

Abstract
The effect of capric acid, secreted by the probiotic yeasts Saccharomyces boulardii, was evaluated on the activities of fluconazole (FLC) and amphotericin B (AMB) against pathogenic Candida albicans fungus. The findings indicated that capric acid may be a promising additive for use in combination with FLC. A FLC-capric acid combination led to reduced efflux activity of multidrug resistance (MDR) transporter Cdr1p by causing it to relocalize from the plasma membrane (PM) to the interior of the cell. The above effect occurred due to inhibitory effect of FLC-capric acid combination of ergosterol biosynthesis. However, capric acid alone stimulated ergosterol production in C. albicans, which in turn generated cross resistance towards AMB and inhibited its action (PM permeabilization and cytoplasm leakage) against C. albicans cells. This concluded that AMB should not be administered among dietary supplements containing capric acid or S. boulardii cells.
Subject terms: Antifungal agents, Fungi, Pathogens
Introduction
Candida albicans is a resident of human microbiota present on the skin, in the gastrointestinal tract and in the genital mucosae1. Under immunosuppression, C. albicans causes opportunistic infections, which may result in sepsis and multiple organ failure2,3.
There is limited range of antifungal drug classes available to combat candidiasis. One such drug class includes azoles, which inhibit ergosterol biosynthesis by targeting cytochrome P-450 lanosterol 14α-demethylase (CYP51A1, Erg11p) encoded by the ERG11 gene4. However, fungi increasingly develop resistance towards azoles, including the most commonly prescribed fluconazole (FLC)5–7. In the case of C. albicans, two major resistance mechanisms have been reported. These involve structural alterations to the target enzyme by overexpression or point mutations of the ERG11 gene8, as well as overproduction of multidrug resistance (MDR) transporters, which remove azoles from the fungal cell9. One of the alternative treatment option involves the use of polyenes, such as amphotericin B (AMB). Polyenes bind to ergosterol in the fungal plasma membrane (PM), inducing permeabilization and cytoplasm leakage10. However, polyenes are toxic towards the mammalian cells and treatment is associated with a number of adverse effects among patients11. Resistance towards polyenes is not common among clinical C. albicans isolates and include downproduction of ergosterol12. However, laboratory-delivered polyene-resistant C. albicans strains overproduce ergosterol13.
Even with antifungal therapy, invasive candidiases results in a 50–70% patient mortality rate14. Therefore, the development of new therapies for fungal diseases remains a challenging priority for modern medicine. The trend towards green chemistry has motivated the quest for natural compounds as novel antimicrobials, such as fatty acids (FAs)15, which have been described to possess antiviral, antibacterial and antifungal properties16. With regards to fungi, FAs are active against molds, dermatophytes, phytopathogens and yeast-like fungal species17. Although FAs are not as effective as polyenes or azoles, it has been reported that pathogenic fungi are less likely to become resistant to FAs17.
FAs are metabolites derived from various sources, including probiotic microorganisms. One such organism, probiotic yeast Saccharomyces boulardii, is known to produce caproic (C6:0), caprylic (C8:0) and capric (C10:0) acids18, the latter of which has been reported to possess high activity against C. albicans19. The toxic effects of capric acid on C. albicans include shrinking of the cytoplasm in this fungus19, as well as inhibition of C. albicans virulence factors, such as adherence and the formation of hyphae18.
Another promising approach to combating antifungal drug resistance is the identification of compounds which synergistically enhance the activity of currently available drugs12. Simultaneously, the use of compounds (such as drugs or dietary supplements) that would antagonistically inhibit the activity of antifungal drugs or have undesirable interactions with them is avoided in clinical settings20,21. In this study we aimed to investigate the effect of capric acid on the antifungal properties of FLC and AMB against C. albicans. We observed that capric acid co-administered with FLC inhibits the activity of Cdr1 transporter, suggesting that this may be a promising synergistic agent in combination with azole drugs. Conversely, we observed that treatment with capric acid leads to overproduction of ergosterol in C. albicans cells, leading to cross resistance towards AMB.
Results
Capric acid has a synergistic effect in combination with fluconazole, but an antagonistic effect in combination with amphotericin B
The effect of capric acid treatment was evaluated against the in-house generated C. albicans KS028 strain, which features an ERG11 gene deletion, depriving the cells of ergosterol (Fig. 1A). The C. albicans KS028 strain was reported to be unaffected by azoles or polyenes due to the absence of their respective targets Erg11p and ergosterol22. Thus, drug screening using the KS028 strain may allow the characterization of new synergistic or antagonistic drug combinations23,24.
Figure 1.

(A) Growth phenotypes of C. albicans CAF2-1 (parental strain) and KS028 (erg11∆/∆) strains after 48 h incubation at 28 °C in YPD medium with capric acid (0–90.6 µg/mL). (B) Percentage of growth of the C. albicans CAF2-1 strain in the presence of fluconazole (0.5–8 μg/mL, chart legend) and in presence of 45.3 μg/mL capric acid (means ± SD, n = 3). (C) Percentage of growth of the C. albicans CAF2-1 strain in the presence of amphotericin B (0.25–8 μg/mL, chart legend) and in presence of 45.3 μg/mL capric acid (means ± SD, n = 3). Statistical analyses were performed by comparing growth at the same fluconazole (B) or amphotericin B (C) concentrations between samples treated and untreated with capric acid. Statistical significance in all cases is presented as follows: **p < 0.01; ***p < 0.001.
The parental C. albicans strain (CAF2-1) continued to proliferate in the presence of all capric acid concentrations used in this study (Fig. 1A). Slight growth inhibition was observed upon treatment with 90.6 µg/mL capric acid. At a capric acid concentration of 22.7 µg/mL, a marked growth inhibition was observed in the KS028 strain. Complete growth inhibition of KS028 was observed upon treatment with 45.3 µg/mL capric acid. Based on these data, a 45.3 µg/mL capric acid concentration was selected for combination with FLC (Fig. 1B) or AMB (Fig. 1C).
Combination with capric acid enhanced the activity of FLC against C. albicans CAF2-1 (Fig. 1B). The growth of C. albicans at FLC concentrations of 2, 4 and 8 μg/mL was 50, 40 and 37%, respectively. Introduction of capric acid resulted in a reduction of CAF2-1 growth to 20, 17 and 16.5% in corresponding concentrations of FLC. In the case of AMB, it was observed that the presence of capric acid almost completely inhibited its activity against C. albicans CAF2-1 (Fig. 1C). Under control conditions, a complete reduction in C. albicans growth was observed at 0.25 µg/mL of AMB. The addition of capric acid resulted in 80–90% growth in the presence of a concentration range of 0.25–4 μg/mL AMB and 65% in the presence of a concentration of 8 μg/mL of AMB (Fig. 1C).
Activity, localization, and expression of the Cdr1 transporter are altered in C. albicans treated with the capric acid or fluconazole–capric acid combination
Expression and activation of MDR transporters is the first line defense of C. albicans towards azole drugs25. Cdr1p belongs to the ATP-binding cassette family and is the most influential of the identified MDR transporters involved in FLC efflux in C. albicans26. The FLC–capric acid combination lead to greater growth inhibition in C. albicans than treatment with FLC alone (Fig. 1). Thus, we aimed to investigate the relative effect of capric acid and FLC-capric acid combination on the expression and activity of CDR1, as well as on the localization of Cdr1p (Fig. 2).
Figure 2.

(A) Relative CDR1 gene expression in the C. albicans CAF2-1. Gene expression levels are reported as means ± SD of 2−ΔΔCT values (n = 3), normalized to 1 for control conditions. (B) Cdr1p-dependent rhodamine 6G (R6G efflux) in the C. albicans CAF2-1 normalized to = 1 for untreated control; means ± SD, n = 3. (C) Fluorescence micrographs of the subcellular localization of the Cdr1-GFP protein in the C. albicans KS052 (CAF2-1 CDR1-GFP). For the experiments C. albicans was grown for 8 h in the following conditions: control without antimicrobial agents, FLC—treated with fluconazole 4 μg/mL; Capric acid—treated with capric acid 45.3 μg/mL; FLC + Capric acid—simultaneously treated with fluconazole 4 μg/mL and capric acid 45.3 μg/mL. Statistical analysis was performed by comparing the CDR1 expression level (A) or R6G fluorescence intensity (B) of cells treated with antimicrobial agent(s) with the corresponding untreated control. Scale bar = 5 μm. Statistical significance was presented as follows: **p < 0.01; ***p < 0.001.
FLC alone induced increased CDR1 expression almost 18-fold and capric acid alone almost fivefold compared with untreated C. albicans cells (Fig. 2A). However, an only 1.3-fold increase in CDR1 expression was identified after treating C. albicans with combination of both compounds.
Cdr1p activity was determined by monitoring the extracellular fluorescence intensity of its fluorescent substrate, R6G27. Treatment with FLC and FLC-capric acid combination resulted in 60% and 70% lower efflux activity of Cdr1, respectively (Fig. 2B). On the other hand, treatment with capric acid alone lead to 2.6-fold higher activity of Cdr1.
We previously reported that Cdr1p localization to the plasma membrane (PM) is critical for its efflux activity22, hence we used CDR1-GFP tagged strain KS05223 to investigate Cdr1p localization after treatment with capric acid or FLC-capric acid combination. Localization of Cdr1-GFP in PM was observed only in C. albicans cells with or without capric acid (Fig. 2C). Upon FLC or FLC-capric acid treatment, mislocalization of Cdr1p from PM to the inside of the cells was observed (Fig. 2C).
Sterol composition is altered in C. albicans treated with capric acid or fluconazole-capric acid combination
The presence of ergosterol in C. albicans’ cells and the fluidity of the membranes are major factors affecting both localization of Cdr1 in PM as well as its efflux activity22. Mislocalization of Cdr1p observed after FLC-capric acid treatment (Fig. 2C) prompted investigation of the content of sterols in the PM, as well as PM fluidity (Table 1).
Table 1.
Sterols (μg/mg dry mass of isolated PM lipids, means ± SD, n = 3) in C. albicans CAF2-1 grown for 8 h in the following conditions: control without antimicrobial agents; FLC—treated with fluconazole 4 μg/mL; Capric acid—treated with capric acid 45.3 μg/mL; FLC + Capric acid—simultaneously treated with fluconazole 4 μg/mL and capric acid 45.3 μg/mL. General polarization values (GP; means ± SD, n = 6) of Laurdan incorporated into PMs of C. albicans CAF2-1 grown for 8 h with antimicrobial agent(s). Statistical analysis was performed by comparing cells treated with antimicrobial agent(s) with the corresponding untreated controls (*p < 0.05; **p < 0.01; ***p < 0.001).
| Control | FLC | Capric acid | FLC + Capric acid | |
|---|---|---|---|---|
| Ergosterol | 35.14 ± 2.84 | 4.53 ± 0.3*** | 54.39 ± 0.9*** | 13.47 ± 1.65*** |
| Lanosterol | 7.19 ± 1.36 | 20.96 ± 2.27* | 8.77 ± 0.72 | 14.37 ± 1.19* |
| GP | −0.26 ± 0.04 | −0.14 ± 0.02* | 0.13 ± 0.05*** | −0.02 ± 0.001** |
The PM of C. albicans cells treated with FLC had an almost eightfold lower ergosterol content, but a threefold higher concentration of lanosterol (Table 1). Capric acid treatment resulted in a 1.6-fold increase in ergosterol concentration in the PM, with no significant differences in lanosterol content. However, the combination of both compounds leads to 2.6-fold decrease in ergosterol content and twofold increase in lanosterol content, compared with untreated cells. Treatment with both compounds appeared to result in higher Laurdan’s general polarization (GP) values, which indicates more rigid PMs. The highest GP values were observed in cells treated with capric acid alone, which indicated almost fourfold more rigid PM, than in case of untreated cells. The treatment with FLC, or FLC-capric acid combination has led to 2- or threefold more rigid PMs, respectively (Table 1).
ERG11 expression is affected differently in C. albicans treated with capric acid or fluconazole-capric acid combination
Alterations in sterol profiles in FLC-resistant C. albicans isolates are mostly the result of over- or underexpression of the ERG11 gene4,28. ERG11 encodes an enzyme, lanosterol 14α-demethylase, which directly converts lanosterol into further ergosterol metabolites29. To understand the variations in sterol content in C. albicans treated with capric acid or an FLC–capric acid combination (Table 1), we evaluated ERG11 gene expression (Fig. 3).
Figure 3.
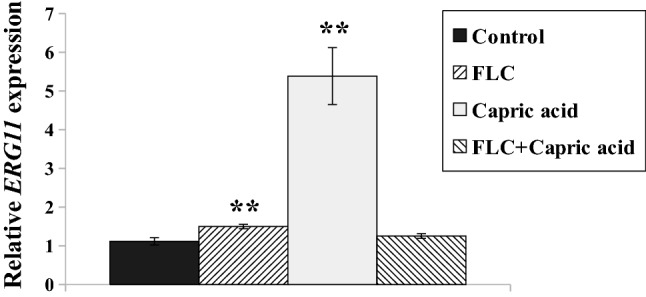
Relative ERG11 gene expression in the C. albicans CAF2-1 grown for 8 h in the following conditions: control without antimicrobial agents, FLC—treated with fluconazole 4 μg/mL; Capric acid—treated with capric acid 45.3 μg/mL; FLC + Capric acid—simultaneously treated with fluconazole 4 μg/mL and capric acid 45.3 μg/mL. Gene expression levels are reported as means ± SD of 2−ΔΔCT values (n = 3), normalized to 1 for control conditions. Statistical analysis was performed by comparing the ERG11 expression level of cells treated with antimicrobial agent(s) with the corresponding untreated control. Statistical significance was presented as follows: **p < 0.01.
An increase in ERG11 expression was observed in response to FLC or capric acid treatment (Fig. 3). FLC treatment lead to only 1.5-fold higher ERG11 gene expression, while capric acid treatment resulted in 5.4-fold higher ERG11 expression. Combining both compounds reverted the effect and ERG11 expression resembled that of untreated cells (Fig. 3).
Ergosterol overproduction induced by treating C. albicans with capric acid inhibits plasma membrane permeabilization and K+ leakage by amphotericin B
Combination treatment with capric acid and AMB lead to an antagonistic effect against C. albicans (Fig. 1C). Treatment with capric acid resulted in higher content of ergosterol in PM (Table 1). As C. albicans may acquire resistance to AMB by overproduction of ergosterol13, we decided to investigate the influence of AMB on C. albicans cells grown in the presence of capric acid.
The mechanism of AMB toxicity is the binding to fungal ergosterol, permeabilization of PM and cytoplasm leakage30,31. Our previous studies indicated that the mechanism of action of AMB also involved delocalization of membrane proteins, including Cdr1p27. Here, we characterized the effect of AMB in C. albicans CAF2-1 cells treated with capric acid (Fig. 4).
Figure 4.

(A) Amphotericin B (AMB) binding to C. albicans CAF2-1 cells (means ± SD, n = 3). (B) PM permeabilization measured by counting propidium iodide (PI) positive cells (means ± SD, n = 6). (C) Fluorescence micrographs of the subcellular localization of the Cdr1-GFP protein in the C. albicans KS052 (CAF2-1 CDR1-GFP) after treatment with AMB. (D) Concentration of extracellular potassium (K+) (means ± SD, n = 3). For the experiments C. albicans was grown for 8 h without capric acid (control) or treated with capric acid 45.3 μg/mL. Following the culture, cells were washed to get rid of capric acid and treated with 2 µg/mL of AMB for 1 h. Statistical analyses (B and D) were performed by comparing the results cells treated with AMB with the corresponding untreated control. Scale bar = 5 μm. Statistical significance was presented as follows: ***p < 0.001.
The binding of AMB to C. albicans cells (Fig. 4A) was monitored by spectrophotometry at 409 nm32. In C. albicans CAF2-1 cells treated for 8 h with 45.3 µg/mL of capric acid, AMB binding to C. albicans were at a similar level (> 95%) as was observed in untreated cells and no statistically significant difference was observed. However, AMB-treatment of C. albicans cells not also treated with capric acid, resulted in permeabilization of PM in ~ 65% of cells (Fig. 4B). In the presence of capric acid, PM was permeabilized in about 10% of AMB-treated cells, which was comparable to untreated cells (Fig. 4B).
C. albicans cells grown with AMB and without capric acid featured dispersion of the Cdr1-GFP protein inside the cells and delocalization from the PM (Fig. 4C). When cells were cultured in the presence of capric acid, PM localization of Cdr1-GFP was observed even after AMB treatment (Fig. 4C).
Cytoplasm leakage was evaluated by measuring the extracellular concentration of potassium ions (K+), using inductively coupled plasma atomic emission spectroscopy (ICP-AES) (Fig. 4D). C. albicans cells grown in the presence of capric acid and treated with AMB were characterized by almost threefold lower extracellular K+ concentration than cells not treated with capric acid. Capric acid used alone did not K+ efflux from the cells, which was similar to entirely untreated C. albicans cells (Fig. 4D).
Discussion
Contemporary medicine is faced with the challenge of the development of resistance among Candida spp. to the common antifungal drug classes including azoles and polyenes. Cytotoxicity of polyenes is another major concern. For these reasons, there is an urgent need for alternative treatment options to the currently available antifungal regimens. Investigating natural compounds in combination with antifungal drugs have recently gained increasing research interest. In this study we report the effect of capric acid on the activities of the most commonly used azole and polyene drugs, FLC and AMB, respectively. Capric acid is a short-chain (C10:0) fatty acid (FA) synthesized by the yeast S. boulardii and is commonly used as an anti-diarrheal dietary supplement18. Our previous studies indicated that capric acid has anti-adhesion activity and inhibits the adhesion of C. albicans cells to abiotic surfaces and intestinal epithelial cells18. Furthermore, not only does capric acid display no cytotoxic effects towards human cells33, but it has also been reported to inhibit proliferation of different tumor cell lines34.
In this study, capric acid was toxic to the C. albicans ERG11 deletion mutant (erg11Δ/Δ) which was as a result negative for ergosterol (Fig. 1A). It has been previously suggested that the presence of ergosterol in fungal cell membranes is crucial for the cellular response to fatty acid treatment17. In addition, it has been shown that fungal species lacking sterols are highly sensitive to heptadecenoic acid35. Similarly, S. cerevisiae cells deficient in ergosterol due to deletion of the ERG4 gene showed increased sensitivity to undecanoic and lauric acids36. The C. albicans erg11Δ/Δ strain is also a model example of azole-treated fungal cells22, which has been previously used in typing synergistic additives to azole drugs24. In our study, the combination of capric acid with FLC was shown to reduce the growth of C. albicans CAF2-1 to a greater extent than treatment with FLC alone (Fig. 1B). Similarly, the addition of capric acid enhanced the effect of FLC on clinical, FLC-resistant C. albicans isolates. The data was included in Supplementary Information (Fig. S1). In case of C. albicans CAF2-1, this difference did not affect the MIC50 value of FLC (MIC50 = 2 µg/mL), but we found a promising effect of the combination FLC and capric acid on Cdr1p (Fig. 2). Separately, either FLC or capric acid induced expression of CDR1 gene (Fig. 2A). However, upon combination of both compounds, the expression of CDR1 reverted to a similar level to that of untreated C. albicans cells (Fig. 2A). This effect, as well as Cdr1p mislocalization (Fig. 2C), lead to highly reduced efflux activity of this protein (Fig. 2B). Detection of Cdr1 proteins, which act as inhibitors of C. albicans, is one of strategies in overcoming drug resistance to azole drugs37. The Cdr1 inhibitors described to date, such as enniatin A and beauvericin, are ionophores which induce the permeability of the cell membrane for ions and display cytotoxic effects towards human cells38. This effect is not observed in the case of capric acid, which does not display cytotoxicity towards human cell lines34.
Mislocalization of Cdr1p after treatment with FLC-capric acid combination was associated with reduced ergosterol concentration in the PM of C. albicans (Table 1). A similar effect was observed in ergosterol-depleted C. albicans erg11Δ/Δ strains, in which Cdr1 delocalized into vacuoles22. Furthermore, we had previously described that reduced ergosterol levels due to FLC treatment leads to mislocalization of Cdr122,23. In our study, upon either FLC or FLC-capric acid treatment, we observed simultaneous mislocalization of Cdr1p (Fig. 2C) and reduced ergosterol concentration (Table 1). Inhibition of ergosterol biosynthesis most likely occurred at the step of lanosterol demethylation, catalyzed by Erg11p, due to higher content of lanosterol in above mentioned cases (Table 1). Interestingly, in the case of treating C. albicans with capric acid alone, we observed high overproduction of ergosterol and rigidification of PM. Such an effect is commonly described in the case of Candida spp. strains resistant towards AMB12. Therefore, the presence of capric acid was found to almost completely reduce the activity of AMB against C. albicans CAF2-1 (Fig. 1C). This is especially important as in clinical practice, apart from looking for additives that would enhance the activity of antifungal drugs, attention is also paid to possible antagonistic interactions. A known example is the interaction of FLC with the anti-cancer drug vincristine, which leads to a neurotoxic effect in patients21. The use of FLC during therapy with drugs whose metabolism is mediated by the human cytochrome P-450 is avoided20. On the other hand, AMB was reported to display antagonistic effects when administered with immunosuppressants (such as cyclosporin A) or diuretics (such as furosemide)20.
Free FAs have been previously reported to directly interact with AMB and inhibit its antifungal activity39. Such a relationship has been described so far for myristic, palmitic and stearic acids39. To eliminate the effect of a potential direct interaction of capric acid with AMB, C. albicans cells were first cultured in the presence of capric acid to an early exponential growth phase of growth and then treated with AMB. It was found that AMB did not cause membrane permeabilization or delocalization of the Cdr1p in C. albicans CAF2-1 cells if grown in presence of capric acid (Fig. 4B, C). At this stage of the study, it was concluded that the reduced PM permeabilization occurs despite complete binding of AMB to C. albicans cells, regardless if cultured with or without capric acid (Fig. 4A). Additionally, lack of differences in extracellular K+ concentration after AMB treatment suggests reduced cytoplasm leakage as a result of AMB treatment. This concludes that, AMB should not be administered among dietary supplements containing capric acid or S. boulardii cells.
Conclusions
Capric acid, produced by S. boulardii probiotic yeast, may be a promising additive for use in combination with azole drugs. Capric acid in the combination with FLC has led to reduced efflux activity of Cdr1p as a result of its relocalization from the PM to the interior of the cell. Due to the crucial role of ergosterol in Cdr1p PM localization, the above effect occurred due to inhibitory effect of FLC-capric acid combination of ergosterol biosynthesis. On the other hand, capric acid alone stimulated ergosterol production in C. albicans, which in turn generated cross resistance towards AMB and inhibited its toxic mechanisms towards C. albicans cells.
Materials and methods
Chemicals
Chemicals and reagents were purchased from the following suppliers: capric acid, fluconazole (FLC), conventional amphotericin B (AMB), 2-deoxy-d-glucose, rhodamine 6G (R6G), laurdan, β-mercaptoethanol (BME), ethylenediaminetetraacetic acid (EDTA), cholesterol, and BSTFA/TMCS (N,O-bis(trimethylsilyl) trifluoroacetamide/trimethylchlorosilane) were purchased from Sigma-Aldrich (St. Louis, MO, USA); D-glucose, bacteriological agar, propidium iodide (PI), zymolyase, D-sorbitol were purchased from Bioshop (Ontario, Canada); peptone and yeast extract (YE) were purchased from Becton Dickinson (Franklin Lakes, NJ, USA); chloroform and methanol were purchased from Chempur (Bangalore, India); Total RNA Mini Kit was purchased from A&A Biotechnology (Gdynia, Poland). All chemicals were high purity grade.
Strains and growth conditions
The C. albicans strains used in this study are listed in Table 2. CAF2-1 was a gift from Professor D. Sanglard (Lausanne, Switzerland)40. KS028 and KS052 were previously generated by our team22,23. Strains were routinely pre-grown at 28 °C on YPD medium (1% YE, 1% peptone, 2% glucose) in a shaking incubator (120 rpm)22,23. Agar in a final concentration of 2% was used for medium solidification22,23.
Table 2.
C. albicans strains used in the study.
For most of the experiments, cells were grown in 20 mL of YPD medium, at 28 °C, shaking at 120 rpm with a starting A600 = 0.122,23. Cells were incubated with not treatment, or with capric acid, FLC, or the combination of both drugs added at t = 0 h until they reached the early logarithmic phase, typically after 8 h. Cells were then centrifuged at 4,500 rpm for 5 min, washed twice with either phosphate-buffered saline (PBS), 50 mM HEPES–NaOH buffer (pH 7.0) or 0.9% saline, and resuspended in either PBS, HEPES–NaOH or 0.9% saline to the target A600.
Phenotypic tests
Experiments were conducted as previously described22. Briefly, CAF2-1 and KS028 strain suspensions (PBS; A600 = 0.7; prepared from overnight YPD cultures) were serially diluted up until 10−3. Of each dilution, from 100 to 10−3, 2 µL were spotted onto YPD agar with or without 22.7, 45.3 and 90.6 µg/mL of capric acid. After cultivation for 48 h at 28 °C, the plates were photographed using a FastGene B/G GelPic imaging box (Nippon Genetics, Tokyo, Japan).
Determination of growth in the presence of antibiotics
Experiments were performed in compliance with the Clinical and Laboratory Standards Institute (2008), 3rd ed. M27-A3 with described modifications24. Briefly, the growth was determined by serially diluting FLC, AMB, or a combination of each of the drug with capric acid in YPD medium using 96-well sterile plates (Sarstedt; Stare Babice, Poland) and then inoculated with CAF2-1 cells at a final A600 of 0.01. After incubating at 28 °C for 24 h, A600 was measured (ASYS UVM 340, Biogenet). The percentage of growth was determined by normalizing A600 in the control experiments (without antimicrobial agents) as 100%.
Microscopic studies of Cdr1-GFP
The strain KS052, grown in the presence of FLC, capric acid, or combination of both drugs was resuspended in PBS, concentrated and observed under a Zeiss Axio Imager A2 microscope equipped with a Zeiss Axiocam 503 mono microscope camera and a Zeiss HBO100 mercury lamp (Carl Zeiss AG, Oberkochen, Germany).
R6G efflux assay
Suspensions of CAF2-1, in 25 mL HEPES–NaOH; (A600 = 1.0) grown in the presence of FLC, capric acid, or combination of both compounds were treated with 2-deoxy-D-glucose and stained with R6G according to the protocol previously described by Szczepaniak et al.27 In each of the conditions, the R6G uptake was estimated to be ≥ 95%. Fluorescence intensities (IFs) were collected 15 min after R6G efflux initiation and normalized to 1 for the efflux activity of untreated CAF2-1 cells.
Real time polymerase chain reaction
RNA was isolated from the CAF2-1 grown in the presence of FLC, capric acid, or combination of both drugs (PBS; A600 = 10) using the Total RNA Mini Kit (A&A Biotechnology). Synthesis of cDNA and calculation of gene expression levels were performed as previously described41. The following gene-specific primers were used: ACT1F (5′-TCCAGCTTTCTACGTTTCCA-3′), ACT1R (5′-GTCAAGTCTCTACCAGCCAA-3′), CDR1F (5′-TTTAGCCAGAACTTTCACTCATGATT-3′), CDR1R (5′-TATTTATTTCTTCATGTTCATATGGATTGA-3′), ERG11F (5′-TTTGGTGGTGGTAGACATA-3′), ERG11R (5′-GAACTATAATCAGGGTCAGG-3′).
Isolation of plasma membranes (PMs) and sterol analysis
PMs were isolated from suspensions of CAF2-1 grown in the presence of FLC, capric acid, or combination of both compounds (PBS; concentrated to A600 = 20) according to the previously reported method23. Briefly, cells were resuspended in lysis medium (1 M sorbitol, 0.1 M EDTA, 1% BME, 3 mg/mL zymolyase) and incubated at 37 °C for 30 min. Protoplasts were then washed with 1.2 M sorbitol, lysed by ice-cold H2Odd shock and disrupted by sonication (5-s cycles for 2 min at 4 °C) using an ultrasonic processor (SONICS Vibra-cell VCX 130). Cell lysate was centrifuged at 10,000 rpm; at 4 °C for 10 min to remove unbroken material, and the supernatant was ultracentrifuged at 100,000 rpm at 4 °C for 60 min using a Micro Ultracentrifuge CS150FNX (Hitachi; Tokyo, Japan). The crude PM pellets were suspended in a chloroform–methanol solution (1:2 v/v). The chloroform layer was concentrated using nitrogen gas after vigorous stirring at 4 °C for 16 h. PM fractions were derivatizated with BSTFA-TMCS and sterol analysis was performed by gas chromatography mass spectrometry (GC–MS) with cholesterol as an internal standard, following previously described protocol22,23.
Membrane fluidity assessment
The membrane fluidity assay was performed according to previously described protocol22, with modifications. Briefly, CAF2-1 were incubated for 8 h with 22.7, 45.3 and 90.6 µg/mL of capric acid in YPD. The cells were then centrifuged at 4,500 rpm for 5 min), washed twice with PBS and resuspended in 3 mL PBS (A600 = 0.1). Suspensions were labeled with Laurdan at a final concentration of 5 × 10−6 M for 20 min at 25 °C in darkness. The probes were excited at 366 nm (Ex slit = 10 nm), and fluorescence spectra were recorded at 400–550 nm (Em slit = 2.5 nm) (PMT voltage = 400 V) using a fluorescence spectrophotometer equipped with a xenon lamp (HITACHI F-4500, Hitachi, Tokyo, Japan). For data analyses, modified general polarization (GP) was calculated as follows: the difference of the sum of IFs from 425 to 450 nm and the sum from 475 to 525 nm, divided by the sum of IFs from 425 to 450 nm and from 475 to 525 nm.
Assessment of plasma membrane (PM) permeability
To assess the effects of AMB on CAF2-1 PM permeability we followed the PI assay42, with modifications. Briefly, suspensions of CAF2-1 grown with or without capric acid (0.9% saline; A600 = 1) were exposed to 2 µg/mL AMB for 1 h at 28 °C, pelleted, washed twice with 0.9% saline and resuspended in 0.9% saline with 6 µM PI for 5 min. The cells were then pelleted by centrifugation, washed twice with 0.9% saline, concentrated and observed under a Zeiss Axio Imager A2 microscope equipped with a Zeiss Axiocam 503 mono microscope camera and a Zeiss HBO100 mercury lamp. The percentage of permeabilization was evaluated by counting PI positive cells out of at least 100 cells in each replicate.
Cytoplasmic potassium ions (K+) leakage
To assess the effects of AMB on CAF2-1, cytoplasm leakage suspensions of CAF2-1 grown with or without capric acid (0.9% saline; A600 = 1) were exposed to 2 µg/mL AMB for 1 h at 28 °C and pelleted. The supernatants were evaluated for the K+ concentrations using inductively coupled plasma atomic emission spectroscopy (ICP-AES).
Determination of AMB binding to cells
The assay was performed according to Skwarecki et al.32, with modifications. Briefly, suspensions of CAF2-1 grown with or without capric acid (0.9% saline; A600 = 1) were exposed to 2 µg/mL AMB for 1 h at 28 °C and pelleted. A409 of the supernatants was determined using ASYS UVM 340 Biogenet. The percentage of AMB binding was evaluated in accordance with 2 µg/mL AMB in 0.9% saline (100%) and supernatant of AMB untreated cells (0%).
Statistical analysis
At least three independent replicates were performed for each experiment. Statistical significance was determined using Student’s t test (binomial, unpaired).
Supplementary Information
Acknowledgements
This work was supported by the National Science Center, Poland, NCN Grant: 2016/23/B/NZ1/01928.
Author contributions
Conceptualization, J.S. and A.K.; methodology and investigation, J.S., D.D., P.B.; writing—original draft preparation, J.S. and D.D.; writing—review and editing, funding and supervision, A.K. All authors have read and agreed to the published version of the manuscript.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
The online version contains supplementary material available at 10.1038/s41598-021-86012-9.
References
- 1.Puel A, et al. Chronic mucocutaneous candidiasis in humans with inborn errors of interleukin-17 immunity. Science (80-) 2011;332:65–68. doi: 10.1126/science.1200439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Whaley SG, et al. Azole antifungal resistance in Candida albicans and emerging non-albicans Candida Species. Front. Microbiol. 2017;7:2173. doi: 10.3389/fmicb.2016.02173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dadar M, et al. Candida albicans - Biology, molecular characterization, pathogenicity, and advances in diagnosis and control – An update. Microb. Pathog. 2018;117:128–138. doi: 10.1016/j.micpath.2018.02.028. [DOI] [PubMed] [Google Scholar]
- 4.Alizadeh F, Khodavandi A, Zalakian S. Quantitation of ergosterol content and gene expression profile of ERG11 gene in fluconazole-resistant Candida albicans. Curr. Med. Mycol. 2017;3:13–19. doi: 10.29252/cmm.3.1.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Beardsley J, Halliday CL, Chen SCA, Sorrell TC. Responding to the emergence of antifungal drug resistance: perspectives from the bench and the bedside. Future Microbiol. 2018;13:1175–1191. doi: 10.2217/fmb-2018-0059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ortiz C, Torres R. Antifungal resistance and its evolution: an increasing concern. Adv. Biotechnol. Microbiol. 2018;10:555777. doi: 10.19080/AIBM.2018.10.555777. [DOI] [Google Scholar]
- 7.Wiederhold NP. Antifungal resistance: current trends and future strategies to combat. Infect. Drug Resist. 2017;10:249–259. doi: 10.2147/IDR.S124918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wu YQ, Gao N, Li C, Gao J, Ying CM. A newly identified amino acid substitution T123I in the 14α-demethylase (Erg11p) of Candida albicans confers azole resistance. FEMS Yeast Res. 2017;17:1–6. doi: 10.1093/femsyr/fox012. [DOI] [PubMed] [Google Scholar]
- 9.Paul S, Moye-Rowley WS. Multidrug resistance in fungi: regulation of transporter-encoding gene expression. Front. Physiol. 2014;5:143. doi: 10.3389/fphys.2014.00143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gray KC, et al. Amphotericin primarily kills yeast by simply binding ergosterol. Proc. Natl. Acad. Sci. U. S. A. 2012;109:2234–2239. doi: 10.1073/pnas.1117280109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gogtay NJ, Kulkarni UP, Panchabhai TS. Adverse reactions to antifungal agents. Adverse Drug React. Bull. 2008;2:963–966. doi: 10.1097/FAD.0b013e32831844cb. [DOI] [Google Scholar]
- 12.Santos GCDO, et al. Candida infections and therapeutic strategies: Mechanisms of action for traditional and alternative agents. Front. Microbiol. 2018;9:1351. doi: 10.3389/fmicb.2018.01351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hamilton-Miller JM. Sterols from polyene-resistant mutants of Candida albicans. J. Gen. Microbiol. 1972;73:201–203. doi: 10.1099/00221287-73-1-201. [DOI] [PubMed] [Google Scholar]
- 14.Salci TP, Negri M, Abadio AKR, Svidzinski TIE, Kioshima ÉS. Targeting Candida spp. to develop antifungal agents. Drug Discov. Today. 2018;23:802–814. doi: 10.1016/j.drudis.2018.01.003. [DOI] [PubMed] [Google Scholar]
- 15.Altieri C, Cardillo D, Bevilacqua A, Sinigaglia M. Inhibition of Aspergillus spp. and Penicillium spp. by fatty acids and their monoglycerides. J. Food Prot. 2007;70:1206–1212. doi: 10.4315/0362-028X-70.5.1206. [DOI] [PubMed] [Google Scholar]
- 16.Carballeira NM. New advances in fatty acids as antimalarial, antimycobacterial and antifungal agents. Prog. Lipid Res. 2008;47:50–61. doi: 10.1016/j.plipres.2007.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pohl CH, Kock JLF, Thibane VS. Antifungal free fatty acids: a review. Sci. Against Microb. Pathog. Curr. Res. Technol. Adv. 2011;1:61–71. [Google Scholar]
- 18.Murzyn A, Krasowska A, Stefanowicz P, Dziadkowiec D, Łukaszewicz M. Capric acid secreted by S. boulardii inhibits C. albicans filamentous growth, adhesion and biofilm formation. PLoS ONE. 2010;5:2–8. doi: 10.1371/journal.pone.0012050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bergsson G, Arnfinnsson J, Steingrímsson Ó, Thormar H. In vitro killing of Candida albicans by fatty acids and monoglycerides. Antimicrob. Agents Chemother. 2001;45:3209–3212. doi: 10.1128/AAC.45.11.3209-3212.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Depont F, et al. Drug–drug interactions with systemic antifungals in clinical practice. Pharmacoepidemiol. Drug Saf. 2007;16:1227–1233. doi: 10.1002/pds.1473. [DOI] [PubMed] [Google Scholar]
- 21.Cornely OA, Kontoyiannis DP. How to prophylax against invasive fungal infections in adult ALL? An unmet need. Mycoses. 2018;61:646–649. doi: 10.1111/myc.12786. [DOI] [PubMed] [Google Scholar]
- 22.Suchodolski J, Muraszko J, Bernat P, Krasowska A. A crucial role for ergosterol in plasma membrane composition, localisation, and activity of Cdr1p and H + -ATPase in Candida albicans. Microorganisms. 2019;7:378. doi: 10.3390/microorganisms7100378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Suchodolski J, Muraszko J, Korba A, Bernat P, Krasowska A. Lipid composition and cell surface hydrophobicity of Candida albicans influence the efficacy of fluconazole-gentamicin treatment. Yeast. 2020;37:117–129. doi: 10.1002/yea.3455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Suchodolski J, Derkacz D, Muraszko J, Panek JJ, Jezierska A. Fluconazole and lipopeptide surfactin interplay during Candida albicans plasma membrane and cell wall remodeling increases fungal immune system exposure. Pharmaceutics. 2020;12:314. doi: 10.3390/pharmaceutics12040314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Prasad R, Nair R, Banerjee A. Multidrug transporters of Candida species in clinical azole resistance. Fungal Genet. Biol. 2019;132:E103252. doi: 10.1016/j.fgb.2019.103252. [DOI] [PubMed] [Google Scholar]
- 26.Prasad R, Balzi E, Banerjee A, Khandelwal NK. All about CDR transporters: Past, present, and future. Yeast. 2019;36:223–233. doi: 10.1002/yea.3356. [DOI] [PubMed] [Google Scholar]
- 27.Szczepaniak J, Cieślik W, Romanowicz A, Musioł R, Krasowska A. Blocking and dislocation of Candida albicans Cdr1p transporter by styrylquinolines. Int. J. Antimicrob. Agents. 2017;50:171–176. doi: 10.1016/j.ijantimicag.2017.01.044. [DOI] [PubMed] [Google Scholar]
- 28.Mukherjee PK, Chandra J, Kuhn DM, Ghannoum MA. Mechanism of fluconazole resistance in Candida albicans biofilms: phase-specific role of efflux pumps and membrane sterols. Infect. Immun. 2003;71:4333–4340. doi: 10.1128/IAI.71.8.4333-4340.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Flowers SA, Colón B, Whaley SG, Schuler MA, David Rogers P. Contribution of clinically derived mutations in ERG11 to azole resistance in Candida albicans. Antimicrob. Agents Chemother. 2015;59:450–460. doi: 10.1128/AAC.03470-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wu HC, et al. Preparation of Amphotericin B-Ergosterol structures and molecular simulation of water adsorption and diffusion. J. Memb. Sci. 2018;545:229–239. doi: 10.1016/j.memsci.2017.09.032. [DOI] [Google Scholar]
- 31.Wu HC, et al. Molecular simulation of a modified amphotericin B-Ergosterol artificial water channel to evaluate structure and water molecule transport performance. J. Memb. Sci. 2019;583:49–58. doi: 10.1016/j.memsci.2019.04.021. [DOI] [Google Scholar]
- 32.Skwarecki AS, et al. Molecular umbrellas modulate the selective toxicity of polyene macrolide antifungals. Bioconjug. Chem. 2018;29:1454–1465. doi: 10.1021/acs.bioconjchem.8b00136. [DOI] [PubMed] [Google Scholar]
- 33.Jóźwiak M, Filipowska A, Fiorino F, Struga M. Anticancer activities of fatty acids and their heterocyclic derivatives. Eur. J. Pharmacol. 2020;871:172937. doi: 10.1016/j.ejphar.2020.172937. [DOI] [PubMed] [Google Scholar]
- 34.Narayanan A, Baskaran SA, Amalaradjou MAR, Venkitanarayanan K. Anticarcinogenic properties of medium chain fatty acids on human colorectal, skin and breast cancer cells in vitro. Int. J. Mol. Sci. 2015;16:5014–5027. doi: 10.3390/ijms16035014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Avis TJ, Bélanger RR. Specificity and mode of action of the antifungal fatty acid cis-9-heptadecenoic acid produced by Pseudozyma flocculosa. Appl. Environ. Microbiol. 2001;67:956–960. doi: 10.1128/AEM.67.2.956-960.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.McDonough V, Stukey J, Cavanagh T. Mutations in erg4 affect the sensitivity of Saccharomyces cerevisiae to medium-chain fatty acids. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2002;1581:109–118. doi: 10.1016/S1388-1981(02)00127-0. [DOI] [PubMed] [Google Scholar]
- 37.Szczepaniak J, Lukaszewicz M, Krasowska A. Detection of inhibitors of Candida albicans Cdr transporters using a diS-C3(3) fluorescence. Front. Microbiol. 2015;6:E176. doi: 10.3389/fmicb.2015.00176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tonshin AA, Teplova VV, Andersson MA, Salkinoja-Salonen MS. The Fusarium mycotoxins enniatins and beauvericin cause mitochondrial dysfunction by affecting the mitochondrial volume regulation, oxidative phosphorylation and ion homeostasis. Toxicology. 2010;276:49–57. doi: 10.1016/j.tox.2010.07.001. [DOI] [PubMed] [Google Scholar]
- 39.Iannitelli RC, Ikawa M. Effect of fatty acids on action of polyene antibiotics. Antimicrob. Agents Chemother. 1980;17:861–864. doi: 10.1128/AAC.17.5.861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Fonzi WA, Irwin MY. Isogenic strain construction and gene mapping in Candida albicans. Genetis. 1993;134:717–728. doi: 10.1093/genetics/134.3.717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Szczepaniak J, Łukaszewicz M, Krasowska A. Estimation of Candida albicans ABC transporter behavior in real-time via fluorescence. Front. Microbiol. 2015;6:E1382. doi: 10.3389/fmicb.2015.01382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Suchodolski J, Feder-Kubis J, Krasowska A. Antifungal activity of ionic liquids based on (−)-menthol: a mechanism study. Microbiol. Res. 2017;197:56–64. doi: 10.1016/j.micres.2016.12.008. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.