

Abstract
Background
Laccases and laccase-like multicopper oxidases (LMCOs) oxidize a vast array of phenolic compounds and amines, releasing water as a byproduct. Their low substrate specificity is responsible for their tremendous biotechnological interest, since they have been used for numerous applications. However, the laccases characterized so far correspond to only a small fraction of the laccase genes identified in fungal genomes. Therefore, the knowledge regarding the biochemistry and physiological role of minor laccase-like isoforms is still limited.
Results
In the present work, we describe the isolation, purification and characterization of two novel LMCOs, PcLac1 and PcLac2, from Pleurotus citrinopileatus. Both LMCOs were purified with ion-exchange chromatographic methods. PcLac2 was found to oxidize a broader substrate range than PcLac1, but both LMCOs showed similar formal potentials, lower than those reported previously for laccases from white-rot fungi. Proteomic analysis of both proteins revealed their similarity with other well-characterized laccases from Pleurotus strains. Both LMCOs were applied to the oxidation of ferulic and sinapic acid, yielding oligomers with possible antioxidant activity.
Conclusions
Overall, the findings of the present work can offer new insights regarding the biochemistry and variability of low-redox potential laccases of fungal origin. Low-redox potential biocatalysts could offer higher substrate selectivity than their high-redox counterparts, and thus, they could be of applied value in the field of biocatalysis.
Supplementary Information
The online version contains supplementary material available at 10.1186/s13068-021-01937-7.
Keywords: Laccase-like multicopper oxidases, Biocatalysis, Phenol oligomers
Background
Multicopper oxidases (MCOs) constitute a group of proteins with a distinctive structure, occupying at least four copper atoms in three different copper centers T1, T2, and T3. Their catalytic mechanism consists of substrate oxidation at a mononuclear copper center (T1) and subsequent electron transfer via a His–Cys–His tripeptide motif to a trinuclear copper center (T1/T2) [1, 2]. Laccases (benzenediol: oxygen oxidoreductases, Lac, EC 1.10.3.2), one of the major MCO superfamilies, act on phenolic substrates, by catalyzing the one-electron oxidation of phenolic hydroxyl groups to phenoxy radicals, reducing O2 to water by a four-electron transfer [3]. They are highly oxidizing enzymes, with E0′ values varying between + 500 and + 800 mV vs NHE (normal hydrogen electrode) [4]. Hence, their substrate spectrum includes mono-, di-, polyphenols, methoxy-substituted phenols, aromatic compounds, and amines, such as acrylamines or aminophenols. A study upon the catalytic efficiency of four laccases from basidiomycetes revealed that in the case of monophenolic substrates, E0′ values of the enzyme play a defining role in substrate transformation, whereas for larger substrates, the structure of the active site is of equal importance [5]. Their biological functions involve lignin biosynthesis and degradation, morphogenesis, stress response, pigment and melanin formation, plant pathogenesis, and insect sclerotization [2, 3].
An interesting characteristic of basidiomycetes is the presence of multiple copies of laccase genes in their genome, also known as laccase multigene families. Such multigene families usually contain four to eight genes and encode a variety of isoenzymes. A typical fungal multigene family has been found to encode up to 17 laccase isoenzymes in Coprinopsis cinerea [6], while a greater variety of laccase isoforms is attributed to post-translational modifications. Moreover, laccases from different strains of the same species can possess distinct properties. Glazunova et al. [7] highlighted the importance of glycosylation at modulating catalytic properties of four laccases from different strains of Steccherinum ochraceum. However, isoenzymes can differ considerably regarding their biochemical characteristics and can also have different roles in the physiology of the microorganism [8]. A typical example of laccase multigene families is the existence of 12 phenol oxidase (laccase) genes in Pleurotus ostreatus [9]. Laccase gene transcription is found to be regulated by culture conditions, while enzymatic activities and gene transcription profiles differ even between closely related strains [9]. Additionally, Park et al. [10] revealed that the expression of those genes differs among developmental stages and under various growth conditions.
With the number of characterized laccase enzymes increasing steadily over the last decades, many counterparts were described in the literature with variable properties, which can differ significantly from those previously known for traditional laccases. Laccase-like MCOs (LMCOs) is a term proposed by Reiss et al. [1], to describe a multigenic family of oxidoreductases with especially variable characteristics, mainly related to substrate specificity, thus confining laccases to MCOs that are active against urushiol, an unsaturated alkyl catechol. LMCOs are found in bacteria, fungi, plant, and insects, and are considered to have similar biological functions to laccases, even though their exact role and substrate spectrum is yet to be clarified. Even though laccase-like enzymes show conserved active site residues, where the copper ligands are organized, they may lack other laccase-specific signature sequences. For example, Kumar et al. [11] described a set of conserved consensus sequences L1–L4 in an effort to develop reliable homology tools to identify ‘true’ laccases. LMCOs usually contain these motifs, but with several amino acid substitutions, as shown for one of the few fungal LMCOs described in the literature, TtLMCO1 from Thermothelomyces thermophila [12].
Laccase multigene families and LMCOs have been studied from various perspectives, including enzyme characterization, transcriptome and secretome analysis, expression, and biological function studies. Regarding fungal LMCOs, Chen et al. [13] indicated the presence of laccase-like genes in ectomycorrhizal basidiomycetes (Piloderma byssinum). LCMOs have also been detected in Trichoderma reesei [14] and Aspergillus niger genomes [15]. Another phylogenetic analysis of a laccase multigene family was conducted on seven Auricularia auricula-judae laccase genes containing the signature sequences L1–L4, indicating their functional differences from other basidiomycete LMCOs [16]. Furthermore, Vasina et al. [17] focused on a laccase multigene family of the basidiomycete Trametes hirsuta strain 072, locating five laccase genes which are not only categorized to different clusters within the genus Trametes, but their production also appears to depend on the stage of biomass degradation.
In the field of Industrial Biotechnology, the contribution of laccases has been undeniable. Food and beverage processing, bioethanol production, bioremediation, paper production, dye decolorization, textile functionalization, and biosensor construction are only some of the applications where laccases have been proven to be valuable [18–21]. Especially for LMCOs, most of the proposed biotechnological applications focus on dye decolorization [22, 23].
Moreover, the exploitation of laccases is gaining interest in the field of organic synthesis (i.e., as green catalysts). For example, laccases catalyze the synthesis or modification of bioactive compounds, such as antioxidants [24], alkaloids [25], amino acid analogues [26], or bioactive polymers [27–30]. Although the value of high-redox potential laccases in the field of biocatalysis is widely accepted, there are only a few references on the synthetic properties of low-redox LMCOs, such is the case of TtLMCO1 from T. thermophila which catalyzed the conversion of 2′,3,4-trihydroxychalcone to 3′,4′-dihydroxy-aurone [12].
The aim of the present study is the isolation and characterization of two novel LCMOs from Pleurotus citrinopileatus (Fungi, Basidiomycota), with high potential for biotechnological applications. Pleurotus species are known to contain laccase multigene families in their genome [9], while the degradative potential of P. citrinopileatus toward lignocellulosic substrates has been previously demonstrated in sugarcane bagasse [31], paper and cardboard [32], and olive oil mill wastewater (OMWW [33]). Two LMCOs were isolated from the culture supernatants of P. citrinopileatus grown on OMWW, and they were fully characterized. Proteomic analyses revealed similarities with major secreted laccases in other Pleurotus species. Based on their low formal potentials and thus high substrate specificity, this work intends to highlight the synthetic properties of PcLac1 and PcLac2, and provide evidence toward their potential use as biocatalysts in the fields of Green Chemistry and Organic Synthesis.
Results
Production and purification of laccases
For the isolation of putative laccase enzymes from P. citrinopileatus, the strain was grown as previously described [33], i.e., in OMWW supplemented with corn steep liquor, for 12 days, until the maximum levels of laccase production were reached, around 1000 U L−1. Then, the culture supernatant was collected and concentrated before purification. Purification with Q sepharose column resulted in two separate fractions with laccase activity, which were further purified separately. After an additional purification step with DEAE sepharose, two novel laccases were isolated, named PcLac1 and PcLac2. The purification results are shown in Additional file 1: Table S1. Their purity was confirmed with SDS-PAGE analysis and by the respective zymograms (Fig. 1). From the SDS-PAGE, it can be concluded that PcLac1 is a 60 kDa protein, while PcLac2 is closer to 75 kDa.
Fig. 1.
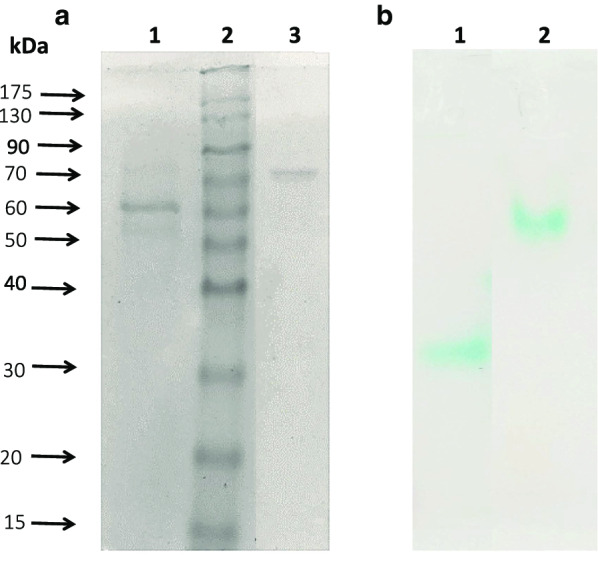
a SDS-PAGE analysis of the isolated proteins. Lane 1: PcLac1, lane 2: protein marker, lane 3: PcLac2. b Zymogram of the isolated proteins, lane 1: PcLac1, lane 2: Pclac2
Proteomic analysis
The identification of in-gel digested protein bands is displayed in Table 1 and Additional file 2. In gel band PcLac1, 22 MSMS spectra matched with 9 peptides providing 18.6% coverage of the sequence Q2VT18_PLEPU Laccase-2 of Pleurotus pulmonarius, which is 96.5% homologous to Q12739 LAC2_PLEOS—Laccase-2 of Pleurotus ostreatus.
Table 1.
Proteomic analysis of PcLac1 and PcLac2
| PcLac1 | PcLac2 | ||
|---|---|---|---|
| Description | > tr|Q2VT18|Q2VT18_PLELaccase 2 from Pleurotus pulmonarius | > tr|A0A067N2X1|A0A067N2X1_PLEOS Uncharacterized protein from Pleurotus ostreatus | > tr|Q2VT19|Q2VT19_PLELaccase 6 from Pleurotus pulmonarius |
| |Log Prob| | 87.07 | 69.7 | 188.3 |
| Best |Log Prob|" | 17.31 | 13.4 | 15.9 |
| Best score | 985.80 | 873.2 | 958.7 |
| Total Intensity | 4.37E + 08 | 4.5E + 08 | 4.1E + 08 |
| # of PSM spectra | 22 | 27 | 65 |
| # of unique peptides | 9 | 11 | 29 |
| # of mod peptides | 1 | 3 | 9 |
| Coverage % | 18.61 | 19.5 | 42.6 |
| # AA's in protein | 532 | 591 | 521 |
| BLAST homology | 96.5% homologous to Q12739 | 61% homologous to V2XEX2 | 99% homologous to A0A067NLM3 |
Gel band of PcLac2 displays 92 MSMS spectra matching in total 40 peptides that belong to two different proteins; a laccase from P. pulmonarius (Q2VT19_PLEPU laccase 6, 29 peptides with 42.6% sequence coverage) that is 99% homologous to laccase 0A067NLM3_PLEOS from P. ostreatus and an uncharacterized protein from P. ostreatus (A0A067N2X1_PLEOS, 11 peptides with 19.5% sequence coverage) that is 61% homologous to copper radical oxidase V2XEX2 from Moniliophthora roreri.
Biochemical characterization of laccases
The optimal temperature and pH conditions were investigated for both laccases, and the results are shown in Fig. 2. Both enzymes have a temperature optimum at 55 °C, but while PcLac2 retains over 90% of its activity in the range 50–60 °C, PcLac1 retains slightly lower than 80% in this same range. In addition, pH optimum was 4 for PcLac2 and 4.5 for PcLac1, while both enzymes retained over 80% of their activity in the range 3–4.5. However, in higher pH, the residual activity dropped abruptly, in pH 5 for PcLac2 and pH 6 for PcLac1. Stability of both laccases was also investigated for a range of temperatures and pH values, and the results are shown in Fig. 3. PcLac1 was more thermostable than PcLac2, which lost almost all activity after 4 h incubation at temperatures over 40 °C. On the contrary, PcLac1 retained 60% of its activity after 4 h at 40 °C, and a little over 40% for the same time at 50 °C. Regarding the pH stability, both laccases were stable in neutral pH, but they lose quickly their activity at pH lower than 4. PcLac1 was also stable at basic pH, but PcLac2 lost some of its activity at pH 8 or higher.
Fig. 2.

Temperature (a) and pH (b) optimum for the laccases PcLac1 (black circles) and PcLac2 (white circles)
Fig. 3.

a Temperature stability of PcLac1 (black circles) and PcLac2 (white circles) after 4 h incubation b pH stability of PcLac1 (black circles) and PcLac2 (white circles) after 24 h incubation
The substrate oxidation spectrum was investigated for both laccases, against a variety of substrates, as shown in Table 2. From the 24 substrates tested, PcLac1 and PcLac2 oxidized unambiguously 8 and 14 substrates, respectively. Both laccases were found to strongly oxidize ABTS, but no activity was detected against ascorbic acid and tyrosine, indicating that the studied laccases do not have ascorbate oxidase or tyrosinase activity. Hydroxybenzenes, such as catechol, hydroquinone, and pyrogallol, were also oxidized by the enzymes, similarly to most known laccases. However, the very low levels of guaiacol oxidation detected for both enzymes are rather unusual for fungal laccases, although both enzymes oxidized 2,6-dimethoxyphenol. Phenol and vanillin were also not oxidized by the studied laccases, indicating a rather low formal potential; however, PcLac2 activity was demonstrated—to a certain extent—against vanillic acid, veratryl alcohol (3,4-dimethoxybenzyl alcohol), and p-cresol. These distinct differences in the substrate oxidation spectrum of the two laccases might indicate a potentially different organization and architecture of the copper cluster site. Moreover, both laccases were found to strongly oxidize most of the hydroxycinnamic acids tested, except p-coumaric acid, but only PcLac2 was able to oxidize hydroxybenzoic acids, such as vanillic and gallic acids. Interestingly, both enzymes strongly oxidized quercetin, which might be an indication of their possible involvement in the production of bioactive compounds and secondary metabolites from P. citrinopileatus.
Table 2.
Substrate oxidation spectrum of PcLac1 and PcLac2
| Substrate | PcLac1 | PcLac2 | |
|---|---|---|---|
| Hydroxybenzenes | |||
| 1 | Phenol | − | − |
| 2 | Catechol | + | + + |
| 3 | Resorcinol | − | − |
| 4 | Hydroquinone | + | + |
| 5 | Chlorocatechol | − | + |
| 6 | Pyrogallol | + | + |
| Methoxyphenols | |||
| 7 | Guaiacol | ± | ± |
| 8 | 2.6-Dimethoxyphenol | + | + |
| Aromatic alcohols | |||
| 9 | 3.4-Dimethoxybenzyl alcohol | − | + |
| Phenethyl alcohols | |||
| 10 | Tyrosol | − | − |
| Aromatic amines | |||
| 11 | Aniline | − | − |
| 12 | Tyrosine | − | − |
| Phenolic aldehydes | |||
| 13 | Vanillin | − | − |
| Flavonoids | |||
| 14 | Quercetin | + + | + + |
| Hydroxycinnamic acids | |||
| 15 | Caffeic acid | + | + |
| 16 | Ferulic acid | + | + |
| 17 | Sinapic acid | + | + |
| 18 | p-Coumaric acid | − | − |
| Hydroxybenzoic acids | |||
| 19 | Vanillic acid | − | + |
| 20 | Gallic acid | − | + |
| Aromatic azo compounds | |||
| 21 | ABTS | + + + | + + + |
| Other acids | |||
| 22 | Ascorbic acid | − | − |
| Cresols | |||
| 23 | o-Cresol | − | − |
| 24 | m-Cresol | − | − |
| 25 | p-Cresol | − | + |
Activity, in terms of absorbance difference in the recorded UV/Vis spectrum between reaction and blank, is indicated with ( +) when positive (indicating absorbance difference in the spectrum maxima), with (−) when negative and with ( ±) when ambiguous. Multiple ( +) signs indicate differences higher than 1 (+ +) or 2 (+ + +) absorbance units
The substrate specificity was determined for both enzymes against some of the tested substrates, and the results are shown in Table 3. ABTS was the substrate most strongly oxidized by both laccases, but PcLac2 showed a significantly higher specific activity than PcLac1. The specificity for the other compounds tested is similar for both enzymes, yet significantly lower. Remarkably, the specific activity of both enzymes against guaiacol was very low, several times lower than in the case of 2,6-DMP, a property rather unusual for laccases from fungal sources.
Table 3.
Substrate specificity of PcLac1 and PcLac2
| Substrate | PcLac1 (Units/mg) | PcLac2 (Units/mg) |
|---|---|---|
| ABTS | 1.48 ± 0.09 | 4.78 ± 0.33 |
| 2.6 DMP | 0.08 ± 0.00 | 0.019 ± 0.003 |
| Catechol | 0.06 ± 0.01 | 0.04 ± 0.01 |
| Pyrogallol | 0.020 ± 0.001 | 0.0390 ± 0.0003 |
| Guaiacol | 0.003 ± 0.000 | 0.0006 ± 0.0002 |
| Hydroquinone | 0.079 ± 0.003 | 0.09 ± 0.02 |
Michaelis–Menten kinetic parameters were calculated for both enzymes on ABTS and 2,6-DMP (Table 4). Both enzymes showed a high affinity for ABTS, similarly to other fungal laccases, but a significantly lower affinity for 2,6-DMP. However, PcLac1 exhibited a much higher affinity for both substrates compared to PcLac2. The turnover number was low in the case of 2,6-DMP oxidation, but considerably higher in the case of ABTS oxidation. PcLac2 showed a very high turnover number for ABTS oxidation in comparison with PcLac1. The same trend was followed for the catalytic efficiency.
Table 4.
Michaelis–Menten kinetic parameters calculated for PcLac1 and PcLac2
| Enzyme | Substrate | Vmax (Units/mg) | kcat (min−1) | KM (μΜ) | Kcat/KM (μM−1 min−1) |
|---|---|---|---|---|---|
| PcLac1 | ABTS | 2.38 ± 0.06 | 95.2 ± 2.3 | 60 ± 7 | 1.57 ± 0.17 |
| 2.6-DMP | 0.36 ± 0.01 | 14.2 ± 0.5 | 672 ± 56 | 0.021 ± 0.002 | |
| PcLac2 | ABTS | 4.6 ± 0.1 | 368.7 ± 8.8 | 165 ± 15 | 2.24 ± 0.21 |
| 2.6-DMP | 0.041 ± 0.002 | 3.3 ± 0.2 | 1070 ± 105 | 0.0030 ± 0.0003 |
The effect of various inhibitors and solvents was also tested on the activity of both PcLac1 and PcLac2, and the results are shown in Fig. 4. EDTA, a known inhibitor of laccase activity, did not seem to affect significantly the activity of both enzymes, at least up to the tested concentrations. On the contrary, NaN3, also a known laccase inhibitor, highly affected the activity of both enzymes; residual activity was lower than 50% for both enzymes, even with the addition of 0.01 mM NaN3. Addition of copper at low concentrations enhanced the activity of PcLac1, while PcLac2 was not affected. NaCl strongly inhibited the activity of PcLac1, but not of PcLac2. Regarding the effect of solvents, PcLac1 was strongly affected in most cases, since the residual activity was lower than 60% in all conditions tested. On the other hand, PcLac2 was found to be more resistant to the presence of solvents. The residual activity observed was higher than 80% with the addition of 10% of either ethanol, methanol, acetone, and DMSO, and over 60% of initial activity was observed with the addition of 10% 1,4-dioxane. The satisfactory stability of PcLac2 in the presence of solvents might be an attractive property for biocatalysis applications, especially in biotransformations of phenolic compounds, where the low water solubility of both substrates and products requires the use of organic solvents.
Fig. 4.

Effect of inhibitors (a) and solvents (b) on the activity of laccases PcLac1 (black bars) and PcLac2 (gray bars)
Determination of redox potential
FTacV was performed for PcLac1 and PcLac2 at different temperatures T and frequencies f, while keeping the scan rate v and the amplitude A0 steady at 50 mV/s and 180 mV, respectively. The temperatures examined were 30, 33, 39, and 43 °C, and the frequencies corresponded to values of 6, 9, 12, and 16 Hz. The 3rd harmonic for the different frequencies at an indicative temperature of 43 °C is presented in Additional file 1: Figure S1. In the third harmonic, the signal is already clear enough with no strong effect of capacitance currents, and thus, a value for the formal potential E0′ of the active site of the enzymes can be determined without having to resort to higher order of harmonics. The potential is calculated from the dominant peak of peak triplets that appear around 250 mV for PcLac1 and 180 mV vs Ag|AgCl, KCl sat. for PcLac2 (Table 5). An average value of the formal potential from the different frequencies was calculated at each temperature together with the respective standard deviation. From the pattern of the 3rd harmonics, it can be assumed that the reduction and oxidation of these enzymes fall into the quasi reversible case, where the value of the dominant peak corresponds to the value of the formal potential [34]. The smaller irreversible peak that appeared at more negative potentials in both cases is not attributed to the enzyme but to other impurities existing in the samples due to incomplete purification and, thus, should not be considered for the present study.
Table 5.
Calculated and corrected values of the apparent formal potential of PcLac1 and PcLac2 at different temperatures
| T (°C) | E0′ vs Ag|AgCl KCl sat. (mV) | E0′corrected vs Ag|AgCl KCl sat. (mV) corrected | E0′corrected vs NHE (mV) | Standard deviation |
|---|---|---|---|---|
| PcLac1 | ||||
| 30 | 261 | 256 | 453 | 1.2 |
| 33 | 262 | 257 | 454 | 0.7 |
| 39 | 260 | 255 | 452 | 1.0 |
| 43 | 270 | 265 | 462 | 7.8 |
| PcLac2 | ||||
| 30 | 182 | 177 | 374 | 3.9 |
| 33 | 187 | 182 | 379 | 2.8 |
| 39 | 188 | 183 | 380 | 1.8 |
| 43 | 182 | 177 | 374 | 10.1 |
Regarding the variance of the formal potential with the temperature, it can be concluded that in the temperature range examined, no considerable effect was observed.
Biocatalytic synthesis of phenolic acid derivatives
PcLac1 and PcLac2 were applied to oxidation reactions of ferulic acid and sinapic acid, with the aim to investigate their potential at oxidative oligomerization of phenolic acids. Samples were drawn at 3 h and 8 h intervals, and analyzed with UHPLC–HRMS/MS. After 24 h reaction, the formation of precipitate was observed, indicating the polymerization of the substrates to insoluble compounds. The reaction mixtures were centrifuged and the supernatant was also analyzed by UHPLC–HRMS/MS.
The results for the bioconversion of ferulic acid are shown in Table 6. The double decarboxylated dimer 1 was found in all samples of PcLac2 as a major constituent, and was detected in PcLac1 samples in traces only. The decarboxylated dimer could be detected in all samples, 3 isomers at Rt 7.07, 7.29, and 7.99 min (2, 4, 5). The dimer 3 was found in all samples. In PcLac1 samples, the dimer was major, while in PcLac2 samples, the decarboxylated dimers were the main constituents. In all samples, trimers were the less intense peaks.
Table 6.
LC–MS identification of the main ions detected during oxidation of ferulic acid by PcLac1 and PcLac2
| Compound | Rt (min) | m/z [M-H]− | MF | MS/MS | Identification | PcLac1 samples | PcLac2 samples |
|---|---|---|---|---|---|---|---|
| Dimers | |||||||
| 1 | 7.07 | 297.111 | C18H17O4 | 146 (C9H6O2) 109 (C6H5O2) | Double-decarboxylated diferulic acid | Minor | Major |
| 2 | 7.07 | 341 | C19H17O6 | 281 (C17H13O4), 267 (C16H11O4), 209 (C14H9O2), 159 (C10H7O2), 146 (C9H6O2) | Decarboxylated diferulic acid | Major | Major |
| 3 | 7.28 | 385 | C20H17O8 | 267 (C16H11O4), 239 (C15H11O3) | Diferulic acid | Major | Major |
| 4 | 7.29 | 341 | C19H17O6 | 281 (C17H13O4), 267 (C16H11O4), 209 (C14H9O2), 159 (C10H7O2), 146 (C9H6O2) | Decarboxylated diferulic acid | Major | Major |
| 5 | 7.99 | 341 | C19H17O6 | 281 (C17H13O4), 267 (C16H11O4), 209 (C14H9O2), 159 (C10H7O2), 146 (C9H6O2) | Decarboxylated diferulic acid | Major | Major |
| Trimers | |||||||
| 6 | 7.40 | 577 | C30H25O12 | 193 | Triferulic acid | Not detected | Major |
| 7 | 7.62 | 533 | C29H26O10 | 193 | Triferulic acid, decarboxylated | Not detected | Major |
| 8 | 7.63 | 577 | C30H25O12 | 193 | Triferulic acid | Not detected | Major |
| 9 | 7.65 | 533 | C29H26O10 | 193 (C10H9O4), | Triferulic acid, decarboxylated | Minor | Major |
| 10 | 7.67 | 533 | C29H26O10 | 193 | Triferulic acid, decarboxylated | Not detected | Major |
| 11 | 8.02 | 533 | C29H26O10 | 193 | Triferulic acid, decarboxylated | Minor | Major |
| 12 | 8.21 | 533 | C29H26O10 | 193 | Triferulic acid, decarboxylated | Minor | Major |
| 13 | 8.35 | 533 | C29H26O10 | 193 | Triferulic acid, decarboxylated | Minor | Major |
| 14 | 8.35 | 565 | C30H29O11 | 193 (C10H9O4), 178 (C9H6O4), 149 (C9H9O2), 134 (C8H6O2) | Unknown triferulic acid | Major | Major |
| 15 | 8.63 | 533 | C29H26O10 | 193 | Triferulic acid, decarboxylated | Not detected | Minor |
| Others | |||||||
| 16 | 7.5 | 551 | C29H28O11 | 193 (C10H9O4), 178 (C9H6O4), 149 (C9H9O2), 134 (C8H6O2) | Triferulic acid, decarboxylated-H2O* | Detected | Detected |
*The MS/MS shows that it is a triferulic acid derivative but it is not obvious how. A carboxyl group and a H2O molecule are missing
The mono decarboxylated triferulic acid comes out to 4 isomers 9, 11, 12, and 13, corresponding to Rt 7.65, 8.02, 8.21, and 8.35 min, respectively. These trimers are most probably position isomers, as they share similar fragmentation patterns. Their structure was confirmed by the MS/MS spectra with a fraction at 193 m/z corresponding to the ferulic acid ion. They were recorded in all samples, but they were less intense in PcLac1 samples. In addition, three other isomers (7, 10, and 15) were detected in PcLac2 samples at Rt 7.62, 7.67, and 8.63 min, respectively. Two isomers of triferulic acid with m/z 577, C30H25O12 (6 and 8) were detected in PcLac2 samples, with Rts 7.40 and 7.63, respectively.
The results for the bioconversion of sinapic acid are shown in Table 7. The decarboxylated dimer 19 was found in PcLac2 but not in PcLac1 samples. The dimer 18 was found in PcLac1 samples corresponding to 3 and 8 h of reaction, and not in PcLac2 reaction. Trimer 22 at m/z 667 Rt 7.37 were found in PcLac2 samples in very small quantities. The decarboxylated trimers 23 and 24 at m/z 623 at Rts 7.10 and 7.56, respectively, were found in PcLac2 reactions only and not in the corresponding PcLac1. A di-decarboxylated trimer (25) at m/z 579 at 8.16 min was found only in PcLac2 reactions and was absent from PcLac1 ones. Furthermore, a tetramer of the sinapic acid (26) was detected in PcLac2 samples.
Table 7.
LC–MS identification of the main ions detected during oxidation of sinapic acid by PcLac1 and PcLac2
| Compound | Rt (min) | m/z [M-H]− | MF | MS/MS | Identification | PcLac1 samples | PcLac2 samples |
|---|---|---|---|---|---|---|---|
| Dimers | |||||||
| 17 | 6.19 | 445 | C22H21O10 | 189 (C11H9O3), 174 (C10H6O3), 161 (C9H5O3), 145 (C9H5O2), 121 (C7H5O2) | Disinapic acid | Minor | Major |
| 18 | 6.69 | 445 | C22H21O10 | 189 (C11H9O3), 174 (C10H6O3), 161 (C9H5O3), 145 (C9H5O2), 121 (C7H5O2) | Disinapic acid | Major | Minor |
| 19 | 6.97 | 401.1220 | C21H21O8 |
189 (C11H9O3), 174 (C10H6O3), 161 (C9H5O3), 145 (C9H5O2), 133 (C8H5O2), 121 (C7H5O2) |
Decarboxylated disinapic acid | Not detected | Detected |
| 20 | 7.21 | 401.1220 | C21H21O8 | 189 (C11H9O3), 174 (C10H6O3), 161 (C9H5O3), 145 (C9H5O2), 133 (C8H5O2), 121 (C7H5O2) | Decarboxylated disinapic acid | Not detected | Detected |
| 21 | 6.26 | 357.060 | C18H13O8 | 271 (C14H7O6), 227 (C13H7O4), 199 (C12H7O2), 167 (C12H7O) | Bis-Decarboxylated disinapic acid | Detected | Detected |
| Trimers | |||||||
| 22 | 7.38 | 667.1643 | C33H31O15 | - | Trisinapic acid | Not detected | Minor |
| 23 | 7.09 | 623.1746 | C32H31O13 | 265 (C16H9O4), 209 (C14H9O2), 121 (C7H5O2) | Trisinapic acid, decarboxylated | Not detected | Detected |
| 24 | 7.55 | 623.1746 | C32H31O13 | 265 (C16H9O4), 209 (C14H9O2), 121 (C7H5O2) | Trisinapic acid, decarboxylated | Not detected | Detected |
| 25 | 8.16 | 579.1846 | C31H31O11 | 339 (C19H15O6) | Trisinapic acid, bis-decarboxylated | Not detected | Detected |
| Tetramers | |||||||
| 26 | 6.98 | 891.2319 | C44H43O20 | 341 (C19H17O6), 189 (C11H9O3), 174 (C10H6O3), 161 (C9H5O3), 145 (C9H5O2), 121 (C7H5O2) | Tetrasinapic acid | Not detected | Detected |
| Others | |||||||
| 27 | 7.99 | 503.1172 | C24H23O12 | 147 (C8H3O3) 119 (C7H3O2) | Unknown compound | Detected | Not detected |
In the bioconversion reactions for both ferulic and sinapic acid, the samples corresponding to 24 h reaction with PcLac1 seemed to be degraded, since only a few of the detected compounds were found in this case. This might be attributed to further polymerization of the oligomers produced in the first hours of the reaction, and their subsequent precipitation from the reaction mixture.
Discussion
In most fungal genomes, many copies of laccase-encoding genes are present, forming laccase multigene families. In the genome of P. ostreatus, 12 laccase genes have been detected, but only 6 of them have been isolated and characterized [8, 9]. Therefore, there is still a significant knowledge gap on the minor laccase enzymes produced by Pleurotus species, which nonetheless contribute in a major way to the decomposition of complex plant materials. A study regarding P. ostreatus genome identified 3 laccase gene groups [10], while the isolated laccase isozymes showed high variability in terms of ABTS oxidation and malachite green decolorization. The same study revealed, apart from the existence of major laccase isozymes with potent dye decolorization activity, the presence of minor laccase isozymes. The genes lacc2 and lacc11, and the corresponding enzymes, were considered ‘defective’ laccases by the authors, due to the absence of the characteristic HXHG and copper-binding HCH motifs, respectively. Lacc2 (POXA3) and Lacc11 did not decolorize malachite green, and their expression in the tested conditions was low, although Castanera et al. [9] reported enhanced expression of Lacc2 in wheat straw-based media. Consequently, their biochemical properties and physiological role were not examined, although their presence in P. ostreatus transcriptome is established. The laccase system of P. citrinopileatus seems to be similar to P. ostreatus, since the two laccases isolated from the supernatant of P. citrinopileatus cultures accounted for 50% of total laccase activity. Furthermore, PcLac1 was found to be similar to LACC2 from P. ostreatus, and PcLac2 was also found to be similar to another P. ostreatus laccase. The relatively low E0′ and narrow substrate range of PcLac1 together with the previously studied biochemical properties of LACC2 of P. ostreatus indicate that the laccase-based ligninolytic system of the two Pleurotus species has many similarities.
In our previous work regarding the degradation of phenolic compounds from OMWW by P. citrinopileatus, we obtained laccase zymograms on both SDS-PAGE and IEF gels with large smear bands, possibly corresponding to multiple laccase enzymes [33]. In the present study, we aimed at the isolation and characterization of laccase isozymes during the growth of the fungus in OMWW. The fractionation of the culture supernatant at the selected conditions resulted in the isolation of two separate peaks with laccase activity. The corresponding enzymes were named PcLac1 and PcLac2. Both enzymes correspond to slightly over 50% of the total laccase activity of the crude supernatant, indicating the existence of other laccases, which were not isolated by the selected chromatographic conditions (Additional file 1: Table S1).
The molecular mass of both PcLac1 and PcLac2 lies within the range of most fungal laccases [3]. They both have acidic pH optima, but PcLac1 was found more thermostable and pH-stable than PcLac2. In addition, the substrate range of the studied laccases shows some interesting features. Both laccases were able to oxidize ortho- and para-substituted phenols (catechol and hydroquinone), but not meta- substituted phenols (resorcinol), similarly to what was reported for many microbial laccases [1]. The existence of ortho- or para- substituents facilitates the electron abstraction from the phenoxy-OH, leading to the formation of the respective radical. As shown previously [35], the apparent Km of laccases is the result of both the enzyme’s affinity for the substrate and the substrate electric properties, such as the redox potential and the ionization energy [1]. An interesting observation regarding the substrate preference of the studied laccases is that they both show higher specific activity for 2,6-DMP than the mono-methoxy- substituted guaiacol. This is unusual for fungal laccases, since guaiacol is considered one of the traditional substrates. In this respect, PcLac1 and PcLac2 are more similar to bacterial laccases, which lack the ability to oxidize guaiacol [1]. Both laccases can oxidize hydroxycinnamic acids, except p-coumaric acid, but only PcLac2 can oxidize hydroxybenzoic acids, such as vanillic and gallic acid. This is an interesting observation, since most known fungal laccases have the ability to oxidize gallic acid. Together with the fact that PcLac2 is able to oxidize vanillic acid, but not vanillin, we may assume the presence of a residue interacting with the carboxyl group, corresponding to Asn264 of T. versicolor laccase [35], and thus facilitating the electron abstraction from substrates bearing the –COOH group. The same authors attributed the oxidation of substrates bearing ortho- substituted methoxy or hydroxyl groups, to the hydrogen bonds formed with His458 and Asp206 in T. versicolor laccase, which stabilize the substrate and facilitate electron abstraction. The kinetic constants calculated for PcLac1 and PcLac2 are also in the range reported for other fungal laccases [3], and specifically for Pleurotus species. Regarding ABTS oxidation, PcLac2 showed a similar Km value to Lacc6 from P. ostreatus [10], while the Km calculated for PcLac1 on ABTS was similar to two laccases isolated from P. ostreatus by Tinoco et al. [36]. These biochemical properties are in accordance with the proteomics data, which illustrate the similarity of P. citrinopileatus laccase system with the corresponding enzymatic system of P. ostreatus.
Regarding the effect of various inhibitors on the activity of PcLac1 and Pclac2, the results were similar to previously described fungal laccases. The addition of low copper concentrations enhanced PcLac1 activity, but no effect was observed on the activity of PcLac2. While many studies reported no effect on laccase activity in the presence of low copper concentrations [20], others reported a certain degree of activity enhancement [37]. This is attributed to the variable architecture of laccase molecules, allowing a certain degree of mobility to the copper ions for some laccases, as in the case of Coprinus cinereus laccase, which is able to lose reversibly the Cu ions in the T2 copper center [38]. Nonetheless, increasing Cu concentration above a certain point usually inhibits laccase activity [20], similarly to PcLac1 and PcLac2. On the other hand, NaN3 strongly inhibited the activity of PcLac1 and PcLac2, similarly to most known multicopper oxidases [12, 20]. NaN3 binds to the copper atoms of laccases, disrupting the electron transfer.
Regarding the effect of organic solvents, PcLac2 was found to be significantly more stable than PcLac1. The stability of PcLac2 is high in the presence of 10% of most studied solvents, similar to LACC6 from P. ostreatus [20]. On the other hand, PcLac1 was found to be severely inhibited by most organic solvents tested, similarly to most fungal laccases. Solvent stability is an important property for oxidases in biocatalysis applications, since the solubility of most phenolic compounds used as substrates for laccases is limited in aqueous-based media. This is reflected in the efforts made—through genetic engineering—to increase their stability in the presence of organic solvents [39–41].
The formal potential of PcLac1 and PcLac2 was calculated with FtacV. PcLac1 was found to have a higher E0′ than PcLac2, a result that is in agreement with the substrate spectrum. Overall, the electrochemical and biochemical data indicated a rather higher oxidative activity for PcLac1. Temperature variations between 30 and 43 °C did not seem to affect significantly the E0′ of both enzymes. The low-E0′ values measured for both laccases are rather unusual for oxidative enzymes from white-rot fungi, since most low-E0′ laccases reported in the literature are from bacterial or plant sources [1]. Indeed, significant research effort has been dedicated in the quest for high-E0′ oxidative enzymes, due to their outstanding performance in biodegradation and detoxification applications, targeting recalcitrant pollutants. However, their use as biocatalysts for ‘green’ synthetic applications has always been hindered by the formation of multiple products, which creates the need for further costly purification steps. Moreover, the use of laccases as tools for biosensor development suffers from the same issue: their wide substrate specificity results in biosensors with low selectivity against target compounds. The discovery of novel laccases with low E0′ could offer a viable alternative to these issues, by providing an array of enzyme options with variable properties, tailored for specific applications.
Regarding the biocatalytic potential of PcLac1 and PcLac2 enzymes, the bioconversion of ferulic and sinapic acid was studied in 24 h experiments. It should be noted that the stability profiles of both PcLac1 and PcLac2 against 10% (v/v) DMSO enabled the addition of this solvent to the bioconversion reactions, thus allowing the introduction of higher concentrations of the starting monomers. This property can be exploited in ‘green’ synthesis applications, involving substrates with low water solubility. Both enzymes were shown to polymerize the substrates, since the formation of precipitates was observed in all cases at the end of the reaction. The water-soluble reaction products were analyzed with UHPLC–HRMS/MS. The product profiles of the two enzymes presented clear differences, reflecting their difference in redox, biochemical and kinetic properties. The most obvious difference between the two enzymes is that the substrates seem to be polymerized from PcLac1, and therefore, they precipitate out of the reaction in prolonged reaction times.
The study of Slagman et al. [42], regarding the oligomerization of 4-hydroxybenzoic acid by T. versicolor laccase, concluded that not all dimers formed by the action of the enzyme are the same. Some of them can be oxidized by the laccase easier than the original monomer, but other dimers might be less reactive, and thus, they are recovered in higher yields. These conclusions may be applicable in the present study, since the radicals generated by laccase oxidation usually react further in a non-enzymatically controlled fashion. Under this view, PcLac1 and PcLac2 should theoretically show similar product profiles. The greater abundance of phenolic acid oligomers shown by PcLac2 is, therefore, the result of a lower redox potential, since these oligomers are rapidly polymerized by PcLac1, and precipitate out of the reaction. These results might be of great significance for applications involving environmentally friendly synthesis routes for novel compounds with significant biotechnological potential.
Significant research efforts have been dedicated to the biological activity of hydroxycinnamic acids [43–45], fully documenting their antioxidant, anticancer, and radical-scavenging activity. Enzymatic synthesis, if implemented in a controlled fashion, could provide novel bioactive molecules with superior properties. Ferulic acid dimers are already tested as possible therapeutic agents [46]. As regards oxidative biotransformations, most characterized laccases are relatively high-redox enzymes, capable of oxidizing a great variety of phenolic compounds. This property is favorable for many applications, such as biodegradation of xenobiotic compounds, but it can pose many problems in biocatalytic applications, due to the formation of highly oxidized, variable products. It is generally acknowledged that the redox potential plays a major role in the substrate selectivity of laccases [35], although substrate affinity is also a significant property. Therefore, low-E0′ oxidative enzymes could offer an interesting alternative in the biocatalytic synthesis of novel compounds with biological activity. In the present study, the production of novel derivatives of hydroxycinnamic acids was shown, including trimer and tetramer compounds. The general laccase-mediated oxidation scheme of hydroxycinnamic acids is shown in Fig. 5, while the possible oligomerization products previously described in the literature are shown in Additional file 1: Figures S2–S5. Dimerization of hydroxycinnamate acids can lead to the formation of 5′-5′ dimers (Additional file 1: Figure S2, compounds 6 and 10, Additional file 1: Figure S3, compound 6), 5-O-4′ dimers (Additional file 1: Figure S2, compounds 2, 7 and 9, Additional file 1: Figure S3, compound 12), β-5 dimers (from 8, 5′ dimerization, Additional file 1: Figure S2, compound 3, Additional file 1: Figure S3, compound 10), 8-O-4′ dimers (Additional file 1: Figure S3, compound 9), or 8,8′ diacids, which can be transformed to either monolactone or dilactone β–β dimers (Additional file 1: Figure S3, compounds 5, 7 and 8) [47, 48]. Hydroxycinnamate oligomers have been chemically synthesized and/or isolated from plants, such as the ferulic acid trimer with mass of 578, isolated from maize bran (Additional file 1: Figure S4, compound 10; [49]).
Fig. 5.

Laccase-mediated oxidation of hydroxycinnamic acids. The O• radical (a) is formed by the action of laccase, resulting in the intermediate radicals β (b) and 5 (c), while only the radicals that contribute to bond-forming events are shown. The R could be either a hydrogen for ferulic acid, or a methoxy group for sinapic acid. The coupling of these intermediates results in the formation of β-β, β-5 β-Ο-4, 5–5, 5-O-4, and O–O dimers. The O–O coupled dimer is unstable, and thus, it is not found among the products, while for sinapic acid oxidation, C5 is occupied by a methoxy group, and therefore does not participate in bond forming [47, 48]
Studies reporting the enzymatic synthesis of hydroxycinnamate oligomers are sparse. Ward et al. [50] reported the lignin peroxidase-mediated synthesis of three major ferulic acid dimers (Additional file 1: Figure S3, compounds 5, 8, and 10) and two trimers (Additional file 1: Figure S4, compounds 1 and 3), one of which showed the same m/z (552) (Additional file 1: Figure S4, compound 3) with one of the trimers obtained in this study (Table 6, compound 16). The synthesis of one sinapic acid tetramer was previously reported [51] by a plant peroxidase, together with a variety of dimers. The characterized tetramer had m/z 831.25 and molecular formula C43H43O17, which is of different molecular mass than the tetramer reported in this study (Table 7, compound 26). Adelakun et al., [24], also reported the laccase-mediated dimerization of ferulic acid in products with MW of 386 (Additional file 1: Figure S3, compounds 8 and 10), similar to product 3 reported here (Table 6).
Conclusions
In the present work, the discovery, purification, and characterization of two new low-redox potential laccases is described, from the white-rot fungus Pleurotus citrinopileatus. Peptide analysis of the purified proteins revealed similarities with the laccase system of P. ostreatus, but this is the first report of their characterization. The satisfactory stability of both enzymes against different organic solvents allowed the use of DMSO in the biotransformation reactions of ferulic and sinapic acids, resulting in improved solubility of the starting monomers. Both laccases are able to oligomerize ferulic and sinapic acid to different degrees, revealing the importance of the low-redox potential laccases in biocatalytic applications where high selectivity is required. Moreover, to the best of our knowledge, this is the first report regarding the enzymatic production of sinapic acid trimers and tetramers. Further studies are necessary to determine their exact structure and bioactivity, but the importance of low-redox enzymes in synthetic chemistry is hereby demonstrated.
Materials and methods
Chemicals and enzymes
2,2′-Azino-bis-(3-ethylbenzthioazoline-6-sulfonic acid) diammonium salt (ABTS) was from Sigma-Aldrich (St. Louis, MO, USA). Other substrates and chemicals were from Applichem (Darmstadt, Germany) or Sigma-Aldrich, and were of the highest purity available.
Microorganism and growth conditions
The strain P. citrinopileatus LGAM 28684 was previously evaluated in respect to the production of ligninolytic enzymes [52]; it is routinely maintained in Potato Dextrose Agar (PDA; Applichem, Germany), and is stored in the fungal culture collection of the Laboratory of General and Agricultural Microbiology (Agricultural University of Athens). P. citrinopileatus was cultivated in an olive mill wastewater-based liquid medium, as described by Zerva et al. [33]. Olive mill wastewater (OMWW) was obtained from an olive oil mill with a three-phase decanter in Kalamata (Peloponnese, S.W. Greece) and was maintained at -20 °C; OMWW composition and pretreatment process prior to fungal growth were as previously described [33]. P. citrinopileatus was grown in 50% (v/v) OMWW supplemented with 3 g L−1 corn steep liquor as nitrogen source, for 25 days, and then, the supernatant was collected and used for the isolation of laccase enzymes.
Enzyme purification
The culture supernatant was collected by centrifugation, filtrated, and concentrated using an Amicon ultrafiltration device Stirred Cell 8400 (membrane cutoff 10 kDa, Merck Millipore). The concentrate was dialyzed against 20 mM piperazine buffer pH 5.7 at 4 °C overnight, and applied to an equilibrated Q sepharose column with a flow of 4 mL min−1. A linear gradient from 0 to 500 mM NaCl was applied with a Biorad Econo gradient pump. Eluted fractions were analyzed for laccase activity, and selected positive fractions were pooled together, and dialyzed again. A second purification step followed, applying the pooled and dialyzed fractions to a properly equilibrated DEAE-Sepharose column. Elution was performed with a linear gradient from 0 to 500 mM NaCl as previously described. Fractions with laccase activity were pooled, concentrated, and dialyzed against 20 mM Bis–Tris buffer pH 7. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed according to Laemmli [53], with 12.5% polyacrylamide gels.
Proteomic analysis
Protein bands were excised from a Coomassie Brilliant Blue (CBB) stained gel. Gel slices were cut into small 1 mm3 cubes and transferred to 1.5 mL Eppendorf tubes. Proteins in the gel cubes were digested with a standard “in-gel digestion” protocol, as follows; gel pieces were washed twice with 0.1 M NH4HCO3:acetonitrile (1:1) and subsequently incubated in 50 μL of 10 mM DTT in 0.1 M NH4HCO3 (15 min at 56 °C). Afterward, gel pieces were washed with 0.1 M NH4HCO3:acetonitrile (1:1) and incubated with 50 μL of 55 mM iodoacetamide in 0.1 M NH4HCO3 (30 min at room temperature in the dark). Subsequently, the buffer was removed, and gel pieces were washed with 0.1 M NH4HCO3: acetonitrile (1:1) and finally with 100% acetonitrile for 5 min. Gel pieces were dried under vacuum for 20 min and rehydrated in digestion buffer with sequencing grade modified porcine trypsin (Promega; 12.5 ng/µL trypsin in 0.1 M NH4HCO3) at 37 °C overnight. Peptides were extracted by water bath sonication incubating with 50 μL water, adding 50 μL 50% acetonitrile 0.1% formic acid to the gel and incubating 15 min, recover supernatant and pool with first extract. Pooled peptide extracts were dried by SpeedVac and re-dissolved in 20 μL 0.1% formic acid.
From the re-dissolved peptide extract, 5 μL were injected on a nanoLC-MS system (EASY-LCII connected to QexactivePLUS, ThermoScientific, USA). Peptides were trapped on a 2 cm × 0.1 mm C18 trap column, and separated on an 8 cm × 0.75 mm C18 analytical column (PepSep, Denmark) using a flow rate of 300 nL/min. Sample loading and trapping were performed in buffer A (0.1% formic acid in water), and elution was performed with a 20 min gradient going from 2 to 30% buffer B (0.1% formic acid in acetonitrile), followed by column regeneration at 80% buffer B and re-equilibration at 2% buffer B. The nanoLC eluate was directly sprayed into the source of Qexactive by Flex-ion nanospray, using PepSep nanospray needle at 2.3 kV ESI potential. MS acquisition was performed using a DDA method with alternating MS1 scan at resolution 70,000 profile mode, AGC target 3e6, maxIT 50 ms, scan range 500–1400 m/z, and subsequently 8 MS2 scans centroid mode, resolution 17500 AGC target 5e4, maxIT100 ms, with isolation window 1.6 m/z at NCE = 28 on with preferred peptide match ions of charges 2, 3, or 4 and a dynamic exclusion window of 30 s. Protein identification was performed using the database matching algorithm of Byonic (ProteinMetrics, Cupertino, USA) using either the Uniprot entries of Pleurotus (taxon 5320, downloaded 8nov 2019).
Enzymatic assays
Laccase activity was routinely assayed with ABTS as the substrate (2 mM, ε420 = 36,000 M−1 cm−1), in 50 mM phosphate—citrate buffer pH 4 at 35 °C. The reaction was monitored for 10 min in a Spectra Max 250 microplate reader (Molecular Devices, CA, USA). The ability of both enzymes to oxidize different substrates was explored in 24 h reactions. The UV/Vis spectra (250–750 nm) of the reactions and their respective controls with heat-inactivated enzyme were recorded, to determine differences in absorbance maxima. Substrate specificity assays were performed by measuring laccase activity against a variety of substrates (2 mM), including catechol (ε450 = 928 M−1 cm−1), 2,6-dimethoxyphenol (2,6-DMP, ε469 = 27,500 M−1 cm−1), pyrogallol (ε420 = 3724 M−1 cm−1), guaiacol (ε456 = 12,100 M−1 cm−1), and hydroquinone (ε247 = 21,300 M−1 cm−1). One unit of enzymatic activity was defined as the amount of enzyme oxidizing 1 μmol of substrate per min. Kinetic measurements of laccase activity were performed for ABTS and 2,6-DMP, in 50 mM phosphate-citrate buffer pH 4. Michaelis–Menten constants were calculated by the respective nonlinear regression curves using the software GraphPad Prism 5 (GraphPad Software, Inc., U.S.A.).
Determination of pH and temperature optima and thermal stability: biochemical characterization
The effect of pH and temperature on enzyme activity and stability was measured using ABTS (2 mM) as the substrate. The optimum pH was calculated by measuring the activity at 40 °C after incubation for 10 min over the pH range 2.0–9.0 using as buffers 0.1 M phosphate (pH 2.0), 0.1 M citrate–phosphate (pH 3.0–5.0), 0.1 M phosphate (pH 6.0–7.0), and 0.1 M Tris–HCl (pH 8.0–9.0). The optimum temperature was calculated by assaying laccase activity over the range of 20–70 °C for 10 min in 0.1 M citrate–phosphate buffer, pH 3 for PcLac1, and pH 4 for PcLac2. The thermostability was estimated after incubation of the purified enzymes at temperatures ranging from 30 to 70 °C and pH 7.0, for selected time intervals, and calculating the residual activity under standard assay conditions. The effects of various inhibitors and organic solvents on the laccase activity were determined by adding the compound at the indicated concentration in the assay mixture and by measuring the residual activity under standard assay conditions.
Determination of formal potential
FTacV experiments [54] were performed in a single compartment three-electrode cell consisting of a disk glassy carbon electrode (GC) with a surface of 0.785 mm as a working electrode, a 1.6 mm-diameter Pt-coated titanium rod as a counter electrode, and an Ag|AgCl, KCl sat. reference electrode (+ 0.197 V vs NHE) [34]. The aqueous solution of about 3 mL consisted of 100 mM tartrate buffer (pH 4) (PENTA) as a supporting electrolyte.
Voltammetric measurements were performed by a PAR 263A Potentiostat connected to an AFG 5101 Tektronix programmable arbitrary function generator. All solutions were deaerated for at least 10 min before the experiments to avoid oxygen reduction on the electrode surface. Nitrogen gas was purged over the solution during measurement, as well. The temperature of the cell was kept steady using a thermostated bath (FALC WB-MD5). The cell temperature was recorded during the measurement.
The glassy carbon electrode was polished on a cloth with the use of 0.3 μm and 0.05 μm Al2O3. Then, washing of the electrode and sonication (5 min) was performed in distilled water. After the sonication, the electrode was washed again with distilled water. An enzyme volume of 1 μL was left on the surface of the electrode to dry for about 15 min at room temperature. The procedure was repeated once more to increase the enzyme surface concentration on the electrode. Subsequently, 1 μL of Nafion was left to dry above the deposited enzyme to complete the immobilization [55, 56]. All the formal potentials were corrected by taking into account the temperature coefficient − 1.01 mV/°C of the Ag/AgCl, KCl sat. reference electrode [57].
Biocatalytic transformation of phenolic acids
For the laccase-mediated biotransformation of phenolic acids, 25 mg of either ferulic or sinapic acid were dissolved in 100 mM citrate–phosphate buffer (pH 4 for PcLac1 and 5 for PcLac2), containing 7.5% (v/v) DMSO. The reaction started with the addition of 1.5 Units of each enzyme to a total volume of 8 mL. Reaction mixtures were incubated in a rotary shaker at 35 °C, 200 rpm for 24 h. At selected time intervals (3, 8, and 24 h), samples were collected and were immediately frozen until further analysis.
Analytical methods
The monitoring of the products of all enzymatic reactions was performed on an UHPLC–HRMS/MS Q-Exactive Orbitrap (Thermo) platform at negative ionization mode. The analytical methodology was optimized for the better separation of the products on a UPLC C18 (2.1 × 150 mm, 1.9 μm) reversed phased column. The gradient phase consisted of solvents A: aqueous 0.1% (v/v) formic acid and B: acetonitrile. The gradient elution was: T = 0 min, 5% B; T = 1 min, 5% B, T = 14 min, 95% B, T = 15 min, 95% B, T = 15.1 min, 5% B; T = 16 min, 5% B. The flow rate was 0.350 mL/min and the injection volume was 5 μL. The column temperature was kept at 40 °C, while the sample tray temperature was set at 4 °C. The HRMS data were acquired at a full-scan mode with mass range 120–1200 m/z. The ionization conditions were as set as follows: capillary temperature, 350 °C; spray voltage, 2.7 kV; S-lense Rf level, 50 V; sheath gas flow, 40 arb. units; aux gas flow, 5 arb. units; aux. gas heater temperature, 50 °C. For the full-scan experiments, the resolution was 70,000. The data-dependent acquisition capability has been also used at 17,500 resolution, allowing for MS/MS fragmentation of the three most intense ions of every peak exceeding the predefined threshold applying a 10 s dynamic exclusion. Normalized collision energy was set at 35%. Data acquisition and analysis have been completed employing Xcalibur (Thermo).
Statistical analysis
All results are expressed as the mean value of three replicates. Error bars represent the standard deviation. Statistical analysis was performed with SigmaPlot v.12.5 software package (Systat Software, Inc., San Jose, CA, USA).
Supplementary Information
Additional file 1. Figure S1. 3rd Harmonic of (a) PcLac1 and (b) PcLac2. Experimental conditions: deaerated tartrate buffer pH 4.0 100 mM, T 43oC, v 50 mV/s, A 180 mV and f 6 (black), 9 (red), 12 (green) and 16 (blue) Hz. Figure S2. Possible products from ferulic acid dimerization. Products shown were previously reported [1–4]. Figure S3. Possible products from ferulic acid dimerization. Products shown were previously reported [1,2,4,5]. Figure S4. Possible products from ferulic acid trimerization. Products shown were previously reported [6–9]. Figure S5. Possible products from sinapic acid dimerization. Products shown were previously reported [4,5,10–12].
Additional file 2. Proteomic analysis of PcLac1 and PcLac2.
Acknowledgements
Not applicable.
Authors’ contributions
AZ: methodology, visualization, and writing—original draft preparation; CP: investigation and visualization; AT: investigation and formal analysis; SKB: investigation; AHPA: investigation and visualization; DZ: investigation and visualization; AK: visualization and formal analysis; GIZ: resources; ET: conceptualization and supervision. All authors contributed to the writing of the manuscript and edited the final version. All authors read and approved the final manuscript.
Funding
This project was supported by the project “INSPIRED-The National Research Infrastructures on Integrated Structural Biology, Drug Screening Efforts and Drug target functional characterization” (MIS 5002550) which is implemented under the Action “Reinforcement of the Research and Innovation Infrastructure”, funded by the Operational Programme “Competitiveness, Entrepreneurship and Innovation” (NSRF 2014–2020) and co-financed by Greece and the European Union (European Regional Development Fund).
Availability of data and materials
All data supporting the conclusions of this article are included in the manuscript and its additional files. Samples of materials produced in the current work are available from the corresponding author upon reasonable request.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Reiss R, Ihssen J, Richter M, Eichhorn E, Schilling B, Thöny-Meyer L. Laccase versus laccase-like multi-copper oxidase: a comparative study of similar enzymes with diverse substrate spectra. PLoS ONE. 2013;8:65633. doi: 10.1371/journal.pone.0065633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kaur K, Sharma A, Capalash N, Sharma P. Multicopper oxidases: Biocatalysts in microbial pathogenesis and stress management. Microbiol Res. 2019;222:1–13. doi: 10.1016/j.micres.2019.02.007. [DOI] [PubMed] [Google Scholar]
- 3.Baldrian P. Fungal laccases—occurrence and properties. FEMS Microbiol Rev. 2006;30:215. doi: 10.1111/j.1574-4976.2005.00010.x. [DOI] [PubMed] [Google Scholar]
- 4.Riva S. Laccases: blue enzymes for green chemistry. Trends Biotechnol. 2006;24:219. doi: 10.1016/j.tibtech.2006.03.006. [DOI] [PubMed] [Google Scholar]
- 5.Glazunova OA, Trushkin NA, Moiseenko K V, Filimonov IS, Fedorova TV. Catalytic efficiency of basidiomycete laccases: redox potential versus substrate-binding pocket structure. Catal. 2018.
- 6.Kilaru S, Hoegger PJ, Kües U. The laccase multi-gene family in Coprinopsis cinerea has seventeen different members that divide into two distinct subfamilies. Curr Genet. 2006;50:45. doi: 10.1007/s00294-006-0074-1. [DOI] [PubMed] [Google Scholar]
- 7.Glazunova OA, Moiseenko KV, Kamenihina IA, Isaykina TU, Yaropolov AI, Fedorova TV. Laccases with variable properties from different strains of Steccherinum ochraceum: does glycosylation matter? Int J Mol Sci. 2019;20:2008. doi: 10.3390/ijms20082008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pezzella C, Lettera V, Piscitelli A, Giardina P, Sannia G. Transcriptional analysis of Pleurotus ostreatus laccase genes. Appl Microbiol Biotechnol. 2013;97:705. doi: 10.1007/s00253-012-3980-9. [DOI] [PubMed] [Google Scholar]
- 9.Castanera R, Pérez G, Omarini A, Alfaro M, Pisabarro AG, Faraco V, et al. Transcriptional and enzymatic profiling of Pleurotus ostreatus laccase genes in submerged and solid-state fermentation cultures. Appl Environ Microbiol. 2012;78:4037–4045. doi: 10.1128/AEM.07880-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Park M, Kim M, Kim S, Ha B, Ro HS. Differential expression of laccase genes in Pleurotus ostreatus and biochemical characterization of laccase isozymes produced in Pichia pastoris. Mycobiology. 2015;43:280–287. doi: 10.5941/MYCO.2015.43.3.280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kumar SVS, Phale PS, Durani S, Wangikar PP. Combined sequence and structure analysis of the fungal laccase family. Biotechnol Bioeng. 2003;83:386–394. doi: 10.1002/bit.10681. [DOI] [PubMed] [Google Scholar]
- 12.Zerva A, Koutroufini E, Kostopoulou I, Detsi A, Topakas E. A novel thermophilic laccase-like multicopper oxidase from Thermothelomyces thermophila and its application in the oxidative cyclization of 2′,3,4-trihydroxychalcone. N Biotechnol. 2019;49:10. doi: 10.1016/j.nbt.2018.12.001. [DOI] [PubMed] [Google Scholar]
- 13.Chen DM, Bastias BA, Taylor AFS, Cairney JWG. Identification of laccase-like genes in ectomycorrhizal basidiomycetes and transcriptional regulation by nitrogen in Piloderma byssinum. New Phytol. 2003;157. [DOI] [PubMed]
- 14.Levasseur A, Saloheimo M, Navarro D, Andberg M, Pontarotti P, Kruus K, et al. Exploring laccase-like multicopper oxidase genes from the ascomycete Trichoderma reesei: a functional, phylogenetic and evolutionary study. 2010. http://genome.jgi-psf. [DOI] [PMC free article] [PubMed]
- 15.Tamayo-Ramos JA, van Berkel WJ, de Graaff LH. Biocatalytic potential of laccase-like multicopper oxidases from Aspergillus niger. Microb Cell Fact. 2012;11:165. doi: 10.1186/1475-2859-11-165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fan X, Zhou Y, Xiao Y, Xu Z, Bian Y. Cloning, expression and phylogenetic analysis of a divergent laccase multigene family in Auricularia auricula-judae. Microbiol Res. 2014;169:453. doi: 10.1016/j.micres.2013.08.004. [DOI] [PubMed] [Google Scholar]
- 17.Vasina DV, Mustafaev ON, Moiseenko KV, Sadovskaya NS, Glazunova OA, Tyurin AA, et al. The Trametes hirsuta 072 laccase multigene family: genes identification and transcriptional analysis under copper ions induction. Biochimie. 2015;116:154. doi: 10.1016/j.biochi.2015.07.015. [DOI] [PubMed] [Google Scholar]
- 18.Osma JF, Toca-Herrera JL, Rodríguez-Couto S. Uses of laccases in the food industry. Enzyme Res. 2010;2010:918761. doi: 10.4061/2010/918761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yang J, Li W, Ng TB, Deng X, Lin J, Ye X. Laccases: production, expression regulation, and applications in pharmaceutical biodegradation. Front Microbiol. 2017;8:832. doi: 10.3389/fmicb.2017.00832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhuo R, Zhang J, Yu H, Ma F, Zhang X. The roles of Pleurotus ostreatus HAUCC 162 laccase isoenzymes in decolorization of synthetic dyes and the transformation pathways. Chemosphere. 2019;234:733–745. doi: 10.1016/j.chemosphere.2019.06.113. [DOI] [PubMed] [Google Scholar]
- 21.Wang J, Huang R, Qi W, Su R, Binks BP, He Z. Construction of a bioinspired laccase-mimicking nanozyme for the degradation and detection of phenolic pollutants. Appl Catal B Environ. 2019;254.
- 22.Jasińska A, Góralczyk A, Soboń A, Długoński J. Novel laccase-like multicopper oxidases from the fungus Myrothecium roridum—production enhancement, identification and application in dyes removal. Acta Biochim Pol. 2018;65:287. doi: 10.18388/abp.2017_2546. [DOI] [PubMed] [Google Scholar]
- 23.Copete LS, Chanagá X, Barriuso J, López-Lucendo MF, Martínez MJ, Camarero S. Identification and characterization of laccase-type multicopper oxidases involved in dye-decolorization by the fungus Leptosphaerulina sp. BMC Biotechnol. 2015;15:74. doi: 10.1186/s12896-015-0192-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Adelakun OE, Kudanga T, Parker A, Green IR, le Roes-Hill M, Burton SG. Laccase-catalyzed dimerization of ferulic acid amplifies antioxidant activity. J Mol Catal B Enzym. 2012;74:29–35. http://www.sciencedirect.com/science/article/pii/S1381117711002359
- 25.Sagui F, Chirivì C, Fontana G, Nicotra S, Passarella D, Riva S, et al. Laccase-catalyzed coupling of catharanthine and vindoline: an efficient approach to the bisindole alkaloid anhydrovinblastine. Tetrahedron. 2009;65.
- 26.Piscitelli A, Amore A, Faraco V. Last Advances in Synthesis of Added Value Compounds and Materials by Laccasemediated Biocatalysis. Curr Org Chem. 2012;16.
- 27.Aljawish A, Chevalot I, Jasniewski J, Revol-Junelles A-M, Scher J, Muniglia L. Laccase-catalysed functionalisation of chitosan by ferulic acid and ethyl ferulate: Evaluation of physicochemical and biofunctional properties. Food Chem. 2014;161:279. doi: 10.1016/j.foodchem.2014.03.076. [DOI] [PubMed] [Google Scholar]
- 28.Lim J, Sana B, Krishnan R, Seayad J, Ghadessy FJ, Jana S, et al. Laccase-catalyzed synthesis of low-molecular-weight lignin-like oligomers and their application as UV-blocking materials. Chem Asian J. 2018;13:284. doi: 10.1002/asia.201701573. [DOI] [PubMed] [Google Scholar]
- 29.Reano AF, Pion F, Domenek S, Ducrot P-H, Allais F. Chemo-enzymatic preparation and characterization of renewable oligomers with bisguaiacol moieties: promising sustainable antiradical/antioxidant additives. Green Chem. 2016;18.
- 30.López J, Hernández-Alcántara JM, Roquero P, Montiel C, Shirai K, Gimeno M, et al. Trametes versicolor laccase oxidation of gallic acid toward a polyconjugated semiconducting material. J Mol Catal B Enzym. 2013;97.
- 31.Pandey VK, Singh MP, Srivastava AK, Vishwakarma SK, Takshak S. Biodegradation of sugarcane bagasse by Pleurotus citrinopileatus. Cell Mol Biol. 2012;58:8–14. [PubMed] [Google Scholar]
- 32.Kulshreshtha S, Mathur N, Bhatnagar P, Kulshreshtha S. Cultivation of Pleurotus citrinopileatus on handmade paper and cardboard industrial wastes. Ind Crops Prod. 2013;41.
- 33.Zerva A, Zervakis GI, Christakopoulos P, Topakas E. Degradation of olive mill wastewater by the induced extracellular ligninolytic enzymes of two wood-rot fungi. J Environ Manage. 2017;203:791. doi: 10.1016/j.jenvman.2016.02.042. [DOI] [PubMed] [Google Scholar]
- 34.Zouraris D, Karantonis A. Large amplitude ac voltammetry: Chief observables for a reversible reaction of free electroactive species. J Electroanal Chem. 2019;847.
- 35.Mehra R, Muschiol J, Meyer AS, Kepp KP. A structural-chemical explanation of fungal laccase activity. Sci Rep. 2018;8:17285. doi: 10.1038/s41598-018-35633-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tinoco R, Pickard MA, Vazquez-Duhalt R. Kinetic differences of purified laccases from six Pleurotus ostreatus strains. Lett Appl Microbiol. 2001;32:331. doi: 10.1046/j.1472-765X.2001.00913.x. [DOI] [PubMed] [Google Scholar]
- 37.Lorenzo M, Moldes D, Rodríguez Couto S, Sanromán M. Inhibition of laccase activity from Trametes versicolor by heavy metals and organic compounds. Chemosphere. 2005;60:1124. doi: 10.1016/j.chemosphere.2004.12.051. [DOI] [PubMed] [Google Scholar]
- 38.Bukh C, Bjerrum MJ. The reversible depletion and reconstitution of a copper ion in Coprinus cinereus laccase followed by spectroscopic techniques. J Inorg Biochem. 2010;104:1029–37. http://www.sciencedirect.com/science/article/pii/S0162013410001285 [DOI] [PubMed]
- 39.Zumárraga M, Bulter T, Shleev S, Polaina J, Martínez-Arias A, Plou FJ, et al. In Vitro Evolution of a Fungal Laccase in High Concentrations of Organic Cosolvents. Chem Biol. 2007;14:1052–64. http://www.sciencedirect.com/science/article/pii/S107455210700289X [DOI] [PubMed]
- 40.Mate D, Garcia-Ruiz E, Camarero S, Alcalde M. Directed Evolution of Fungal Laccases. Curr Genomics. 2011;12:113. doi: 10.2174/138920211795564322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pardo I, Rodríguez-Escribano D, Aza P, de Salas F, Martínez AT, Camarero S. A highly stable laccase obtained by swapping the second cupredoxin domain. Sci Rep. 2018;8:15669. doi: 10.1038/s41598-018-34008-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Slagman S, Escorihuela J, Zuilhof H, Franssen MCR. Characterization of the laccase-mediated oligomerization of 4-hydroxybenzoic acid. RSC Adv. 2016;6:99367–99375. doi: 10.1039/C6RA23040G. [DOI] [Google Scholar]
- 43.Nićiforović N, Abramovič H. Sinapic acid and its derivatives: natural sources and bioactivity. Compr Rev Food Sci Food Saf. 2014;13:34. doi: 10.1111/1541-4337.12041. [DOI] [PubMed] [Google Scholar]
- 44.Mura F, Silva T, Castro C, Borges F, Zuñiga MC, Morales J, et al. New insights into the antioxidant activity of hydroxycinnamic and hydroxybenzoic systems: Spectroscopic, electrochemistry, and cellular studies. Free Radic Res. 2014;48:1473. doi: 10.3109/10715762.2014.965702. [DOI] [PubMed] [Google Scholar]
- 45.Damasceno SS, Dantas BB, Ribeiro-Filho J, Antônio M, Araújo D, Galberto M, da Costa JK. Chemical properties of caffeic and ferulic acids in biological system: implications in cancer therapy. A review. Curr Pharm Des. 2017;23:3015. doi: 10.2174/1381612822666161208145508. [DOI] [PubMed] [Google Scholar]
- 46.Priebe A, Hunke M, Tonello R, Sonawane Y, Berta T, Natarajan A, et al. Ferulic acid dimer as a non-opioid therapeutic for acute pain. J Pain Res. 2018;11:1075. doi: 10.2147/JPR.S161161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Grúz J, Pospíšil J, Kozubíková H, Pospíšil T, Doležal K, Bunzel M, et al. Determination of free diferulic, disinapic and dicoumaric acids in plants and foods. Food Chem. 2015;171:280–286. doi: 10.1016/j.foodchem.2014.08.131. [DOI] [PubMed] [Google Scholar]
- 48.Carunchio F. Oxidation of ferulic acid by laccase: identification of the products and inhibitory effects of some dipeptides. Talanta. 2001;55:189. doi: 10.1016/S0039-9140(01)00417-9. [DOI] [PubMed] [Google Scholar]
- 49.Bunzel M, Ralph J, Funk C, Steinhart H. Structural elucidation of new ferulic acid-containing phenolic dimers and trimers isolated from maize bran. Tetrahedron Lett. 2005;46:5845–50. http://www.sciencedirect.com/science/article/pii/S0040403905014218
- 50.Ward G, Hadar Y, Bilkis I, Konstantinovsky L, Dosoretz CG. Initial steps of ferulic acid polymerization by lignin peroxidase. J Biol Chem. 2001;276:18734–18741. doi: 10.1074/jbc.M009785200. [DOI] [PubMed] [Google Scholar]
- 51.Liu HL, Wan X, Huang XF, Kong LY. Biotransformation of sinapic acid catalyzed by Momordica charantia peroxidase. J Agric Food Chem. 2007;55:1003–1008. doi: 10.1021/jf0628072. [DOI] [PubMed] [Google Scholar]
- 52.Koutrotsios G, Zervakis GI. Comparative examination of the olive mill wastewater biodegradation process by various wood-rot Macrofungi. Biomed Res Int. 2014;2014:482937. doi: 10.1155/2014/482937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Laemmli UK. Cleavage of structural proteins during the assembly of the Head of Bacteriophage T4. Nature. 1970;227:680. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 54.Zouraris D, Karantonis A. Determination of kinetic and thermodynamic parameters from large amplitude Fourier transform ac voltammetry of immobilized electroactive species. J Electroanal Chem. 2020;876:114729. doi: 10.1016/j.jelechem.2020.114729. [DOI] [Google Scholar]
- 55.Muraleedharan MN, Zouraris D, Karantonis A, Topakas E, Sandgren M, Rova U, et al. Effect of lignin fractions isolated from different biomass sources on cellulose oxidation by fungal lytic polysaccharide monooxygenases. Biotechnol Biofuels. 2018;11:296. doi: 10.1186/s13068-018-1294-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Zouraris D, Dimarogona M, Karnaouri A, Topakas E, Karantonis A. Direct electron transfer of lytic polysaccharide monooxygenases (LPMOs) and determination of their formal potentials by large amplitude Fourier transform alternating current cyclic voltammetry. Bioelectrochemistry. 2018;124:149. doi: 10.1016/j.bioelechem.2018.07.009. [DOI] [PubMed] [Google Scholar]
- 57.Sawyer DT, Sobkowiak A, Roberts JL. Electrochemistry for chemists. 2. New York: Wiley; 1995. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1. Figure S1. 3rd Harmonic of (a) PcLac1 and (b) PcLac2. Experimental conditions: deaerated tartrate buffer pH 4.0 100 mM, T 43oC, v 50 mV/s, A 180 mV and f 6 (black), 9 (red), 12 (green) and 16 (blue) Hz. Figure S2. Possible products from ferulic acid dimerization. Products shown were previously reported [1–4]. Figure S3. Possible products from ferulic acid dimerization. Products shown were previously reported [1,2,4,5]. Figure S4. Possible products from ferulic acid trimerization. Products shown were previously reported [6–9]. Figure S5. Possible products from sinapic acid dimerization. Products shown were previously reported [4,5,10–12].
Additional file 2. Proteomic analysis of PcLac1 and PcLac2.
Data Availability Statement
All data supporting the conclusions of this article are included in the manuscript and its additional files. Samples of materials produced in the current work are available from the corresponding author upon reasonable request.