloxacin; Levofl – levofloxacin; Col – colistin; Aztr – aztreonam; Pip/taz – piperacillin/ tazobactam; Dor – doripenem; Imip – imipenem; Mer – meropenem; Ceft – ceftazidime; Cef – cefepime. Polyp – polypeptide class; Monob – monobactams class. S – susceptible; I – intermediate susceptible; R– resistant isolates. Bolded letters represent resistant values
MICs for ceftazidime, cefepime, and meropenem confirmed these findings and singled out meripenem as the most effective inhibitor, as 72 isolates (76.6 %) were highly susceptible to meropenem with MIC below 1 μg/mL (Table 3). The most resistant isolates were obtained from wounds, urine, and sputum.
Table 3.
Minimum inhibitory concentration (MIC, expressed in μg/mL) of inpatient and outpatient Pseudomonas spp. isolates and their potential to produce beta-lactamases
| Inpatients | Outpatients | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Isolate coded name | Specimen | Mer | Ceft | Cef | Enzyme produced | Isolate name coded | Specimen | Mer | Ceft | Cef | Enzyme produced |
| 5660 | Throat | <1 | 4 | 8 | - | 2609 | Ear | <1 | <2 | <2 | - |
| 5661 | Throat | <1 | <2 | <2 | - | 4278 | Ear | <1 | <2 | <2 | - |
| 1183 | Urine | >16 | 16 | 4 | - | 5518 | Ear | >16 | 16 | 32 | MBL |
| 2638 | Urine | >16 | >32 | >32 | CRP | 7546 | Ear | <1 | <2 | <2 | - |
| 2844 | Urine | <1 | <2 | 16 | - | 294 | Sputum | <1 | <2 | <2 | - |
| 3853 | Urine | 4 | 16 | 4 | - | 2124 | Sputum | 2 | >32 | >32 | - |
| 611 | Wound | <1 | <2 | <2 | - | 2941 | Sputum | <1 | <2 | <2 | - |
| 821 | Wound | >16 | 4 | 2 | - | 2966 | Sputum | <1 | 2 | 4 | - |
| 971 | Wound | <1 | 32 | 16 | - | 3496 | Sputum | <1 | >32 | >32 | - |
| 1087 | Wound | >16 | >32 | >32 | CRP | 3919 | Sputum | <1 | 4 | 4 | - |
| 1416 | Wound | <1 | 4 | 4 | - | 8142 | Sputum | <1 | 4 | 4 | - |
| 2064 | Wound | <1 | 16 | <2 | - | 9921 | Sputum | 2 | 16 | 16 | - |
| 2073 | Wound | >16 | >32 | >32 | CRP | 11838 | Sputum | <1 | <2 | <2 | - |
| 2581 | Wound | <1 | 16 | 8 | MBL | 1863 | Throat | <1 | <2 | <2 | - |
| 2645 | Wound | >16 | >32 | >32 | CRP | 2005 | Throat | <1 | 2 | 2 | - |
| 2900 | Wound | >16 | >32 | >32 | CRP | 2047 | Throat | <1 | <2 | <2 | - |
| 3122 | Wound | <1 | 8 | 8 | - | 3238 | Throat | <1 | 4 | 4 | - |
| 3322 | Wound | 16 | 2 | 2 | MBL | 3477 | Throat | <1 | 2 | 2 | - |
| 3451 | Wound | <1 | 16 | 8 | - | 3540 | Throat | <1 | 4 | 2 | - |
| 3563 | Wound | <1 | 4 | 4 | - | 3864 | Throat | <1 | 2 | 2 | - |
| 3595 | Wound | <1 | 4 | 4 | - | 4087 | Throat | <1 | 4 | 4 | - |
| 3658 | Wound | <1 | 4 | 2 | - | 4646 | Throat | <1 | 2 | 2 | - |
| 3714 | Wound | >16 | 32 | >32 | MBL | 9642 | Throat | <1 | <2 | <2 | - |
| 3883 | Wound | 2 | <2 | <2 | - | 10412 | Throat | <1 | 4 | 4 | - |
| 4071 | Wound | <1 | 4 | 4 | - | 2383 | Tongue | <1 | 4 | 4 | - |
| 4082 | Wound | <1 | 4 | 4 | - | 2967 | Tongue | <1 | 16 | 32 | - |
| 4211 | Wound | <1 | 16 | 8 | - | 1408 | Urine | <1 | <2 | <2 | - |
| 4212 | Wound | <1 | 4 | 4 | - | 1790 | Urine | <1 | 4 | 4 | - |
| 4312 | Wound | <1 | 16 | 2 | - | 2285 | Urine | <1 | 2 | 2 | - |
| 4314 | Wound | <1 | 2 | 2 | - | 2588 | Urine | 16 | 2 | 4 | - |
| 4354 | Wound | <1 | 2 | 2 | - | 2597 | Urine | <1 | 4 | 16 | ESBL |
| 4473 | Wound | <1 | 8 | 8 | - | 2711 | Urine | <1 | 4 | 16 | MBL |
| 4541 | Wound | <1 | <2 | <2 | - | 2875 | Urine | <1 | 2 | 2 | - |
| 5348 | Wound | <1 | 2 | 4 | - | 3199 | Urine | <1 | 4 | 4 | - |
| 5774 | Wound | 4 | 4 | 16 | - | 3214 | Urine | 1 | >32 | 16 | - |
| 5797 | Wound | 2 | >32 | 2 | - | 3777 | Urine | >16 | 4 | 16 | CRP |
| 6412 | Wound | <1 | 2 | 2 | - | 4188 | Urine | >16 | 4 | 32 | CRP |
| 6982 | Wound | <1 | <2 | <2 | - | 4362 | Urine | <1 | 16 | 16 | - |
| 7543 | Wound | 1 | 16 | 16 | ESBL | 5586 | Urine | <1 | <2 | <2 | - |
| 7698 | Wound | <1 | 4 | 4 | - | 8599 | Urine | >16 | 32 | 8 | - |
| 7881 | Wound | <1 | 2 | 2 | - | 10019 | Urine | <1 | 8 | 16 | - |
| 8283 | Wound | <1 | <2 | <2 | - | 10600 | Urine | <1 | 16 | 4 | - |
| 8702 | Wound | <1 | <2 | <2 | - | 2671 | Vag. swab | <1 | 2 | 2 | - |
| 8913 | Wound | <1 | <2 | <2 | - | 2689 | Vag. swab | <1 | 16 | 8 | - |
| 10195 | Wound | <1 | >32 | >32 | - | ||||||
| 10336 | Wound | <1 | <2 | <2 | - | ||||||
| 10708 | Wound | <1 | 4 | 2 | - | ||||||
| 10800 | Wound | <1 | 32 | 8 | CRP | ||||||
| 11947 | Wound | <1 | 2 | 2 | - | ||||||
| 19677 | Wound | <1 | 4 | 4 | - |
Mer – meropenem (S<2, I4-8, R>8); Ceft – ceftazidime (S<8; R>8); Cef – cefepime (S<8; R>8); CRP – carbapenemase-producnig Pseudomonas sp.; MBL – metallo-beta-lactamase; ESBL – extended spectrum beta-lactamase. Bolded numbers represent resistant MIC values. The results are expressed in μg/mL
Only two isolates – one from inpatient wound and the other from outpatient urine – produced ESBL, which is the likely reason for resistance to piperacillin/tazobactam, cephalosporins, and aztreonam (Tables 2 and 3). Eight isolates – five wound and three urine – produced carbapenemases. All of these inpatient wound isolates were resistant to carbapenems, and all isolates showed multidrug resistance, except 10800 (Table 2). Five isolates produced MBLs (three from inpatient wounds and two from outpatient ear and urine specimens) (Table 3). Two wound isolates were resistant to the piperacillin/tazobactam combination, while all isolates were susceptible to doripenem (Table 2).
Inpatient isolates generally showed much stronger resistance than outpatient, which is in line with earlier reports (35). Similar was also the prevalence of aminoglycoside-resistant P. aeruginosa isolates (around 30 %) reported earlier for tobramycin and amikacin (36, 37). In contrast, a group of Bulgarian scientists (38) reported more than two times higher resistance to gentamicin (79.7 %) and netilmicin (69.6 %) than we have. It is encouraging, though, that susceptibility to colistin, meropenem, and imipenem has not dropped in Serbia over the last five years, judging by a 2015 report (4). However, resistance to fluoroquinolones seems to have increased, most likely as a consequence of enhanced antibiotic extrusion through efflux pump as reported before (39). Also, frequent use of ceftazidime and cefepime has resulted in associated resistance and limited use (40). A different rate of resistance to ceftazidime has been reported across Europe, including patients from Lithuania (13.9 %), Italy (13.4 %), Spain (15 %), Greece (25.5 %) and Belgium (28.5 %) (41). Lutz et al. (42) reported resistance to ceftazidime and meropenem of 14.1 % and 9.4 %, respectively, which proved meropenem to be the best antibiotic, as we have. The difference in the resistance to ceftazidime and cefepime was not significant in many studies. Most often it was attributed to intrinsic resistance that Pseudomonas isolates naturally possess, usually utilising enzymes to destroy the drug. Another reason is the MexCD-OprJ efflux system, which is selective for the 4th generation of cephalosporins (43). Extended- spectrum beta-lactamases are responsible for the hydrolysis of most penicillins, extended-spectrum cephalosporins, and aztreonam and their production also differs in some parts of the world (44). In Western Europe, the production of ESBL does not exceed 1 %, while it is much larger in China and other parts of the world (7), which was not the case in our study. The use of carbapenems, as the only choice against ESBL-producing isolates, leads to an increased resistance to antibiotics from the 4th generation of cephalosporins. Thus, the detection of carbapenemase is of great importance for proper therapy administration as well as infection control. Although discovered relatively early, carbapenemases are a major problem in the world, since their encoding genes, mostly found on transmissible genetic elements, are associated with other resistance genes, which leads to uncontrolled spread of resistance (45). The occurrence of metallo-beta-lactamase-producing isolates in hospitals and their spreading among bacteria is a serious problem for future therapy. Low prevalence of MBL producing P. aeruginosa isolates was reported in the countries of the Balkan region, but these strains belonged to the epidemic clones spread all over the Mediterranean Europe (7). In addition, Serbia and the entire Balkan region confirmed the presence of the New Delhi MBL 1 (NDM-1) in P. aeruginosa clinical isolates for the first time nearly a decade ago (6).
Relationship between antibiotic resistance/susceptibility and phenotypic characteristics of clinical isolates
Table 4 shows that 29 (18 inpatient and 11 outpatient) of our 94 isolates were resistant to up to three or more antibiotic classes. Fourteen originated from wounds (48.3 %) and 12 from urine (41.4 %), while only a few came from sputum and tongue. Twenty-four MDR isolates (82.8 %) produced both pyoverdine and pyocyanin (no other pigments were observed), all were non-typeable, while the P6 and P11 serotypes were equally distributed among all MDR isolates. Nine of the MDR isolates – mostly inpatient wound – were moderate biofilm producers, while 12 showed weak biofilm formation. Only one from inpatient urine formed a strong biofilm. As many as 13 MDR isolates (44.8 %) produced antibiotic-metabolising enzymes, while only two were in the group of isolates resistant to up to two antibiotic classes (Tables 3 and 4). Among these, only five (35.7 %) had the P6 serotype and produced moderate or weak biofilm and both pigments. Six isolates from the same group produced only pyoverdine and weak biofilm or none at all. In contrast, most isolates resistant to one antibiotic class were non-typeable (six isolates, 42.9 %) and had the P11 serotype (four isolates, 28.6 %). Most were moderate or strong biofilm- and pigment producers (Table 4). Thirty-seven isolates (39.4 %) were completely susceptible to all tested antibiotic classes. P1 and P6 were the most common monovalent serotypes among them (27.7 % and 24.3 %, respectively), with weak or non-biofilm formation, while the production of both pigments was again predominant.
Table 4.
Multidrug resistant Pseudomonas spp. isolates and their pigmentation, serotype, and biofilm formation potential
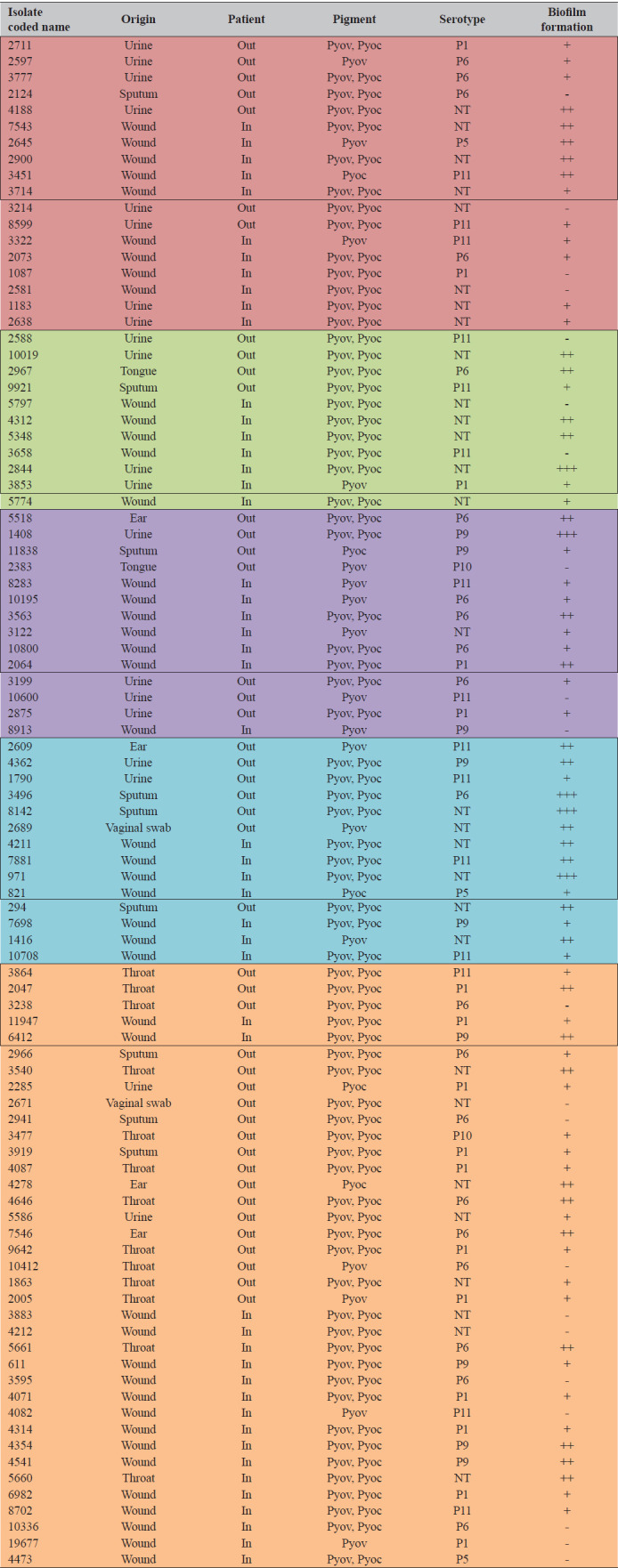 |
Pyov – Pyoverdine; Pyoc – Pyocyanin; NT – non-tippable; No biofilm producer (category -); Weak biofilm producer (category +); Moderate biofilm producer (category ++); Strong biofilm producer (category +++); Out – outpatient; In – inpatient;  Resistant up to four antibiotic classes or more;
Resistant up to four antibiotic classes or more;  Resistant up to three antibiotic classes;
Resistant up to three antibiotic classes;  Resistant up to two antibiotic classes;
Resistant up to two antibiotic classes;  Resistant up to one antibiotic class;
Resistant up to one antibiotic class;  Susceptible to all tested antibiotic classes. Framed isolates have been selected for molecular genetic characterization
Susceptible to all tested antibiotic classes. Framed isolates have been selected for molecular genetic characterization
In general, no direct correlation was observed between antibiotic susceptibility and virulence-associated phenotypic characteristics, serotype distribution in particular. In studies described earlier, P6 and P11 were the most abundant serogroups in all types of P. aeruginosa-caused infections (46), while the P1 serotype isolates showed susceptibility to the majority of the tested antibiotics (47), which was in accordance with our results. Thrane et al. (48) showed that serotypes do not correlate to antibiotic resistance, as they switch through recombination and can mislead isolate antibiotic susceptibility classification. Similar lack of correlation between antibiotic resistance and pigment production has been established by Finlayson and Brown (49). There is, however, a correlation between biofilm formation and the pattern of antibiotic resistance/susceptibility. Evidence by Corehtash et al. (50) that resistant isolates produce moderate or strong biofilm seems to corroborate our own findings.
Antibiotic resistance/susceptibility in relation to molecular genetic characterisation
Based on the results of pigmentation, serotyping, biofilm formation, enzyme production, and antibiotic resistance/susceptibility, a total of 45 isolates were selected {Table 4 [Group I – resistant to up to four antibiotic classes or more (groups 1-1 to 1-10); Group II – resistant to up to three antibiotic classes (groups 2-1 to 2-10); Group III – resistant to up to two antibiotic classes (groups 3-1 to 3-10); Group IV – resistant to one antibiotic class (groups 4-1 to 4-10); and Group V – susceptible to all tested antibiotic classes (groups 5-1 to 5-5)]} for further rep-PCR and RAPD PCR molecular genetic analysis (Table 5) to see it genetic clusterisation of different isolates in relation to phenotypic characteristics.
Table 5.
Selected isolates for further molecular genetic analysis
| Group I | Group II | Group III | Group IV | Group V | |||||
|---|---|---|---|---|---|---|---|---|---|
| Resistant up to 4 antibiotic classes or more | Resistant up to 3 antibiotic classes | Resistant up to 2 antibiotic classes | Resistant up to 1 antibiotic class | Susceptible to all tested antibiotic classes | |||||
| Isolate coded name | Group cipher | Isolate coded name | Group cipher | Isolate coded name | Group cipher | Isolate coded name | Group cipher | Isolate coded name | Group cipher |
| 2711 | 1-1 | 5797 | 2-1 | 8283 | 3-1 | 2609 | 4-1 | 3864 | 5-1 |
| 7543 | 1-2 | 4312 | 2-2 | 5518 | 3-2 | 821 | 4-2 | 2047 | 5-2 |
| 2597 | 1-3 | 5348 | 2-3 | 10195 | 3-3 | 4362 | 4-3 | 6412 | 5-3 |
| 3714 | 1-4 | 10019 | 2-4 | 3563 | 3-4 | 1790 | 4-4 | 3238 | 5-4 |
| 2645 | 1-5 | 3658 | 2-5 | 3122 | 3-5 | 7881 | 4-5 | 11947 | 5-5 |
| 3777 | 1-6 | 2967 | 2-6 | 10800 | 3-6 | 4211 | 4-6 | ||
| 2900 | 1-7 | 2844 | 2-7 | 2064 | 3-7 | 3496 | 4-7 | ||
| 3451 | 1-8 | 3853 | 2-8 | 11838 | 3-8 | 8142 | 4-8 | ||
| 2124 | 1-9 | 2588 | 2-9 | 1408 | 3-9 | 971 | 4-9 | ||
| 4188 | 1-10 | 9921 | 2-10 | 2383 | 3-10 | 2689 | 4-10 | ||
Primers 272 and 208, which were used for the RAPD PCR, created 3 to 15 fingerprinting patterns, whose range differed from 100 bp to 5000 bp. These results show that different primers within the cluster with the largest number of isolates yield similar grouping (Figure 2). Primer 272 allocated 12 clusters, while 208 allocated eight clusters. Extraordinary heterogeneity was noticed with both primers within the clusters. With primer 208 the largest cluster was extremely heterogeneous and included all but the fourth group of tested isolates was excluded. With primer 272, that same cluster was divided into as many as seven clusters. For both RAPD primers used, there was a noticeable pattern uniformity among different groups of antibiotic resistance. The case in point is pattern uniformity of the first, second, fourth, and fifth group. Similar uniformity was observed for isolate patterns of the third, first, and second group. In contrast, pattern dissimilarity (based on genetic distances) was observed between the isolates from the fourth and second group, especially with primer 272. The most genetically distant and therefore most diverse patterns were obtained for the isolates from the susceptible group, in the form of separate monophyletic branch for isolates 5-1 (primer 272) and 5-4 (primer 208).
Figure 2.
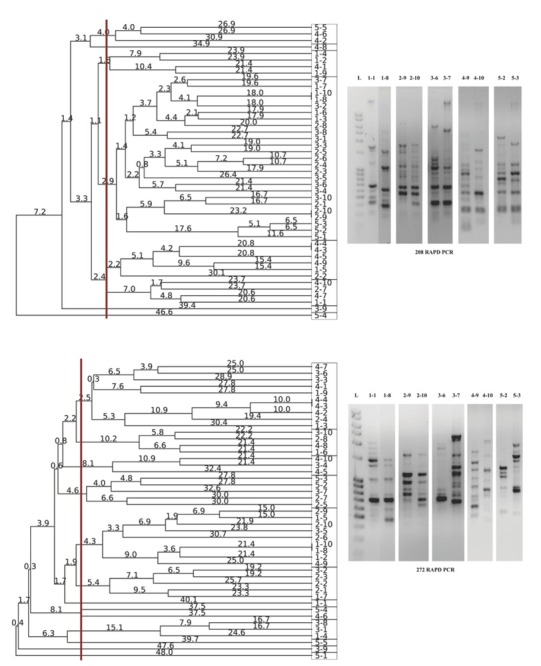
Dendrogram obtained by UPGMA analysis of the selected Pseudomonas isolates using 208 (top) and 272 (bottom) primers, based on RADP profile (right). Genetic distances were presented with numbers placed on the branches. Rectangles are representing the isolate grouping according to obtained RAPD patterns
Additionally, BOX and ERIC primers used in rep-PCR analysis created between five and 17 fingerprinting patterns of different sizes, ranging from 100 bp to 5000 bp. BOX PCR allocated a total of 10 clusters, indicating a good primer-discriminatory power, while ERIC PCR provided a total of eight clusters (Figure 3). As in RAPD analysis, the BOX PCR clusters were extremely heterogeneous, except for a few homogeneous clusters. The results of the BOX PCR analysis, like in RADP profiling, showed pattern uniformity of different antibiotic resistance groups, but certain clusters were heterogeneous. For instance, BOX PCR split the largest cluster obtained with primer 272 into four separate clusters. Pattern uniformity between isolates from the same resistance group within some clusters was also verified. The similarity between BOX and 272 primers was observed in terms of pattern uniformity between isolates from the first MDR group and those from the second and fourth group, as well as between the isolates from the fourth and fifth group. Another correlation was observed in pattern dissimilarity between isolates from the fourth and the second group, while isolate 5-1 was the most distant one genetically, as in BOX PCR analysis. ERIC PCR analysis generated more homogeneous isolate clustering from different antibiotic groups (mainly the second and the third group), and allocated a total of eight clusters. Unlike other tests, ERIC PCR showed pattern uniformity between the most susceptible isolates (the fifth group) and the second, third, and fourth group. Since RAPD272 PCR analysis provided the best characterisation with 12 clusters, we selected a few representative isolates from each cluster for 16S rRNA gene identification and all were identified as P. aeruginosa strains, which confirmed their genetic heterogeneity (Figure 4).
Figure 3.
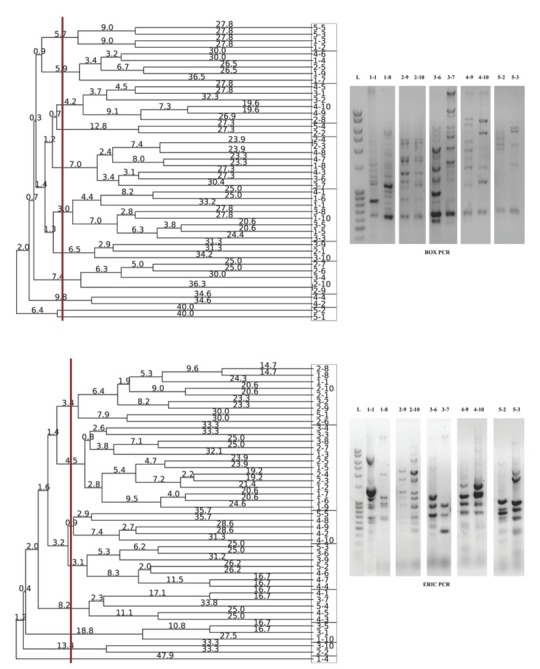
Dendrogram obtained by UPGMA analysis of the selected Pseudomonas isolates using BOX (top) and ERIC (bottom) primers, based on rep-PCR profile (right). Genetic distances were presented with numbers placed on the branches. Rectangles are representing the isolate grouping according to obtained rep-PCR patterns
Figure 4.
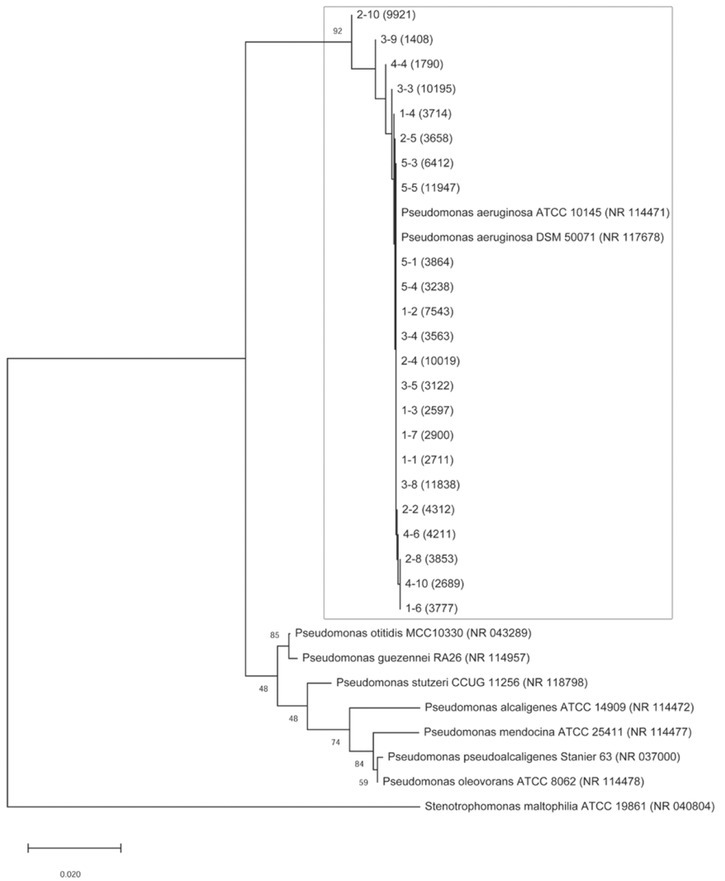
Phylogenetic relationships of Pseudomonas aeruginosa clinical isolates from all 12 RAPD272 clusters based on the sequence of 16S rRNA. Phylogenetic tree was constructed by the neighbour-joining method and the distances were calculated with the Kimura two-parameter model. Bootstrap values are given for each node, with 1000 replicates. Stenotrophomonas maltophilia was used as an outgroup. The horizontal bar indicates a genetic distance of 0.02
The use of 272 and 208 primers was successful in discrimination of different Pseudomonas isolates described before (28). Although Nagaveni et al. (51) showed that ERIC PCR provided an exceptionally qualitative discrimination for pseudomonads, the results of our and similar studies (52, 53), showed the advantage of BOX PCR, as it confirmed the results obtained with 272 RADP PCR.
We did not find a correlation between RAPD and rep-PCR patterns and antibiotic resistance/susceptibility or other phenotype characteristics (production of pigments, distribution of serotypes and biofilm formation). The uniformity of genetic patterns of P. aeruginosa isolates in different antibiotic groups suggests that other molecular methods such as whole genome sequencing should also be used to find a correlation with other phenotype characteristics.
Conclusion
This is the first comprehensive study of genotype and phenotype characteristics of P. aeruginosa isolates from Southern Serbia. RAPD PCR analysis with the 272 primer proved to be the best discriminatory fingerprinting technique. Although we did not find any correlation between genotype patterns and phenotype characteristics (antibiotic resistance, pigmentation, serotypes, and biofilm formation), our findings call for other molecular combinations and a larger number of isolates to reveal potential connections between them.
Acknowledgments
This work was supported by the Ministry of Education, Science and Technological Development of Serbia [Contract No. 451-03-68/2020-14/200178]. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. We are grateful to Ms Milica Milovanović for proofreading the manuscript.
Footnotes
Conflict of interest
None to declare.
References
- 1.Lister PD, Wolter DJ, Hanson ND. Antibacterial-resistant Pseudomonas aeruginosa: clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin Microbiol Rev. 2009:582–610. doi: 10.1128/CMR.00040-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Brooks FG, Btel JS, Morse SA. Jawetz, Melnick and Adelberg’s Medical Microbiology. 21st ed. Stamford Connecticut (USA): Appleton and Lange; 1998. [Google Scholar]
- 3.Driscoll JA, Brody SL, Kollef MH. The epidemiology, pathogenesis and treatment of Pseudomonas aeruginosa infections. Drugs. 2007;67:351–68. doi: 10.2165/00003495-200767030-00003. [DOI] [PubMed] [Google Scholar]
- 4.Stanković Nedeljković N, Tiodorović B, Kocić B, Ćirić V, Milojković M, Waisi H.. Pseudomonas aeruginosa serotypes and resistance to antibiotics from wound swabs. Vojnosanit Pregl. 2015;72:996–1003. doi: 10.2298/vsp131224108s. [DOI] [PubMed] [Google Scholar]
- 5.Magiorakos AP, Srinivasan A, Carey RB, Carmeli Y, Falagas ME, Giske CG, Harbarth S, Hindler JF, Kahlmeter G, Olsson-Liljequist B, Paterson DL, Rice LB, Stelling J, Struelens MJ, Vatopoulos A, Weber JT, Monnet DL. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18:268–81. doi: 10.1111/j.1469-0691.2011.03570.x. [DOI] [PubMed] [Google Scholar]
- 6.Jovcic B, Lepsanovic Z, Suljagic V, Rackov G, Begovic J, Topisirovic L, Kojic M. Emergence of NDM-1 metallo-β-lactamase in Pseudomonas aeruginosa clinical isolates from Serbia. Antimicrob Agents Chemother. 2011;55:3929–31. doi: 10.1128/AAC.00226-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sardelic S, Bedenic B, Colinon-Dupuich C, Orhanovic S, Bosnjak Z, Plecko V, Cournoyer B, Rossolini GM. Infrequent finding of metallo-β-lactamase VIM-2 in carbapenem-resistant Pseudomonas aeruginosa strains from Croatia. Antimicrob Agents Chemother. 2012;56:2746–9. doi: 10.1128/AAC.05212-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bert F, Lambert-Zechovsky N. Antibiotic resistance patterns in Pseudomonas aeruginosa an 8-year surveillance study in French hospital. Int J Antimicrob Agents. 1997;9:107–12. doi: 10.1016/S0924-8579(97)00033-2. [DOI] [PubMed] [Google Scholar]
- 9.Cornaglia G, Giamarellou H, Rossolini GM. Metallo-β-lactamases: a last frontier for β-lactams? Lancet Infect Dis. 2011;11:381–93. doi: 10.1016/S1473-3099(11)70056-1. [DOI] [PubMed] [Google Scholar]
- 10.Coenye T, Nelis HJ.. In vitro and in vivo model systems to study microbial biofilm formation. J Microbiol Methods. 2010;83:89–105. doi: 10.1016/j.mimet.2010.08.018. [DOI] [PubMed] [Google Scholar]
- 11.Kirketerp-Møller K, Jensen PØ, Fazli M, Madsen KG, Pedersen J, Moser C, Tolker-Nielsen T, Høiby N, Givskov M, Bjarnsholt T. Distribution, organization, and ecology of bacteria in chronic wounds. J Clin Microbiol. 2008;46:2717–22. doi: 10.1128/JCM.00501-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Woo JK, Webb JS, Kirov SM, Kjelleberg S, Rice SA. Biofilm dispersal cells of a cystic fibrosis Pseudomonas aeruginosa isolate exhibit variability in functional traits likely to contribute to persistent infection. FEMS Immunol Med Microbiol. 2012;66:251–64. doi: 10.1111/j.1574-695X.2012.01006.x. [DOI] [PubMed] [Google Scholar]
- 13.Raymond CK, Sims EH, Kas A, Spencer DH, Kutyavin TV, Ivey RG, Zhou Y, Kaul R, Clendenning JB, Olson MV. Genetic variation at the O antigen biosynthetic locus in Pseudomonas aeruginosa. J Bacteriol. 2002;184:3614–22. doi: 10.1128/JB.184.13.3614-3622.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Syrmis MW, O’Carroll MR, Sloots TP, Coulter C, Wainwright CE, Bell SC, Nissen MD. Rapid genotyping of Pseudomonas aeruginosa isolates harbored by adult and pediatric patients with cystic fibrosis using repetitive-element-based PCR assays. J Med Microbiol. 2004;53:1089–96. doi: 10.1099/jmm.0.45611-0. [DOI] [PubMed] [Google Scholar]
- 15.Gevers D, Huys G, Swings J. Applicability of rep-PCR fingerprinting for identification of Lactobacillus species. FEMS Microbiol Lett. 2001;205:31–6. doi: 10.1111/j.1574-6968.2001.tb10921.x. [DOI] [PubMed] [Google Scholar]
- 16.Stepanović S, Ćirković I, Mijač V, Švabić-Vlahović M. Influence of the incubation temperature, atmosphere and dynamic conditions on biofilm formation by Salmonella spp. Food Microbiol. 2003;20:339–43. doi: 10.1016/S0740-0020(02)00123-5. [DOI] [Google Scholar]
- 17.Wikler MA, Cockerill FR, Craig WA, Dudley MN, Eliopoulos GM, Hecht DW, Hindler JF, Ferraro MJ, Swenson JM, Low DE, Sheehan DJ. M02-A12: Performance Standards for Antimicrobial Disk Susceptibility Tests; Approved Standard. Wayne Pennsylvania (USA): Clinical and Laboratory Standards Institute; 2015. 12th ed. [Google Scholar]
- 18.The European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint tables for interpretation of MICs and zone diameters. Vers. 7.1. 2017 http://www.eucast.org [displayed 28 July 2020]. Available at. [Google Scholar]
- 19.ISO (International Organization for Standardization). Clinical laboratory testing and in vitro diagnostic test systems–Susceptibility testing of infectious agents and evaluation of performance of antimicrobial susceptibility test devices–Part 1: Reference method for testing the in vitro activity of antimicrobial agents against rapidly growing aerobic bacteria involved in infectious diseases. ISO 20776-1. 2006 https://www.iso.org/standard/41630.html 19 [displayed 28 July 2020] Available at. [Google Scholar]
- 20.The European Committee on Antimicrobial Susceptibility Testing (EUCAST). EUCAST guideline for the detection of resistance mechanisms and specific resistances of clinical and/or epidemiological importance. Vers. 2.0. 2017 http://www.eucast.org/resistance_mechanisms/ [displayed 28 July 2020]. Available at. [Google Scholar]
- 21.Fonseca AP, Extremina C, Fonseca AF, Sousa JC. Effect of subinhibitory concentration of piperacillin/tazobactam on Pseudomonas aeruginosa. J Med Microbiol. 2004;53:903–10. doi: 10.1099/jmm.0.45637. [DOI] [PubMed] [Google Scholar]
- 22.Jesudason MV, Kandathil AJ. Balaji. Comparison of two methods to detect carbapenemase & metallo-beta-lactamase production in clinical isolates. Indian J Med Res. 2005;121:780–3. PMID: 16037624. [PubMed] [Google Scholar]
- 23.Janakiev T, Dimkić I, Bojić S, Fira D, Stanković S, Berić T. Bacterial communities of plum phyllosphere and characterization of indigenous antagonistic Bacillus thuringiensis R3/3 isolate. J Appl Microbiol. 2019;128:528–43. doi: 10.1111/jam.14488. [DOI] [PubMed] [Google Scholar]
- 24.Dawson SL, Fry JC, Dancer BN. A comparative evaluation of five typing techniques for determining the diversity of fluorescent pseudomonads. J Microbiol Methods. 2002;50:9–22. doi: 10.1016/S0167-7012(02)00003-9. [DOI] [PubMed] [Google Scholar]
- 25.Mahenthiralingam E, Campbell ME, Foster J, Lam JS, Speert DP. Random amplified polymorphic DNA typing of Pseudomonas aeruginosa isolates recovered from patients with cystic fibrosis. J Clin Microbiol. 1996;34:1129–35. doi: 10.1128/JCM.34.5.1129-1135.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pavel AB, Vasile CI. PyElph-a software tool for gel images analysis and phylogenetics. BMC Bioinformatics. 2012;13:9. doi: 10.1186/1471-2105-13-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Stanković Nedeljković N, Kocić B, Tiodorović B, Branković S, Mladenović Antić S. Serotipizacija i analiza vrsta proizvedenih pigmenata kliničkih izolata Pseudomonas aeruginosa [Serotyping and analysis of produced pigments kinds by Pseudomonas aeruginosa clinical isolates, in Serbian] Vojnosanit Pregl. 2011;68:923–9. doi: 10.2298/vsp1111923s. [DOI] [PubMed] [Google Scholar]
- 28.Hafiane A, Ravaoarinoro M. Characterization of Pseudomonas aeruginosa isolated from cystic fibrosis patients by different typing methods. Pathol Biol. 2011;59:109–14. doi: 10.1016/j.patbio.2009.06.002. [DOI] [PubMed] [Google Scholar]
- 29.Estahbanati HK, Kashani PP, Ghanaatpisheh F. Frequency of Pseudomonas aeruginosa serotypes in burn wound infections and their resistance to antibiotics. Burns. 2002;28:340–8. doi: 10.1016/S0305-4179(02)00024-4. [DOI] [PubMed] [Google Scholar]
- 30.Loureiro MM, de Moraes BA, Mendonca VL, Quadra MR, Pinheiro GS, Asensi MD.. Pseudomonas aeruginosa Study of antibiotic resistance and molecular typing in hospital infection cases in a neonatal intensive care unit from Rio de Janeiro City, Brazil. Mem Inst Oswaldo Cruz. 2002;97:387–94. doi: 10.1590/S0074-02762002000300020. [DOI] [PubMed] [Google Scholar]
- 31.Vasiljević Z, Jovčić B, Ćirković I, Đukić S. An examination of potential differences in biofilm production among different genotypes of Pseudomonas aeruginosa. Arch Biol Sci. 2014;66:117–21. doi: 10.2298/ABS1401117V. [DOI] [Google Scholar]
- 32.da Costa Lima JL, Alves LR, de Araujo Jacome PRL, Neto JPB, Maciel MAV, de Morais MMC. Biofilm production by clinical isolates of Pseudomonas aeruginosa and structural changes in LasR protein of isolates non biofilm-producing. Braz J Infect Dis. 2018;22:129–36. doi: 10.1016/j.bjid.2018.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Nasirmoghadas P, Yadegari S, Moghim S, Esfahani BN, Fazeli H, Poursina F, Hosseininassab SA, Safaei HG. Evaluation of biofilm formation and frequency of multidrug-resistant and extended drug-resistant strain in Pseudomonas aeruginosa isolated from burn patients in Isfahan. Adv Biomed Res. 2018;7:61. doi: 10.4103/abr.abr_37_17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Falagas ME, Kasiakou SK. Toxicity of polymyxins: a systematic review of the evidence from old and recent studies. Crit Care. 2006;10:R27. doi: 10.1186/cc3995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Livermore DM. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa our worst nightmare? Clin Infect Dis. 2002;34:634–40. doi: 10.1086/338782. [DOI] [PubMed] [Google Scholar]
- 36.Higgins PG, Fluit AC, Milatovic D, Verhoef J, Schmitz FJ. Mutations in GyrA, ParC, MexR and NfxB in clinical isolates of Pseudomonas aeruginosa. Int J Antimicrob Agents. 2003;21:409–13. doi: 10.1016/S0924-8579(03)00009-8. [DOI] [PubMed] [Google Scholar]
- 37.Teixeira B, Rodulfo H, Carreno N, Guzman M, Salazar E, De Donato MD. Aminoglycoside resistance genes in Pseudomonas aeruginosa isolates from Cumana, Venezuela. Rev Inst Med Trop Sao Paulo. 2016;58:13. doi: 10.1590/S1678-9946201658013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Strateva T, Ouzounova-Raykova V, Markova B, Todorova A, Marteva-Proevska Y, Mitov I. Problematic clinical isolates of Pseudomonas aeruginosa from the university hospitals in Sofia, Bulgaria: current status of antimicrobial resistance and prevailing resistance mechanisms. J Med Microbiol. 2007;56:956–63. doi: 10.1099/jmm.0.46986-0. [DOI] [PubMed] [Google Scholar]
- 39.Masuda N, Sakagawa E, Ohya S, Gotoh N, Tsujimoto H, Nishino T. Substrate specificities of MexAB-OprM, MexCDOprJ, and MexXY-OprM efflux pumps in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2000;44:3322–7. doi: 10.1128/AAC.44.12.3322-3327.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Goossens H. Susceptibility of multi-drug-resistant Pseudomonas aeruginosa in intensive care units: results from the European MYSTIC study group. Clin Microbiol Infect. 2003;9:980–3. doi: 10.1046/j.1469-0691.2003.00690.x. [DOI] [PubMed] [Google Scholar]
- 41.Adukauskienė D, Vitkauskaitė A, Skrodenienė E, Dambrauskienė A, Vitkauskienė A. Changes in antibiotic resistance level of nosocomial Pseudomonas aeruginosa isolates in the largest university hospital of Lithuania. Medicina. 2011;47:39. doi: 10.3390/medicina47050039. [DOI] [PubMed] [Google Scholar]
- 42.Lutz L, Pereira DC, Paiva RM, Zavascki AP, Barth AL. Macrolides decrease the minimal inhibitory concentration of anti-pseudomonal agents against Pseudomonas aeruginosa from cystic fibrosis patients in biofilm. BMC Microbiol. 2012;12:196. doi: 10.1186/1471-2180-12-196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Van Eldere J. Multicentre surveillance of Pseudomonas aeruginosa susceptibility patterns in nosocomial infections. J Antimicrob Chemother. 2003;51:347–52. doi: 10.1093/jac/dkg102. [DOI] [PubMed] [Google Scholar]
- 44.Singleton A, Cluck D. The pharmacists role in treating extended-spectrum beta-lactamase infections. US Pharmacist. 2019;44:HS2–6. [Google Scholar]
- 45.Biswas S, Brunel JM, Dubus JC, Reynaud-Gaubert M, Rolain JM. Colistin: an update on the antibiotic of the 21st century. Expert Rev Anti Infect Ther. 2012;10:917–34. doi: 10.1586/eri.12.78. [DOI] [PubMed] [Google Scholar]
- 46.Tielen P, Narten M, Rosin N, Biegler I, Haddad I, Hogardt M, Neubauer R, Schobert M, Wiehlmann L, Jahn D. Genotypic and phenotypic characterization of Pseudomonas aeruginosa isolates from urinary tract infections. Int J Med Microbiol. 2011;301:282–92. doi: 10.1016/j.ijmm.2010.10.005. [DOI] [PubMed] [Google Scholar]
- 47.Bubonja-Sonje M, Matovina M, Skrobonja I, Bedenic B, Abram M. Mechanisms of carbapenem resistance in multidrug-resistant clinical isolates of Pseudomonas aeruginosa from a Croatian hospital. Microb Drug Resist. 2015;21:261–9. doi: 10.1089/mdr.2014.0172. [DOI] [PubMed] [Google Scholar]
- 48.Thrane SW, Taylor VL, Freschi L, Kukavica-Ibrulj I, Boyle B, Laroche J, Pirnay JP, Lévesque RC, Lam JS, Jelsbak L. The widespread multidrug-resistant serotype O12 Pseudomonas aeruginosa clone emerged through concomitant horizontal transfer of serotype antigen and antibiotic resistance gene clusters. mBio. 2015;6(5):e01396–15. doi: 10.1128/mBio.01396-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Finlayson EA, Brown PD. Comparison of antibiotic resistance and virulence factors in pigmented and non-pigmented Pseudomonas aeruginosa. West Indian Med J. 2011;60:24–32. PMID: 21809707. [PubMed] [Google Scholar]
- 50.Corehtash ZG, Ahmad Khorshidi FF, Akbari H, Aznaveh AM. Biofilm formation and virulence factors among Pseudomonas aeruginosa isolated from burn patients. Jundishapur J Microbiol. 2015;8(10):e22345. doi: 10.5812/jjm.22345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Nagaveni S, Rajeshwari H, Kumar Oli A, Patil SA, Kelmani Chandrakanth R. Widespread emergence of multidrug resistant Pseudomonas aeruginosa isolated from CSF samples. Indian J Microbiol. 2011;51:2–7. doi: 10.1007/s12088-011-0062-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wolska K, Kot B, Jakubczak A, Rymuza K. BOX-PCR is an adequate tool for typing of clinical Pseudomonas aeruginosa isolates. Folia Histochem Cytobiol. 2011;49:734–8. doi: 10.5603/FHC.2011.0098. [DOI] [PubMed] [Google Scholar]
- 53.Kareem AA, Salman Hassan S. Determination the genotyping diversity between biofilm forming and collagenase producing Pseudomonas aeruginosa strains. J Nat Sci Res. 2014;4:178–85. [Google Scholar]