

Abstract
Capsaicin, the pungent ingredient in chili peppers, produces intense burning pain in humans. Capsaicin selectively activates the transient receptor potential vanilloid 1 (TRPV1), which is enriched in nociceptive primary afferents, and underpins the mechanism for capsaicin-induced burning pain. Paradoxically, capsaicin has long been used as an analgesic. The development of topical patches and injectable formulations containing capsaicin has led to application in clinical settings to treat chronic pain conditions, such as neuropathic pain and the potential to treat osteoarthritis. More detailed determination of the neurobiological mechanisms of capsaicin-induced analgesia should provide the logical rationale for capsaicin therapy and help to overcome the treatment’s limitations, which include individual differences in treatment outcome and procedural discomfort. Low concentrations of capsaicin induce short-term defunctionalization of nociceptor terminals. This phenomenon is reversible within hours and, hence, likely does not account for the clinical benefit. By contrast, high concentrations of capsaicin lead to long-term defunctionalization mediated by the ablation of TRPV1-expressing afferent terminals, resulting in long-lasting analgesia persisting for several months. Recent studies have shown that capsaicin-induced Ca2+/calpain-mediated ablation of axonal terminals is necessary to produce long-lasting analgesia in a mouse model of neuropathic pain. In combination with calpain, axonal mitochondrial dysfunction and microtubule disorganization may also contribute to the longer-term effects of capsaicin. The analgesic effects subside over time in association with the regeneration of the ablated afferent terminals. Further determination of the neurobiological mechanisms of capsaicin-induced analgesia should lead to more efficacious non-opioidergic analgesic options with fewer adverse side effects.
Keywords: neuropathic pain, resiniferatoxin, capsaicin, osteoarthritis, TRPV1, analgesia, calpain, microtubule
1. Introduction
Chronic pain, defined as pain that lasts for longer than three months after the onset of the initial injury or disease, affects approximately 20% of Americans (Dahlhamer et al., 2018). The high prevalence of pain and pain-related diseases is a leading cause of disability and a major contributor to the global disease burden. Chronic pain is challenging to treat due to the limited efficacy and adverse side effects of first-line therapies. Although opioids are clinically useful when appropriately indicated, opioid use may lead to addiction. Moreover, opioid use carries major risks. Tachyphylaxis is common and paradoxical opioid-induced hyperalgesia may occur (Hoffman et al., 2017). In particular, the rising opioid overdose death rate has raised questions about the practice of prescribing opioids, and current guidelines discourage opioid use to treat chronic pain. Thus, the need for novel nonopioid, non-addictive treatments for the effective management of chronic pain is paramount.
Capsaicin (trans-8-methyl-N-vanillyl-6-nonenamide) is an active ingredient found in chili peppers and similar plants of the capsicum plant family. These plants contain between 0.1% and 2.0% capsaicin, and are common food additives. Topical application of capsaicin produces burning pain in humans, and has been used as a valuable tool for producing standardized experimental pain in humans and experimental animals. Acute and long-term pharmacological actions of systemically and locally administered capsaicin in multiple species have been thoroughly reviewed (Holzer, 1991; Szallasi and Blumberg, 1999). In particular, early studies using capsaicin established fundamental concepts in pain neurobiology, such as classification of subtypes of primary afferents, concepts of primary and secondary hyperalgesia, and central sensitization signaling in spinal cord (Treede et al., 1992; Sluka and Willis, 1997; O’Neill et al., 2012). Topical capsaicin has also been used as a reliable model for testing the efficacy of newly developed analgesic compounds (Wang et al., 2008; Wallace et al., 2016). Importantly, despite its strong algogenic effects, capsaicin has long been used as an analgesic agent—as early as 7000 BC (Lembeck, 1987; Watson, 1994). The first formal report of the pain-reducing properties of topical hot alcoholic pepper extract concerned toothache treatment (Turnbull, 1850; Szallasi and Blumberg, 1999). After decades of research and development, topical capsaicin has been identified as a valuable non-opioidergic drug for alleviating pain with few side effects, adding to its role in imbuing food with piquancy and in producing pain with pungency (Schumacher, 2010; Anand and Bley, 2011; O’Neill et al., 2012; Szallasi and Sheta, 2012; Szolcsanyi and Pinter, 2013; Chung and Campbell, 2016). In addition, the therapeutic application of topical capsaicin has been extended to numerous other conditions, such as itch in psoriasis, obesity, urological, airway, cardiovascular and gastrointestinal disorders, and cancer (Basith et al., 2016; Fattori et al., 2016).
Herein, we provide an overview of capsaicin’s major therapeutic applications to treat chronic pain. We discuss neurobiological and pharmacological mechanisms of capsaicin’s long-lasting analgesic effects on chronic pain.
2. Development of topical formula of capsaicin
Since capsaicin’s pharmacological adoption began in the mid-19th century, topical capsaicin has been available in several different concentrations and formulations including gels, creams, sprays, and patches (Hall et al., 2020). Formulas of topical capsaicin during early development contained low concentrations (<1%), and required frequent and continuous administration for prolonged periods of time, e.g., 3 or 4 applications daily for a minimum of six weeks (Watson et al., 1993) were necessary. Nevertheless, studies showed inconclusive efficacy for the use of low concentrations of capsaicin in treating chronic pain (Derry and Moore, 2012). The development of patches containing a high concentration of capsaicin (8%) (Qutenza developed by NeurogesX and currently supplied by Grünenthal) greatly enhanced the utility of capsaicin as an analgesic (Backonja et al., 2008). The therapeutic utility of Qutenza for chronic pain treatment has been extensively reviewed previously (Anand and Bley, 2011; Mou et al., 2013; Uceyler and Sommer, 2014; Bonezzi et al., 2020). Compared to a 0.4% capsaicin control patch, Qutenza produces long-lasting analgesia for self-reported pain in patients with postherpetic neuralgia (PHN). A single 60-min application produces analgesia lasting for 3 months, and the patient can be re-treated every 3 months. After topical application of Qutenza, systemic absorption of capsaicin is limited, and elimination is rapid with a half-life of 1.6 hours (Babbar et al., 2009). Absorbed capsaicin is metabolized by multiple cytochrome P450 enzymes in liver (Reilly et al., 2003). In the range of systemic concentrations achieved following topical application (<58 nM), capsaicin does not induce or inhibit cytochrome P450 enzymes in liver and is unlikely to affect metabolism of other drugs (Babbar et al., 2010). Thus, topical capsaicin has not shown drug interactions, and can be safely used in combination with other commonly used analgesics. Topical capsaicin shows only modest adverse side effects, the most common of which involve a small transient elevation of blood pressure, and local reactions, such as burning pain, erythema, and pruritus at the site of application (Simpson et al., 2010). Repeated treatments were well tolerated by patients during 52 weeks of observation (Galvez et al., 2017). Nonetheless, the repeated administration of capsaicin raised a concern, because metabolism of capsaicin in skin is slow (Chanda et al., 2008), and capsaicin has been suggested to have carcinogenic potential (Hwang et al., 2010; Bley et al., 2012). However, the carcinogenic potential of capsaicin appears to be related to impurities in capsaicin extracts, and evidence for carcinogenic effects of pure capsaicin is lacking. Moreover, capsaicin-induced anti-cancer effects are better established (Bley et al., 2012). Based on these therapeutic efficacy and safety data, Qutenza was approved by the US Food and Drug Administration in 2009 for the management of neuropathic pain associated with PHN, and was approved in 2020 for the management of neuropathic pain associated with diabetic peripheral neuropathy. In the European Union, Qutenza has been approved for the treatment of peripheral neuropathic pain in adults either alone or in combination with other medicinal products for pain.
Currently other formulations of capsaicin are also under development. CNTX-4975 by Centrexion is an injectable capsaicin for treating Morton’s neuroma and pain associated with osteoarthritis of the knee (Campbell et al., 2016; Stevens et al., 2019). A liquid form of high-concentration capsaicin (CGS-200 by Propella Therapeutics) is also under development for treating painful knee joint osteoarthritis.
3. Therapeutic efficacy of topical capsaicin for chronic pain
Currently approved therapeutic applications and peer-reviewed publications from clinical trials of topical capsaicin are summarized in Table 1.
Table 1.
Therapeutic applications and trials of topical capsaicin for treating pain
| Application | Administration | Efficacy | References |
|---|---|---|---|
| Post herpetic neuralgia* | Single 60 min 8% capsaicin patch or a 0.04% capsaicin control patch | Reduction of NPRS score by 29.9%-36.5% during 2w to 8w or 12w (20.4%-29.9% in control) | (Backonja et al., 2008) (Irving et al., 2011) (Webster et al., 2010a) |
| Reduction of NPRS score by 25.0% during 2w to 12w (14.7% in control). Mean pain reductions in 90-, 60-, or 30-min groups by 26.1%, 24.4%, and 24.4%, respectively. | (Webster et al., 2010b) | ||
| Single 8% capsaicin patch or oral pregabalin | Reduction of NPRS score by 55% in both capsaicin and pregabalin group. Shorter time-to-onset of pain relief by capsaicin (7.5d) than pregabalin (36d). Greater reduction of intensity and area of dynamic mechanical allodynia by capsaicin than by pregabalin in patients with peripheral neuropathic pain due to PHN, peripheral nerve injury or nondiabetic painful polyneuropathy. | (Haanpaa et al., 2016; Cruccu et al., 2018) | |
| 0.075% capsaicin cream for 6w | Reduction of VAS by 21% (6% in control). | (Watson et al., 1993) | |
| Diabetic peripheral neuropathy* | Single 30 min 8% capsaicin or placebo patch | Reduction of NPRS score by 27.4% during 2w to 8w (20.9% in control). | (Simpson et al., 2017) |
| Single 30 min or 60 min 8% capsaicin patch plus SOC, or SOC alone | Reduction of BPI by 67.3% in 30-min and 67.5% in 60-min group (40.6% in SOC alone) after 52 weeks. 28.6% of patients in 30-min group and 22.6% in 60-min group achieved a 30% response (14.3% in SOC alone). Interval for capsaicin retreatment was 68d. | (Vinik et al., 2016) | |
| Single 60-90 min 8% capsaicin patch | Reduction of NPRS score by 30.5% at 12w. 46.8% and 33.8% of patients show ≥30% and ≥50% reduction, respectively. | (Martini et al., 2012) | |
| HIV-associated neuropathy* | Single 30 min or 60 min 8% or 0.04% (control) capsaicin patch | Reduction of NPRS score from baseline to 2w to 12w in the 30 min group by 27% (16% in control). | (Brown et al., 2013) |
| Chemotherapy-induced neuropathy | 8% Capsaicin patch encouraged to wear for as long as possible | Reduction of NPRS by 84% and 97% in patients with high and low sensitivity to oxaliplatin, respectively, after 12 w. | (Filipczak-Bryniarska et al., 2017) |
| Single 30 min 8% capsaicin patch | Reduction of NPRS (−1.27), touch-evoked pain (−1.823) and cold-evoked pain (−1.456). | (Anand et al., 2019) | |
| 8% Capsaicin patch | Reduction of BPI by ≥30% in 37% of patients after 4w. | (Ramnarine et al., 2016) | |
| post-traumatic/post-surgical neuropathic pain | Single 8% capsaicin patch for 30 min to the feet and for 60 min to other areas of the body | Median reduction of NPRS by 36% in foot and by 41.2% in other areas after 1-2w. | (Tenreiro Pinto et al., 2018) |
| 8% Capsaicin patch (1-5 times) | 76.3% of patients had post-traumatic/post-surgical NP. ≥ 30% Reduction of NPRS in 22% of patients. | (Lanteri-Minet and Perrot, 2019) | |
| Phantom limb pain | Single 60 min 8% capsaicin patch | Reduction of NPRS scores for spontaneous stump pain by 1.01, evoked stump pain by 2.03, and phantom limb pain by 1.41 after 4w. Reduction in the areas of brush allodynia (by 80%) and pinprick hypersensitivity (by 72%) in the amputation stump. | (Privitera et al., 2017) |
| Knee joint osteoarthritis | 0.025% Capsaicin or vehicle cream four times per day for 12w | Reduction of VAS by ~50 % at 4w through 12w (~25% in control). | (Altman et al., 1994) |
| Single intraarticular injection of capsaicin (0.5 mg or 1.0 mg) or placebo. | Improvement of WOMAC and knee OA–associated pain through 12w in 0.5 mg group and 24w in 1.0 mg group. | (Stevens et al., 2019) | |
| Morton’s neuroma | Single injection of 0.1 mg capsaicin or placebo | Reduction in BPI at 1w and 4w | (Campbell et al., 2016) |
| Post-hemiotomy pain | Single intraoperative wound instillation of 1 mg capsaicin | Reduction in VAS during the first 3 days but not for the entire 1 or 4w after treatment. |
(Aasvang et al., 2008) |
| Oral neuropathic pain | 0.025% Capsaicin cream four times a day for 4w | Reduction in VAS in 63% of patients. | (Epstein and Marcoe, 1994) |
| Temporomandibular disorders | Single 8% Capsaicin or vehicle cream for 2 h on skin overlying affected TMJ and superficial masseter muscle | Reduction of VAS during 1w following treatment. Reduction in HPT 2h after capsaicin application for both the control and TMD groups, which was resolved within a week. No effect on PPT or mechanical sensitivity in both TMD and healthy individuals. | (Campbell et al., 2017) |
| Post-operative pain after tooth extraction | Single submucosal injection of 100 μg capsaicin or vehicle one day before surgery | Reduction in VAS at 60, 80, and 100 min after surgery. No difference between groups at 12, 24, and 48h post-operatively. | (Neubert et al., 2017) |
| Cluster headache | Total 50 μg capsaicin or vehicle liquid to both nostrils once daily for 7d | Reduction in the number of headaches from baseline to post-treatment during 1d through 7d | (Saper et al., 2002) |
BPI, brief pain inventory; d, day; HIV, human immunodeficiency virus; HPT, heat pain threshold; NPRS, numerical pain rating scale; PHN, post-herpetic neuralgia; PPT, pressure pain threshold; SOC, standard of care; TMD, temporomandibular disorders; TMJ, temporomandibular joint; w, week, WOMAC, Western Ontario and McMaster Universities Osteoarthritis Index; VAS, visual analog scale;
, clinical applications in which topical 8% capsaicin patch is approved to use in the US or EU.
3.1. Neuropathic pain in humans
Neuropathic pain is chronic pain that arises as a direct consequence of a lesion or disease affecting the somatosensory system. Approximately 7–10% of the population suffers from pain with neuropathic characteristics (van Hecke et al., 2014). Neuropathic pain may be associated with diabetic neuropathy, PHN, amputation (i.e., phantom limb pain), surgery or trauma, stroke or spinal cord injury, trigeminal neuralgia, HIV infection, chemotherapy-induced peripheral neuropathy, and other disorders. Current pharmacotherapy for neuropathic pain is primarily based on systemic medications (tricyclic antidepressants, serotonin-/norepinephrine-reuptake inhibitor antidepressants, pregabalin, gabapentin, etc.) (Finnerup et al., 2015; Bannister et al., 2020). However, systemic therapies are marred by potentially serious adverse effects (e.g., cognitive impairment and dizziness), and most patients have only modest pain relief (Bannister et al., 2020). Therefore, topical treatments (e.g., capsaicin and local anesthetics) have been used to provide relief for neuropathic pain due to abnormalities in the peripheral nervous system (Pickering et al., 2017). Capsaicin is recommended as a second-line drug for treating peripheral neuropathic pain by the Neuropathic Pain Special Interest Group of the International Association for the Study of Pain (Finnerup et al., 2015). The German Society for Neurology guidelines have recommended the capsaicin 8% dermal patch as a first-line option for localized neuropathic pain and as a second-line option for neuropathic pain of any cause (Binder and Baron, 2016).
To date, more than 200 randomized controlled trials (RCT) for the medical treatment of neuropathic pain have been published, and many systematic reviews of pharmacologic treatments for patients with neuropathic pain have been conducted to evaluate the efficacy of topical capsaicin in different modalities of neuropathic pain, such as post-herpetic neuralgia, post-traumatic or post-surgical nerve injury, and HIV-induced neuropathy (Mou et al., 2013; Finnerup et al., 2015; Derry et al., 2017; Galvez et al., 2017; van Nooten et al., 2017; Anand et al., 2019). A single application of capsaicin relieves PHN and HIV-associated neuropathic pain beginning from approximately three days following treatment, and lasting approximately five months (Brown et al., 2013; Mou et al., 2014). Aside from self-reported pain, topical capsaicin attenuates mechanical allodynia in patients with peripheral neuropathy (Moller et al., 2006; Kennedy et al., 2010; Campbell et al., 2016), and its efficacy has been shown to be superior even to pregabalin (Cruccu et al., 2018). The FDA has also approved the application of Qutenza in painful diabetic neuropathy. A network meta-analysis of 25 RCTs showed considerable efficacy of the capsaicin 8% patch compared to placebo and compared to centrally acting agents in patients with diabetic neuropathic pain (van Nooten et al., 2017). The capsaicin 8% patch provides efficacious pain relief similar to duloxetine, pregabalin, or gabapentin. Importantly, the oral medications were associated with significantly elevated risk of somnolence, dizziness, and discontinuation compared with placebo; none of these events was reported in association with the capsaicin 8% patch (van Nooten et al., 2017). In patients with chronic painful chemotherapy-induced peripheral neuropathy, the capsaicin 8% patch also reduced spontaneous pain, mechanical allodynia, and cold-evoked pain (Anand et al., 2019). Topical capsaicin also shows efficacy in patients with phantom limb pain by reducing spontaneous pain from stump and phantom pain, and decreasing the area of static and dynamic allodynia (Privitera et al., 2017). Trials are currently underway to determine the effects of topical capsaicin on neuropathic pain in patients with cancer, spinal cord injury, or breast surgery (clinicaltrials.gov; NCT03317613, NCT02869867, NCT03794388).
3.2. Knee osteoarthritis in humans
Osteoarthritis (OA) is one of the most prevalent and disabling chronic degenerative diseases; 250 million people worldwide have symptomatic knee OA, and this is predicted to increase sevenfold by 2030 (Dobson et al., 2018). Despite the condition’s clinical and financial burdens, no approved disease-modifying OA drugs are currently available. Pain is among the most prevalent symptoms of knee OA. However, it remains unclear what drives pain in knee OA, nor is it clear why some people develop pain while others do not. For treatment of OA pain, acetaminophen is used as first-line therapy in symptomatic OA, and second-line agents include topical agents (capsaicin and topical nonsteroidal anti-inflammatory drugs) (Nelson et al., 2014).
A systematic review cited efficacy of topical capsaicin cream (0.025% or 0.075%, four times daily) compared to placebo for pain from OA of multiple joints (Laslett and Jones, 2014). More recently, a liquid form of high-concentration topical capsaicin (5%; CGS-200-5 by Propella Therapeutics) is also under development for treating painful knee joint OA. In a phase 2 study (clinicaltrials.gov; NCT03528369), one hour application of CGS-200-5 on skin around knees on 4 consecutive days decreased the Western Ontario and McMaster Universities Osteoarthritis Index (WOMAC) OA total scores. A recent potential advance in OA pain management is the development of an intra-articular capsaicin formulation, thereby overcoming likely limited permeability of topical capsaicin into the knee joint (Stevens et al., 2019). Capsaicin injection into the knee joint was well tolerated and provided dose-dependent improvement in knee OA pain with walking. Onset of improvement occurred as early as one week after treatment, and capsaicin injection (1.0 mg) produced a significant decrease in OA knee pain through 24 weeks. A 0.5 mg capsaicin intraarticular dose also significantly improved pain, albeit to a lesser degree, at 12 weeks. Phase 3 trials are in progress.
3.3. Other chronic pain conditions
Some painful conditions are highly predictable, and pre-emptive treatment can be useful. Clinical trials have supported the utility of preoperative submucosal injection of capsaicin (0.1 mg) for attenuating acute post-operative pain following third molar extraction (Neubert et al., 2017). Perioperative administration of capsaicin decreases post-operative acute pain after herniotomy or total knee arthroplasty (Aasvang et al., 2008; Hartrick et al., 2011). Capsaicin also showed efficacy in idiopathic pain conditions, such as cluster headaches (Saper et al., 2002), temporomandibular disorders (Campbell et al., 2017), and burning mouth syndrome (Imamura et al., 2019). It is noteworthy that intrathecal delivery of resiniferatoxin (RTX), a highly potent vanilloid analog of capsaicin, is under development to treat intractable cancer pain in humans (Sapio et al., 2018) (clinicaltrials.gov; NCT00804154).
4. Ablation of TRPV1+ nerve terminals by capsaicin: a major contributor to analgesic treatment for chronic pain
4.1. Desensitization vs defunctionalization
Following more than 100 years of dormancy since the first formal report of the pain-reducing properties of topical hot alcoholic pepper extract for toothache (Turnbull, 1850), mechanistic studies using capsaicin were revitalized in the wake of the noted report of the activation of nociceptive sensory nerves by the application of capsaicin, and of reduced nerve responsiveness subsequent to topical application of capsaicin (Jancso N and Jancso A, 1949; Jancso G and Santha, 2015). Pioneering work by the Jancso group showed that capsaicin reducec the responsiveness of sensory nerve endings to chemical irritants in rodents; they found that repeated application of capsaicin into eyes, skin, or airways led to gradual insensitivity to subsequently applied capsaicin or other irritants, which lasted for days, a phenomenon they termed ‘desensitization’ (Jancso N and Jancso A, 1949; Jancso G and Santha, 2015). Subsequent studies in humans showed that topical application of capsaicin induced the loss of sensitivity not only to following applications of capsaicin, but also to various other chemical stimuli, such as histamine, bradykinin, and menthol (Wallengren and Hakanson, 1992; Green and Shaffer, 1993; Cliff and Green, 1996; Lo Vecchio et al., 2018). The quality and duration of sensory loss appears to depend primarily on the dose of capsaicin, which led to development of nomenclatures to indicate the effects of topical capsaicin on neural responses more accurately (Holzer, 1991). When the dose of topical capsaicin in initial administration is low, subsequent neural responses are reduced to the following administration of capsaicin, but not to other irritants, and is reversible within a couple of hours. This condition is referred to ‘specific desensitization’ (Holzer, 1991) or ‘tachyphylaxis’ (Szolcsanyi and Pinter, 2013). By contrast, when the dose of topical capsaicin during initial application is high, subsequent neural responses are reduced not only to capsaicin but also to other irritants or stimuli. This condition is referred to ‘non-specific desensitization’ or ‘defunctionalization’ (Holzer, 1991). The term ‘desensitization’ as initially used by Jancso and Jancso (Jancso N and Jancso A, 1949) encompasses both specific and nonspecific desensitization. In the literature, the term ‘desensitization’ has been used to indicate either the original meaning or only specific desensitization. Here, we will use the term ‘specific desensitization’ to indicate tachyphylaxis and ‘defunctionalization’ to indicate non-specific desensitization (Fig. 1A). We will further classify defunctionalization into two types based on capsaicin dose, duration, reversibility, and putative biological mechanisms. A relatively low dose range of capsaicin induces defunctionalization that is reversible within hours. This ‘short-term’ defunctionalization is likely due to functional impairment of primary afferents without structural changes (Fig. 1A). In contrast, high doses of topical capsaicin induce loss of sensation that is not readily reversible but lasts for months (Fig. 1A). This ‘long-term’ defunctionalization is likely mediated by structural ablation of afferent terminals. Of note, specific desensitization, reversible defunctionalization, and defunctionalization with ablation of afferent terminals are not always discretely segregated, but rather exist in a continuum with considerable overlap. For example, high dose capsaicin may cause initial functional impairment followed by structural ablation. Low dose capsaicin can cause both specific desensitization and short-term defunctionalization at the same time. Multiple repeated topical applications of low doses of capsaicin can result in an accumulation of capsaicin due to its slow metabolism in skin (Chanda et al., 2008), and thus might gradually produce short-term as well as long-term defunctionalization.
Figure 1. Potential contributors to capsaicin-induced analgesia.
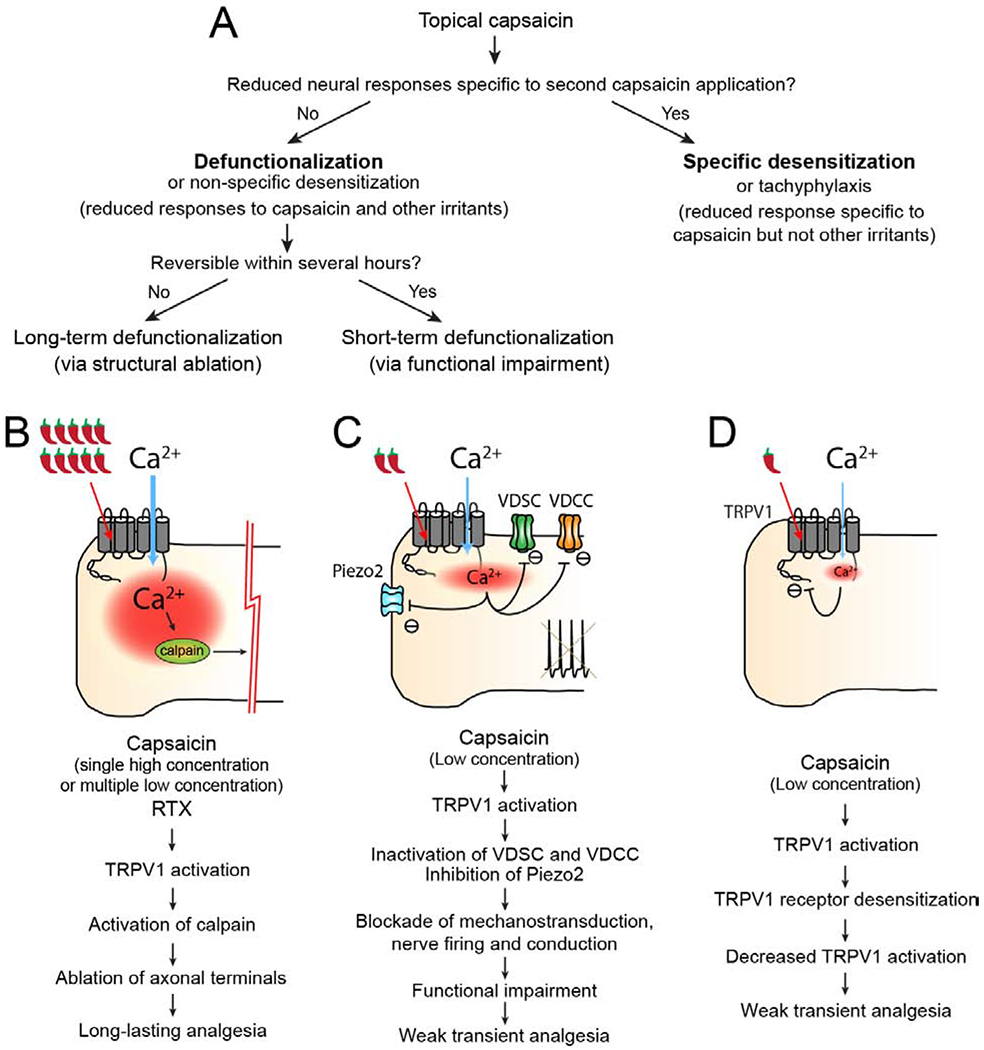
A. Diagrammatic depiction of neurobiological effects following topical administration of capsaicin. In this review, topical capsaicin-induced neural effects are classified based on the dose of capsaicin, analgesic quality, duration and reversibility, and mechanistically classified as long-term defunctionalization via structural ablation, short-term defunctionalization via functional impairment and specific desensitization. Each phenomenon has apparent mechanistic linkages with the processes shown in B, C, or D.
B. High concentrations of capsaicin or RTX activate TRPV1 and mediate influx of large amounts of Ca2+, leading to the activation of Ca2+-dependent protease calpain. Ablation of axonal terminals by calpain activation results in long-term defunctionalization of TRPV1+ afferents and mediates long-lasting analgesia for chronic pain.
C. Low concentrations of capsaicin activate TRPV1 to induce Ca2+ influx and subsequent inactivation of voltage-dependent sodium channels (VDSC), voltage-dependent calcium channels (VDCC), and Piezo2, a mechanosensitive ion channel. Inactivation of these channels results in blockade of transduction of sensory stimuli, initiation and conduction of action potentials, which produces short-term defunctionalization via functional impairment, and leads to transient analgesia.
D. Low concentrations of capsaicin produce TRPV1 activation and Ca2+ influx, which leads to receptor desensitization of TRPV1 ion channels. TRPV1 receptor desensitization decreases the response of TRPV1 to endogenous ligands or inflammatory mediators, and in turn reduces TRPV1-dependent hyperalgesia.
The relative contribution and evidence supporting the contribution to analgesia for chronic pain is strongest in A and weakest in C.
Understanding of neurobiological mechanisms underlying effects of capsaicin was greatly enhanced by the discovery of transient receptor potential vanilloid subfamily member 1 (TRPV1), the gene encoding the capsaicin receptor (Caterina et al., 1997). TRPV1 is a selective target of capsaicin and RTX. TRPV1 is a polymodal receptor that is critical to the sensing of various stimuli, such as noxious heat, protons, and endogenous lipid ligands (Schumacher, 2010; Chung et al., 2011). TRPV1 is a non-selective cationic ion channel, and its activation by agonists results in an influx of sodium and calcium through TRPV1, producing depolarization of neurons and action potential firing. The TRPV1 receptor is highly expressed primarily on C and some Aδ nociceptive sensory nerves, particularly those that detect painful or noxious sensations (nociceptors). Therefore, activation of TRPV1 results in the perception of pain, a process known as nociception. Indeed, knockout of TRPV1 in mice completely abolished pain behaviors in response to capsaicin or RTX (Caterina et al., 2000), and the discovery of TRPV1 has offered a mechanistic explanation as to how capsaicin produces burning pain. Further studies established that TRPV1 is a key molecule in peripheral sensitization associated with inflammation and injury, which rationalized the development of specific antagonists of TRPV1 as potential analgesics (Holzer, 2008; Schumacher, 2010; Chung et al., 2011; Szallasi and Sheta, 2012; Brederson et al., 2013). Studies have also revealed detailed mechanisms of neural responses following the topical administration of capsaicin (Holzer, 2008; Schumacher, 2010; O’Neill et al., 2012; Chung and Campbell, 2016). Specific desensitization following topical capsaicin is likely due to receptor desensitization of TRPV1 (Fig. 1D). Short-term defunctionalization via functional impairment is largely mediated by inactivation or inhibition of neuronal ion channels following activation of TRPV1 by capsaicin (Fig. 1C). Long-term defunctionalization via structural ablation is mediated by TRPV1 and Ca2+-dependent localized ablation of afferent terminals (Fig. 1B). In the following sections, we will discuss potential contributions of these TRPV1-dependent mechanisms to the analgesic effects of topical capsaicin.
4.2. Topical capsaicin mediates ablation of nociceptive nerve terminals localized at the site of capsaicin treatment
Capsaicin was found not only to activate but also to induce degeneration of a subset of primary afferent neurons (Jancso et al., 1977). An electron microscopic study showed that a single systemic injection of high-dose capsaicin (50 mg/kg) caused loss of a large proportion of primarily small-diameter neurons and axonal terminals in the spinal cord dorsal horn of neonatal rats. In adult rats, the systemic administration of capsaicin at a higher dose (100 mg/kg) also induced the degeneration of dorsal root ganglia (DRG) neurons and unmyelinated axons (Jancso et al., 1985). However, in adult rats, the extent to which degeneration of saphenous nerve fibers occurred was 45%, which is considerably less than the 70% observed in neonatal rats (Jancso et al., 1977; Jancso et al., 1985). RTX is more potent than capsaicin in producing excitotoxic effects; intraperitoneal injection of RTX (200 μg/kg) ablated TRPV1+ DRG neurons and central terminals in dorsal horn, and depleted unmyelinated fibers in the sciatic nerves by >90% (Pan et al., 2003). In addition to systemic injection, local delivery of capsaicin or RTX into the sensory ganglia can also produce ablation of primary afferents. For example, intraganglionic or peri-ganglionic injection of RTX can produce ablation of TRPV1+ afferents in different species (Karai et al., 2004; Tender et al., 2005; Brown et al., 2015; Unger et al., 2019; Wang et al., 2019). On the other hand, injection of relatively low doses of capsaicin or RTX close to the skin’s peripheral terminals produces reversible ablation restricted to peripheral terminals without the ablation of somata located in the sensory ganglia (Neubert et al., 2003; Yu and Premkumar, 2015; Wang et al., 2017b). Likewise, intrathecal injection of capsaicin also ablates the central terminals within the spinal cord without affecting TRPV1+ afferent soma in DRG (Cavanaugh et al., 2009). In humans, topical application or localized injection of capsaicin is unlikely to induce excitotoxicity in soma (i.e., capsaicin does not cause destruction of TRPV1+ afferent somata within sensory ganglia) because the capsaicin dose is not high enough to produce systemic effects. For example, the capsaicin 8% patch contains a total of 179 mg of capsaicin, which corresponds to a dose of approximately 3 mg/kg when the entire amount is absorbed. In a pharmacokinetic study, the maximum detectable plasma concentration in patients after a 60 min exposure to topical 8% capsaicin patch was 17.8 ng/ml (mean 1.9 ng/ml) (Babbar et al., 2009). Nonetheless, this is not comparable to the dose (>100 mg/kg) required to destroy TRPV1+ afferent somata within ganglia in adult rats. The amounts of capsaicin used for injection into a knee joint (1 mg per injection) or Morton’s neuroma (0.1 mg) are even lower (Campbell et al., 2016; Stevens et al., 2019). Therefore, in humans, it is reasonable to postulate that topical application or local injection of capsaicin exerts its effects by ablating only the peripheral terminals at the site of treatment without further proximal effects (Fig. 1A).
4.3. Analgesia for neuropathic pain by the ablation of epidermal nerve fibers: More complicated than it sounds like
Robust reduction of epidermal nerve fiber density (ENFD) after the application of high concentrations of capsaicin in human skin was shown by staining skin biopsies with a pan-axonal marker, PGP 9.5 (Simone et al., 1998; Polydefkis et al., 2004; Rage et al., 2010). In healthy volunteers, the 8% capsaicin patch produces an 80% reduction in ENFD after one week (Kennedy et al., 2010). Intraepidermal fibers were ablated as early as one day following intradermal injection of capsaicin in humans (Simone et al., 1998; Rage et al., 2010). Long-term effects of capsaicin on somatosensation are not straightforward, and its effects on thermal and mechanical sensitivity and pain are inconsistent in different studies (Table 2). Nonetheless, it is well accepted that a high concentration of capsaicin decreases heat sensation and heat pain for a prolonged period of time. Reduction of ENFD may underpin such reduced warmth detection, laser-evoked heat pain, and flare response (Landmann et al., 2016; Lo Vecchio et al., 2018). By contrast, capsaicin application did not affect tactile sensitivity or cold pain, and caused only a marginal decrease in mechanical pain (Lo Vecchio et al., 2018). The correlation between reduced ENFD and loss of heat pain appears to represent straightforward evidence for capsaicin-induced analgesia. However, capsaicin-induced reduction of ENFD does not necessarily guarantee analgesia for neuropathic pain. ENFD reduction is not necessarily associated with loss of sensation to mechanical stimuli. A noted paradox in the study of neuropathic pain relates to the association of a decrease in denervation with an increase in pain. In principle, reduced innervation should be associated with loss of function, i.e., loss of pain. However, multiple neuropathic pain conditions, such as diabetic neuropathy and chemotherapy-induced neuropathy, or idiopathic chronic pain conditions, such as fibromyalgia, are accompanied by reductions in ENFD (Caro and Winter, 2014; Landowski et al., 2016) (Fig. 2A, B). Under these conditions, ENFD reduction is often associated with increased rather than decreased pain. One can have loss of sensation in conjunction with mechanical hyperalgesia as demonstrated in psychophysical studies in human patients. For example, stroking pain (dynamic allodynia) can be prominent whereas punctate hyperalgesia (static hyperalgesia) is less obvious. The paradox of “hyperalgesia associated with denervation” can be explained by sensitization of uninjured remaining afferents (Campbell and Meyer, 2006). For example, in the spared nerve injury where one of the overlapping nerves that innervates the foot is left intact while the others are axotomized, striking hyperalgesia emerges, and mechanical allodynia is associated with sensitization of the uninjured nerves (Chen et al., 2018). One potential explanation of this paradox is that Wallerian degeneration of injured fibers leads to an increase in pronociceptive factors, such as nerve growth factor (NGF) and tumor necrosis factor, which in turn can cause sensitization of the remaining fibers (Wu et al., 2002; Schafers et al., 2003; Obata et al., 2004) (Fig. 2B).
Table 2.
Long-term effects of topical capsaicin on somatosensation in healthy volunteers
| Site | Administration | Affected sensation | References |
|---|---|---|---|
| Forearm | 0.075% Capsaicin or control cream four times daily for 6w | Increase in HPT and decrease in heat pain intensity over 6w. No effects on tactile threshold, pinprick pain, MDT, CDT, and histamine-induced itch. | (Simone and Ochoa, 1991) |
| Upper arm | Intradermal injection of capsaicin (0.2, 2, or 20 μg) | Reduction in heat pain and mechanical pain after 1w. No effects on cold sensation or tactile threshold. | (Simone et al., 1998) |
| Volar forearm | 0.075% Capsaicin four times daily for 3w | Reduction in pain intensity to heat, cold and mechanical stimulation. Increase in latency of heat pain detection by 5-fold and latency to cold by 3-fold. Increase in tactile threshold. Started ~2w of application, lasted ~3w after termination of application, and recovered at 6w after termination of application. | (Nolano et al., 1999) |
| Inner thigh | 8% Capsaicin (30, 60 and 120 min), 0.04% (30 min), or placebo | Increase in WDT in 8% for 60 or 120 min groups after 1w. No effects on CDT. | (Malmberg et al., 2004) |
| Anteromedial thighs | 8% Capsaicin for 1h | Reduction in sharp mechanical pain by 15% and increase in tactile threshold by 8% after 1w. No effects on CDT and HPT. All changes recovered at 12w. | (Kennedy et al., 2010) |
| Volar forearms | 8% Capsaicin or vehicle patches for 1 and 24h | 24h group: Increase in WDT and HPT. Marginal increase in MPT. Reduction in histamine-induced itch and neurogenic flare. No effects on MDT, CDT, CPT, VDT. Functional recovery after 21d. 1h group: modest increase in WDT and HPT after 1d. | (Lo Vecchio et al., 2018) |
CDT, cold detection threshold, CPT, cold pain threshold; HPT, heat pain threshold; MDT, mechanical detection threshold; VDT, vibration detection threshold; WDT, warmth detection threshold.
Figure 2. Hypothesis of capsaicin-induced analgesia for neuropathic pain.
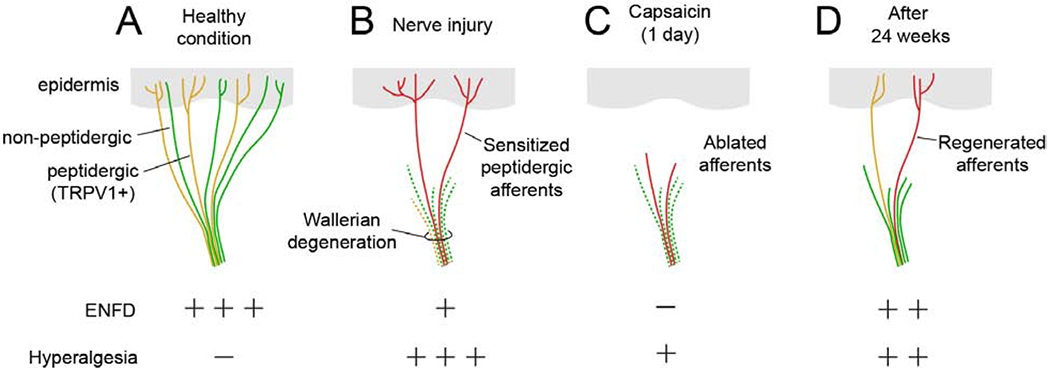
A. Epidermal nerve fibers are composed of non-peptidergic and peptidergic TRPV1+ nociceptors.
B. Partial nerve injury induces preferential degeneration of non-peptidergic afferents, which reduces epidermal nerve fiber density (ENFD). Wallerian degeneration and inflammation sensitize uninjured remaining peptidergic afferents.
C. The remaining subpopulation of afferents is sensitive to capsaicin, and high concentrations of topical capsaicin begin to ablate afferent terminals within 1 day to produce analgesia for neuropathic pain.
D. Capsaicin triggers regenerative processes in TRPV1+ afferents, and the ablated afferents are regenerated in 24 weeks (8 weeks in mice), at which point neuropathic pain returns. Retreatment with capsaicin induces analgesia in both humans and mice.
As severity of neuropathy increases or is characterized by more chronic progression, degeneration of TRPV1+ epidermal fibers increases, and topical capsaicin is unlikely to produce analgesia.
Investigations of cutaneous nerves have demonstrated preferential degeneration of fibers with different neurochemical identities (Bechakra et al., 2017): At 2 weeks following partial ligation of the sciatic nerve, the density of epidermal nerve fibers expressing PGP9.5, a panneuronal marker, or P2X3, a marker of non-peptidergic nociceptors, was decreased, whereas the density of calcitonin gene-related peptide (CGRP)-expressing peptidergic afferents in the footpad did not differ (Fig. 2A, B). In other studies, peptidergic afferents were reduced one to two weeks after the nerve injury, but demonstrated early regeneration (Grelik et al., 2005; Taylor et al., 2012). Several other nerve injury or nerve reconstruction studies have demonstrated similar preferential regeneration of peptidergic afferents compared to non-peptidergic afferents (Peleshok and Ribeiro-da-Silva, 2011; Duraku et al., 2013; Kambiz et al., 2015; Tsuboi et al., 2015). Consequently, more peptidergic afferents remain in skin than non-peptidergic afferents after nerve injury. In a recent study, patients with painful diabetic neuropathy showed increased density of dermal peptidergic fibers compared to patients without pain or healthy controls (Karlsson et al., 2020). Interestingly, in rats with peripheral neuropathy, mechanical hyperalgesia correlated with a reduction of P2X3-expressing epidermal nociceptors (Bechakra et al., 2017). Selective ablation of non-peptidergic afferents using isolectin B4 conjugated with saporin increased mechanical hyperalgesia in mice with CCI (Taylor et al., 2012). This is in contrast with the contribution of non-peptidergic afferents to mechanical hyperalgesia under inflammation (Pinto et al., 2019). A plausible explanation is that the greater degeneration of non-peptidergic epidermal nociceptors leads to greater sensitization of the remaining peptidergic afferents, and these remaining peptidergic fibers maintain mechanical hyperalgesia (Fig. 2B). Indeed, in a spinal nerve ligation model, mechanical hypersensitivity following injury of lumbar 5 afferents were maintained by lumbar 4 (uninjured) afferents, and capsaicin treatment on lumbar 4 afferents abolished mechanical hypersensitivity (Jang et al., 2007). Since peptidergic afferents are enriched with TRPV1, selective effects of capsaicin on the remaining and sensitized peptidergic fibers may explain the therapeutic benefits of capsaicin in neuropathic pain (Fig. 2C).
4.4. Capsaicin provides analgesia for neuropathic pain only in cases where hyperalgesia is maintained by TRPV1+ epidermal nerve fibers
Based on the aforementioned morphological and functional studies, we postulate that TRPV1+ peptidergic afferents likely persist after nerve injury, and that peptidergic nociceptive afferents likely become sensitized and contribute to the maintenance of neuropathic pain. Indeed, it has long been hypothesized that a subpopulation of patients with painful neuropathy have abnormal sensitization of unmyelinated nociceptors (“irritable nociceptors”) (Fields et al., 1998). In patients with painful neuropathy, polymodal and mechanically insensitive C-nociceptors are hyperexcitable, which correlates with mechanical hyperalgesia and spontaneous burning pain (Ochoa et al., 2005). Patients with painful neuropathy show greater capsaicin-induced vasoactive reactions than patients without pain or healthy volunteers (Forstenpointner et al., 2019). Since mechanically insensitive C-nociceptors mediate axon reflex flare, burning pain, and secondary hyperalgesia through central sensitization in humans (Schmelz et al., 2000b; Schmelz et al., 2000a; Sauerstein et al., 2018), these reports suggest the possibility that a subset of TRPV1+ afferents are hyperexcitable, and contribute to the maintenance of chronic pain in peripheral neuropathy. In preclinical studies, however, the contribution of TRPV1-expressing afferents to neuropathic pain, especially mechanical allodynia, is unclear. Multiple studies have shown that TRPV1 is involved in thermal hyperalgesia following nerve injury (King et al., 2011; Urano et al., 2012; Hirai et al., 2014; Abooj et al., 2016), which is consistent with the idea that TRPV1 and TRPV1+ nociceptors primarily mediate heat pain (Cavanaugh et al., 2009). By contrast, most reports have disputed their involvement in neuropathic mechanical allodynia—inhibition, knockout, or knockdown of TRPV1 does not affect mechanical hyperalgesia following neuropathy (Caterina et al., 2000; Urano et al., 2012; Hirai et al., 2014). Genetic ablation of TRPV1+ neurons or chemical ablation of TRPV1+ afferents neither prevented nor attenuated mechanical hyperalgesia following neuropathy (Ossipov et al., 1999; King et al., 2011; Mishra et al., 2011; Nakao et al., 2012; Zhang et al., 2014; Ma et al., 2015; Abooj et al., 2016). These data contrast sharply with the aforementioned clinical data, as well as with multiple preclinical studies implicating TRPV1 and TRPV1+ afferents in mechanical hyperalgesia following neuropathy (Pomonis et al., 2003; Rashid et al., 2003; Kanai et al., 2005; Culshaw et al., 2006; Christoph et al., 2007; Kissin et al., 2007; Kim et al., 2008; Watabiki et al., 2011; Labuz et al., 2016). The source of this discrepancy is unclear. Diverse pathophysiological mechanisms among the various peripheral neuropathy conditions may be a factor. Presumably, TRPV1 contributes to mechanical hyperalgesia in cases where TRPV1+ nociceptors are relatively intact and abnormally sensitized (Fig. 2B). In severe or advanced neuropathy conditions that accompany substantial loss of TRPV1+ afferents, TRPV1 may not contribute to the maintenance of neuropathic pain, and capsaicin therapy is likely to be less effective.
Recent studies have suggested that TRPV1 and TRPV1+ afferents are clearly involved in orofacial mechanical hyperalgesia following chronic constriction injury of the infraorbital nerve (ION-CCI) in mice (Kim et al., 2014; Wang et al., 2020). Systemic treatment with RTX prevented development of mechanical allodynia following ION-CCI. Importantly, focal injection of capsaicin into facial skin attenuates mechanical hyperalgesia lasting for >two weeks in this model (Wang et al., 2020). As well, capsaicin-induced analgesia for mechanical hyperalgesia is abolished by the inhibition of capsaicin-induced ablation of afferent terminals in skin (Wang et al., 2020). These data indicate that TRPV1+ nerve terminals remaining after ION-CCI maintain neuropathic pain, and that capsaicin-induced ablation of TRPV1+ afferent terminals is necessary to induce analgesia for mechanical hyperalgesia. This notion is further supported by the fact that chemogenetic inhibition of TRPV1-lineage afferents in skin attenuates mechanical hyperalgesia (Wang et al., 2020). Therefore, reduced ENFD following capsaicin treatment in this model produces analgesia for neuropathic pain (Fig. 2C). Notably, in ION-CCI, the TRPV1 channel itself also contributes to the maintenance of neuropathic pain in a site-specific manner (Fig. 3); inhibition of TRPV1 channels in the central terminals within the trigeminal subnucleus caudalis attenuates mechanical hyperalgesia, whereas inhibition of TRPV1 channel in the peripheral terminals does not (Kim et al., 2014; Wang et al., 2020). Therefore, we conclude that sensitization of TRPV1 in the central terminals mediates mechanical hyperalgesia in mice with ION-CCI, whereas TRPV1 expressed in the peripheral terminals enables these structures to respond to topical capsaicin and produces analgesia (Fig. 3).
Figure 3. The effects of manipulating TRPV1+ afferents in contrast to blockage of the TRPV1 channel in a trigeminal neuropathic pain model.
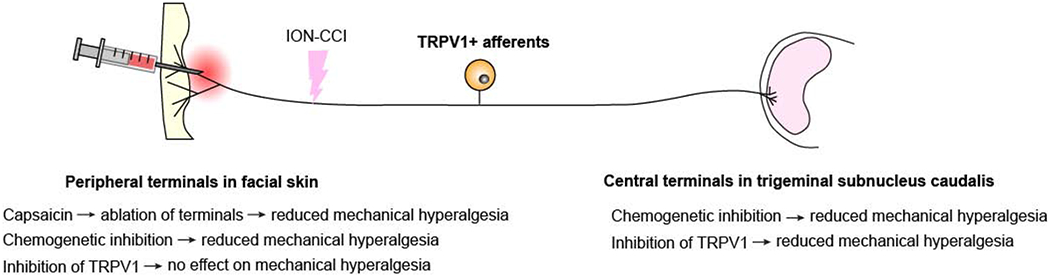
In mice with chronic constriction injury of infraorbital nerve (ION-CCI), chemogenetic inhibition of peripheral or central terminals of TRPV1+ afferents reduces mechanical hyperalgesia. Chemogenetic inhibition was performed by the administration of clozapine-N oxide to TRPV1Cre mice expressing inhibitory DREADD receptor. Pharmacological inhibition of TRPV1 channel function at central terminals within the trigeminal subnucleus caudalis attenuates mechanical hyperalgesia, whereas inhibition at peripheral terminals has no effect. Capsaicin administration at peripheral terminals activates TRPV1 and induces ablation of terminals of TRPV1+ afferents to produce analgesia for mechanical hyperalgesia. Summary of results from two publications (Kim et al., 2014; Wang et al., 2020).
TRPV1+ afferents also contribute to spontaneous ongoing pain in mice with peripheral inflammation (Wang et al., 2017a). Administration of capsaicin to facial skin attenuates ongoing non-evoked pain in mice with ION-CCI, and chemogenetic inhibition of TRPV1+ afferents in mice with ION-CCI likewise reduces ongoing pain (Wang et al., 2020). Nonetheless, the mechanisms whereby topical capsaicin reduces spontaneous pain are not clear. In humans, peripheral neuropathy patients show increased spontaneous firing of C-nociceptors (Ochoa et al., 2005; Serra et al., 2012). In rats, spontaneous activity from uninjured C fibers occurs as injured fibers degenerate (Wu et al., 2002). However, there is no evidence supporting the notion that topical capsaicin decreases such spontaneous firing of nociceptors. Even so, it is curious how capsaicin-induced ablation of afferent terminals can reduce spontaneous firing of nociceptors that is apparently initiated at the site of neuroma rather than nerve terminals. One possibility is that capsaicin-induced ablation of afferent terminals deprives nociceptors of the supply of factors contributing to the hyperactivity, such as NGF. NGF plays a profound role in modulation of nociceptor activity under conditions of tissue and nerve injury (Barker et al., 2020). Localized administration of neutralizing anti-NGF antibody into the hindpaw attenuates mechanical and thermal hyperalgesia in rats with neuropathic injury (Dai et al., 2020). It is possible that capsaicin-induced ablation of ENFD reduces the effects of NGF supplied from skin, which leads to the reduction in hyperactivity of nociceptors, as suggested previously (Anand and Bley, 2011).
4.5. Capsaicin-induced analgesia for other pathological pain conditions
The effect of capsaicin-induced ablation on pain associated with knee OA is less complicated considering the clear roles of intra-articular TRPV1+ afferents and TRPV1 in OA pain. Pharmacological or genetic inhibition of TRPV1 attenuates OA-induced hyperalgesia, such as weight-bearing imbalance, in rodent models (Barton et al., 2006; Honore et al., 2009; Helyes et al., 2010). TRPV1 also contributes to inflammation-induced edema of the knee joint (Szabo et al., 2005; Hoffmeister et al., 2011). Intra-articular injection of JNJ-17203212, a TRPV1 antagonist, reduced inflammation-induced sensitization of knee joint afferents to mechanical stimuli (Kelly et al., 2015). Hyaluronic acid inhibits TRPV1 in knee joint afferents, which likely mediates analgesia (Caires et al., 2015). Treatment with TRPV1 antagonist alleviates stair-climbing-induced pain in patients with knee OA (Manitpisitkul et al., 2018). Consistently, intra-articular injection of RTX or capsaicin improves weight-distribution behavior in knee arthritis models in rats, mice and dogs (Kissin et al., 2005a; Kalff et al., 2010; Abdullah et al., 2016; Kim et al., 2016). In humans, capsaicin-induced ablation of TRPV1+ afferent terminals within the knee joint can relieve hyperalgesia associated with OA, as was shown in a clinical trial (Stevens etal., 2019).
Recent results released from a clinical trial (clinicaltrials.gov; NCT03528369), yet unpublished, showed that repeated topical application of high concentrations of a liquid form of capsaicin on skin around the knee joint reduced pain from OA. This effect is surprising and puzzling, since it is not known if penetration and accumulation of capsaicin inside the knee joint is high enough to produce comparable effects on intra-articular TRPV1+ nerves as injectable capsaicin could do. If skin and joint afferents are innervated by the same primary afferents—which has not yet been demonstrated—ablation of cutaneous afferents might affect sensitivity of intra-articular afferents through unknown mechanisms (e.g., deprivation of NGF supply from skin). Although afferent inputs from knee joint and overlying skin converge in the spinal cord, it is unclear if transdermal capsaicin-induced ablation of cutaneous afferents reduces central sensitization in spinal cord.
Pain management in cases of burn injury is also challenging. RTX has shown efficacy in a rat model of full-thickness burn injury-induced hyperalgesia. RTX injection at the site of the burn injury reverses thermal and mechanical hyperalgesia throughout the recovery period, suggesting the role of TRPV1+ afferents in thermal and mechanical hyperalgesia in this model (Salas et al., 2017). In a partial-thickness burn injury rat hindpaw model, administration of TRPV1 antagonist to the hindpaw did not attenuate mechanical hyperalgesia, whereas intrathecal administration did, robustly (Green et al., 2016). Such site-specific effects of TRPV1 and the contribution of TRPV1+ afferents to mechanical hyperalgesia are reminiscent of the results observed in ION-CCI studies illustrated in Fig. 3.
The effects of capsaicin on preventing post-surgical hyperalgesia appear to be conflicting. Capsaicin instillation prior to incision in rat hindpaw prevents the development of spontaneous foot lifting, guarding behaviors, and thermal hyperalgesia, which correlated with the reduction of epidermal nerve fibers (Hamalainen et al., 2009; Uhelski et al., 2020). In these studies, capsaicin treatment did not affect the development of mechanical hyperalgesia. By contrast, the instillation of higher concentrations of capsaicin or injection of RTX prevented the development not only of thermal but also of mechanical hyperalgesia (Kissin et al., 2005b; Pospisilova and Palecek, 2006), suggesting a conflicting role for TRPV1+ afferents in mechanical hyperalgesia under these conditions.
4.6. Regeneration of ablated nerve terminals may determine duration of analgesia
Capsaicin-induced analgesia for neuropathic pain is not permanent, and the capsaicin 8% patch may require reapplication at 12-week intervals (Mou et al., 2013). The capsaicin 8% patch reduces ENFD by 80% at week 1, which is decreased to 20% at Week 12, and is almost fully recovered at week 24 (Kennedy et al., 2010). In mice, capsaicin-induced reduction of epidermal nerve fibers recovers by 8 weeks (Wang et al., 2017b). Capsaicin-induced analgesia for mechanical hyperalgesia in mice with ION-CCI disappears by approximately 7 weeks (Wang et al., 2020). Following complete recovery of mechanical hyperalgesia, a second capsaicin injection produces similar analgesia. Therefore, in both humans and rodents, the duration of capsaicin-induced analgesia may depend on the regeneration of ablated afferent terminals and reinnervation of the target area (Fig. 2D). A key question that has yet to be answered is what are the consequences of nerve regeneration following capsaicin-induced ablation under chronic pain conditions. Does nerve regeneration lead to recovery of physiological function (normal sensitivity) or pathological function (allodynia or spontaneous pain)? A partial answer is suggested by the finding that a patient with small fiber neuropathy treated with topical capsaicin showed reduced pain accompanied by normalization of ENFD (Trouvin and Perrot, 2019). A recent study in patients with painful chemotherapy-induced peripheral neuropathy also suggested that the regeneration of afferents following capsaicin treatment normalizes sensory functions (Anand et al., 2019). The reduction of ENFD in painful chemotherapy-induced peripheral neuropathy is partially restored 3 months after a single application with 8% capsaicin and is accompanied by reductions in spontaneous pain and mechanical allodynia (Anand et al., 2019). Therefore, it is reasonable to assume that some regenerated fibers are normal nerve fibers without pathological hypersensitivity (Fig. 2D).
Mechanisms of regeneration of nerve terminals ablated by capsaicin are not fully understood. Peptidergic afferent terminals denervated by capsaicin treatment undergo regeneration as evidenced by the expression of growth associated protein 43, which is facilitated by treatment with NGF (Schicho et al., 1999). Interestingly, recent studies suggest that capsaicin induces preferential regeneration of TRPV1+ afferents. Exposure of dissociated DRG neurons to capsaicin (10 μM) for more than 24 hours enhanced axonal outgrowth (Frey et al., 2018; Poitras et al., 2019). This process was Ca2+- and TRPV1-dependent, and involved the activation of protein kinase A and pro-regenerative transcriptional factor Cyclic adenosine monophosphate Response Element-Binding Protein (Frey et al., 2018). Therefore, topical capsaicin induces ablation of nociceptive terminals, and is likely to trigger regenerative machinery in TRPV1+ afferents simultaneously so that TRPV1+ afferents are preferentially regenerated compared to TRPV1-negative nociceptors. Determining more detailed neurobiological mechanisms for the regeneration of afferent terminals that are affected by both peripheral neuropathy and capsaicin should help to improve capsaicin therapy. For example, selective interference with the regeneration of the hypersensitive afferents may increase the duration of analgesia. Alternatively, selectively facilitating regeneration of normal afferents may attenuate hyperalgesia while enhancing the restoration of physiological sensation.
5. Other neurobiological contributors to capsaicin-induced analgesia
5.1. Capsaicin-induced functional impairment of TRPV1+ nerve terminals may induce analgesia
In humans, topical capsaicin initially increases thermal sensitivity but this is followed by a decrease in thermal sensitivity (Carpenter and Lynn, 1981; Simone and Ochoa, 1991). Intradermal injection of capsaicin induces hyposensitivity to pinprick stimuli as early as 15 min after injection and lasting for less than 24 hours (LaMotte et al., 1991). Electrically evoked potentials from human skin are reduced two hours after the application of topical capsaicin, but is recovered after 3 days (Papagianni et al., 2018). In rats, RTX injection to the hindpaws initially induces thermal hyperalgesia followed by thermal hypoalgesia approximately 2–3 hours after injection (Neubert et al., 2003; Neubert et al., 2008). Mechanical hyperalgesia induced by complete Freund’s adjuvant is modestly reduced after 2 hours following intraplantar injection of capsaicin (100 μg) and persists over 24 hours (Ma et al., 2015). Electrophysiological studies have shown that capsaicin inhibits axonal conduction (Such and Jancso, 1986) (Fig. 1B). Upon perineural application, capsaicin (100 μM) produces nerve discharge of Aδ and C fibers, followed by blockade of axonal impulse propagation. Since this inhibitory effect is reversible, and the capsaicin concentration is low, it is unlikely that the inhibitory effect is mediated by axonal ablation, but the finding suggests that capsaicin defunctionalizes axons through functional impairment without structural ablation. Capsaicin (1 μM) also inhibits action potential firing in dissociated sensory neurons from rodents (Liu et al., 2001; Ma et al., 2015). This inhibitory effect was abolished in neurons from TRPV1 knockout mice and was dependent on the influx of extracellular Ca2+ (Ma et al., 2015). In rat DRG neurons, capsaicin inhibits voltage-dependent Na+ currents (Su et al., 1999; Liu et al., 2001), and this effect depends on TRPV1 (Onizuka et al., 2011). Capsaicin also decreases high voltage-activated Ca2+ currents, which is prevented by a TRPV1 antagonist (Wu et al., 2005). Therefore, capsaicin-induced activation of TRPV1 leads to the inhibition of voltage-dependent Na+ and Ca2+ currents in the afferent terminals, which in turn inhibits action potential initiation and conduction in TRPV1+ afferents resulting in short-term defunctionalization (Fig. 1B). A recent study showed that capsaicin also inhibits Piezo2, a mechanosensitive ion channel, in sensory neurons through Ca2+-dependent depletion of membrane phospholipid (Borbiro et al., 2015). Since Piezo2 colocalizes with TRPV1 in DRG and contributes to mechanical pain in mice and humans (Murthy et al., 2018; Szczot et al., 2018), capsaicin-induced inhibition of Piezo2 can contribute to functional inhibition of nociceptors (Fig. 1B), which may underpin the mechanical insensitivity to pinprick (“analgesic bleb”) lasting for approximately 22h after capsaicin injection in humans (LaMotte et al., 1991).
The relative contribution of capsaicin-induced short-term defunctionalization to analgesia for chronic pain in humans may be called into question. Since the ablation of afferent terminals following the application of capsaicin in high doses occurs as early as 1 day after treatment in both humans and rodents (Simone et al., 1998; Neubert et al., 2003; Wang et al., 2017b), analgesia lasting longer than 1 day may be entirely attributable to the structural ablation (Fig. 1A, 2C). Under these conditions, capsaicin-induced functional impairments may contribute to transient analgesia immediately following capsaicin application before ablation of nerve terminals occurs. In a mouse model of neuropathic pain (Wang et al., 2020), capsaicin-induced attenuation of mechanical hyperalgesia was evident 1 day after capsaicin injection, but was not evident after 6 hours, suggesting that the acute functional effects of capsaicin on nerve excitability and conduction might not contribute to analgesia for neuropathic pain. However, in the case where capsaicin concentration is not high enough to produce ablation of afferent terminals, analgesia can be attributed to functional impairment of nociceptive afferents. A single 2-hour application of a patch containing 0.04% capsaicin does not reduce ENFD (Malmberg et al., 2004). Therefore, a single application of a low concentration of capsaicin (e.g., over-the-counter cream containing 0.025% capsaicin) might produce analgesia resulting from short-term defunctionalization via functional impairment. Since short-term defunctionalization following administration of a low concentration of capsaicin is transient and reversible (Such and Jancso, 1986), any analgesia achieved by a low concentration of capsaicin is also likely to be transient and reversible. Therefore, frequent applications over long periods would be necessary for effective analgesia. However, repeated application of 0.075% capsaicin reduces ENFD (Nolano et al., 1999), suggesting that repeated application of low concentrations of capsaicin will likely produce analgesia primarily through ablation of afferent terminals (Fig. 1C) rather than through functional impairment of the nociceptive nerve terminals. Thus, the ablation achieved by repeated low doses of capsaicin must be localized to cutaneous nerve terminals, as shown by the fact that 10 weeks of daily treatment with topical capsaicin (0.75%) in rat hindpaw does not affect neuropeptide content of nerves nor the numbers of DRG neurons (McMahon et al., 1991).
5.2. Desensitization of TRPV1+ afferents via receptor desensitization of TRPV1 likely produces transient analgesia
Capsaicin-induced analgesia is also attributable to a diminished TRPV1 channel function following capsaicin-induced activation. Upon activation with capsaicin, an influx of extracellular Ca2+ through TRPV1 triggers multiple Ca2+-dependent processes, including activation of calmodulin and calcineurin, leading to diminished responses of TRPV1 to prolonged or subsequent applications of capsaicin (Koplas et al., 1997; Numazaki et al., 2003; Vyklicky et al., 2008; Joseph et al., 2013). This diminished response is reversible (Vyklicky et al., 2008). This phenomenon is called “desensitization of TRPV1”. Receptor desensitization of TRPV1 likely contributes to reduced responses of TRPV1 located at nerve endings following multiple capsaicin applications (Fig. 1C). Since persistent TRPV1 activation leads to inflammatory hyperalgesia in multiple injury conditions (Honore et al., 2009; Helyes et al., 2010; Wang et al., 2018a), capsaicin-induced receptor desensitization of TRPV1 likely contributes to transient analgesia for inflammatory pain. In electrophysiological recordings in a rat ex vivo spinal cord-tail preparation (Dray et al., 1990), application of low concentration of capsaicin (<2 μM) diminished responses to subsequent capsaicin applications but not to bradykinin or heat. This desensitizing effect of capsaicin disappears within several hours, likely mediated by receptor desensitization of TRPV1. In contrast, a higher concentration of capsaicin (20 μM) impairs responses not only to capsaicin but also to other stimuli, suggesting the involvement of short-term or long-term defunctionalization. Again, however, it is difficult to dissect the relative contribution to capsaicin-induced analgesia in humans of desensitization of TRPV1+ afferents through receptor desensitization of TRPV1 or through short-term or long-term defunctionalization. In contrast, since RTX produces no TRPV1 receptor desensitization, RTX-induced analgesia is unlikely to be attributable to TRPV1 receptor desensitization (Caterina et al., 1997).
5.3. Release of anti-nociceptive neuropeptides may contribute to analgesia
Systemic capsaicin injection in adult rats reduces the amount of substance P by approximately 50% in spinal dorsal horn (Jessell et al., 1978). Intrathecal capsaicin also reduces substance P in rat spinal cord and reduces thermal sensitivity, as well (Yaksh et al., 1979). These reductions were not accompanied by reductions in morphine binding in spinal dorsal horn (Jessell et al., 1978), which suggested the depletion of the releasable pool of substance P without degeneration of primary afferents. Based on the proposal that substance P is an excitatory transmitter associated with nociception in the dorsal horn (Henry, 1976), capsaicin-induced depletion of substance P was suggested as a potential mechanism of capsaicin-induced desensitization effects. However, systemic and intrathecal capsaicin can cause degeneration of central terminals of primary afferents even without destroying soma (Chung et al., 1990; Cavanaugh et al., 2009), and it is unlikely that substance P is depleted by capsaicin without the occurrence of structural changes in the central terminals of primary afferents. Although preclinical studies show some evidence supporting the role of substance P in nociception, clinical trials of neurokinin 1 receptor in humans did not show analgesic effects (Hill, 2000). Therefore, capsaicin-induced depletion of substance P from TRPV1+ afferents does not sufficiently explain clinical analgesia. On the other hand, capsaicin treatment induces the release of somatostatin—an anti-nociceptive neuropeptide—from TRPV1+ neurons to produce systemic analgesia for inflammatory pain (Szolcsanyi et al., 1998; Helyes et al., 2004; Petho et al., 2017). In patients with lower-back pain, topical capsaicin decreases pain, and is accompanied by a threefold increase in plasma somatostatin (Horvath et al., 2014). Since a somatostatin receptor 4 agonist attenuates neuropathic pain (Kantas et al., 2019), capsaicin-induced analgesia for chronic neuropathic pain might also be attributable to somatostatin released from primary afferents, especially during the initial period immediately after administration of both low and high doses of capsaicin.
5.4. Conditioned pain modulation probably does not contribute to analgesia for chronic pain
Administration of capsaicin produces widespread activation of brain circuits. A study using functional magnetic resonance imaging comparing awake rats with or without TRPV1 expression showed that capsaicin administration activates circuitry involved in sensory and affective pain as well as pain modulation, including the anterior cingulate, insular, somatosensory cortex, amygdala, parabrachial nucleus, and periaqueductal gray (Yee et al., 2015). A key question is whether the capsaicin-induced activation of brain circuits produces analgesia in patients. For example, capsaicin can produce conditioned pain modulation: the application of capsaicin to cheek skin attenuates pain caused by electrical stimulation of tooth pulp in humans (Kemppainen et al., 1997). Such ‘pain inhibition by pain’ paradigm has been used in rodent models for studying mechanisms of descending noxious inhibitory control (DNIC). In this model, mechanical sensitivity measured from hindpaw is reduced by capsaicin injection into a forepaw as a conditioning stimulus (Da Silva et al., 2018; Phelps et al., 2019; Tobaldini et al., 2019; Yoneda et al., 2020). Capsaicin-induced DNIC involves capsaicin-induced altered functions of limbic and descending pain modulatory circuitry, such as the anterior cingulate cortex and prelimbic cortex, nucleus accumbens, amygdala, periaqueductal gray, and rostral ventromedial medulla (Da Silva et al., 2018; Phelps et al., 2019; Tobaldini et al., 2019). Capsaicin injection into the hindpaw reduces the jaw opening reflex, which is prevented by the injection of opioid antagonists or dopamine antagonists into the nucleus accumbens (Gear et al., 1999). Although it is reasonable to postulate that capsaicin-induced DNIC contributes to capsaicin-induced analgesia for pathological pain, the supporting evidence for this is meager. Since capsaicin-induced DNIC lasts for about one hour (Da Silva et al., 2018), it is possible that DNIC is involved in transient analgesia achieved with a low concentration of capsaicin rather than weeks of long-lasting analgesia achieved using a high concentration of capsaicin. Furthermore, in rats with neuropathic pain or knee OA pain, capsaicin-induced DNIC is impaired (Phelps et al., 2019; Yoneda et al., 2020), suggesting that capsaicin is unlikely to produce DNIC effects in patients with OA or neuropathic pain. However, analgesics such as duloxetine and tapentadol are known to reverse DNIC impairment in rats (Lockwood and Dickenson, 2019; Yoneda et al., 2020), suggesting that in patients taking these analgesics capsaicin treatment may enhance analgesia immediately after the application of capsaicin. Kappa opioid agonists or tyrosine receptor kinase B agonists (Kato et al., 2019; Phelps et al., 2019) also normalize capsaicin-induced DNIC, and may improve the analgesic effects of capsaicin. Since topical capsaicin acutely induces altered function of cortical and subcortical structures involved in pain perception and modulation (Da Silva et al., 2018; Phelps et al., 2019; Tobaldini et al., 2019), and placebo analgesia share common brain structures (Geuter et al., 2017), it is also noteworthy that analgesia achieved by topical capsaicin may include expectancy-induced analgesia. Further study on the central mechanisms of capsaicin-induced analgesia is warranted.
5.5. Capsaicin-induced procedural pain is not necessary for analgesia
Among the barriers to the wider application of capsaicin treatment is the intense burning pain that patients experience during the procedure. In patients with Morton’s neuroma or OA pain, the extent of procedural pain upon capsaicin injection is not associated with the extent of analgesia (Campbell et al., 2016; Stevens et al., 2019). Therefore, capsaicin-induced procedural pain is not likely associated with its therapeutic effects, and a reduction in capsaicin-induced discomfort may increase patient tolerance during the procedure. However, except for nerve block anesthesia, which is not applicable in most cases, no effective means of preventing capsaicin-induced procedural pain is currently known. Topical lidocaine administered prior to the application of the capsaicin 8% patch does not greatly attenuate burning pain (Knolle et al., 2013; Kern et al., 2014). One possibility is exposure of the skin at the site of capsaicin application to cold, which has been found to attenuate burning pain (Knolle et al., 2013). An ongoing trial is testing the effects of refrigerated cushions on capsaicin-induced procedural pain in patients with neuropathic pain (clinicaltrials.gov; NCT02869867). Cold reduces action potential conduction from nerve endings by inactivating voltage-gated sodium channels (Zimmermann et al., 2007). One potential concern in cold application is whether decreased temperature at the site of capsaicin application might affect the therapeutic efficacy of capsaicin due to the inhibitory effects of cold temperature on capsaicin-induced TRPV1 activation (Chung and Wang, 2011). However, cold temperature (20°C) does not affect the reduction of ENFD by topical capsaicin in humans (Knolle et al., 2013), suggesting that pre-emptive cooling of the site for local application of capsaicin may offer a means of attenuating procedural pain in patients. Further studies of the mechanisms of capsaicin-induced pain and capsaicin-induced ablation may lead to the development of new methods to selectively attenuate pain without compromising the efficacy of capsaicin-induced ablation.
It is possible that capsaicin-induced burning pain and analgesia are driven by a specific subset of TRPV1+ afferents. Capsaicin-induced burning pain in humans correlates with sustained discharge of mechano-insensitive, heat-insensitive C fibers rather than polymodal nociceptors (Schmelz et al., 2000a). If the subset of mechano-insensitive C fibers can be selectively silenced, it may eradicate procedural pain. Recent single-cell transcriptomic studies subclassified TRPV1+ primary afferents from DRG into different classes (Usoskin et al., 2015; Li et al., 2016). In one study, TRPV1+ afferents were clustered into seven groups, most of which were mechanoheat-sensitive polymodal nociceptors (Li et al., 2016). In deep somatic afferents, acetylcholine receptor subunit alpha-3 (Charna3) labels mechano-insensitive nociceptors (Prato et al., 2017), which may account for procedural pain upon injection of capsaicin into the knee joint. Therefore, selective silencing of Charna3+ nerves in knee joint may attenuate procedural pain upon injection of capsaicin. However, Charna3+ mechano-insensitive afferents are not found in skin (Prato et al., 2017), and the molecular signature of the mechano-insensitive heat-insensitive C fibers in skin remains unknown.
Non-pungent capsaicin analogs are under development with the aim of reducing procedural pain. For example, olvanil, arvanil, or palvanil activates TRPV1 with slower kinetics than capsaicin, and produces only modest eye-wiping responses upon corneal application in rodents (Ursu et al., 2010; De Petrocellis et al., 2011). Olvanil reduces capsaicin-induced thermal hyperalgesia in vivo, and induces receptor desensitization in dissociated sensory neurons in vitro (Alsalem et al., 2016). Palvanil pretreatment attenuates capsaicin-induced eye wiping and this effect lasts less than 6 hours (De Petrocellis et al., 2011). These reports suggest that analgesic mechanisms operate through receptor desensitization of TRPV1. Olvanil may also induce short-term defunctionalization, since olvanil leads to inhibition of VDCC through a TRPV1/Ca2+/calmodulin-dependent process in dissociated sensory neurons (Wu et al., 2006). However, it is unclear if the non-pungent capsaicin-like compounds readily produce long-term defunctionalization. Systemic daily treatment with palvanil is necessary to attenuate thermal and mechanical hyperalgesia following spared nerve injury (Luongo et al., 2012). NE-21610, a capsaicin analogue, produces brief moderately intense pain in humans, which is followed by profound heat hypoalgesia for a week (Davis et al., 1995a). Pretreatment with NE-21610 prevents the development of heat and mechanical hyperalgesia from burn injury in humans (Davis et al., 1995b). With further development, non-pungent vanilloid analogues that produce efficient ‘non-painful ablation’ should be assessed for their potential therapeutic utility.
5.6. Capsaicin is not for everyone: Individual differences of capsaicin-induced analgesia
Individual differences characterize the experience of pain and have been a topic of keen interest to pain researchers for many years (Fillingim, 2017). The conditions and the pathophysiological states that determine the onset of neuropathic pain are diverse, and clinical neuropathic pain conditions are heterogenous with individual differences. In addition, the complexity and multiplicity of underlying pathophysiological processes occurring in individual patients provoke different responses to a single treatment. Individual patients also experience differences in the therapeutic efficacy of capsaicin, and not all patients benefit from capsaicin treatment (Martini et al., 2012; Martini et al., 2013; Treede et al., 2013). Approximately 44% of patients with PHN have been shown to experience pain relief of 30% or more, and 10% of patients exhibit complete pain relief in response to capsaicin treatment (Mou et al., 2014). Therefore, it is important to identify reliable predictors of patient responses to capsaicin treatment.
Baseline pain prior to treatment can be a predictor. Variability of pain prior to treatment, baseline pain scores, and time with preexisting pain are associated with analgesic effects (Martini et al., 2013; Hoper et al., 2014; Maihofner and Heskamp, 2014; Katz et al., 2015; Serrano et al., 2017) (Table 3). In a meta-analysis of 1,248 patients across four RCTs, responses to the capsaicin 8% patch were better in a group characterized by younger female patients with a shorter duration of pain prior to the treatment, low baseline pain, and variability in baseline pain score (Martini et al., 2013). Other studies also reported better responses to topical capsaicin in patients with shorter duration of neuropathic pain and lower baseline pain intensity (Hoper et al., 2014; Katz et al., 2014; Maihofner and Heskamp, 2014). Importantly, a favorable response to topical capsaicin was predicted in patients showing analgesia by topical local anesthetics (Martini et al., 2013). These results suggest that capsaicin likely produces greater analgesia if patients do not have a rigid and fully manifested chronic pain process associated with a severe central plastic change.
Table 3.
Potential predictors of positive responders to topical capsaicin among patients with neuropathic pain
| Potential Predictors | References | |
|---|---|---|
| Sex and age | Female>male; younger>older | (Martini et al., 2013) |
| History of pain before treatment | Shorter pain duration | (Martini et al., 2013; Hoper et al., 2014; Maihofner and Heskamp, 2014) |
| Greater variability of pain intensity before treatment | (Martini et al., 2013) | |
| Lower baseline intensity of pain | (Martini et al., 2013; Katz et al., 2014) | |
| No concomitant opioid use | (Martini et al., 2013) | |
| Pain symptoms | Presence of cold and pinprick hyperalgesia | (Mainka et al., 2016) |
| Absence of allodynia | (Katz et al., 2014) | |
| Nonhomogeneous heat pain and warmth detection | (Serrano et al., 2017) | |
| Relief by topical lidocaine | (Martini et al., 2013) | |
Other studies suggest that sensory phenotypes defined through standardized testing can also be used as a surrogate marker of underlying mechanisms of pain generation and as a predictor of responses to capsaicin treatment. Patients with neuropathic pain syndromes show a wide spectrum of sensory symptoms that depend upon multiple yet entirely unidentified factors involving diverse underlying central and peripheral mechanisms (Maier et al., 2010; Baron et al., 2017). An unbiased clustering of ~900 peripheral neuropathy patients with diverse etiologies based on quantitative sensory profiles showed that approximately one third of patients clustered to a group showing significant sensory gain (without sensory loss) (Baron et al., 2017). This group of patients showed heat, cold, and mechanical hyperalgesia, which may have been mediated by sensitized nociceptors (Fields et al., 1998). Indeed, responders to capsaicin treatment demonstrated more intensive cold and mechanical pinprick hyperalgesia than non-responders, though warmth detection and heat pain were the same (Mainka et al., 2016). Consistently, absence of allodynia is associated with responders (Katz et al., 2015). High sensory neuropathic abnormality scores as well as the presence of burning and pressure-evoked pain were weakly but not significantly associated with treatment response (Hoper et al., 2014). A recent study suggested that thermal sensitivity may also be a predictor of response to capsaicin (Serrano et al., 2017). Patients with significant differences in only one of either the warm sensation threshold or the heat pain threshold between the neuropathic pain area and the contralateral side were more likely to respond than patients with significant differences in both the warm sensation threshold or the heat pain threshold, regardless of increase or decrease. These reports suggest that patients with localized neuropathic pain maintained by sensitized peripheral nociceptors with partial damage of peripheral nerves will respond better to capsaicin than patients with widespread centralized pain and severe nerve damage. This may be due to the dynamic change of TRPV1+ afferents during the progression of neuropathy. For example, progression of diabetic neuropathy decreases TRPV1+ ENFD (Narayanaswamy et al., 2012). Results of autopsy of patients with painful neuropathy shows reduced TRPV1+ ENFD compared to control skin (Lauria et al., 2006). In advanced lesions, TRPV1+ afferents are denervated and topical capsaicin cannot reach the target site. Therefore, density of TRPV1+ ENFD is suggested to be a predictor of capsaicin efficacy (Bonezzi et al., 2020), an intriguing possibility that needs to be validated.
Variation of capsaicin-induced analgesia is also evident in preclinical models, and the effects of capsaicin or RTX treatment on neuropathic pain are variable (Ossipov et al., 1999; Rashid et al., 2003; Kissin et al., 2007; Kim et al., 2008; King et al., 2011; Mishra et al., 2011; Nakao et al., 2012; Zhang et al., 2014; Ma et al., 2015; Abooj et al., 2016). Multiple factors may explain this variation. As in humans, it is possible that TRPV1+ afferents contribute to mechanical hyperalgesia to different extents in different peripheral neuropathy models. Presumably, TRPV1 contributes to mechanical hyperalgesia in cases where nociceptors are relatively intact and abnormally sensitized. For example, the ION-CCI model involves use of chromic gut for loose ligation of ION (Wang et al., 2020), which induces strong inflammatory responses (Ma et al., 2012). Since TRPV1-dependent thermal hyperalgesia is evident (Urano et al., 2012), this model likely involves a greater role for TRPV1+ nociceptors and, hence, exhibits a greater degree of capsaicin-induced analgesia. Determining the pathophysiological mechanisms underlying differential analgesia by capsaicin in various models may yield important clues with respect to individual variation among patients.
It is also possible that genetic and epigenetic variations may contribute to individual differences. TRPV1 variants are known to affect activation and receptor desensitization of TRPV1 in vitro, and single amino acid variations impact capsaicin-induced responses in rodents (Wang et al., 2016; Joseph et al., 2019). In humans, single nucleotide polymorphism of TRPV1 affects pain sensitivity (Valdes et al., 2011; Forstenpointner et al., 2017; Okamoto et al., 2018). However, it is not known whether TRPV1 variants affect capsaicin-induced analgesic effects. Furthermore, TRPV1 expression is distinct depending upon the sex and strain of mice (Ono et al., 2015; Payrits et al., 2017). The expression of TRPV1 is also under the control of epigenetic regulation (Crow et al., 2015), and capsaicin causes the epigenetic regulation of other genes (Wang et al., 2018b). Aside from TRPV1, capsaicin-induced analgesia is likely affected by variations of a number of other genes involved in neuronal excitability and ablation of axonal terminals. The aggregate effects of these genetic and epigenetic factors may contribute to individual differences in capsaicin-induced pain and analgesia.
5.7. Non-neuronal TRPV1-expressing cells may not affect therapeutic efficacy and toxicity
TRPV1 is functionally expressed in non-neuronal cells in humans, such as keratinocytes (Denda et al., 2001) and synoviocytes (Kochukov et al., 2006). Keratinocytes are in proximity to nociceptors, and a recent study indicates that TRPV1 in keratinocytes can transduce nociceptive information (Pang et al., 2015). Although mouse keratinocytes do not express TRPV1 endogenously, transgenic overexpression of TRPV1 using a keratin promoter in TRPV1 KO mice showed that activation of TRPV1 limited to keratinocytes induces c-fos expression in the ipsilateral spinal cord, and is associated with acute paw-licking nocifensive behaviors (Pang et al., 2015). Since topical application of capsaicin simultaneously affects TRPV1 in nerve terminals and in keratinocytes, we may ask whether TRPV1 in keratinocytes contributes to analgesic effects or side effects following topical application of capsaicin. However, the supporting evidence for this is meager. It is not known whether TRPV1 in keratinocytes contributes to the maintenance of chronic pain. Clinical studies report no side effects of topical capsaicin with respect to skin integrity aside from localized skin reactions, such as erythema, pruritus, and pain (Derry et al., 2013). Pre-emptive injection of capsaicin before skin incision does not interfere with wound healing (Uhelski et al., 2020). Primary keratinocytes from human skin biopsies exhibit lower levels of TRPV1 mRNA compared to sensory ganglia (Facer et al., 2007). Moreover, neither capsaicin nor RTX induces cytotoxicity in the HaCaT human keratinocyte cell line or in primary keratinocytes from skin biopsies (Pecze et al., 2008). Interestingly, human keratinocytes contain a splice variant form of TRPV1—TRPV1b (Pecze et al., 2008)—which functions as a dominant negative of TRPV1 (Vos et al., 2006). Therefore, it is unlikely that topical application of capsaicin produces analgesia or adverse side effects through the activation or receptor desensitization of TRPV1 in keratinocytes or through cell death of keratinocytes.
TRPV1 is expressed in human synovium, and is upregulated in synovial samples taken from patients with OA (Kochukov et al., 2006; Engler et al., 2007; Kelly et al., 2015). Activation of TRPV1 in synoviocytes leads to the generation of reactive oxygen species and proinflammatory cytokines from synoviocytes (Engler et al., 2007; Hu et al., 2008; Westlund et al., 2010; Terenzi et al., 2013). These reports suggest that intra-articular injection of capsaicin may either aggravate or alleviate osteoarthritic conditions. However, it is not known whether TRPV1 in synovium contributes to capsaicin-induced analgesia or synovial changes following local administration of capsaicin. A single intra-articular injection of RTX in osteoarthritic dogs improves gait and weight bearing for 4 months or longer, with no detectable signs of accelerated osteoarthritic progression (Iadarola et al., 2018). Likewise, a single intra-articular injection of capsaicin does not impair healing of tendon or joint articular cartilage in rabbits (Friel et al., 2015). Therefore, intra-articular capsaicin appears to be a safe and effective treatment for OA pain, despite its effects on the function or structure of synoviocytes.
6. Mechanisms of vanilloid-induced ablation of nociceptive nerve terminals
As discussed above, long-lasting analgesia achieved with high doses of capsaicin primarily depends on axonal ablation without the degeneration of soma in the sensory ganglia. Most studies regarding capsaicin-induced toxicity have focused on mechanisms of cell death (Holzer, 1991; Zhai et al., 2020), and the mechanisms of ablation of nociceptive terminals remain poorly understood. In this section, we will discuss potential mechanisms of capsaicin-induced ablation of nociceptor terminals.
6.1. Cytosolic increase in Ca2+
Axotomy-induced degeneration of axons from sensory nerves in explant cultures of DRG depends on extracellular Ca2+ (George et al., 1995). To determine the contribution of Ca2+-dependent mechanisms underlying capsaicin-induced ablation of axonal terminals, a novel in vitro preparation was developed to quantify capsaicin-induced effects in real-time (Wang et al., 2017b). Using dissociated DRG neurons from TRPV1-lineage reporter mice in a multicompartmental microfluidic chamber, the effects of capsaicin application restricted to the axons (avoiding exposure to soma) were monitored (Fig. 4A). In this assay, capsaicin induced beading and ablation of GFP labeled axonal terminals as early as 1 min (Fig. 4B–D). This approach excludes indirect axonal ablation as a consequence of cell death. Ca2+ influx through TRPV1 is necessary for capsaicin-induced beading and ablation of axonal terminals (Fig 3E), shown by the fact that chelation of extracellular Ca2+ or TRPV1 knockout prevents capsaicin-induced ablation. In addition to Ca2+-influx through the TRPV1 pore, other sources of Ca2+ increase can contribute to capsaicin-induced axonal ablation. Interestingly, however, pharmacological inhibition of the voltage-gated Ca2+ channels and depletion of Ca2+ stores does not affect capsaicin-induced ablation (Wang et al., 2017b) (Fig. 4F), and neither does inhibition of Ca2+-activated Cl− channels. Therefore, Ca2+ influx through the TRPV1 pore is the dominant source of Ca2+ in the production of capsaicin-induced axonal ablation (Wang et al., 2017b).
Figure 4. Ca2+-dependent calpain-mediated capsaicin-induced ablation.
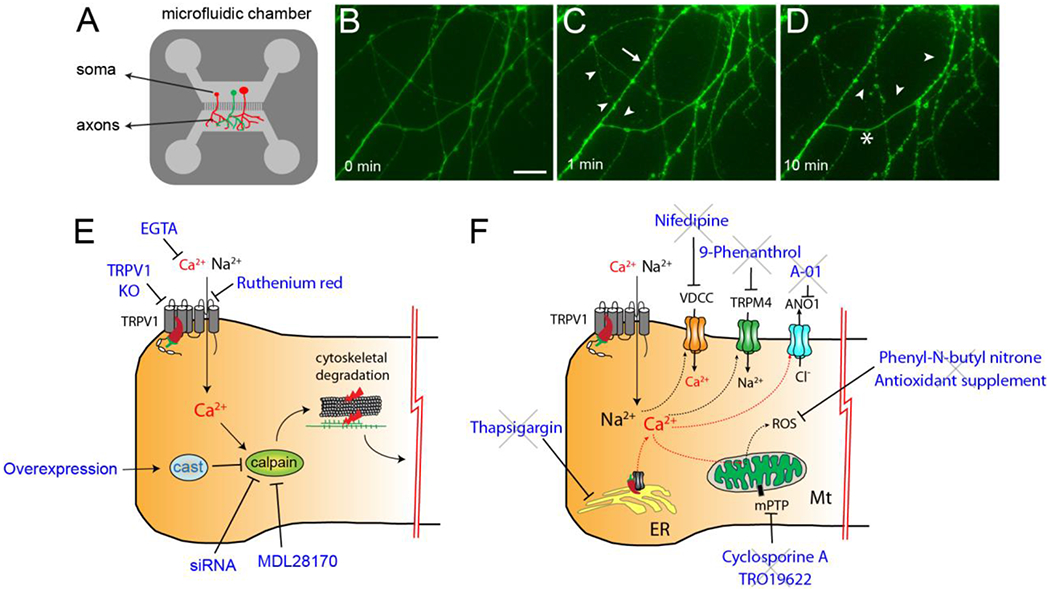
A. A schematic of multicompartmental culture of DRG neurons in a microfluidic chamber (MFC). Primary afferent neurons were dissociated from TRPV1-GFP reporter mice and plated in one side of the MFC. Axonal terminals cross the barrier and reach the compartment on the other side (axonal compartment). Capsaicin application was restricted to the axonal compartment, where time-lapse imaging was performed.
B-D. Representative time-lapse imaging of axonal terminals from MFC. Fluorescence signals from GFP were monitored before and after the axonal compartment was exposed to capsaicin (100 μM). Scale bar in B, 10 μm; arrow heads, ablated fibers; arrows, beaded fibers; asterisk, fibers not affected.
E. TRPV1-mediated Ca2+ influx through TRPV1 pore is necessary for capsaicin-induced ablation of axonal terminals. Chelation of extracellular Ca2+ (EGTA), genetic knockout, or pore blocking (by ruthenium red) of TRPV1 prevents capsaicin-induced ablation of TRPV1-lineage axons. In addition, capsaicin-induced ablation is decreased by pharmacological inhibition of calpain, a Ca2+-dependent protease, by MDL28170, by siRNA knockdown of calpain, or by overexpression of calpastatin (cast), an endogenous inhibitor of calpain. Activation of calpain likely induces subsequent cytoskeletal degradation, which leads to ablation of axonal terminals.
F. Factors/mechanisms that do not contribute to capsaicin-induced ablation: voltage-dependent Ca2+ channels (VDCC, inhibited by nifedipine); transient receptor potential subfamily M member 4 (TRPM4, a Ca2+-activated non-selective cationic channel, inhibited by 9-Phenanthrol); anoctamin 1 (ANO1, a Ca2+-activated Cl− channel inhibited by A-01); mitochondrial permeabilization transition pore (mPTP, inhibited by cyclosporine A or TRO19622); Ca2+ from endoplasmic reticulum (ER, depleted by thapsigargin); or reactive oxygen species (ROS, scavenged by PBN or antioxidant supplement). A-D, reproduced from Wang et al., 2017 (Wang et al., 2017b). E and F, summary of results from Wang et al., 2017 (Wang et al., 2017b).
6.2. Mitochondrial dysfunction
Mitochondrial effects likely contribute to capsaicin-induced ablation of nociceptor terminals. Injection of capsaicin into skin also increases mobility of mitochondria toward peripheral terminals (Sajic et al., 2013). Consequently, mitochondrial redistribution may result in the accumulation of mitochondria in axonal terminals in the skin to better supply energy and to maintain Ca2+ homeostasis at the site of capsaicin injection. However, mitochondrial dysfunction at the capsaicin injection site contributes to axonal degeneration (Chiang et al., 2015). In adult mice, repeated intradermal injection of 1% capsaicin into the hindpaw induces axonal swelling and reduction in density after 7 days, suggesting ablation of afferent terminals. Capsaicin-treated unmyelinated axons contained swollen and shortened mitochondria as well as decreased neurofilament densities. In dissociated DRG neurons, capsaicin was observed to induce axonal swelling, which was accompanied by a reduction in the length of mitochondria and motility within axons. These changes in mitochondria were attenuated by Ca2+ chelators or capsazepine. Transfection with a dominant negative mutant Drp1—a mitochondrial protein responsible for mitochondrial fission—attenuated capsaicin-induced decreases in mitochondrial length as well as axonal swelling and degeneration in vitro (Chiang et al., 2015). This study suggests that capsaicin-induced fission and loss of function of mitochondria are associated with axonal swelling and degeneration following application of capsaicin. It is possible that capsaicin-induced fission of mitochondria reduces the functional capacity of mitochondria to buffer cytosolic Ca2+ increase, which in turn may augment other, as yet undefined, Ca2+-dependent pathways. Alternatively, mitochondrial fission accompanies other detrimental changes in mitochondria, such as the opening of the mitochondrial permeability transition pore (mPTP). Degeneration of explanted sciatic nerve, a model of Wallerian degeneration, induces mitochondrial swelling and axonal degeneration. In addition, pharmacological inhibition or knocking down of cyclophilin D, a component of mPTP, attenuates axonal degeneration, suggesting that mPTP contributes to axonal degeneration of the sciatic nerves (Barrientos et al., 2011). At the site of capsaicin application, mitochondrial swelling occurs at the axonal terminals in vivo (Szolcsanyi et al., 1975; Chiang et al., 2015). Since the capsaicin-induced increase in cytosolic Ca2+ is sequestered to mitochondria (Wang et al., 2017b), and mitochondrial Ca2+ overloading triggers the opening of mPTP (Duchen, 2004), capsaicin may lead to the opening of mPTP to induce axonal ablation. Furthermore, capsaicin-induced death of DRG neurons is attenuated by pharmacological inhibition of mPTP (Shin et al., 2003). The opening of mPTP is enhanced in the presence of reactive oxygen species (ROS); however, dissipation of the mitochondrial membrane potential and inhibition of mPTP or ROS are not critical contributors to capsaicin-induced ablation of TRPV1-expressing nerve terminals (Wang et al., 2017b) (Fig. 4F). Of note, capsaicin also directly interferes with mitochondrial function. In isolated mitochondria from rat liver, capsaicin inhibits respiration (Chudapongse and Janthasoot, 1976). This direct toxicity of capsaicin on mitochondria occurs at high concentrations, the IC50 is approximately 20 μg/mg protein, and the concentration of capsaicin in reacting solutions is approximately 88 μM. Nevertheless, it would be difficult to unambiguously delineate the contribution of direct or indirect effects of capsaicin on mitochondria, or to connect these effects to capsaicin-induced nerve terminal ablation in vivo. Consequently, the role of mitochondria in capsaicin-induced nerve terminal ablation remains an open question.
6.3. Disruption of axonal cytoskeleton
Disruption of cytoskeleton in axons contributes to axonal degeneration and pathology following axonal injury. The axonal cytoskeleton is mainly composed of neurofilaments and microtubules. Axonal microtubules are stiff cytoskeletal structures that contribute to the maintenance of structural integrity and intracellular molecular traffic (Fletcher and Mullins, 2010). Neurofilaments are intermediate filaments and are less stiff than microtubules. Neurofilaments are crosslinked to one another, forming a structural matrix of axons. Axonal degeneration involves degradation of axonal neurofilaments as well as disruption of microtubules (Ma, 2013). The mechanisms of microtubule degradation likely involve Ca2+-dependent processes. The polymerization of microtubules is sensitive to Ca2+, and high concentrations of Ca2+ enhance depolymerization, presumably by increasing the GTP hydrolysis rate (O’Brien et al., 1997).
Capsaicin inhibits anterograde and retrograde axoplasmic transport by causing disorganization of axonal microtubules and neurofilaments in dissociated sensory neurons. This disorganization is accompanied by an increase in axoplasmic Ca2+ (Kawakami et al., 1993). Capsaicin mediates Ca2+ influx through TRPV1 activation, which likely triggers the Ca2+-dependent processes that enhance depolymerization and disrupt the axonal cytoskeleton. Close interactions of TRPV1 with microtubules likely enhance disruption of microtubules following TRPV1 activation. The carboxy terminal domain of TRPV1 interacts with and stabilizes microtubules (Goswami et al., 2004); activation of TRPV1 results in the disruption of dynamic microtubules containing tyrosinated tubulins but not of stable microtubules or neurofilaments (Goswami et al., 2006). TRPV1-mediated reorganization of microtubules is not entirely dependent on Ca2+ influx (Goswami and Hucho, 2007; Goswami et al., 2007; Goswami et al., 2011), suggesting that interactions between microtubules and TRPV1 are complicated. Overall, these reports suggest that vanilloid-induced activation of TRPV1 may result in cytoskeleton disorganization, and therefore may induce ablation of axons.
6.4. Activation of calpains
Calpains are Ca2+-dependent cysteine proteases that fulfill various critical physiological functions (Goll et al., 2003). Aberrant activation of calpains has been associated with multiple disorders, such as traumatic brain injury, spinal cord injury, neurodegenerative diseases, and peripheral neuropathies (Ma, 2013). Among 15 mammalian calpains, the best characterized are calpain-1 (μ-calpain) and calpain-2 (m-calpain). These two subtypes show different Ca2+ sensitivities, and calpain-2 requires a higher Ca2+ concentration for proteolytic activity than calpain-1 (Goll et al., 2003). The association between calpains and axotomy-induced axonal degeneration is established. Axonal degeneration of sensory nerves following axonal injury or damage by chemotherapeutic agents is attenuated by pharmacological inhibition of calpains (George et al., 1995; Wang et al., 2004). Regulation of calpain activity by calpastatin, an endogenous inhibitor of calpain, determines the extent of axonal degeneration following axonal injury (Ma et al., 2013; Yang et al., 2013). Shear stress injury in cultured neurons induces focal increases in intra-axonal Ca2+ and calpain; these foci produce axonal beadings. These findings suggest a causal relationship between localized axonal increases in Ca2+, calpain dynamics, and the formation of axonal beading (Kilinc et al., 2009). Because cytoskeletal components, such as neurofilaments and microtubules, are proteolytic targets of calpains, C2+-dependent activation of calpains is thought to contribute to axonal degeneration following axonal injury (Ma, 2013). Both calpain-1 and calpain-2 are expressed in sensory ganglia neurons. Calpain-2 is expressed both in large myelinated neurons and CGRP-positive neurons in DRG, and is upregulated following neuropathic injury (Zang et al., 2015). Calpain-1 is upregulated in DRG neurons in a rat model of diabetic neuropathy, and calpain inhibitors can attenuate capsaicin-induced death of dissociated DRG neurons (Chard et al., 1995). Multiple lines of evidence suggest that calpains play a key role in capsaicin-induced axonal ablation (Wang et al., 2017b): (1) MDL28170, an inhibitor of the Ca2+-dependent calpain protease activity, and overexpression of calpastatin or knockdown of calpain-2 diminishes capsaicin-induced axonal beading and ablation. (2) Histological assessment or longitudinal in vivo imaging of epidermal nerve terminals in the hindpaws of TRPV1-lineage reporter mice shows that MDL28170 prevents capsaicin-induced ablation (Wang et al., 2017b; Wang et al., 2020) (Fig. 5A). (3) Capsaicin-induced calpain-dependent ablation of TRPV1+ nerve terminals is necessary for analgesia (Wang et al., 2020). In mice with ION-CCI, capsaicin-induced hyperalgesia for neuropathic pain is substantially prevented by pretreatment with MDL28170 (Wang et al., 2020) (Fig. 5B). In aggregate, these findings indicate that TRPV1/Ca2+/calpain-dependent signaling is a dominant contributor to capsaicin-induced analgesia for neuropathic pain.
Figure 5. Capsaicin-induced calpain-mediated ablation is necessary for capsaicin-induced analgesia of neuropathic pain.
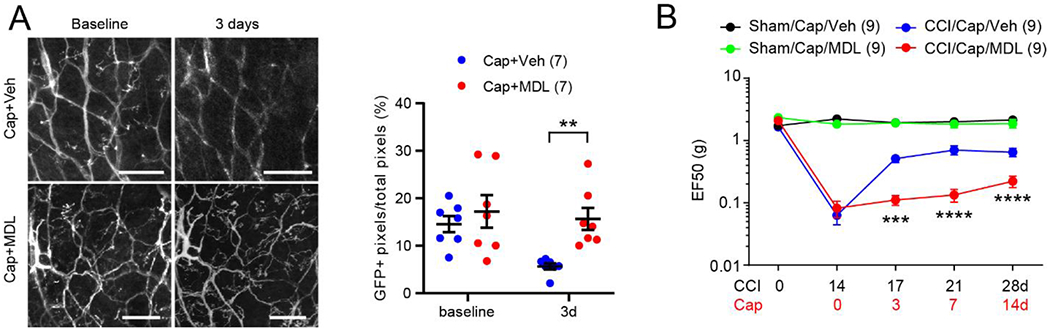
A. Longitudinal in vivo two-photon imaging of GFP+ afferent terminals in intact hindpaw of a mouse expressing membrane-bound GFP from the TRPV1 locus. (Left) To assess changes in GFP+ afferent terminals, the same site was imaged repeatedly – before (Baseline) and 3 days after intraplantar injection of vehicle (Veh) or MDL28170 (MDL; a calpain inhibitor) followed by capsaicin. (Right) The proportion of GFP+ pixels/total pixels in the imaged area was calculated. N=7 hindpaws from 5 mice per group. **p<0.01 in Sidak posthoc assay following two-way ANOVA.
B. Mechanical hyperalgesia in facial skin following chronic constriction injury of infraorbital nerve (ION-CCI) is attenuated by intradermal injection of capsaicin. Co-administration of capsaicin with MDL abolishes capsaicin-induced analgesia (compare CCI/Cap/MDL (red) with CCI/Cap/Veh (blue)). Two-way RM ANOVA followed by Bonferroni post-test (CCI/Cap/Veh vs CCI/Cap/MDL); ***p<0.0005, ****<0.0001. Reproduced from Wang et al., 2020 (Wang et al., 2020).
Despite the dominant role of calpain, it is likely that effects mediated by microtubules, mitochondria, and calpains cooperate to induce axonal degeneration. In shear stress injury of cultured neurons, calpain activity, microtubule disruption, and mitochondria co-localize within localized areas of axonal swelling (Kilinc et al., 2008, 2009). Microtubule stability is also affected by calpains (Billger et al., 1988). Although it is unclear whether calpains degrade microtubules, it is known that calpains degrade microtubule-associated proteins that regulate the stability of microtubules, which results in proteolysis of tubulin (Billger et al., 1988). Therefore, enhanced activation of calpains may indirectly alter the stability of microtubules. Microtubules and mitochondria also interact with one another during axotomy-induced axonal degeneration (Park et al., 2013). Energy depletion is an early phenomenon following axotomy, and is associated with axonal microtubule organization. Such destabilization of microtubules in turn induces mitochondrial swelling. Therefore, microtubules, mitochondria, and calpains may dynamically interact with one another throughout the axonal degenerative processes following axotomy, and similar interactions may also occur in capsaicin-induced axonal ablation.
Despite their potential similarity, the mechanisms underlying capsaicin-induced axonal ablation may be distinct from those of Wallerian degeneration, and therefore, must be determined independently. Vanilloid-induced axonal ablation apparently occurs more rapidly than axotomy-induced Wallerian degeneration (Ma et al., 2013). Ablation of epidermal nerve terminals occurs as early as 6 hours following the injection of capsaicin into the hindpaw, and peaks after 1 day (Wang et al., 2017b). In contrast, axotomy-induced axonal degeneration involves more complex initiation processes during the 24 hour latency phase before the onset of axonal degeneration. Swift elevation of Ca2+ following capsaicin application is likely to induce a relatively rapid ‘chemical axotomy’ in localized regions of axonal terminals, which may be followed by Wallerian degeneration-like processes within ablated axonal fragments. Therefore, it is possible that the same factors that contribute to capsaicin-induced ablation may also act to induce degenerative processes, as suggested in axotomy-induced Wallerian degeneration. From a therapeutic standpoint, it is important to identify the mechanisms underlying early chemical axotomy following capsaicin application. Understanding these mechanisms may be helpful for developing novel approaches aimed at improving analgesia through enhancing selective ablation by vanilloids with less pungency and greater efficacy.
7. Conclusion
The medical benefits of capsaicin have been suggested for more than a hundred and fifty years, and numerous studies have been conducted in animal and human populations to understand its clinical uses. High concentrations of topical capsaicin have been found to be effective for treating chronic pain, and the 8% capsaicin patch is approved in the US and EU for post-herpetic neuralgia, HIV-associated painful neuropathy, and painful diabetic neuropathy. Topical capsaicin is particularly effective for treating conditions in which hyperalgesia is localized and driven by hyperexcitable nociceptors. Topical capsaicin treats not only ongoing spontaneous pain, but also mechanical hyperalgesia and mechanical allodynia. The efficacy is comparable or even superior to oral analgesics, such as pregabalin. The safety of topical capsaicin is well validated, and adverse side effects, such as procedural pain, erythema, or reduced heat sensitivity, are considered to be minor issues. The patch form is optimal for targeting large areas of skin (e.g., post-herpetic neuralgia or diabetic neuropathy), whereas the injectable form is under development for precise delivery of capsaicin to focal lesions (e.g., Morton’s neuroma or intraoral post-operative pain). Understanding neurobiological mechanisms of capsaicin-induced analgesia is important for further improving therapy by decreasing procedural pain, attenuating pathological pain without interfering with physiological sensation, and establishing criteria for prediction of patient populations who will respond well to the therapy. Capsaicin-induced analgesia is also an excellent model for investigating the contribution of nociceptive primary afferents to the development and maintenance of chronic pain conditions. Different doses of capsaicin produce different degrees and durations of analgesia through mechanistically distinct effects on the function and structure of primary afferent terminals. Transient modest analgesia achieved using low doses of capsaicin involves the desensitization of capsaicin receptors and short-term defunctionalization of nerve endings, whereas long-lasting analgesia that persists for several months achieved by high doses of capsaicin is mediated by long-term defunctionalization via ablation of afferent terminals. TRPV1/Ca2+/calpain-mediated ablation of axonal terminals by capsaicin is necessary for analgesic treatment of neuropathic pain. In combination with calpain, axonal mitochondrial dysfunction and microtubule disorganization may also contribute. Elucidating the molecular and neural circuitry mechanisms underlying capsaicin-induced analgesia will enhance our understanding of peripheral mechanisms of neuropathic pain.
Acknowledgment
This work was supported by the National Institutes of Health R01 DE027731 and R35 DE030045 to MKC.
Abbreviations
- CGRP
calcitonin gene-related peptide
- DNIC
descending noxious inhibitory control
- DRG
dorsal root ganglia
- ENFD
epidermal nerve fiber density
- FDA
U.S. Food and Drug Administration
- GFP
green fluorescence protein
- HIV
human immunodeficiency virus
- ION-CCI
chronic constriction injury of infraorbital nerve
- OA
osteoarthritis
- mPTP
mitochondrial permeability transition pore
- NGF
nerve growth factor
- PHN
postherpetic neuralgia
- RCT
randomized controlled trials
- RTX
resiniferatoxin
- TRPV1
transient receptor potential vanilloid subfamily member 1
- VDCC
voltage-dependent calcium channel
- VDSC
voltage-dependent sodium channels
- WOMAC
Western Ontario and McMaster Universities Osteoarthritis Index
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Conflict of interest statement
James N. Campbell is on the Board of Directors and is an employee of Centrexion Therapeutics. Centrexion is involved in the commercial development of capsaicin for the treatment of pain. The other authors have no conflicts of interest to declare.
References
- Aasvang EK, Hansen JB, Malmstrom J, Asmussen T, Gennevois D, Struys MM, Kehlet H (2008) The effect of wound instillation of a novel purified capsaicin formulation on postherniotomy pain: a double-blind, randomized, placebo-controlled study. Anesth Analg 107:282–291. [DOI] [PubMed] [Google Scholar]
- Abdullah M, Mahowald ML, Frizelle SP, Dorman CW, Funkenbusch SC, Krug HE (2016) The effect of intra-articular vanilloid receptor agonists on pain behavior measures in a murine model of acute monoarthritis. J Pain Res 9:563–570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abooj M, Bishnoi M, Bosgraaf CA, Premkumar LS (2016) Changes in Spinal Cord Following Inflammatory and Neuropathic Pain and the Effectiveness of Resiniferatoxin. Open Pain J 9:1–4. [Google Scholar]
- Alsalem M, Millns P, Altarifi A, El-Salem K, Chapman V, Kendall DA (2016) Anti-nociceptive and desensitizing effects of olvanil on capsaicin-induced thermal hyperalgesia in the rat. BMC Pharmacol Toxicol 17:31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altman RD, Aven A, Holmburg CE, Pfeifer LM, Sack M, Young GT (1994) Capsaicin cream 0.025% as monotherapy for osteoarthritis: a double-blind study. Seminars in arthritis and rheumatism. Seminars in Arthritis and Rheumatism 23:25–33. [Google Scholar]
- Anand P, Bley K (2011) Topical capsaicin for pain management: therapeutic potential and mechanisms of action of the new high-concentration capsaicin 8% patch. Br J Anaesth 107:490–502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anand P, Elsafa E, Privitera R, Naidoo K, Yiangou Y, Donatien P, Gabra H, Wasan H, Kenny L, Rahemtulla A, Misra P (2019) Rational treatment of chemotherapy-induced peripheral neuropathy with capsaicin 8% patch: from pain relief towards disease modification. J Pain Res 12:2039–2052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Babbar S, Chanda S, Bley K (2010) Inhibition and induction of human cytochrome P450 enzymes in vitro by capsaicin. Xenobiotica 40:807–816. [DOI] [PubMed] [Google Scholar]
- Babbar S, Marier JF, Mouksassi MS, Beliveau M, Vanhove GF, Chanda S, Bley K (2009) Pharmacokinetic analysis of capsaicin after topical administration of a high-concentration capsaicin patch to patients with peripheral neuropathic pain. Ther Drug Monit 31:502–510. [DOI] [PubMed] [Google Scholar]
- Backonja M, Wallace MS, Blonsky ER, Cutler BJ, Malan P, Jr., Rauck R, Tobias J, Group N-CS (2008) NGX-4010, a high-concentration capsaicin patch, for the treatment of postherpetic neuralgia: a randomised, double-blind study. Lancet Neurol 7:1106–1112. [DOI] [PubMed] [Google Scholar]
- Bannister K, Sachau J, Baron R, Dickenson AH (2020) Neuropathic Pain: Mechanism-Based Therapeutics. Annu Rev Pharmacol Toxicol 60:257–274. [DOI] [PubMed] [Google Scholar]
- Barker PA, Mantyh P, Arendt-Nielsen L, Viktrup L, Tive L (2020) Nerve Growth Factor Signaling and Its Contribution to Pain. J Pain Res 13:1223–1241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baron R et al. (2017) Peripheral Neuropathic Pain: A mechanism-related organizing principle based on sensory profiles. Pain 158:261–272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrientos SA, Martinez NW, Yoo S, Jara JS, Zamorano S, Hetz C, Twiss JL, Alvarez J, Court FA (2011) Axonal degeneration is mediated by the mitochondrial permeability transition pore. J Neurosci 31:966–978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barton NJ, McQueen DS, Thomson D, Gauldie SD, Wilson AW, Salter DM, Chessell IP (2006) Attenuation of experimental arthritis in TRPV1R knockout mice. Exp Mol Pathol 81:166–170. [DOI] [PubMed] [Google Scholar]
- Basith S, Cui M, Hong S, Choi S (2016) Harnessing the Therapeutic Potential of Capsaicin and Its Analogues in Pain and Other Diseases. Molecules 21:966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bechakra M, Schuttenhelm BN, Pederzani T, van Doorn PA, de Zeeuw CI, Jongen JLM (2017) The reduction of intraepidermal P2X3 nerve fiber density correlates with behavioral hyperalgesia in a rat model of nerve injury-induced pain. J Comp Neurol 525:3757–3768. [DOI] [PubMed] [Google Scholar]
- Billger M, Wallin M, Karlsson JO (1988) Proteolysis of tubulin and microtubule-associated proteins 1 and 2 by calpain I and II. Difference in sensitivity of assembled and disassembled microtubules. Cell Calcium 9:33–44. [DOI] [PubMed] [Google Scholar]
- Binder A, Baron R (2016) The Pharmacological Therapy of Chronic Neuropathic Pain. Dtsch Arztebl Int 113:616–625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bley K, Boorman G, Mohammad B, McKenzie D, Babbar S (2012) A comprehensive review of the carcinogenic and anticarcinogenic potential of capsaicin. Toxicol Pathol 40:847–873. [DOI] [PubMed] [Google Scholar]
- Bonezzi C, Costantini A, Cruccu G, Fornasari DMM, Guardamagna V, Palmieri V, Polati E, Zini P, Dickenson AH (2020) Capsaicin 8% dermal patch in clinical practice: an expert opinion. Expert Opin Pharmacother 21:1377–1387. [DOI] [PubMed] [Google Scholar]
- Borbiro I, Badheka D, Rohacs T (2015) Activation of TRPV1 channels inhibits mechanosensitive Piezo channel activity by depleting membrane phosphoinositides. Sci Signal 8:ra15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brederson JD, Kym PR, Szallasi A (2013) Targeting TRP channels for pain relief. Eur J Pharmacol 716:61–76. [DOI] [PubMed] [Google Scholar]
- Brown JD, Saeed M, Do L, Braz J, Basbaum AI, Iadarola MJ, Wilson DM, Dillon WP (2015) CT-guided injection of a TRPV1 agonist around dorsal root ganglia decreases pain transmission in swine. Sci Transl Med 7:305ra145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown S, Simpson DM, Moyle G, Brew BJ, Schifitto G, Larbalestier N, Orkin C, Fisher M, Vanhove GF, Tobias JK (2013) NGX-4010, a capsaicin 8% patch, for the treatment of painful HIV-associated distal sensory polyneuropathy: integrated analysis of two phase III, randomized, controlled trials. AIDS Res Ther 10:5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caires R, Luis E, Taberner FJ, Fernandez-Ballester G, Ferrer-Montiel A, Balazs EA, Gomis A, Belmonte C, de la Pena E (2015) Hyaluronan modulates TRPV1 channel opening, reducing peripheral nociceptor activity and pain. Nat Commun 6:8095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campbell BK, Fillingim RB, Lee S, Brao R, Price DD, Neubert JK (2017) Effects of High-Dose Capsaicin on TMD Subjects: A Randomized Clinical Study. JDR Clin Trans Res 2:58–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campbell CM, Diamond E, Schmidt WK, Kelly M, Allen R, Houghton W, Brady KL, Campbell JN (2016) A randomized, double blind, placebo controlled trial of injected capsaicin for pain in Morton’s neuroma. Pain 157:1297–1304. [DOI] [PubMed] [Google Scholar]
- Campbell JN, Meyer RA (2006) Mechanisms of neuropathic pain. Neuron 52:77–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caro XJ, Winter EF (2014) Evidence of abnormal epidermal nerve fiber density in fibromyalgia: clinical and immunologic implications. Arthritis Rheumatol 66:1945–1954. [DOI] [PubMed] [Google Scholar]
- Carpenter SE, Lynn B (1981) Vascular and sensory responses of human skin to mild injury after topical treatment with capsaicin. Br J Pharmacol 73:755–758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D (1997) The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 389:816–824. [DOI] [PubMed] [Google Scholar]
- Caterina MJ, Leffler A, Malmberg AB, Martin WJ, Trafton J, Petersen-Zeitz KR, Koltzenburg M, Basbaum AI, Julius D (2000) Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 288:306–313. [DOI] [PubMed] [Google Scholar]
- Cavanaugh DJ, Lee H, Lo L, Shields SD, Zylka MJ, Basbaum AI, Anderson DJ (2009) Distinct subsets of unmyelinated primary sensory fibers mediate behavioral responses to noxious thermal and mechanical stimuli. Proc Natl Acad Sci U S A 106:9075–9080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chanda S, Bashir M, Babbar S, Koganti A, Bley K (2008) In vitro hepatic and skin metabolism of capsaicin. Drug Metab Dispos 36:670–675. [DOI] [PubMed] [Google Scholar]
- Chard PS, Bleakman D, Savidge JR, Miller RJ (1995) Capsaicin-induced neurotoxicity in cultured dorsal root ganglion neurons: involvement of calcium-activated proteases. Neuroscience 65:1099–1108. [DOI] [PubMed] [Google Scholar]
- Chen W, Chi YN, Kang XJ, Liu QY, Zhang HL, Li ZH, Zhao ZF, Yang Y, Su L, Cai J, Liao FF, Yi M, Wan Y, Liu FY (2018) Accumulation of Cav3.2 T-type Calcium Channels in the Uninjured Sural Nerve Contributes to Neuropathic Pain in Rats with Spared Nerve Injury. Front Mol Neurosci 11:24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiang H, Ohno N, Hsieh YL, Mahad DJ, Kikuchi S, Komuro H, Hsieh ST, Trapp BD (2015) Mitochondrial fission augments capsaicin-induced axonal degeneration. Acta Neuropathol 129:81–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christoph T, Gillen C, Mika J, Grunweller A, Schafer MK, Schiene K, Frank R, Jostock R, Bahrenberg G, Weihe E, Erdmann VA, Kurreck J (2007) Antinociceptive effect of antisense oligonucleotides against the vanilloid receptor VR1/TRPV1. Neurochem Int 50:281–290. [DOI] [PubMed] [Google Scholar]
- Chudapongse P, Janthasoot W (1976) Studies on the effect of capsaicin on metabolic reactions of isolated rat liver mitochondria. Toxicol Appl Pharmacol 37:263–270. [DOI] [PubMed] [Google Scholar]
- Chung K, Klein CM, Coggeshall RE (1990) The receptive part of the primary afferent axon is most vulnerable to systemic capsaicin in adult rats. Brain Res 511:222–226. [DOI] [PubMed] [Google Scholar]
- Chung MK, Wang S (2011) Cold Suppresses Agonist-induced Activation of TRPV1. J Dent Res 90:1098–1102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung MK, Campbell JN (2016) Use of Capsaicin to Treat Pain: Mechanistic and Therapeutic Considerations. Pharmaceuticals (Basel) 9:E66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung MK, Jung SJ, Oh SB (2011) Role of TRP Channels in Pain Sensation. Adv Exp Med Biol 704:615–636. [DOI] [PubMed] [Google Scholar]
- Cliff MA, Green BG (1996) Sensitization and desensitization to capsaicin and menthol in the oral cavity: interactions and individual differences. Physiol Behav 59:487–494. [DOI] [PubMed] [Google Scholar]
- Crow M, Khovanov N, Kelleher JH, Sharma S, Grant AD, Bogdanov Y, Wood JN, McMahon SB, Denk F (2015) HDAC4 is required for inflammation-associated thermal hypersensitivity. FASEB J 29:3370–3378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cruccu G, Nurmikko TJ, Ernault E, Riaz FK, McBride WT, Haanpaa M (2018) Superiority of capsaicin 8% patch versus oral pregabalin on dynamic mechanical allodynia in patients with peripheral neuropathic pain. Eur J Pain 22:700–706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Culshaw AJ et al. (2006) Identification and biological characterization of 6-aryl-7-isopropylquinazolinones as novel TRPV1 antagonists that are effective in models of chronic pain. J Med Chem 49:471–474. [DOI] [PubMed] [Google Scholar]
- Da Silva JT, Zhang Y, Asgar J, Ro JY, Seminowicz DA (2018) Diffuse noxious inhibitory controls and brain networks are modulated in a testosterone-dependent manner in Sprague Dawley rats. Behav Brain Res 349:91–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dahlhamer J, Lucas J, Zelaya C, Nahin R, Mackey S, DeBar L, Kerns R, Von Korff M, Porter L, Helmick C (2018) Prevalence of Chronic Pain and High-Impact Chronic Pain Among Adults - United States, 2016. MMWR Morb Mortal Wkly Rep 67:1001–1006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dai WL, Yan B, Bao YN, Fan JF, Liu JH (2020) Suppression of peripheral NGF attenuates neuropathic pain induced by chronic constriction injury through the TAK1-MAPK/NF-kappaB signaling pathways. Cell Commun Signal 18:66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis KD, Meyer RA, Turnquist JL, Filloon TG, Pappagallo M, Campbell JN (1995a) Cutaneous injection of the capsaicin analogue, NE-21610, produces analgesia to heat but not to mechanical stimuli in man. Pain 61:17–26. [DOI] [PubMed] [Google Scholar]
- Davis KD, Meyer RA, Turnquist JL, Filloon TG, Pappagallo M, Campbell JN (1995b) Cutaneous pretreatment with the capsaicin analog NE-21610 prevents the pain to a burn and subsequent hyperalgesia. Pain 62:373–378. [DOI] [PubMed] [Google Scholar]
- De Petrocellis L, Guida F, Moriello AS, De Chiaro M, Piscitelli F, de Novellis V, Maione S, Di Marzo V (2011) N-palmitoyl-vanillamide (palvanil) is a non-pungent analogue of capsaicin with stronger desensitizing capability against the TRPV1 receptor and anti-hyperalgesic activity. Pharmacol Res 63:294–299. [DOI] [PubMed] [Google Scholar]
- Denda M, Fuziwara S, Inoue K, Denda S, Akamatsu H, Tomitaka A, Matsunaga K (2001) Immunoreactivity of VR1 on epidermal keratinocyte of human skin. Biochem Biophys Res Commun 285:1250–1252. [DOI] [PubMed] [Google Scholar]
- Derry S, Moore RA (2012) Topical capsaicin (low concentration) for chronic neuropathic pain in adults. Cochrane Database Syst Rev: CD010111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derry S, Sven-Rice A, Cole P, Tan T, Moore RA (2013) Topical capsaicin (high concentration) for chronic neuropathic pain in adults. Cochrane Database Syst Rev: CD007393. [DOI] [PubMed] [Google Scholar]
- Derry S, Rice AS, Cole P, Tan T, Moore RA (2017) Topical capsaicin (high concentration) for chronic neuropathic pain in adults. Cochrane Database Syst Rev 1:CD007393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dobson GP, Letson HL, Grant A, McEwen P, Hazratwala K, Wilkinson M, Morris JL (2018) Defining the osteoarthritis patient: back to the future. Osteoarthritis Cartilage 26:1003–1007. [DOI] [PubMed] [Google Scholar]
- Dray A, Bettaney J, Forster P (1990) Actions of capsaicin on peripheral nociceptors of the neonatal rat spinal cord-tail in vitro: dependence of extracellular ions and independence of second messengers. Br J Pharmacol 101:727–733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duchen MR (2004) Roles of mitochondria in health and disease. Diabetes 53 Suppl 1:S96–102. [DOI] [PubMed] [Google Scholar]
- Duraku LS, Hossaini M, Schuttenhelm BN, Holstege JC, Baas M, Ruigrok TJ, Walbeehm ET (2013) Reinnervation patterns by peptidergic Substance-P, non-peptidergic P2X3, and myelinated NF-200 nerve fibers in epidermis and dermis of rats with neuropathic pain. Exp Neurol 241:13–24. [DOI] [PubMed] [Google Scholar]
- Engler A, Aeschlimann A, Simmen BR, Michel BA, Gay RE, Gay S, Sprott H (2007) Expression of transient receptor potential vanilloid 1 (TRPV1) in synovial fibroblasts from patients with osteoarthritis and rheumatoid arthritis. Biochem Biophys Res Commun 359:884–888. [DOI] [PubMed] [Google Scholar]
- Epstein JB, Marcoe JH (1994) Topical application of capsaicin for treatment of oral neuropathic pain and trigeminal neuralgia. Oral Surg Oral Med Oral Pathol 77:135–140. [DOI] [PubMed] [Google Scholar]
- Facer P, Casula MA, Smith GD, Benham CD, Chessell IP, Bountra C, Sinisi M, Birch R, Anand P (2007) Differential expression of the capsaicin receptor TRPV1 and related novel receptors TRPV3, TRPV4 and TRPM8 in normal human tissues and changes in traumatic and diabetic neuropathy. BMC Neurol 7:11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fattori V, Hohmann MS, Rossaneis AC, Pinho-Ribeiro FA, Verri WA (2016) Capsaicin: Current Understanding of Its Mechanisms and Therapy of Pain and Other Pre-Clinical and Clinical Uses. Molecules 21:844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fields HL, Rowbotham M, Baron R (1998) Postherpetic neuralgia: irritable nociceptors and deafferentation. Neurobiol Dis 5:209–227. [DOI] [PubMed] [Google Scholar]
- Filipczak-Bryniarska I, Krzyzewski RM, Kucharz J, Michalowska-Kaczmarczyk A, Kleja J, Woron J, Strzepek K, Kazior L, Wordliczek J, Grodzicki T, Krzemieniecki K (2017) High-dose 8% capsaicin patch in treatment of chemotherapy-induced peripheral neuropathy: single-center experience. Med Oncol 34:162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fillingim RB (2017) Individual differences in pain: understanding the mosaic that makes pain personal. Pain 158 Suppl 1:S11–S18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finnerup NB, Attal N, Haroutounian S, McNicol E, Baron R, Dworkin RH, Gilron I, Haanpaa M, Hansson P, Jensen TS, Kamerman PR, Lund K, Moore A, Raja SN, Rice AS, Rowbotham M, Sena E, Siddall P, Smith BH, Wallace M (2015) Pharmacotherapy for neuropathic pain in adults: a systematic review and meta-analysis. Lancet Neurol 14:162–173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fletcher DA, Mullins RD (2010) Cell mechanics and the cytoskeleton. Nature 463:485–492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forstenpointner J, Naleschinski D, Wasner G, Hullemann P, Binder A, Baron R (2019) Sensitized vasoactive C-nociceptors: key fibers in peripheral neuropathic pain. Pain Rep 4:e709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forstenpointner J, Forster M, May D, Hofschulte F, Cascorbi I, Wasner G, Gierthmuhlen J, Baron R (2017) Short Report: TRPV1-polymorphism 1911 A>G alters capsaicin-induced sensory changes in healthy subjects. PLoS One 12:e0183322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frey E, Karney-Grobe S, Krolak T, Milbrandt J, DiAntonio A (2018) TRPV1 Agonist, Capsaicin, Induces Axon Outgrowth after Injury via Ca(2+)/PKA Signaling. eNeuro 5: e0095–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friel NA, McNickle AG, DeFranco MJ, Wang F, Shewman EF, Verma NN, Cole BJ, Bach BR Jr., Chubinskaya S, Kramer SM, Wang VM (2015) Effect of highly purified capsaicin on articular cartilage and rotator cuff tendon healing: An in vivo rabbit study. J Orthop Res 33:1854–1860. [DOI] [PubMed] [Google Scholar]
- Galvez R, Navez ML, Moyle G, Maihofner C, Stoker M, Ernault E, Nurmikko TJ, Attal N (2017) Capsaicin 8% Patch Repeat Treatment in Nondiabetic Peripheral Neuropathic Pain: A 52-Week, Open-Label, Single-Arm, Safety Study. Clin J Pain 33:921–931. [DOI] [PubMed] [Google Scholar]
- Gear RW, Aley KO, Levine JD (1999) Pain-induced analgesia mediated by mesolimbic reward circuits. J Neurosci 19:7175–7181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- George EB, Glass JD, Griffin JW (1995) Axotomy-induced axonal degeneration is mediated by calcium influx through ion-specific channels. J Neurosci 15:6445–6452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geuter S, Koban L, Wager TD (2017) The Cognitive Neuroscience of Placebo Effects: Concepts, Predictions, and Physiology. Annu Rev Neurosci 40:167–188. [DOI] [PubMed] [Google Scholar]
- Goll DE, Thompson VF, Li H, Wei W, Cong J (2003) The calpain system. Physiol Rev 83:731–801. [DOI] [PubMed] [Google Scholar]
- Goswami C, Hucho T (2007) TRPV1 expression-dependent initiation and regulation of filopodia. J Neurochem 103:1319–1333. [DOI] [PubMed] [Google Scholar]
- Goswami C, Schmidt H, Hucho F (2007) TRPV1 at nerve endings regulates growth cone morphology and movement through cytoskeleton reorganization. FEBS J 274:760–772. [DOI] [PubMed] [Google Scholar]
- Goswami C, Dreger M, Otto H, Schwappach B, Hucho F (2006) Rapid disassembly of dynamic microtubules upon activation of the capsaicin receptor TRPV1. J Neurochem 96:254–266. [DOI] [PubMed] [Google Scholar]
- Goswami C, Dreger M, Jahnel R, Bogen O, Gillen C, Hucho F (2004) Identification and characterization of a Ca2+ -sensitive interaction of the vanilloid receptor TRPV1 with tubulin. J Neurochem 91:1092–1103. [DOI] [PubMed] [Google Scholar]
- Goswami C, Kuhn J, Dina OA, Fernandez-Ballester G, Levine JD, Ferrer-Montiel A, Hucho T (2011) Estrogen destabilizes microtubules through an ion-conductivity-independent TRPV1 pathway. J Neurochem 117:995–1008. [DOI] [PubMed] [Google Scholar]
- Green BG, Shaffer GS (1993) The sensory response to capsaicin during repeated topical exposures: differential effects on sensations of itching and pungency. Pain 53:323–334. [DOI] [PubMed] [Google Scholar]
- Green D, Ruparel S, Gao X, Ruparel N, Patil M, Akopian A, Hargreaves K (2016) Central activation of TRPV1 and TRPA1 by novel endogenous agonists contributes to mechanical allodynia and thermal hyperalgesia after burn injury. Mol Pain 12:1744806916661725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grelik C, Bennett GJ, Ribeiro-da-Silva A (2005) Autonomic fibre sprouting and changes in nociceptive sensory innervation in the rat lower lip skin following chronic constriction injury. Eur J Neurosci 21:2475–2487. [DOI] [PubMed] [Google Scholar]
- Haanpaa M, Cruccu G, Nurmikko TJ, McBride WT, Docu Axelarad A, Bosilkov A, Chambers C, Ernault E, Abdulahad AK (2016) Capsaicin 8% patch versus oral pregabalin in patients with peripheral neuropathic pain. Eur J Pain 20:316–328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall OM, Broussard A, Range T, Carroll Turpin MA, Ellis S, Lim VM, Cornett EM, Kaye AD (2020) Novel Agents in Neuropathic Pain, the Role of Capsaicin: Pharmacology, Efficacy, Side Effects, Different Preparations. Curr Pain Headache Rep 24:53. [DOI] [PubMed] [Google Scholar]
- Hamalainen MM, Subieta A, Arpey C, Brennan TJ (2009) Differential effect of capsaicin treatment on pain-related behaviors after plantar incision. J Pain 10:637–645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartrick CT, Pestano C, Carlson N, Hartrick S (2011) Capsaicin instillation for postoperative pain following total knee arthroplasty: a preliminary report of a randomized, double-blind, parallel-group, placebo-controlled, multicentre trial. Clin Drug Investig 31:877–882. [DOI] [PubMed] [Google Scholar]
- Helyes Z, Szabo A, Nemeth J, Jakab B, Pinter E, Banvolgyi A, Kereskai L, Keri G, Szolcsanyi J (2004) Antiinflammatory and analgesic effects of somatostatin released from capsaicin-sensitive sensory nerve terminals in a Freund’s adjuvant-induced chronic arthritis model in the rat. Arthritis Rheum 50:1677–1685. [DOI] [PubMed] [Google Scholar]
- Helyes Z, Sandor K, Borbely E, Tekus V, Pinter E, Elekes K, Toth DM, Szolcsanyi J, McDougall JJ (2010) Involvement of transient receptor potential vanilloid 1 receptors in protease-activated receptor-2-induced joint inflammation and nociception. Eur J Pain 14:351–358. [DOI] [PubMed] [Google Scholar]
- Henry JL (1976) Effects of substance P on functionally identified units in cat spinal cord. Brain Res 114:439–451. [DOI] [PubMed] [Google Scholar]
- Hill R (2000) NK1 (substance P) receptor antagonists--why are they not analgesic in humans? Trends Pharmacol Sci 21:244–246. [DOI] [PubMed] [Google Scholar]
- Hirai T, Enomoto M, Kaburagi H, Sotome S, Yoshida-Tanaka K, Ukegawa M, Kuwahara H, Yamamoto M, Tajiri M, Miyata H, Hirai Y, Tominaga M, Shinomiya K, Mizusawa H, Okawa A, Yokota T (2014) Intrathecal AAV serotype 9-mediated delivery of shRNA against TRPV1 attenuates thermal hyperalgesia in a mouse model of peripheral nerve injury. Mol Ther 22:409–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman EM, Watson JC, St Sauver J, Staff NP, Klein CJ (2017) Association of Long-term Opioid Therapy With Functional Status, Adverse Outcomes, and Mortality Among Patients With Polyneuropathy. JAMA Neurol 74:773–779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffmeister C, Trevisan G, Rossato MF, de Oliveira SM, Gomez MV, Ferreira J (2011) Role of TRPV1 in nociception and edema induced by monosodium urate crystals in rats. Pain 152:1777–1788. [DOI] [PubMed] [Google Scholar]
- Holzer P (1991) Capsaicin: cellular targets, mechanisms of action, and selectivity for thin sensory neurons. Pharmacol Rev 43:143–201. [PubMed] [Google Scholar]
- Holzer P (2008) The pharmacological challenge to tame the transient receptor potential vanilloid-1 (TRPV1) nocisensor. Br J Pharmacol 155:1145–1162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honore P et al. (2009) Repeated dosing of ABT-102, a potent and selective TRPV1 antagonist, enhances TRPV1-mediated analgesic activity in rodents, but attenuates antagonist-induced hyperthermia. Pain 142:27–35. [DOI] [PubMed] [Google Scholar]
- Hoper J, Helfert S, Heskamp ML, Maihofner CG, Baron R (2014) High concentration capsaicin for treatment of peripheral neuropathic pain: effect on somatosensory symptoms and identification of treatment responders. Curr Med Res Opin 30:565–574. [DOI] [PubMed] [Google Scholar]
- Horvath K, Boros M, Bagoly T, Sandor V, Kilar F, Kemeny A, Helyes Z, Szolcsanyi J, Pinter E (2014) Analgesic topical capsaicinoid therapy increases somatostatin-like immunoreactivity in the human plasma. Neuropeptides 48:371–378. [DOI] [PubMed] [Google Scholar]
- Hu F, Sun WW, Zhao XT, Cui ZJ, Yang WX (2008) TRPV1 mediates cell death in rat synovial fibroblasts through calcium entry-dependent ROS production and mitochondrial depolarization. Biochem Biophys Res Commun 369:989–993. [DOI] [PubMed] [Google Scholar]
- Hwang MK, Bode AM, Byun S, Song NR, Lee HJ, Lee KW, Dong Z (2010) Cocarcinogenic effect of capsaicin involves activation of EGFR signaling but not TRPV1. Cancer Res 70:6859–6869. [DOI] [PubMed] [Google Scholar]
- Iadarola MJ, Sapio MR, Raithel SJ, Mannes AJ, Brown DC (2018) Long-term pain relief in canine osteoarthritis by a single intra-articular injection of resiniferatoxin, a potent TRPV1 agonist. Pain 159:2105–2114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imamura Y, Shinozaki T, Okada-Ogawa A, Noma N, Shinoda M, Iwata K, Wada A, Abe O, Wang K, Svensson P (2019) An updated review on pathophysiology and management of burning mouth syndrome with endocrinological, psychological and neuropathic perspectives. J Oral Rehabil 46:574–587. [DOI] [PubMed] [Google Scholar]
- Irving GA, Backonja MM, Dunteman E, Blonsky ER, Vanhove GF, Lu SP, Tobias J, Group N-CS (2011) A multicenter, randomized, double-blind, controlled study of NGX-4010, a high-concentration capsaicin patch, for the treatment of postherpetic neuralgia. Pain Med 12:99–109. [DOI] [PubMed] [Google Scholar]
- Jancso G, Santha P (2015) The foundation of sensory pharmacology: Nicholas (Miklos) Jancso and the Szeged contribution. Temperature (Austin) 2:152–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jancso G, Kiraly E, Jancso-Gabor A (1977) Pharmacologically induced selective degeneration of chemosensitive primary sensory neurones. Nature 270:741–743. [DOI] [PubMed] [Google Scholar]
- Jancso G, Kiraly E, Joo F, Such G, Nagy A (1985) Selective degeneration by capsaicin of a subpopulation of primary sensory neurons in the adult rat. Neurosci Lett 59:209–214. [DOI] [PubMed] [Google Scholar]
- Jancso N, Jancso A (1949) Desensitization of sensory nerve endings. Orvostudomány 2:1–43. [Google Scholar]
- Jang JH, Kim KH, Nam TS, Lee WT, Park KA, Kim DW, Leem JW (2007) The role of uninjured C-afferents and injured afferents in the generation of mechanical hypersensitivity after partial peripheral nerve injury in the rat. Exp Neurol 204:288–298. [DOI] [PubMed] [Google Scholar]
- Jessell TM, Iversen LL, Cuello AC (1978) Capsaicin-induced depletion of substance P from primary sensory neurones. Brain Res 152:183–188. [DOI] [PubMed] [Google Scholar]
- Joseph J, Wang S, Lee J, Ro JY, Chung MK (2013) Carboxyl-terminal Domain of Transient Receptor Potential Vanilloid 1 Contains Distinct Segments Differentially Involved in Capsaicin- and Heat-induced Desensitization. J Biol Chem 288:35690–35702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joseph J, Qu L, Wang S, Kim M, Bennett D, Ro J, Caterina M, Chung MK (2019) Phosphorylation of TRPV1 S801 contributes to modality-specific hyperalgesia in mice. J Neurosci 39:9954–9966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalff KM, El Mouedden M, van Egmond J, Veening J, Joosten L, Scheffer GJ, Meert T, Vissers K (2010) Pre-treatment with capsaicin in a rat osteoarthritis model reduces the symptoms of pain and bone damage induced by monosodium iodoacetate. Eur J Pharmacol 641:108–113. [DOI] [PubMed] [Google Scholar]
- Kambiz S, Duraku LS, Baas M, Nijhuis TH, Cosgun SG, Hovius SE, Ruigrok TJ, Walbeehm ET (2015) Long-term follow-up of peptidergic and nonpeptidergic reinnervation of the epidermis following sciatic nerve reconstruction in rats. J Neurosurg 123:254–269. [DOI] [PubMed] [Google Scholar]
- Kanai Y, Nakazato E, Fujiuchi A, Hara T, Imai A (2005) Involvement of an increased spinal TRPV1 sensitization through its up-regulation in mechanical allodynia of CCI rats. Neuropharmacology 49:977–984. [DOI] [PubMed] [Google Scholar]
- Kantas B, Borzsei R, Szoke E, Banhegyi P, Horvath A, Hunyady A, Borbely E, Hetenyi C, Pinter E, Helyes Z (2019) Novel Drug-Like Somatostatin Receptor 4 Agonists are Potential Analgesics for Neuropathic Pain. Int J Mol Sci 20:6245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karai L, Brown DC, Mannes AJ, Connelly ST, Brown J, Gandal M, Wellisch OM, Neubert JK, Olah Z, Iadarola MJ (2004) Deletion of vanilloid receptor 1-expressing primary afferent neurons for pain control. J Clin Invest 113:1344–1352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlsson P, Provitera V, Caporaso G, Stancanelli A, Saltalamacchia AM, Borreca I, Manganelli F, Santoro L, Jensen TS, Nolano M (2020) Increased peptidergic fibers as a potential cutaneous marker of pain in diabetic small fiber neuropathy. Pain (in press). [DOI] [PubMed] [Google Scholar]
- Kato D, Suto T, Obata H, Saito S (2019) Spinal Activation of Tropomyosin Receptor Kinase-B Recovers the Impaired Endogenous Analgesia in Neuropathic Pain Rats. Anesth Analg 129:578–586. [DOI] [PubMed] [Google Scholar]
- Katz NP, Mou J, Paillard FC, Turnbull B, Trudeau J, Stoker M (2014) Predictors of Response in Patients with Post-herpetic Neuralgia and HIV-associated Neuropathy Treated with the 8% Capsaicin Patch (Qutenza(R)). Clin J Pain 31:859–866. [DOI] [PubMed] [Google Scholar]
- Katz NP, Mou J, Paillard FC, Turnbull B, Trudeau J, Stoker M (2015) Predictors of Response in Patients With Postherpetic Neuralgia and HIV-Associated Neuropathy Treated With the 8% Capsaicin Patch (Qutenza). Clin J Pain 31:859–866. [DOI] [PubMed] [Google Scholar]
- Kawakami T, Hikawa N, Kusakabe T, Kano M, Bandou Y, Gotoh H, Takenaka T (1993) Mechanism of inhibitory action of capsaicin on particulate axoplasmic transport in sensory neurons in culture. J Neurobiol 24:545–551. [DOI] [PubMed] [Google Scholar]
- Kelly S, Chapman RJ, Woodhams S, Sagar DR, Turner J, Burston JJ, Bullock C, Paton K, Huang J, Wong A, McWilliams DF, Okine BN, Barrett DA, Hathway GJ, Walsh DA, Chapman V (2015) Increased function of pronociceptive TRPV1 at the level of the joint in a rat model of osteoarthritis pain. Ann Rheum Dis 74:252–259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kemppainen P, Waltimo A, Waltimo T, Kononen M, Pertovaara A (1997) Differential effects of noxious conditioning stimulation of the cheek by capsaicin on human sensory and inhibitory masseter reflex responses evoked by tooth pulp stimulation. J Dent Res 76:1561–1568. [DOI] [PubMed] [Google Scholar]
- Kennedy WR, Vanhove GF, Lu SP, Tobias J, Bley KR, Walk D, Wendelschafer-Crabb G, Simone DA, Selim MM (2010) A randomized, controlled, open-label study of the long-term effects of NGX-4010, a high-concentration capsaicin patch, on epidermal nerve fiber density and sensory function in healthy volunteers. J Pain 11:579–587. [DOI] [PubMed] [Google Scholar]
- Kern KU, Nowack W, Poole C (2014) Treatment of neuropathic pain with the capsaicin 8% patch: is pretreatment with lidocaine necessary? Pain Pract 14:E42–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilinc D, Gallo G, Barbee KA (2008) Mechanically-induced membrane poration causes axonal beading and localized cytoskeletal damage. Exp Neurol 212:422–430. [DOI] [PubMed] [Google Scholar]
- Kilinc D, Gallo G, Barbee KA (2009) Mechanical membrane injury induces axonal beading through localized activation of calpain. Exp Neurol 219:553–561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim SM, Kim J, Kim E, Hwang SJ, Shin HK, Lee SE (2008) Local application of capsaicin alleviates mechanical hyperalgesia after spinal nerve transection. Neurosci Lett 433:199–204. [DOI] [PubMed] [Google Scholar]
- Kim Y, Kim EH, Lee KS, Lee K, Park SH, Na SH, Ko C, Kim J, Yooon YW (2016) The effects of intra-articular resiniferatoxin on monosodium iodoacetate-induced osteoarthritic pain in rats. Korean J Physiol Pharmacol 20:129–136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim YS, Chu Y, Han L, Li M, Li Z, Lavinka PC, Sun S, Tang Z, Park K, Caterina MJ, Ren K, Dubner R, Wei F, Dong X (2014) Central terminal sensitization of TRPV1 by descending serotonergic facilitation modulates chronic pain. Neuron 81:873–887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- King T, Qu C, Okun A, Mercado R, Ren J, Brion T, Lai J, Porreca F (2011) Contribution of afferent pathways to nerve injury-induced spontaneous pain and evoked hypersensitivity. Pain 152:1997–2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kissin EY, Freitas CF, Kissin I (2005a) The effects of intraarticular resiniferatoxin in experimental knee-joint arthritis. Anesth Analg 101:1433–1439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kissin I, Davison N, Bradley EL, Jr. (2005b) Perineural resiniferatoxin prevents hyperalgesia in a rat model of postoperative pain. Anesth Analg 100:774–780. [DOI] [PubMed] [Google Scholar]
- Kissin I, Freitas CF, Bradley EL Jr. (2007) Perineural resiniferatoxin prevents the development of hyperalgesia produced by loose ligation of the sciatic nerve in rats. Anesth Analg 104:1210–1216. [DOI] [PubMed] [Google Scholar]
- Knolle E, Zadrazil M, Kovacs GG, Medwed S, Scharbert G, Schemper M (2013) Comparison of cooling and EMLA to reduce the burning pain during capsaicin 8% patch application: a randomized, double-blind, placebo-controlled study. Pain 154:2729–2736. [DOI] [PubMed] [Google Scholar]
- Kochukov MY, McNearney TA, Fu Y, Westlund KN (2006) Thermosensitive TRP ion channels mediate cytosolic calcium response in human synoviocytes. Am J Physiol Cell Physiol 291:C424–432. [DOI] [PubMed] [Google Scholar]
- Koplas PA, Rosenberg RL, Oxford GS (1997) The role of calcium in the desensitization of capsaicin responses in rat dorsal root ganglion neurons. J Neurosci 17:3525–3537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Labuz D, Spahn V, Celik MO, Machelska H (2016) Opioids and TRPV1 in the peripheral control of neuropathic pain--Defining a target site in the injured nerve. Neuropharmacology 101:330–340. [DOI] [PubMed] [Google Scholar]
- LaMotte RH, Shain CN, Simone DA, Tsai EF (1991) Neurogenic hyperalgesia: psychophysical studies of underlying mechanisms. J Neurophysiol 66:190–211. [DOI] [PubMed] [Google Scholar]
- Landmann G, Lustenberger C, Schleinzer W, Schmelz M, Stockinger L, Rukwied R (2016) Short lasting transient effects of a capsaicin 8% patch on nociceptor activation in humans. Eur J Pain 20:1443–1453. [DOI] [PubMed] [Google Scholar]
- Landowski LM, Dyck PJ, Engelstad J, Taylor BV (2016) Axonopathy in peripheral neuropathies: Mechanisms and therapeutic approaches for regeneration. J Chem Neuroanat 76:19–27. [DOI] [PubMed] [Google Scholar]
- Lanteri-Minet M, Perrot S (2019) QAPSA: post-marketing surveillance of capsaicin 8% patch for long-term use in patients with peripheral neuropathic pain in France. Curr Med Res Opin 35:417–426. [DOI] [PubMed] [Google Scholar]
- Laslett LL, Jones G (2014) Capsaicin for osteoarthritis pain. Prog Drug Res 68:277–291. [DOI] [PubMed] [Google Scholar]
- Lauria G, Morbin M, Lombardi R, Capobianco R, Camozzi F, Pareyson D, Manconi M, Geppetti P (2006) Expression of capsaicin receptor immunoreactivity in human peripheral nervous system and in painful neuropathies. J Peripher Nerv Syst 11:262–271. [DOI] [PubMed] [Google Scholar]
- Lembeck F (1987) Columbus, Capsicum and capsaicin: past, present and future. Acta Physiol Hung 69:265–273. [PubMed] [Google Scholar]
- Li CL, Li KC, Wu D, Chen Y, Luo H, Zhao JR, Wang SS, Sun MM, Lu YJ, Zhong YQ, Hu XY, Hou R, Zhou BB, Bao L, Xiao HS, Zhang X (2016) Somatosensory neuron types identified by high-coverage single-cell RNA-sequencing and functional heterogeneity. Cell Res 26:967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu L, Oortgiesen M, Li L, Simon SA (2001) Capsaicin inhibits activation of voltage-gated sodium currents in capsaicin-sensitive trigeminal ganglion neurons. J Neurophysiol 85:745–758. [DOI] [PubMed] [Google Scholar]
- Lo Vecchio S, Andersen HH, Arendt-Nielsen L (2018) The time course of brief and prolonged topical 8% capsaicin-induced desensitization in healthy volunteers evaluated by quantitative sensory testing and vasomotor imaging. Exp Brain Res 236:2231–2244. [DOI] [PubMed] [Google Scholar]
- Lockwood SM, Dickenson AH (2019) A combination pharmacotherapy of tapentadol and pregabalin to tackle centrally driven osteoarthritis pain. Eur J Pain 23:1185–1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luongo L, Costa B, D’Agostino B, Guida F, Comelli F, Gatta L, Matteis M, Sullo N, De Petrocellis L, de Novellis V, Maione S, Di Marzo V (2012) Palvanil, a non-pungent capsaicin analogue, inhibits inflammatory and neuropathic pain with little effects on bronchopulmonary function and body temperature. Pharmacol Res 66:243–250. [DOI] [PubMed] [Google Scholar]
- Ma F, Zhang L, Lyons D, Westlund KN (2012) Orofacial neuropathic pain mouse model induced by Trigeminal Inflammatory Compression (TIC) of the infraorbital nerve. Mol Brain 5:44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma M (2013) Role of calpains in the injury-induced dysfunction and degeneration of the mammalian axon. Neurobiol Dis 60:61–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma M, Ferguson TA, Schoch KM, Li J, Qian Y, Shofer FS, Saatman KE, Neumar RW (2013) Calpains mediate axonal cytoskeleton disintegration during Wallerian degeneration. Neurobiol Dis 56:34–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma XL, Zhang FX, Dong F, Bao L, Zhang X (2015) Experimental evidence for alleviating nociceptive hypersensitivity by single application of capsaicin. Mol Pain 11:22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maier C et al. (2010) Quantitative sensory testing in the German Research Network on Neuropathic Pain (DFNS): somatosensory abnormalities in 1236 patients with different neuropathic pain syndromes. Pain 150:439–450. [DOI] [PubMed] [Google Scholar]
- Maihofner CG, Heskamp ML (2014) Treatment of peripheral neuropathic pain by topical capsaicin: Impact of pre-existing pain in the QUEPP-study. Eur J Pain 18:671–679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mainka T, Malewicz NM, Baron R, Enax-Krumova EK, Treede RD, Maier C (2016) Presence of hyperalgesia predicts analgesic efficacy of topically applied capsaicin 8% in patients with peripheral neuropathic pain. Eur J Pain 20:116–129. [DOI] [PubMed] [Google Scholar]
- Malmberg AB, Mizisin AP, Calcutt NA, von Stein T, Robbins WR, Bley KR (2004) Reduced heat sensitivity and epidermal nerve fiber immunostaining following single applications of a high-concentration capsaicin patch. Pain 111:360–367. [DOI] [PubMed] [Google Scholar]
- Manitpisitkul P, Flores CM, Moyer JA, Romano G, Shalayda K, Tatikola K, Hutchison JS, Mayorga AJ (2018) A multiple-dose double-blind randomized study to evaluate the safety, pharmacokinetics, pharmacodynamics and analgesic efficacy of the TRPV1 antagonist JNJ-39439335 (mavatrep). Scand J Pain 18:151–164. [DOI] [PubMed] [Google Scholar]
- Martini C, Yassen A, Olofsen E, Passier P, Stoker M, Dahan A (2012) Pharmacodynamic analysis of the analgesic effect of capsaicin 8% patch (Qutenza) in diabetic neuropathic pain patients: detection of distinct response groups. J Pain Res 5:51–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martini CH, Yassen A, Krebs-Brown A, Passier P, Stoker M, Olofsen E, Dahan A (2013) A novel approach to identify responder subgroups and predictors of response to low- and high-dose capsaicin patches in postherpetic neuralgia. Eur J Pain 17:1491–1501. [DOI] [PubMed] [Google Scholar]
- McMahon SB, Lewin G, Bloom SR (1991) The consequences of long-term topical capsaicin application in the rat. Pain 44:301–310. [DOI] [PubMed] [Google Scholar]
- Mishra SK, Tisel SM, Orestes P, Bhangoo SK, Hoon MA (2011) TRPV1-lineage neurons are required for thermal sensation. EMBO J 30:582–593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moller AT, Feldt-Rasmussen U, Rasmussen AK, Sommer C, Hasholt L, Bach FW, Kolvraa S, Jensen TS (2006) Small-fibre neuropathy in female Fabry patients: reduced allodynia and skin blood flow after topical capsaicin. J Peripher Nerv Syst 11:119–125. [DOI] [PubMed] [Google Scholar]
- Mou J, Paillard F, Turnbull B, Trudeau J, Stoker M, Katz NP (2013) Efficacy of Qutenza(R) (capsaicin) 8% patch for neuropathic pain: a meta-analysis of the Qutenza Clinical Trials Database. Pain 154:1632–1639. [DOI] [PubMed] [Google Scholar]
- Mou J, Paillard F, Turnbull B, Trudeau J, Stoker M, Katz NP (2014) Qutenza (capsaicin) 8% patch onset and duration of response and effects of multiple treatments in neuropathic pain patients. Clin J Pain 30:286–294. [DOI] [PubMed] [Google Scholar]
- Murthy SE, Loud MC, Daou I, Marshall KL, Schwaller F, Kuhnemund J, Francisco AG, Keenan WT, Dubin AE, Lewin GR, Patapoutian A (2018) The mechanosensitive ion channel Piezo2 mediates sensitivity to mechanical pain in mice. Sci Transl Med 10:eaat9897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakao A, Takahashi Y, Nagase M, Ikeda R, Kato F (2012) Role of capsaicin-sensitive C-fiber afferents in neuropathic pain-induced synaptic potentiation in the nociceptive amygdala. Mol Pain 8:51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narayanaswamy H, Facer P, Misra VP, Timmers M, Byttebier G, Meert T, Anand P (2012) A longitudinal study of sensory biomarkers of progression in patients with diabetic peripheral neuropathy using skin biopsies. J Clin Neurosci 19:1490–1496. [DOI] [PubMed] [Google Scholar]
- Nelson AE, Allen KD, Golightly YM, Goode AP, Jordan JM (2014) A systematic review of recommendations and guidelines for the management of osteoarthritis: The chronic osteoarthritis management initiative of the U.S. bone and joint initiative. Semin Arthritis Rheum 43:701–712. [DOI] [PubMed] [Google Scholar]
- Neubert JK, Karakunnel JJ, Brahim J, dionne RA (2017) Analgesic Effects of Vanilloid Receptor Desensitization by Capsaicin in the Oral Surgery Model. Applied Clinical Pharmacology and Toxicology ACPT-101. [Google Scholar]
- Neubert JK, Karai L, Jun JH, Kim HS, Olah Z, Iadarola MJ (2003) Peripherally induced resiniferatoxin analgesia. Pain 104:219–228. [DOI] [PubMed] [Google Scholar]
- Neubert JK, Mannes AJ, Karai LJ, Jenkins AC, Zawatski L, Abu-Asab M, Iadarola MJ (2008) Perineural resiniferatoxin selectively inhibits inflammatory hyperalgesia. Mol Pain 4:3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nolano M, Simone DA, Wendelschafer-Crabb G, Johnson T, Hazen E, Kennedy WR (1999) Topical capsaicin in humans: parallel loss of epidermal nerve fibers and pain sensation. Pain 81:135–145. [DOI] [PubMed] [Google Scholar]
- Numazaki M, Tominaga T, Takeuchi K, Murayama N, Toyooka H, Tominaga M (2003) Structural determinant of TRPV1 desensitization interacts with calmodulin. Proc Natl Acad Sci U S A 100:8002–8006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Brien ET, Salmon ED, Erickson HP (1997) How calcium causes microtubule depolymerization. Cell Motil Cytoskeleton 36:125–135. [DOI] [PubMed] [Google Scholar]
- O’Neill J, Brock C, Olesen AE, Andresen T, Nilsson M, Dickenson AH (2012) Unravelling the mystery of capsaicin: a tool to understand and treat pain. Pharmacol Rev 64:939–971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obata K, Yamanaka H, Dai Y, Mizushima T, Fukuoka T, Tokunaga A, Yoshikawa H, Noguchi K (2004) Contribution of degeneration of motor and sensory fibers to pain behavior and the changes in neurotrophic factors in rat dorsal root ganglion. Exp Neurol 188:149–160. [DOI] [PubMed] [Google Scholar]
- Ochoa JL, Campero M, Serra J, Bostock H (2005) Hyperexcitable polymodal and insensitive nociceptors in painful human neuropathy. Muscle Nerve 32:459–472. [DOI] [PubMed] [Google Scholar]
- Okamoto N, Okumura M, Tadokoro O, Sogawa N, Tomida M, Kondo E (2018) Effect of single-nucleotide polymorphisms in TRPV1 on burning pain and capsaicin sensitivity in Japanese adults. Mol Pain 14:1744806918804439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Onizuka S, Yonaha T, Tamura R, Hosokawa N, Kawasaki Y, Kashiwada M, Shirasaka T, Tsuneyoshi I (2011) Capsaicin indirectly suppresses voltage-gated Na+ currents through TRPV1 in rat dorsal root ganglion neurons. Anesth Analg 112:703–709. [DOI] [PubMed] [Google Scholar]
- Ono K, Ye Y, Viet CT, Dang D, Schmidt BL (2015) TRPV1 expression level in isolectin B(4)-positive neurons contributes to mouse strain difference in cutaneous thermal nociceptive sensitivity. J Neurophysiol 113:3345–3355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ossipov MH, Bian D, Malan TP Jr., Lai J, Porreca F (1999) Lack of involvement of capsaicin-sensitive primary afferents in nerve-ligation injury induced tactile allodynia in rats. Pain 79:127–133. [DOI] [PubMed] [Google Scholar]
- Pan HL, Khan GM, Alloway KD, Chen SR (2003) Resiniferatoxin induces paradoxical changes in thermal and mechanical sensitivities in rats: mechanism of action. J Neurosci 23:2911–2919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pang Z, Sakamoto T, Tiwari V, Kim YS, Yang F, Dong X, Guler AD, Guan Y, Caterina MJ (2015) Selective keratinocyte stimulation is sufficient to evoke nociception in mice. Pain 156:656–665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papagianni A, Siedler G, Sommer C, Uceyler N (2018) Capsaicin 8% patch reversibly reduces A-delta fiber evoked potential amplitudes. Pain Rep 3:e644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park JY, Jang SY, Shin YK, Koh H, Suh DJ, Shinji T, Araki T, Park HT (2013) Mitochondrial swelling and microtubule depolymerization are associated with energy depletion in axon degeneration. Neuroscience 238:258–269. [DOI] [PubMed] [Google Scholar]
- Payrits M, Saghy E, Cseko K, Pohoczky K, Bolcskei K, Ernszt D, Barabas K, Szolcsanyi J, Abraham IM, Helyes Z, Szoke E (2017) Estradiol Sensitizes the Transient Receptor Potential Vanilloid 1 Receptor in Pain Responses. Endocrinology 158:3249–3258. [DOI] [PubMed] [Google Scholar]
- Pecze L, Szabo K, Szell M, Josvay K, Kaszas K, Kusz E, Letoha T, Prorok J, Koncz I, Toth A, Kemeny L, Vizler C, Olah Z (2008) Human keratinocytes are vanilloid resistant. PLoS One 3:e3419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peleshok JC, Ribeiro-da-Silva A (2011) Delayed reinnervation by nonpeptidergic nociceptive afferents of the glabrous skin of the rat hindpaw in a neuropathic pain model. J Comp Neurol 519:49–63. [DOI] [PubMed] [Google Scholar]
- Petho G, Bolcskei K, Furedi R, Botz B, Bagoly T, Pinter E, Szolcsanyi J (2017) Evidence for a novel, neurohumoral antinociceptive mechanism mediated by peripheral capsaicin-sensitive nociceptors in conscious rats. Neuropeptides 62:1–10. [DOI] [PubMed] [Google Scholar]
- Phelps CE, Navratilova E, Dickenson AH, Porreca F, Bannister K (2019) Kappa opioid signaling in the right central amygdala causes hind paw specific loss of diffuse noxious inhibitory controls in experimental neuropathic pain. Pain 160:1614–1621. [DOI] [PubMed] [Google Scholar]
- Pickering G, Martin E, Tiberghien F, Delorme C, Mick G (2017) Localized neuropathic pain: an expert consensus on local treatments. Drug Des Devel Ther 11:2709–2718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinto LG, Souza GR, Kusuda R, Lopes AH, Sant’Anna MB, Cunha FQ, Ferreira SH, Cunha TM (2019) Non-Peptidergic Nociceptive Neurons Are Essential for Mechanical Inflammatory Hypersensitivity in Mice. Mol Neurobiol 56:5715–5728. [DOI] [PubMed] [Google Scholar]
- Poitras T, Chandrasekhar A, McCoy L, Komirishetty P, Areti A, Webber CA, Zochodne DW (2019) Selective Sensory Axon Reinnervation and TRPV1 Activation. Mol Neurobiol 56:7144–7158. [DOI] [PubMed] [Google Scholar]
- Polydefkis M, Hauer P, Sheth S, Sirdofsky M, Griffin JW, McArthur JC (2004) The time course of epidermal nerve fibre regeneration: studies in normal controls and in people with diabetes, with and without neuropathy. Brain 127:1606–1615. [DOI] [PubMed] [Google Scholar]
- Pomonis JD, Harrison JE, Mark L, Bristol DR, Valenzano KJ, Walker K (2003) N-(4-Tertiarybutylphenyl)-4-(3-cholorphyridin-2-yl)tetrahydropyrazine -1(2H)-carbox-amide (BCTC), a novel, orally effective vanilloid receptor 1 antagonist with analgesic properties: II. in vivo characterization in rat models of inflammatory and neuropathic pain. J Pharmacol Exp Ther 306:387–393. [DOI] [PubMed] [Google Scholar]
- Pospisilova E, Palecek J (2006) Post-operative pain behavior in rats is reduced after single high-concentration capsaicin application. Pain 125:233–243. [DOI] [PubMed] [Google Scholar]
- Prato V, Taberner FJ, Hockley JRF, Callejo G, Arcourt A, Tazir B, Hammer L, Schad P, Heppenstall PA, Smith ES, Lechner SG (2017) Functional and Molecular Characterization of Mechanoinsensitive “Silent” Nociceptors. Cell Rep 21:3102–3115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Privitera R, Birch R, Sinisi M, Mihaylov IR, Leech R, Anand P (2017) Capsaicin 8% patch treatment for amputation stump and phantom limb pain: a clinical and functional MRI study. J Pain Res 10:1623–1634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rage M, Van Acker N, Facer P, Shenoy R, Knaapen MW, Timmers M, Streffer J, Anand P, Meert T, Plaghki L (2010) The time course of CO2 laser-evoked responses and of skin nerve fibre markers after topical capsaicin in human volunteers. Clin Neurophysiol 121:1256–1266. [DOI] [PubMed] [Google Scholar]
- Ramnarine S, Laird B, Williams LJ, Fallon MT (2016) Investigating high-concentration 8% capsaicin patch in chronic cancer-treatment related peripheral neuropathic pain. Journal of Clinical Oncology 34:208.26628476 [Google Scholar]
- Rashid MH, Inoue M, Kondo S, Kawashima T, Bakoshi S, Ueda H (2003) Novel expression of vanilloid receptor 1 on capsaicin-insensitive fibers accounts for the analgesic effect of capsaicin cream in neuropathic pain. J Pharmacol Exp Ther 304:940–948. [DOI] [PubMed] [Google Scholar]
- Reilly CA, Ehlhardt WJ, Jackson DA, Kulanthaivel P, Mutlib AE, Espina RJ, Moody DE, Crouch DJ, Yost GS (2003) Metabolism of capsaicin by cytochrome P450 produces novel dehydrogenated metabolites and decreases cytotoxicity to lung and liver cells. Chem Res Toxicol 16:336–349. [DOI] [PubMed] [Google Scholar]
- Sajic M, Mastrolia V, Lee CY, Trigo D, Sadeghian M, Mosley AJ, Gregson NA, Duchen MR, Smith KJ (2013) Impulse conduction increases mitochondrial transport in adult mammalian peripheral nerves in vivo. PLoS Biol 11:e1001754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salas MM, Clifford JL, Hayden JR, Iadarola MJ, Averitt DL (2017) Local Resiniferatoxin Induces Long-Lasting Analgesia in a Rat Model of Full Thickness Thermal Injury. Pain Med 18:2453–2465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saper JR, Klapper J, Mathew NT, Rapoport A, Phillips SB, Bernstein JE (2002) Intranasal civamide for the treatment of episodic cluster headaches. Arch Neurol 59:990–994. [DOI] [PubMed] [Google Scholar]
- Sapio MR, Neubert JK, LaPaglia DM, Maric D, Keller JM, Raithel SJ, Rohrs EL, Anderson EM, Butman JA, Caudle RM, Brown DC, Heiss JD, Mannes AJ, Iadarola MJ (2018) Pain control through selective chemo-axotomy of centrally projecting TRPV1+ sensory neurons. J Clin Invest 128:1657–1670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sauerstein K, Liebelt J, Namer B, Schmidt R, Rukwied R, Schmelz M (2018) Low-Frequency Stimulation of Silent Nociceptors Induces Secondary Mechanical Hyperalgesia in Human Skin. Neuroscience 387:4–12. [DOI] [PubMed] [Google Scholar]
- Schafers M, Lee DH, Brors D, Yaksh TL, Sorkin LS (2003) Increased sensitivity of injured and adjacent uninjured rat primary sensory neurons to exogenous tumor necrosis factor-alpha after spinal nerve ligation. J Neurosci 23:3028–3038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schicho R, Schuligoi R, Sirinathsinghji DJ, Donnerer J (1999) Increased expression of GAP-43 in small sensory neurons after stimulation by NGF indicative of neuroregeneration in capsaicin-treated rats. Regul Pept 83:87–95. [DOI] [PubMed] [Google Scholar]
- Schmelz M, Schmid R, Handwerker HO, Torebjork HE (2000a) Encoding of burning pain from capsaicin-treated human skin in two categories of unmyelinated nerve fibres. Brain 123 Pt 3:560–571. [DOI] [PubMed] [Google Scholar]
- Schmelz M, Michael K, Weidner C, Schmidt R, Torebjork HE, Handwerker HO (2000b) Which nerve fibers mediate the axon reflex flare in human skin? Neuroreport 11:645–648. [DOI] [PubMed] [Google Scholar]
- Schumacher MA (2010) Transient receptor potential channels in pain and inflammation: therapeutic opportunities. Pain Pract 10:185–200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serra J, Bostock H, Sola R, Aleu J, Garcia E, Cokic B, Navarro X, Quiles C (2012) Microneurographic identification of spontaneous activity in C-nociceptors in neuropathic pain states in humans and rats. Pain 153:42–55. [DOI] [PubMed] [Google Scholar]
- Serrano A, Torres D, Veciana M, Caro C, Montero J, Mayoral V (2017) Quantitative Thermal Testing Profiles as a Predictor of Treatment Response to Topical Capsaicin in Patients with Localized Neuropathic Pain. Pain Res Treat 2017:7425907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shin CY, Shin J, Kim BM, Wang MH, Jang JH, Surh YJ, Oh U (2003) Essential role of mitochondrial permeability transition in vanilloid receptor 1-dependent cell death of sensory neurons. Mol Cell Neurosci 24:57–68. [DOI] [PubMed] [Google Scholar]
- Simone DA, Ochoa J (1991) Early and late effects of prolonged topical capsaicin on cutaneous sensibility and neurogenic vasodilatation in humans. Pain 47:285–294. [DOI] [PubMed] [Google Scholar]
- Simone DA, Nolano M, Johnson T, Wendelschafer-Crabb G, Kennedy WR (1998) Intradermal injection of capsaicin in humans produces degeneration and subsequent reinnervation of epidermal nerve fibers: correlation with sensory function. J Neurosci 18:8947–8959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson DM, Gazda S, Brown S, Webster LR, Lu SP, Tobias JK, Vanhove GF, Group N-CS (2010) Long-term safety of NGX-4010, a high-concentration capsaicin patch, in patients with peripheral neuropathic pain. J Pain Symptom Manage 39:1053–1064. [DOI] [PubMed] [Google Scholar]
- Simpson DM, Robinson-Papp J, Van J, Stoker M, Jacobs H, Snijder RJ, Schregardus DS, Long SK, Lambourg B, Katz N (2017) Capsaicin 8% Patch in Painful Diabetic Peripheral Neuropathy: A Randomized, Double-Blind, Placebo-Controlled Study. J Pain 18:42–53. [DOI] [PubMed] [Google Scholar]
- Sluka KA, Willis WD (1997) The effects of G-protein and protein kinase inhibitors on the behavioral responses of rats to intradermal injection of capsaicin. Pain 71:165–178. [DOI] [PubMed] [Google Scholar]
- Stevens RM, Ervin J, Nezzer J, Nieves Y, Guedes K, Burges R, Hanson PD, Campbell JN (2019) Randomized, Double-Blind, Placebo-Controlled Trial of Intraarticular Trans-Capsaicin for Pain Associated With Osteoarthritis of the Knee. Arthritis Rheumatol 71:1524–1533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su X, Wachtel RE, Gebhart GF (1999) Capsaicin sensitivity and voltage-gated sodium currents in colon sensory neurons from rat dorsal root ganglia. Am J Physiol 277:G1180–1188. [DOI] [PubMed] [Google Scholar]
- Such G, Jancso G (1986) Axonal effects of capsaicin: an electrophysiological study. Acta Physiol Hung 67:53–63. [PubMed] [Google Scholar]
- Szabo A, Helyes Z, Sandor K, Bite A, Pinter E, Nemeth J, Banvolgyi A, Bolcskei K, Elekes K, Szolcsanyi J (2005) Role of transient receptor potential vanilloid 1 receptors in adjuvant-induced chronic arthritis: in vivo study using gene-deficient mice. J Pharmacol Exp Ther 314:111–119. [DOI] [PubMed] [Google Scholar]
- Szallasi A, Blumberg PM (1999) Vanilloid (Capsaicin) receptors and mechanisms. Pharmacol Rev 51:159–212. [PubMed] [Google Scholar]
- Szallasi A, Sheta M (2012) Targeting TRPV1 for pain relief: limits, losers and laurels. Expert Opin Investig Drugs 21:1351–1369. [DOI] [PubMed] [Google Scholar]
- Szczot M, Liljencrantz J, Ghitani N, Barik A, Lam R, Thompson JH, Bharucha-Goebel D, Saade D, Necaise A, Donkervoort S, Foley AR, Gordon T, Case L, Bushnell MC, Bonnemann CG, Chesler AT (2018) PIEZO2 mediates injury-induced tactile pain in mice and humans. Sci Transl Med 10:eaat9892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szolcsanyi J, Pinter E (2013) Transient receptor potential vanilloid 1 as a therapeutic target in analgesia. Expert Opin Ther Targets 17:641–657. [DOI] [PubMed] [Google Scholar]
- Szolcsanyi J, Jancso-Gabor A, Joo F (1975) Functional and fine structural characteristics of the sensory neuron blocking effect of capsaicin. Naunyn Schmiedebergs Arch Pharmacol 287:157–169. [DOI] [PubMed] [Google Scholar]
- Szolcsanyi J, Helyes Z, Oroszi G, Nemeth J, Pinter E (1998) Release of somatostatin and its role in the mediation of the anti-inflammatory effect induced by antidromic stimulation of sensory fibres of rat sciatic nerve. Br J Pharmacol 123:936–942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor AM, Osikowicz M, Ribeiro-da-Silva A (2012) Consequences of the ablation of nonpeptidergic afferents in an animal model of trigeminal neuropathic pain. Pain 153:1311–1319. [DOI] [PubMed] [Google Scholar]
- Tender GC, Walbridge S, Olah Z, Karai L, ladarola M, Oldfield EH, Lonser RR (2005) Selective ablation of nociceptive neurons for elimination of hyperalgesia and neurogenic inflammation. J Neurosurg 102:522–525. [DOI] [PubMed] [Google Scholar]
- Tenreiro Pinto J, Pereira FC, Loureiro MC, Gama R, Fernandes HL (2018) Efficacy Analysis of Capsaicin 8% Patch in Neuropathic Peripheral Pain Treatment. Pharmacology 101:290–297. [DOI] [PubMed] [Google Scholar]
- Terenzi R, Romano E, Manetti M, Peruzzi F, Nacci F, Matucci-Cerinic M, Guiducci S (2013) Neuropeptides activate TRPV1 in rheumatoid arthritis fibroblast-like synoviocytes and foster IL-6 and IL-8 production. Ann Rheum Dis 72:1107–1109. [DOI] [PubMed] [Google Scholar]
- Tobaldini G, Sardi NF, Guilhen VA, Fischer L (2019) Pain Inhibits Pain: an Ascending-Descending Pain Modulation Pathway Linking Mesolimbic and Classical Descending Mechanisms. Mol Neurobiol 56:1000–1013. [DOI] [PubMed] [Google Scholar]
- Treede RD, Meyer RA, Raja SN, Campbell JN (1992) Peripheral and central mechanisms of cutaneous hyperalgesia. Prog Neurobiol 38:397–421. [DOI] [PubMed] [Google Scholar]
- Treede RD et al. (2013) Mechanism- and experience-based strategies to optimize treatment response to the capsaicin 8% cutaneous patch in patients with localized neuropathic pain. Curr Med Res Opin 29:527–538. [DOI] [PubMed] [Google Scholar]
- Trouvin AP, Perrot S (2019) Functional and histological improvements of small nerve neuropathy after high-concentration capsaicin patch application: A case study. Pain Rep 4:e761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuboi Y, Honda K, Bae YC, Shinoda M, Kondo M, Katagiri A, Echizenya S, Kamakura S, Lee J, Iwata K (2015) Morphological and functional changes in regenerated primary afferent fibres following mental and inferior alveolar nerve transection. Eur J Pain 19:1258–1266. [DOI] [PubMed] [Google Scholar]
- Turnbull A (1850) Tincture of capsaicin as a remedy for chilblains and toothache. Dublin Free Press 1:95–96. [Google Scholar]
- Uceyler N, Sommer C (2014) High-Dose Capsaicin for the Treatment of Neuropathic Pain: What We Know and What We Need to Know. Pain Ther 3:73–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uhelski ML, McAdams B, Johns ME, Kabadi RA, Simone DA, Banik RK (2020) Lack of relationship between epidermal denervation by capsaicin and incisional pain behaviours: A laser scanning confocal microscopy study in rats. Eur J Pain 24:1197–1208. [DOI] [PubMed] [Google Scholar]
- Unger MD, Pleticha J, Steinauer J, Kanwar R, Diehn F, LaVallee KT, Banck MS, Jones B, Yaksh TL, Maus TP, Beutler AS (2019) Unilateral Epidural Targeting of Resiniferatoxin Induces Bilateral Neurolysis of Spinal Nociceptive Afferents. Pain Med 20:897–906. [DOI] [PubMed] [Google Scholar]
- Urano H, Ara T, Fujinami Y, Hiraoka BY (2012) Aberrant TRPV1 expression in heat hyperalgesia associated with trigeminal neuropathic pain. Int J Med Sci 9:690–697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ursu D, Knopp K, Beattie RE, Liu B, Sher E (2010) Pungency of TRPV1 agonists is directly correlated with kinetics of receptor activation and lipophilicity. Eur J Pharmacol 641:114–122. [DOI] [PubMed] [Google Scholar]
- Usoskin D, Furlan A, Islam S, Abdo H, Lonnerberg P, Lou D, Hjerling-Leffler J, Haeggstrom J, Kharchenko O, Kharchenko PV, Linnarsson S, Ernfors P (2015) Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat Neurosci 18:145–153. [DOI] [PubMed] [Google Scholar]
- Valdes AM et al. (2011) The Ile585Val TRPV1 variant is involved in risk of painful knee osteoarthritis. Ann Rheum Dis 70:1556–1561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Hecke O, Austin SK, Khan RA, Smith BH, Torrance N (2014) Neuropathic pain in the general population: a systematic review of epidemiological studies. Pain 155:654–662. [DOI] [PubMed] [Google Scholar]
- van Nooten F, Treur M, Pantiri K, Stoker M, Charokopou M (2017) Capsaicin 8% Patch Versus Oral Neuropathic Pain Medications for the Treatment of Painful Diabetic Peripheral Neuropathy: A Systematic Literature Review and Network Meta-analysis. Clin Ther 39:787–803 e718. [DOI] [PubMed] [Google Scholar]
- Vinik AI, Perrot S, Vinik EJ, Pazdera L, Jacobs H, Stoker M, Long SK, Snijder RJ, van der Stoep M, Ortega E, Katz N (2016) Capsaicin 8% patch repeat treatment plus standard of care (SOC) versus SOC alone in painful diabetic peripheral neuropathy: a randomised, 52-week, open-label, safety study. BMC Neurol 16:251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vos MH, Neelands TR, McDonald HA, Choi W, Kroeger PE, Puttfarcken PS, Faltynek CR, Moreland RB, Han P (2006) TRPV1b overexpression negatively regulates TRPV1 responsiveness to capsaicin, heat and low pH in HEK293 cells. J Neurochem 99:1088–1102. [DOI] [PubMed] [Google Scholar]
- Vyklicky L, Novakova-Tousova K, Benedikt J, Samad A, Touska F, Vlachova V (2008) Calcium-dependent desensitization of vanilloid receptor TRPV1: a mechanism possibly involved in analgesia induced by topical application of capsaicin. Physiol Res 57 Suppl 3:S59–68. [DOI] [PubMed] [Google Scholar]
- Wallace M, Duan R, Liu W, Locke C, Nothaft W (2016) A Randomized, Double-Blind, Placebo-Controlled, Crossover Study of the T-Type Calcium Channel Blocker ABT-639 in an Intradermal Capsaicin Experimental Pain Model in Healthy Adults. Pain Med 17:551–560. [DOI] [PubMed] [Google Scholar]
- Wallengren J, Hakanson R (1992) Effects of capsaicin, bradykinin and prostaglandin E2 in the human skin. Br J Dermatol 126:111–117. [DOI] [PubMed] [Google Scholar]
- Wang H, Bolognese J, Calder N, Baxendale J, Kehler A, Cummings C, Connell J, Herman G (2008) Effect of morphine and pregabalin compared with diphenhydramine hydrochloride and placebo on hyperalgesia and allodynia induced by intradermal capsaicin in healthy male subjects. J Pain 9:1088–1095. [DOI] [PubMed] [Google Scholar]
- Wang MS, Davis AA, Culver DG, Wang Q, Powers JC, Glass JD (2004) Calpain inhibition protects against Taxol-induced sensory neuropathy. Brain 127:671–679. [DOI] [PubMed] [Google Scholar]
- Wang S, Joseph J, Diatchenko L, Ro JY, Chung MK (2016) Agonist-dependence of functional properties for common nonsynonymous variants of human transient receptor potential vanilloid 1. Pain 157:1515–1524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang S, Brigoli B, Lim J, Karley A, Chung MK (2018a) Roles of TRPV1 and TRPA1 in Spontaneous Pain from Inflamed Masseter Muscle. Neuroscience 384:290–299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang S, Lim J, Joseph J, Wei F, Ro JY, Chung MK (2017a) Spontaneous and Bite-Evoked Muscle Pain Are Mediated by a Common Nociceptive Pathway With Differential Contribution by TRPV1. J Pain 18:1333–1345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang S, Kim M, Ali Z, Ong K, Pae EK, Chung MK (2019) TRPV1 and TRPV1-Expressing Nociceptors Mediate Orofacial Pain Behaviors in a Mouse Model of Orthodontic Tooth Movement. Front Physiol 10:1207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang S, Asgar J, Joseph J, Ro JY, Wei F, Campbell JN, Chung MK (2017b) Ca2+ and Calpain Mediate Capsaicin-induced Ablation of Axonal Terminals Expressing Transient Receptor Potential Vanilloid 1. J Biol Chem 292:8291–8303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang S, Bian C, Yang J, Arora V, Gao Y, Wei F, Chung MK (2020) Ablation of TRPV1+ afferent terminals by capsaicin mediates long-lasting analgesia for trigeminal neuropathic pain. eNeuro 7:0118–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang SE, Ko SY, Kim YS, Jo S, Lee SH, Jung SJ, Son H (2018b) Capsaicin upregulates HDAC2 via TRPV1 and impairs neuronal maturation in mice. Exp Mol Med 50:e455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watabiki T, Kiso T, Kuramochi T, Yonezawa K, Tsuji N, Kohara A, Kakimoto S, Aoki T, Matsuoka N (2011) Amelioration of neuropathic pain by novel transient receptor potential vanilloid 1 antagonist AS1928370 in rats without hyperthermic effect. J Pharmacol Exp Ther 336:743–750. [DOI] [PubMed] [Google Scholar]
- Watson CP (1994) Topical capsaicin as an adjuvant analgesic. J Pain Symptom Manage 9:425–433. [DOI] [PubMed] [Google Scholar]
- Watson CP, Tyler KL, Bickers DR, Millikan LE, Smith S, Coleman E (1993) A randomized vehicle-controlled trial of topical capsaicin in the treatment of postherpetic neuralgia. Clin Ther 15:510–526. [PubMed] [Google Scholar]
- Webster LR, Tark M, Rauck R, Tobias JK, Vanhove GF (2010a) Effect of duration of postherpetic neuralgia on efficacy analyses in a multicenter, randomized, controlled study of NGX-4010, an 8% capsaicin patch evaluated for the treatment of postherpetic neuralgia. BMC Neurol 10:92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webster LR, Malan TP, Tuchman MM, Mollen MD, Tobias JK, Vanhove GF (2010b) A multicenter, randomized, double-blind, controlled dose finding study of NGX-4010, a high-concentration capsaicin patch, for the treatment of postherpetic neuralgia. J Pain 11:972–982. [DOI] [PubMed] [Google Scholar]
- Westlund KN, Kochukov MY, Lu Y, McNearney TA (2010) Impact of central and peripheral TRPV1 and ROS levels on proinflammatory mediators and nociceptive behavior. Mol Pain 6:46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu G, Ringkamp M, Murinson BB, Pogatzki EM, Hartke TV, Weerahandi HM, Campbell JN, Griffin JW, Meyer RA (2002) Degeneration of myelinated efferent fibers induces spontaneous activity in uninjured C-fiber afferents. J Neurosci 22:7746–7753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu ZZ, Chen SR, Pan HL (2005) Transient receptor potential vanilloid type 1 activation down-regulates voltage-gated calcium channels through calcium-dependent calcineurin in sensory neurons. J Biol Chem 280:18142–18151. [DOI] [PubMed] [Google Scholar]
- Wu ZZ, Chen SR, Pan HL (2006) Signaling mechanisms of down-regulation of voltage-activated Ca2+ channels by transient receptor potential vanilloid type 1 stimulation with olvanil in primary sensory neurons. Neuroscience 141:407–419. [DOI] [PubMed] [Google Scholar]
- Yaksh TL, Farb DH, Leeman SE, Jessell TM (1979) Intrathecal capsaicin depletes substance P in the rat spinal cord and produces prolonged thermal analgesia. Science 206:481–483. [DOI] [PubMed] [Google Scholar]
- Yang J, Weimer RM, Kallop D, Olsen O, Wu Z, Renier N, Uryu K, Tessier-Lavigne M (2013) Regulation of axon degeneration after injury and in development by the endogenous calpain inhibitor calpastatin. Neuron 80:1175–1189. [DOI] [PubMed] [Google Scholar]
- Yee JR, Kenkel W, Caccaviello JC, Gamber K, Simmons P, Nedelman M, Kulkarni P, Ferris CF (2015) Identifying the integrated neural networks involved in capsaicin-induced pain using fMRI in awake TRPV1 knockout and wild-type rats. Front Syst Neurosci 9:15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoneda S, Kasai E, Matsuo M, Tamano R, Sakurai Y, Asaki T, Fujita M (2020) Duloxetine ameliorates the impairment of diffuse noxious inhibitory control in rat models of peripheral neuropathic pain and knee osteoarthritis pain. Neurosci Lett 729:134990. [DOI] [PubMed] [Google Scholar]
- Yu S, Premkumar LS (2015) Ablation and Regeneration of Peripheral and Central TRPV1 Expressing Nerve Terminals and the Consequence of Nociception. Open Pain J 8:1–9. [Google Scholar]
- Zang Y, Chen SX, Liao GJ, Zhu HQ, Wei XH, Cui Y, Na XD, Pang RP, Xin WJ, Zhou LJ, Liu XG (2015) Calpain-2 contributes to neuropathic pain following motor nerve injury via up-regulating interleukin-6 in DRG neurons. Brain Behav Immun 44:37–47. [DOI] [PubMed] [Google Scholar]
- Zhai K, Liskova A, Kubatka P, Busselberg D (2020) Calcium Entry through TRPV1: A Potential Target for the Regulation of Proliferation and Apoptosis in Cancerous and Healthy Cells. Int J Mol Sci 21:4177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang K, Ramamurthy S, Prihoda TJ, Eckmann MS (2014) Effect of delayed intrathecal administration of capsaicin on neuropathic pain induced by chronic constriction injury of the sciatic nerve in rats. J Pain Res 7:547–554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmermann K, Leffler A, Babes A, Cendan CM, Carr RW, Kobayashi J, Nau C, Wood JN, Reeh PW (2007) Sensory neuron sodium channel Nav1.8 is essential for pain at low temperatures. Nature 447:855–858. [DOI] [PubMed] [Google Scholar]