

Abstract
The interleukins (ILs) are a pluripotent cytokine family that have been reported to regulate ischemic stroke and cerebral ischemia/reperfusion (I/R) injury. IL-22 is a member of the IL-10 superfamily and plays important roles in tissue injury and repair. However, the effects of IL-22 on ischemic stroke and cerebral I/R injury remain unclear. In the current study, we provided direct evidence that IL-22 treatment decreased infarct size, neurological deficits, and brain water content in mice subjected to cerebral I/R injury. IL-22 treatment remarkably reduced the expression of inflammatory cytokines, including IL-1β, monocyte chemotactic protein- (MCP-) 1, and tumor necrosis factor- (TNF-) α, both in serum and the ischemic cerebral cortex. In addition, IL-22 treatment also decreased oxidative stress and neuronal apoptosis in mice after cerebral I/R injury. Moreover, IL-22 treatment significantly increased Janus tyrosine kinase (JAK) 2 and signal transducer and activator of transcription (STAT) 3 phosphorylation levels in mice and PC12 cells, and STAT3 knockdown abolished the IL-22-mediated neuroprotective function. These findings suggest that IL-22 might be exploited as a potential therapeutic agent for ischemic stroke and cerebral I/R injury.
1. Introduction
A report from the Global Burden of Disease (GBD) 2016 Stroke Collaborators showed that although the prevalence and mortality of stroke have decreased in the past 20 years, stroke remains the second leading cause of death and long-term disability worldwide [1, 2]. Among them, ischemic stroke is the most common type and occurs when cerebral arteries are occluded [3, 4]. Currently, restoring blood perfusion is an approved therapy for cerebral ischemic injury, including intravenous thrombolytic and endovascular therapy [5–8]. However, the degree of brain injury may be further aggravated following the reperfusion process, which is called cerebral ischemia/reperfusion (I/R) injury [9, 10]. Increasing evidence has shown that cerebral I/R can cause secondary brain injury, including cerebral hemorrhage, cerebral edema, and even death [11, 12]. Thus, it is necessary to clarify the pathological mechanism underlying cerebral I/R injury and explore novel therapeutic agents for ischemic stroke and cerebral I/R injury.
The interleukins (ILs) are a pluripotent cytokine family that have been reported to regulate ischemic stroke and cerebral I/R injury [13, 14]. In clinical experiments, higher IL-33 levels in acute ischemic stroke patients were positively correlated with better prognosis and could be used to predict outcomes and recurrences in acute ischemic stroke patients [15]. In addition, an IL-1 receptor antagonist (IL-1Ra) significantly decreased plasma concentrations of IL-6 and C-reactive protein in patients with ischemic stroke, indicating that IL-1Ra can improve clinical outcomes by reducing inflammation [16]. IL-35 pretreatment significantly reduced brain infarction and neurological deficits after cerebral I/R injury [17]. In addition, inhibition of IL-32 significantly reduced the infarct volume and neurological deficits following cerebral I/R injury by suppressing proinflammatory cytokine secretion [18].
IL-22 is a member of the IL-10 superfamily and plays important roles in tissue injury and repair [19, 20]. IL-22 has been reported to participate in various biological processes, including the inflammatory response, oxidative stress, endoplasmic reticulum stress, autophagy, apoptosis, and cell death [21–24]. Takahashi et al. reported that IL-22 treatment ameliorated I/R-induced myocardial injury and apoptosis by activating the signal transducer and activator of transcription (STAT) 3 signaling pathway [25]. Xu et al. also reported that IL-22 treatment or IL-22 overexpression prevented renal injury and inflammation after renal I/R in mice [26]. However, it remains unclear whether IL-22 is involved in ischemic stroke and cerebral I/R injury. Thus, the aim of this study was to determine the roles of IL-22 in cerebral I/R injury and to explore the underlying mechanism.
2. Materials and Methods
2.1. Animals and Animal Model
Male C57BL/6J mice were purchased from Beijing HFK Biotechnology Co., Ltd. (Beijing, China). All mice were maintained in standard housing conditions under a 12 h light-dark cycle and were allowed free access to standard rodent food and water. Animal care and procedures were conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals and approved by the Animal Ethics Committee of Anhui Medical University.
A middle cerebral artery occlusion (MCAO) model was generated according to previous research [27]. After 45 min of ischemia, the suture was removed to initiate reperfusion. The sham-operated mice underwent the same procedures but did not receive sutures. Thirty minutes before reperfusion, the mice were injected intraperitoneally with recombinant mouse IL-22 protein (rIL-22). The doses and times were selected according to our pilot experiments and previous research. The 90 mice were randomly allocated into the following three groups (n = 30/group): sham group, MCAO group, and rIL-22 group.
2.2. Neurological Impairment Scores
After 24 h of reperfusion, neurological impairment was evaluated as previously described [28]. The neurological scoring system ranged from 0 (no neurological deficits) to 4 (inability to walk spontaneously).
2.3. Measurement of Infarct Area
After neurological evaluation, the brains were rapidly removed and subsequently cut into coronal sections and then incubated with 2,3,5-triphenyltetrazolium chloride (TTC) at 37°C for 20 min. The sections were fixed in 4% paraformaldehyde and photographed using an HD camera. The infarct area was analyzed using ImageJ, and the infarct volume was calculated as previously described [29].
2.4. Brain Water Content
After 24 h of reperfusion, the mice were sacrificed and the brain tissues were rapidly removed and then immediately weighed to obtain the wet weight. Subsequently, the brain tissues were dried in a desiccating oven at 105°C to obtain the dry weight. The brain water content was calculated according to the previous described [30].
2.5. Cell Culture and Treatment
PC12 cells procured from the Culture Collection of the Chinese Academy of Science (Shanghai, China) were cultured in DMEM containing 10% fetal bovine serum and 1% penicillin/streptomycin. Oxygen and glucose deprivation/reperfusion (OGD/R) was established by culturing the cells in glucose-free DMEM and hypoxic conditions with 95%N2/5%CO2. After 2 h of hypoxia, the cells were transferred back to full culture medium under normal atmosphere and incubated for 24 h. At 3h before OGD/R, rIL-22 (100 ng/mL) was administrated. To knockdown JAK2 and STAT3 expression, PC12 cells were transfected with si-JAK2 and si-STAT3, respectively, using Lipofectamine 2000 according to the manufacturer's recommendation.
2.6. ELISA
After 24 h of reperfusion, blood specimens were obtained from mice and centrifuged to separate the serum. The levels of IL-1β, monocyte chemotactic protein- (MCP-) 1, and tumor necrosis factor- (TNF-) α were measured by ELISA kits (R&D Systems, USA) according to the manufacturer's instructions.
2.7. Oxidative Stress Detection
After 24 h of reperfusion, the brain tissues were rapidly removed and prepared as homogenates, and then, the supernatants were collected. For the cells, PC12 cells were harvested and lysed, and then, the supernatants were collected. The activity of total superoxide dismutase (SOD) and glutathione (GSH) and the concentration of malondialdehyde (MDA) were measured by commercial assay kits (Beyotime Biotechnology, China) according to the manufacturer's instructions.
2.8. Apoptosis Assay
Cell apoptosis was measured by a terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) assay kit, as previously described [31]. Briefly, the slices were incubated with TUNEL reagents, and DAPI solution was prepared according to the manufacturer's instructions. The number and ratio of TUNEL-positive cells were calculated based on the apoptosis index evaluated by an investigator blinded to the experiment.
2.9. Quantitative Real-Time RT-PCR
Total RNA was extracted from the ischemic hemisphere and PC12 cells using a TRIzol reagent and then reverse transcribed into cDNA according to the manufacturer's protocol. Real-time PCR analysis was performed using a LightCycler 480 qPCR System. The relative expression of target genes was normalized against β-actin mRNA. The primer sequences are presented in Table 1.
Table 1.
Primer sequences for RT-PCR assays.
| Gene | Species | Sequence (5′-3′) | |
|---|---|---|---|
| IL-1β | Mouse | Forward | GGGCCTCAAAGGAAAGAATC |
| Reverse | TACCAGTTGGGGAACTCTGC | ||
| IL-1β | Rat | Forward | GTGCTGTCTGACCCATGTGA |
| Reverse | CACAGGGATTTTGTCGTTGCT | ||
| MCP-1 | Mouse | Forward | GAGGTCACTCCTATCCTCTGG |
| Reverse | GCCATTTCCTCCGACTTTTCTC | ||
| MCP-1 | Rat | Forward | AGCATCCACGTGCTGTCTC |
| Reverse | GATCATCTTGCCAGTGAATGAG | ||
| TNF-α | Mouse | Forward | CCCAGGGACCTCTCTCTAATC |
| Reverse | ATGGGCTACAGGCTTGTCACT | ||
| TNF-α | Rat | Forward | CTACTCCCAGGTTCTCTTCAA |
| Reverse | GCTGACTTTCTCCTGGTATGA | ||
| β-Actin | Mouse | Forward | TATTGGCAACGAGCGGTTCC |
| Reverse | GGCATAGAGGTCTTTACGGATGT | ||
| β-Actin | Rat | Forward | CAAGAAGGTGGTGAAGCAG |
| Reverse | AAAGGTGGAAGAATGGGAG | ||
2.10. Western Blotting
Protein was extracted from the ischemic hemisphere and PC12 cells and then separated using SDS-PAGE. The proteins were transferred onto an Immobilon-P membrane (Millipore, USA). The membranes were incubated with primary antibodies against Bax, Bcl-2, p-JAK2, JAK2, p-STAT3, STAT3, and β-actin, followed by incubation with the secondary antibody. Finally, proteins on the membranes were detected using an Odyssey infrared imaging system (LI-COR, USA), and the protein expression levels were normalized to that of β-actin.
2.11. Statistical Analysis
Statistical analyses were performed using SPSS software. Normally distributed data are expressed as the mean ± standard deviation (SD). One-way analysis of variance (ANOVA) was used for comparisons among multiple groups, and when the differences were statistically significant, a post hoc Tukey test was carried out. Nonnormally distributed data are expressed as the median and quartiles. The Kruskal-Wallis H test was used for comparisons among multiple groups, and when the differences were statistically significant, the Mann-Whitney U test was carried out followed by Bonferroni correction. The Bonferroni correction is α′ = 0.05/K, where K is the number of comparisons. P values less than 0.05 were considered statistically significant.
3. Results
3.1. IL-22 Treatment Ameliorated Cerebral I/R Injury
After ischemia-reperfusion, significant cerebral infarction was observed in the MCAO group, but IL-22 treatment significantly decreased the infarct volume of mice (Figures 1(a) and 1(b)). In addition, IL-22 administration significantly ameliorated neurological deficits and brain water content after cerebral I/R injury (Figures 1(c) and 1(d)).
Figure 1.
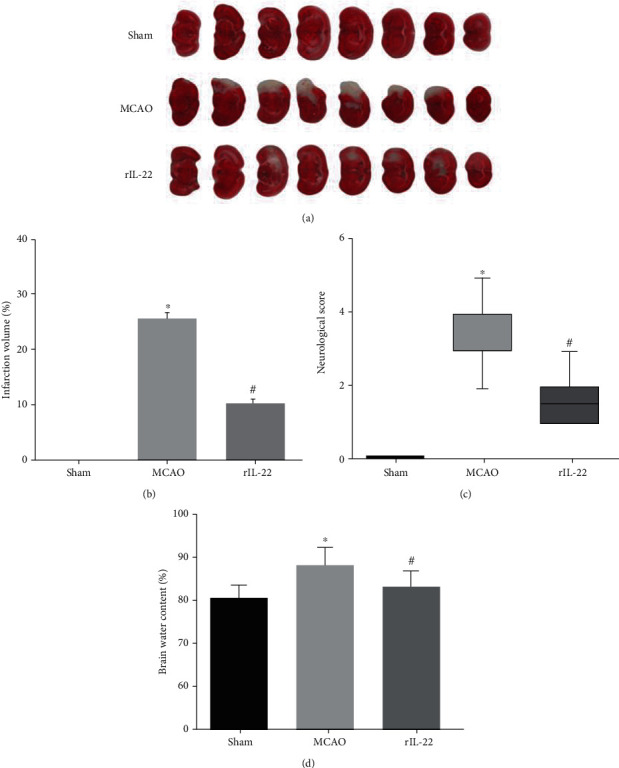
IL-22 treatment ameliorated cerebral I/R injury. (a) Representative sections of TTC staining in each group (n = 6). (b) Quantification of infarct volume in each group (n = 6). (c) Neurological deficits were assessed in each group (n = 8). (d) Brain water content was calculated in each group (n = 6). ∗P < 0.05 vs sham group; #P < 0.05 vs MCAO group.
3.2. IL-22 Treatment Inhibited the Inflammatory Response after Cerebral I/R Injury
Compared with the sham group, serum levels of inflammatory cytokines, including IL-1β, MCP-1, and TNF-α, in the MCAO group were significantly increased, while IL-22 treatment reduced the serum levels of these cytokines (Figures 2(a)–2(c)). Furthermore, IL-22 treatment also decreased the mRNA expression of IL-1β, MCP-1, and TNF-α in the ischemic cerebral cortex (Figures 2(d)–2(f)).
Figure 2.
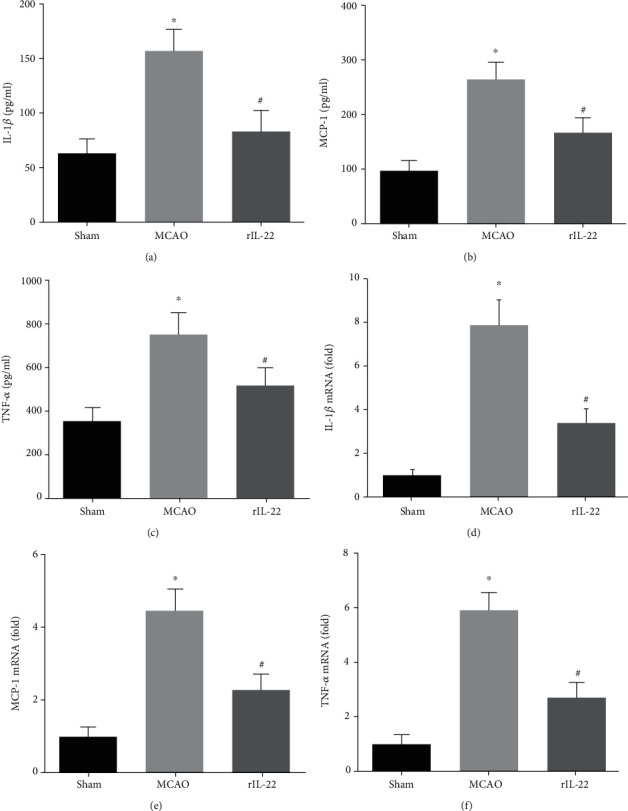
IL-22 treatment inhibited the inflammatory response after cerebral I/R injury. Serum levels of IL-1β (a), MCP-1 (b), and TNF-α (c) were measured by ELISA (n = 8). The mRNA expression of IL-1β (d), MCP-1 (e), and TNF-α (f) was detected in brain tissues (n = 8). ∗P < 0.05 vs. sham group; #P < 0.05 vs. MCAO group.
3.3. IL-22 Treatment Attenuated Oxidative Stress and Neuronal Apoptosis after Cerebral I/R Injury
Compared with the sham group, the activities of SOD and GSH in the MCAO group were significantly decreased and the levels of MDA were significantly increased after cerebral I/R injury (Figures 3(a)–3(c)). Interestingly, the activities of SOD and GSH in the brain tissues were significantly increased, and the levels of MDA were significantly reduced in the rIL-22 group compared with those in the MCAO group (Figures 3(a)–3(c)). The TUNEL staining results also showed that IL-22 treatment significantly decreased neuronal apoptosis after cerebral I/R (Figure 3(d)).
Figure 3.
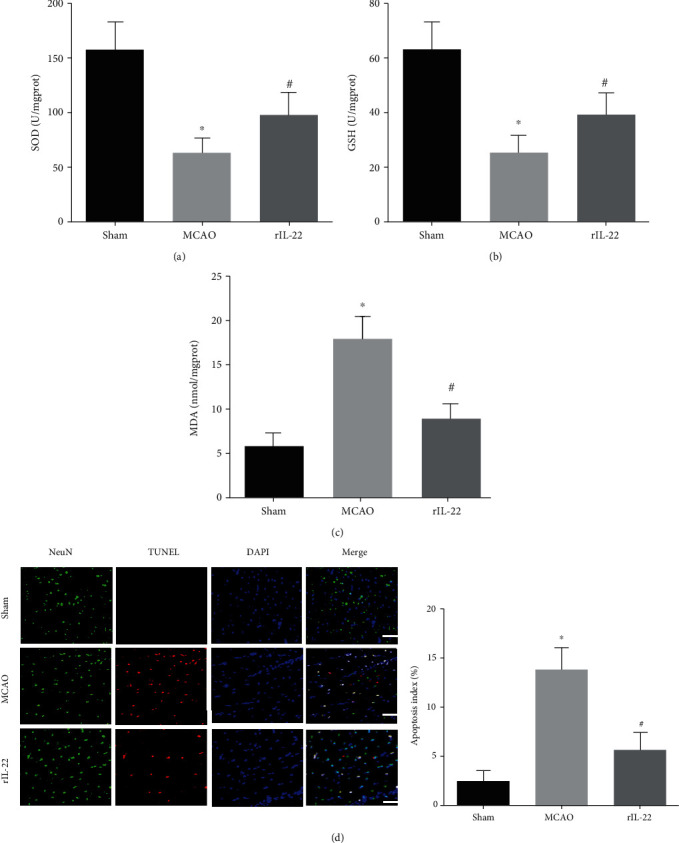
IL-22 treatment attenuated oxidative stress and neuronal apoptosis after cerebral I/R injury. The levels of SOD (a), GSH (b), and MDA (c) were detected in brain tissues (n = 6). (d) Neuronal apoptosis was detected by TUNEL staining combined with immunostaining for NeuN (n = 5, scale bar = 75 μm). ∗P < 0.05 vs. sham group; #P < 0.05 vs. MCAO group.
3.4. IL-22 Treatment Inhibited OGD/R-Induced Inflammation and Oxidative Stress
Our results showed that IL-22 treatment significantly decreased the mRNA expression of IL-1β, MCP-1, and TNF-α after OGD/R (Figures 4(a)–4(c)). In addition, IL-22 treatment significantly increased the activities of SOD and GSH and reduced the levels of MDA compared with the OGD/R group (Figures 4(d)–4(f)).
Figure 4.
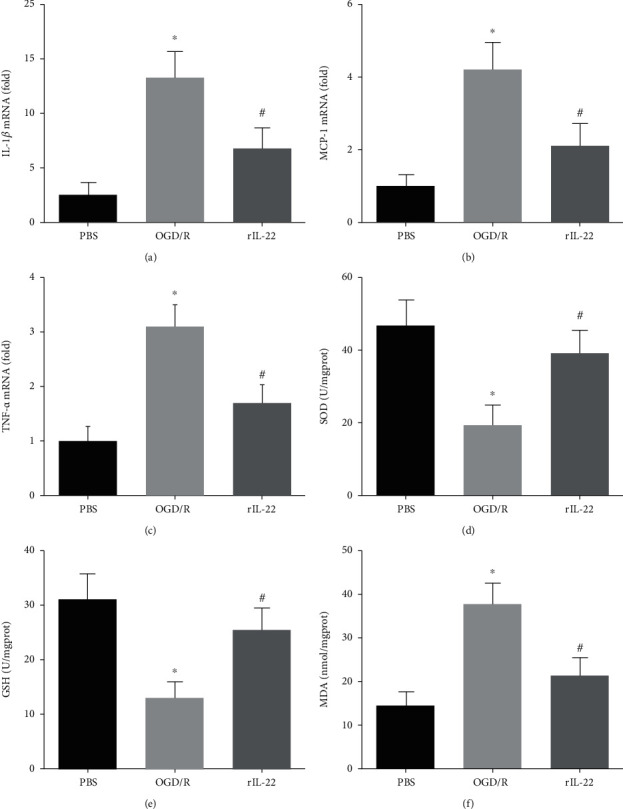
IL-22 treatment inhibited OGD/R-induced inflammation and oxidative stress. The mRNA expression of IL-1β (a), MCP-1 (b), and TNF-α (c) was detected in PC12 cells (n = 6). The levels of SOD (d), GSH (e), and MDA (f) were detected in PC12 cells (n = 6). ∗P < 0.05 vs. PBS group; #P < 0.05 vs. OGD/R group.
3.5. IL-22 Treatment Attenuated OGD/R-Induced Neuronal Apoptosis
The TUNEL staining results showed that the apoptotic index in the OGD/R group was significantly lower than that in the PBS group, while IL-22 treatment significantly diminished OGD/R-induced cell apoptosis (Figure 5(a)). In addition, IL-22 treatment significantly attenuated the restored Bcl-2 expression and reduced Bax expression after OGD/R insult (Figure 5(b)).
Figure 5.
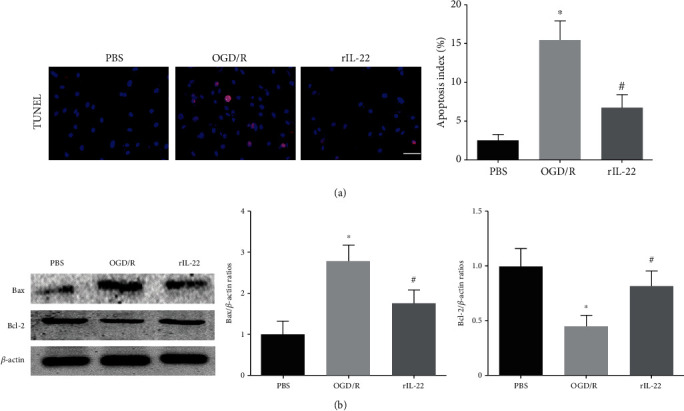
IL-22 treatment attenuated OGD/R-induced neuronal apoptosis. (a) Neuronal apoptosis was detected by TUNEL staining (n = 5, scale bar = 50 μm). (b) The expression of Bax, Bcl-2, and β-actin was detected by western blotting (n = 4). ∗P < 0.05 vs. PBS group; #P < 0.05 vs. OGD/R group.
3.6. IL-22 Treatment Activated the JAK2/STAT3 Signaling Pathway
Our results showed that the phosphorylation levels of JAK2 and STAT3 in the MCAO group were significantly higher than those in the sham group, and IL-22 treatment further increased JAK2 and STAT3 phosphorylation levels (Figure 6(a)). In addition, our results also showed that IL-22 treatment upregulated JAK2 and STAT3 phosphorylation levels in PC12 cells after OGD/R insult (Figure 6(b)).
Figure 6.
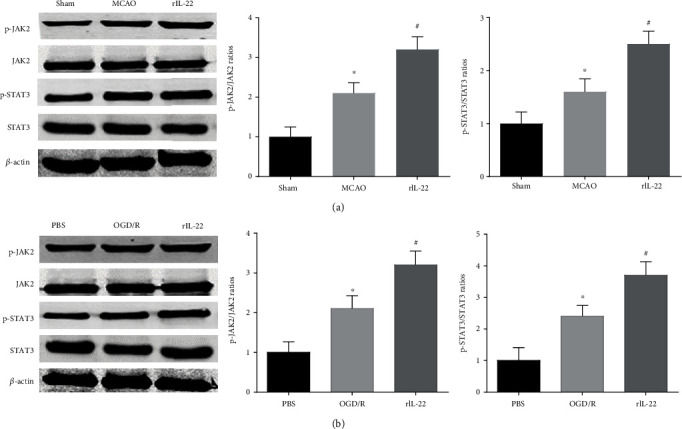
IL-22 treatment activated the JAK2/STAT3 signaling pathway. (a) The expression of p-JAK2, JAK2, p-STAT3, STAT3, and β-actin in the brain was detected by western blotting (n = 4). (b) The expression of p-JAK2, JAK2, p-STAT3, STAT3, and β-actin in PC12 cells was detected by western blotting (n = 4). ∗P < 0.05 vs. the sham or PBS group; #P < 0.05 vs. the MCAO or OGD/R group.
3.7. JAK2 and STAT3 Knockdown Abolished IL-22-Mediated Neuroprotection
To further confirm the effect of the JAK2/STAT3 signaling pathway in IL-22-mediated neuroprotection, transfection with si-JAK2 and si-STAT3 was performed to knock down JAK2 and STAT3 expression in vitro, respectively. The results showed that JAK2 and STAT3 knockdown abolished the IL-22-mediated anti-inflammatory effects by increasing IL-1β, MCP-1, and TNF-α mRNA expression (Figures 7(a)–7(c)). In addition, JAK2 and STAT3 knockdown also attenuated IL-22-mediated antioxidative stress and antiapoptotic effects (Figures 7(d)–7(g)). The above results revealed that the JAK2/STAT3 pathway plays a central role in IL-22-mediated neuroprotective effects.
Figure 7.
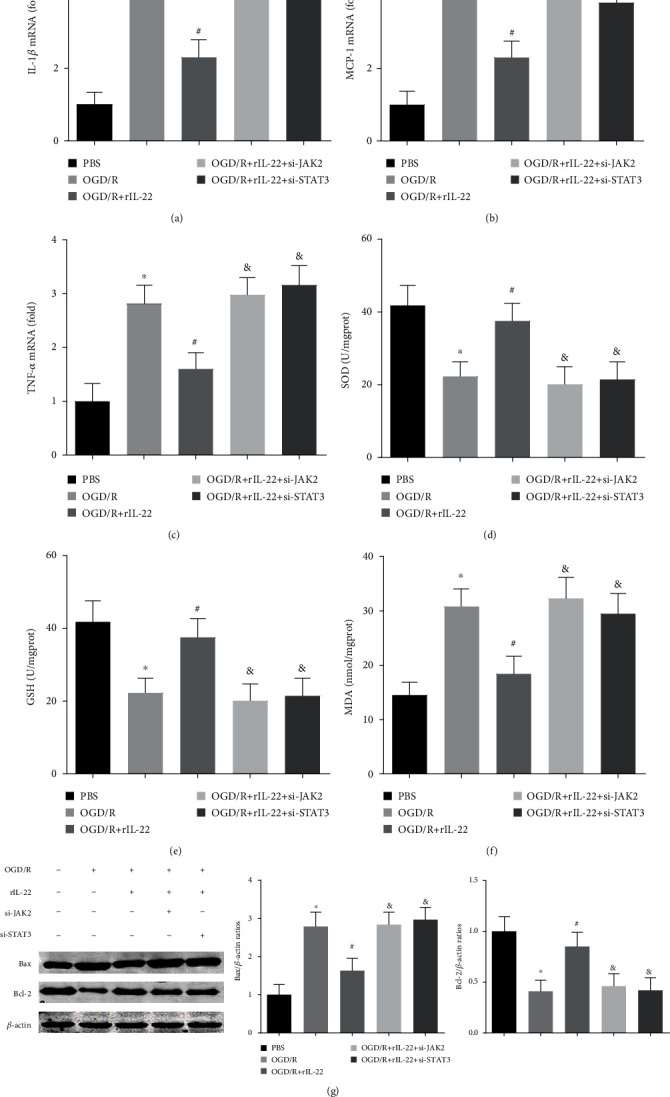
JAK2 and STAT3 knockdown abolished IL-22-mediated neuroprotection. The mRNA expression of IL-1β (a), MCP-1 (b), and TNF-α (c) was detected in PC12 cells (n = 6). The levels of SOD (d), GSH (e), and MDA (f) were detected in PC12 cells (n = 6). (g) The expression of Bax, Bcl-2, and β-actin was detected by western blotting (n = 4). ∗P < 0.05 vs. PBS group; #P < 0.05 vs. OGD/R group; &P < 0.05 vs. OGD/R+rIL-22 group.
4. Discussion
In the current study, we investigated the protective effect of IL-22 against cerebral I/R injury. We provided direct evidence that IL-22 treatment decreased infarct size, neurological deficits, and brain water content in mice subjected to cerebral I/R injury. IL-22 treatment remarkably attenuated the inflammatory response, oxidative stress, and neuronal apoptosis after cerebral I/R injury. In addition, IL-22 treatment decreased the inflammatory response, oxidative stress, and apoptosis of PC12 cells after OGD/R insult. Moreover, IL-22 treatment significantly increased JAK2 and STAT3 phosphorylation levels in mice and PC12 cells, and STAT3 knockdown abolished the IL-22-mediated neuroprotective function. These findings suggest that IL-22 could be exploited as a potential therapeutic agent for ischemic stroke and cerebral I/R injury.
Based on similarities in structure and receptor subunits, the IL-10 family comprises six members, including IL-10, IL-19, IL-20, IL-22, IL-24, and IL-26 [32–34]. As an anti-inflammatory cytokine, the neuroprotective effect of IL-10 on cerebral I/R injury has been identified in numerous studies [35, 36]. In addition, IL-19 administration also reduced ischemia-induced brain infarct and neurological deficits in mice after experimental ischemic stroke, indicating that IL-19 is a novel therapeutic target for cerebral I/R injury [37]. However, IL-20 expression was upregulated in the serum and brain tissue of rats after cerebral I/R, and anti-IL-20 neutralizing antibody administration ameliorated MCAO-induced brain infarction in rats [38]. IL-22 was first identified as a product of CD4+ T cell subsets, and subsequent studies demonstrated that IL-22 is also secreted by macrophage, natural killer cells, and natural killer T cells [24, 39]. Previous studies reported that IL-22 was expressed in human brain tissue and mouse brain, and IL-22 treatment protected nutriment-deprived astrocytes from cell death [40, 41]. Liu et al. also reported that IL-22 treatment significantly inhibited serum starvation-induced PC12 cell death, indicating that IL-22 may confer a neuroprotective function [42].
The results showed that rIL-22 administered in advance significantly decreased the infarct volume and ameliorated neurological deficits and brain water content after cerebral I/R injury in mice. In addition, IL-22 treatment also diminished OGD/R-induced neuronal injury and apoptosis in vitro. These findings demonstrated that IL-22 exerts a neuroprotective effect on cerebral ischemic injury.
The inflammatory response plays a pivotal role in the pathophysiology of I/R-induced cerebral injury. After stroke, the interruption and reperfusion of blood flow in brain tissue trigger inflammatory cell infiltration and cause a robust inflammatory response, which induces neuronal apoptosis and death [43, 44]. Multiple inflammatory-related cytokines are released in ischemic brain injury and participate in the damage and repair process of brain tissue, including ILs, TNF, interferon, and chemokines [45, 46]. In addition, numerous data suggest that the inflammatory response is closely related to oxidative stress and aggravating I/R-induced cerebral injury [47, 48]. Thus, it is clear that therapeutic drugs targeting the inflammatory response and oxidative stress can be very effective in improving cerebral I/R injury. Previous research has shown that IL-22 is an inflammation-related cytokine and has anti-inflammatory and antioxidative stress effects. Thus, we investigated whether IL-22 affects the inflammatory response and oxidative stress in MCAO-induced cerebral I/R injury.
Our results showed that serum levels of inflammatory cytokines, including IL-1β, MCP-1, and TNF-α, in the MCAO group were significantly higher than those in the sham group, while IL-22 treatment significantly reduced the serum levels of these cytokines. In addition, IL-22 treatment also decreased the mRNA expression of IL-1β, MCP-1, and TNF-α in brain tissue after cerebral I/R injury. To assess the effects of IL-22 on cerebral I/R-induced oxidative stress, we measured the SOD and GSH activities and MDA contents in brain tissues. The results showed that the activities of SOD and GSH in the brain tissues were significantly increased and the levels of MDA were significantly reduced in the rIL-22 group compared with those in the MCAO group.
Signaling through the JAK/STAT pathway is important for the progression of neurological diseases, including stroke, traumatic brain injury, status epilepticus, brain tumors, and neurodegenerative diseases [49, 50]. Many lines of evidence have indicated that JAK2/STAT3 signaling is activated in the early stage of cerebral ischemia and mediates oxidative stress, the inflammatory response, and neuronal apoptosis [49, 51]. Kinouchi et al. reported that pioglitazone protects against cerebral I/R injury by activating the JAK2/STAT3 signaling pathway [52]. Liu et al. reported that diosmin inhibits neuronal apoptosis by activating the JAK2/STAT3 signaling pathway after cerebral ischemia in mice [53]. Accumulating evidence suggests that JAK2/STAT3 is a major downstream signal of IL-22 and mediates its hepatoprotective and cardioprotective functions [25, 39].
In the current study, we investigated the role of IL-22 treatment in JAK2/STAT3 signaling after cerebral I/R. The results showed that the phosphorylation levels of JAK2 and STAT3 were upregulated in mice after cerebral I/R injury and in PC12 cells following OGD/R. In addition, IL-22 treatment further increased JAK2 and STAT3 phosphorylation levels, and STAT3 knockdown abolished the IL-22-mediated neuroprotective function. These findings indicate that the neuroprotective actions of IL-22 are related to the JAK2/STAT3 signaling pathway.
Over the last decade, evidence supporting combination therapies has been obtained from a variety of studies in many types of animal models [54, 55]. The combination of thrombolysis and neuroprotection has been considered a promising approach for the treatment of acute ischemic stroke [55, 56]. Tissue plasminogen activator (tPA) is the only treatment approved by the USA FDA for acute ischemic stroke; it dissolves the obstructive clot to restore cerebral blood flow [57]. Neuroprotective agents attenuate the inflammatory response and suppress molecules that mediate thrombosis and blood-brain barrier disruption induced by ischemia such that the benefits of tPA may be extended [55, 56]. Our study shows that IL-22 may be a promising neuroprotective agent; however, the combined effect of IL-22 and tPA is still unclear and will be the focus of our next study.
In conclusion, our findings provide preliminary evidence demonstrating the roles of IL-22 in cerebral I/R injury. IL-22 treatment prevented I/R-induced cerebral injury and neurological deficits by alleviating the inflammatory response, oxidative stress, and neuronal apoptosis. Our data indicate that IL-22 may serve as an attractive therapeutic target for treating ischemic stroke and cerebral I/R injury.
Acknowledgments
This study was supported by grants from the Fundamental Research Funds for the Central Universities (Grant No. WK9110000036).
Data Availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request in compliance with ethical standards.
Conflicts of Interest
No conflicts of interests are declared by the authors.
Authors' Contributions
Chaoshi Niu was involved in the design and execution of experiments, data analysis, and manuscript writing. Yongfei Dong and Chengyun Hu were involved in the design of the study, the execution of experiments, data analysis, and manuscript writing. Chunxia Huang, Jie Gao, Wanxiang Niu, Di Wang, and Yang Wang were all involved in the execution of experiments and data analysis. All authors provided final approval of the version to be submitted. Yongfei Dong and Chengyun Hu contributed equally to this work.
References
- 1.Johnson C. O., Nguyen M., Roth G. A., et al. Global, regional, and national burden of stroke, 1990-2016: a systematic analysis for the Global Burden of Disease Study 2016. The Lancet Neurology. 2019;18(5):439–458. doi: 10.1016/S1474-4422(19)30034-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gorelick P. B. The global burden of stroke: persistent and disabling. The Lancet Neurology. 2019;18(5):417–418. doi: 10.1016/S1474-4422(19)30030-4. [DOI] [PubMed] [Google Scholar]
- 3.Amani H., Mostafavi E., Alebouyeh M. R., et al. Would colloidal gold nanocarriers present an effective diagnosis or treatment for ischemic stroke? International Journal of Nanomedicine. 2019;14:8013–8031. doi: 10.2147/IJN.S210035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Xiao G., Lyu M., Wang Y., et al. Ginkgo flavonol glycosides or ginkgolides tend to differentially protect myocardial or cerebral ischemia-reperfusion injury via regulation of TWEAK-Fn14 signaling in heart and brain. Frontiers in Pharmacology. 2019;10:p. 735. doi: 10.3389/fphar.2019.00735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Khan M. S., Khan A., Ahmad S., et al. Inhibition of JNK alleviates chronic hypoperfusion-related ischemia induces oxidative stress and brain degeneration via Nrf2/HO-1 and NF-κB signaling. Oxidative Medicine and Cellular Longevity. 2020;2020:18. doi: 10.1155/2020/5291852.5291852 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wu F., Ling Y., Yang L., Cheng X., Dong Q., Cao W. High level of serum tissue kallikrein is associated with favorable outcome in acute ischemic stroke patients. Disease Markers. 2019;2019:6. doi: 10.1155/2019/5289715.5289715 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cao Y., Cui C., Zhao H., et al. Plasma osteoprotegerin correlates with stroke severity and the occurrence of microembolic signals in patients with acute ischemic stroke. Disease Markers. 2019;2019:7. doi: 10.1155/2019/3090364.3090364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zhang W., Song J., Li W., et al. Salvianolic acid D alleviates cerebral ischemia-reperfusion injury by suppressing the cytoplasmic translocation and release of HMGB1-triggered NF-κB activation to inhibit inflammatory response. Mediators of Inflammation. 2020;2020:15. doi: 10.1155/2020/9049614.9049614 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bernstock J. D., Peruzzotti-Jametti L., Leonardi T., et al. SUMOylation promotes survival and integration of neural stem cell grafts in ischemic stroke. eBioMedicine. 2019;42:214–224. doi: 10.1016/j.ebiom.2019.03.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Geng W., Cai L., Han K., et al. Electroacupuncture pretreatment alleviates cerebral ischemia-reperfusion injury by increasing GSK-3β phosphorylation level via adenosine A1 receptor. BioMed Research International. 2020;2020:9. doi: 10.1155/2020/6848450.6848450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Huang Y., Liu Z., Tan F., Hu Z., Lu M. Effects of the insulted neuronal cells-derived extracellular vesicles on the survival of umbilical cord-derived mesenchymal stem cells following cerebral ischemia/reperfusion injury. Oxidative Medicine and Cellular Longevity. 2020;2020:26. doi: 10.1155/2020/9768713.9768713 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zeng J., Zhu L., Liu J., et al. Metformin protects against oxidative stress injury induced by ischemia/reperfusion via regulation of the lncRNA-H19/miR-148a-3p/Rock2 axis. Oxidative Medicine and Cellular Longevity. 2019;2019:18. doi: 10.1155/2019/8768327.8768327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhang B., Zhang H. X., Shi S. T., et al. Interleukin-11 treatment protected against cerebral ischemia/reperfusion injury. Biomedicine & Pharmacotherapy. 2019;115, article ??? doi: 10.1016/j.biopha.2019.108816. [DOI] [PubMed] [Google Scholar]
- 14.Semple B. D., Dill L. K., O'Brien T. J. Immune challenges and seizures: how do early life insults influence epileptogenesis? Frontiers in Pharmacology. 2020;11:p. 2. doi: 10.3389/fphar.2020.00002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Liu J., Xing Y., Gao Y., Zhou C. Changes in serum interleukin-33 levels in patients with acute cerebral infarction. Journal of Clinical Neuroscience. 2014;21(2):298–300. doi: 10.1016/j.jocn.2013.04.036. [DOI] [PubMed] [Google Scholar]
- 16.Smith C. J., Hulme S., Vail A., et al. SCIL-STROKE (subcutaneous interleukin-1 receptor antagonist in ischemic stroke): a randomized controlled phase 2 trial. Stroke. 2018;49(5):1210–1216. doi: 10.1161/STROKEAHA.118.020750. [DOI] [PubMed] [Google Scholar]
- 17.Xu C., Zhu H., Shen R., Feng Q., Zhou H., Zhao Z. IL-35 is a protective immunomodulator in brain ischemic injury in mice. Neurochemical Research. 2018;43(7):1454–1463. doi: 10.1007/s11064-018-2560-5. [DOI] [PubMed] [Google Scholar]
- 18.Liu C., Xu X., Huang C., Shang D., Zhang L., Wang Y. Inhibition of IL-32 expression ameliorates cerebral ischemia-reperfusion injury via the NOD/MAPK/NF-κB signaling pathway. Journal of Molecular Neuroscience. 2020;70(11):1713–1727. doi: 10.1007/s12031-020-01557-0. [DOI] [PubMed] [Google Scholar]
- 19.Xie M. H., Aggarwal S., Ho W. H., et al. Interleukin (IL)-22, a novel human cytokine that signals through the interferon receptor-related proteins CRF2-4 and IL-22R∗. The Journal of Biological Chemistry. 2000;275(40):31335–31339. doi: 10.1074/jbc.M005304200. [DOI] [PubMed] [Google Scholar]
- 20.Dudakov J. A., Hanash A. M., van den Brink M. R. Interleukin-22: immunobiology and pathology. Annual Review of Immunology. 2015;33(1):747–785. doi: 10.1146/annurev-immunol-032414-112123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Weidenbusch M., Rodler S., Anders H. J. Interleukin-22 in kidney injury and regeneration. American Journal of Physiology. Renal Physiology. 2015;308(10):F1041–F1046. doi: 10.1152/ajprenal.00005.2015. [DOI] [PubMed] [Google Scholar]
- 22.Liu Y., Verma V. K., Malhi H., et al. Lipopolysaccharide downregulates macrophage-derived IL-22 to modulate alcohol-induced hepatocyte cell death. American Journal of Physiology. Cell Physiology. 2017;313(3):C305–C313. doi: 10.1152/ajpcell.00005.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hu M., Yang S., Yang L., Cheng Y., Zhang H. Interleukin-22 alleviated palmitate-induced endoplasmic reticulum stress in INS-1 cells through activation of autophagy. PLoS One. 2016;11, article e146818 doi: 10.1371/journal.pone.0146818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Shi L., Ji Q., Liu L., et al. IL-22 produced by Th22 cells aggravates atherosclerosis development in ApoE−/−mice by enhancing DC-induced Th17 cell proliferation. Journal of Cellular and Molecular Medicine. 2020;24(5):3064–3078. doi: 10.1111/jcmm.14967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Takahashi J., Yamamoto M., Yasukawa H., et al. Interleukin-22 directly activates myocardial STAT3 (signal transducer and activator of transcription-3) signaling pathway and prevents myocardial ischemia reperfusion injury. Journal of the American Heart Association. 2020;9:p. e14814. doi: 10.1161/JAHA.119.014814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Xu M. J., Feng D., Wang H., Guan Y., Yan X., Gao B. IL-22 ameliorates renal ischemia-reperfusion injury by targeting proximal tubule epithelium. Journal of the American Society of Nephrology. 2014;25(5):967–977. doi: 10.1681/ASN.2013060611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Guan X., Wang Y., Kai G., et al. Cerebrolysin ameliorates focal cerebral ischemia injury through neuroinflammatory inhibition via CREB/PGC-1α pathway. Frontiers in Pharmacology. 2019;10:p. 1245. doi: 10.3389/fphar.2019.01245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Longa E. Z., Weinstein P. R., Carlson S., Cummins R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke. 1989;20(1):84–91. doi: 10.1161/01.STR.20.1.84. [DOI] [PubMed] [Google Scholar]
- 29.Li J., Zhang K., Zhang Q., et al. PPAR-γ mediates Ta-VNS-induced angiogenesis and subsequent functional recovery after experimental stroke in rats. BioMed Research International. 2020;2020:12. doi: 10.1155/2020/8163789.8163789 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tang C., Hu Y., Lyu H., et al. Neuroprotective effects of 1-O-hexyl-2,3,5-trimethylhydroquinone on ischaemia/reperfusion-induced neuronal injury by activating the Nrf2/HO-1 pathway. Journal of Cellular and Molecular Medicine. 2020;24(18):10468–10477. doi: 10.1111/jcmm.15659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jiang L., Gong Y., Hu Y., et al. Peroxiredoxin-1 overexpression attenuates doxorubicin-induced cardiotoxicity by inhibiting oxidative stress and cardiomyocyte apoptosis. Oxidative Medicine and Cellular Longevity. 2020;2020:11. doi: 10.1155/2020/2405135.2405135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Burmeister A. R., Marriott I. The interleukin-10 family of cytokines and their role in the CNS. Frontiers in Cellular Neuroscience. 2018;12:p. 458. doi: 10.3389/fncel.2018.00458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sziksz E., Pap D., Lippai R., et al. Fibrosis related inflammatory mediators: role of the IL-10 cytokine family. Mediators of Inflammation. 2015;2015:15. doi: 10.1155/2015/764641.764641 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mocan M., Mocan Hognogi L. D., Anton F. P., et al. Biomarkers of inflammation in left ventricular diastolic dysfunction. Disease Markers. 2019;2019:14. doi: 10.1155/2019/7583690.7583690 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Nakajima M., Nito C., Sowa K., et al. Mesenchymal stem cells overexpressing interleukin-10 promote neuroprotection in experimental acute ischemic stroke. Molecular Therapy - Methods & Clinical Development. 2017;6:102–111. doi: 10.1016/j.omtm.2017.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chen M., Yang Y. A meta-analysis on associations of IL-6 and IL-10 polymorphisms with susceptibility to ischemic stroke. Journal of Neuroimmunology. 2019;335, article 577004 doi: 10.1016/j.jneuroim.2019.577004. [DOI] [PubMed] [Google Scholar]
- 37.Xie W., Fang L., Gan S., Xuan H. Interleukin-19 alleviates brain injury by anti-inflammatory effects in a mice model of focal cerebral ischemia. Brain Research. 2016;1650:172–177. doi: 10.1016/j.brainres.2016.09.006. [DOI] [PubMed] [Google Scholar]
- 38.Chen W. Y., Chang M. S. IL-20 is regulated by hypoxia-inducible factor and up-regulated after experimental ischemic stroke. Journal of Immunology. 2009;182(8):5003–5012. doi: 10.4049/jimmunol.0803653. [DOI] [PubMed] [Google Scholar]
- 39.Xiang X., Hwang S., Feng D., Shah V. H., Gao B. Interleukin-22 in alcoholic hepatitis and beyond. Hepatology International. 2020;14(5):667–676. doi: 10.1007/s12072-020-10082-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Perriard G., Mathias A., Enz L., et al. Interleukin-22 is increased in multiple sclerosis patients and targets astrocytes. Journal of Neuroinflammation. 2015;12(1):p. 119. doi: 10.1186/s12974-015-0335-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Dumoutier L., Louahed J., Renauld J. C. Cloning and characterization of IL-10-related T cell-derived inducible factor (IL-TIF), a novel cytokine structurally related to IL-10 and inducible by IL-9. Journal of Immunology. 2000;164(4):1814–1819. doi: 10.4049/jimmunol.164.4.1814. [DOI] [PubMed] [Google Scholar]
- 42.Liu Y., Pan W., Yang S., et al. Interleukin-22 protects rat PC12 pheochromocytoma cells from serum deprivation-induced cell death. Molecular and Cellular Biochemistry. 2012;371(1-2):137–146. doi: 10.1007/s11010-012-1430-8. [DOI] [PubMed] [Google Scholar]
- 43.Wang G., Han B., Shen L., et al. Silencing of circular RNA HIPK2 in neural stem cells enhances functional recovery following ischaemic stroke. eBioMedicine. 2020;52, article 102660 doi: 10.1016/j.ebiom.2020.102660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wang J., Mao J., Wang R., Li S., Wu B., Yuan Y. Kaempferol protects against cerebral ischemia reperfusion injury through intervening oxidative and inflammatory stress induced apoptosis. Frontiers in Pharmacology. 2020;11:p. 424. doi: 10.3389/fphar.2020.00424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Pawluk H., Woźniak A., Grześk G., et al. The role of selected pro-inflammatory cytokines in pathogenesis of ischemic stroke. Clinical Interventions in Aging. 2020;15:469–484. doi: 10.2147/CIA.S233909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Zhang L., Liu C., Huang C., Xu X., Teng J. miR-155 knockdown protects against cerebral ischemia and reperfusion injury by targeting MafB. BioMed Research International. 2020;2020:11. doi: 10.1155/2020/6458204.6458204 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Yu J., Wang W.-N., Matei N., et al. Ezetimibe attenuates oxidative stress and neuroinflammation via the AMPK/Nrf2/TXNIP pathway after MCAO in rats. Oxidative Medicine and Cellular Longevity. 2020;2020:14. doi: 10.1155/2020/4717258.4717258 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wattanathorn J., Ohnon W., Thukhammee W., Muchmapura S., Wannanon P., Tong-Un T. Cerebroprotective effect against cerebral ischemia of the combined extract of Oryza sativa and Anethum graveolens in metabolic syndrome rats. Oxidative Medicine and Cellular Longevity. 2019;2019:19. doi: 10.1155/2019/9658267.9658267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Nicolas C. S., Amici M., Bortolotto Z. A., et al. The role of JAK-STAT signaling within the CNS. JAK-STAT. 2014;2(1, article e22925) doi: 10.4161/jkst.22925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Li L., Sun L., Qiu Y., Zhu W., Hu K., Mao J. Protective effect of stachydrine against cerebral ischemia-reperfusion injury by reducing inflammation and apoptosis through P65 and JAK2/STAT3 signaling pathway. Frontiers in Pharmacology. 2020;11:p. 64. doi: 10.3389/fphar.2020.00064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Liang Z., Wu G., Fan C., et al. The emerging role of signal transducer and activator of transcription 3 in cerebral ischemic and hemorrhagic stroke. Progress in Neurobiology. 2016;137:1–16. doi: 10.1016/j.pneurobio.2015.11.001. [DOI] [PubMed] [Google Scholar]
- 52.Kinouchi T., Kitazato K. T., Shimada K., et al. Activation of signal transducer and activator of transcription-3 by a peroxisome proliferator-activated receptor gamma agonist contributes to neuroprotection in the peri-infarct region after ischemia in oophorectomized rats. Stroke. 2012;43(2):478–483. doi: 10.1161/STROKEAHA.111.618926. [DOI] [PubMed] [Google Scholar]
- 53.Liu X., Zhang X., Zhang J., et al. Diosmin protects against cerebral ischemia/reperfusion injury through activating JAK2/STAT3 signal pathway in mice. Neuroscience. 2014;268:318–327. doi: 10.1016/j.neuroscience.2014.03.032. [DOI] [PubMed] [Google Scholar]
- 54.Poustchi F., Amani H., Ahmadian Z., et al. Combination therapy of killing diseases by injectable hydrogels: from concept to medical applications. Advanced Healthcare Materials. 2021;10(3, article e2001571) doi: 10.1002/adhm.202001571. [DOI] [PubMed] [Google Scholar]
- 55.Wang Y., Li Q., Wang J., Zhuang Q. K., Zhang Y. Y. Combination of thrombolytic therapy and neuroprotective therapy in acute ischemic stroke: is it important? European Review for Medical and Pharmacological Sciences. 2015;19(3):416–422. [PubMed] [Google Scholar]
- 56.Fukuta T., Asai T., Yanagida Y., et al. Combination therapy with liposomal neuroprotectants and tissue plasminogen activator for treatment of ischemic stroke. The FASEB Journal. 2017;31(5):1879–1890. doi: 10.1096/fj.201601209R. [DOI] [PubMed] [Google Scholar]
- 57.Thiebaut A. M., Gauberti M., Ali C., et al. The role of plasminogen activators in stroke treatment: fibrinolysis and beyond. The Lancet Neurology. 2018;17(12):1121–1132. doi: 10.1016/S1474-4422(18)30323-5. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request in compliance with ethical standards.