

Abstract
The pathology and physiology of breast cancer (BC), including metastasis, and drug resistance, is driven by multiple signaling pathways in the tumor microenvironment (TME), which hamper antitumor immunity. Recently, long non-coding RNAs have been reported to mediate pathophysiological develop-ments such as metastasis as well as immune suppression within the TME. Given the complex biology of BC, novel personalized therapeutic strategies that address its diverse pathophysiologies are needed to improve clinical outcomes. In this review, we describe the advances in the biology of breast neoplasia, including cellular and molecular biology, heterogeneity, and TME. We review the role of novel molecules such as long non-coding RNAs in the pathophysiology of BC. Finally, we provide an up-to-date overview of anticancer compounds extracted from marine microorganisms, crustaceans, and fishes and their synergistic effects in combination with other anticancer drugs. Marine compounds are a new discipline of research in BC and offer a wide range of anti-cancer effects that could be harnessed to target the various pathways involved in BC development, thus assisting current therapeutic regimens.
Keywords: Breast cancer, Tumor microenvironment, Long non-coding RNAs, Growth, Metastasis, Angiogenesis, Marine compounds
Core Tip: Breast cancer (BC) is an aggressive and heterogenous disease. The BC tumor microenvironment contributes to immune evasion and chemoresistance in BC. Long non-coding RNAs contribute to BC pathophysiology and are potential BC biomarkers. Marine compounds display promising anticancer activity against BC. The use of novel bioengineering tools will enormously help in improving their production and clinical development for the treatment of BC and other cancers.
INTRODUCTION
According to GLOBOCAN (2018), 2.1 million females were diagnosed with breast cancer (BC) in 2018, accounting for 11.6 % of total cancer patients. As per this report, BC accounts for nearly 1 in 4 cancer cases among women, illustrating its high frequency. In fact, the incidence of BC ranks first among women-specific cancers worldwide. The elevated prevalence of BC compared to other cancers is due to its risk factors associated with menstruation, reproduction, intake of hormones, nutrition, and anthropometry, postponement of childbearing, etc. Additionally, the prevalence rate of BC increases in successive generations of high-risk populations due to inherited genetic predisposition[1].
The characterization of pathophysiology and treatment of BC is highly challenging due to its heterogeneous nature. BC displays rapid visceral as well as distant metastatic ability. Some BC subtypes can be highly aggressive and exhibit unique proliferation markers, endothelial growth factor receptor (EGFR) and endocrine receptors. They differ in metabolism, cell proliferation, protein synthesis, and communication between cells. The combined action of mutations, as well as changes in copy number, define the type of BC, while alterations in epigenetic regulation primarily initiate the development of cancer[2]. BC patients develop drug resistance, recurrence, or therapy failure. The high mortality of BC patients is mainly due to aggressive nature, heterogeneity, metastasis, drug resistance, and recurrence as well as late diagnosis.
Breast tumors consist of cancer cells, surrounding stromal cells, and blood cells in the tumor microenvironment (TME), which are critical for cancer progression and metastasis. For example, suppressor immune cells, soluble factors, and remodeled extracellular matrix (ECM) collectively hamper the anti-tumor immunity but help metastasis. Stromal cells with altered molecular mechanisms as well as abnormal signaling pathways and their interplay with other components of the TME are associated with poor prognosis of BC. Additionally, the ECM can induce changes in biochemical and molecular mechanisms. Furthermore, the molecular signature of the stroma-ECM influences BC progression as well as impacts the drug resistance cancer phenotypes towards hormone-based as well as cytotoxic based therapeutics. Similarly, the cancer-associated stroma (CAS) overexpresses genes encoding ECM, cell cycle-related proteins, matrix metalloproteinases (MMPs) (MMP-2, -11 and -14), and mitochondrial ribosomal proteins during the change to invasive phenotype from pre-invasive phenotype. CAS also correlates with enhanced expression of genes related to immune response[3]. The molecular mechanisms promoting obesity-associated changes in the TME support BC growth by altering functions of adipose tissue, inducing adipocyte demise, and imposing low-grade chronic inflammation. The inflamed TME allows remodeling and infiltration of immune cells and stimulates protumor after-effects to sustain irreversible metastatic progression[4].
Tumor angiogenesis is a hallmark of solid tumors and an alternative target of metastatic BC. Metastatic breast tumors create unusual vascularization around the tumor by secreting abnormal levels of proangiogenic factors. The vascular network in tumors is highly disorganized and permeable. The abnormal vascularization leads to the development of poorly perfused tumors in the hypoxic TME. This hypoxic environment selects highly invasive and aggressive cancer cells with the ability to escape from tumor-destroying immune cells for metastasis. Furthermore, vascular endothelial growth factor (VEGF) increases angiogenesis by suppressing immunity. In addition, abnormal perfusion decreases the efficacy of chemotherapy and radiation therapy. Apart from cancer cells, tumor-associated leukocytes and stromal cells also promote tumor angiogenesis by secreting extracellular proteases, chemokines, and cytokines as well as exosomes.
Given the complex biology of BC, novel personalized therapeutic strategies that address its diverse pathophysiologies are needed to improve clinical outcomes. Marine sources offer tremendous potential for the detection and advancement of new therapeutic regimens that can assist in BC treatment. Since 1980, advances in biotechnology have enabled new avenues of research exploring marine organisms and their therapeutic potential for novel drug development. This research is dynamic and utilizes advanced technological tools[5], and has led to an increased interest in exploiting marine microbial, crustacean, and natural fish product scaffolds for rational drug discovery. Marine bioactive compounds represent an accessible source of chemical entities, novel drugs as well as drug leads. They are the basis for nearly 80% of the chemotherapeutics approved by the United States Food and Drug Administration (FDA), and more than half of all drugs. Marine compounds are currently used for the treatment of 87% of diseases, including cancer as they can induce cytotoxicity in cancer cells by targeting mediators of oncogenic transduction pathways or by inhibiting the growth of cancer cells[6,7]. Recent advances in technology and research focusing on marine compounds have allowed the discovery of novel anticancer agents, which are currently undergoing clinical trials.
In the present review, we describe advances in the biology of breast neoplasia, including cellular and molecular biology and heterogeneity as well as epidemiology. Furthermore, we summarize recent knowledge of BC pathophysiology, including the role of TME components in metastasis, angiogenesis, and drug resistance. We then provide an up-to-date review of anticancer compounds originating from marine microorganisms, crustaceans, and fishes and their synergistic effect on anticancer drugs. Finally, we present a summary of the pathophysiology of BC and delineate future directions in the development of marine-based therapeutics for BC.
BC BIOLOGY
The biology of BC encompasses a unique signature of histopathology, cellular and molecular heterogeneity and responses, which allows a deeper understanding of BC pathophysiology. The biology of breast tumors in women varies with age. BC in younger women is highly complex and accounts for 15% of all BC subtypes in developing countries. Breast tumors in younger women are classified as high-frequency subtypes with unique signatures of histopathology, genetics, molecular biology, and genomics. Substantial literature validates the function of patient immunosurveillance in affecting response to treatment in some subtypes of BC like triple-negative breast cancer (TNBC) and human epidermal growth factor receptor 2 (HER2)+, but its role in estrogen receptor (ER)+ remains unclear because it is less immunogenic[8], highlighting the need for in-depth clinical studies to improving management strategies in ER+ BC.
AGE SIGNATURE OF BC PATIENTS
BC in younger women exhibits differential gene expression, definite genomic signs, and elevated genetic susceptibility, compared to aged females with BC[9]. In addition, differences in breast stroma, changes in the breast during pregnancy or breastfeeding contribute to the difference in the biology of tumors in younger women[10]. Furthermore, BCs in younger women are multicentric, grade 3, triple-negative, and have a high proliferative index, Ki-67[11]. Furthermore, radiation treatment following BCS can decrease local recurrence from 19.5% to 10.2% in younger BC patients[12]. Clinical investigations evaluating five years of endocrine therapy in TEXT and SOFT trials and extended therapy with tamoxifen in ATTom and ATLAS trials suggest that adjuvant endocrine therapy is an effective treatment for younger BC patients[13].
BCs in adolescents account for 5.6% of all invasive BC and are likely to be driven by genes responsible for familial cancer predisposition, which cause larger tumors with distinct metastasis. These patients need a multidisciplinary approach of care and treatment regimens to minimize premature menopause, as well as osteoporosis and sexual health[14]. In aged women, BCs found at an initial stage are treatable and have prolonged disease-free survival. Furthermore, adjuvant chemotherapy, radiation or endocrine therapy can reduce the danger of BC recurrence and improve survival[15]. Population-based studies conducted in Asia have recognized that adjuvant modalities, including endocrine and chemotherapy regimens, have significantly improved the overall survival of older women with BC[16], thus illustrating the beneficial role of clinical studies in ameliorating patient outcomes.
CELLULAR AND MOLECULAR BIOLOGY OF BC
The role of pathological examination in predicting etiology, pathogenesis, and clinicopathological correlations of BCs is the gold standard. Despite many biochemical and histochemical biomarkers, ER, progesterone receptor (PR), and HER-2 are the vital components of clinical examination. Initially, these markers were used for the prognosis of BCs, but also for prediction of treatment outcome. However, high throughput experimentation shows molecular heterogeneity in histologically similar subtypes. For example, high throughput data has facilitated the classification of invasive ductal carcinoma into four categories such as luminal, HER2 amplification type, basal-like and normal breast-like. Among the subtypes, 75% of BCs belong to the luminal type, which exhibits highly complex endocrine resistance due to genetic and epigenetic mechanisms. Epigenetic changes in receptors or their signaling pathways, such as the pattern of DNA methylation, histone modifications, and altered expression of miRNAs, are heritable and reversible in luminal BCs[17]. Lobular carcinoma of invasive (ILC) type of BC accounts for 5%-15% of ductal carcinoma of invasive type. This subtype shows an endocrine response but responds poorly to chemotherapy. Some studies suggest that systemic therapy might be useful for patients with ILC[18].
Genomic analysis has identified seven BC epitypes (ET1 to ET7). One subtype is characterized by BRCA1 mutations and is linked to basal-like tumors, while another subtype has a complex genome with several additional amplifications such as ERBB2 amplification. A third subtype exhibits a normal epithelial cell-like methylation profile. In contrast, the remaining four epigenetic subtypes look like luminal BCs but differ in genomic instability, promoter, and global hypermethylation as well as the rate of proliferation[19]. Furthermore, high throughput studies have uncovered an interaction between genetic and epigenetic alterations among BC subtypes[20]. The diverse and complex biology of these various BC subtypes highlights the need for personalized targeted therapies that address the specificities of each subtype to enhance patient outcomes. In an effort to identify new biomarkers of each subtype, a microarray study by Zheng et al[21] found differentially expressed genes associated with immune response, cell differentiation, and cell adhesion in various BC types. The genes engage in a protein-protein network with EGFR and spleen-associated tyrosine kinase, which are critical for BC metastasis, and could thus serve as promising novel biomarkers[21] that could improve early BC diagnosis.
CELLULAR AND MOLECULAR HETEROGENEITY OF BC
As the previous section shows, breast tumor consists of genetically, pathologically, and clinically distinct subtypes. Based on the characteristics of histopathology, BC can be classified into three subtypes, including BCs positive for ER/PR, TNBC, and amplified with HER2. TNBCs account for 15% of all BCs, enriched with heterogeneity in terms of BRCA1/2 mutations, expression of androgen receptor (AR) along with rare genomic changes, and a high degree of lymphocyte infiltration. Furthermore, gene expression studies have demonstrated six clinically distinct subsets of TNBC, counting basal-like-1 and -2, immunomodulatory, mesenchymal, luminal AR and mesenchymal stem-like[3]. Breast tumors display hierarchical heterogeneity due to the presence of a high degree of cell plasticity elicited by activated oncogenes, which allows the cells to differentiate into diverse cell types. Also, the differentiation of tumor-initiating cells or stem cells, such as the CD44high/+/CD24low subtypes, in breast tumors underlies tumor heterogeneity and the emergence of aggressive and resistant neoplastic niches[22].
A BC subclone carrying mutations in AKT, FGFR, PIK3CA, and TP53 and increased chemoresistance, invasiveness, and metastatic ability has been characterized using whole genome and targeted genome sequencing. Single-cell technology has identified a unique claudin-low subclone displaying an epithelial stem cell phenotype and high resistance to chemotherapy[23]. Additionally, a change in the copy number can cause intratumor heterogeneity in BC. For example, HER2 copy numbers can vary in different BC patients. The accumulation of copy number variations due to complex genomic rearrangements or bursts of chromosome changes can induce the development of metastable, pseudodiploid or chromosomic subpopulations in TNBC[24]. Copy number changes are also responsible for the emergence of immune suppressive phenotypes in some T-and B-lymphocytes as well as M2 macrophages[25].
Molecular heterogeneity in ER has been described in estrogen and androgen hormone responsive BCs, which differ in distant recurrence risk and disease-free survival. Mutational analysis uncovered ER+/PR- BC subpopulations enriched with mutations in TP53, GATA3, CDH1, HER2, and BRAF[26]. Whole exome sequencing studies also reported heterogeneity in ER+/HER2- BCs, which differ in mutational alleles in TP53, CDH1, MYC and have poor overall survival[27]. As these studies show, cellular and molecular heterogeneity pose great challenges in treating metastatic BCs and overcoming drug resistance.
EPIDEMIOLOGY OF BC
Epidemiological investigations have established unmodifiable risk factors such as race, family history of BC, genetic makeup and ethnicity, as well as variable factors such as consumption of alcohol, high-fat diet, female reproductive factors, hormone therapy, obesity as BC causative factors. Additionally, menarche at a younger age, pregnancy at a late age, parity, and nulliparity can also influence BC risk via by affecting sex hormones. Genetic variants and gene mutations in proteins tangled in DNA repair have been implicated as additional risk promoters in BC[28]. Molecular and genetic studies have identified high-risk BC patients with mutations in BRCA1 and BRCA2, along with moderate change of BC-susceptibility genes including PTEN and TP53[29]. Furthermore, GWAS have established the combination of genetic weakness loci, reproductive risk factors, the interaction of gene-environment with obesity and lifestyle as a potential risk factor. Additionally, gene-environment interaction studies have identified BC subtypes in high-risk women[30]. Physical activity can enhance anti-tumor immunity in BC patients by increasing cytotoxic monocytes, natural killer (NK) cells, and cytokines[31]. Recent technological advances have thus allowed an improved characterization of the various BC subtypes and their molecular and genetic underpinnings, which can enable enhanced detection strategies.
PATHOPHYSIOLOGY OF BC
The current understanding of the pathophysiology of the breast TME is that the aggressive and metastatic phenotypes arise through a series of molecular alterations at the cellular and molecular level. Advanced studies have also elucidated the role of sustained angiogenesis in the pathophysiology of TME and the involvement of CSCs and redox biology in drug resistance in BC. The following sections explore the role of the TME in the development and progression of breast tumors.
TME components in BC metastasis
Metastasis is the last step of BC progression and is a common cause of BC mortality. Recent studies show that the components of the TME, including immune suppressive cells, soluble factors, and remodeled ECM, can reciprocally communicate with each other to promote BC metastasis (Figure 1). The breast TME is characterized by unusual signaling pathways and characteristic molecular changes. Tumor associated stomal cell interactions with BC cells promote metastasis through the induction of cytokine (CXCL8) production, which activates the expression of notch1 via tumor necrosis factor (TNF) α-induced p65 activation[32]. Cancer-associated fibroblasts (CAFs) are the most prominent stromal cells of the breast TME and accelerate metastasis by associating with other TME cells. Zhou et al[33] reported that CAFs promote lymph node metastasis of TNBC cells by interacting with tumor-associated macrophages (TAMs). This study also demonstrated that activated CAFs can increase lymphatic metastasis by increasing the intrusion of polarized macrophages in TNBC patients. CAFs promoted lung metastasis of TNBC cells in the homograft tumor model by activating transforming growth factor (TGF) β[34], thus indicating their key role in BC progression.
Figure 1.
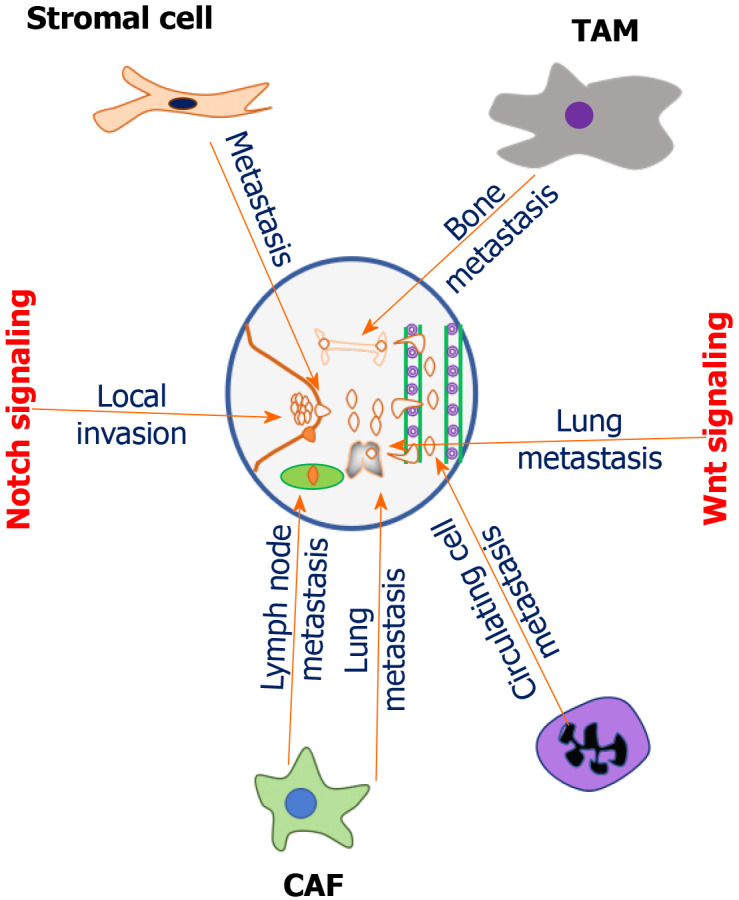
Role of tumor microenvironment components in promoting breast cancer metastasis. Breast tumor microenvironment is typified with different types of components with typical cellular and molecular functions. Tumor associated stromal cells promote metastasis through the induction of cytokine (CXCL8) production by activating the expression of notch1 via TNFα-induced p65 activation. Cancer-associated fibroblasts (CAFs) accelerate lymph node metastasis of triple-negative breast cancer (TNBC) cells by increasing the intrusion of polarized macrophages in TNBC patients. Tumor associated macrophages promote bone metastasis by secreting interleukin 12. CAFs promoted lung metastasis of TNBC cells in the homograft tumor model by activating transforming growth factor β. Cancer associated neutrophils mediates metastasis by leukotrienes. Neutrophils enhance metastatic ability of circulating breast cancer (BC) cells. Notch signaling promotes local invasion of BC cells. Wnt signaling modulates metastasis of BC cells to the lung. TAM: Tumor-associated macrophage; CAF: Cancer-associated fibroblast.
Neutrophils can also accelerate BC metastasis. Coffelt et al[35] reported that neutrophils are expanded and polarized by interleukin (IL) 17 from γδT-cells of breast tumor in mice. The polarized neutrophils promote metastasis by suppressing CD8+ cytotoxic T cells. IL-17 neutralization using specific antibodies reduced the lung metastasis of BC cells by decreasing the T-cell suppressive neutrophils[35]. Another study reported that neutrophils can also mediate BC metastasis through leukotrienes. This study observed that blocking leukotriene synthesizing enzyme 5-lipooxygenase can reduce metastasis by abrogating the prometastatic activity of neutrophils[36]. Additionally, neutrophils can increase the metastatic ability of BC cells by eliciting cell cycle progression via modulating the expression of cytokine receptors and genes associated with cell-cell junctions[37]. Notch signaling also promotes local invasion of BC cells. Furthermore, the Wnt signaling component LGR4 (the fourth member of the leucine-rich repeat-containing GPCR family) is over expressed in BC stem cells (BCSC) and can modulate the metastasis of BC cells to the lung by promoting epithelial to mesenchymal transition (EMT) via increasing the expression of N-cadherin and transcription factor snail and decreasing the expression of E-cadherin[38]. Therapeutic strategies that target and reprogram the TME are thus required to improve treatment outcomes in BC patients.
TME components in BC angiogenesis
In the TME, angiogenesis is modulated by innate immune cells and other communicators (Figure 2), including neutrophils, macrophages, dendritic cells, myeloid-derived cells, γδTcells, mast cells, and NK cells[39]. Lin et al[40] reported that TAMs help angiogenesis after infiltrating into the breast TME. This study correlated the expression of CCL18, CD34, and microvascular density by double immuno-histochemical staining of breast tumor tissues. It also demonstrated the induction of angiogenesis by chemokine, CCL18 in the coculture of TAMs, and endothelial cells. Additionally, the study reported that blocking of CCL18 putative receptor (PITPNM3) inhibited CCL18-promoted migration of endothelial cells as well as tube formation. In contrast, the treatment of endothelial cells with CCL18 stimulated the transition of endothelial cells to mesenchymal cells by activating the Akt/GSK-3β/Snail and ERK (extracellular regulated protein kinases) signaling[40]. In the TME, TAMs induced angiogenesis by remodeling ECM for promoting invasion and BC cell modeling into stem cells for evading host immunity as well as recruiting immune-suppressive immune cells[41]. This study indicates that TAMs can induce angiogenesis and promote the progression of BC.
Figure 2.
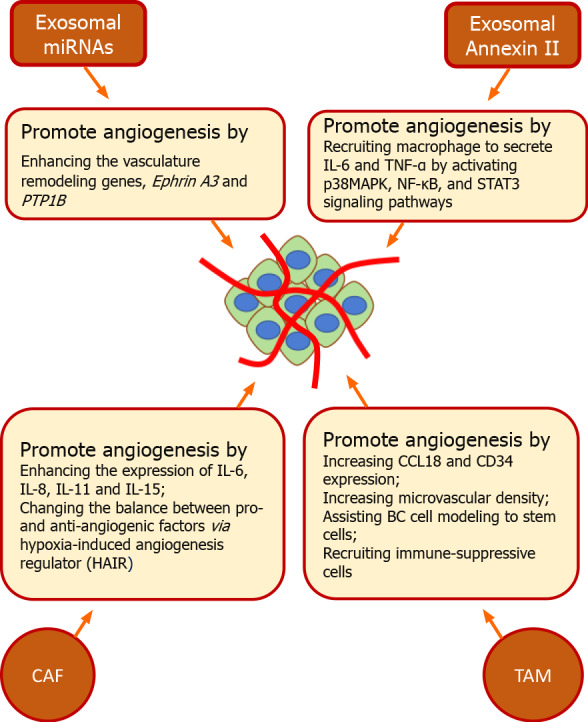
Modulation of angiogenesis by innate immune cells and other communicators in breast tumor microenvironment. In breast tumor microenvironment, innate immune cells and other communicators modulate angiogenesis. Tumor-associated macrophages promote angiogenesis by increasing the expression of CCL18, CD34, and microvascular density and assisting breast cancer cell modeling into stem cells for recruitment of immune-suppressive cells. Cancer-associated fibroblasts promote tumor angiogenesis by enhancing the expression of interleukin (IL) 6, IL-8, IL-11 and IL-15 as well as by changing the balance between pro-and anti-angiogenic factors via hypoxia-induced angiogenesis regulator. Tumor-derived exosomal Annexin II induce angiogenesis by recruiting macrophages to secrete IL-6 and tumor necrosis factor-α by activating p38MAPK, nuclear factor-κappa beta, and STAT3 signaling pathways. Exosomal miRNAs contribute to the development of tumor angiogenesis by enhancing the vasculature remodeling genes, Ephrin A3 and PTP1B. TAM: Tumor-associated macrophage; CAF: Cancer-associated fibroblast; HAIR: Hypoxia-induced angiogenesis regulator; NF-κβ: Nuclear factor-κappa beta; TNF: Tumor necrosis factor; IL: Interleukin; BC: Breast cancer.
Similar to TAMs, CAFs promote tumor angiogenesis by various mechanisms. Du et al[42] demonstrated that miRNA-205/YAP1 signaling induces tumor formation as well as the sprouting of endothelial cells by promoting the transformation of normal fibroblasts into CAFs via enhancing the expression of IL11 and IL15. However, blocking of miRNA-205 in CAFs inhibited angiogenesis. Additionally, the study reported that CAFs release IL11 and 15 by stimulating STAT3 signaling in endothelial cells. However, blocking of IL11 and 15 halted CAF-induced angiogenesis as well as BC metastasis by inactivating STAT3 signaling[42], thus highlighting its therapeutic potential in BC. Furthermore, Eiro et al[43] reported that CAFs can enhance the levels of invasion and angiogenesis-associated genes in BC. The study demonstrated that CAFs derived from MMP11 positive stromal mononuclear inflammatory cells increased the transcript levels of IL-6 and -8 in BC cells in the co-culture model[43]. However, in co-culture of hypoxic CAFs and endothelial cells, CAFs induced abnormal blood vessels formation by changing the balance between anti- and pro-angiogenic factors. This study also revealed that hypoxia altered the expression of hypoxia-induced angiogenesis regulator (HAIR) in CAFs. Furthermore, knockdown of HAIR inhibited CAF-induced migration of endothelial cells by reducing the release of VEGF as well as inhibiting VEGF/VEGFR signaling[44]. These various studies show that CAFs can strongly influence BC development, suggesting that therapeutic strategies that target CAFs in the TME could improve therapeutic responses in BC patients.
Tumor-derived exosomes have emerged as key mediators of tumor-induced angiogenesis. Annexin II (Anx II) is one of the most highly expressed exosomal proteins associated with BC pathogenesis. The exosomal Anx II (Exo-AnxII) reshapes the environment for supporting metastasis, including promoting tPA-mediated angiogenesis. This study reported that Exo-Anx II induces macrophages to secrete IL6 and TNF-α by activating p38 MAPK (mitogen-activated protein kinase), nuclear factor-κappa beta (NF-κB), and STAT3 signaling pathways[45]. Also, exosomal miRNAs from hypoxic tumor cells contribute to the development of tumor angiogenesis. They promote angiogenesis by enhancing the vasculature remodeling genes, including Ephrin A3 and PTP1B, via transferring miRNA-210 to adjacent tumor cells in TME[46]. Exosomes orchestrate angiogenesis for BC metastasis after activation by Notch signaling. The interaction of aspartate β-hydroxylase (ASPH) with Notch signaling components promotes the synthesis and release of exosomes in murine BC models[47]. Exosomes from CXCR4 overexpressing BC cells enhance the expression of stemness-related genes in recipient cells and increase their proliferative, migratory, and invasive ability[48]. In contrast, exosomes from mesenchymal stem cells can reduce angiogenesis in BC by decreasing the VEGF expression via targeting miRNA-16[49]. These studies support the role of exosomes in the modulation of angiogenesis in BC and highlight their importance as novel potentials diagnostic and therapeutic tools in BC.
TME components in drug resistance of BC
In BC, drug resistance and recurrence are mediated by BCSC as well as redox molecules (Figure 3). A fine orchestration between BCSC and TME cells is involved in the process of drug resistance. This interaction promotes drug resistance by upregulating BC resistance protein (BCRP), which mediates the drug efflux mechanism in BC cells. Stem cells from bone marrow, adipose tissue, and fibroblasts also enhance the metastatic potential of BC cells by activating developmental pathways via secreting chemokines and cytokines. Wnt signaling of BCSC along with glutathione overexpressing genes mediates resistance[50]. These mechanisms can reshape the TME leading to increased drug evasion and survival, thus challenging the success of current therapeutic regimens.
Figure 3.
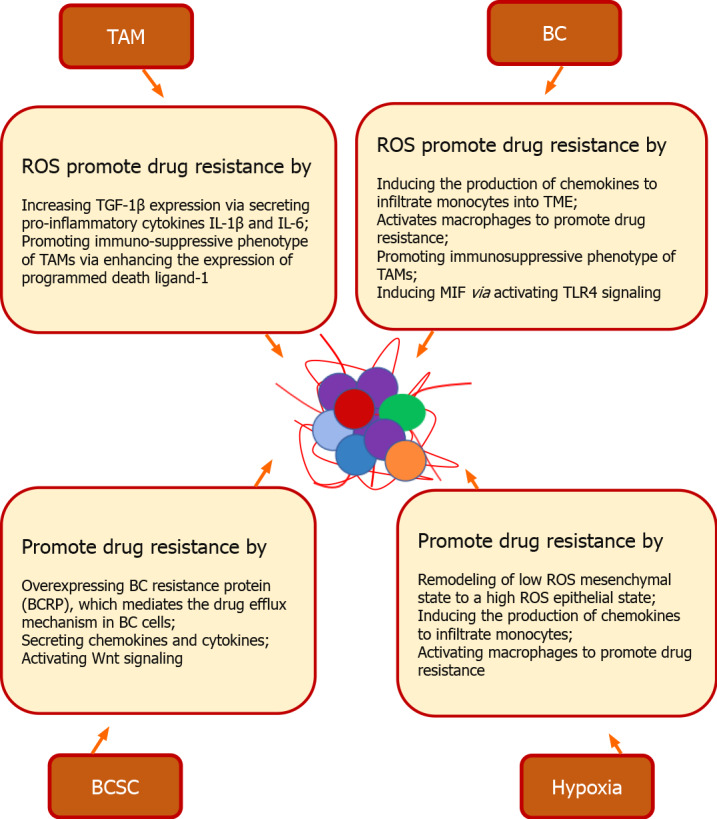
Cancer stem cell and redox mediated drug resistance in breast cancer. In breast tumor microenvironment (TME), a fine orchestration between breast cancer (BC) stem cells (BCSC) and TME cells promotes drug resistance. BCSC promote drug resistance by overexpressing BC resistance protein, which mediates the drug efflux mechanism in BC cells. Stem cells derived from bone marrow, adipose tissue, and fibroblast enhances the drug resistance by activating developmental pathways via secreting chemokines and cytokines. Wnt signaling of BCSC along with glutathione overexpressing genes mediates resistance. Hypoxic TME promotes drug resistance by remodeling of cells from a low reactive oxygen species (ROS) mesenchymal state to a high ROS epithelial state. High ROS from cancer cells induces the production of chemokines to infiltrate monocytes into TME and activates macrophages to promote drug resistance. ROS also induces drug resistance by promoting the immunosuppressive phenotype of tumor-associated macrophages via enhancing the expression of programmed death ligand-1 via NF-κB (nuclear factor-κappa beta) signaling. Multinucleated cells promote drug resistance by increasing the secretion of vascular endothelial growth factor and macrophage migration inhibitory factor (MIF) via RAS/MAPK pathway-dependent hypoxia-inducible factor-1α. Macrophage derived ROS induces drug resistance by increasing transforming growth factor 1β expression via secreting pro-inflammatory cytokines interleukin (IL)-1β and IL-6. ROS also facilitates drug resistance by inducing MIF via promoting phosphorylation of ERK. via activating TLR4 (toll-like receptor 4) signaling. TAM: Tumor-associated macrophage; BC: Breast cancer; ROS: Reactive oxygen species; TGF: Transforming growth factor; IL: Interleukin; TME: Tumor microenvironment; MIF: Macrophage migration inhibitory factor; TLR: Toll-like receptor; BCSC: Breast cancer stem cell.
Additionally, emerging scenarios have recognized that redox signals promote TME mediated drug resistance in BC. Redox mechanisms orchestrate pathophysiology of BC to promote abnormal proliferation, metastasis, and drug resistance through reactive oxygen species (ROS) signals. The hypoxic condition in the TME promotes drug resistance by inducing the remodeling of cells from a low ROS mesenchymal state to a high ROS epithelial state. Cancer cells with high ROS induce drug resistance by producing chemokines that elicit the infiltration of monocytes into the TME and activate macrophages to promote angiogenesis[51]. ROS also induces drug resistance by promoting the immunosuppressive phenotype of TAMs, which release immuno-suppressive chemokines by increasing the levels of programmed death ligand-1 (PD-L1) via NF-κB signaling[52]. In the breast TME, multinucleated cells promote drug resistance by increasing VEGF secretion and macrophage migration inhibitory factor (MIF) via RAS/MAPK pathway-dependent hypoxia-inducible factor (HIF)-1α[53]. Macrophages induce TGF-1β expression in cancer cells by secreting IL-1β and IL-6. These cytokines promote EMT and migration of tumor cells by activating pCREB signaling via ROS. ROS facilitates drug resistance in metastatic cells by inducing MIF via promoting phosphorylation of ERK, which promotes the release of high-mobility group box 1 (HMGB1) into the cytosol from the nucleus by inducing caveolin-1 phosphorylation. The released HMGB1 then phosphorylates NF-κB via activating TLR4 (toll-like receptor 4) signaling. The activated NF-κB activates MMP2 and induces transcription of Snail and Twist. This MIF-mediated HMGB1 signaling regulates the CD11b+ immune cell recruitment in the breast TME and promotes drug resistance[54]. As these various studies show, the TME plays a key role in BC development and metastasis. Hence, novel therapeutic regimens that target the various components of the TME to overcome resistance mechanisms could unleash the native immune system against BCs and improve therapeutic outcomes.
LONG NON-CODING RNAS IN THE PATHOPHYSIOLOGY OF BC
Recently, long non-coding RNAs (lncRNAs) have been reported to mediate pathophysiological developments such as metastasis as well as immune suppression within the TME (Figure 4). Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is a lncRNAs enriched in the nucleus and helps cancer growth and progression by regulating gene expression and post-translational modifications. In TNBC cells, MALAT1 drives tumor progression and metastasis. The expression levels are correlated with reduced disease-free survival in both HER2 and TNBC patients, who are negative for ER and lymph node[55]. MALAT1interacts with ER, and its increased expression levels correlate with ER and PR positive status in BCs with prognostic significance (cut-off value 75%) associated with relapse. It also promotes invasion and migration of TNBCs by targeting tumor suppressor miRNA-129-5p, but its silencing induced cell cycle arrest at G0/G1phage[56]. Hence, MALAT1 could be a potential diagnostic tool for BC (Figure 4)[57].
Figure 4.

Role of long non-coding RNAs in the pathophysiology of breast cancer. Long non-coding RNAs are reported to mediate pathophysiological developments such as metastasis as well as immune suppression within the tumor microenvironment. Metastasis-associated lung adenocarcinoma transcript 1 promotes growth and metastasis by controlling gene expression and post-translational modifications, invasion and migration by targeting tumor suppressor miRNA-129-5p, metastasis by transcriptionally regulating disruptor of telomeric silencing 1-like and promoting the interaction of hypoxia-inducible factor-2α. HOTAIR (HOX antisense intergenic RNA) increases invasiveness and metastasis by enhancing the expression of polycomb repressive complex 2 and altering the epigenome by histone H3 lysine 27 methylation, enhancing cell migration and invasiveness and inhibiting apoptosis through targeting high mobility AT-hook 2 via miRNA-20a-5p. MEG3 reduced angiogenesis by decreasing the expression of proangiogenic molecules as well as blocking Akt signaling. NKILA [nuclear factor-κappa beta (NF-κB) interacting lncRNA] negatively regulates tumor angiogenesis by decreasing the interleukin-dependent expression of vascular endothelial growth factor (VEGF)-A and VEGF-R through inhibiting NF-κB signaling and promotes antitumor immunity by inducing immune evasion of tumor cells via sensitizing T-cells through activation-induced cell death mechanism[57]. LINC00968 inhibits capillary formation by downregulating miRNA-423-5p mediated PROX1. LINC01133 promotes stem cell phenotypes by triggering miRNA-199a dependent FOXP2 signaling via modulation of Kruppel-like factor 4. Silencing of lncRNA-21 in tumor-associated macrophages induced apoptosis and reduced cell migration and invasion. LINK-A promotes antitumor immunity by inducing loss of antigenicity through PIP3 and inhibitory G-protein coupled receptor pathway as well as attenuating protein kinase A-dependent phosphorylation of E3 ubiquitin ligase TRIM71 and degradation of antigen peptide loading complex in triple-negative breast cancer. MALAT1: Metastasis-associated lung adenocarcinoma transcript 1; DOT1L: Disruptor of telomeric silencing 1-like; HIF: Hypoxia-inducible factor; HOTAIR: HOX antisense intergenic RNA; PRC2: Polycomb repressive complex 2; HMGA2: High mobility AT-hook 2; TNBC: Triple-negative breast cancer; IL: Interleukin; VEGF: Vascular endothelial growth factor.
Silencing of MALAT1 reduced cancer growth and progression by increasing cell adhesion and decreasing migration of cancer cells in mouse mammary carcinoma model via altering the expression pattern of genes implicated in differentiation and protumorigenic signaling pathways[58]. The expression patterns of MALAT1 and transcription factor BACH1 were substantially associated with TNM stage, metastatic ability, and pathological stage as well as the survival of patients[59]. MALAT1 can possibly promote metastasis in association with ZEB2 by transcriptionally regulating disruptor of telomeric silencing 1-like in BC[60]. Furthermore, hypoxia induces MALAT1, which also enhances the metastasis of BCs by promoting the interaction of HIF-2α with the MALAT1 gene between the promoter region and putative enhancer-3 as well as chromatin interactions with putative enhancer-7[61]. In contrast, Kim et al[62] reported that overexpression of MALAT1 can reduce metastasis of BC cells in the xenograft model by hindering the interaction of TEAD with YAP via transcriptional inactivation. However, the silencing of MALAT1 enhances the metastatic ability of BC cells.
Another important lncRNA that promotes metastasis of BCs by reprogramming chromatin state is HOX antisense intergenic RNA (HOTAIR). It is highly expressed in both primary as well as metastatic breast tumors. Its expression is strongly correlated with lymph node metastasis and the expression of the AR. Mechanistically, HOTAIR upregulation using overexpressing plasmid enhanced the expression of polycomb repressive complex 2 (PRC2) and altered the epigenome by histone H3 lysine 27 methylation. It also enhanced invasiveness and metastasis in BCs, while its silencing inhibited invasiveness in PRC2 overexpressing BCs[63]. HOTAIR can also promote progression by increasing migration and invasiveness and inhibiting apoptosis through targeting high mobility AT-hook 2 (HMGA2) via miRNA-20a-5p in BC cells[64].
The overexpression of MEG3 reduced angiogenesis in BC by reducing the capillary formation of endothelial cells via decreasing the expression of proangiogenic molecules as well as blocking Akt signaling[65]. However, overexpression of lnc RNA NKILA (NF-κB interacting lncRNA) significantly reduced tumor angiogenesis by reducing the IL-dependent VEGF-A and VEGF-R expressions through inhibiting NF-κB signaling and IkBα phosphorylation and nuclear translocation[66]. Recently, Sun et al[67] reported that overexpression of lncRNA (LINC00968) reduced capillary formation by downregulating miRNA-423-5p mediated PROX1 in BC cells. Another lnRNA, LINC01133 promoted stem cell phenotypes in TNBC cells by triggering miRNA-199a dependent FOXP2 signaling via modulation of Kruppel-like factor 4[68]. TME induced stemness in BC cells by silencing lncRNA-HAL through chromatin level and transcriptional regulation[69].
Additionally, lncRNAs can regulate tumor immunity through various mechanisms. Huang et al[57] reported that NKILA promotes antitumor immunity by inducing immune evasion of tumor cells via sensitizing T-cells through activation-induced cell death mechanism. Silencing of lncRNA-21 in TAMs induced apoptosis in BC cells and reduced metastasis[70]. In contrast, lncRNA (LINK-A) promotes antitumor immunity by inducing loss of antigenicity through PIP3 and IGPCR pathway in TNBC[71]. Hence, lncRNAs can modulate metastasis at different molecular levels, including epigenetic modification and multiple signaling pathways. Future studies could lead to the development of clinical applications based on lncRNAs as potential diagnostic biomarkers or drug targets.
BC THERAPY
As the previous sections show, the complex pathophysiology poses a challenge to the development of effective therapeutic regimens. Novel personalized strategies that target the BC TME are required to improve clinical responses. Natural compounds (e.g., cytarabine, eribulin mesylate) from marine organisms have been approved by the FDA for cancer treatment and could represent valuable clinical modalities for BC. Several marine compounds with antineoplastic activity are currently undergoing different phases of evaluation in clinical trials (e.g., plinabulin). Additionally, the anticancer activity of more than 1500 marine compounds has been evaluated using in vivo models while the activity of 10000 compounds has been tested using in vitro models[72]. Marine compounds malformin A (cyclic pentapeptide), kuanoniamine D (pyridoacridine alkaloid), hymenialdisine (C11–N5 alkaloid), and gallic acid (GA, phenolic acid) exert potentially high anticancer activity against MCF-7 BC cells by inducing apoptosis and promoting nuclear fragmentation, membrane protrusion, blebbing and chromatin segregation[73]. The following paragraphs will examine some of the best-studied marine compounds and explore their relevance to BC treatment.
Marine compounds with potential anti-BC activity from microorganisms and algae
Marine compounds from microorganisms have potential anticancer activity (Table 1). For example, GA can stimulate apoptosis by changing p53, Mcl, and p21 expression. GA can also stimulate cell cycle arrest in BC cells by altering the cyclin-dependent kinases (CDKs) expression. Mechanistically, MAP38 kinase is intricated in GA-stimulated cell cycle arrest and apoptosis via downregulating cyclin Da/CDK4 and cyclin E/CDK2. However, a combination of GA with curcumin stimulated apoptosis in BC cells by overexpressing the expression of Bax as well as activating caspase 3 and poly (ADP ribose) polymerase (PARP) and decreasing Bcl2 along with arresting at sub-G1 phase[74]. In addition, a nanocomposite of iron oxide magnetite nanoparticles (NPs) and polyethylene glycol (PEG) enhanced the anticancer activity of GA. Moreover, GA conjugated Gold NPs suppressed the metastasis of MDA-MB cells by blocking EGF dependent MMP-9 expression via suppressing stabilization of p300 as well as activating NF-κB/c-Jun pathway. However, overexpression of MEK1 and Akt reduced the inhibitory activity of GA on the EGF-dependent upregulation of MMP-9[75]. Crambescidin 800 is a heteropenta cyclic guanidine alkaloid. Crambescidin 800 stimulated arrest at the G2M phase by decreasing the cyclin D1, CDK-4, and -6 expression in TNBC cells via modulating the Akt/NF-κB/MAPK pathway[76] and thus warrants further clinical investigation.
Table 1.
Marine compounds with potential anticancer activity against breast cancer from microorganisms and algae
|
Marine compound
|
Chemical nature
|
Mechanism of action
|
Ref.
|
| Gallic acid (GA) | Phenolic compound | (1) Altered the expression of P53, Mcl and p21 as well as cell cycle regulators; and (2) MAP38 kinase involved in GA induced cell cycle arrest and apoptosis via downregulating cyclin Da/CDK4 and cyclin E/CDK2 | Moghtaderi et al[74] |
| GA | Phenolic compound | (1) In combination with curcumin stimulated apoptosis by increasing the Bax expression, activating PARP and caspase 3; (2) Decreased Bcl2 expression; and (3) Arrested at sub-G1 stage | |
| GA | Phenolic compound | Conjugated to Gold NPs, suppressed metastasis by blocking EGF dependent MMP-9 expression via suppressing stabilization of p300 and activation of NF-κB/c-Jun pathway | Chen et al[75] |
| Crambescidin 800 | Heteropenta cyclic guanidine alkaloid | Induced cell cycle at the G2M phase by decreasing the cyclin D1, CDK-4, and -6 expression in TNBC cells via modulating Akt/NF-κB/MAPK pathway | Moon et al[102] |
| EPS11 | Polysaccharide | (1) Inhibited lung metastasis by inhibiting cell adhesion protein CD99; and (2) Inhibited cancer cell growth by inducing anoikis via inducing Akt pathway-dependent expression of βIII-tubulin | Cao et al[77] |
| SWP1 and SWP2 | Polysaccharide | Inhibited proliferation by inducing apoptosis, activating caspase 3/9 and disrupting the mitochondrial membrane via generation of ROS | Vaikundamoorthy et al[78] |
| Carrageenan | Polysaccharide | Induced apoptosis via promoting condensation of the nucleus and fragmentation of DNA as well as activating caspase 8, an extrinsic apoptotic protein | Murad et al[79] |
| Exopolysaccharide | Polysaccharide | (1) Inhibited the cell growth by decreasing the cyclin D1 and E expression; and (2) Induced the proliferation of B-cells and decreased production of IL-6 and TNF-1α in T-cells | Park et al[80] |
| Ilmycin C | Cyclic peptide | Inhibited migration and invasion by inducing apoptosis via Bax/Bcl-2 dependent caspases as well as inhibiting MMP-2 and -9 via blocking IL-6 dependent phosphorylation of STAT3 | Xie et al[81] |
| Inhibited growth by inducing apoptosis through activation of SR stress and reducing Bcl2 in a CHOP dependent manner | Zhou et al[82] | ||
| Molassamide | Cyclic depsipeptide | (1) Abrogated elastase-dependent migration of highly metastatic TNBC cells; and (2) Inhibited the activity of elastase and the migration of TNBC cells by targeting the expression of ICAM-1 via inhibiting the NF-κB pathway | Al-Awadhi et al[83] |
| Kempopeptin C | Cyclic depsipeptide | Inhibited invasion and migration by decreasing the cleavage of matriptase substrates CDCP1 and sesmoglein-2 | Al-Awadhi et al[84] |
| Cyclic leucylproline | Cyclic peptide | Inhibited migration by inhibiting cell proliferation, inducing cell arrest via DNA damage. Mechanistically, CLP induced cell cycle arrest by blocking the expression of cyclin C, CDK4, PAK, RAC1, and p27kiP1 via targeting CD151 and EGFR signaling axis in TNBC cells | Kgk et al[85] |
| Galaxamide | Cyclic pentapeptide | Elicited apoptosis in BC cells by arresting at the G1 phase as well as reducing mitochondrial membrane potential via the generation of ROS | Lunagariya et al[86] |
| Brintonamide D | Linear peptide | Reduced the CCL27 and stimulated proliferation and progression of metastatic BC cells by targeting serine protease kallikrein 7 (KLK7). This study reported that brintonamide D targeted KLK7 by modulating CCR10, the receptor of CCL27 in BC cells | Al-Awadhi et al[87] |
| Iturin A | Lipopeptide | (1) Induced apoptosis by increasing sub-G1 cell population, fragmentation of DNA via inhibiting FGF-mediated phosphorylation of Akt, FoxO3a and GSK3β; (2) And reduced tumor growth by promoting translocation of FoxO3a via downregulating MAPK and Akt kinase in the xenograft model | Dey et al[88] |
| Halilectin-3 | Sugar-binding lectin protein | Inhibited proliferation by inducing arrest at the G1 phase and apoptosis by increasing the activity of caspase 9 and autophagy by inducing the expression of light chain 3 | do Nascimento-Neto et al[89] |
| Sinularin | Terpenoid | Reduced cell viability by halting at the G2M phase and stimulating apoptosis through activation of caspase-3 and -8 as well as PARP. In addition, it also induced DNA damage by generating ROS via stimulating oxidative stress | Huang et al[90] |
| Sipholenol A | Triterpene | Reduced the metastatic ability of TNBC cells by inhibiting protein tyrosine kinase 6, a key mediator of growth factor-dependent migration | Foudah et al[91] |
| Agelasine B | Diterpene alkaloid | (1) Induced apoptosis by inhibiting ER Ca2+ -ATPase (SERCA) activity via releasing Ca2+ from ER and inducing DNA fragmentation; (2) Reduced the Bcl2 expression and enhanced the caspase 8 expression; and (3) Induced cell death in an ER-mediated extrinsic apoptotic pathway | Pimentel et al[92] |
| Hirsutanol A | Sesquiterpene | (1) Reduced cell growth by inhibiting proliferation; (2) Induced apoptosis, and autophagy via generating ROS; and (3) Silenced Atg7 with siRNA and blockade of autophagy using bafilomycin A1 synergistically increased the efficacy of hirsutanol A in inducing apoptosis and inhibiting cell proliferation | Yang et al[93] |
| Dehydrothyrsiferol | Triterpenoid | Induced apoptosis by causing DNA fragmentation and arrest at S-phase and G2M phase | Pec et al[94] |
| Sodwanone | Triterpene | (1) Induced cytotoxicity to BC cells; and (2) Inhibited hypoxia-induced HIF-1α | Dai et al[95] |
| Pseudopterosin | Diterpene glycoside | (1) Reduced the production of IL-6, TNF-1α, and MCP-1 via blocking p65 and IkB phosphorylation; and (2) Promoted translocation of glucocorticoid receptor from nucleus to cytosol | Sperlich et al[96] |
| Quinazoline | Heterocyclic compound | (1) Induced apoptosis in HER+ve BC cells by reducing the Bcl2 expression and increasing the Bax expression; and (2) Promoted cell death via ROS-dependent extrinsic or intrinsic apoptotic pathways without systemic toxicity in the mouse model | De et al[97] |
| (3β)-Cholest-5-en-3-ol | Cholesterol | Induced cell death by activating caspase 3 and 8 as well as increasing the Bax expression and decreasing the Bcl2 expression | Sharifi et al[98] |
| 3β,11-dihydroxy-9,11-secogorgost-5-en-9-one | Sterol | (1) Inhibited cell growth by inducing apoptosis via activation of caspase 3 and PARP and cell cycle arrest via targeting cyclin D1 and CDK6 through blocking the p38/ERK signaling pathway; and (2) Induced autophagy via generating ROS and DNA damage by increasing the expression of H2AX | Weng et al[99] |
| 4-methyenedioxy-β-nitrostyrene | β-nitrostyrene derivatives | Inhibited migration by disrupting the focal adhesion complex as well as a network of actin stress fibers via reducing β1 integrin-dependent phosphorylation of FAK and paxillin | Chen et al[100] |
CDK: Cyclin-dependent kinase; PAPR: Poly (ADP ribose) polymerase; NPs: Nanoparticles; EGF: Endothelial growth factor; MMP: Matrix metalloproteinase; NF-κβ: nuclear factor-κappa beta; TNBC: Triple-negative breast cancer; MAPK: Mitogen-activated protein kinase; ROS: Reactive oxygen species; IL: Interleukin; TNF: Tumor necrosis factor; ICAM-1: Intercellular adhesion molecule-1; CDCP1: CUB-domain containing protein 1; CLP: Cyclic dipeptide of leucine and proline; EGFR: Endothelial growth factor receptor; HIF: Hypoxia-inducible factor; MCP-1: Monocyte chemotactic protein-1; BC: Breast cancer; HER: Human epidermal growth factor receptor; FAK: Focal adhesion kinase.
The polysaccharide EPS11, isolated from marine bacteria, inhibits lung metastasis by inhibiting the cell adhesion protein CD99. It also inhibits cancer cell growth by inducing anoikis via inducing the Akt pathway-dependent expression of βIII-tubulin[77]. The polysaccharide fractions, SWP1, and SWP2 from brown seaweed decreased the proliferation of BC cells by stimulating apoptosis and activating caspase 3/9 and disrupting the mitochondrial membrane via generation of ROS[78]. Carrageenan, a polysaccharide from red algae, inhibited BC cells at 50 μg by inducing apoptosis via promoting condensation of the nucleus and fragmentation of DNA as well as activating caspase 8, an extrinsic apoptotic protein[79]. Exopolysaccharides from marine algae inhibited the growth of BC cells by reducing the cyclin D1 and E transcript levels, while inducing the proliferation of B-cells and decreasing the IL-6 and TNF-1α production in T-cells[80]. Hence this compound displays promising immunomodulatory effects that warrant further investigation.
The marine compound, Ilmycin C, is a cyclic peptide, that inhibits the invasion and migration of BC cells by inducing apoptosis via Bax/Bcl-2 dependent caspases as well as inhibiting MMP-2 and -9 via blocking IL-6 dependent phosphorylation of STAT3[81]. Ilamycin E can also inhibit TNBC growth by inducing apoptosis through activating SR stress and reducing Bcl2 in a CHOP dependent manner[82]. Cyclic depsipeptide, molassamide can abrogate elastase-dependent migration of highly metastatic TNBC cells. It can also inhibit the activity of elastase and the migration of TNBC cells by targeting the expression of ICAM-1 (intercellular adhesion molecule-1) via decreasing NF-κB[83]. Another cyclic depsipeptide, kempopeptin C from marine cyanobacteria can inhibit the invasion and migration of TNBC cells by reducing the cleavage of matriptase substrates CDCP1 (CUB-domain containing protein 1) and sesmoglein-2[84]. However, cyclic dipeptide of leucine and proline (CLP) decreased TNBC cells migration by inhibiting proliferation and inducing G2/M arrest via DNA damage. Mechanistically, CLP induced cell cycle arrest by blocking cyclin C, CDK4, PAK, RAC1, and p27kiP1 expression via targeting CD151 and EGFR signaling axis in TNBC cells[85]. The marine cyclic pentapeptide galaxamide and its analogs elicited apoptosis in BC cells by arresting the G0/G1 phase as well as reducing mitochondrial membrane potential via the generation of ROS[86]. These various studies illustrate the wide ranging molecular and immunomodulatory effects of marine compounds, highlighting their versatility as potential sources of TME modulating molecules with therapeutic benefits.
Brintonamide D is a linear peptide that can reduce CCL27-stimulated proliferation and migration of metastatic BC cells by targeting serine protease kallikrein 7 (KLK7). This study reported that brintonamide D targeted KLK7 by modulating CCR10, the receptor of CCL27 in BC cells[87]. Iturin A, a marine bacterium derived lipopeptide, stimulated apoptosis in BC cells by increasing sub-G1 cell population, fragmentation of DNA via inhibiting FGF-mediated phosphorylation of Akt, FoxO3a and GSK3β. This study also reported that Iturin A reduces tumor growth by promoting translocation of FoxO3a via inactivating MAPK and Akt kinase in the xenograft model[88]. Furthermore, Halilectin-3, a sugar-binding lectin protein from marine sponges, inhibited BC cell proliferation by stimulating the G0/G1 arrest. It also promoted apoptosis by increasing the activity of caspase 9 and autophagy by inducing the expression of the light chain 3[89]. These various marine compounds display promising anti-tumor activity that could be harnessed to target the BC TME in a non-invasive and non-toxic fashion.
The marine terpenoid sinularin can decrease the viability of TNBC cells by halting the the G2/M phase and inducing apoptosis through activation of caspase-3 and -8 and PARP. In addition, it can also induce DNA damage by generating ROS via stimulating oxidative stress[90]. Sipholenol A, a triterpene from marine sponges, reduced the metastatic ability of TNBC cells by inhibiting protein tyrosine kinase 6, a key mediator of growth factor-dependent migration[91]. Agelasine B is a diterpene alkaloid from marine sponges that can stimulate apoptosis in BC cells by inhibiting ER Ca2+ -ATPase (SERCA) activity via releasing Ca2+ from ER and inducing DNA fragmentation. It also reduced Bcl2 expression and increased caspase 8 levels. This study demonstrated that Agelasine B can induce cell death in BC cells through an ER-mediated extrinsic apoptotic pathway[92]. Hirsutanol A, a sesquiterpene from marine fungus, also reduced the growth of BC cells by inhibiting proliferation and stimulating autophagy and apoptosis via generating ROS. Silencing of Atg7 with siRNA and blockade of autophagy using bafilomycin A1 synergistically increased the efficacy of hirsutanol A in inducing apoptosis and inhibiting cell proliferation. This study indicates that hirsutanol A can induces autophagy and programmed cell death (PCD) by enhancing ROS accumulation in BC cells[93]. Dehydrothyrsiferol (DT) is a triterpenoid that promotes apoptosis in BC cells by DNA fragmentation and arrest at S-phase and G2M phase[94]. Sodwanone, a marine triterpene, is highly toxic to BC cells and can inhibit hypoxia-induced HIF-1α[95]. Pseudopterosin, a marine diterpene glycoside, also decreased the production of IL-6, TNF-1α, and MCP-1 (monocyte chemotactic protein-1) in BC cells via blocking p65 and IkB phosphorylation as well as promoting translocation of glucocorticoid receptor from nucleus to cytosol[96].
Quinazoline, a six-membered nitrogen-containing heterocyclic compound from marine sponge Hyrtios erectus, stimulated apoptosis in HER-positive BC cells by decreasing Bcl2 expression and increasing Bax. This study reported that quinazoline promoted cell death via ROS-dependent extrinsic or intrinsic apoptotic pathways without systemic toxicity in the mouse model[97]. Sharifi et al[98] showed that marine compounds (3β)-Cholest-5-en-3-ol (cholesterol), 2-hexadecanol and hexadecanoic acid from sea pen stimulated cell death in TNBC cells by activating caspase 3 and 8 as well as increasing Bax expression and reducing Bcl 2 levels. Furthermore, a sterol from soft corals inhibited BC cell growth by stimulating apoptosis via activation of caspase 3 and PARP. It also elicited Go/G1 arrest by reducing cyclin D1 and CDK6 and blocking p38/ERK signaling. Furthermore, it induced autophagy via generating ROS and DNA damage through H2AX upregulation[99]. Another marine compound, 4-methyenedioxy-β-nitrostyrene (MNS) also inhibited the migration of TNBC cells by disrupting the focal adhesion complex as well as a network of actin stress fibers via reducing β1 integrin-dependent phosphorylation of FAK (focal adhesion kinase) and paxillin[100]. Yang et al[101] reported that leucine aminopeptidase-3 (LAP-3) can inhibit the growth, invasion and migration of TNBC cells by targeting LAP-3 dependent expression of fascin and MMP-2/9. As these studies illustrate, marine compounds display a wide range of antineoplastic effects that could be exploited to develop novel drug modalities for BC.
Marine compounds with potential anti-BC activity from crustaceans
The crustacean shell is a rich source of chitin. Crab, the major crustacean, is highly recommended for cancer treatment in traditional medicine due to chitin, carotenoids (astaxanthin, AST), lutein, and β-carotene (Table 2)[102]. The hydro-alcoholic extract of crab shells can reduce the proliferation of MCF-7 cells by stimulating apoptosis and decreasing nitric oxide production. This study reported that chitosan and its oligosaccharides, AST as well as selenium could be the principle compounds accountable for the anticancer activity of hydro-alcoholic extract of crab[103]. Chitosan from shrimp shell waste reduced the proliferation of MCF-7 cells without affecting normal fibroblasts by stimulating G2/M arrest and apoptosis, decreasing Bcl2 expression and upregulating p53 levels[104,105]. AST from shrimps, a marine crustacean, induced PCD in BC cells via alteration in expression of cyclin D1, Bax, p53, and Bcl2 through arrest at G0/G1 phase[106]. Chondroitin sulfate from shrimps inhibited angiogenesis by reducing tube formation via inhibiting the expression of VEGF[107]. β-carotene stimulated apoptosis in BC cells by inducing the release of cytochrome C, increasing PPAR-γ (peroxisome proliferator-activated receptor gamma), and p21(WAF1/CIP1) expression and decreasing cyclooxygenase-2 expression through ROS generation[108]. These studies suggest that bioactive compounds from marine crustaceans are potential targets of BC.
Table 2.
Marine compounds with potential anticancer activity against breast cancer from crustaceans
|
Marine compound
|
Chemical nature
|
Mechanism of action
|
Ref.
|
| Hydro-alcoholic extract of crab | Chitosan and Astaxanthin | Reduced proliferation by inducing apoptosis and decreasing nitric oxide production | Moghtaderi et al[74] |
| Chitosan | Polysaccharide | (1) Reduced proliferation without affecting normal fibroblasts by inducing arrest at the G2M phase; and (2) Induced apoptosis by decreasing the expression of Bcl2 via elevation of p53 level | Resmi et al[104] and Mohamed et al[105] |
| Astaxanthin | Oligosaccharide | Induced PCD in BC cells via alteration in the cyclin D1, p53, Bax, and Bcl2 expression through inducing arrest at G0/G1 stage | Resmi et al[104] and Mohamed et al[105] |
| Chondroitin sulfate | Muco-polysaccharide | Inhibited angiogenesis by reducing tube formation via inhibiting the expression of VEGF | Chen et al[75] |
| β-carotene | Terpenoids | Stimulated apoptosis in BC cells by inducing the release of cytochrome C, increasing PPAR-γ, and p21 (WAF1/CIP1) expression and decreasing cyclooxygenase-2 expression through ROS generation | Moon et al[102] |
PCD: Programmed cell death; BC: Breast cancer; VEGF: Vascular endothelial growth factor; ROS: Reactive oxygen species; PPAR-γ: Peroxisome proliferator-activated receptor gamma; WAF1/CIP1: Cyclin-dependent kinase inhibitor p21.
Marine compounds with potential anti-BC activity from marine fishes
The muscle and live tissues of marine fishes have been described as a source of anticancer compounds (Table 3). Epidemiological studies show that high consumption of fish and seafood in Asian countries correlates with a low prevalence of BC. A meta-analysis studies reported that high consumption of marine fish can reduce the risk of BC due to the presence of n-3 polyunsaturated fatty acids (PUFA) as well as α-linolenic acid[109]. Another epidemiological study reported a positive association between the reduction of invasive BC risk and intake of fatty fish but not with the intake of lean fish[110]. Some in vitro and animal studies also confirmed the inhibitory effect of eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA), and n-3 PUFA from marine fish on BC growth. The intake of fish containing EPA and DHA showed a dose-dependent reduction of BC risk[111]. Two peptides (K092A and K092B) from marine fish stimulated apoptosis in BC cells by altering the cytoskeleton via targeting actin and tubulin, halting at the G2M phase and decreasing mitochondrial activity[112]. A tetrahydroisoquinoline alkaloid, trabectedin from tunicate, induced death receptor-mediated apoptosis by increasing the expression of TNF-related apoptosis-inducing ligand (TRAIL)-R1, -R2, Fas, TNF RI, and FADD (fas-associated protein with death domain) in BC cells[113]. Hence, marine compounds extracted from marine fish can display a wide and divers’ range of anti-cancer effects and warrant further study to exploit their potential therapeutic properties in the context of BC.
Table 3.
Marine compounds with potential anticancer activity from marine fishes and invertebrates against breast cancer
|
Marine compound
|
Chemical nature
|
Mechanism of action
|
Ref.
|
| n-3 PUFA, α-linolenic acid | Fatty acids | Decreased the risk of BC | Moghtaderi et al[74] |
| EPA, DHA, n-3 fatty acids | Fatty acids | (1) Decreased the risk of BC; and (2) Inhibited growth BC cells | Resmi et al[104] and Mohamed et al[105] |
| K092A and K092B | Peptides | Induced apoptosis by altering the cytoskeleton via targeting actin and tubulin and halting cell cycle at the G2M and decreasing mitochondrial activity | Resmi et al[104] and Mohamed et al[105] |
| Tetrahydro-isoquinoline | Alkaloid | (1) Induced death receptor-mediated apoptosis by increasing the expression of TRAIL-R1, -R2, Fas, TNF RI, and FADD; and (2) Induced mitochondrial-mediated apoptosis by decreasing the of Bcl2 and Bcl-XL expression and increasing the Bax, Bad, cytochrome C and caspase 3 expression via increasing the ROS generation | Chen et al[75] |
BC: Breast cancer; EPA: Eicosapentaenoic acid; DHA: Docosahexaenoic acid; PUFA: Polyunsaturated fatty acids; TRAIL: Tumor necrosis factor-related apoptosis-inducing ligand; TNF: Tumor necrosis factor; FADD: Fas-associated protein with death domain; ROS: Reactive oxygen species.
Combination of marine compounds and chemotherapeutics
Marine compounds can resensitize resistant BC cells to chemotherapy (Table 4). For instance, marine-derived cyclic peptide, elisidepsin, and cisplatin or paclitaxel showed synergistic toxicity on BC cells by reducing the phosphorylation of Akt and inhibiting the MAPK pathway via targeting ERBB expression[114]. Sipholenol A increased the sensitivity of paclitaxel in BC cells by inhibiting P-glycoprotein (P-gp) and multidrug-resistant associated protein (MRP1)[115]. Permethyl ningalin B, resensitized BC cells to paclitaxel by inhibiting drug efflux activity of P-gp and increasing drug accumu-lation[116]. Preclinical studies reported that trabectedin (ET-743) used with a combination of cisplatin or paclitaxel or doxorubicin displayed an additive effect in the preclinical system[117]. Moreover, marine polysaccharides, in combination with cisplatin, synergistically inhibited TNBC cells growth and migration by blocking MMP-2 and MMP-9 expression[118]. Iturin A, combined with docetaxel, substantially sensitized docetaxel-resistant TNBC cells by reducing proliferation via massive arrest at the G1 phase, increasing caspase 3 levels and inhibiting Akt and its downstream signaling pathways[119]. Renieramycin M, a tetrahydroisoquinoline from blue sponges, sensitized MCF-7 cells synergistically to doxorubicin by promoting doxorubicin-induced DNA damage, cell cycle arrest, and apoptosis via downregulating ERBB/PI3K-Akt, integrin, and focal adhesion signaling[120]. Zhang et al[121] demon-strated that marine sulfated polysaccharide in a combination of pH-sensitive doxorubicin releasing nanosystem inhibited growth and progression of BC cells in tumor-bearing mice. Furthermore, papuamine, a marine compound from sponges, inhibited the colony-forming ability of BC cells at a subtoxic concentration by targeting activation of JNK[122]. Hence, marine compounds can resensitize refractory BC cells to chemotherapy and display promising potential as therapeutic companion tools to overcome drug resistance in BC.
Table 4.
Marine compounds with potential anticancer activity and chemotherapeutics against breast cancer
|
Marine compound
|
Chemical nature
|
Mechanism of action
|
Ref.
|
| Elisidepsin | Cyclic peptide | With combination of cisplatin or paclitaxel showed synergistic toxicity on BC cells by reducing the phosphorylation of Akt and inhibiting the MAPK pathway via targeting ErbB expression | Moghtaderi et al[74] |
| Sipholenol A | Triterpene | Increased the sensitivity of paclitaxel in BC cells by inhibiting P-gp and MRP1 | Resmi et al[104] and Mohamed et al[105] |
| Permethyl ningalin B | Pyrrole-containingcompound | Sensitized BC cells to paclitaxel by inhibiting drug efflux activity of P-gp and increasing drug accumulation | Resmi et al[104] and Mohamed et al[105] |
| Trabectedin | Alkaloid | Combination with cisplatin or paclitaxel or doxorubicin showed an additive effect in the preclinical system | Chen et al[75] |
| Marine polysaccharide | Polysaccharide | In combination with cisplatin, synergistically inhibited the proliferation and migration by blocking the MMP-2 and MMP-9 expression | Chen et al[75] |
| Iturin A | Cyclo-lipopeptide | In combination with docetaxel, substantially sensitized docetaxel-resistant TNBC cells by reducing proliferation via massive arresting at the G1 stage and activating caspase 3 as well as inhibiting Akt and its downstream signaling pathways | Dey et al[119] |
| Renieramycin M | Tetrahydro-isoquinoline | Sensitized MCF-7 cells synergistically to doxorubicin by promoting doxorubicin-induced DNA damage, cell cycle arrest, and apoptosis via downregulating ErbB/PI3K-Akt, integrin, and focal adhesion signaling | Tun et al[120] |
| Sulfated polysaccharide | - | In combination with pH-sensitive DOX releasing nanosystem inhibited growth and metastasis of BC cells in tumor-bearing mice | Zhang et al[121] |
| Papuamine | Pentacyclic alkaloid | Inhibited colony formation of BC cells by targeting activation of JNK | Kanno et al[122] |
BC: Breast cancer; MAPK: Mitogen-activated protein kinase; P-gp: P-glycoprotein; MRP: Multidrug-resistant associated protein; MMP: Matrix metalloproteinase; TNBC: Triple-negative breast cancer; DOX: Doxorubicin; JNK: c-Jun N-terminal kinase.
CONCLUSION
As this in-depth review shows, BC is a highly complex and heterogenous disease and remains one of the major causes of female mortality worldwide. Hence, novel diagnostic tools and drug targets are needed to develop effective management and treatment regimens that overcome chemoresistance and increase survival. The search for novel treatment BC strategies has prompted the clinical investigation of various anticancer compounds extracted from marine microorganisms. As per our detailed survey, marine compounds display promising antineoplastic therapeutic potential for BC. In fact, their enormous genetic diversity supports a wide range of anti-cancer and immunomodulatory mechanisms of action that could be harnessed for BC treatment. Future pharmaceutical development is required to improve the pharmacokinetic properties of these compounds and evaluate their efficacy. Although marine compounds represent a relatively young discipline, their chemical diversity, eco-friendliness and therapeutic properties render them a promising new source of anticancer drugs. The use of novel bioengineering tools will enormously help in improving their production and clinical development for the treatment of BC and other cancers.
Footnotes
Conflict-of-interest statement: None.
Manuscript source: Invited manuscript
Corresponding Author's Membership in Professional Societies: American Association for Cancer Research, No. 229378.
Peer-review started: December 23, 2020
First decision: January 11, 2021
Article in press: February 20, 2021
Specialty type: Biochemistry and molecular biology
Country/Territory of origin: United States
Peer-review report’s scientific quality classification
Grade A (Excellent): 0
Grade B (Very good): B
Grade C (Good): C
Grade D (Fair): 0
Grade E (Poor): 0
P-Reviewer: Gazouli M S-Editor: Gao CC L-Editor: A P-Editor: Yuan YY
Contributor Information
Rama Rao Malla, Department of Biochemistry and Bioinformatics, GITAM (Deemed to be University), Visakhapatnam 530045, AP, India.
Batoul Farran, Department of Hematology and Medical Oncology, Emory University, Atlanta, GA 30322, United States.
Ganji Purnachandra Nagaraju, Department of Hematology and Medical Oncology, Winship Cancer Institute, Emory University, Atlanta, GA 30322, United States. pganji@emory.edu.
References
- 1.Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424. doi: 10.3322/caac.21492. [DOI] [PubMed] [Google Scholar]
- 2.Byler S, Goldgar S, Heerboth S, Leary M, Housman G, Moulton K, Sarkar S. Genetic and epigenetic aspects of breast cancer progression and therapy. Anticancer Res. 2014;34:1071–1077. [PubMed] [Google Scholar]
- 3.Deepak KGK, Vempati R, Nagaraju GP, Dasari VR, S N, Rao DN, Malla RR. Tumor microenvironment: Challenges and opportunities in targeting metastasis of triple negative breast cancer. Pharmacol Res. 2020;153:104683. doi: 10.1016/j.phrs.2020.104683. [DOI] [PubMed] [Google Scholar]
- 4.Iyengar NM, Gucalp A, Dannenberg AJ, Hudis CA. Obesity and Cancer Mechanisms: Tumor Microenvironment and Inflammation. J Clin Oncol. 2016;34:4270–4276. doi: 10.1200/JCO.2016.67.4283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Newman DJ, Cragg GM. Drugs and Drug Candidates from Marine Sources: An Assessment of the Current "State of Play". Planta Med. 2016;82:775–789. doi: 10.1055/s-0042-101353. [DOI] [PubMed] [Google Scholar]
- 6.Shinde P, Banerjee P, Mandhare A. Marine natural products as source of new drugs: a patent review (2015-2018) Expert Opin Ther Pat. 2019;29:283–309. doi: 10.1080/13543776.2019.1598972. [DOI] [PubMed] [Google Scholar]
- 7.Khalifa SAM, Elias N, Farag MA, Chen L, Saeed A, Hegazy MF, Moustafa MS, Abd El-Wahed A, Al-Mousawi SM, Musharraf SG, Chang FR, Iwasaki A, Suenaga K, Alajlani M, Göransson U, El-Seedi HR. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar Drugs. 2019;17 doi: 10.3390/md17090491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Luen S, Virassamy B, Savas P, Salgado R, Loi S. The genomic landscape of breast cancer and its interaction with host immunity. Breast. 2016;29:241–250. doi: 10.1016/j.breast.2016.07.015. [DOI] [PubMed] [Google Scholar]
- 9.Gómez-Flores-Ramos L, Castro-Sánchez A, Peña-Curiel O, Mohar-Betancourt A. Molecular Biology In Young Women With Breast Cancer: From Tumor Gene Expression To DNA Mutations. Rev Invest Clin. 2017;69:181–192. doi: 10.24875/ric.17002225. [DOI] [PubMed] [Google Scholar]
- 10.Azim HA Jr, Partridge AH. Biology of breast cancer in young women. Breast Cancer Res. 2014;16:427. doi: 10.1186/s13058-014-0427-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Erić I, Petek Erić A, Kristek J, Koprivčić I, Babić M. BREAST CANCER IN YOUNG WOMEN: PATHOLOGIC AND IMMUNOHISTOCHEMICAL FEATURES. Acta Clin Croat. 2018;57:497–502. doi: 10.20471/acc.2018.57.03.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ribnikar D, Ribeiro JM, Pinto D, Sousa B, Pinto AC, Gomes E, Moser EC, Cardoso MJ, Cardoso F. Breast cancer under age 40: a different approach. Curr Treat Options Oncol. 2015;16:16. doi: 10.1007/s11864-015-0334-8. [DOI] [PubMed] [Google Scholar]
- 13.Rosenberg SM, Partridge AH. Management of breast cancer in very young women. Breast. 2015;24 Suppl 2:S154–S158. doi: 10.1016/j.breast.2015.07.036. [DOI] [PubMed] [Google Scholar]
- 14.Johnson RH, Anders CK, Litton JK, Ruddy KJ, Bleyer A. Breast cancer in adolescents and young adults. Pediatr Blood Cancer. 2018;65:e27397. doi: 10.1002/pbc.27397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Jolly TA, Williams GR, Bushan S, Pergolotti M, Nyrop KA, Jones EL, Muss HB. Adjuvant treatment for older women with invasive breast cancer. Womens Health (Lond) 2016;12:129–45; quiz 145. doi: 10.2217/whe.15.92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Jeon YW, You SH, Lee JE, Youn HJ, Lim W, Han JH, Park CH, Kim YS. Optimal treatment of breast cancer in women older than 75 years: a Korea Breast Cancer Registry analysis. Breast Cancer Res Treat. 2019;178:693–701. doi: 10.1007/s10549-019-05426-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Abdel-Hafiz HA, Horwitz KB. Role of epigenetic modifications in luminal breast cancer. Epigenomics. 2015;7:847–862. doi: 10.2217/epi.15.10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Guiu S, Wolfer A, Jacot W, Fumoleau P, Romieu G, Bonnetain F, Fiche M. Invasive lobular breast cancer and its variants: how special are they for systemic therapy decisions? Crit Rev Oncol Hematol. 2014;92:235–257. doi: 10.1016/j.critrevonc.2014.07.003. [DOI] [PubMed] [Google Scholar]
- 19.Holm K, Staaf J, Lauss M, Aine M, Lindgren D, Bendahl PO, Vallon-Christersson J, Barkardottir RB, Höglund M, Borg Å, Jönsson G, Ringnér M. An integrated genomics analysis of epigenetic subtypes in human breast tumors links DNA methylation patterns to chromatin states in normal mammary cells. Breast Cancer Res. 2016;18:27. doi: 10.1186/s13058-016-0685-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dumitrescu RG. Interplay Between Genetic and Epigenetic Changes in Breast Cancer Subtypes. Methods Mol Biol. 2018;1856:19–34. doi: 10.1007/978-1-4939-8751-1_2. [DOI] [PubMed] [Google Scholar]
- 21.Zheng T, Wang A, Hu D, Wang Y. Molecular mechanisms of breast cancer metastasis by gene expression profile analysis. Mol Med Rep. 2017;16:4671–4677. doi: 10.3892/mmr.2017.7157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rodilla V, Fre S. Cellular Plasticity of Mammary Epithelial Cells Underlies Heterogeneity of Breast Cancer. Biomedicines. 2018;6 doi: 10.3390/biomedicines6040103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yeo SK, Guan JL. Breast Cancer: Multiple Subtypes within a Tumor? Trends Cancer. 2017;3:753–760. doi: 10.1016/j.trecan.2017.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gao R, Davis A, McDonald TO, Sei E, Shi X, Wang Y, Tsai PC, Casasent A, Waters J, Zhang H, Meric-Bernstam F, Michor F, Navin NE. Punctuated copy number evolution and clonal stasis in triple-negative breast cancer. Nat Genet. 2016;48:1119–1130. doi: 10.1038/ng.3641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chung W, Eum HH, Lee HO, Lee KM, Lee HB, Kim KT, Ryu HS, Kim S, Lee JE, Park YH, Kan Z, Han W, Park WY. Single-cell RNA-seq enables comprehensive tumour and immune cell profiling in primary breast cancer. Nat Commun. 2017;8:15081. doi: 10.1038/ncomms15081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lopez G, Costanza J, Colleoni M, Fontana L, Ferrero S, Miozzo M, Fusco N. Molecular Insights into the Classification of Luminal Breast Cancers: The Genomic Heterogeneity of Progesterone-Negative Tumors. Int J Mol Sci. 2019;20 doi: 10.3390/ijms20030510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ma D, Jiang YZ, Liu XY, Liu YR, Shao ZM. Clinical and molecular relevance of mutant-allele tumor heterogeneity in breast cancer. Breast Cancer Res Treat. 2017;162:39–48. doi: 10.1007/s10549-017-4113-z. [DOI] [PubMed] [Google Scholar]
- 28.Coughlin SS. Epidemiology of Breast Cancer in Women. Adv Exp Med Biol. 2019;1152:9–29. doi: 10.1007/978-3-030-20301-6_2. [DOI] [PubMed] [Google Scholar]
- 29.Kleibl Z, Kristensen VN. Women at high risk of breast cancer: Molecular characteristics, clinical presentation and management. Breast. 2016;28:136–144. doi: 10.1016/j.breast.2016.05.006. [DOI] [PubMed] [Google Scholar]
- 30.Ito H, Matsuo K. Molecular epidemiology, and possible real-world applications in breast cancer. Breast Cancer. 2016;23:33–38. doi: 10.1007/s12282-015-0609-8. [DOI] [PubMed] [Google Scholar]
- 31.Schmidt T, van Mackelenbergh M, Wesch D, Mundhenke C. Physical activity influences the immune system of breast cancer patients. J Cancer Res Ther. 2017;13:392–398. doi: 10.4103/0973-1482.150356. [DOI] [PubMed] [Google Scholar]
- 32.Liubomirski Y, Lerrer S, Meshel T, Rubinstein-Achiasaf L, Morein D, Wiemann S, Körner C, Ben-Baruch A. Tumor-Stroma-Inflammation Networks Promote Pro-metastatic Chemokines and Aggressiveness Characteristics in Triple-Negative Breast Cancer. Front Immunol. 2019;10:757. doi: 10.3389/fimmu.2019.00757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zhou J, Wang XH, Zhao YX, Chen C, Xu XY, Sun Q, Wu HY, Chen M, Sang JF, Su L, Tang XQ, Shi XB, Zhang Y, Yu Q, Yao YZ, Zhang WJ. Cancer-Associated Fibroblasts Correlate with Tumor-Associated Macrophages Infiltration and Lymphatic Metastasis in Triple Negative Breast Cancer Patients. J Cancer. 2018;9:4635–4641. doi: 10.7150/jca.28583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Takai K, Le A, Weaver VM, Werb Z. Targeting the cancer-associated fibroblasts as a treatment in triple-negative breast cancer. Oncotarget. 2016;7:82889–82901. doi: 10.18632/oncotarget.12658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Coffelt SB, Kersten K, Doornebal CW, Weiden J, Vrijland K, Hau CS, Verstegen NJM, Ciampricotti M, Hawinkels LJAC, Jonkers J, de Visser KE. IL-17-producing γδ T cells and neutrophils conspire to promote breast cancer metastasis. Nature. 2015;522:345–348. doi: 10.1038/nature14282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wculek SK, Malanchi I. Neutrophils support lung colonization of metastasis-initiating breast cancer cells. Nature. 2015;528:413–417. doi: 10.1038/nature16140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Szczerba BM, Castro-Giner F, Vetter M, Krol I, Gkountela S, Landin J, Scheidmann MC, Donato C, Scherrer R, Singer J, Beisel C, Kurzeder C, Heinzelmann-Schwarz V, Rochlitz C, Weber WP, Beerenwinkel N, Aceto N. Neutrophils escort circulating tumour cells to enable cell cycle progression. Nature. 2019;566:553–557. doi: 10.1038/s41586-019-0915-y. [DOI] [PubMed] [Google Scholar]
- 38.Yue Z, Yuan Z, Zeng L, Wang Y, Lai L, Li J, Sun P, Xue X, Qi J, Yang Z, Zheng Y, Fang Y, Li D, Siwko S, Li Y, Luo J, Liu M. LGR4 modulates breast cancer initiation, metastasis, and cancer stem cells. FASEB J. 2018;32:2422–2437. doi: 10.1096/fj.201700897R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Albini A, Bruno A, Noonan DM, Mortara L. Contribution to Tumor Angiogenesis From Innate Immune Cells Within the Tumor Microenvironment: Implications for Immunotherapy. Front Immunol. 2018;9:527. [Google Scholar]
- 40.Lin L, Chen YS, Yao YD, Chen JQ, Chen JN, Huang SY, Zeng YJ, Yao HR, Zeng SH, Fu YS, Song EW. CCL18 from tumor-associated macrophages promotes angiogenesis in breast cancer. Oncotarget. 2015;6:34758–34773. doi: 10.18632/oncotarget.5325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Choi J, Gyamfi J, Jang H, Koo JS. The role of tumor-associated macrophage in breast cancer biology. Histol Histopathol. 2018;33:133–145. doi: 10.14670/HH-11-916. [DOI] [PubMed] [Google Scholar]
- 42.Du YE, Tu G, Yang G, Li G, Yang D, Lang L, Xi L, Sun K, Chen Y, Shu K, Liao H, Liu M, Hou Y. MiR-205/YAP1 in Activated Fibroblasts of Breast Tumor Promotes VEGF-independent Angiogenesis through STAT3 Signaling. Theranostics. 2017;7:3972–3988. doi: 10.7150/thno.18990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Eiro N, González L, Martínez-Ordoñez A, Fernandez-Garcia B, González LO, Cid S, Dominguez F, Perez-Fernandez R, Vizoso FJ. Cancer-associated fibroblasts affect breast cancer cell gene expression, invasion and angiogenesis. Cell Oncol (Dordr) 2018;41:369–378. doi: 10.1007/s13402-018-0371-y. [DOI] [PubMed] [Google Scholar]
- 44.Kugeratski FG, Atkinson SJ, Neilson LJ, Lilla S, Knight JRP, Serneels J, Juin A, Ismail S, Bryant DM, Markert EK, Machesky LM, Mazzone M, Sansom OJ, Zanivan S. Hypoxic cancer-associated fibroblasts increase NCBP2-AS2/HIAR to promote endothelial sprouting through enhanced VEGF signaling. Sci Signal. 2019;12 doi: 10.1126/scisignal.aan8247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Maji S, Chaudhary P, Akopova I, Nguyen PM, Hare RJ, Gryczynski I, Vishwanatha JK. Exosomal Annexin II Promotes Angiogenesis and Breast Cancer Metastasis. Mol Cancer Res. 2017;15:93–105. doi: 10.1158/1541-7786.MCR-16-0163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Jung KO, Youn H, Lee CH, Kang KW, Chung JK. Visualization of exosome-mediated miR-210 transfer from hypoxic tumor cells. Oncotarget. 2017;8:9899–9910. doi: 10.18632/oncotarget.14247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Lin Q, Chen X, Meng F, Ogawa K, Li M, Song R, Zhang S, Zhang Z, Kong X, Xu Q, He F, Bai X, Sun B, Hung MC, Liu L, Wands J, Dong X. ASPH-notch Axis guided Exosomal delivery of Prometastatic Secretome renders breast Cancer multi-organ metastasis. Mol Cancer. 2019;18:156. doi: 10.1186/s12943-019-1077-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Rodríguez M, Silva J, Herrera A, Herrera M, Peña C, Martín P, Gil-Calderón B, Larriba MJ, Coronado MJ, Soldevilla B, Turrión VS, Provencio M, Sánchez A, Bonilla F, García-Barberán V. Exosomes enriched in stemness/metastatic-related mRNAS promote oncogenic potential in breast cancer. Oncotarget. 2015;6:40575–40587. doi: 10.18632/oncotarget.5818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lee JK, Park SR, Jung BK, Jeon YK, Lee YS, Kim MK, Kim YG, Jang JY, Kim CW. Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells. PLoS One. 2013;8:e84256. doi: 10.1371/journal.pone.0084256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Nguyen NP, Almeida FS, Chi A, Nguyen LM, Cohen D, Karlsson U, Vinh-Hung V. Molecular biology of breast cancer stem cells: potential clinical applications. Cancer Treat Rev. 2010;36:485–491. doi: 10.1016/j.ctrv.2010.02.016. [DOI] [PubMed] [Google Scholar]
- 51.Kubli SP, Bassi C, Roux C, Wakeham A, Göbl C, Zhou W, Jafari SM, Snow B, Jones L, Palomero L, Thu KL, Cassetta L, Soong D, Berger T, Ramachandran P, Baniasadi SP, Duncan G, Lindzen M, Yarden Y, Herranz C, Lazaro C, Chu MF, Haight J, Tinto P, Silvester J, Cescon DW, Petit A, Pettersson S, Pollard JW, Mak TW, Pujana MA, Cappello P, Gorrini C. AhR controls redox homeostasis and shapes the tumor microenvironment in BRCA1-associated breast cancer. Proc Natl Acad Sci U S A. 2019;116:3604–3613. doi: 10.1073/pnas.1815126116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Roux C, Jafari SM, Shinde R, Duncan G, Cescon DW, Silvester J, Chu MF, Hodgson K, Berger T, Wakeham A, Palomero L, Garcia-Valero M, Pujana MA, Mak TW, McGaha TL, Cappello P, Gorrini C. Reactive oxygen species modulate macrophage immunosuppressive phenotype through the up-regulation of PD-L1. Proc Natl Acad Sci U S A. 2019;116:4326–4335. doi: 10.1073/pnas.1819473116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Parekh A, Das S, Parida S, Das CK, Dutta D, Mallick SK, Wu PH, Kumar BNP, Bharti R, Dey G, Banerjee K, Rajput S, Bharadwaj D, Pal I, Dey KK, Rajesh Y, Jena BC, Biswas A, Banik P, Pradhan AK, Das SK, Das AK, Dhara S, Fisher PB, Wirtz D, Mills GB, Mandal M. Multi-nucleated cells use ROS to induce breast cancer chemo-resistance in vitro and in vivo. Oncogene. 2018;37:4546–4561. doi: 10.1038/s41388-018-0272-6. [DOI] [PubMed] [Google Scholar]
- 54.Lv W, Chen N, Lin Y, Ma H, Ruan Y, Li Z, Li X, Pan X, Tian X. Macrophage migration inhibitory factor promotes breast cancer metastasis via activation of HMGB1/TLR4/NF kappa B axis. Cancer Lett. 2016;375:245–255. doi: 10.1016/j.canlet.2016.02.005. [DOI] [PubMed] [Google Scholar]
- 55.Jadaliha M, Zong X, Malakar P, Ray T, Singh DK, Freier SM, Jensen T, Prasanth SG, Karni R, Ray PS, Prasanth KV. Functional and prognostic significance of long non-coding RNA MALAT1 as a metastasis driver in ER negative lymph node negative breast cancer. Oncotarget. 2016;7:40418–40436. doi: 10.18632/oncotarget.9622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Arun G, Spector DL. MALAT1 Long non-coding RNA and breast cancer. RNA Biol. 2019;16:860–863. doi: 10.1080/15476286.2019.1592072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Huang D, Chen J, Yang L, Ouyang Q, Li J, Lao L, Zhao J, Liu J, Lu Y, Xing Y, Chen F, Su F, Yao H, Liu Q, Su S, Song E. NKILA lncRNA promotes tumor immune evasion by sensitizing T cells to activation-induced cell death. Nat Immunol. 2018;19:1112–1125. doi: 10.1038/s41590-018-0207-y. [DOI] [PubMed] [Google Scholar]
- 58.Arun G, Diermeier S, Akerman M, Chang KC, Wilkinson JE, Hearn S, Kim Y, MacLeod AR, Krainer AR, Norton L, Brogi E, Egeblad M, Spector DL. Differentiation of mammary tumors and reduction in metastasis upon Malat1 LncRNA loss. Genes Dev. 2016;30:34–51. doi: 10.1101/gad.270959.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ou X, Gao G, Bazhabayi M, Zhang K, Liu F, Xiao X. MALAT1 and BACH1 are prognostic biomarkers for triple-negative breast cancer. J Cancer Res Ther. 2019;15:1597–1602. doi: 10.4103/jcrt.JCRT_282_19. [DOI] [PubMed] [Google Scholar]
- 60.Duan Y, Zhang X, Yang L, Dong X, Zheng Z, Cheng Y, Chen H, Lan B, Li D, Zhou J, Xuan C. Disruptor of telomeric silencing 1-like (DOT1L) is involved in breast cancer metastasis via transcriptional regulation of MALAT1 and ZEB2. J Genet Genomics. 2019;46:591–594. doi: 10.1016/j.jgg.2019.11.008. [DOI] [PubMed] [Google Scholar]
- 61.Stone JK, Kim JH, Vukadin L, Richard A, Giannini HK, Lim SS, Tan M, Ahn EE. Hypoxia induces cancer cell-specific chromatin interactions and increases MALAT1 expression in breast cancer cells. J Biol Chem. 2019;294:11213–11224. doi: 10.1074/jbc.RA118.006889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kim J, Piao HL, Kim BJ, Yao F, Han Z, Wang Y, Xiao Z, Siverly AN, Lawhon SE, Ton BN, Lee H, Zhou Z, Gan B, Nakagawa S, Ellis MJ, Liang H, Hung MC, You MJ, Sun Y, Ma L. Long noncoding RNA MALAT1 suppresses breast cancer metastasis. Nat Genet. 2018;50:1705–1715. doi: 10.1038/s41588-018-0252-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Gupta RA, Shah N, Wang KC, Kim J, Horlings HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, Wang Y, Brzoska P, Kong B, Li R, West RB, van de Vijver MJ, Sukumar S, Chang HY. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010;464:1071–1076. doi: 10.1038/nature08975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Zhao W, Geng D, Li S, Chen Z, Sun M. LncRNA HOTAIR influences cell growth, migration, invasion, and apoptosis via the miR-20a-5p/HMGA2 axis in breast cancer. Cancer Med. 2018;7:842–855. doi: 10.1002/cam4.1353. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 65.Zhang CY, Yu MS, Li X, Zhang Z, Han CR, Yan B. Overexpression of long non-coding RNA MEG3 suppresses breast cancer cell proliferation, invasion, and angiogenesis through AKT pathway. Tumour Biol. 2017;39:1010428317701311. doi: 10.1177/1010428317701311. [DOI] [PubMed] [Google Scholar]
- 66.Luo LH, Rao L, Luo LF, Chen K, Ran RZ, Liu XL. Long non-coding RNA NKILA inhibited angiogenesis of breast cancer through NF-κB/IL-6 signaling pathway. Microvasc Res. 2020;129:103968. doi: 10.1016/j.mvr.2019.103968. [DOI] [PubMed] [Google Scholar]
- 67.Sun X, Huang T, Zhang C, Zhang S, Wang Y, Zhang Q, Liu Z. Long non-coding RNA LINC00968 reduces cell proliferation and migration and angiogenesis in breast cancer through up-regulation of PROX1 by reducing hsa-miR-423-5p. Cell Cycle. 2019;18:1908–1924. doi: 10.1080/15384101.2019.1632641. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 68.Tu Z, Schmöllerl J, Cuiffo BG, Karnoub AE. Microenvironmental Regulation of Long Noncoding RNA LINC01133 Promotes Cancer Stem Cell-Like Phenotypic Traits in Triple-Negative Breast Cancers. Stem Cells. 2019;37:1281–1292. doi: 10.1002/stem.3055. [DOI] [PubMed] [Google Scholar]
- 69.García-Venzor A, Mandujano-Tinoco EA, Lizarraga F, Zampedri C, Krötzsch E, Salgado RM, Dávila-Borja VM, Encarnación-Guevara S, Melendez-Zajgla J, Maldonado V. Microenvironment-regulated lncRNA-HAL is able to promote stemness in breast cancer cells. Biochim Biophys Acta Mol Cell Res. 2019;1866:118523. doi: 10.1016/j.bbamcr.2019.118523. [DOI] [PubMed] [Google Scholar]
- 70.Zhou L, Tian Y, Guo F, Yu B, Li J, Xu H, Su Z. LincRNA-p21 knockdown reversed tumor-associated macrophages function by promoting MDM2 to antagonize* p53 activation and alleviate breast cancer development. Cancer Immunol Immunother. 2020;69:835–846. doi: 10.1007/s00262-020-02511-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Hu Q, Ye Y, Chan LC, Li Y, Liang K, Lin A, Egranov SD, Zhang Y, Xia W, Gong J, Pan Y, Chatterjee SS, Yao J, Evans KW, Nguyen TK, Park PK, Liu J, Coarfa C, Donepudi SR, Putluri V, Putluri N, Sreekumar A, Ambati CR, Hawke DH, Marks JR, Gunaratne PH, Caudle AS, Sahin AA, Hortobagyi GN, Meric-Bernstam F, Chen L, Yu D, Hung MC, Curran MA, Han L, Lin C, Yang L. Oncogenic lncRNA downregulates cancer cell antigen presentation and intrinsic tumor suppression. Nat Immunol. 2019;20:835–851. doi: 10.1038/s41590-019-0400-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Dyshlovoy SA, Honecker F. Marine Compounds and Cancer: 2017 Updates. Mar Drugs. 2018;16 doi: 10.3390/md16020041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Dobretsov S, Tamimi Y, Al-Kindi MA, Burney I. Screening for Anti-Cancer Compounds in Marine Organisms in Oman. Sultan Qaboos Univ Med J. 2016;16:e168–e174. doi: 10.18295/squmj.2016.16.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Moghtaderi H, Sepehri H, Delphi L, Attari F. Gallic acid and curcumin induce cytotoxicity and apoptosis in human breast cancer cell MDA-MB-231. Bioimpacts. 2018;8:185–194. doi: 10.15171/bi.2018.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Chen YJ, Lee YC, Huang CH, Chang LS. Gallic acid-capped gold nanoparticles inhibit EGF-induced MMP-9 expression through suppression of p300 stabilization and NFκB/c-Jun activation in breast cancer MDA-MB-231 cells. Toxicol Appl Pharmacol. 2016;310:98–107. doi: 10.1016/j.taap.2016.09.007. [DOI] [PubMed] [Google Scholar]
- 76.Shrestha S, Sorolla A, Fromont J, Blancafort P, Flematti GR. Crambescidin 800, Isolated from the Marine Sponge Monanchora viridis, Induces Cell Cycle Arrest and Apoptosis in Triple-Negative Breast Cancer Cells. Mar Drugs. 2018;16 doi: 10.3390/md16020053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Cao R, Jin W, Shan Y, Wang J, Liu G, Kuang S, Sun C. Marine Bacterial Polysaccharide EPS11 Inhibits Cancer Cell Growth via Blocking Cell Adhesion and Stimulating Anoikis. Mar Drugs. 2018;16 doi: 10.3390/md16030085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Vaikundamoorthy R, Krishnamoorthy V, Vilwanathan R, Rajendran R. Structural characterization and anticancer activity (MCF7 and MDA-MB-231) of polysaccharides fractionated from brown seaweed Sargassum wightii. Int J Biol Macromol. 2018;111:1229–1237. doi: 10.1016/j.ijbiomac.2018.01.125. [DOI] [PubMed] [Google Scholar]
- 79.Murad H, Ghannam A, Al-Ktaifani M, Abbas A, Hawat M. Algal sulfated carrageenan inhibits proliferation of MDA-MB-231 cells via apoptosis regulatory genes. Mol Med Rep. 2015;11:2153–2158. doi: 10.3892/mmr.2014.2915. [DOI] [PubMed] [Google Scholar]
- 80.Park GT, Go RE, Lee HM, Lee GA, Kim CW, Seo JW, Hong WK, Choi KC, Hwang KA. Potential Anti-proliferative and Immunomodulatory Effects of Marine Microalgal Exopolysaccharide on Various Human Cancer Cells and Lymphocytes In Vitro. Mar Biotechnol (NY) 2017;19:136–146. doi: 10.1007/s10126-017-9735-y. [DOI] [PubMed] [Google Scholar]
- 81.Xie Q, Yang Z, Huang X, Zhang Z, Li J, Ju J, Zhang H, Ma J. Ilamycin C induces apoptosis and inhibits migration and invasion in triple-negative breast cancer by suppressing IL-6/STAT3 pathway. J Hematol Oncol. 2019;12:60. doi: 10.1186/s13045-019-0744-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Zhou W, Fang H, Wu Q, Wang X, Liu R, Li F, Xiao J, Yuan L, Zhou Z, Ma J, Wang L, Zhao W, You H, Ju J, Feng J, Chen C. Ilamycin E, a natural product of marine actinomycete, inhibits triple-negative breast cancer partially through ER stress-CHOP-Bcl-2. Int J Biol Sci. 2019;15:1723–1732. doi: 10.7150/ijbs.35284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Al-Awadhi FH, Paul VJ, Luesch H. Structural Diversity and Anticancer Activity of Marine-Derived Elastase Inhibitors: Key Features and Mechanisms Mediating the Antimetastatic Effects in Invasive Breast Cancer. Chembiochem. 2018;19:815–825. doi: 10.1002/cbic.201700627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Al-Awadhi FH, Salvador LA, Law BK, Paul VJ, Luesch H. Kempopeptin C, a Novel Marine-Derived Serine Protease Inhibitor Targeting Invasive Breast Cancer. Mar Drugs. 2017;15 doi: 10.3390/md15090290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kgk D, Kumari S, G S, Malla RR. Marine natural compound cyclo(L-leucyl-L-prolyl) peptide inhibits migration of triple negative breast cancer cells by disrupting interaction of CD151 and EGFR signaling. Chem Biol Interact. 2020;315:108872. doi: 10.1016/j.cbi.2019.108872. [DOI] [PubMed] [Google Scholar]
- 86.Lunagariya J, Liao X, Long W, Zhong S, Bhadja P, Li H, Zhao B, Xu S. Cytotoxicity Study of Cyclopentapeptide Analogues of Marine Natural Product Galaxamide towards Human Breast Cancer Cells. Oxid Med Cell Longev. 2017;2017:8392035. doi: 10.1155/2017/8392035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Al-Awadhi FH, Gao B, Rezaei MA, Kwan JC, Li C, Ye T, Paul VJ, Luesch H. Discovery, Synthesis, Pharmacological Profiling, and Biological Characterization of Brintonamides A-E, Novel Dual Protease and GPCR Modulators from a Marine Cyanobacterium. J Med Chem. 2018;61:6364–6378. doi: 10.1021/acs.jmedchem.8b00885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Dey G, Bharti R, Dhanarajan G, Das S, Dey KK, Kumar BN, Sen R, Mandal M. Marine lipopeptide Iturin A inhibits Akt mediated GSK3β and FoxO3a signaling and triggers apoptosis in breast cancer. Sci Rep. 2015;5:10316. doi: 10.1038/srep10316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.do Nascimento-Neto LG, Cabral MG, Carneiro RF, Silva Z, Arruda FVS, Nagano CS, Fernandes AR, Sampaio AH, Teixeira EH, Videira PA. Halilectin-3, a Lectin from the Marine Sponge Haliclona caerulea, Induces Apoptosis and Autophagy in Human Breast Cancer MCF7 Cells Through Caspase-9 Pathway and LC3-II Protein Expression. Anticancer Agents Med Chem. 2018;18:521–528. doi: 10.2174/1871520617666171114094847. [DOI] [PubMed] [Google Scholar]
- 90.Huang HW, Tang JY, Ou-Yang F, Wang HR, Guan PY, Huang CY, Chen CY, Hou MF, Sheu JH, Chang HW. Sinularin Selectively Kills Breast Cancer Cells Showing G2/M Arrest, Apoptosis, and Oxidative DNA Damage. Molecules. 2018;23 doi: 10.3390/molecules23040849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Foudah AI, Jain S, Busnena BA, El Sayed KA. Optimization of marine triterpene sipholenols as inhibitors of breast cancer migration and invasion. ChemMedChem. 2013;8:497–510. doi: 10.1002/cmdc.201200516. [DOI] [PubMed] [Google Scholar]
- 92.Pimentel AA, Felibertt P, Sojo F, Colman L, Mayora A, Silva ML, Rojas H, Dipolo R, Suarez AI, Compagnone RS, Arvelo F, Galindo-Castro I, De Sanctis JB, Chirino P, Benaim G. The marine sponge toxin agelasine B increases the intracellular Ca(2+) concentration and induces apoptosis in human breast cancer cells (MCF-7) Cancer Chemother Pharmacol. 2012;69:71–83. doi: 10.1007/s00280-011-1677-x. [DOI] [PubMed] [Google Scholar]
- 93.Yang F, Chen WD, Deng R, Li DD, Wu KW, Feng GK, Li HJ, Zhu XF. Hirsutanol A induces apoptosis and autophagy via reactive oxygen species accumulation in breast cancer MCF-7 cells. J Pharmacol Sci. 2012;119:214–220. doi: 10.1254/jphs.11235fp. [DOI] [PubMed] [Google Scholar]
- 94.Pec MK, Aguirre A, Moser-Thier K, Fernández JJ, Souto ML, Dorta J, Diáz-González F, Villar J. Induction of apoptosis in estrogen dependent and independent breast cancer cells by the marine terpenoid dehydrothyrsiferol. Biochem Pharmacol. 2003;65:1451–1461. doi: 10.1016/s0006-2952(03)00123-0. [DOI] [PubMed] [Google Scholar]
- 95.Dai J, Fishback JA, Zhou YD, Nagle DG. Sodwanone and yardenone triterpenes from a South African species of the marine sponge Axinella inhibit hypoxia-inducible factor-1 (HIF-1) activation in both breast and prostate tumor cells. J Nat Prod. 2006;69:1715–1720. doi: 10.1021/np060278q. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Sperlich J, Kerr R, Teusch N. The Marine Natural Product Pseudopterosin Blocks Cytokine Release of Triple-Negative Breast Cancer and Monocytic Leukemia Cells by Inhibiting NF-κB Signaling. Mar Drugs. 2017;15 doi: 10.3390/md15090262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.De AK, Muthiyan R, Mondal S, Mahanta N, Bhattacharya D, Ponraj P, Muniswamy K, Kundu A, Kundu MS, Sunder J, Karunakaran D, Bera AK, Roy SD, Malakar D. A Natural Quinazoline Derivative from Marine Sponge Hyrtios erectus Induces Apoptosis of Breast Cancer Cells via ROS Production and Intrinsic or Extrinsic Apoptosis Pathways. Mar Drugs. 2019;17 doi: 10.3390/md17120658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Sharifi S, Mostafavi PG, Tarasi R, Moradi AM, Givianrad MH, Farimani MM, Ebrahimi SN, Hamburger M, Niknejad H. Purified compounds from marine organism sea pen induce apoptosis in human breast cancer cell MDA-MB-231 and cervical cancer cell Hela. Eur J Pharmacol. 2020;877:173075. doi: 10.1016/j.ejphar.2020.173075. [DOI] [PubMed] [Google Scholar]
- 99.Weng JR, Chiu CF, Hu JL, Feng CH, Huang CY, Bai LY, Sheu JH. A Sterol from Soft Coral Induces Apoptosis and Autophagy in MCF-7 Breast Cancer Cells. Mar Drugs. 2018;16 doi: 10.3390/md16070238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Chen IH, Chang FR, Wu YC, Kung PH, Wu CC. 3,4-Methylenedioxy-β-nitrostyrene inhibits adhesion and migration of human triple-negative breast cancer cells by suppressing β1 integrin function and surface protein disulfide isomerase. Biochimie. 2015;110:81–92. doi: 10.1016/j.biochi.2015.01.006. [DOI] [PubMed] [Google Scholar]
- 101.Yang H, Dai G, Wang S, Zhao Y, Wang X, Zhao X, Zhang H, Wei L, Zhang L, Guo S, Song W, Guo L, Fang C. Inhibition of the proliferation, migration, and invasion of human breast cancer cells by leucine aminopeptidase 3 inhibitors derived from natural marine products. Anticancer Drugs. 2020;31:60–66. [Google Scholar]
- 102.Moon DO, Choi YH, Moon SK, Kim WJ, Kim GY. Gossypol decreases tumor necrosis factor-α-induced intercellular adhesion molecule-1 expression via suppression of NF-κB activity. Food Chem Toxicol. 2011;49:999–1005. doi: 10.1016/j.fct.2011.01.006. [DOI] [PubMed] [Google Scholar]
- 103.Rezakhani L, Rashidi Z, Mirzapur P, Khazaei M. Antiproliferatory Effects of Crab Shell Extract on Breast Cancer Cell Line (MCF7) J Breast Cancer. 2014;17:219–225. doi: 10.4048/jbc.2014.17.3.219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Resmi R, Yoonus J, Beena B. Anticancer and antibacterial activity of chitosan extracted from shrimp shell waste. Materials Today: Proceedings 2020. [Google Scholar]
- 105.Mohamed A, Bakaar A, Ghaly CAF. In vitro assessment of anticancer activity of shrimp derived chitosan and related apoptotic profile alteration. 2018. [Google Scholar]
- 106.Sowmya PR, Arathi BP, Vijay K, Baskaran V, Lakshminarayana R. Astaxanthin from shrimp efficiently modulates oxidative stress and allied cell death progression in MCF-7 cells treated synergistically with β-carotene and lutein from greens. Food Chem Toxicol. 2017;106:58–69. doi: 10.1016/j.fct.2017.05.024. [DOI] [PubMed] [Google Scholar]
- 107.Palhares LCGF, Barbosa JS, Scortecci KC, Rocha HAO, Brito AS, Chavante SF. In vitro antitumor and anti-angiogenic activities of a shrimp chondroitin sulfate. Int J Biol Macromol. 2020;162:1153–1165. doi: 10.1016/j.ijbiomac.2020.06.100. [DOI] [PubMed] [Google Scholar]
- 108.Cui Y, Lu Z, Bai L, Shi Z, Zhao WE, Zhao B. beta-Carotene induces apoptosis and up-regulates peroxisome proliferator-activated receptor gamma expression and reactive oxygen species production in MCF-7 cancer cells. Eur J Cancer. 2007;43:2590–2601. doi: 10.1016/j.ejca.2007.08.015. [DOI] [PubMed] [Google Scholar]
- 109.Zheng JS, Hu XJ, Zhao YM, Yang J, Li D. Intake of fish and marine n-3 polyunsaturated fatty acids and risk of breast cancer: meta-analysis of data from 21 independent prospective cohort studies. BMJ. 2013;346:f3706. doi: 10.1136/bmj.f3706. [DOI] [PubMed] [Google Scholar]
- 110.Engeset D, Alsaker E, Lund E, Welch A, Khaw KT, Clavel-Chapelon F, Thiébaut A, Chajès V, Key TJ, Allen NE, Amiano P, Dorronsoro M, Tjønneland A, Stripp C, Peeters PH, van Gils CH, Chirlaque MD, Nagel G, Linseisen J, Ocké MC, Bueno-de-Mesquita HB, Sacerdote C, Tumino R, Ardanaz E, Sánchez MJ, Panico S, Palli D, Trichopoulou A, Kalapothaki V, Benetou V, Quirós JR, Agudo A, Overvad K, Bjerregaard L, Wirfält E, Schulz M, Boeing H, Slimani N, Riboli E. Fish consumption and breast cancer risk. The European Prospective Investigation into Cancer and Nutrition (EPIC) Int J Cancer. 2006;119:175–182. doi: 10.1002/ijc.21819. [DOI] [PubMed] [Google Scholar]
- 111.Patterson RE, Flatt SW, Newman VA, Natarajan L, Rock CL, Thomson CA, Caan BJ, Parker BA, Pierce JP. Marine fatty acid intake is associated with breast cancer prognosis. J Nutr. 2011;141:201–206. doi: 10.3945/jn.110.128777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Bosseboeuf A, Baron A, Duval E, Gautier A, Sourdaine P, Auvray P. K092A and K092B, Two Peptides Isolated from the Dogfish (Scyliorhinus canicula L.), with Potential Antineoplastic Activity Against Human Prostate and Breast Cancer Cells. Mar Drugs. 2019;17 doi: 10.3390/md17120672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Atmaca H, Bozkurt E, Uzunoglu S, Uslu R, Karaca B. A diverse induction of apoptosis by trabectedin in MCF-7 (HER2-/ER+) and MDA-MB-453 (HER2+/ER-) breast cancer cells. Toxicol Lett. 2013;221:128–136. doi: 10.1016/j.toxlet.2013.06.213. [DOI] [PubMed] [Google Scholar]
- 114.Teixidó C, Arguelaguet E, Pons B, Aracil M, Jimeno J, Somoza R, Marés R, Ramón Y Cajal S, Hernández-Losa J. ErbB3 expression predicts sensitivity to elisidepsin treatment: in vitro synergism with cisplatin, paclitaxel and gemcitabine in lung, breast and colon cancer cell lines. Int J Oncol. 2012;41:317–324. doi: 10.3892/ijo.2012.1425. [DOI] [PubMed] [Google Scholar]
- 115.Abraham I, Jain S, Wu CP, Khanfar MA, Kuang Y, Dai CL, Shi Z, Chen X, Fu L, Ambudkar SV, El Sayed K, Chen ZS. Marine sponge-derived sipholane triterpenoids reverse P-glycoprotein (ABCB1)-mediated multidrug resistance in cancer cells. Biochem Pharmacol. 2010;80:1497–1506. doi: 10.1016/j.bcp.2010.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Wang Z, Wong IL, Li FX, Yang C, Liu Z, Jiang T, Jiang TF, Chow LM, Wan SB. Optimization of permethyl ningalin B analogs as P-glycoprotein inhibitors. Bioorg Med Chem. 2015;23:5566–5573. doi: 10.1016/j.bmc.2015.07.027. [DOI] [PubMed] [Google Scholar]
- 117.D'Incalci M, Jimeno J. Preclinical and clinical results with the natural marine product ET-743. Expert Opin Investig Drugs. 2003;12:1843–1853. [PubMed] [Google Scholar]
- 118.Liu HZ, Xiao W, Gu YP, Tao YX, Zhang DY, Du H, Shang JH. Polysaccharide from Sepia esculenta ink and cisplatin inhibit synergistically proliferation and metastasis of triple-negative breast cancer MDA-MB-231 cells. Iran J Basic Med Sci. 2016;19:1292–1298. doi: 10.22038/ijbms.2016.7913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Dey G, Bharti R, Das AK, Sen R, Mandal M. Resensitization of Akt Induced Docetaxel Resistance in Breast Cancer by 'Iturin A' a Lipopeptide Molecule from Marine Bacteria Bacillus megaterium. Sci Rep. 2017;7:17324. doi: 10.1038/s41598-017-17652-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Tun JO, Salvador-Reyes LA, Velarde MC, Saito N, Suwanborirux K, Concepcion GP. Synergistic Cytotoxicity of Renieramycin M and Doxorubicin in MCF-7 Breast Cancer Cells. Mar Drugs. 2019;17 doi: 10.3390/md17090536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Zhang T, Liu H, Li Y, Li C, Wan G, Chen B, Wang Y. A pH-sensitive nanotherapeutic system based on a marine sulfated polysaccharide for the treatment of metastatic breast cancer through combining chemotherapy and COX-2 inhibition. Acta Biomater. 2019;99:412–425. doi: 10.1016/j.actbio.2019.09.001. [DOI] [PubMed] [Google Scholar]
- 122.Kanno SI, Yomogida S, Tomizawa A, Yamazaki H, Ukai K, Mangindaan RE, Namikoshi M, Ishikawa M. Combined effect of papuamine and doxorubicin in human breast cancer MCF-7 cells. Oncol Lett. 2014;8:547–550. doi: 10.3892/ol.2014.2218. [DOI] [PMC free article] [PubMed] [Google Scholar]