

Abstract
Ecosystems integrity and services are threatened by anthropogenic global changes. Mitigating and adapting to these changes require knowledge of ecosystem functioning in the expected novel environments, informed in large part through experimentation and modelling. This paper describes 13 advanced controlled environment facilities for experimental ecosystem studies, herein termed ecotrons, open to the international community. Ecotrons enable simulation of a wide range of natural environmental conditions in replicated and independent experimental units while measuring various ecosystem processes. This capacity to realistically control ecosystem environments is used to emulate a variety of climatic scenarios and soil conditions, in natural sunlight or through broad‐spectrum lighting. The use of large ecosystem samples, intact or reconstructed, minimizes border effects and increases biological and physical complexity. Measurements of concentrations of greenhouse trace gases as well as their net exchange between the ecosystem and the atmosphere are performed in most ecotrons, often quasi continuously. The flow of matter is often tracked with the use of stable isotope tracers of carbon and other elements. Equipment is available for measurements of soil water status as well as root and canopy growth. The experiments ran so far emphasize the diversity of the hosted research. Half of them concern global changes, often with a manipulation of more than one driver. About a quarter deal with the impact of biodiversity loss on ecosystem functioning and one quarter with ecosystem or plant physiology. We discuss how the methodology for environmental simulation and process measurements, especially in soil, can be improved and stress the need to establish stronger links with modelling in future projects. These developments will enable further improvements in mechanistic understanding and predictive capacity of ecotron research which will play, in complementarity with field experimentation and monitoring, a crucial role in exploring the ecosystem consequences of environmental changes.
Keywords: biodiversity, controlled environment facilities, ecosystem functioning, ecosystem process measurements, environmental simulations, experimentation, global change, research infrastructures
Experimentation and modelling are necessary to predict ecosystem functioning under future environments and to develop mitigating and adapting strategies. This paper describes 13 advanced controlled environment facilities, called ecotrons, open to the international community. An ecotron comprises a set of replicated enclosures designed to host ecosystems samples and enable realistic simulations of above‐ and belowground environmental conditions, while simultaneously and automatically measuring ecosystem processes. The characteristics of these infrastructures are given as well as examples of collaborative projects hosted so far.

1. INTRODUCTION
In the face of rapid climate change and biodiversity loss, the goods and services provided by ecosystems are under increasing threat (Pecl et al., 2017; Scheffers et al., 2016), and securing their future delivery is one of today's most pressing challenges (Arneth et al., 2019; Challinor et al., 2017; Díaz et al., 2019; Wheeler & Von Braun, 2013). To do so, we need a better understanding of the fundamental processes underpinning ecosystem functions and services and of how these processes will be altered in the novel environments of the future. This understanding will foster the development of mitigating management strategies through innovation and adaptation. Ecosystem science is developing at a fast rate, taking advantage of progress in other scientific disciplines (e.g. genomics, metabolomics, phenomics, spectronomics, etc.) and of development of new technologies (e.g. metabarcoding, new laser gas/isotope analysers, high‐resolution proximate and remote sensing, etc.). What is now expected from ecosystem science is a stronger adoption of interdisciplinary approaches connecting theory, experiments, field observations, modelling and simulation to address pressing questions on the future of ecosystems and societal welfare (Hanson & Walker, 2020; Mauser et al., 2013) and the complexity of biodiversity–ecosystem feedbacks (Abiven et al., 2017).
In this context, controlled environment facilities (CEFs), such as growth chambers and advanced greenhouses, have become standard tools to simulate different environmental conditions and disentangle their influences on ecosystem functioning. These have been used, for example, to reveal the underlying mechanisms of observed overall responses, for model parametrization and for theory testing (Clobert et al., 2018; Hanson & Walker, 2020; Kreyling et al., 2014). CEFs have been steadily improved through the use of better lighting systems, the regulation of additional parameters, such as CO2 and ozone, user‐friendly computerized environmental control and the possibility of remote operation and security checks. During the last three decades, however, a more innovative step forward has been achieved through the development of a more heavily instrumented type of CEF: herein termed ecotrons. We define an ecotron as an experimental facility comprising a set of replicated enclosures designed to host ecosystems samples, enabling realistic simulations of aboveground and belowground environmental conditions while automatically measuring ecosystem processes. Therefore, ecotrons provide continuous information on ecosystem functioning (fluxes of energy and matter).
This principle of using enclosures (a lysimeter for the soil and an aerial compartment around the canopy) for simultaneous environmental control and process measurement has been pioneered, at the canopy level, by field physiologists as early as the 1930s (Thomas & Hill, 1937), but most of these sunlit facilities were developed from the 1960s (Liu et al., 2000). Starting with the München ExpoSCREEN (Payer et al., 1986), the Imperial College ecotron in Silwood Park (Lawton, 1993) and the Desert Research Institute EcoCELLs in Reno (Griffin et al., 1996), larger permanent infrastructures, open to national and international collaboration, were constructed. Four ecotrons were built between 1985 and 2006, eight between 2010 and 2020 and two more are under construction. A more thorough historical background with the etymology of the word ecotron is provided in the Supporting Information file ‘Ecotron‐related facilities’. These ecotrons can be seen as a new means of performing ecological research through centralized, shared and heavily instrumented research facilities mirroring practices in other disciplines such as astronomy and physics (Granjou & Walker, 2016; Rineau et al., 2019).
This paper reviews the characteristics of existing ecotrons (or ecotrons under construction), focusing on their environmental control capacities and the design and technology underpinning ecosystem process measurements. Since most of them are open to national and international collaboration, we also outline the advantages and prospects of using the listed ecotrons. An analysis of the experiments conducted so far reveals the large range of research topics that can be addressed in these infrastructures, but we also address their limitations, emphasizing the necessary complementarity between ecotrons and other experimental or observational facilities for the pursuit of predicting, mitigating and adapting to ongoing global environmental changes. Finally, we discuss the perspectives on the future development of ecotrons and the need to combine their experiments with modelling efforts.
2. THE FEATURES OF ECOTRONS
Advantages of ecotrons are increasingly acknowledged by the scientific community and funding bodies, as indicated by the growing investment of research institutes and universities in such new facilities. However, part of the scientific community is not up to date with the more recently developed features and is not fully aware of the advantages and trade‐offs of ecotron experiments relative to greenhouse or field experimental approaches. Throughout this section, we discuss the features of ecotrons showing that in addition to developing specific technologies, their strength is to cumulate many of the advantages found in some of these other experimental facilities.
2.1. More realistic experiments, across a broad range of environmental conditions
Ecosystem experiments can be conducted in settings that vary in realism, environmental control and replication of experimental units, along previously described trade‐offs (De Boeck et al., 2015; Diaz et al., 2003; Stewart et al., 2013). Here we define realism as providing conditions as close as possible to the complexity of natural environments, whether in the past, present or predicted future. This implies the capability to simulate natural ranges, dynamics and combinations of abiotic and biotic variables. Especially in global change research, realism also includes the need to impose experimental treatments going beyond the historical record (Hanson & Walker, 2020), and even beyond the current model‐predicted climate change scenarios (De Boeck et al., 2020).
Generally, confined ecosystems have a reduced spatial and biological complexity, are surrounded by walls and have modified physics (e.g. energy exchange), compared with natural ecosystems. Ecotrons deal with these issues via a series of features that set them apart from typical growth chambers and which render the experimental conditions closer to field conditions. One such feature is using large ecosystem samples hosted in large enclosed atmospheric volumes (see details on the features in Section 3), thus incorporating more aboveground and belowground biological complexity and spatial heterogeneity. Whenever possible and suitable, intact soil monoliths are extracted in situ and inserted in the ecotron enclosures, thus preserving soil physicochemical properties, soil biota and vegetation. Getting closer to realistic outdoor conditions is important, as there is accumulating evidence that the use of small and simplified systems such as pots or small containers brings identified biases (Poorter et al., 2012) or unidentified laboratory‐specific artefacts (e.g. Massonnet et al., 2010; Milcu et al., 2018) that may generate results with less external validity (i.e. results which can be generalized with less confidence; Poorter et al., 2016).
Another feature that improves experimental realism is the capacity of most ecotrons to simulate a wider range of environmental parameters than is usually the case in growth chambers, or in some of the earlier ecotrons (e.g. Silwood Park ecotron; Lawton et al., 1993). The following conditions can be reached, although not in all facilities (details in the section referred to above): freezing or near‐freezing air temperatures maybe achieved through the use of refrigeration, such as compressed gas expansion within the air circuit; very low air relative humidity achieved by injecting dry air; replicating the in situ soil matrix potential at the bottom of the soil column that affects evapotranspiration (Groh et al., 2016) and replicating the in situ soil temperature gradient that affects soil respiration as well as plant growth (Füllner et al., 2012). To improve realism, some ecotrons take advantage of natural sunlight while only reducing light intensity to a minor extent including in the UV range, while others combine light‐emitting diodes (LED) or a mixture of metal halide/quartz halogen lamps and fluorescent tubes (as cited in Ghirardo et al., 2020) to achieve a radiation spectrum approaching that of the sun. Taken together, these features considerably step up the realism of environmental control and allow for the simulation of past or future environmental conditions and climatic extremes with improved accuracy and precision. As a result of the realism of the simulated environmental conditions combined with the incorporation of more aboveground and belowground complexity, ecotron experiments are often much closer to field experiments than most typical CEF experiments.
2.2. Disentangling ecosystem mechanisms through confinement and replication
The ecotrons' capacity to independently manipulate biotic as well as abiotic variables comes with many advantages, the most important one being the disentanglement of the ecological effects of variables that often co‐vary in natural settings. For example, drought and co‐varying factors such as temperature, vapour pressure deficit and sunlight (De Boeck & Verbeeck, 2011), or atmospheric CO2 and its effect on leaf and canopy temperature (Leuzinger & Körner, 2007), can be independently controlled and their impacts untwined. Similarly, treatments that manipulate soil biota presence and diversity can be relatively easily established in ecotrons, a manipulation challenging in the field without disturbing the ecosystem and risking contamination from the surroundings. Combining soil sterilization techniques and subsequent inoculation of specific species or groups of species, ecotrons are a powerful tool for exploring the effects of specific biota on ecosystems (Bradford et al., 2002). Another overlooked feature of ecotrons is their inherent capacity to incorporate environment‐biotic feedbacks as well as the possibility to impose feedbacks as experimental treatments. For the first aspect, we emphasize the fact that the large size of the experimental systems will inherently incorporate more of the natural biological diversity of the model system. Hence, more key taxa that will be present and will realistically respond to the experimental treatments and feedback on soil properties or other taxa, a response likely less to occur with smaller pot‐size systems. For the second aspect, while some of these feedbacks can be performed in the field and in classical CEFs (e.g. plant–soil biota feedbacks; Van der Putten et al., 2013), other feedbacks require greater control over the environmental variables. For example, using materially closed systems in the former Silwood Park ecotron, Milcu et al. (2012) established CO2‐temperature feedback treatments in a simplified physical model of the terrestrial C cycle. The temperature of the experimental systems was continuously adjusted depending on the emerging CO2 concentration of the units (using the most likely CO2‐temperature sensitivity) resulting from the combination of simulated anthropogenic emissions, photosynthesis and plant and soil respiration.
Disentangling the ecological effects of different variables requires multiple identical and independently controlled experimental units. Lack of replication at the unit level can lead to biased parameter estimates (Porter et al., 2015), because any confounding chamber effect is not taken into account (Potvin & Tardif, 1988). Given that the high construction cost can limit the number of ecotron experimental units, several facilities opted for a minimum of 12 units, since it allows the establishment of treatments with 6, 4 and 3 replicates per treatment combination for experimental designs with 2, 3 and 4 treatment combinations, respectively. Such a series of experimental units also suits gradient experiments with many different, un‐replicated treatment levels, to which a regression‐type analysis is applied instead of an analysis of variance requiring replication. Gradient experiment is an underused methodology appropriate for identifying thresholds, tipping points and response functions (Kreyling et al., 2018).
2.3. A major focus on measurements of ecosystem processes
Next to the capacity to simulate and measure multiple environmental conditions, the most compelling characteristic of ecotrons is their focus on non‐destructive, automatic, real‐time measurements of ecosystem‐level processes. Some ecotrons are specifically designed to use the confinement of the ecosystem as large gas exchange chambers (e.g. Barton et al., 2010; Milcu et al., 2016), analogous to leaf chambers in portable photosynthesis systems. This allows the measurement of the net ecosystem exchange (NEE) between the terrestrial compartment and the atmosphere for various molecules (CO2, N2O, CH4, H2O, O3, NOx, volatile organic compounds [VOCs]) using either a static non‐steady state or a dynamic (flow‐through) steady‐state approach. Both approaches are feasible but they require that one single ecosystem is hosted within each chamber, in stark contrast with CEFs where different model systems (set‐ups in pots or containers) are incubated side by side. These NEE rates are measured at high frequency (every 10–20 min), capturing both the short‐term and the cumulative long‐term responses of elemental budgets with a high degree of confidence (e.g. Roy et al., 2016). Ecotrons offer the possibility to balance all fluxes: the inflow, stock and outflow can be precisely quantified for most components of the soil/plant/atmosphere system, including energy. Furthermore, the recent availability of multi‐gas and multi‐isotopologue laser analysers enables simultaneous measurements of the molecules listed above, as well as their isotopologues (e.g. Braden‐Behrens et al., 2017; Braendholt et al., 2019; McManus et al., 2015). Examples of processes estimated by the measurements of isotopic fractionation and isotopomers include canopy conductance and respiration as well as the coupling of CO2 and H2O cycles via δ13C and δ18O of CO2 (Harwood et al., 1999), nitrification and denitrification processes via δ15N, δ18O and isotopomers of 15N, that is, the 15N site preference (SP) in N2O (Baggs, 2008; Butterbach‐Bahl et al., 2013), and tracing ecosystem water fluxes and disentangling evapotranspiration via δ2H, δ18O of H2O (liquid and vapour; Oerter & Bowen, 2017).
Other ecotrons focus on automatic measurements of ecosystem properties that are not related to ecosystem gas exchange. These include root growth using minirhizotrons (Möller et al., 2019), invertebrate and plant community composition using novel imaging techniques like computerized trap systems, video cameras or radio frequency identification (Dell et al., 2014; Dombos et al., 2017), thermography to analyse the heterogeneity of transpiration, and hyperspectral reflectance for canopy biomass and chemical content (Tan et al., 2018; Xie et al., 2020). Similar to what is done in other experimental settings, in all ecotron experiments the automatic measurements are complemented by low‐frequency samplings of soil, plants, soil solution, leachate, etc., for further analysis of fauna and microbe diversity, elemental and isotopic composition of soil, water and plant material, delivering a more complete understanding of the impact of the experimental treatments. These complementary analyses are generally performed by the hosted teams and are often not the responsibility of the ecotron facility.
2.4. Experimental flexibility
While generally costly to build, run and maintain, ecotrons offer significant experimental flexibility. First, they can host many different types of treatments (climate, atmospheric composition and pressure, pollution, soil types, trophic levels, biodiversity within trophic levels, ecosystem management, etc.). For some of these treatments which are outside of the range of current environmental conditions, specific regulations have been installed, for example decreased CO2 in air using scrubbing molecular sieves to reach pre‐industrial CO2 concentrations; ozone fumigation produced by an ozone generator plugged on pure oxygen gas bottles; low oxygen concentration through dilution with nitrogen and simultaneous readjustment of CO2 concentration. Furthermore, unlike field facilities which are bound to a specific ecosystem, in ecotrons the targeted model system can change from one experiment to the other, ranging from agricultural systems to grasslands, peatlands, shrublands and regenerating forest (saplings), essentially any ecosystem type where plant stature fits the height of the units. However, the flexibility to host various ecosystem types in consecutive experiments often trades off against their duration. One solution is to bring ecosystem samples extracted from long‐term field experiments to the ecotron for short‐term, more thorough, physiological measurements (i.e. use the ecotron as an ecosystem analyser, e.g. Milcu et al., 2014) or for applying complementary treatments. Ecotrons are often also flexible in terms of dimensions. The size of the lysimeters and the height of the canopy enclosures can often be tuned to the particular experiment. In some ecotrons, the main experimental unit can be divided into subunits to vary their connectivity, thus allowing the study of spatial and meta‐population dynamics (Eisenhauer & Türke, 2018). Some ecotron platforms can work with either sunlight or artificial light (e.g. Resco de Dios et al., 2016). Another element of flexibility is the option for hosted research teams to temporarily install supplementary costly instruments in the ecotron air circuits, such as VOCs or NOx analysers to bring added value by answering additional questions. The length of the experiments carried out in ecotrons so far is variable as it depends on the addressed scientific question. It can be relatively short (e.g. 4 months), especially when samples from long‐term field experiments are used (see section Complementarity between ecotron and field experiments) or can last up to 3 years. The average length of experiments run so far is 1 year.
2.5. Open access to the infrastructure
The physical sciences traditionally share their state of the art, large and costly infrastructures with hundreds of scientists from all over the world. Since ecotrons are also costly to build and run, albeit to a lesser scale, the experiments often include international teams assembled in large consortia. This arrangement facilitates the interaction among scientists with the complementary expertise needed to perform interdisciplinary research projects, for example, plant and animal ecologists, hydrologists, microbiologists, chemists, modellers, data scientists, etc. Therefore, most ecotrons work with open access calls. In most cases, ecotron experiments do not require a permanent presence of personnel from the external teams running an experiment: most of the environmental controls and key process measurements are automated, and the ecotron personnel regularly check the proper functioning of the instruments. Moreover, data are accessible in quasi real‐time via dedicated web interfaces. The external team is mainly involved in setting up the experiment and specific measurement campaigns. The most ambitious experiments, however, rely on a dedicated post‐doc and/or technician located at the ecotron.
Although open to private companies, most ecotrons are primarily running projects involving researchers from universities and/or research institutes and supported by public funding. The facilities charters may impose an open access data policy after a short embargo period. This access is often organized at the national level (nodes coordinating sets of ecosystem experimental facilities) or international level (e.g. the European ESFRI infrastructure AnaEE—Analysis and Experimentation on Ecosystems—www.anaee.eu, which includes several ecotrons). In Europe, the Cluster of Environmental Research Infrastructures (ENVRI) is developing a project (https://envri.eu/home‐envri‐fair) to feed the data from its constituent infrastructures (AnaEE and others) to the European Open Science Cloud.
2.6. Choosing between running experiments in CEF, field or ecotron
In the context of the former Silwood Park ecotron, Lawton (1995) argued that ‘model laboratory systems (real organisms interacting in the laboratory) are a halfway house between mathematical models and the full complexity of the field’. Current ecotron facilities, through their improved realism, are much closer to experimental field conditions. However, a comparison among experimental systems should include multiple criteria to inform choice of the facility best fit for the purpose: hypotheses that require environmental conditions technically difficult to achieve in the field and/or intensive process measurements would be better tested in ecotron experiments, as long as the number of drivers remains low. Hypotheses testing that requires a high level of realism and a large number of treatments or replicates would be best done in field experiments, at least when the application of treatments does not require expensive technology. CEF experiments appear to have considerably more limitations, but are certainly needed for rapid preliminary trials or when field or ecotron experiments are not available or are too costly to run.
3. THE CHARACTERISTICS OF CURRENT ECOTRONS
We identified 13 facilities having the defining features of ecotrons, including 11 facilities in Europe, one in North America and one in Australia (Tables 1A and 1B; Figure 1). Two were operational since the 1980s and 1990s, but most of them opened between 2010 and 2020, and two additional ones are currently being built. The Supporting Information file ‘Ecotrons description’ describes each facility, with website links, pictures, contextual information, specific technical details, contacts for collaboration and a short list of key publications. In another Supporting Information file (‘Ecotron‐related facilities’), some CEFs are described which paved the way for the development of current ecotrons (the phytotrons of the 1950s and 1960s, the sunlit growth chambers, the Closed Ecological Life Support Systems, the early ecotrons) or facilities which are now being developed for aquatic ecosystem research and plant phenotyping.
TABLE 1A.
Administrative and structural characteristics of the ecotrons. Additional information on each facility (including, in some cases, specific capacities of a subgroup of experimental units) are given in the Supporting Information file ‘Ecotrons descriptions'
| Ecotron short name | Owner | Town, country | Opening year | Access | Staff | No. climate controlled cells | Area each cell m2 |
|---|---|---|---|---|---|---|---|
| ExpoSCREEN, München | Helmholtz Zentrum München | Neuherberg, Germany | 1985 | ● | 3 | 4 | 6 |
| EcoCELLs, Reno | Desert Research Institute | Reno, Nevada USA | 1995 | ● | 0.25 | 4 | 40.5 |
| Whole Tree Chambers, Richmond | Western Sydney University | Richmond, NSW, Australia | 2006 | ○ | 1 | 12 | 8.3 |
| Montpellier Ecotron, Macrocosms | CNRS (INEE) | Montferrier sur Lez, France | 2010 | ● | 2.7 | 12 | 20 |
| IledeFrance Ecotron Ecolab, Nemours | CNRS (INEE) | Saint‐Pierre‐lès‐Nemours, France | 2017 | ● | 3.5 | 15 | 4.5 |
| iDiv Ecotron, Leipzig | iDiv, Leipzig University | Bad Lauchstädt, Germany | 2017 | ○ | 1.5 | 24 | 2 |
| TUMmesa, München | Technical University Munich | Freising, Germany | 2017 | ○ | 1 | 8 | 8 |
| UHasselt Ecotron, Hasselt | Hasselt University | Maasmechelen, Belgium | 2018 | ○ | 2.7 | 12 | 19 |
| TERRA Ecotron, Gembloux | Liège University | Gembloux, Belgium | 2018 | ● | 2 | 6 | 20.3 |
| Montpellier Ecotron, Mesocosms | CNRS (INEE) | Montferrier sur lez, France | 2018 | ● | 2.7 | 18 | 1 |
| TerraXcube, Bolzano | Eurac Research | Bolzano/Bozen, Italy | 2020 | ● | 2 | 4 | 9 |
| AGRASIM, Jülich | Forschungszentrum Jülich | Jülich, Germany | 2021–2022 | ○ | 3 | 4 | 2.6 |
| Antwerp Ecotron | University of Antwerp | Antwerp, Belgium | 2021 −2022 | ● | 1 | Not set | 4 |
Access, ● open calls (see Supporting Information file ‘Ecotrons individual descriptions’ for links to these calls) with occasional in‐house collaborative projects; o in‐house projects with external collaborations; Staff, Number of persons/year (permanent or on temporary contract) fully dedicated to the functioning of the ecotron. (e.g. 1 means that there is the equivalent of one technical person working 12 months full time for the facility).
Number of cells, The cells provide (independently from each other) the climate control over a single lysimeter (they are then called enclosures) or over several lysimeters (in that case they are then called chambers).
Area cell, Area of each enclosure or chamber allowing the climate control. It is the lysimeter(s) area plus, if present, a walking area around the lysimeter(s).
TABLE 1B.
Table 1 Continued
| Ecotron short name | No. lysimeters/cell* | Area each lysimeter m2 | Air volume** m3 | External air flux*** m3/min | Air internal recirculation m3/min | Plant height max m | Soil depth max m | Soil weight/lysimeter | Biosafety level |
|---|---|---|---|---|---|---|---|---|---|
| ExpoSCREEN, München | 4 | 0.6 | 0.5** | 1.3 to 2.7 | None | 0.8 | ≤0.7 | 0.5 t | 1 |
| EcoCELLs, Reno | 1 | 9 or 11 | 130 | 13 to 130 | ~660 | 2.4 | 1.8 | 36 t | 1 |
| Whole Tree Chambers, Richmond | 1 | 8.3 | 53 | 0.6 | 180 | 9 | 1 | In situ | 1 |
| Montpellier Ecotron, Macrocosms | 1 | 2 or 4 or 5 | 35 | 2.7 | 70 | 3 | 0.6–2 | 3 to 15 t | 1 |
| IledeFrance Ecotron Ecolab, Nemours | 1* | 1.3 | 13 | 0 to 200 | 0–1.25 | 1.5 | 0.8 | ≤ 2 t | 1 |
| iDiv Ecotron, Leipzig | 1* | 0.2 | 3 | <6 | None | 1.5 (1.2) | 0.8 | 0.2 t | 1 |
| TUMmesa, München | 4* | 0.38 | 36 | 1.9 | 83 | 1.5 | 0.8 | 238 kg | 1 |
| UHasselt Ecotron, Hasselt | 1 | 3.14 | 222 | 0 | 60 | 2.5 | 1.4 | 5–12 t | 1 |
| TERRA Ecotron, Gembloux | 1* | 2 | 65 | 45 | 1–3.2 | 1.5 | 1.5 | 6 t | 2 |
| Montpellier Ecotron, Mesocosms | 1 | 1 | 4 | 0–0.5 | 10 | 1.8 | 1 | 0.3–2 t | 1 |
| TerraXcube, Bolzano | 4 | 0.13 | 27 | 1.7 | 60 | 2.5 | 0.4 | 60 kg | 2 |
| AGRASIM, Jülich | 1 | 1 | 6.9 | 0.02–1 | ≤27 | 2.5 | 1.4 | ≤3 t | 1 |
| Antwerp Ecotron | 1 | 1 | 7 | 0.3–2.5 | 25 | 1 | 1 | ≤2 t | 1 |
Biosafety: Level 1: washing hands upon entering and exiting the lab; potentially infectious material decontaminated before disposal; lab must have a door which can be locked to limit access.
Level 2: items of level 1 plus: advanced training for personnel and scientists; limited access to the laboratory; extreme precautions to be taken with contaminated items; use of physical containment equipment when aerosols or splashes may occur.
A higher number of smaller lysimeters can also be used. When available (*) this option is detailed in the Supporting Information file ‘Ecotrons individual descriptions’.
The volume of the chamber permanently enclosing the canopy above each lysimeter, otherwise indicates the volume of the whole cell.
The facilities with 0 external air flux works (or can work) in a close system mode while the other facilities work in an open system mode (cf. principles of gas exchange measurements).
FIGURE 1.

Left to right and top to bottom: UHasselt ecotron (Hasselt Belgium), EcoCELLs (Reno USA), Whole Tree Chambers (Richmond Australia), iDiv Ecotron (Leipzig Germany), TUMmesa (München Germany), IleDeFrance ecotron Ecolab (Nemours France), ExpoSCREEN (München Germany), TERRA ecotron (Gembloux Belgium), TerraXcube (Bolzano Italy), and Macrocosms and Mesocosms platforms of the European Montpellier Ecotron (France)
Design differences among ecotrons reflect the scientific and strategic objectives of the funding organizations. The average construction cost of the most recent (2010 and after) ecotron platforms is 6 M€ with a large range of variation (3–10 M€) revealing differences in the number or size of the experimental units or in their control and measurement capacities. For a given amount of available funds, there are unavoidable trade‐offs among (i) the number of controlled environmental parameters (with light quality and isotopes being the most complex/expensive ones to control); (ii) the number of processes measured in real time by automated systems (with soil respiration, trace gas emission, isotope fractionation and faunal activity being the most complex/expensive ones to measure); (iii) the number of replicated units, and (iv) the size of these units. The ecotrons planning phase took 2.5 years in average and the building phase (including tests) 3.5 years. Average annual running costs in 2019 were ≈140 k€ (with 80 k€ for the maintenance and small improvements and 60 k€ for the consumables of which electricity, with a consumption around 600 MWh, partly green, constitutes the major part). Personnel annual cost averaged ≈160 k€ (for an average 2 full‐time equivalent).
3.1. Controlled environmental parameters
The environmental parameters controlled in these ecotrons are summarized in Tables 2A and 2B. Among the climatic conditions, air temperature and relative humidity are regulated, as in most CEFs, with low temperature (below 5°C) being achievable in almost all facilities, and freezing temperature and air humidity below 30% attainable in nine out of 13 facilities, thus allowing the simulation of winter climatic conditions of most temperate regions when needed. Since light intensity and quality have been a major concern regarding the external validity of CEFs, these parameters have been given high priority in ecotron design. High light levels are achieved in the sunlit ecotrons, thanks to high overall transmissivity (≈0.9 on average) of the canopy enclosures. This transmissivity, calculated over 24 h with sensors inside and outside the enclosures, is very high compared to glasshouses. This is due to a high transmissivity of the covering material, a low inter‐cell shading, a dome like shape and very light supporting structures. In facilities with artificial light, when the ground‐lamp distance is adjustable, the photosynthetic photon flux density (PPFD) averaged across ecotrons is 1100 µmol m−2 s−1 at 50 cm from the light sources. Otherwise, when the distance is not adjustable, the PPFD range across ecotrons is 340–1300 µmol m−2 s−1 at 50 cm above the ground level. Maximum PPFD outside can be significantly higher on clear summer days, but while PPFD is the most relevant variable for instantaneous photosynthesis, many plant traits at higher levels of integration are better related to the daily light integral (DLI), the PPFD integrated over a day (Poorter et al., 2019 and references therein). In June, the highest average DLI is 45 mol m−2 day−1 at a latitude of 40°N (Poorter et al., 2019). This can be obtained in ecotrons with only 11.5 h of constant light at a PPFD of 1100 µmol m−2 s−1. In addition, all ecotrons running with artificial lights have dimmable lamps or step switching of the lamps to simulate typical daily light courses. Although Poorter et al. (2016) emphasized the impossibility to reach, in growth chambers, the high photothermal ratio found outside in spring at most latitudes (high light at low temperatures), photothermal ratios were found to match these field values both in a sunlit ecotron (Montpellier Macrocosms) and in an artificially lit ecotron (Gembloux Terra ecotron), for example on April days with a DLI > 20.
TABLE 2A.
Controlled environmental parameters in each of the ecotrons*
| Ecotron short name | Opening date | Air T °C* | Air T °C < 5* | Air T °C < 0* | Air RH %* | Air RH <30%* | Soil boundary T°C* | No. sensors T°C soil V × H | Watering S, D, M* | SWC%* | No. sensors SWC V × H | Soil boundary ψm | No. sensors soil ψm V × H |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ExpoSCREEN, München | 1985 | ● | ● | ● | ● | ● | □ | D, M | □ | □ | |||
| EcoCELLs, Reno | 1995 | ● | ● | ● | ● | ● | ● | 5 × 15 | S, D, M | ● | 5 × 6 | □ | |
| Whole Tree Chambers, Richmond | 2006 | ● | ● | 3 × 1 | S, D, M | 3 × 3 | — | ||||||
| Montpellier Ecotron, Macrocosms | 2010 | ● | ● | ● | ● | ● | 4 × 3 | S, D, M | ● | 4 × 3 | ○ | 1 × 1 | |
| Ile de France EcoLab, Nemours | 2014 | ● | ● | ● | ● | ● | ● | 1 × 1 | S, D, M | ● | 4 × 10 | □ | 1 × 1 |
| iDiv Ecotron, Leipzig | 2017 | ● | ● | 3 × 4 | S, M | ● | 3 × 4 | 3 × 4 | |||||
| TUMmesa, München | 2017 | ● | ● | ● | 4 × 1 | S, D, M | ● | 4 × 1 | □ | ||||
| UHasselt Ecotron, Hasselt | 2018 | ● | ● | ● | ● | ● | ● | 5 × 3 | S, M | ● | 5 × 3 | ● | 5 × 3 |
| TERRA Ecotron, Gembloux | 2018 | ● | ● | ● | ● | ● | 5 × 1 | S, M | ● | 5 × 1 | ● | 3 × 1 | |
| Montpellier Ecotron, Mesocosms | 2018 | ● | ● | ● | ● | ● | ● | 4 × 2 | S, D | ● | 4 × 2 | ○ | 1 × 1 |
| TerraXcube, Bolzano | 2020 | ● | ● | ● | ● | ● | ○ | 3 × 1 | S, D, M | ● | 3 × 1 | ● | 3 × 1 |
| AGRASIM, Jülich | 2021–2022 | x | x | x | x | x | x | 7 × 2 | D, M | x | 7 × 2 | x | 7 × 1 |
| Antwerp Ecotron | 2021–2022 | x | x | x | x | x | x | 3 × 1 | S, D, M | x | 3 × 1 | x | 1 × 1 |
*, controlled parameters have their name in bold.
●, existing automatic control and/or measurement.
○, automatic control and/or measurement being installed.
□, measurement/sampling done manually at frequencies to be determined (optional, upon negotiation).
x, planned regulation and measurements (in facilities being built).
—, indicates that this parameter is not relevant (sunlight transmission for indoors facilities, adjustable lamp intensity for sunlit facilities).
TABLE 2B.
Table 2 continued
| Ecotron short name | No. sensors soil EC V × H | Sunlight transmission T vis, T UV | PPFD µmol/m2/s | CV PPFD% | Adjustable lamps intensity* | UV radiation % | Red/far‐red | Canopy air speed m/s | CO2 > 400 ppm* | CO2 pre‐industrial* | δ13C CO2* | O3* | NOx* |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ExpoSCREEN, München | — | 600 | ≤10 | ● | 4–6.5 | 1.3–1.7 | <2 | ● | ● | ● | ● | ● | |
| EcoCELLs, Reno | □ | 0.8–0.9 | s.l. | ≤15 | — | s.l. | s.l. | <2 | ● | ● | |||
| Whole Tree Chambers, Richmond | 0.93, 0.93 | s.l. | Variable | — | s.l. | s.l. | 0.3 | ● | ● | ||||
| Montpellier Ecotron, Macrocosms | 0.92, 0.8 | s.l. | 10 | — | s.l. | s.l. | 1 | ● | ● | ||||
| Ile de France Ecotron EcoLab, Nemours | □ | — | 1000 | <10 | ● | 2 | 1.2 | 0.1–20 | ● | ● | ○ | □ | □ |
| iDiv Ecotron, Leipzig | — | 340 | 10 | ● | 0.3% | 1.0 | <0.7 | □ | □ | ||||
| TUMmesa, München | — | 1070 | 4 | ● | 8.6 | 3.5 | <0.25 | ● | □ | ● | ● | ||
| UHasselt Ecotron, Hasselt | 5 × 3 | 0.95, 0.95 | s.l. | <10 | — | s.l. | s.l. | 1.52 | ● | ● | |||
| TERRA Ecotron, Gembloux | — | 1200 | 7 | ● | 4.7 | 1.2 | 0.2 | ● | ● | ||||
| Montpellier Ecotron, Mesocosms | 0.9, 0.8 | s.l. | <10 | — | s.l. | s.l. | <0.7 | ● | ● | ● | |||
| TerraXcube, Bolzano | — | 1300 | <10 | ● | tbd | tbd | 2 | ● | ○ | ○ | |||
| AGRASIM, Jülich | — | 1200 | tbd | x | tbd | tbd | 0–10 | x | x | x | x | ||
| Antwerp Ecotron | 0.9, 0.9 | s.l. | tbd | — | s.l. | s.l. | tbd | x |
s.l., sunlit facility; tbd, to be determined.
T°C, RH%, temperatures and relative humidity refers to day time or when the light are on. Some performances can only be reached under adequate outside climatic conditions (e.g. negative air temperatures not reachable in summer).
T°C, ψm, control of the soil temperature or matrix potential at the bottom of the lysimeter. It recreates near natural soil temperature and matrix potential profiles.
V × H, V is the number of positions of sensors vertically, H is the average number of sensors placed horizontally at each vertical position.
S, D, M, watering can be done by spray (S) with nozzle(s) above the canopy or by drip (D) with drippers on the soil surface, or manually (M).
SWC%, soil water content controlled via watering after measurements of water loss (weighing the lysimeters) or via soil humidity sensors in the soil profile.
T vis, T UV, sun light transmission by the containment structure in the visible range T vis and in the UV range T UV.
PPFD, maximum photosynthetic photon flux density at 50 cm below lamps when soil‐lamp distance is adjustable, or 50 cm above soil level when this distance is not adjustable.
CV PPFD, light homogeneity: variation coefficient of PPFD measured at several points uniformly distributed over the canopy area.
UV, (ratio radiation photons 280–400 nm/radiation photons 280–700 nm) × 100. This percentage is 6.6 for solar radiation (based on the standard solar spectrum AM1.5 expressed in photons).
red:far red ratio radiation photons 600–700 nm/radiation photons 700–800 nm. This percentage is 1.1 for solar radiation (based on the standard solar spectrum AM1.5 expressed in photons).
δ, (delta): ratio of stable isotopes in a given molecule (here 13C/12C in CO2) in reference to a standard (Pee Dee Belemnite).
Although rarely documented, light homogeneity across the horizontal plane is often also of concern in CEFs due to lamp positions or light interception by greenhouses structures. In the listed ecotrons, the variation coefficient of PPFD is typically below 10%. With artificial light, the vertical extinction profile of light in the canopy is much stronger than in nature due to the quadratic loss of light with increasing distance from the lamps. In tall canopies, supplemental vertical strings of LEDs can compensate for this. Surrounding the sides of the canopy with a vertical shading cloth of adjustable height and transmissivity is recommended to prevent light and turbulent air from entering the canopy sideways. Aboveground edge effects can be minimized that way, but belowground edge effects are unavoidable and are only lessened by the large area to circumference ratio of typical ecotron lysimeters. Light quality, which also strongly impacts the external validity of CEF studies, is considerably improved in some artificially lit ecotrons using LED arrays providing a continuous light spectrum close to the solar spectrum with most of the physiologically active wavelength in adequate proportions (although UVB is often still missing). Given the increasing recognition of the importance of UV radiation (Ulm & Jenkins, 2015; Verdaguer et al., 2017), in addition to the red: far red ratio, for the growth and development of plants (Galvão & Fankhauser, 2015) and for some trophic interactions (e.g. Moreno et al., 2009), these light quality parameters are reported in Tables 2A and 2B. The average proportion of UV in the UV +PPFD spectrum, calculated from data in µmol m−2 s−1, is 3.8% in the ecotrons with artificial light (range: 0.3–8.6) compared with 6.6% in the standard AM1.5 solar spectrum, and the red: far red ratio is on average 1.7 (range: 1–3.5) compared to 1.1 in the AM1.5 solar spectrum.
Despite relatively high air internal recirculation in most ecotrons (often two to three cell volumes per min), air speed at the canopy level is generally below 2 m s−1. Such values are common over short statured vegetation such as agricultural crops or tree nurseries (mostly between 0.5 and 1 m s−1, and rarely exceeding 3 m s−1; Day & Parkinson, 1979; Barnard & Bauerle, 2016), which are the vegetation types typically studied in ecotrons.
Air CO2 concentration is nowadays controlled in most CEFs and in all the ecotrons, except one. This control is important not only for studies simulating future CO2 concentrations but also to prevent variation in daytime CO2 concentration when plant photosynthesis is active (Bernier et al., 1994; Romer, 2001). The one ecotron lacking routine control of air CO2 concentration overcomes this by a high external air flux to cell air volume ratio. Pre‐industrial CO2 concentrations can be simulated in half the ecotrons by scrubbing the incoming air with a CO2 removal system. This opens the opportunity to label the organic matter synthesized during the whole experiment by continuously injecting CO2 with a specific δ13C signature (depleted or enriched). Ozone concentration is controlled in three ecotrons while NOx is controlled in a single ecotron.
Due to the high content of dissolved nitrogen often found in tap water, all ecotrons, except 3, are using deionized water to simulate rainfall. Two of them have the capacity to add specific ions to the deionized water and two can alternatively use stored rainwater. Dew is generally not observed on the ecotron canopies. Snow cannot be generated within ecotrons, but can be brought in from outside.
3.2. In‐house measured processes in standard operation mode
The in‐house measured processes in the ecotrons are summarized in Table 3. Overall, 18 different ecosystem processes are assessed, with an average of 10 per ecotron among which five as routine high‐frequency measurements. Seven of the 18 processes refer to emitted or absorbed gases at the ecosystem scale. Evaporated and transpired water as well as photosynthesized and respired CO2 are key in understanding and measuring primary productivity. Emphasis is on the three main greenhouse gases (CO2, CH4 and N2O) with six ecotrons capable of measuring their fluxes, enabling calculation of metrics for the global warming potential of ecosystems (Neubauer & Megonigal, 2015). Measurements of the stable isotopes in the CO2, H2O and N2O molecules are being developed. Routine or on‐demand measurements of δ13C and δ18O of CO2 (providing information on canopy conductance, respiration and coupling of cycles) are possible in six and four of the ecotrons, respectively. Measurements of δ15 N, δ18O and the intramolecular SP (isotopomers) of 15N in N2O (providing information on nitrification and denitrification) can be done in four ecotrons, and measurements of δ2H, δ18O of H2O (providing information on the water cycle) in two ecotrons.
TABLE 3.
In‐house process measurements done automatically (continuously or at high frequency) as services offered routinely by the facility to its internal or external users or done manually at a frequency to be determined after negotiation. Measurements at scales smaller than the ecosystem (leaf level for example) as well as measurements which are usually done externally on soil plant or air samples are not considered in this table
| Ecotron short name | Opening year | ET (H2O) | NEE (CO2) | δ13C in CO2 | Δ18O in CO2 | Soil respiration | CH4 emission | N2O emission | O3 emission |
|---|---|---|---|---|---|---|---|---|---|
| ExpoSCREEN, München | 1985 | ●ge | ● | ○ | ● | ||||
| EcoCELLs, Reno | 1995 | ● | ● | □ | □ | ● | □ | □ | |
| Whole Tree Chambers, Richmond | 2006 | ●ge | ● | □ | □ | ● | □ | □ | |
| Montpellier Ecotron, Macrocosms | 2010 | ● | ● | ● | ● | □ | ● | ● | |
| Ile de France Ecotron, EcoLab | 2014 | ● | ● | ○ | ● | □ | □ | ● | |
| iDiv Ecotron, Leipzig | 2017 | □ | □ | □ | |||||
| TUMmesa, München | 2017 | ● | □ | ||||||
| UHasselt Ecotron, Hasselt | 2018 | ● | ○ | ○ | ○ | ||||
| TERRA ecotron, Gembloux | 2018 | ● | ● | ● | ● | ||||
| Montpellier Ecotron, Mesocosms | 2018 | ● | ● | ● | ● | □ | ● | ● | |
| TerraXcube, Bolzano | 2020 | ● | ○ | □ | |||||
| AGRASIM, Jülich | 2021–2022 | x | x | x | x | x | x | x | |
| Antwerp Ecotron | 2021–2022 | x | x | x | x | x |
| Ecotron short name | VOCs fluxes | δ15 N, δ18O in N2O | δ2H, δ18O in H2O | Drainage flux | Soil solution sampling ** | Root growth *** | LAI*** | Canopy leaf T °C *** | Hyper‐ spec *** | Fauna tracking *** |
|---|---|---|---|---|---|---|---|---|---|---|
| ExpoSCREEN, München | □ | □ | □ | |||||||
| EcoCELLs, Reno | □ | ● | □ | □ | □ | □ | □ | |||
| Whole Tree Chambers, Richmond | □ | □ | □ | □ | □ | ● | □ | □ | ||
| Montpellier Ecotron, Macrocosms | ● | □ | □ | □ | ||||||
| Ile de France Ecotron, EcoLab | □ | □ | □ | ● | ● | □ | □ | □ | ||
| iDiv Ecotron, Leipzig | □ | ○ | □ | □ | □ | □ | □ | ○ed,rf | ||
| TUMmesa, München | ○ | □ | □ | □ | □ | |||||
| UHasselt Ecotron, Hasselt | ● | ● | ||||||||
| TERRA ecotron, Gembloux | ● | □ | □ | ●st | □ | □ | ||||
| Montpellier Ecotron, Mesocosms | ● | □ | □ | □ | ||||||
| TerraXcube, Bolzano | ● | □ | ○ | ○ | □ | |||||
| AGRASIM, Jülich | x | x | x | x | x | x | x | |||
| Antwerp Ecotron | x | x | x | x |
●: existing automatic continuous (or at high frequency) measurement provided to the hosted teams.
○:automatic continuous (or at high frequency) measurement being installed; □, measurement/sampling done manually (by the facility staff or by the hosted team), often using available portable devices, at frequencies to be determined (optional, upon negotiation).
ET: evapotranspiration, measured by weighing (●) or by gas exchange (●ge).
NEE:net ecosystem exchange of CO2 (balance between canopy photosynthesis and canopy and soil respiration).
δ: isotopic difference resulting from fractionation within the ecosystem (δ value difference of a given gas before and after going through an ecosystem in an ecotron unit).
Root growth: measured with minirhizotrons (ingrowth cores measurements are not indicated here since they do not require instrument investment by the facility and are usually done by the hosted team).
LAI: leaf area index (it often gives also canopy transmittance, but this transmittance can also be obtained with simpler light sensors); ●st LAI measured by stereoscopic cameras;
Hyperspec: Canopy hyperspectral reflectance.
Fauna: tracking done by real time detection number and size measurement of catched soil microarthropods (Edapholog system) (○ed) or/and RFID monitoring of beetle movements (○rf).
**Sampling, but no measurements on these samples (measurements often done externally).
***The facilities own the (often portable) equipement to run the raw initial measurements but since finalising results requires a lot of man power and/or very specific skills (i.e. analysis of the roots pictures, parametrization of the hyperspectral models), these final results are often not offered routinely to external users of the facility.
Drainage fluxes and soil solution sampling are provided routinely or on demand in most ecotrons. Minirhizotrons are available in half of the ecotrons, but the root images are usually not analysed by the facility personnel. LAI meters are generally available, and one ecotron is equipped with an automatic stereoscopic measurement of LAI. Canopy temperature infrared measurement can be done in half the ecotrons (two automated at high frequency). Hyperspectral reflectance measurements are being developed for non‐destructive measurement of chemical contents and biomass of canopies. Equipment for such measurements is available in half the ecotrons. Fauna activity is analysed automatically in only one ecotron through cameras and radio frequency identification, in addition to a computerized trap system for the activity of soil microarthropods.
4. TRACK RECORD OF ECOTRONS
4.1. Environmental control and process measurements
Examples of environmental controls achieved in ecotrons are shown in Figure 2. The sunlit ecotron of Hasselt demonstrates the capacity to track the rapidly fluctuating field conditions at the nearby ICOS station. The Gembloux ecotron, operating with artificial (LED) lights, shows the capacity to simulate ranges of light and temperature derived from a model using minimum and maximum temperatures and total solar insolation from a weather station, combined with astronomical and heat transfer data.
FIGURE 2.
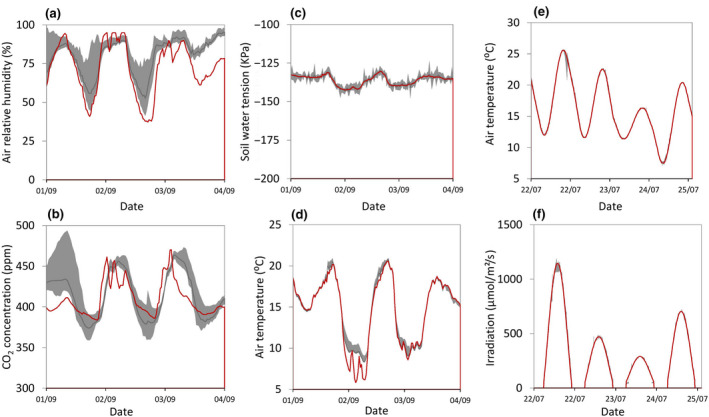
Examples of ecotron environmental controls: Simulation of outside air relative humidity (a), CO2 concentration (b), soil water tension (c) and air temperature (d) measured at the Maasmechelen, Belgium ICOS station and reproduced in the UHasselt sunlit Ecotron (unpublished data), and simulation of air temperature (e) and photosynthetically active radiation (f) derived from a model and reproduced in the Terra Ecotron Gembloux with artificial lights (unpublished data). Red lines: conditions to be simulated, grey area: range of variation of the parameters across 12 (a–d) or 3 (e, f) experimental units, dark grey line: average for the 12 or 3 units
To exemplify the capabilities in terms of isotopic labelling and process measurements, we show the successive measurements in a single experimental unit while 12 units (Montpellier) or 16 units (München) are labelled or measured simultaneously with single analysers multiplexed across these units (Figure 3).
FIGURE 3.
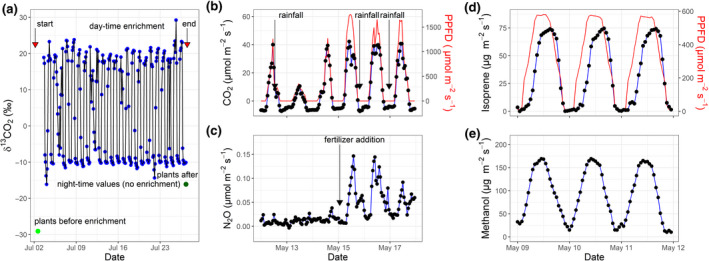
Examples of ecotron isotopic labelling and process measurements: Air 13CO2 enrichment and plant13C labelling in one macrocosm of the Montpellier Ecotron (a); measurements of net ecosystem CO2 exchange (b) and net ecosystem N2O exchange (c) with photosynthetic photon flux density (PPFD) in one macrocosm of the same ecotron; measurements of the emission of two volatile organic compounds (isoprene and methanol) with PPFD in one of the sub‐chambers of the München ExpoSCREEN facility (d, e)
4.2. Scientific achievements
A cluster analysis of the published ecotron papers based on the applied experimental treatments reveals three main categories: investigating ecosystem response to abiotic global change drivers (‘novel environments’ experiments sensu Hanson & Walker, 2020) (55% of the papers), deepening our understanding of ecosystem processes (27%) and understanding biodiversity–ecosystem functioning relationships (18%; Figure 4a). Since more than half of the ecotrons presented in the current paper have opened too recently to have published results, we included the experimental results papers from the Silwood Park ecotron to document more broadly the research areas. The Silwood Park data represent 27 of a total of 126 papers analysed in Figure 4a. In addition, we also analysed in the same way the running or recently completed, but not yet published, experiments in the newly open ecotrons (Figure 4b).
FIGURE 4.
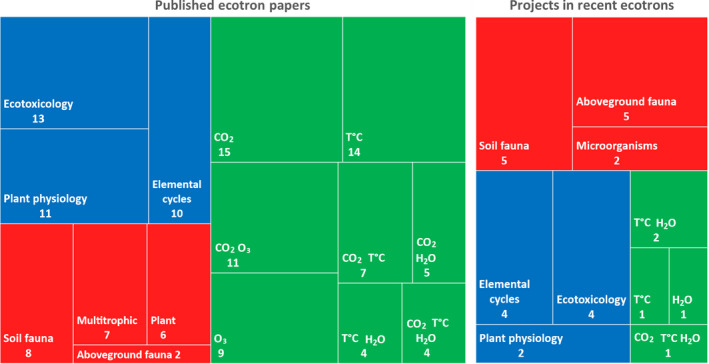
Treemap diagrams showing the research areas covered by the ecotrons in the published ecotron papers (left) and in the running or recently completed projects in the newly open ecotrons (right). Blue: experiments designed to better understand specific ecosystem processes; green: global change experiments simulating ‘novel’ future environments; red: biodiversity experiments with manipulations at various trophic levels. Numbers indicate the number of papers/experiments in each sub‐category
Half of the papers on novel environments analyses the interactions between at least two environmental drivers. CO2 and temperature were the most studied drivers (61% and 42% of 69 papers, respectively). Among the papers on ecosystem process understanding, ecotoxicology is well represented with several papers on mercury circulation in ecosystems published by one ecotron. Plant physiology and elemental cycles are also well represented. Most experiments addressing the role of biodiversity in ecosystem functioning were conducted using grassland ecosystems. Interestingly, manipulations of soil fauna and multi‐trophic systems are as represented as plant manipulations. The clustering of the newly completed or running projects in the recently open ecotrons shows an increase in biodiversity experiments, especially related to fauna, as a result of the opening of the German ecotrons. Novel environment experiments are proportionally less studied in these recent ecotrons. The list of published papers and recent projects is shown in the Supporting Information file ‘Ecotron published papers and recent projects’.
Below we describe selected experiments showing how the environmental control and process measurement capacities in ecotrons led to remarkable findings in each of the three main scientific areas of Figure 4.
4.2.1. Global change experiments
Drake et al. (2018) studied how an extreme heatwave affects the physiological performance of forests. They grew whole Eucalyptus parramattensis trees (6 m tall) in a field setting with the Richmond Whole Tree Chambers (WTC), and crossed the heatwave with a warming treatment. The WTC controlled T air, vapour pressure deficit and CO2 concentration while measuring net CO2 and H2O exchange of the entire canopy every 15 min. Additional measurements were leaf temperature, fluorescence and water potential, and a leaf‐level photosynthetic model was used. The heatwave reduced canopy photosynthesis more strongly than transpiration, which maintained canopy cooling. This decoupling is not captured in the standard photosynthetic models and consequently is not considered in climate models, overestimating the negative impact of heatwaves. This result, as well as an observed increase in leaf thermal tolerance during the heatwave, was identical in both the ambient and warmed treatments. Using similar high‐frequency ecosystem gas exchange measurements, Roy et al. (2016) showed in the Montpellier ecotron Macrocosms that elevated CO2 buffered the impact of an extreme drought and heat on intact grassland monoliths, mostly owing to very strong recovery in autumn under this treatment. Also using continuous measurements of CO2 NEE, Arnone III et al. (2008) demonstrated in the Reno EcoCells that the reduction of CO2 uptake in intact tallgrass prairie monoliths by an anomalously warm year was carried over to the next year because soil biota respiration was stimulated. Using real‐time measurements of NEE and VOCs, Ghirardo et al. (2020) showed in the München ExpoSCREEN that global warming decreases carbon sequestration in subarctic tundra ecosystems via reducing NEE and increasing VOC emissions. The use of 13CO2‐labeling experiments further allowed coupling the atmospheric carbon dioxide to VOCs biosynthesis and emissions.
4.2.2. Biodiversity–ecosystem functioning experiments
Recent findings of an nearly 75% decline in flying insect biomass over the past 27 years (Hallmann et al., 2017) motivated the investigation of the repercussions of such a decline for ecosystem processes. Using the 24 experimental EcoUnits of the Leipzig iDiv Ecotron, nine projects investigated how the decline in invertebrate biomass in grassland ecosystems affects the biotic interactions between aboveground (insects, plants and bacteria) and belowground organisms and thus the associated ecosystem functions and services. Artificial grassland communities consisting of 12 central European species were established in the EcoUnits. To mimic invertebrates decline, live aboveground invertebrates were introduced at two abundance levels (100% and 25%) in eight EcoUnits each while another eight EcoUnits received no fauna. The invertebrate treatment had a large impact on the extent of an accidental aphid infestation which occurred in all 24 EcoUnits. The strongest infestation occurred in the EcoUnits without additional invertebrates and the weakest in the 100%‐invertebrate EcoUnits, underlining the importance of natural pest control by predators. Invertebrate densities also shifted plant species abundances and phenology. For example, the dominant species in the grassland community, Trifolium pratense, declined in abundance with invertebrates present (Ulrich et al., 2020).
4.2.3. Process understanding experiments
In the Macrocosms platform of the Montpellier Ecotron, the effect of circadian rhythm on the diurnal gas exchange of leaves and canopies was investigated (Resco de Dios et al., 2017). Such an effect is studied by maintaining all environmental parameters constant (with light or in the dark) after a few days of ‘entrainment’ during which light, temperature and water vapour pressure deficit follow typical outdoor conditions while gas exchange is measured throughout every 12 min. The sunlit macrocosms were planted with either bean or cotton. After 1 month of growth, a completely opaque cover was fitted on each macrocosm dome and light was then controlled by dimmable plasma lamps. Under these field‐like conditions, circadian regulation was observed to exert control over net CO2 exchange that was of similar magnitude to the controls exerted by direct physiological responses to temperature and vapour pressure deficit (Resco de Dios et al., 2016, 2017). Circadian rhythm also induced contrasting changes in the photosynthetic pigments and photochemical efficiency in bean vs. cotton, calling into question the extrapolation of the response of model plants to other species (García‐Plazaola et al., 2017). Night‐time dark respiration showed a circadian oscillation at both leaf and canopy level, but light‐enhanced dark respiration was under circadian control only in cotton, suggesting that circadian controls may help explain temporal variability in ecosystem respiration (Gessler et al., 2017).
5. COMPLEMENTARITY BETWEEN ECOTRON EXPERIMENTS, FIELD EXPERIMENTS AND IN‐NATURA OBSERVATIONS
The earlier‐mentioned fundamental trade‐off between internal and external validity dictates that no single approach can span the entire validity gradient (De Boeck et al., 2015). Ecotrons can elucidate mechanisms underpinning responses to an array of potential changes in the environment, yet their inherent limitations (small spatial scale, island effect, reduced biological complexity, low number of replicates, etc.) warrant that outcomes of ecotron studies be considered in conjunction with results from other approaches.
Some ecosystems do not allow to take representative samples that could be transferred to ecotrons. This is the case for mature forests, one of the most critical ecosystems with regard to the regulation of the global climate (carbon sequestration, water cycle) and the preservation of the Earth's biodiversity. To study the mature forest ecosystems, in‐situ experimental platforms exist where certain features of the environment are changed, for example through FACE systems (Norby et al., 2016), through the use of extended rainout shelters above (Misson et al., 2011) or below (Hoover et al., 2018) the canopy or through natural phenomena, such as in the FORHOT study in Iceland, where seismic activity is warming the soil under an existing Sitka spruce forest (Sigurdsson et al., 2016). Furthermore, natural fluctuations in ambient weather (warm years, dry summers, etc.) enable the use of data from observational studies, including the vast array of eddy flux covariance towers (e.g. Schwalm et al., 2010). Ecotrons studies can complement studies of the early stages of forest growth and in particular determine how trees respond to interactive factors (CO2 and drought e.g. Crous et al., 2012, CO2 and temperature e.g. Crous et al., 2013, drought and temperature e.g. Drake et al., 2018, 2019). As the climate becomes more extreme, this type of research gains interest from both managers and conservationists interested in establishment success and growth of forests with a species or ecotype composition that is ‘climate proof’ (e.g. Coomes et al., 2014).
Another strategy that combines ecotron and field studies can circumvent the fact that ecotron experiment cannot be run for many years. This strategy is illustrated by the Jena Experiment, initiated in 2002 and still running. For the main experiment in the field, herbaceous plant communities of one to 60 plant species and one to four plant functional groups were sown on a former arable site. A high number of plots (82 plots 20 m × 20 m each) was necessary to allow the partitioning of biodiversity effects on ecosystem functioning into the effects of species richness, functional group richness and the contribution of particular functional groups (Weisser et al., 2017). Once the diversity effects had been well established in the field, underlying physiological mechanisms were studied in the Macrocosms platform of the Montpellier ecotron. Large soil monoliths with vegetation (2 m2, 2 m deep, 8 t) were sampled in plots of four and 16 species 9 years after sowing and inserted in the ecotron for an entire growing season. Automatic measurements of ecosystem evapotranspiration, net carbon exchange and night‐time respiration together with tracer studies (13C, 15N, 2H), modelling and a final destructive harvest enabled the determination of day respiration and photosynthesis, the partitioning of evapotranspiration into evaporation and transpiration, efficiency of water, nitrogen and light use, carbon allocation to soil compartments, patterns of root water uptake and the relative effect sizes of biodiversity components (Guderle et al., 2018; Lange et al., 2015; Mellado‐Vázquez et al., 2016; Milcu et al., 2014, 2016; Roscher et al., 2019). Only field experiments can provide such large plots for many experimental treatments and that can be used for decades, but the ecotron was necessary to answer questions arising from field observation but requiring additional measurement capacities difficult to implement in the field. Since the ecotron was used as an ecosystem analyser of treatment plots manipulated in the field for 9 years, the transient effects following the start of the treatments are considered to have faded. In this case, the length of the experiment in the ecotron could then be limited to a single growing season, another advantage of the complementarity between ecotron and field studies.
In ecotrons, the size limits also constrain the complexity of the ecosystem under study. Not all the drivers and trophic levels found in nature are typically represented in ecotrons, even though they may be relevant or even critical for the process under scrutiny. For example, it is well known that herbivores and predators can play a major role in some ecosystem responses to climate change. For example, bark beetle outbreaks often coincide with drought as trees suffering from drought stress tend to have reduced defences against pathogens and herbivores, leading to improved feeding opportunities for these insects and their offspring (Kolb et al., 2019; Marini et al., 2017). Such pest outbreaks can have even more profound effects on tree growth and mortality than the initial drought (Fettig et al., 2019). Although these secondary effects cannot be fully explored in ecotrons, experiments where targeted herbivore species or other relevant taxonomic groups of animals are introduced in a controlled manner are possible (e.g. Stevnbak et al., 2012; Van De Velde et al., 2017). The iDiv Ecotron was specifically built to study aboveground–belowground multitrophic communities of invertebrates and their impacts on ecosystem processes (Eisenhauer & Türke, 2018). Although such studies must be supplemented by observations in the field to increase the external validity, they are valuable for testing specific hypotheses and uncovering particular mechanisms as part of a larger research strategy (e.g. Thakur et al., 2020).
Ecotrons can also provide a critical contribution in verifying seedling‐scale phenomena obtained in greenhouses and in testing the robustness of scaling relationships from leaf to ecosystem. For example, the WTC were used to investigate the role of heatwaves on trees physiology, complementing glasshouse‐based studies of potted seedlings (Aspinwall et al., 2019). This provided a rare test of integrated canopy gas exchange responses to an experimental heatwave on large trees. In turn, the WTC study provided evidence to underpin ecosystem‐scale assessments of the uncoupling of photosynthesis and transpiration during heatwaves using eddy covariance flux data (De Kauwe et al., 2019). In addition, WTC studies elucidated the mechanisms of temperature responses of tree growth and thermal acclimation of carbon exchange under field conditions (Aspinwall et al., 2016; Drake et al., 2016), complementing findings determined in glasshouse studies of eucalypt seedlings (Drake et al., 2015, 2017). It also provided robust tests of temperature effects in response to diurnal and seasonal temperature changes under ambient and warmed treatments.
Ecotrons can also be used to complement studies in controlled laboratory environments. For example, laboratory experiments investigating patterns of gene expression and resulting physiological processes are often performed under highly constant conditions. However, models need to be developed to translate laboratory knowledge to applications in agriculture. A direct approach incorporating field fluctuations into models that predict transcriptome changes demonstrated an impact of air temperature, humidity, solar radiation and wind (Nagano et al., 2012). Nevertheless, further development of such models would benefit from studying transcriptomes in ecotrons, where fluctuations in the environment can be controlled. This would enable improved parametrization of phenomenological models, such as the ones developed by Nagano et al. (2012) and to develop biophysical ones.
6. FURTHER IMPROVEMENTS
Apart from deploying additional instrumentation for environmental control and process measurements, intrinsic operational improvements of ecotrons are underway. With respect to environmental control, simulating the climate of a distant location or a preceding year remains challenging in sunlit ecotrons, as the natural correlation between temperature and light intensity is difficult to reproduce when local day‐to‐day light conditions are not predictable. Another challenge relates to the incorporation of feedbacks between drought and warming. When drought decreases evapotranspiration, sensible heat flux increases compared to latent heat flux, resulting in landscape‐wide temperature increases (De Boeck & Verbeeck, 2011). This drought impact on temperature should be incorporated into the temperature set‐points of experiments. With many regulated and measured parameters, involving hundreds to thousands of sensors, the early detection of sensor malfunction or deviation from target climatic conditions also remains challenging. This is alleviated by automated alarms on parameter thresholds or by duplicating sensors to indicate sensor drift or failure. However, the use of parameter thresholds may not detect small but deleterious deviations, especially in parameters with large daily fluctuations. Additional alarms based on algorithms taking into account the correlation between replicates and between parameters, as well as on models, need to be tested and deployed to ease experiments human supervision. In terms of improving process measurements, the investigation of soil functioning is lagging behind in ecotrons like in any other experimental facility. Much could come from non‐invasive soil process measurements via automatic gas sampling in the soil. Such sampling is installed in some ecotrons to measure greenhouse gas concentrations, but this needs to be combined with soil physical properties in models calculating soil greenhouse gas emission rates. Furthermore, the analysis of volatile organic compounds (VOCs) emitted from soils via ultra‐sensitive proton transfer reaction mass spectrometers is starting to reveal valuable information on function and biodiversity (e.g. Abis et al., 2018, 2020). The chemotyping of fungi growing on medium can currently be done for some functional groups (Guo et al., 2020; Muller et al., 2013), but a more comprehensive soil VOCs chemotyping, requiring strong international cooperative investments, is needed to advance research on this topic.
A major improvement of ecotron research will also come from a thorough blending of experiments with modelling. Hanson and Walker (2020) wrote ‘From the outset, these studies must be informed by and integrated with ecosystem models that provide quantitative predictions from their embedded mechanistic hypotheses’. Although some ecotron results have been informing model processes (Guderle et al., 2018; Jiang et al., 2020; Milcu et al., 2016), systematic association between models and experiments in ecotrons has not yet been implemented. In particular, recent modelling approaches based on data assimilation and forecasting based on inverse modelling and forward prediction (Huang et al., 2019; Luo et al., 2011) have not been explored with ecotron‐produced data despite ecotrons being ideal for such approaches. Quasi real‐time data assimilation (e.g. once a week or once a month) could be implemented since many ecosystem parameters are acquired at high frequency. This could be used to improve predictions of ecosystem states during the course of experiments in order, for example, to optimize the dates on which to conduct soil or gas samplings for costly and labour‐intensive analyses. Real‐time model outputs could also be used to manage experiments. Often the experimental treatments defined at the start of an experiment (e.g. temperature or precipitation levels) fail to bring the ecosystem into the optimum state to uncover specific ecosystem responses. Real‐time modelling and data assimilation would thus allow to estimate whether or not the ecosystem will reach the target state, and if not, the treatment levels could be adjusted accordingly.
7. PERSPECTIVES
Hanson and Walker (2020) suggested future directions in global change biology, emphasizing the need for large‐scale experiments that incorporate most biochemical and biodiversity feedbacks. They also advocate that a full range of methodological approaches, including smaller spatial scales, will continue to be needed to further mechanistic understanding. This is where the ecotrons will continue to play a significant role, especially through experiments with elevated CO2 atmospheres, anticipated warming or drought scenarios which take us beyond the historical record.
With their versatility and advanced analytical capacities, especially through isotopic approaches, ecotrons should also continue to be used to address not only global change questions but also fundamental questions in ecology, agronomy and environmental science. Moreover, being open to the scientific community at large, we anticipate that new experiments will be created, including those not directly related to ecosystem science (e.g. parametrizing sub‐models of the earth's atmospheric cycles; testing epigenetic effects in specific environments, etc.).
Extended collaborations with other disciplines will be key to take further advantage of the research capacities of ecotrons (Rineau et al., 2019). This will be accomplished in particular through international infrastructure projects. So far, six of the ecotrons described in this paper participate in the Analysis and Experimentation on Ecosystems European infrastructure (ESFRI AnaEE; www.anaee.eu), which is providing a data and modelling centre to facilitate the blending of modelling and experiments. AnaEE, together with ICOS, LTER and LifeWatch, is a constituent infrastructure of the ENVRI consortium (https://envri.eu/), where interactions between research on the life, air, land and water components of the Earth System are developed.
This paper presents the ecotrons, a small part of the national and international efforts to serve environmental research in the context of unprecedented global changes. Its aim is to inform researchers, especially those in the ecology and agronomy fields, about the possibilities offered by these recently built experimental facilities and to encourage their cooperative use. With their high degree of environmental control and exceptional process measurement capacities, the ecotrons described here offer realistic experimental conditions that are much closer to field conditions than those of CEFs in general. Alongside field experiments and observational sites, through their complementary features, the ecotrons can play a pivotal role in uncovering mechanisms and supplying parametrization of ecosystem processes while fostering transnational collaboration. These infrastructures will bring key contributions to the prediction and maintenance of ecosystem services in the context of current environmental changes.
Supporting information
Supplementary Material
Supplementary Material
Supplementary Material
ACKNOWLEDGEMENTS
This study made the most of discussions within the European infrastructure project AnaEE. This study benefited from the CNRS resources allocated to the French ECOTRONS Research Infrastructure, from the Occitanie Region and FEDER investments as well as from the state allocation ‘Investissement d'Avenir’ AnaEE‐France ANR‐11‐INBS‐0001. F.R. thanks the Flemish government (Hercules Foundation big infrastructure and FSR Flanders project G0H4117N), Limburg Sterk Merk (project 271), Hasselt University (project BOF12BR01 and Methusalem project 08M03VGRJ) and the Regional Landscape Kempen and Maasland for funding and support for the building of the UHasselt Ecotron. H.J.D.B. and I.N. were supported by the Research Foundation‐Flanders (FWO, project G0H4117N and I000719N) and the Flemish Government (HERMES Fund).
DATA AVAILABILITY STATEMENT
Data available on request due to privacy/ethical restrictions.
REFERENCES
- Abis, L. , Loubet, B. , Ciuraru, R. , Lafouge, F. , Dequiedt, S. , Houot, S. , Maron, P. A. , & Bourgeteau‐Sadet, S. (2018). Profiles of volatile organic compound emissions from soils amended with organic waste products. Science of The Total Environment, 636, 1333–1343. 10.1016/j.scitotenv.2018.04.232 [DOI] [PubMed] [Google Scholar]
- Abis, L. , Loubet, B. , Ciuraru, R. , Lafouge, F. , Houot, S. , Nowak, V. , Tripied, J. , Dequiedt, S. , Maron, P. A. , & Sadet‐Bourgeteau, S. (2020). Reduced microbial diversity induces larger volatile organic compound emissions from soils. Scientific Reports, 10, 6104. 10.1038/s41598-020-63091-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abiven, S. , Altermatt, F. , Backhaus, N. , Deplazes‐Zemp, A. , Furrer, R. , Korf, B. , Niklaus, P. A. , Schaepman‐Strub, G. , Shimizu, K. K. , Zuppinger‐Dingley, D. , & Petchey, O. L. (2017). Integrative research efforts at the boundary of biodiversity and global change research. Current Opinion in Environmental Sustainability, 29, 215–222. 10.1016/j.cosust.2018.04.016 [DOI] [Google Scholar]
- Arneth, A. , Barbosa, H. , Benton, T. , Calvin, K. , Calvo, E. , Connors, S. , Cowie, A. , Davin, E. , Denton, F. , van Diemen, R. , & Ferrat, M. (2019). IPCC SR: Climate change and land. An IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems. Retrieved from https://www.ipcc.ch/srccl/chapter/summary‐for‐policymakers/ [Google Scholar]
- Arnone III, J. A. , Verburg, P. S. J. , Johnson, D. W. , Larsen, J. D. , Jasoni, R. L. , Lucchesi, A. J. , Batts, C. M. , von Nagy, C. , Coulombe, W. G. , Schorran, D. E. , & Buck, P. E. (2008). Prolonged suppression of ecosystem carbon dioxide uptake after an anomalously warm year. Nature, 455, 383–386. 10.1038/nature07296 [DOI] [PubMed] [Google Scholar]
- Aspinwall, M. J. , Drake, J. E. , Campany, C. , Vårhammar, A. , Ghannoum, O. , Tissue, D. T. , Reich, P. B. , & Tjoelker, M. G. (2016). Convergent acclimation of leaf photosynthesis and respiration to prevailing ambient temperatures under current and warmer climates in Eucalyptus tereticornis . The New Phytologist, 212, 354–367. 10.1111/nph.14035 [DOI] [PubMed] [Google Scholar]
- Aspinwall, M. J. , Pfautsch, S. , Tjoelker, M. G. , Vårhammar, A. , Possell, M. , Drake, J. E. , Reich, P. B. , Tissue, D. T. , Atkin, O. K. , Rymer, P. D. , & Dennison, S. (2019). Range size and growth temperature influence Eucalyptus species responses to an experimental heatwave. Global Change Biology, 25, 1665–1684. 10.1111/gcb.14590 [DOI] [PubMed] [Google Scholar]
- Baggs, E. M. (2008). A review of stable isotope techniques for N2O source partitioning in soils: Recent progress, remaining challenges and future considerations. Rapid Communications in Mass Spectrometry, 22, 1664–1672. [DOI] [PubMed] [Google Scholar]
- Barnard, D. M. , & Bauerle, W. L. (2016). Seasonal variation in canopy aerodynamics and the sensitivity of transpiration estimates to wind velocity in broadleaved deciduous species. Journal of Hydrometeorology, 17, 3029–3043. 10.1175/jhm-d-16-0049.1 [DOI] [Google Scholar]
- Barton, C. V. M. , Ellsworth, D. S. , Medlyn, B. E. , Duursma, R. A. , Tissue, D. T. , Adams, M. A. , Eamus, D. , Conroy, J. P. , McMurtrie, R. E. , Parsby, J. , & Linder, S. (2010). Whole‐tree chambers for elevated atmospheric CO2 experimentation and tree scale flux measurements in south‐eastern Australia: The Hawkesbury Forest Experiment. Agricultural and Forest Meteorology, 150, 941–951. 10.1016/j.agrformet.2010.03.001 [DOI] [Google Scholar]
- Bernier, P. Y. , Stewart, J. D. , & Hogan, G. D. (1994). Quantifying the uncontrolled CO2 dynamics of growth chambers. Journal of Experimental Botany, 45, 1143–1146. 10.1093/jxb/45.8.1143 [DOI] [Google Scholar]
- Braden‐Behrens, J. , Yan, Y. , & Knohl, A. (2017). A new instrument for stable isotope measurements of 13C and 18O in CO2 – Instrument performance and ecological application of the Delta Ray IRIS analyzer. Atmospheric Measurement Techniques, 10, 4537–4560. 10.5194/amt-10-4537-2017 [DOI] [Google Scholar]
- Bradford, M. A. , Jones, T. H. , Bardgett, R. D. , Black, H. I. , Boag, B. , Bonkowski, M. , Cook, R. , Eggers, T. , Gange, A. , & Grayston, S. J. (2002). Impacts of soil faunal community composition on model grassland ecosystems. Science, 298, 615–618. 10.1126/science.1075805 [DOI] [PubMed] [Google Scholar]
- Braendholt, A. , Ibrom, A. , Ambus, P. , Larsen, K. S. , & Pilegaard, K. (2019). Combining a quantum cascade laser spectrometer with an automated closed‐chamber system for δ13C measurements of forest soil, tree stem and tree root CO2 fluxes. Forests, 10, 1–19. [Google Scholar]
- Butterbach‐Bahl, K. , Baggs, E. M. , Dannenmann, M. , Kiese, R. , & Zechmeister‐Boltenstern, S. (2013). Nitrous oxide emissions from soils: How well do we understand the processes and their controls? Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 368, 20130122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Challinor, A. J. , Adger, W. N. , & Benton, T. G. (2017). Climate risks across borders and scales. Nature Climate Change, 7, 621–623. 10.1038/nclimate3380 [DOI] [Google Scholar]
- Clobert, J. , Chanzy, A. , Le, G. J. F. , Chabbi, A. , Greiveldinger, L. , Caquet, T. , Loreau, M. , Mougin, C. , Pichot, C. , Roy, J. , & Saint‐André, L. (2018). How to integrate experimental research approaches in ecological and environmental studies: AnaEE France as an example. Frontiers in Ecology and Evolution, 6, 43. 10.3389/fevo.2018.00043 [DOI] [Google Scholar]
- Coomes, D. A. , Flores, O. , Holdaway, R. , Jucker, T. , Lines, E. R. , & Vanderwel, M. C. (2014). Wood production response to climate change will depend critically on forest composition and structure. Global Change Biology, 20, 3632–3645. 10.1111/gcb.12622 [DOI] [PubMed] [Google Scholar]
- Crous, K. Y. , Quentin, A. G. , Lin, Y. S. , Medlyn, B. E. , Williams, D. G. , Barton, C. V. M. , & Ellsworth, D. S. (2013). Photosynthesis of temperate Eucalyptus globulus trees outside their native range has limited adjustment to elevated CO2 and climate warming. Global Change Biology, 19, 3790–3807. 10.1111/gcb.12314 [DOI] [PubMed] [Google Scholar]
- Crous, K. Y. , Zaragoza‐Castells, J. , Ellsworth, D. S. , Duursma, R. A. , Löw, M. , Tissue, D. T. , & Atkin, O. K. (2012). Light inhibition of leaf respiration in field‐grown Eucalyptus saligna in whole‐tree chambers under elevated atmospheric CO2 and summer drought. Plant, Cell and Environment, 35, 966–981. 10.1111/j.1365-3040.2011.02465.x [DOI] [PubMed] [Google Scholar]
- Day, W. , & Parkinson, K. J. (1979). Importance to gas exchange of mass flow of air through leaves. Plant Physiology, 64, 345–346. 10.1104/pp.64.2.345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Boeck, H. J. , Bloor, J. M. G. , Aerts, R. , Bahn, M. , Beier, C. , Emmett, B. A. , Estiarte, M. , Grünzweig, J. M. , Halbritter, A. H. , Holub, P. , & Jentsch, A. (2020). Understanding ecosystems of the future will require more than realistic climate change experiments – A response to Korell et al. Global Change Biology, 26, e6–e7. 10.1111/gcb.14854 [DOI] [PubMed] [Google Scholar]
- De Boeck, H. J. , & Verbeeck, H. (2011). Drought‐associated changes in climate and their relevance for ecosystem experiments and models. Biogeosciences, 8, 1121–1130. 10.5194/bg-8-1121-2011 [DOI] [Google Scholar]
- De Boeck, H. J. , Vicca, S. , Roy, J. , Nijs, I. , Milcu, A. , Kreyling, J. , Jentsch, A. , Chabbi, A. , Campioli, M. , Callaghan, T. , & Beierkuhnlein, C. (2015). Global change experiments: Challenges and opportunities. BioScience, 65, 922–931. [Google Scholar]
- De Kauwe, M. G. , Medlyn, B. E. , Pitman, A. J. , Drake, J. E. , Ukkola, A. , Griebel, A. , Pendall, E. , Prober, S. , & Roderick, M. (2019). Examining the evidence for decoupling between photosynthesis and transpiration during heat extremes. Biogeosciences, 16, 903–916. 10.5194/bg-16-903-2019 [DOI] [Google Scholar]
- Dell, A. I. , Bender, J. A. , Branson, K. , Couzin, I. D. , de Polavieja, G. G. , Noldus, L. P. J. J. , Pérez‐Escudero, A. , Perona, P. , Straw, A. D. , Wikelski, M. , & Brose, U. (2014). Automated image‐based tracking and its application in ecology. Trends in Ecology & Evolution, 29, 417–428. 10.1016/j.tree.2014.05.004 [DOI] [PubMed] [Google Scholar]
- Díaz, S. , Settele, J. , Brondízio, E. , Ngo, H. T. , Guèze, M. , Agard, J. , Arneth, A. , Balvanera, P. , Brauman, K. , Butchart, S. , & Chan, K. (2019). Summary for policymakers of the global assessment report on biodiversity and ecosystem services. IPBES secretariat, 56 pp. 10.5281/zenodo.3553579 [DOI] [Google Scholar]
- Diaz, S. , Symstad, A. J. , Chapin, F. S. , Wardle, D. A. , & Huenneke, L. F. (2003). Functional diversity revealed by removal experiments. Trends in Ecology & Evolution, 18, 140–146. 10.1016/s0169-5347(03)00007-7 [DOI] [Google Scholar]
- Dombos, M. , Kosztolányi, A. , Szlávecz, K. , Gedeon, C. , Flórián, N. , Groó, Z. , Dudás, P. , & Bánszegi, O. (2017). EDAPHOLOG monitoring system: Automatic, real‐time detection of soil microarthropods. Methods in Ecology and Evolution, 8, 313–321. 10.1111/2041-210x.12662 [DOI] [Google Scholar]
- Drake, J. E. , Aspinwall, M. J. , Pfautsch, S. , Rymer, P. D. , Reich, P. B. , Smith, R. A. , Crous, K. Y. , Tissue, D. T. , Ghannoum, O. , & Tjoelker, M. G. (2015). The capacity to cope with climate warming declines from temperate to tropical latitudes in two widely distributed Eucalyptus species. Global Change Biology, 21, 459–472. [DOI] [PubMed] [Google Scholar]
- Drake, J. E. , Tjoelker, M. G. , Aspinwall, M. J. , Reich, P. B. , Barton, C. V. M. , Medlyn, B. E. , & Duursma, R. A. (2016). Does physiological acclimation to climate warming stabilize the ratio of canopy respiration to photosynthesis? New Phytologist, 211, 850–863. 10.1111/nph.13978 [DOI] [PubMed] [Google Scholar]
- Drake, J. E. , Tjoelker, M. G. , Aspinwall, M. J. , Reich, P. B. , Pfautsch, S. , & Barton, C. V. M. (2019). The partitioning of gross primary production for young Eucalyptus tereticornis trees under experimental warming and altered water availability. New Phytologist, 222, 1298–1312. 10.1111/nph.15629 [DOI] [PubMed] [Google Scholar]
- Drake, J. E. , Tjoelker, M. G. , Vårhammar, A. , Medlyn, B. E. , Reich, P. B. , Leigh, A. , Pfautsch, S. , Blackman, C. J. , López, R. , Aspinwall, M. J. , & Crous, K. Y. (2018). Trees tolerate an extreme heatwave via sustained transpirational cooling and increased leaf thermal tolerance. Global Change Biology, 24, 2390–2402. 10.1111/gcb.14037 [DOI] [PubMed] [Google Scholar]
- Drake, J. E. , Vårhammar, A. , Kumarathunge, D. , Medlyn, B. E. , Pfautsch, S. , Reich, P. B. , Tissue, D. T. , Ghannoum, O. , & Tjoelker, M. G. (2017). A common thermal niche among geographically diverse populations of the widely distributed tree species Eucalyptus tereticornis: No evidence for adaptation to climate‐of‐origin. Global Change Biology, 23, 5069–5082. 10.1111/gcb.13771 [DOI] [PubMed] [Google Scholar]
- Eisenhauer, N. , & Türke, M. (2018). From climate chambers to biodiversity chambers. Frontiers in Ecology and the Environment, 16, 136–137. 10.1002/fee.1784 [DOI] [Google Scholar]
- Fettig, C. J. , Mortenson, L. A. , Bulaon, B. M. , & Foulk, P. B. (2019). Tree mortality following drought in the central and southern Sierra Nevada, California, U.S. Forest Ecology and Management, 432, 164–178. [Google Scholar]
- Füllner, K. , Temperton, V. M. , Rascher, U. , Jahnke, S. , Rist, R. , Schurr, U. , & Kuhn, A. J. (2012). Vertical gradient in soil temperature stimulates development and increases biomass accumulation in barley. Plant, Cell and Environment, 35, 884–892. 10.1111/j.1365-3040.2011.02460.x [DOI] [PubMed] [Google Scholar]
- Galvão, V. C. , & Fankhauser, C. (2015). Sensing the light environment in plants: Photoreceptors and early signaling steps. Current Opinion in Neurobiology, 34, 46–53. 10.1016/j.conb.2015.01.013 [DOI] [PubMed] [Google Scholar]
- García‐Plazaola, J. I. , Fernández‐Marín, B. , Ferrio, J. P. , Alday, J. G. , Hoch, G. , Landais, D. , Milcu, A. , Tissue, D. T. , Voltas, J. , Gessler, A. , & Roy, J. (2017). Endogenous circadian rhythms in pigment composition induce changes in photochemical efficiency in plant canopies. Plant, Cell and Environment, 40, 1153–1162. 10.1111/pce.12909 [DOI] [PubMed] [Google Scholar]
- Gessler, A. , Roy, J. , Kayler, Z. , Ferrio, J. P. , Alday, J. G. , Bahn, M. , del Castillo, J. , Devidal, S. , García‐Muñoz, S. , Landais, D. , & Martín‐Gomez, P. (2017). Night and day – Circadian regulation of night‐time dark respiration and light‐enhanced dark respiration in plant leaves and canopies. Environmental and Experimental Botany, 137, 14–25. 10.1016/j.envexpbot.2017.01.014 [DOI] [Google Scholar]
- Ghirardo, A. , Lindstein, F. , Koch, K. , Buegger, F. , Schloter, M. , Albert, A. , Michelsen, A. , Winkler, J. B. , Schnitzler, J. P. , & Rinnan, R. (2020). Origin of volatile organic compound emissions from subarctic tundra under global warming. Global Change Biology, 26, 1908–1925. 10.1111/gcb.14935 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Granjou, C. , & Walker, J. (2016). Promises that matter: Reconfiguring ecology in the ecotrons. Science and Technology Studies, 29, 49–67. [Google Scholar]
- Griffin, K. L. , Ross, P. D. , Sims, D. A. , Luo, Y. , Seemann, J. R. , Fox, C. A. , & Ball, J. T. (1996). EcoCELLs: Tools for mesocosm scale measurements of gas exchange. Plant, Cell and Environment, 19, 1210–1221. 10.1111/j.1365-3040.1996.tb00437.x [DOI] [PubMed] [Google Scholar]
- Groh, J. , Vanderborght, J. , Pütz, T. , & Vereecken, H. (2016). How to control the lysimeter bottom boundary to investigate the effect of climate change on soil processes? Vadose Zone Journal, 15, vzj2015.08.0113. 10.2136/vzj2015.08.0113 [DOI] [Google Scholar]
- Guderle, M. , Bachmann, D. , Milcu, A. , Gockele, A. , Bechmann, M. , Fischer, C. , Roscher, C. , Landais, D. , Ravel, O. , Devidal, S. , & Roy, J. (2018). Dynamic niche partitioning in root water uptake facilitates efficient water use in more diverse grassland plant communities. Functional Ecology, 32, 214–227. 10.1111/1365-2435.12948 [DOI] [Google Scholar]
- Guo, Y. , Jud, W. , Ghirardo, A. , Antritter, F. , Benz, J. P. , Schnitzler, J. , & Rosenkranz, M. (2020). Sniffing fungi – Phenotyping of volatile chemical diversity in Trichoderma species. New Phytologist, 227, 244–259. 10.1111/nph.16530 [DOI] [PubMed] [Google Scholar]
- Hallmann, C. A. , Sorg, M. , Jongejans, E. , Siepel, H. , Hofland, N. , Schwan, H. , Stenmans, W. , Müller, A. , Sumser, H. , Hörren, T. , & Goulson, D. (2017). More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS One, 12, e0185809. 10.1371/journal.pone.0185809 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanson, P. J. , & Walker, A. P. (2020). Advancing global change biology through experimental manipulations: Where have we been and where might we go? Global Change Biology, 26, 287–299. 10.1111/gcb.14894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harwood, K. G. , Gillon, J. S. , Roberts, A. , & Griffiths, H. (1999). Determinants of isotopic coupling of CO2 and water vapour within a Quercus petraea forest canopy. Oecologia, 119, 109–119. 10.1007/s004420050766 [DOI] [PubMed] [Google Scholar]
- Hoover, D. L. , Wilcox, K. R. , & Young, K. E. (2018). Experimental droughts with rainout shelters: A methodological review. Ecosphere, 9, 1. 10.1002/ecs2.2088 [DOI] [Google Scholar]
- Huang, Y. , Stacy, M. , Jiang, J. , Sundi, N. , Ma, S. , Saruta, V. , Jung, C. G. , Shi, Z. , Xia, J. , Hanson, P. J. , & Ricciuto, D. (2019). Realized ecological forecast through an interactive Ecological Platform for Assimilating Data (EcoPAD, v1.0) into models. Geoscientific Model Development, 12, 1119–1137. 10.5194/gmd-12-1119-2019 [DOI] [Google Scholar]
- Jiang, M. , Medlyn, B. E. , Drake, J. E. , Duursma, R. A. , Anderson, I. C. , Barton, C. V. M. , Boer, M. M. , Carrillo, Y. , Castañeda‐Gómez, L. , Collins, L. , & Crous, K. Y. (2020). The fate of carbon in a mature forest under carbon dioxide enrichment. Nature, 580, 227–231. [DOI] [PubMed] [Google Scholar]
- Kolb, T. , Keefover‐Ring, K. , Burr, S. J. , Hofstetter, R. , Gaylord, M. , & Raffa, K. F. (2019). Drought‐mediated changes in tree physiological processes weaken tree defenses to bark beetle attack. Journal of Chemical Ecology, 45, 888–900. 10.1007/s10886-019-01105-0 [DOI] [PubMed] [Google Scholar]
- Kreyling, J. , Jentsch, A. , & Beier, C. (2014). Beyond realism in climate change experiments: Gradient approaches identify thresholds and tipping points. Ecology Letters, 17, 10–11. 10.1111/ele.12193 [DOI] [PubMed] [Google Scholar]
- Kreyling, J. , Schweiger, A. H. , Bahn, M. , Ineson, P. , Migliavacca, M. , Morel‐Journel, T. , Christiansen, J. R. , Schtickzelle, N. , & Larsen, K. S. (2018). To replicate, or not to replicate – That is the question: How to tackle nonlinear responses in ecological experiments. Ecology Letters, 21, 1629–1638. 10.1111/ele.13134 [DOI] [PubMed] [Google Scholar]
- Lange, M. , Eisenhauer, N. , Sierra, C. A. , Bessler, H. , Engels, C. , Griffiths, R. I. , Mellado‐Vázquez, P. G. , Malik, A. A. , Roy, J. , Scheu, S. , & Steinbeiss, S. (2015). Plant diversity increases soil microbial activity and soil carbon storage. Nature Communications, 6, 6707. 10.1038/ncomms7707 [DOI] [PubMed] [Google Scholar]
- Lawton, J. H. (1993). The Ecotron: A controlled environmental facility for the investigation of population and ecosystem processes. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 341, 181–194. 10.1098/rstb.1993.0102 [DOI] [Google Scholar]
- Lawton, J. H. (1995). Ecological experiments with model systems. Science, 269, 328–331. 10.1126/science.269.5222.328 [DOI] [PubMed] [Google Scholar]
- Lawton, J. H. , Naeem, S. , Woodfin, R. , Brown, V. K. , Gange, A. , Godfray, H. J. C. , Heads, P. A. , Lawler, S. , Magda, D. , Thomas, C. , & Thompson, L. J. (1993). The Ecotron: A controlled environmental facility for the investigation of population and ecosystem processes. Philosophical Transactions of the Royal Society B: Biological Sciences, 341, 181–194. 10.1098/rstb.1993.0102 [DOI] [Google Scholar]
- Leuzinger, S. , & Körner, C. (2007). Water savings in mature deciduous forest trees under elevated CO2 . Global Change Biology, 13, 2498–2508. 10.1111/j.1365-2486.2007.01467.x [DOI] [Google Scholar]
- Liu, L. , Hoogenboom, G. , & Ingram, K. T. (2000). Controlled‐environment sunlit plant growth chambers. Critical Reviews in Plant Sciences, 19, 347–375. 10.1080/07352680091139268 [DOI] [Google Scholar]
- Luo, Y. , Ogle, K. , Tucker, C. , Fei, S. , Gao, C. , LaDeau, S. , Clark, J. S. , & Schimel, D. S. (2011). Ecological forecasting and data assimilation in a data‐rich era. Ecological Applications, 21, 1429–1442. 10.1890/09-1275.1 [DOI] [PubMed] [Google Scholar]
- Marini, L. , Økland, B. , Jönsson, A. M. , Bentz, B. , Carroll, A. , Forster, B. , Grégoire, J. C. , Hurling, R. , Nageleisen, L. M. , Netherer, S. , & Ravn, H. P. (2017). Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography, 40, 1426–1442. 10.1111/ecog.02769 [DOI] [Google Scholar]
- Massonnet, C. , Vile, D. , Fabre, J. , Hannah, M. A. , Caldana, C. , Lisec, J. , Beemster, G. T. S. , Meyer, R. C. , Messerli, G. , Gronlund, J. T. , & Perkovic, J. (2010). Probing the reproducibility of leaf growth and molecular phenotypes: A comparison of three Arabidopsis accessions cultivated in ten laboratories. Plant Physiology, 152, 2142–2157. 10.1104/pp.109.148338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mauser, W. , Klepper, G. , Rice, M. , Schmalzbauer, B. S. , Hackmann, H. , Leemans, R. , & Moore, H. (2013). Transdisciplinary global change research: The co‐creation of knowledge for sustainability. Current Opinion in Environmental Sustainability, 5, 420–431. 10.1016/j.cosust.2013.07.001 [DOI] [Google Scholar]
- McManus, J. B. , Nelson, D. D. , & Zahniser, M. S. (2015). Design and performance of a dual‐laser instrument for multiple isotopologues of carbon dioxide and water. Optics Express, 23, 6569. 10.1364/oe.23.006569 [DOI] [PubMed] [Google Scholar]
- Mellado‐Vázquez, P. G. , Lange, M. , Bachmann, D. , Gockele, A. , Karlowsky, S. , Milcu, A. , Piel, C. , Roscher, C. , Roy, J. , & Gleixner, G. (2016). Plant diversity generates enhanced soil microbial access to recently photosynthesized carbon in the rhizosphere. Soil Biology and Biochemistry, 94, 122–132. 10.1016/j.soilbio.2015.11.012 [DOI] [Google Scholar]
- Milcu, A. , Eugster, W. , Bachmann, D. , Guderle, M. , Röscher, C. , Gockele, A. , Landais, D. , Ravel, O. , Gessler, A. , Ebeling, A. , Weisser, W. W. , Roy, J. , Hildebrandt, A. , & Buchmann, N. (2016). Plant functional diversity increases grassland productivity‐ related water vapor fluxes: An ecotron and modeling approach. Ecology, 97, 2044–2054. 10.1890/15-1110.1 [DOI] [PubMed] [Google Scholar]
- Milcu, A. , Lukac, M. , Subke, J. A. , Manning, P. , Heinemeyer, A. , Wildman, D. , Anderson, R. , & Ineson, P. (2012). Biotic carbon feedbacks in a materially closed soil‐vegetation‐atmosphere system. Nature Climate Change, 2, 281–284. [Google Scholar]
- Milcu, A. , Puga‐Freitas, R. , Ellison, A. M. , Blouin, M. , Scheu, S. , Freschet, G. T. , Rose, L. , Barot, S. , Cesarz, S. , Eisenhauer, N. , Girin, T. , Assandri, D. , Bonkowski, M. , Buchmann, N. , Butenschoen, O. , Devidal, S. , Gleixner, G. , Gessler, A. , Gigon, A. , … Roy, J. (2018). Genotypic variability enhances the reproducibility of an ecological study. Nature Ecology and Evolution, 2, 279–287. 10.1038/s41559-017-0434-x [DOI] [PubMed] [Google Scholar]
- Milcu, A. , Roscher, C. , Gessler, A. , Bachmann, D. , Gockele, A. , Guderle, M. , Landais, D. , Piel, C. , Escape, C. , Devidal, S. , & Ravel, O. (2014). Functional diversity of leaf nitrogen concentrations drives grassland carbon fluxes. Ecology Letters, 17, 435–444. 10.1111/ele.12243 [DOI] [PubMed] [Google Scholar]
- Misson, L. , Degueldre, D. , Collin, C. , Rodriguez, R. , Rocheteau, A. , Ourcival, J. M. , & Rambal, S. (2011). Phenological responses to extreme droughts in a Mediterranean forest. Global Change Biology, 17, 1036–1048. 10.1111/j.1365-2486.2010.02348.x [DOI] [Google Scholar]
- Möller, B. , Chen, H. , Schmidt, T. , Zieschank, A. , Patzak, R. , Türke, M. , Weigelt, A. , & Posch, S. (2019). RhizoTrak: A flexible open source Fiji plugin for user‐friendly manual annotation of time‐series images from minirhizotrons. Plant and Soil, 444, 519–534. 10.1007/s11104-019-04199-3 [DOI] [Google Scholar]
- Moreno, J. E. , Tao, Y. , Chory, J. , & Ballaré, C. L. (2009). Ecological modulation of plant defense via phytochrome control of jasmonate sensitivity. Proceedings of the National Academy of Sciences of the United States of America, 106, 4935–4940. 10.1073/pnas.0900701106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muller, A. , Faubert, P. , Hagen, M. , zu Castell, W. , Polle, A. , Schnitzler, J. P. , & Rosenkranz, M. (2013). Volatile profiles of fungi – Chemotyping of species and ecological functions. Fungal Genetics and Biology, 54, 25–33. 10.1016/j.fgb.2013.02.005 [DOI] [PubMed] [Google Scholar]
- Nagano, A. J. , Sato, Y. , Mihara, M. , Antonio, B. A. , Motoyama, R. , Itoh, H. , Nagamura, Y. , & Izawa, T. (2012). Deciphering and prediction of transcriptome dynamics under fluctuating field conditions. Cell, 151, 1358–1369. 10.1016/j.cell.2012.10.048 [DOI] [PubMed] [Google Scholar]
- Neubauer, S. C. , & Megonigal, J. P. (2015). Moving beyond global warming potentials to quantify the climatic role of ecosystems. Ecosystems, 18, 1000–1013. 10.1007/s10021-015-9879-4 [DOI] [Google Scholar]
- Norby, R. J. , De Kauwe, M. G. , Domingues, T. F. , Duursma, R. A. , Ellsworth, D. S. , Goll, D. S. , Lapola, D. M. , Luus, K. A. , MacEnzie, A. R. , Medlyn, B. E. , Pavlick, R. , Rammig, A. , Smith, B. , Thomas, R. , Thonicke, K. , Walker, A. P. , Yang, X. , & Zaehle, S. (2016). Model‐data synthesis for the next generation of forest free‐air CO2 enrichment (FACE) experiments. New Phytologist, 209, 17–28. 10.1111/nph.13593 [DOI] [PubMed] [Google Scholar]
- Oerter, E. J. , & Bowen, G. (2017). In situ monitoring of H and O stable isotopes in soil water reveals ecohydrologic dynamics in managed soil systems. Ecohydrology, 10, 1–13. 10.1002/eco.1841 [DOI] [Google Scholar]
- Payer, H.‐D. , Blank, L. W. , Bosch, C. , Gnatz, G. , Schmolke, W. , & Schramel, P. (1986). Simultaneous exposure of forest trees to pollutants and climatic stress. Water, Air, and Soil Pollution, 31, 485–491. 10.1007/bf00630866 [DOI] [Google Scholar]
- Pecl, G. T. , Araújo, M. B. , Bell, J. D. , Blanchard, J. , Bonebrake, T. C. , Chen, I. C. , Clark, T. D. , Colwell, R. K. , Danielsen, F. , Evengård, B. , & Falconi, L. (2017). Biodiversity redistribution under climate change: Impacts on ecosystems and human well‐being. Science, 355, 6332. [DOI] [PubMed] [Google Scholar]
- Poorter, H. , Bühler, J. , Van Dusschoten, D. , Climent, J. , & Postma, J. A. (2012). Pot size matters: A meta‐analysis of the effects of rooting volume on plant growth. Functional Plant Biology, 39, 839–850. 10.1071/fp12049 [DOI] [PubMed] [Google Scholar]
- Poorter, H. , Fiorani, F. , Pieruschka, R. , Wojciechowski, T. , van der Putten, W. H. , Kleyer, M. , Schurr, U. , & Postma, J. (2016). Pampered inside, pestered outside? Differences and similarities between plants growing in controlled conditions and in the field. New Phytologist, 212, 838–855. 10.1111/nph.14243 [DOI] [PubMed] [Google Scholar]
- Poorter, H. , Niinemets, Ü. , Ntagkas, N. , Siebenkäs, A. , Mäenpää, M. , Matsubara, S. , & Pons, T. L. (2019). A meta‐analysis of plant responses to light intensity for 70 traits ranging from molecules to whole plant performance. New Phytologist, 223, 1073–1105. 10.1111/nph.15754 [DOI] [PubMed] [Google Scholar]
- Porter, A. S. , Evans‐Fitz Gerald, C. , McElwain, J. C. , Yiotis, C. , & Elliott‐Kingston, C. (2015). How well do you know your growth chambers? Testing for chamber effect using plant traits. Plant Methods, 11, 1–10. 10.1186/s13007-015-0088-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Potvin, C. , & Tardif, S. (1988). Sources of variability and experimental designs in growth chambers. Functional Ecology, 2, 123–130. 10.2307/2389472 [DOI] [Google Scholar]
- Resco de Dios, V. , Gessler, A. , Ferrio, J. P. , Alday, J. , Bahn, M. , del Castillo, J. , Devidal, S. , García‐Muñoz, S. , Kayler, Z. , Landais, D. , & Martín‐Gómez, P. (2017). Circadian rhythms regulate the environmental responses of net CO2 exchange in bean and cotton canopies. Agricultural and Forest Meteorology, 239, 185–191. 10.1016/j.agrformet.2017.03.014 [DOI] [Google Scholar]
- Resco de Dios, V. , Gessler, A. , Ferrio, J. P. , Alday, J. G. , Bahn, M. , del Castillo, J. , Devidal, S. , García‐Muñoz, S. , Kayler, Z. , Landais, D. , & Martín‐Gómez, P. (2016). Circadian rhythms have significant effects on leaf‐to‐canopy scale gas exchange under field conditions. GigaScience, 5, 43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rineau, F. , Malina, R. , Beenaerts, N. , Arnauts, N. , Bardgett, R. D. , Berg, M. P. , Boerema, A. , Bruckers, L. , Clerinx, J. , Davin, E. L. , & De Boeck, H. J. (2019). Towards more predictive and interdisciplinary climate change ecosystem experiments. Nature Climate Change, 9, 809–816. [Google Scholar]
- Romer, M. (2001). Carbon dioxide within controlled environments the commonly neglected variable. In Proceedings of the International Conference: Controlled environments in the New Millenium, Norwich, United Kingdom. Retrieved from https://ncera.phytotron.ncsu.edu/wp‐content/uploads/sites/6/2017/06/pa‐romer.pdf [Google Scholar]
- Roscher, C. , Karlowsky, S. , Milcu, A. , Gessler, A. , Bachmann, D. , Jesch, A. , Lange, M. , Mellado‐Vázquez, P. , Strecker, T. , Landais, D. , & Ravel, O. (2019). Functional composition has stronger impact than species richness on carbon gain and allocation in experimental grasslands. PLoS One, 14, e0204715. 10.1371/journal.pone.0204715 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy, J. , Picon‐Cochard, C. , Augusti, A. , Benot, M.‐L. , Thiery, L. , Darsonville, O. , Landais, D. , Piel, C. , Defossez, M. , Devidal, S. , & Escape, C. (2016). Elevated CO2 maintains grassland net carbon uptake under a future heat and drought extreme. Proceedings of the National Academy of Sciences of the United States of America, 113, 6224–6229. 10.1073/pnas.1524527113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheffers, B. R. , De Meester, L. , Bridge, T. C. L. , Hoffmann, A. A. , Pandolfi, J. M. , Corlett, R. T. , Butchart, S. H. M. , Pearce‐Kelly, P. , Kovacs, K. M. , Dudgeon, D. , & Pacifici, M. (2016). The broad footprint of climate change from genes to biomes to people. Science, 354(6313). 10.1126/science.aaf7671 [DOI] [PubMed] [Google Scholar]
- Schwalm, C. R. , Williams, C. A. , Schaefer, K. , Arneth, A. , Bonal, D. , Buchmann, N. , Chen, J. , Law, B. , Lindroth, A. , Luyssaert, S. , & Reichstein, M. (2010). Assimilation exceeds respiration sensitivity to drought: A FLUXNET synthesis. Global Change Biology, 16, 657–670. 10.1111/j.1365-2486.2009.01991.x [DOI] [Google Scholar]
- Sigurdsson, B. D. , Leblans, N. I. W. , Dauwe, S. , Gudmundsdóttir, E. , Gundersen, P. , Gunnarsdóttir, G. E. , Holmstrup, M. , Ilieva‐Makulec, K. , Kätterer, T. , Marteinsdóttir, B. , & Maljanen, M. (2016). Geothermal ecosystems as natural climate change experiments: The ForHot research site in Iceland as a case study. Icelandic Agricultural Sciences, 29, 53–71. 10.16886/ias.2016.05 [DOI] [Google Scholar]
- Stevnbak, K. , Scherber, C. , Gladbach, D. J. , Beier, C. , Mikkelsen, T. N. , & Christensen, S. (2012). Interactions between above‐ and belowground organisms modified in climate change experiments. Nature Climate Change, 2, 805–808. 10.1038/nclimate1544 [DOI] [Google Scholar]
- Stewart, R. I. A. , Dossena, M. , Bohan, D. A. , Jeppesen, E. , Kordas, R. L. , Ledger, M. E. , Meerhoff, M. , Moss, B. , Mulder, C. , Shurin, J. B. , & Suttle, B. (2013). Mesocosm experiments as a tool for ecological climate‐change research. Advances in Ecological Research, 48, 71–181. 10.1016/b978-0-12-417199-2.00002-1 [DOI] [Google Scholar]
- Tan, C. , Du, Y. , Zhou, J. , Wang, D. , Luo, M. , Zhang, Y. , & Guo, W. (2018). Analysis of different hyperspectral variables for diagnosing leaf nitrogen accumulation in wheat. Frontiers in Plant Science, 9, 674. 10.3389/fpls.2018.00674 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thakur, M. P. , Künne, T. , Unsicker, S. B. , Biere, A. , Ferlian, O. , Pruschitzki, U. , Thouvenot, L. , Türke, M. , & Eisenhauer, N. (2020). Invasive earthworms reduce chemical defense and increase herbivory and pathogen infection in native trees. Journal of Ecology, 10, 1111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas, M. D. , & Hill, G. R. (1937). The continuous measurement of photosynthesis, respiration, and transpiration of alfalfa and wheat browing under field conditions. Plant Physiology, 12, 285–307. 10.1104/pp.12.2.285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulm, R. , & Jenkins, G. I. (2015). Q&A: How do plants sense and respond to UV‐B radiation? BMC Biology, 13, 4–9. 10.1186/s12915-015-0156-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulrich, J. , Bucher, S. , Eisenhauer, N. , Schmidt, A. , Türke, M. , Gebler, A. , Barry, K. , Lange, M. , & Römermann, C. (2020). Invertebrate decline leads to shifts in plant species abundance and phenology. Frontiers in Plant Science, 11, 542125. 10.3389/fpls.2020.542125 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van De Velde, H. , Nijs, I. , & Bonte, D. (2017). Warming affects different components of plant–herbivore interaction in a simplified community but not net interaction strength. Oikos, 126(2). 10.1111/oik.03415 [DOI] [Google Scholar]
- Van der Putten, W. H. , Bardgett, R. D. , Bever, J. D. , Bezemer, T. M. , Casper, B. B. , Fukami, T. , Kardol, P. , Klironomos, J. N. , Kulmatiski, A. , Schweitzer, J. A. , & Suding, K. N. (2013). Plant‐soil feedbacks: The past, the present and future challenges. Journal of Ecology, 101, 265–276. [Google Scholar]
- Verdaguer, D. , Jansen, M. A. K. , Llorens, L. , Morales, L. O. , & Neugart, S. (2017). UV‐A radiation effects on higher plants: Exploring the known unknown. Plant Science, 255, 72–81. 10.1016/j.plantsci.2016.11.014 [DOI] [PubMed] [Google Scholar]
- Weisser, W. W. , Roscher, C. , Meyer, S. T. , Ebeling, A. , Luo, G. , Allan, E. , Beßler, H. , Barnard, R. L. , Buchmann, N. , Buscot, F. , & Engels, C. (2017). Biodiversity effects on ecosystem functioning in a 15‐year grassland experiment: Patterns, mechanisms, and open questions. Basic and Applied Ecology, 23, 1–73. [Google Scholar]
- Wheeler, T. , & Von Braun, J. (2013). Climate change impacts on global food security. Science, 341, 508–513. [DOI] [PubMed] [Google Scholar]
- Xie, Y. , Wang, C. , Yang, W. , Feng, M. , Qiao, X. , & Song, J. (2020). Canopy hyperspectral characteristics and yield estimation of winter wheat (Triticum aestivum) under low temperature injury. Scientific Reports, 10, 244. 10.1038/s41598-019-57100-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Material
Supplementary Material
Supplementary Material
Data Availability Statement
Data available on request due to privacy/ethical restrictions.