

Abstract
Growing evidence suggests that ABO blood group may play a role in the immunopathogenesis of SARS‐CoV‐2 infection, with group O individuals less likely to test positive and group A conferring a higher susceptibility to infection and propensity to severe disease. The level of evidence supporting an association between ABO type and SARS‐CoV‐2/COVID‐19 ranges from small observational studies, to genome‐wide‐association‐analyses and country‐level meta‐regression analyses. ABO blood group antigens are oligosaccharides expressed on red cells and other tissues (notably endothelium). There are several hypotheses to explain the differences in SARS‐CoV‐2 infection by ABO type. For example, anti‐A and/or anti‐B antibodies (e.g. present in group O individuals) could bind to corresponding antigens on the viral envelope and contribute to viral neutralization, thereby preventing target cell infection. The SARS‐CoV‐2 virus and SARS‐CoV spike (S) proteins may be bound by anti‐A isoagglutinins (e.g. present in group O and group B individuals), which may block interactions between virus and angiotensin‐converting‐enzyme‐2‐receptor, thereby preventing entry into lung epithelial cells. ABO type‐associated variations in angiotensin‐converting enzyme‐1 activity and levels of von Willebrand factor (VWF) and factor VIII could also influence adverse outcomes, notably in group A individuals who express high VWF levels. In conclusion, group O may be associated with a lower risk of SARS‐CoV‐2 infection and group A may be associated with a higher risk of SARS‐CoV‐2 infection along with severe disease. However, prospective and mechanistic studies are needed to verify several of the proposed associations. Based on the strength of available studies, there are insufficient data for guiding policy in this regard.
Keywords: COVID‐19, SARS‐CoV‐2, ABO blood groups, disease susceptibility, disease severity
Introduction
The COVID‐19 pandemic spurred a crisis that is unprecedented in modern times [1]. The disease course varies substantially among individuals, from mild or even subclinical infection to severe disease [2]. Indeed, more than 1 million COVID‐19‐related deaths have been reported globally. There is interest in potential risk factors that affect susceptibility to infection and disease progression. Multiple medical (e.g. diabetes, hypertension) and sociodemographic (e.g. sex, age and race/ethnicity) risk factors for severe outcomes were already established [2]. Growing evidence suggests that the ABO blood group may also play a role in the immunopathogenesis of SARS‐CoV‐2 infection, with group O being protective and group A conferring risks of higher disease susceptibility and severity [3, 4, 5, 6, 7].
An international group of experts in transfusion medicine and haematology were assembled by the International Society of Blood Transfusion (ISBT) to review and summarize the literature with a view of offering recommendations pertaining to ABO type and COVID‐19. To this end, we provide an overview of the ABO blood group system, ABO population frequencies and distributions, its role as a histo‐blood group antigen, not just a blood group antigen, and the known associations between ABO type and various infectious and non‐infectious diseases. Finally, we present a scoping review of the literature on the associations of ABO type with COVID‐19 and propose mechanistic pathways that could potentially explain these observations.
Search strategy and selection criteria
A combination of searching using Medline’s controlled vocabulary/indexing, [Mesh]/[Supplemental Concept], and keyword searching titles and abstracts [tiab] features was used to search articles on ABO blood group and infectious diseases and ABO blood group and COVID‐19 or SARS‐CoV‐2 to (a) pull articles either specifically indexed about our topic/concept as well as (b) pull articles wherever our topic/concept of interest was mentioned. Furthermore, we also searched all the Web of Science (WOS) databases using the same keyword searching as for PubMed/Medline. WOS in addition supports the NEAR operator (searches for words that appear near one another in the text without requiring an exact phrase match). The search was confined to English‐language articles that were published prior to 10 November 2020. Pre‐print articles were also included on a case‐by‐case basis.
ABO blood group overview
Of the 39 blood group systems and 350 antigens recognized by the ISBT [8], the ABO system is clinically the most important [9]. The A and B antigens are inherited co‐dominantly over O [10]. The ABH antigens (H antigen defines the O blood type) are oligosaccharides exposed on RBCs and other cells; they are also found in body secretions. The A and B antigens are determined by allelic genes encoding glycosyltransferases that transfer monosaccharides to the non‐reducing ends of specific glycans on glycoproteins and glycolipids. For A and B, this monosaccharide is N‐acetyl‐D‐galactosamine and D‐galactose, respectively. In group O individuals, the corresponding A and B glycosyltransferases are either not present or have been inactivated by one of various polymorphisms, such that the non‐reducing ends of the corresponding glycans express the H antigen.
Antibodies in this system (i.e. anti‐A and anti‐B) develop in the first few months of life; they are typically ‘naturally occurring’ antibodies produced after contact with non‐self A and/or B antigens, often found in food and micro‐organisms, notably the gut microbiota [11]. Anti‐A and anti‐B, typically of the IgM isotype, circulate in almost all healthy individuals who lack the corresponding antigen; IgG anti‐A,B is often found in group O individuals [12, 13, 14]. Transfusion of ABO‐incompatible RBCs can provoke acute haemolytic transfusion reactions because the corresponding IgM antibodies bind complement efficiently, causing intravascular hemolysis of the transfused RBCs and activation of coagulation. IgG antibodies can also cause severe intravascular hemolysis in this setting, because of the very high density of ABH antigens on RBCs, leading to close proximity of anti‐A and/or anti‐B IgG molecules on the RBC surface with subsequent complement activation [15, 16]. If untreated, this medical emergency can induce acute renal failure, disseminated intravascular coagulation and death. Therefore, always transfusing ABO‐compatible RBCs is a central focus of modern blood banking processes and procedures.
Population frequency of ABO blood groups
ABO blood group frequencies vary among human populations, suggesting that migration and the selective advantage of particular blood groups, perhaps relating to exposure to specific pathogens, might have contributed to these variations (Table 1). For example, group O is the most common globally, followed by A, then B, and then AB. Group O may have originated in Africa before early human migration, because it may have provided a selective advantage against malaria [17]. In contrast, group O individuals are at higher risk of developing severe cholera, perhaps explaining the lower prevalence of group O and higher prevalence of group B, in the Ganges Delta region of Bangladesh [18].
Table 1.
Table of ABO Geographic Distributions for the native and contemporary populations
| Region | Native population | Current population | |||||
|---|---|---|---|---|---|---|---|
| Type A % | Type O % | Type B % | Type A % | Type O % | Type B % | ||
| North America | Canada | Up to 40 | 80–100 | 0–5 | 40+ | 40+ | 9 |
| United States | 0–15* | 80–100 | 0–5 | 40+ | 40+ | ˜10 | |
| Central and South America | Absent | 90–100 | 0–5 | 10–30 | 50–80 | ˜10 | |
| Greenland | Up to 40+ | ||||||
| Australia | Up to 40+ † | 60–80 (North) | 0–5 | 38 | 49 | ˜10 | |
| Africa | 15–20 | 60–80 | 10–20 | ˜20–25 ¥ | Up to 60 | West > 20 | |
| Middle East | 15–20 | 60–80 | 5–15 | ˜25 | >40 | >20 | |
| Europe | Scandinavia | 25–40 ‡ | 50–70 | 0–10 | 40+ | ˜40 | 10 |
| Western Europe | 25–30 | 60–70 | 5–10 | 30–40 | 30–40 | ˜10 | |
| Eastern Europe | 25–30 | 50–60 | 10–20 | 30–40 | 30–40 | ˜10 | |
| Russia | 15–20 | 50–60 | 15–30 | ˜35 | ˜35 | ˜10 | |
| Asia | China | 20–25 | 60–70 | 15–25 | ˜30 | ˜50 | ˜20 |
| Japan | 15–25 | 50–70 | 10–15 | 40 | 30 | 20 | |
| Pacific | 15–20 | 60–70 | 15–25 | 25–30 | >40 | ˜30 | |
| India | 15–20 | 56–60 | Up to 30 | 22 | 29 | 38 | |
Blackfoot of Montana: 30–35%.
Aboriginal Australians: 40–53%.
Lapp: 50–90%.
Cameroon 38, Uganda 39, South Africa 32%.
Indigenous populations show widely divergent ABO blood group distributions. For example, those in Australia and North and South America almost completely lack group B, whereas those in Asia show the highest rates of group B. In addition, group A is almost non‐existent in indigenous populations of South and Central America, but more prevalent (>30%) in Canada, Scandinavia and Central Europe, perhaps due to selective pressure provided by smallpox [18]. In contrast, ABO distributions of current populations in these same countries demonstrate the impact of migration; for example, although group A is virtually absent in indigenous populations in Central and South America, its current overall population frequency is as high as 30% [18].
ABO is not just a blood group antigen
Each RBC expresses ˜2 million copies of its genetically encoded ABH blood group antigens on its surface, although the density varies by antigen type. Other blood cells (e.g. platelets and lymphocytes) also adsorb ABH‐expressing glycosphingolipids from plasma, where they circulate attached to lipoproteins. In addition, ABH antigens are synthesized and expressed on endothelial cells and certain epithelial cells. Thus, although some blood group antigens are only on RBCs, ABH antigens are in various cells, body fluids and secretions. Therefore, they are more correctly denoted as ‘histo‐blood group antigens’ (HBGA), not solely as blood group antigens [11, 19, 20, 21, 22, 23].
In addition to serving as antigenic barriers during transfusion, transplantation and pregnancy, ABH oligosaccharides physiologically influence hemostasis and, therefore, confer disease risks in this setting. For example, A and B glycosyltransferases modify H‐active glycans on von Willebrand factor (VWF) [24]. Interestingly, VWF in group O individuals has a shorter half‐life, accompanied by 25–30% reduced VWF and Factor VIII levels, as compared to group A or B individuals. However, independent of the ABO blood group, glycosyltransferase activity was also decreased in patients with venous thromboembolism, as compared to healthy controls [25]. In addition, higher VWF and factor VIII levels are associated with increased risks for coronary heart disease, arterial thrombosis and venous thrombosis [26, 27]. Therefore, perhaps not surprisingly, recent genome‐wide association studies (GWAS) demonstrated that ABO locus variants correlate with increased plasma lipid and inflammatory marker levels [25, 28].
ABO expression may also not be stable over time, with lower levels in healthy children <2 years old [29] and changes in various diseases (e.g. necrotizing infection, thalassemia, malignancy) [20, 30]. In addition, as compared to group O, group A individuals have a higher prevalence of gastric cancer, and group A, AB or B individuals have a higher prevalence of pancreatic cancer; possible mechanisms include ABO blood group influences on regulating proinflammatory [31] and adhesion molecules [32, 33], and the role of VWF in angiogenesis and apoptosis [22, 25, 34, 35].
Associations between blood groups and infectious diseases
HBGAs are implicated in the pathogenesis of multiple infections. In particular, the ABO blood type has been associated with, for example, tuberculosis, malaria, cholera, norovirus, retrovirus, Chikungunya virus, Helicobacter pylori (H. pylori) and Escherichia coli [36, 37]. The underlying mechanisms range from simple (e.g. receptor‐ligand interactions) to complex and may be limited to a specific pathogenic product, strain or disease state. For ABO, possible explanations include ABH antigens as receptors for pathogens, natural antibodies and lectins as inhibitors, and molecular mimicry by blood group antigens between pathogen and host.
One specific example involves the P antigen in the Globoside blood group. This antigen is necessary, but not sufficient [38], for parvovirus B19 entry into RBCs, requiring a co‐receptor for infection [39, 40]. The distribution of P antigen, including relatively high expression by RBCs and their precursors, is consistent with parvovirus B19 clinical syndromes, including aplastic anaemia [41]. Furthermore, individuals lacking the P antigen (i.e. the p phenotype) are resistant to this infection [42].
For other infections, HBGAs can be receptors for toxins, virulence factors or other pathogenic products without directly binding the implicated pathogen itself. In addition, HBGAs in secretions, body fluids or non‐erythroid tissues can contribute to pathogenesis. For example, adhesion and colonization by H. pylori, the aetiologic agent of peptic ulcer disease and some forms of gastric cancer, are facilitated by Leb/H oligosaccharides on gastric epithelial mucins [43, 44] with certain strains having higher affinity for Leb/H [45]; in addition, group O individuals, by expressing more H antigen, are more likely to be infected [46].
In summary, the relationship between HBGAs and a specific infection should ideally satisfy Koch’s postulates. Nonetheless, this is often difficult to document fully in human studies. Typically, one can only show that individuals expressing a specific blood group or HBGA are more susceptible to infection, whereas individuals without it are completely resistant or, at least, protected from severe disease. Multiple examples demonstrate that HBGAs can interact with pathogens at initial infection, or alter disease progression/severity, or affect clinical presentation (Table 2).
Table 2.
Potential mechanisms for relationships between histo‐blood group antigen (HBGA) and infection
| Action as a receptor or co‐receptor for a given pathogen |
| Functions as a receptor for a virulence factor, toxin, or other pathogenic product |
| Interaction with a pathogen that is limited to a specific strain, specific organ system or disease state |
| Modification of a key target cell surface glycoprotein or glycolipid, thereby affecting important cellular functions (e.g. endocytosis, phagocytosis, signal transduction) in response to infection |
ABO blood group and susceptibility to SARS‐CoV‐2
During the severe acute respiratory syndrome coronavirus (SARS‐CoV‐1) epidemic, several observations suggested that ABO type may contribute to disease, with less susceptibility in group O individuals [47]. This was also observed for SARS‐CoV‐2 (Table 3). Most studies identified a higher proportion of group A, and a lower proportion of group O, among COVID‐19 patients, as compared to healthy controls [5, 7, 12, 48, 49, 50, 51, 52]. These studies included patients with SARS‐CoV‐2 pneumonia ranging in severity from mild to critically ill requiring mechanical ventilation or intensive care unit admission [48, 51]. For example, in one study, the proportion of group A infected patients was significantly higher than in healthy controls (38% vs. 32·2 %, P < 0·001), whereas group O was significantly lower (25·7% vs. 33·8%, P < 0·001); however, group A patients had higher frequencies of underlying comorbidities [7]. Another retrospective study had similar findings, but did not describe comorbidities [52]. Another study described a higher rate of infection in group AB patients and a lower rate in group O patients [50]. In contrast, an additional study did not find any correlation between group A status and COVID‐19; nonetheless, group O individuals had a lower risk of COVID‐19 and group B and AB individuals had a higher risk [6]. One potential reason for these varying results is that many such studies did not account for various confounders (e.g. age), including comorbidities. Another potential confounder for some of the studies could be the use of randomly selected volunteer blood donors as controls, because of the risk of group O epidemiological predominance due to blood collectors selectively recruiting group O donors. Importantly, volunteer blood donors are not necessarily representative of general populations; although convenient, their use as a control group is not optimal [53, 54].
Table 3.
Summary of reported studies assessing the association between blood groups and SARS‐CoV‐2 infection
| Ref. Author (country) | COVID‐19 study population | Controls(if applicable) |
% group A patients (vs. control) P value (when applicable) |
% group O patients (vs. control) P value (when applicable) |
Blood group susceptibility to SARS‐CoV‐2 infection | Association with clinical outcomes and risk of death |
|---|---|---|---|---|---|---|
|
Zhao J et al. [5] China |
1775 patients | 3694 normal individuals |
37·75 (32·16) P < 0·001 |
25·80 (33·84) P < 0·001 |
Yes, group A | Group A associated with higher risk of mortality than non group A |
|
Li J et al. [7] China |
2153 patients with COVID‐19 pneumonia | 3694 healthy controls |
38·0 (32·2) P < 0·001 |
25·7 (33·8) P < 0·001 |
Yes, group A |
Group A patients at higher risk of hospitalization following SARS‐CoV‐2 infection. Association with risk of mortality not assessed. |
|
Zeng X. et al. [51] China |
137 patients with mild pneumonia 97 patients with severe pneumonia |
Nil, Chinese population data used for comparison |
35·76 (28·39) 39·22 (28·39) |
32·45 (33·20) 26·47(33·20) |
Yes, group A |
Blood group A more susceptible to SARS‐CoV‐2. Blood groups not relevant to acute respiratory distress syndrome, acute kidney injury and mortality. |
|
Zietz M & Tatonetti [56] USA |
Observational data on 14,112 individuals tested for SARS‐CoV‐2 | None | 32·7 (32·7) | 46·9 (48·2) | Yes, group B and Rh(D) |
Risk of intubation decreased among group A and increased among groups AB and B. Risk of morality increased for group AB and decreased for groups A and B. Rh‐negative blood type protective for mortality. |
|
GÖKER H, et al . [49] Turkey |
186 patients | 1881 healthy controls |
57 (38) P < 0·001 |
24·8 (37·2) P = 0·001 |
Yes, group A | No significant effect of ABO and RhD on clinical outcomes including intubation, ICU stay and mortality |
|
Wu et al. [52] China |
187 patients | 1991 non‐COVID‐19 hospitalized patients |
36·9 (27·47) P = 0·006 |
21·92(30·19) P = 0·018 |
Yes, group A | Group A influenced clinical outcomes but no association with mortality |
|
Leaf RK et al. [48] USA |
561 critically ill patients. |
Nil, local population data used for comparison | 45·1 (39·8) | 37·8 (45·2) | Yes, group A | No association with any ABO phenotype and mortality |
|
Latz CA et al. [6] USA |
1289 patients | Nil | 34·2 (NA) | 45·5 (NA) | Yes, positive correlation with group B , AB & Rh(D)Negative with group O | No association with risk of intubation, peak of inflammatory markers and death |
|
Gerard C et al. [12] |
1175 patients | 3694 controls |
37·7 (32·2) P < 0·001 |
49·4 (57·6) |
Yes, presence of anti‐A antibodies in serum and more specifically IgG anti‐Aassociated with higher susceptibility to SARS‐CoV‐2 infection. |
Mortality risk not assessed |
|
Abdollahi A et al. [50] Iran |
397 patients | 500 normal controls |
40·3 (36) P = 0·19 |
28 (38) P = 0·002 |
Yes, group AB with higher susceptibility than other groups. |
No association of ABO or RHD phenotype with severity of disease. No association of ABO or RHD with mortality assessed. |
|
Hoiland et al. [4] Canada |
125 critically ill patients admitted to ICU | Nil, Comparison of blood group distributions between blood donor data was performed. |
37 (35) p‐0·60 No difference from blood donors |
Group O 43% (n = 41) No difference from blood donors |
Yes, group A and AB |
Higher proportion of COVID‐19 patients with blood group A or AB required mechanical ventilation, and continous renal replacement therapy and had longer ICU stay compared with patients with blood group O or B. |
|
Barnkob et al. [3] Denmark |
7422 COVID positive patients among 473 654 individuals tested |
466 232 COVID‐negative individuals |
More A (P <0·001), B (P = 0·011), and AB (P = 0·091) individuals were COVID positive. | 38·41% (95% CI, 37·30–39·50) group O compared with 41·70% (41·60–41·80) in controls | Yes, Decreased infection risk in group O |
ABO blood group as a risk factor for SARS‐CoV‐2 infection but not for hospitalization or death from COVID‐19. |
It has also been hypothesized that anti‐A and anti‐B antibodies could interfere with virus–cell interactions. In a secondary analysis of data from ˜1900 patients with COVID‐19, subjects with circulating anti‐A were significantly less represented in the disease group as compared to those lacking anti‐A. In addition, anti‐A in group O individuals was more protective than anti‐A in group B individuals; this may relate to the increased presence of IgG anti‐A,B in group O plasma [13].
One study attempted a meta‐regression analysis of 101 nations using their known blood group distributions, including ˜9‐million COVID‐19 cases and ˜450 000 deaths in a total population of ˜7 billion. Although there was no association of group A or B with overall mortality, group O significantly correlated with lower mortality (p = 0·02). The authors proposed that COVID‐19 mortality was lower in nations with higher group O prevalence because overall population ABO blood group prevalence was analysed as the control [55].
Studies have also examined the relationship between the Rhesus blood group (e.g. Rh(D) type) and COVID‐19. One study suggested that Rh(D)‐positive individuals were more likely to test positive for SARS‐CoV‐2 [6]. Another study found significant associations between Rh(D) blood group status, group B, and SARS‐CoV‐2 [56].
In summary, these mixed findings may be ascribed to the different populations, the controls that were used for comparison, the geographical locations and the confounders that were considered. The latter include age, comorbidities and using volunteer blood donors as controls.
ABO and COVID‐19 disease severity
The effect of ABO type on COVID‐19 disease severity also warrants analysis. As such, in one study, group A patients had a higher risk of hospitalization for SARS‐CoV‐2 infection, whereas group O patients were at lower risk [7]. However, the group A patients had more comorbid risk factors for severe disease, which were not adjusted for using a multivariate analysis. Another group performed a GWAS of COVID‐19 patients with respiratory failure [57], detecting a statistically significant cross‐replicating association at 9q34. The 9q34 signal was located at the ABO blood group locus and a blood type‐specific analysis showed a higher risk of severe COVID‐19 with respiratory failure for group A individuals and a protective effect for group O.
As another example, in a nested prospective observational study of critically ill patients with COVID‐19 in Canada, using a multivariable adjustment of various risk factors, patients with blood group A or AB had an increased risk of requiring mechanical ventilation, continuous renal replacement therapy and prolonged intensive care unit admission, as compared to group O or B patients [4]. Another recent retrospective cohort analysis including nearly half a million Danish individuals tested for SARS‐CoV‐2, also showed reduced prevalence of SARS‐CoV‐2 infection in blood group O individuals. This study identified ABO blood group as a risk factor for SARS‐CoV‐2 infection but not for hospitalization or death from COVID‐19 [3].
Taken together, these studies suggest that the risk of infection with SARS‐CoV‐2 and the risk of severe COVID‐19 disease may be lower in group O individuals than non‐group O individuals. Nonetheless, these results are not definitive and further studies are warranted.
Mechanisms for associations between ABO blood group and COVID‐19
Several pathophysiological mechanisms were proposed to explain the association between ABO type and SARS‐CoV‐2 infection (Fig. 1, Table 4). Anti‐A and/or anti‐B antibodies might bind to A and/or B antigens expressed on the viral envelope, thereby preventing infection of target cells; that is, these naturally occurring antibodies could function as viral neutralizing antibodies. If true, this would help explain differences in initial susceptibility for SARS‐CoV‐2 infection. For example, an anti‐A viral neutralizing antibody in a potentially susceptible group O host would bind the A antigen on virus produced by, and inhaled from, an infected group A (or group AB) host [58]. Why this mechanism would be relevant to disease severity per se is less obvious, because subsequent rounds of viral proliferation in a group O host would produce virus expressing the H antigen on its envelope. However, assuming that disease severity relates to the size of the infecting inoculum and yielding the subsequent viral load, a neutralizing isoagglutinin (e.g. anti‐A) could attenuate infection, if not preventing infection altogether. Finally, the entry barrier for this virus is the epithelium of the respiratory tract and, possibly, the digestive tract. Thus, to prevent infection, circulating antibodies may need to reach these cell surfaces; although, presumably, the most effective antibodies for this purpose are of the secretory IgA isotype, to date, no data are available about the IgA isotype for either anti‐A and/or anti‐B in this regard.
Fig. 1.
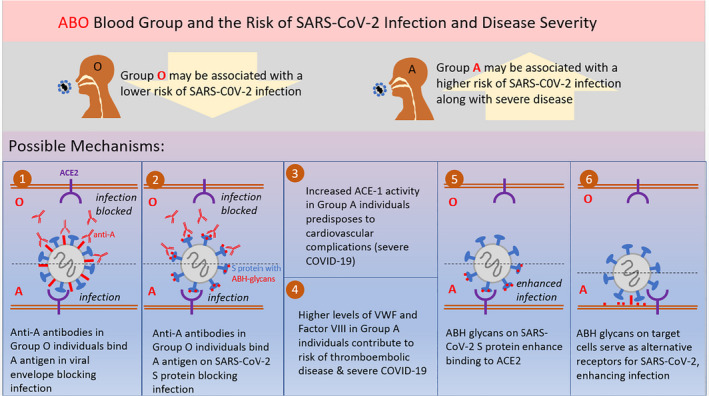
Graphical summary of proposed mechanisms for association Between ABO blood groups and SARS‐CoV‐2 infection.
Table 4.
Proposed mechanisms, theoretical pathways and suggested experiments for studying the association Between ABO blood groups and SARS‐CoV‐2 infection
| Proposed mechanisms for association between ABO blood type and SARS‐CoV‐2 infection |
| Anti‐A and/or anti‐B antibodies serve as viral neutralizing antibodies by binding to A and/or B antigens expressed on the viral envelope, thereby preventing infection of target cells |
| The SARS‐CoV‐2 S protein is bound by human anti‐A antibodies, which may block the interaction between the virus and ACE2R, thereby preventing entry into the lung epithelium |
| An increase in ACE‐1 activity in group A individuals predisposes to cardiovascular complications, accounting for severe COVID‐19 |
| Variation of VWF and Factor VIII levels by ABO type with higher levels in group A individuals contributing to risk of thromboembolic disease and severe COVID‐19 |
| ABH glycans, if present on SARS‐CoV‐2 S protein, may modify the affinity of SARS‐CoV‐2 for ACE2R, its cellular receptor. |
| ABH glycans on target cells could serve as alternative, lower‐affinity receptors for SARS‐CoV‐2 S protein or bind other viral envelope structures. |
| Suggested experiments to confirm the proposed associations between ABO blood groups and SARS‐CoV‐2 infection |
| In vitro production of recombinant SARS‐CoV‐2 S protein in cell lines that can synthesize ABH glycans to determine whether these are, indeed, on the recombinant protein. |
| Culturing SARS‐CoV‐2 in cell lines capable of synthesizing ABH glycans, isolating the virus and determining whether anti‐A and anti‐B prevent infection of cell lines that do not express the corresponding ABH antigen. These experiments would also allow testing whether IgM and IgG anti‐A (or anti‐B) antibodies were equally effective. |
| Producing recombinant SARS‐CoV‐2 S protein in identical host cell lines (e.g. by transfecting in the relevant glycosyltransferases) and then quantifying the affinity of the purified proteins for their receptor. This can determine if ABH glycans are present on SARS‐CoV‐2 S protein and if they modify the affinity of SARS‐CoV‐2 for ACE2R. |
| Produce SARS‐CoV‐2 in vitro in ABH‐expressing cell lines; and quantify the infectivity of a given target cell line with purified virus. |
| Quantify IgA anti‐A and/or anti‐B and correlate with risk of susceptibility to infection by SARS‐CoV‐2 and severity COVID‐19 illness. |
Glycan structures at various N‐glycosylation sites of the SARS‐CoV S protein were previously described [59, 60, 61]. In addition, N‐glycans of recombinant SARS‐CoV‐2 S protein were recently characterized [62]; although ABH antigen structures were not described, this may be due to the cell line used to produce the recombinant protein. Interestingly, the receptor‐binding domains of the SARS‐CoV‐2 and SARS‐CoV S proteins are structurally nearly identical [63]; in addition, glycosylation yields S trimers in which the receptor‐binding domains are covered by N‐glycans. Thus, it is conceivable that SARS‐CoV‐2 S protein could be specifically bound by human anti‐A antibodies, which could then block the interaction between the virus and the angiotensin‐converting enzyme 2 receptor (ACE2R), thereby preventing entry into the lung epithelium. Relevant to this hypothesis, monoclonal or naturally occurring anti‐A antibodies dose‐dependently inhibited interaction between SARS‐CoV S protein and ACE2R, in a model where the A antigen was associated with S protein [64]. Thus, if this is also true for SARS‐CoV‐2, it would support the hypothesis that anti‐A blocks viral attachment and/or entry [65]. Although not definitive, currently available data could support the above mechanism. There is also emerging evidence that receptor‐binding domain (RBD) of SARS‐CoV‐2 may share sequence similarity to an ancient lectin family known to bind blood group antigens. SARS‐CoV‐2 RBD binds the blood group A expressed on respiratory epithelial cells, which could explain the linkage between blood group A and SARS‐CoV‐2 [66].
Another potential mechanism for explaining an association between group A and severe COVID‐19 is an increase in angiotensin‐converting enzyme 1 (ACE‐1) activity, with a predisposition to cardiovascular complications. Severe outcomes could also be explained by higher levels of VWF and factor VIII in group A individuals. Furthermore, VWF is an acute phase reactant with infection inducing even higher levels in group A individuals. Given that anti‐A and anti‐B antibody titres are highly variable among individuals [67, 68], the potential neutralizing effect of such antibodies is also expected to be highly variable [69], possibly obscuring the ‘signal’ in large population studies. This variability may be further compounded by the significantly higher binding affinity of SARS‐CoV‐2 S protein for ACE2R, as compared to SARS‐CoV [70].
Studies to confirm this hypothesis in vitro would involve producing recombinant SARS‐CoV‐2 S protein in cell lines that can synthesize terminal ABH oligosaccharides to determine whether these are, indeed, on the recombinant protein. In addition, it is important to culture SARS‐CoV‐2 in cell lines capable of synthesizing ABH oligosaccharides, isolate the virus and determine whether anti‐A and anti‐B prevent infection of cell lines that do not express the corresponding ABH antigen. If successful, these experiments would also allow testing whether IgM, IgA or IgG anti‐A (or anti‐B) antibodies were equally effective [71]. To our knowledge, there are no published studies to date that address this.
Other potential mechanisms may explain the epidemiological results. For example, if ABH oligosaccharides are on SARS‐CoV‐2 S protein, they may modify the affinity of SARS‐CoV‐2 for ACE2R, its cellular receptor. This could be evaluated formally by producing recombinant S protein in otherwise identical host cell lines (e.g. by transfecting in the relevant glycosyltransferases) and then quantifying the affinity of the purified proteins for their receptor. Analogously, if the virus could be produced in vitro in these ABH‐expressing cell lines and purified, the infectivity of a given target cell line could then be quantified. Given the published human population data, one might expect ‘group O virions’ to be less infectious in these experiments, thereby correlating with decreased COVID‐19 disease severity.
A different, but not mutually exclusive, mechanism may involve ACE2R, which is also a glycoprotein and may express ABH glycans. It is possible that these glycans affect SARS‐CoV‐2 viral binding to ACE2R, the number of ACE2R proteins on a given cell surface, and/or the efficacy of internalization of the virus: receptor complex. In this case, ACE2R expressing H‐antigen glycans may not be as effective at binding and internalizing SARS‐CoV‐2 produced by any source, irrespective of ABO type. This could also underlie COVID‐19 disease severity.
It is also possible that the ABH glycans themselves could serve as (alternative) lower‐affinity receptors for SARS‐CoV‐2 S protein or bind other viral envelope structures. Although current evidence suggests that this is unlikely, if it were relevant, then ABH glycan levels on cell surfaces, in plasma, and in secretions would be important and could affect initial infection and disease severity. For this purpose, determining the ‘secretor phenotype’ and Lewis blood group types would be helpful [37, 72]. Moreover, secretor status and Lewis antigen frequencies can affect host immunity [37, 72].
Because COVID‐19 severity relates significantly to cardiovascular [73], thromboembolic [74, 75] and inflammatory complications, the patient’s ABO type may be a surrogate for these effects and have nothing to do with blood type per se, the presence of naturally occurring anti‐A and/or anti‐B or viral‐target cell interactions. For example, as described above, ABO type influences circulating VWF and factor VIII levels, which influence cardiovascular risk and hemostatic function, even in the absence of infection [76].
Although an association between ABO blood group and the risk of susceptibility to COVID‐19 disease or disease severity is compelling, the practical significance is uncertain. COVID‐19 convalescent plasma (i.e. plasma collected from those who recover from COVID‐19) is being employed as an investigational therapy for treating COVID‐19 [77, 78]. One theoretical ramification of an association with ABO type could affect such a therapy; specifically, donors with higher titres of antibodies could be recruited selectively based on blood type [79]. Nonetheless, this would still require definitive proof of differential titres by ABO type. Routine ABO testing of COVID‐19 patients could also guide decision‐making, for example, by lowering thresholds for escalating care with higher risk blood groups. However, realistically, this is unlikely to happen given that risk, if present, is not sufficiently convincing to alter population‐based care.
Conclusions
The role of ABO blood group in SARS‐CoV‐2 infectivity and COVID‐19 disease severity requires additional study; however, accumulating evidence suggests that, at biochemical and physiological levels, there may be a contribution of ABO blood type to disease biology. It also must be recognized that host factors already identified as contributing to COVID‐19 severity, play a dominant role, coupled with timely access to appropriate medical care. By contrast, the role of ABO type is likely secondary and non‐modifiable.
Conflict of interest
RG serves on the medical advisory board of Rigel and reports personal consulting fees from Alexion Pharmaceuticals and TERUMO BCT outside of the submitted work. EMB reports personal fees and non‐financial support from Terumo BCT, personal fees and non‐financial support from Grifols Diagnostics Solutions and Abbott Laboratories, outside of the submitted work; EMB is a member of the United States Food and Drug Administration (FDA) blood Products Advisory Committee. Any views or opinions that are expressed in this manuscript are that of the author’s, based on his own scientific expertise and professional judgement; they do not necessarily represent the views of either the Blood Products Advisory Committee or the formal position of FDA and also do not bind or otherwise obligate or commit either Advisory Committee or the Agency to the views expressed. DVD serves on the advisory board of Macopharma. SLS serves on the advisory board of Hemanext, Inc. is a consultant with Tioma, Inc., and is the Executive Director of the Worldwide Initiative for Rh Disease Eradication (WIRhE).
Funding
EMB’s effort is supported in part by the National Institutes of Health (K23 HL151826). SLS’s effort is supported in part by the National Institutes of Health (R01 HL133049, R01 HL139489, R01 HL148151 and N26819 HB00003R).
References
- 1. Zhu N, Zhang D, Wang W, et al.: A Novel Coronavirus from patients with pneumonia in China, 2019. N Engl J Med 2020; 382:727–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Wiersinga WJ, Rhodes A, Cheng AC, et al.: Pathophysiology, transmission, diagnosis, and treatment of coronavirus disease 2019 (COVID‐19): a review. JAMA 2020; 324:782–93. [DOI] [PubMed] [Google Scholar]
- 3. Barnkob MB: Reduced prevalence of SARS‐CoV‐2 infection in ABO blood group O. Blood Adv 2020; 4(20):4990–4993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Hoiland RL: The association of ABO blood group with indices of disease severity and multiorgan dysfunction in COVID‐19. Blood Adv 2020; 4(20):4981–4989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Zhao J, Yang Y, Huang H‐P, et alRelationship between the ABO Blood Group and the COVID‐19 Susceptibility. medRxiv 2020.
- 6. Latz CA, DeCarlo C, Boitano L, et al.: Blood type and outcomes in patients with COVID‐19. Ann Hematol 2020; 1–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Li J, Wang X, Chen J, et al.: Association between ABO blood groups and risk of SARS‐CoV‐2 pneumonia. Br J Haematol 2020; 190:24–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Piccinni MP, Vultaggio A, Scaletti C, et al.: Type 1 T helper cells specific for Candida albicans antigens in peripheral blood and vaginal mucosa of women with recurrent vaginal candidiasis. J Infect Dis 2002; 186:87–93. [DOI] [PubMed] [Google Scholar]
- 9. Yamamoto F, Clausen H, White T, et al.: Molecular genetic basis of the histo‐blood group ABO system. Nature 1990; 345:229–33. [DOI] [PubMed] [Google Scholar]
- 10. Blood groups and red cell antigens. Dean L., Bethesda (MD): National Center for Biotechnology Information (US). 2005.
- 11. Holodick NE, Rodriguez‐Zhurbenko N, Hernandez AM: Defining natural antibodies. Front Immunol 2017; 8:872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Gérard C, Maggipinto G, Minon JM: COVID‐19 & ABO blood group: another viewpoint. Br J Haematol 2020; 190:e93–e94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Stussi G, Huggel K, Lutz H, et al.: Isotype‐specific detection of ABO blood group antibodies using a novel flow cytometric method. Br J Haematol 2005; 130:954–63. [DOI] [PubMed] [Google Scholar]
- 14. Daniele F: Anti‐A Isohemagglutinin titers and SARS‐CoV2 neutralization: implications for children and convalescent plasma selection. Br J Haematol 2020; 190:e148–e150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Zimring JC, Spitalnik SL: Pathobiology of transfusion reactions. Annu Rev Pathol 2015; 10:83–110. [DOI] [PubMed] [Google Scholar]
- 16. Branch DR: Anti‐A and anti‐B: what are they and where do they come from? Transfusion 2015; 55(Suppl 2):S74–9. [DOI] [PubMed] [Google Scholar]
- 17. Mourant AE, Kopec AC, Domaniewska‐Sobczak K: The Distribution of Human Blood Groups and Other Polymorphisms. London, Oxford University Press, 1976. [Google Scholar]
- 18. Estrada‐Mena B, Estrada FJ, Ulloa‐Arvizu R, et al.: Blood group O alleles in Native Americans: implications in the peopling of the Americas. Am J Phys Anthropol 2010; 142:85–94. [DOI] [PubMed] [Google Scholar]
- 19. Clausen H, Hakomori S: ABH and related histo‐blood group antigens; immunochemical differences in carrier isotypes and their distribution. Vox Sang 1989; 56:1–20. [DOI] [PubMed] [Google Scholar]
- 20. Dean L: Blood Groups and Red Cell Antigens. Bethesda (MD), National Center for Biotechnology Information (US), 2005. [Google Scholar]
- 21. Brockhausen I: Crossroads between bacterial and mammalian glycosyltransferases. Front Immunol 2014; 5:492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Zouine S, Marnissi F, Otmani N, et al.: Expression of histo‐blood group antigens in tumor and adjacent normal breast tissues as prognostic markers of breast carcinoma. J Breast Cancer 2020; 23:69–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Gampa A, Engen PA, Shobar R, et al.: Relationships between gastrointestinal microbiota and blood group antigens. Physiol Genomics 2017; 49:473–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Sousa NC, Anicchino‐Bizzacchi JM, Locatelli MF, et al.: The relationship between ABO groups and subgroups, factor VIII and von Willebrand factor. Haematologica 2007; 92:236–9. [DOI] [PubMed] [Google Scholar]
- 25. Liumbruno GM, Franchini M: Beyond immunohaematology: the role of the ABO blood group in human diseases. Blood Transfus 2013; 11:491–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Franchini M, Capra F, Targher G, et al.: Relationship between ABO blood group and von Willebrand factor levels: from biology to clinical implications. Thromb J 2007; 5:14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Ibrahim‐Kosta M, Bailly P, Silvy M, et al.: ABO blood group, glycosyltransferase activity and risk of venous thromboembolism. Thromb Res 2020; 193:31–5. [DOI] [PubMed] [Google Scholar]
- 28. Wu O, Bayoumi N, Vickers MA, et al.: ABO(H) blood groups and vascular disease: a systematic review and meta‐analysis. J Thromb Haemost 2008; 6:62–9. [DOI] [PubMed] [Google Scholar]
- 29. Grundbacher FJ: Changes in the human a antigen of erythrocytes with the individual's age. Nature 1964; 204:192–4. [DOI] [PubMed] [Google Scholar]
- 30. Ravn V, Dabelsteen E: Tissue distribution of histo‐blood group antigens. Apmis 2000; 108:1–28. [DOI] [PubMed] [Google Scholar]
- 31. Rummel SK, Ellsworth RE: The role of the histoblood ABO group in cancer. Future Sci OA 2016; 2:FSO107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Karakas M, Baumert J, Kleber ME, et al.: A variant in the ABO gene explains the variation in soluble E‐selectin levels‐results from dense genotyping in two independent populations. PLoS One 2012; 7:e51441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Pare G, Chasman DI, Kellogg M, et al.: Novel association of ABO histo‐blood group antigen with soluble ICAM‐1: results of a genome‐wide association study of 6,578 women. PLoS Genet 2008; 4:e1000118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Franchini M, Lippi G: The intriguing relationship between the ABO blood group, cardiovascular disease, and cancer. BMC Med 2015; 13:7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Randi AM, Laffan MA: Von Willebrand factor and angiogenesis: basic and applied issues. J Thromb Haemost 2017; 15:13–20. [DOI] [PubMed] [Google Scholar]
- 36. Eder AF, Spitalnik SL: Blood Group Antigens as Receptors for Pathogens; in: Blancher A, Klein J, Socha W (eds): Molecular Biology and Evolution of Blood Group and MHC Antigens in Primates. Berlin, Heidelberg, Springer, Berlin Heidelberg, 1997:268–304. [Google Scholar]
- 37. Cooling L: Blood groups in infection and host susceptibility. Clin Microbiol Rev 2015; 28:801–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Kaufmann B, Baxa U, Chipman PR, et al.: Parvovirus B19 does not bind to membrane‐associated globoside in vitro. Virology 2005; 332:189–98. [DOI] [PubMed] [Google Scholar]
- 39. Munakata Y, Saito‐Ito T, Kumura‐Ishii K, et al.: Ku80 autoantigen as a cellular coreceptor for human parvovirus B19 infection. Blood 2005; 106:3449–56. [DOI] [PubMed] [Google Scholar]
- 40. Weigel‐Kelley KA, Yoder MC, Srivastava A: Alpha5beta1 integrin as a cellular coreceptor for human parvovirus B19: requirement of functional activation of beta1 integrin for viral entry. Blood 2003; 102:3927–33. [DOI] [PubMed] [Google Scholar]
- 41. Cooling LL, Koerner TA, Naides SJ: Multiple glycosphingolipids determine the tissue tropism of parvovirus B19. J Infect Dis 1995; 172:1198–205. [DOI] [PubMed] [Google Scholar]
- 42. Brown KE, Hibbs JR, Gallinella G, et al.: Resistance to parvovirus B19 infection due to lack of virus receptor (erythrocyte P antigen). N Engl J Med 1994; 330:1192–6. [DOI] [PubMed] [Google Scholar]
- 43. Borén T, Falk P, Roth KA, et al.: Attachment of Helicobacter pylori to human gastric epithelium mediated by blood group antigens. Science (New York, NY) 1993; 262:1892–5. [DOI] [PubMed] [Google Scholar]
- 44. Rossez Y, Maes E, Lefebvre Darroman T, et al.: Almost all human gastric mucin O‐glycans harbor blood group A, B or H antigens and are potential binding sites for Helicobacter pylori. Glycobiology 2012; 22:1193–206. [DOI] [PubMed] [Google Scholar]
- 45. Aspholm‐Hurtig M, Dailide G, Lahmann M, et al.: Functional adaptation of BabA, the H. pylori ABO blood group antigen binding adhesin. Science (New York, NY) 2004; 305:519–22. [DOI] [PubMed] [Google Scholar]
- 46. Chakrani Z, Robinson K, Taye B: Association between ABO blood groups and Helicobacter pylori infection: a meta‐analysis. Scientific Reports 2018; 8:17604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Cheng Y, Cheng G, Chui CH, et al.: ABO blood group and susceptibility to severe acute respiratory syndrome. Jama 2005; 293:1450–1. [DOI] [PubMed] [Google Scholar]
- 48. Leaf RK, Al‐Samkari H, Brenner SK, et al.: ABO phenotype and death in critically Ill patients with COVID‐19. Br J Haematol 2020; 190:e204–e208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Göker H, Karakulak EA, Demiroğlu H, et al.: The effects of blood group types on the risk of COVID‐19 infection and its clinical outcome. Turkish J Med Sci 2020; 50:679–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Abdollahi A, Mahmoudi‐Aliabadi M, Mehrtash V, et al.: The novel coronavirus SARS‐CoV‐2 vulnerability association with ABO/Rh blood types. Iran J Pathol 2020; 156–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Zeng X, Fan H, Lu D, et al.: Association between ABO blood groups and clinical outcome of coronavirus disease 2019: Evidence from two cohorts. medRxiv 2020.
- 52. Wu Y, Feng Z, Li P, et al.: Relationship between ABO blood group distribution and clinical characteristics in patients with COVID‐19. Clin Chim Acta 2020; 509:220–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Patel EU, Bloch EM, Grabowski MK, et al.: Sociodemographic and behavioral characteristics associated with blood donation in the United States: a population‐based study. Transfusion 2019; 59:2899–907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Golding J, Northstone K, Miller LL, et al.: Differences between blood donors and a population sample: implications for case‐control studies. Int J Epidemiol 2013; 42:1145–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Takagi H: Down the Rabbit‐Hole of blood groups and COVID‐19. Br J Haematol 2020; 190:e268–e270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Zietz M, Tatonetti NP: Testing the association between blood type and COVID‐19 infection, intubation, and death. medRxiv 2020. [DOI] [PMC free article] [PubMed]
- 57. Ellinghaus D, Degenhardt F, Bujanda L, et al.: Genomewide Association Study of Severe Covid‐19 with Respiratory Failure. N Engl J Med 2020; 383:1522–1534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Breiman A, Ruven‐Clouet N, Le Pendu J: Harnessing the natural anti‐glycan immune response to limit the transmission of enveloped viruses such as SARS‐CoV‐2. PLoS Pathog 2020; 16:e1008556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Li F, Li W, Farzan M, et al.: Structure of SARS coronavirus spike receptor‐binding domain complexed with receptor. Science 2005; 309:1864–8. [DOI] [PubMed] [Google Scholar]
- 60. Krokhin O, Li Y, Andonov A, et al.: Mass spectrometric characterization of proteins from the SARS virus: a preliminary report. Mol Cell Proteomics 2003; 2:346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Ying W, Hao Y, Zhang Y, et al.: Proteomic analysis on structural proteins of Severe Acute Respiratory Syndrome coronavirus. Proteomics 2004; 4:492–504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Watanabe Y, Allen JD, Wrapp D, et al.: Site‐specific glycan analysis of the SARS‐CoV‐2 spike. Science 2020; 369:330–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Lan J, Ge J, Yu J, et al.: Structure of the SARS‐CoV‐2 spike receptor‐binding domain bound to the ACE2 receptor. Nature 2020; 581:215–20. [DOI] [PubMed] [Google Scholar]
- 64. Guillon P, Clément M, Sébille V, et al.: Inhibition of the interaction between the SARS‐CoV spike protein and its cellular receptor by anti‐histo‐blood group antibodies. Glycobiology 2008; 18:1085–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Breiman A, Ruvën‐Clouet N, Le Pendu J: Harnessing the natural anti‐glycan immune response to limit the transmission of enveloped viruses such as SARS‐CoV‐2. PLoS Pathogens 2020; 16:e1008556‐e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Wu S, Arthur CM, Wang J, et al.: SARS‐CoV‐2 Receptor Binding Domain Preferentially Recognizes Blood Group A. Blood Adv. 2020. (in press). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. de França NDG, Poli MCC, Ramos PGdA, et al.: Titers of ABO antibodies in group O blood donors. Rev Bras Hematol Hemoter 2011; 33:259–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Tendulkar AA, Jain PA, Velaye S: Antibody titers in Group O platelet donors. Asian J Transfus Sci 2017; 11:22–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Gallian P, Pastorino B, Morel P, et al.: Lower prevalence of antibodies neutralizing SARS‐CoV‐2 in group O French blood donors. Antiviral Res 2020; 181:104880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Wrapp D, Wang N, Corbett KS, et al.: Cryo‐EM structure of the 2019‐nCoV spike in the prefusion conformation. Science 2020; 367:1260–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Rieben R, Buchs JP, Fluckiger E, et al.: Antibodies to histo‐blood group substances A and B: agglutination titers, Ig class, and IgG subclasses in healthy persons of different age categories. Transfusion 1991; 31:607–15. [DOI] [PubMed] [Google Scholar]
- 72. Anstee D: The relationship between blood groups and disease. Blood 2010; 115:4635–43. [DOI] [PubMed] [Google Scholar]
- 73. Dai X: ABO blood group predisposes to COVID‐19 severity and cardiovascular diseases. Eur J Prev Cardiol 2020; 27(13):1436–1437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Ackermann M, Verleden SE, Kuehnel M, et al.: Pulmonary vascular endothelialitis, thrombosis, and angiogenesis in Covid‐19. N Engl J Med 2020; 383:120–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Bilaloglu S, Aphinyanaphongs Y, Jones S, et al.: Thrombosis in hospitalized patients with COVID‐19 in a New York City health system. JAMA 2020; 324:799–801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. O'Donnell J, Laffan MA: The relationship between ABO histo‐blood group, factor VIII and von Willebrand factor. Transfus Med 2001; 11:343–51. [DOI] [PubMed] [Google Scholar]
- 77. Bloch EM, Goel R, Wendel S, et al.: Guidance for the procurement of COVID‐19 convalescent plasma: differences between high‐ and low‐middle‐income countries. Vox Sang 2020. (in press). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Al‐Riyami AZ, Schafer R, van den Berg K, et al.: Clinical use of Convalescent Plasma in the COVID‐19 pandemic: a transfusion‐focussed gap analysis with recommendations for future research priorities. Vox Sang 2020. (in press). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Bloch EM, Patel EU, Marshall C, Littlefield K, Goel R, Grossman BJ, Winters JL, Shrestha R, Burgess I, Laeyendecker O, Shoham S, Sullivan D, Gehrie EA, Redd AD, Quinn TC, Casadevall A, Pekosz A, Tobian AAR: ABO blood group and SARS‐CoV‐2 antibody response in a convalescent donor population. Accepted. Vox Sang 2020. (in press). [DOI] [PMC free article] [PubMed] [Google Scholar]