

Abstract
The rodent hippocampus generates new neurons throughout life. This process, named adult hippocampal neurogenesis (AHN), is a striking form of neural plasticity that occurs in the brains of numerous mammalian species. Direct evidence of adult neurogenesis in humans has remained elusive, although the occurrence of this phenomenon in the human dentate gyrus has been demonstrated in seminal studies and recent research that have applied distinct approaches to birthdate newly generated neurons and to validate markers of adult-born neurons. Our data point to the persistence of AHN until the 10th decade of human life, as well as to marked impairments in this process in patients with Alzheimer's disease. Moreover, our work demonstrates that the methods used to process and analyze postmortem human brain samples can limit the detection of various markers of AHN to the point of making them undetectable. In this Dual Perspectives article, we highlight the critical methodological aspects that should be strictly controlled in human studies and the robust evidence that supports the occurrence of AHN in humans. We also put forward reasons that may account for current discrepancies on this topic. Finally, the unresolved questions and future challenges awaiting the field are highlighted.
Keywords: adult neurogenesis, controversy, human, methodology, hippocampus, immature neuron
Introduction
Adult neurogenesis is a striking form of plasticity that persists throughout life. The addition of new neurons can potentially rewire an entire neural circuit. How the delicate balance between synaptic plasticity and memory persistence is shaped by an apparently disruptive element, such as an uninstructed newly born neuron, is still an enigma for modern neuroscience. Notably, adult neurogenesis is restricted to selected brain regions, including the neurogenic niches of the hippocampal dentate gyrus (DG) (Altman and Das, 1965) and the ventricular-subventricular zone/olfactory bulb (Lois and Alvarez-Buylla, 1994; Lois et al., 1996; Doetsch et al., 1999). Of these, adult hippocampal neurogenesis (AHN) has attracted particularly great interest because of the involvement of the hippocampus in learning, memory, and mood regulation.
The history of AHN has always been touched by controversy. Indeed, the first reports of AHN by Altman and Das (Altman, 1963; Altman and Das, 1965) were initially disbelieved by the scientific community. In these seminal studies, the authors demonstrated selective incorporation of 3H-thymidine in the adult rat subgranular zone, thereby clearly demonstrating the presence of adult-born proliferating cells in this structure. Subsequent work conducted by the group of Nottebohm (Nottebohm, 1981; Alvarez-Buylla et al., 1988), demonstrating seasonal changes of volume in the song control nuclei of canary birds, provided a physiological perspective to the phenomenon of adult neurogenesis and returned momentum to the field. This was probably the first time that a putative physiological role was attributed to this process (Paton and Nottebohm, 1984). The hypothesis that the addition of new neurons was related to increased plasticity, learning, and memory was embraced with enthusiasm by the field.
The seminal studies by Altman and Das were confirmed by Kaplan and Hinds (1977) using electron microscopy as well as by Stanfield and Trice (1988) using retrograde tracers. Moreover, Nowakowski et al. (1989) and Gould, Cameron, and McEwen (Gould et al., 1992; Cameron and Gould, 1994) pioneered the use of thymidine analogs, such as BrdU, combined with the immunolabeling of various specific markers, to demonstrate the neuronal phenotype of newly generated dentate granule cells in vivo. Subsequently, the group of Alvarez-Buylla demonstrated that newborn dentate granule cells are originated from a population of resident radial glia-like astrocytes (Seri et al., 2001). Gould and collaborators demonstrated that stress is one of the most potent negative regulators of AHN (Cameron and Gould, 1994). By showing that AHN could be modulated extrinsically, these studies paved the way for further development of the field. For instance, Kuhn et al. (1996) showed an age-dependent reduction of AHN rate in rodents, a finding that was later demonstrated in humans (Spalding et al., 2013) and nonhuman primates (Gould et al., 1999). Subsequent studies showed that physical exercise, environmental enrichment, and learning also regulate AHN by promoting the maturation and survival of newly generated cells in rodents (van Praag et al., 1999; Kempermann et al., 2000; van Praag et al., 2000). Throughout the following years, newly generated dentate granule cells were demonstrated to show long-term survival (Kempermann et al., 2003), and the sequential maturation stages encompassed by AHN were precisely defined using the expression of specific cell markers (Kempermann et al., 1997, 2003, 2004; Brandt et al., 2003; Brown et al., 2003; Filippov et al., 2003; Rhodes et al., 2003; Couillard-Despres et al., 2005; Laplagne et al., 2006).
Pioneering work by Luskin et al. (1988) using retroviruses to label newly generated progenitor cells in the rodent ventricular-subventricular zone was complemented by extensive studies of the DG (Seri et al., 2001; van Praag et al., 2002; Zhao et al., 2006; Toni et al., 2007, 2008; Kelsch et al., 2008), and retroviral labeling was combined with electrophysiological recordings (van Praag et al., 2002) and electron microscopy (Toni et al., 2008). Newly generated neurons were found to be much more plastic than their fully mature counterparts (Schmidt-Hieber et al., 2004; Bischofberger, 2007; Ge et al., 2007; Piatti et al., 2011; Marin-Burgin et al., 2012) and eager to learn (or possibly to forget) (Akers et al., 2014), so to speak. Immature dentate granule cells have unique electrophysiological properties (Schmidt-Hieber et al., 2004; Bischofberger, 2007; Marin-Burgin et al., 2012; Heigele et al., 2016; Li et al., 2017) and a distinct innervation pattern (Overstreet Wadiche et al., 2005; Marin-Burgin et al., 2012; Temprana et al., 2015; Luna et al., 2019). Thanks to these unconventional features, these cells profoundly impact hippocampal functioning (Gu et al., 2012; Nakashiba et al., 2012; McAvoy et al., 2016; Anacker and Hen, 2017). Several fascinating papers showed that immature neurons are instructed by mature granule cells (and, as recently suggested, that they also instruct the latter) (Luna et al., 2019) in memory formation and information processing (Schinder and Gage, 2004). In rodents, newly generated neurons not only play important roles in mood regulation (Revest et al., 2009; Snyder et al., 2011; Hill et al., 2015; Anacker et al., 2018), but also are crucial in learning and memory, particularly in pattern separation (Sahay et al., 2011; Tronel et al., 2015; Ishikawa et al., 2016).
AHN in distinct mammalian species
The outstanding refinement and controlled methodology that the field of AHN has reached in rodent studies have not, for evident (technical and ethical) reasons, been achieved in other species. Despite exhaustive examination of AHN in particular mammalian species, systematic studies across the whole mammalian phylogenetic scale have not been conducted to date. Given that only captured wild specimens can be examined for some species, heterogeneity between such studies may increase, not only because the exact age and life history of the animals may be unknown, but also because capture and manipulation of the animals may alter AHN (Chawana et al., 2014; Wiget et al., 2017). For instance, several studies described interspecies differences in the expression of AHN markers in bats (Amrein et al., 2007; Gatome et al., 2010), but the expression of these markers has been proposed to be influenced by post-capture stress (Chawana et al., 2014). Moreover, a detailed description of tissue processing techniques and the inclusion of large numbers of specimens are required for the quantitative and qualitative comparison of adult neurogenesis data between distinct mammalian species and studies.
Despite these difficulties, AHN has been observed in the brains of most of the mammalian species examined to date, including several species of primates. Seminal work using both thymidine analogs and markers of AHN revealed the persistence of adult neurogenesis in various nonhuman primate species, thereby firmly paving the way toward the study of adult neurogenesis in humans (Gould et al., 1999; Kornack and Rakic, 1999; Jabes et al., 2010; Kordower et al., 2010; Kohler et al., 2011; Ngwenya et al., 2015). These studies have revealed that primate and rodent AHN share similarities, but also show marked differences (Miller et al., 2013; Fasemore et al., 2018). Perhaps one of the most remarkable differences, elegantly brought to light by Kohler et al. (2011), is the slower maturation of newly generated cells in nonhuman primates than in rodents. This finding has profound physiological implications since the period characterized by the highest excitability of these cells may be considerably extended in long-living organisms. This prolonged period of higher excitability has been proposed to confer an important evolutionary advantage to these species by permitting increased cognitive flexibility (Kohler et al., 2011; Yuan et al., 2014).
The particular case of human adult neurogenesis
The first study addressing whether AHN occurs in humans was published by Eriksson et al. (1998). Those authors showed the incorporation of BrdU into the adult human DG. Several months after administration, BrdU+ cells coexpressed several neuronal markers, thus demonstrating the presence of a restricted population of adult-born dentate granule cells in humans. The conclusions of that study were supported by Spalding et al. (2013), who used another innovative approach to quantify the number of cells newly generated in the human brain. The method was based on the measurement of the transient increase in the availability of 14C during nuclear assays, as well as on the incorporation of this isotope by dividing cells. Using this technology, sophisticated mathematical models, and the injection of thymidine analogs as a positive control, they corroborated the existence of neurons with a record of division in the adult human brain. These two studies took advantage of the higher postmortem stability of nucleic acids compared with proteins, paving the way for the future development of the field. Over the following years, studies by other groups, mainly based on the use of immunohistochemistry, contributed to building a large body of evidence supporting the occurrence of AHN in humans (Boldrini et al., 2009; Crews et al., 2010; Knoth et al., 2010; Ernst et al., 2014) (for a full list of these studies, see Table 1).
Table 1.
Studies addressing the occurrence of human AHNa
| Study | Methods | Age | Tissue | Markers | Main results | Tissue processing |
|---|---|---|---|---|---|---|
| Eriksson et al., 1998 | IHC | 67-72 yr | Postmortem | BrdU, calbindin, NSE, GFAP, NeuN | Incorporation of BrdU into the adult human DG | 4% PFA for 24 h, sliding microtome sectioning |
| Blümcke et al., 2001 | IHC | 4 GW-46 yr | Biopsy (epilepsy and controls) and postmortem (controls) | Nestin, vimentin, Tuj1, Ki67, MAP1b/5, MAP2a-d, NF-L, NeuN, S100β, GFAP, calbindin, CD68, CD45 | Increased number of Nestin+ and Ki67+ cells in epileptic patients | 4% PFA for 24 h (biopsy) or formalin for > 2 w (autopsies); vibratome sectioning |
| Jin et al., 2004 | IHC, WB | 13-90 yr | Postmortem | DCX, PSA-NCAM, TUC4, NeuroD1 | Increased expression levels of markers of immature neurons in AD patients measured by means of WB | Frozen samples (WB), or PFA 4% (unknown fixation time) plus embedding in paraffin |
| Boekhoorn et al., 2006 | IHC | 63-70 yr (controls), 63-69 yr (AD) | Postmortem | DCX, Ki67, GFAP | Unaltered AHN in presenile AD cases | 10% formalin for 30-646 d; microtome sectioning |
| Manganas et al., 2007 | Functional magnetic spectroscopy | — | Living humans | — | Identification of NSCs in the adult human hippocampus | NA |
| Monje et al., 2007 | IHC | 10 mo-63 yr (controls), 10 mo-61 yr (Cancer) | Postmortem | DCX, Ki67, Olig2, CD68, CD20, CD3 | AHN is abolished by cancer treatment | Undisclosed fixation protocol and duration; embedding in paraffin |
| Liu et al., 2008 | IHC, WB, RT-PCR | 18-78 yr (controls), 13-57 yr (epilepsy) | Biopsy (epilepsy) and postmortem (controls) | DCX, PCNA, MCM2, PSA-NCAM,Tuj1, NeuN, Reelin, calretinin, calbindin | Increased AHN in epilepsy | Perfusion/immersion in 15% formalin (following Waldvogel et al., 2006) |
| Boldrini et al., 2009 | IHC | 17-53 yr (controls), 34-62 yr (MDD), 24-61 yr (MDDT-SSRI), 28-61 yr (MDDT-TCA) | Postmortem | Nestin, Ki67, NeuN, GFAP | Decreased number of progenitor cells with age; increased number of progenitor cells in females; antidepressant treatment increased the number of progenitor cells | Frozen tissue, fixed in 4% PFA for 1 wk and freezing |
| Mattiesen et al., 2009 | IHC | 35-81 yr (controls), 35-85 yr (HIE) | Postmortem | TUC-4, calretinin, PCNA | Increased AHN and apoptosis after hypoxic-ischemic encephalopathy | Formalin fixation (undisclosed time and concentration) and embedding in paraffin |
| Geha et al., 2010 | IHC | 22-35 yr (epileptic), 25-66 yr (control) | Biopsy (epilepsy and controls) and postmortem (controls) | Ki67, MCM2, Tuj1, MAP2, NeuN, calretinin, GFAP, Nestin, vimentin | Fixation affects cycle-related immunostaining; persistence of cycling cells in the adult SGZ; these cells were more abundant in epileptic patients | Formalin-zinc (formol 5%; zinc 3 g/L; sodium chloride 8 g/L) for 3 mo (postmortem) or 16 h (surgical), and embedding in paraffin |
| Lucassen et al., 2010 | IHC | 45-85 yr | Postmortem | MCM2, PH3 | Reduction in the number of MCM2+ cells in depressed patients | 4% formaldehyde for 4-5 wk, and embedding in paraffin |
| Knoth et al., 2010 | IHC, ISH, WB | 11 GW-100 yr | Postmortem | DCX, PCNA, Ki67, MCM2, Sox2, Nestin, TUC4, Tuj1, Prox1, PSA-NCAM, NeuroD1, GFAP, calretinin, NeuN | Murine features of human AHN; mild decrease of AHN with age | Formalin fixation (undisclosed fixation protocol) embedding in paraffin |
| Crews et al., 2010 | IHC, qRT-PCR | 87 ± 4.6 yr (controls), 86.1 ± 1.7 yr (early/moderate AD), 80 ± 1.9 yr (severe AD) | Postmortem | DCX, Sox2 | Reduction in the number of DCX+ and Sox2+ cells in patients with AD | 4% PFA (undisclosed duration); vibratome sectioning |
| D'Alessio et al., 2010 | IHC | 40.1 ± 6 yr (epileptic), 45.8 ± 14 yr (controls) | Biopsy (epilepsy) and postmortem (controls) | DCX | Reduced number of DCX+ cells in patients with epilepsy | Formalin for 5 d (undisclosed concentration), and embedding in paraffin; microtome sectioning |
| Johnson et al., 2011 | IHC | 71-101 yr (controls), 75-87 yr (LBD) | Postmortem | PCNA, DCX, GFAP, Musashi | Decrease in the number of Musashi+ and increase in the number of DCX+ cells in the GCL in patients with Lewy body dementia | Undisclosed fixation protocol; embedding in paraffin; microtome sectioning |
| Perry et al., 2012 | IHC | 81.2 ± 7 yr (AD), 80.9 ± 8.5 yr (controls) | Postmortem | Musashi, Nestin, PSA-NCAM, DCX, Tuj1 | Reduced numbers of Musashi+ cells, increased numbers of Nestin+ and PSA-NCAM+ cells, and unchanged numbers of DCX+ and Tuj1+ cells in patients with AD; the expression of DCX was specifically increased in the GCL | 4% formaldehyde for 4 wk and embedding in paraffin; microtome sectioning |
| Boldrini et al., 2012 | IHC | 41.8 ± 14.6 yr (Control), 43.6 ± 13.3 yr (MD), 38.8 ± 13.8 yr (MD-SSRI), 43.2 ± 17.1 yr (MD-TCA) | Postmortem | Ki67, Nestin | Decreased number of Nestin+ cells with age; SSRI increased the number of Nestin+ cells in the DG of depressed patients | 4% formaldehyde (undisclosed duration) and freezing; freezing microtome sectioning |
| Epp et al., 2013 | IHC | 46.8 ± 3.49 yr (Control), 42.83 ± 2.92 yr (MD), 41.5 ± 3.47 yr (MD-Psy) | Postmortem | DCX, p21, NeuN | Increased number of DCX+ cells in depressed patients | Frozen tissue, fixed for 15 min in 4% formaldehyde |
| Spalding et al., 2013 | 14C dating, IHC | 19-92 yr | Postmortem | — | Mild decrease in the AHN rate with aging | Frozen tissue; undisclosed fixation protocol |
| Gómez-Nicola et al., 2014 | IHC | 20-34 yr (Creutzfeldt-Jakob), 58-76 yr (AD), 20-35 yr or 58-79 yr (controls) | Postmortem | Ki67, Sox2, calretinin | Increased number of Ki67+ and calretinin+ cells in patients with Creutzfeldt-Jakob and ADs; decrease in these cell populations in aged controls | Formalin fixation (undisclosed fixation protocol) and embedding in paraffin |
| D'Alessio et al., 2015 | IHC | 22-60 yr (controls), 22-51 yr (TLE) | Biopsy (epilepsy) and postmortem (controls) | Nestin | Reduced number of Nestin+ cells in patients with epilepsy | Formalin (undisclosed concentration) for 5 d, and embedding in paraffin; microtome sectioning |
| Bayer et al., 2015 | IHC | 17-45 yr | Postmortem | Musashi, Nestin, Ki67, calretinin, GFAP, NeuN, Tuj1, DCX | Reduced number of Musashi+ and Nestin+ progenitor and proliferating cells in heroin-addict patients | 4% formaldehyde for 48 h-5 yr and embedding in paraffin; microtome sectioning |
| Dennis et al., 2016 | IHC | 0.2-59 yr | Postmortem | Ki67, DCX, Tuj1, Olig2, EGFR, GFAP, PCNA | Marked decrease in proliferation with age; only microglial cells were identified among the proliferating cell population | 15%-20% formalin for 2-3 wk, and embedding in paraffin |
| Allen et al., 2016 | IHC | 21-81 yr (controls), 55–75 yr (schizophrenia) | Postmortem | Ki67, NeuN | Reduced number of Ki67+ cells in patients with schizophrenia | Undisclosed fixation protocol |
| Le Maître et al, 2018 | IHC | 24-78 yr | Postmortem | NeuN, DCX, Sox2, Ki67 | Reduction in AHN markers in alcohol addicts | Flash-frozen tissue, fixed in 4% formaldehyde for 15-30 min |
| Mathews et al., 2017 | qPCR, IHC | 18-88 yr | Postmortem | Ki67, DCX, GFAP, Tbr2 | Reduced mRNA expression of DCX and Ki67 with aging | Frozen tissue (qPCR); fixation in formalin (undisclosed fixation protocol) and embedding in paraffin (IHC) |
| Liu et al., 2018 | IHC | 12 GW-76 yr | Biopsy (epilepsy) and postmortem (epilepsy and controls) | Nestin, DCX, Mushashi, Tuj1, NeuN, GFAP, MAP2, Olig2, MCM2 | Increased densities of Nestin+ cells in patients with epilepsy | Formalin fixation (undisclosed fixation protocol) and embedding in paraffin |
| Cipriani et al., 2018 | IHC | 13 GW-72 yr (controls), 74-89 yr (AD) | Postmortem | Nestin, GFAP, DCX, Ki67, MCM2, Sox2, Pax6, Tbr2, vimentin, Tuj1 | Sharp reduction of radial glia-like, proliferative, and DCX+ cells in the adult human DG | Frozen tissue was fixed in 4% PFA (undisclosed duration); other tissues were fixed in formalin (undisclosed duration) embedded in paraffin |
| Sorrells et al., 2018 | IHC, EM | 14 GW-77 yr | Biopsy (epilepsy) and postmortem (controls) | Ascl1, BLBP, DCX, BrdU, GFAP, Hopx, Ki67, MCM2, Nestin, NeuN, NeuroD, Olig2, Pax6, Prox1, PSA-NCAM, Sox1, Sox2, Tbr2, Tuj1, vimentin | Absence of AHN markers in the adult human DG | Perfusion with 4% PFA, immersion in either 4% PFA or 10% formalin (undisclosed duration); additional fixation in 4% PFA for 2 d |
| Boldrini et al., 2018 | IHC | 14-79 yr | Postmortem | PSA-NCAM, Sox2, Nestin, Ki67, DCX, NeuN, GFAP, BLBP | Preserved numbers of progenitor and immature cells throughout aging | Flash-frozen tissue, fixed in 4% PFA (undisclosed duration) |
| Tartt et al., 2018 | IHC, RNAscope | 19-67 yr | Postmortem | DCX, NF, Sox2, PSA-NCAM, NeuN | Detection of AHN markers by means of RNAscope | Flash-frozen tissue and short fixation (undisclosed fixation protocol) |
| Stepień et al., 2018 | IHC | 70 ± 6.03 yr (nonhemorragic stroke), 64.75 ± 12.23 yr (hemorrhagic), 64 ± 10.95 yr (controls) | Postmortem | GFAP, PH3 | Increased number of PH3+ cells in patients with hemorrhagic stroke | Undisclosed fixation method and embedding in paraffin; rotary microtome sectioning |
| Moreno-Jiménez et al., 2019 | IHC | 43-97 yr | Postmortem | DCX, PH3, GFAP, Prox1, Tau, NeuN, calretinin, calbindin, Tuj1, PSA-NCAM | Persistence of AHN markers in neurologically healthy control subjects, mild decrease in AHN markers with aging, and sharp decrease in AD patients | 4% PFA for 24 h and vibratome sectioning |
| Tobin et al., 2019 | IHC | 79-99 yr | Postmortem | DCX, PCNA, Nestin, Sox2, Ki67 | Persistence of AHN markers during aging and AD patients; correlation between AHN and cognitive scores | Undisclosed fixation method and embedding in paraffin |
| Seki et al., 2019 | IHC | 9-49 yr | Biopsy | PSA-NCAM, Ki67, HuB, DCX | Reduced numbers of Ki67+/HuB+/DCX+ cells in the adult DG | 4% PFA for 3 d and freezing |
| Flor-García et al., 2020 | IHC | 43-97 yr | Postmortem | DCX, NeuN, PSA-NCAM, MAP2, calbindin, Prox1, calretinin, PH3, GFAP, Tuj1, Tau | Reduced expression of NeuN in DCX+ compared with fully mature DGCs | 4% PFA for 24 h and vibratome sectioning |
aFor each study, the first author's last name, journal, year of publication, methodological approach, age of the subjects studied, type of samples, markers analyzed, main results obtained, and tissue processing protocol are indicated. As shown, most of these studies used postmortem human hippocampal samples and immunohistochemistry. PFA, Paraformaldehyde; IHC, Immunohistochemistry; ISH, in situ hydridization; RT-PCR, real time-polymerase chain reaction; qPCR, quantitative polymesare chain reaction; WB, Western blot; SGZ, subgranular zone; EM, electron microscopy; GCL, granule cell layer; GW, gestational week; MDD, major depression; MDDT-SSRI, MD patients treated with selective serotonin reuptake inhibitors; MDDT-TCA, MD patients treated with tricyclic antidepressants; MD-Psy, MD patients with psychotic symptoms; HIE, hypoxic-ischemic encephalopathy; LBD, Lewy body dementia; TLE, temporal lobe epilepsy; NSE, neuron-specific enolase; NeuN, neuronal nuclei; PCNA, proliferating cell nuclear antigen; MCM2, minichromosome maintenance complex component 2; Sox2, SRY (sex determining region Y)-box 2; Olig2, oligodendrocyte transcription factor; EGFR, epidermal growth factor receptor; Tbr2, T-box brain protein 2; Pax6, paired box protein 6; Ascl1, achaete-scute family BHLH transcription factor 1; BLBP, brain lipid-binding protein; NF-L, neurofilament light chains; PH3, phosphorylated histone 3; MAP2, microtubule-associated protein 2; and MAP1, microtubule-associated protein 1; NF, Neurofilament; NSE, Neuron-Specific Enolase; GFAP, Glial fibrillary acidic protein; DCX, Doublecortin; PSA-NCAM, Polysialylated-neural cell adhesion molecule; Prox1, Prospero homeobox protein 1; HuB; ELAV-like protein 2..
Despite the evidence described above, the existence of human AHN has been recently challenged. Dennis et al. (2016), Sorrells et al. (2018), and Cipriani et al. (2018) reported scarce or absent staining with markers of AHN in the adult human DG, starting from the early second decade of life. Seki et al. (2019) described relatively low numbers of proliferative cells and immature neurons in the adult human DG, thus supporting the previous studies. In contrast, contemporaneous work showed robust persistence of markers of AHN in the human DG throughout the lifetime (Boldrini et al., 2018; Moreno-Jiménez et al., 2019; Tobin et al., 2019). Intriguingly, recent studies that applied apparently equivalent immunohistochemistry methods have drawn opposite conclusions. During 2018 and 2019, these discrepancies raised important technical concerns in the field, as reflected by numerous review papers and opinion pieces (Kempermann et al., 2018; Paredes et al., 2018; Tartt et al., 2018; Lucassen et al., 2020). The human issue is probably a unique chapter within the controversial story of AHN. Our experience working with postmortem human brain samples has taught us that this tissue is particularly sensitive to subtle variations in preservation and histologic procedures, and that methodology is critical for the study of human AHN. The next section summarizes our view of the general limitations of the studies performed on postmortem material of human origin and possible explanations for the discrepancies between human AHN studies.
Limitations of human studies
The most notable limitation that pertains to most research into human AHN is related to the postmortem nature of the samples used. Consequently, information about the past life history of the subjects from whom the samples have been obtained is usually scarce. Digging into the details of drug consumption, medication, and lifestyle habits, including physical activity, sleep patterns, and diet, as well as the existence of either known or unknown comorbidities is not always possible after brain donation. This consideration applies not only to adult neurogenesis studies, but potentially to any analysis performed on postmortem material of human origin.
The second major limitation associated with the use of human samples is the incompatibility between the particular tissue processing methodologies required to perform certain studies and the optimized workflow followed at most brain banks. The methodologies routinely used in these banks, which include either long-term fixation of the samples in formalin and their subsequent embedding into paraffin, or flash-freezing, generate tissues of high quality to perform a great number of biochemical, molecular, and histologic investigations. However, studies of adult neurogenesis require special processing techniques that are not fully compatible with this standard workflow (Moreno-Jiménez et al., 2019; Flor-García et al., 2020). Therefore, researchers working on human AHN are sometimes required to start customized prospective collections of samples that fulfill their needs, and this is not only extremely time- and resource-consuming but not always offered by biobanks.
A third major limitation is related to difficulties in precisely tracking the fixation procedure and duration to which each sample of a single study has been subjected. This is an essential requirement for guaranteeing methodological homogeneity within and between studies, but it is not always fulfilled. Notably, recent work by our group has revealed that subtle differences in the tissue fixation procedure can critically affect the detection of markers of AHN in the human DG (Moreno-Jiménez et al., 2019; Flor-García et al., 2020), as further detailed in the next section.
Another important limitation to human studies concerns the wide variety of specialized technical expertise required by the whole research team to be able to tackle this type of research in a rigorous manner. Using external anatomic cues, neuropathologists performing the autopsies have to be extremely accurate in identifying the brain region intended to be studied to ensure the reliability of the neuropathological examination, accurate classification of the patients, and homogeneity of the brain region analyzed for all the subjects. Moreover, tissue must be extracted with minimal manipulation. Compliance with these requirements not only guarantees the precision and rapidness of the dissection process but also contributes to preserving the macroscopic quality of the tissue. In addition to obtaining properly prepared samples, basic researchers must receive specialized training on performing and interpreting immunohistochemistry on human brain tissue because the evaluation of both positive and negative immunohistochemical signals can be biased by the presence of high autofluorescence and nonspecific staining that characterizes aged human brain tissue. In view of this, studies conducted in humans call for even more restrictive criteria than those used for rodents (Flor-García et al., 2020). The identification of false-positive and -negative results is a crucial task that untrained personnel may not always find straightforward.
A final limitation that applies to any histologic analysis of human tissue is the inherent observational nature of these studies. This includes, but is not limited to, the need to use “proxy markers” for any phenomenon of interest. Given that humans cannot be subjected to interventional studies, careful validation experiments are crucial. Consequently, positive and, in particular, negative results should always be interpreted with caution.
Given the aforementioned limitations of working with postmortem human samples, biopsy specimens from temporal lobe resection surgeries emerge as a potentially promising alternative. These samples have a major advantage over postmortem samples in that they are immediately fixed after surgical dissection. However, the results obtained from these samples must also be interpreted with care as these surgeries are performed almost exclusively on patients with intractable epilepsy or brain tumors. These health conditions have deleterious effects on AHN in rodents (Parent et al., 1997; Jessberger et al., 2005; Overstreet-Wadiche et al., 2006). Although several studies suggest that the consequences of seizures in humans are similar to those observed in mice (Table 1), epilepsy may not affect human AHN in the same way as in mice. Therefore, special emphasis should be placed on not extrapolating the conclusions obtained from patients with these diseases to the general population a priori. Moreover, the methodologies used to process tissues obtained from biopsies should be as precisely controlled as for postmortem samples.
Methodological considerations for the study of adult neurogenesis in humans
Figure 1 shows a schematic representation of distinct techniques routinely implemented to process postmortem human brain samples. As shown, these approaches are not only divergent but also mutually incompatible. Our recent work (Moreno-Jiménez et al., 2019; Flor-García et al., 2020) revealed the great extent to which certain tissue processing methods hinder the detection of markers of AHN in the human DG. Of the steps shown in Figure 1, we focused our attention on determining the impact of a seemingly trivial but, indeed, critical factor, namely, tissue fixation, on the qualitative and quantitative detection of markers of AHN in this region of the brain. To this end, we obtained the whole hippocampus from several neurologically healthy control subjects. Subsequently, hippocampi were divided into several fragments, each of which was fixed for a different period (ranging between 1 and 48 h) in freshly prepared 4% PFA. After completing the fixation step, we counted the number of cells expressing doublecortin (DCX+) or polysialylated neural cell adhesion molecule (PSA-NCAM+), the two gold-standard markers for immature neurons in the adult brain (Rao and Shetty, 2004), in the different fragments from each subject. Fixation for longer than 12 h increased background intensity and nonspecific staining, thus impeding the detection of both DCX+ and PSA-NCAM+ cells, which were clearly observed with shorter fixation times. When fixation was prolonged to up to 48 h, we found that the loss of the DCX and PSA-NCAM signals was reversible using specific histologic procedures (Moreno-Jiménez et al., 2019; Flor-García et al., 2020). In samples fixed for 24 h in 4% PFA, 5 of 9 anti-DCX antibodies produced reliable staining (Flor-García et al., 2020). In contrast, when fixation with commercial formalin was prolonged for several months, which is a routine method used at most brain banks worldwide, none of those 9 anti-DCX antibodies detected the presence of positive cells in the same patients. Thus, we concluded that samples fixed in formalin for long periods are, at least in our hands, totally inadequate for the detection of immature neurons in the human DG (Flor-García et al., 2020). Given the clear-cut consequences that the aforementioned subtle changes in tissue processing methods have on the detection of markers of AHN in the human DG, we believe that it should be mandatory for all human histologic studies to report on the following: the fixation procedure and/or duration; whether histologic or nonhistologic pretreatments were performed; and what controls were used to validate the antibody signal.
Figure 1.

Variety of methods routinely used to process postmortem human brain samples. During autopsy, the brain is removed from the skull (A). After dissecting the area of interest, tissue can be either frozen or fixed. Various chemical fixatives can be used. After fixation, samples can be embedded in resin, paraffin, or a mixture of agarose/sucrose (B) to favor tissue preservation and sectioning. Depending on the embedding support, a cryostat, microtome, or vibratome (C) sectioning method should be used. Staining is performed following previously established immunohistochemical, histologic, or electron immunohistochemical protocols. Human hippocampal sections subjected to immunohistochemistry staining before (D), during (E), and after (F) the final autofluorescence elimination step are shown. Stained sections can be observed under conventional epifluorescence, electron, or confocal microscopes. Blue represents the protocol followed in our laboratory. A complete description of this protocol can be found in Moreno-Jiménez et al. (2019) and Flor-García et al. (2020).
Interestingly, several reflective Dual Perspectives papers (Kempermann et al., 2018; Lucassen et al., 2020) pointed to prolonged postmortem delay, namely, the time elapsed between death and sample immersion in fixative, as a potential explanation for the absence of staining with distinct markers of AHN in several human studies. We carefully analyzed the impact of variable postmortem delay intervals on the quantitative and qualitative staining of several of these markers. We compared samples obtained from 58 individuals whose postmortem delay intervals fell between 2.5 and 38 h (Moreno-Jiménez et al., 2019). We did not observe variations in the number of DCX+ or PSA-NCAM+ cells, staining quality, or cellular distribution of these markers between samples with short and long postmortem delay. Although loss of DCX staining in the distal dendrites and confinement of DCX signal to the nuclear and somatic compartments has been reported in rats after long postmortem delay intervals, we did not qualitatively observe any of these phenomena in our samples. Nevertheless, it should be noted that the postmortem delay of our samples was relatively short compared with those of other studies. Future research should address the impact of longer postmortem delay intervals or other potentially important factors, such as temperature, on the detection of markers of AHN in the human brain.
Due consideration should be given to the apparent disparity in the number of positive cells reported by different studies. Importantly, various recent studies conducted in humans (Knoth et al., 2010; Boldrini et al., 2018; Moreno-Jiménez et al., 2019; Tobin et al., 2019; Flor-García et al., 2020) applied unbiased stereological criteria to perform cell counts in the DG. Of note, not only the microscopy equipment (confocal, conventional, or epifluorescence microscopes), but also the unbiased stereological method used to count cells (i.e., either physical or optical dissector, or optical fractionator) differed between these studies. Moreover, only a few studies have systematically reported total numbers of cells in the entire DG (Boldrini et al., 2018; Tobin et al., 2019). A cautionary note concerns the comparison of total numbers of cells between those studies and others, such as ours (Moreno-Jiménez et al., 2019; Flor-García et al., 2020), in which only cell densities were estimated. We calculated cell densities by dividing the number of cells counted within each stack of confocal images by the reference volume, which was defined as the volume of the granule cell layer included in that stack (Moreno-Jiménez et al., 2019; Flor-García et al., 2020), following the previously described physical dissector stereological method coupled to confocal microscopy (Llorens-Martin et al., 2006). When comparing data from different studies, it should be noted that changes in the structure or substructure used for calculation of the reference volume (e.g., including, or not, the portion of the hilus that appears in the image), or differences in the precision achieved when manually delineating the reference volume, might account for differences of at least one order of magnitude in the total number of cells reported. Moreover, it should be noted that both Boldrini et al. (2018) and Tobin et al. (2019) examined the whole rostrocaudal extent of the human DG, whereas only the posterior pole of the anterior hippocampus was examined by Moreno-Jiménez et al. (2019). The differences in the precise anatomic regions examined and how the reference volume was calculated in different studies may add disparity to the number of cells reported. Therefore, the results of the aforementioned studies should not be considered contradictory but rather calculated and reported in a dissimilar manner.
A final remark in this section regards the need to apply strict criteria to assign a given cell the attribute of “positive” for a certain marker. For human studies based on histologic determinations, exhaustive control experiments for each type of sample and marker of interest are particularly important. These controls should include systematic consultation of the available literature, the use of several antibodies raised against distinct domains of the same protein (Moreno-Jiménez et al., 2019; Flor-García et al., 2020), specific synthetic blocking peptides to rule out nonspecific staining (Moreno-Jiménez et al., 2019), biological validation of the results using positive and negative controls (i.e., fetal vs adult non-neurogenic regions, respectively), and the careful examination of autofluorescence phenomena (Flor-García et al., 2020). Similarly, when negative results are obtained, each step of the method used should be subjected to exhaustive evaluation. For instance, 4 of 9 anti-DCX antibodies that provide quality staining of murine tissue did not show specificity on high-quality human samples that had been fixed for 24 h in PFA (Flor-García et al., 2020). In contrast, another 5 distinct antibodies worked appropriately in the same samples. In light of these results, the correct interpretation of negative results gains further relevance in the context of research into human AHN.
Human AHN in physiology and pathology
As previously mentioned, most of the studies that have either questioned or supported the persistence of AHN in the human DG throughout lifetime are based on the histologic analysis of proxy markers of AHN (Table 1). The two exceptions were those by Eriksson et al. (1998) and Spalding et al. (2013), which examined cell division through the incorporation of either BrdU or 14C into dividing cells. Importantly, these two robust studies provided key quantitative data strongly supporting the occurrence of AHN in the adult human DG. The latter study determined that ∼700 new dentate granule cells are incorporated into the adult human DG per brain hemisphere per day (Spalding et al., 2013). These data are potentially in agreement with the quantification of proliferation markers reported previously (Boldrini et al., 2009, 2012; Knoth et al., 2010). Moreover, the work by Spalding et al. (2013) also showed a mild decrease in the rate of human AHN with aging. We examined markers of AHN in 13 neurologically healthy control subjects aged between 43 and 87 yrs old. Although we did not quantify the absolute number of proliferative cells, we observed a mild decrease in the number of DCX+ immature dentate granule cells with age (Moreno-Jiménez et al., 2019). All the aforementioned observations are consistent with the findings of numerous studies performed on rodents, which also support a reduction of AHN rate with age (Kempermann et al., 2002). Because of significant differences in the lifetimes of humans and rodents, calibration is required to quantitatively compare AHN between these species (Snyder, 2019). In this regard, although the seminal studies by Eriksson et al. (1998) demonstrated that newly generated dentate granule cells achieve full maturation in humans, there is still no precise information regarding the time required by these cells to complete their maturation in humans. Given the high phylogenetic proximity between humans and nonhuman primates, it has been hypothesized that newly generated dentate granule cells in humans, like those in other primates, mature at a slower rate than those of rodents (Kohler et al., 2011; Sorrells et al., 2018).
Despite not providing information on the timing of this maturation, our data support the notion that the population of DCX+ cells is not static but instead continuously undergoes a dynamic maturation process. Indeed, DCX+ cells at different maturation stages are present in the adult human DG, with some subpopulations expressing additional well-characterized cell markers of immature neurons, such as PSA-NCAM and calretinin (Fig. 2A), whereas others express markers of more advanced stages of maturation, such as NeuN, calbindin, and Tau (Moreno-Jiménez et al., 2019). For instance, Figure 2A illustrates the immature morphology of a triple-labeled DCX+/calretinin+/PSA-NCAM+ dentate granule cell. In the subpopulation of cells that coexpress DCX and markers of advanced differentiation stages, we carefully analyzed the expression of those markers. We observed that the expression of NeuN in these cells was lower than in fully mature surrounding DCX– dentate granule cells (Flor-García et al., 2020). Moreover, DCX+ cells that expressed calbindin, a calcium-binding protein expressed by dentate granule cells at advanced maturation stages, showed slightly smaller somas than fully mature calbindin+ DCX– dentate granule cells (Flor-García et al., 2020). These data strongly support the notion that DCX+ dentate granule cells are a dynamic cell population that undergoes progressive maturation and whose features clearly differ from those of fully differentiated mature dentate granule cells.
Figure 2.
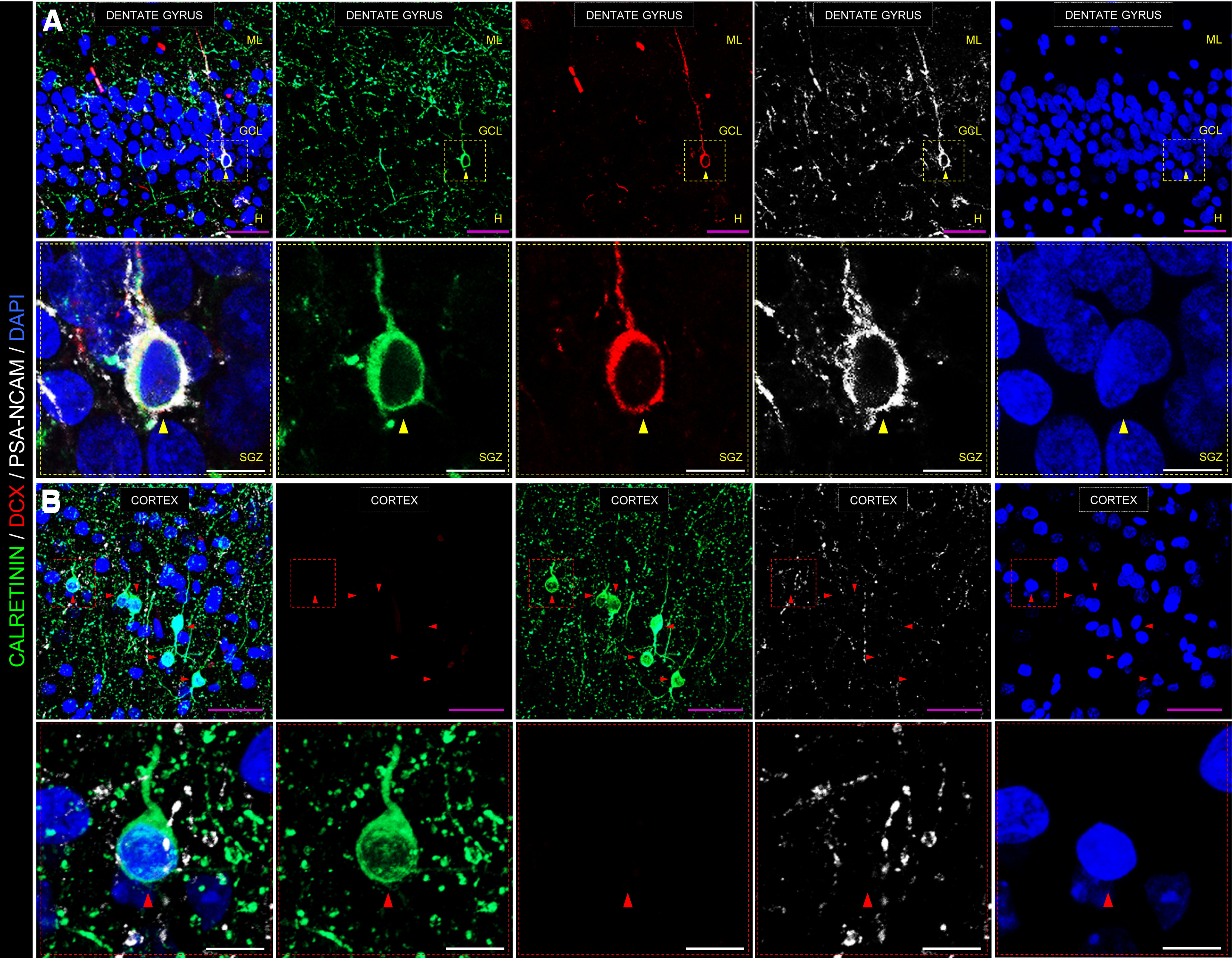
Representative images showing an immunohistochemical staining performed on postmortem human hippocampal samples. A, Low- and high-power magnification images show the anatomic organization of the human DG. In high-power magnification images, a triple-labeled DCX+/calretinin+/PSA-NCAM+ immature dentate granule cell is shown. B, Representative image of the human cortex is shown as a negative control for DCX staining. Note the abundance of calretinin+ interneurons, which is in sharp contrast to the absence of DCX+ cells in this structure. GCL, Granule cell layer; ML, molecular layer; H, hilus; SGZ, subgranular zone. Scale bars: magenta, 50 μm; white, 10 μm. Yellow triangles represent DCX+/calretinin+/PSA-NCAM+ immature dentate granule cell. Red triangles represent DCX–/calretinin+/PSA-NCAM– cortical interneuron. Immunohistochemistry was performed following a previously published protocol (Moreno-Jiménez et al., 2019; Flor-García et al., 2020), which included sodium borohydride (NaBH4), heat-mediated citrate antigen retrieval (HC-AR), and autofluorescence elimination steps. Low-power confocal stacks of images were obtained in a LSM800 Carl Zeiss confocal microscope, equipped with three GaAsP detectors and using a 40× immersion oil objective. A, 0.8 zoom, XY dimensions: 199.66 µm, Z interval: 1.5 µm, pinhole dimensions: 0.9 Airy units. B, 1.1 zoom, XY dimensions: 145.2 µm, Z interval: 1.5 µm, pinhole dimensions: 0.9 Airy units. High-power magnification images were obtained in the same confocal microscope using a 63× immersion oil objective (2.2 zoom, XY dimensions: 46 µm, Z interval: 0.5 µm, pinhole dimensions: 0.9 Airy units). All the images shown in this figure correspond to postmortem human samples obtained from a neurologically healthy control subject. Fixation time was limited to 24 h in freshly prepared 4% PFA at 4°C. The epidemiological data of this subject are the following: gender, male; age, 43; postmortem delay, 5 h; Braak-Tau stage, 0; CERAD stage, 0; and Braak α-synuclein stage, 0. Brain tissue donation, processing, and use for research were in compliance with published protocols (Martinez-Martin et al., 2010), which include the obtaining of informed consent for brain tissue donation from living donors, and approval of the whole donation process by the Ethical Committee of the Spanish Research Council (Committee Approval Reference 025-2020#).
In addition to the physiological variations in AHN observed throughout human life, solid evidence supporting the dynamic nature of this phenomenon in humans comes from the direct examination of markers of AHN in patients with various diseases (Table 1). As has been systematically reported in animal studies, AHN appears to be altered in patients with epilepsy, ischemic and hemorrhagic stroke, major depression, schizophrenia, ethanol or heroin addiction, cancer, and neurodegenerative conditions, such as Alzheimer's disease (AD), Creutzfeldt-Jakob disease, and Lewy body dementia (for a full list of these studies, see Table 1). Of particular importance are the following observations. First, cancer treatment selectively eliminates cells positive for markers of AHN in humans (Monje et al., 2007). Second, reduced numbers of progenitor and proliferating cells are observed in patients with major depression (Boldrini et al., 2009; Lucassen et al., 2010). Importantly, antidepressants increase AHN in mice (Santarelli et al., 2003) and also in humans (Boldrini et al., 2009, 2012) and nonhuman primates (Perera et al., 2007). Finally, a specific decrease in the number of immature neurons (Moreno-Jiménez et al., 2019), an altered number of progenitor and proliferative cells (Crews et al., 2010; Gomez-Nicola et al., 2014; Tobin et al., 2019), and a blockade in the maturation of neuroblasts (Moreno-Jiménez et al., 2019) have been reported in patients with AD, while numbers of fully mature dentate granule cells are unchanged. Our data obtained from these patients suggest that, in addition to the physiological decrease in AHN rate caused by aging, independent neuropathological mechanisms drive this decrease in AHN in AD (Moreno-Jiménez et al., 2019). This hypothesis is particularly relevant because the decrease in the number of immature neurons was found to start during early-stage AD, although AHN impairments were accentuated as the disease progressed (Moreno-Jiménez et al., 2019). Moreover, the expression of proxy markers of AHN correlated with cognitive scores in these patients (Tobin et al., 2019). Together, these data might provide insight into how the neurogenic potential of the human hippocampus contributes to cognition and mood regulation.
Future directions and unsolved questions
Despite the outstanding development of the human AHN research field in recent years, numerous key questions remain unanswered.
First, most human AHN studies published in the last decade have been limited to the postmortem observational analysis of markers of AHN. Despite the extensive validation of these markers in rodents and other mammalian species, the generally poor performance of most antibodies on human tissue has been a major obstacle for the further development of the human AHN research field and has prevented the detailed reconstruction of the whole differentiation process of newly generated neurons. Generally speaking, the efficacy of various markers of immature neurons on human tissue is satisfactory. In contrast, the poor performance of numerous markers that characterize the initial stages of AHN, namely, those in which radial glia-like cells give rise to progenitor cells with a variable commitment to the neuronal lineage, has hindered the detailed reconstruction of these early stages. Alternative techniques to immunohistochemistry, such as those at the single-cell level, are expected to shed light on the field in the coming years. These techniques may help to identify novel markers, or to cluster new subpopulations of cells or intermediate differentiation stages. Indeed, one of the next key challenges in the field concerns the identification of a stem cell signature in the human DG.
Another crucial piece of information that is currently lacking is the tempo of AHN in humans. Current techniques do not allow the determination of the time required for a new neuron to fully mature. Nonmutagenic approaches to replace the use of BrdU and capable of birthdating new cells are needed to achieve a precise reconstruction of the maturational time course of these cells. We also need to determine whether this time course is modified in response to physiological aging or pathologic conditions. In this regard, Trinchero et al. (2017) demonstrated that newborn dentate granule cells matured more slowly in aged rodents than in young animals. Understanding the fine details of the timing of human AHN throughout physiological and pathologic aging might provide novel ways to preserve and stimulate the survival and maturation of new neurons under certain pathologic conditions.
The field also requires the standardization and exchange of protocols to precisely define the minimal requirements that human samples must fulfill to be considered valid for AHN studies. Of note, the vast majority of samples stored at brain banks worldwide are subjected to several weeks/months fixation in formalin. As discussed above, in our hands and to date, this methodology appears to be incompatible with the detection of markers of AHN. The development of a method that can work with these samples in the context of human AHN studies would be an extremely valuable contribution to the field.
Finally, although postmortem observational studies have an enormous potential to increase our basic knowledge about human AHN, they are of limited therapeutic or diagnostic use. Therefore, great hope is placed on methodologies capable of visualizing AHN in living organisms (Manganas et al., 2007; Sierra et al., 2011) or exploring pattern separation capabilities as a proxy for human AHN (Suwabe et al., 2018; Bernstein and McNally, 2019; Riphagen et al., 2020). Breakthroughs in this regard are likely to be the only way through which the role played by newborn dentate granule cells in the human brain will be unraveled. Demonstration that this cell population in humans plays similar roles to that in rodents may lead to the use of noninvasive methodologies to diagnose (or even to prevent) certain pathologic or neurodegenerative conditions and would turn AHN into a relevant biomarker for these conditions.
In conclusion, evidence discussed here demonstrates that slight differences in either tissue processing or histologic methodologies do limit the detection of several markers of adult neurogenesis in the human hippocampus, to the point of making them undetectable. In contrast, when high-quality tissues are subjected to short fixation times and to the appropriate histologic pretreatments, thousands of immature neurons can be observed in the DG until the 10th decade of human life. These data are in line with previous evidence, including that based on immunohistochemistry, BrdU incorporation, and carbon dating. Moreover, they indicate that adult neurogenesis is a robust phenomenon that occurs in the human hippocampus during physiological and pathologic aging.
AHN is an unquestionable source of plasticity in rodents. Given that a significant number of new neurons seem to be added to the human healthy hippocampus and that AHN sharply decreases in patients with AD, research efforts should be channeled into finding ways to protect these cells before the onset of neurodegeneration. In the long run, unraveling the mechanisms that control newborn neuron maturation and synaptic integration under physiological and pathologic conditions is expected to pave the way toward the promotion of the regenerative potential of the human brain, potentially helping to preserve this unique source of plasticity throughout lifetime.
Footnotes
This work was supported by Spanish Ministry of Economy and Competitiveness SAF-2017-82185-R and RYC-2015-171899 to M.L.-M., Alzheimer's Association 2015-NIRG-340709 and AARG-17-528125 to M.L.-M., Association for Frontotemporal Degeneration 2016 Basic Science Pilot Grant Award to M.L.-M., and Comunidad de Madrid PEJD-2017-PRE/BMD-3439 to M.L.-M. Institutional grants from Fundación Ramón Areces and Banco de Santander to Centro de Biología Molecular “Severo Ochoa” are also acknowledged. The salary of E.M.J. was supported by Beca Predoctoral en Neurociencias Fundación Tatiana Pérez de Guzmán fellowship. The salary of J.T.-R. was supported by Universidad Autónoma de Madrid Doctorate fellowship FPI-UAM 2017 program. The salary of M.F.-G. was supported by Formación de personal Investigador contract, associated with Grant SAF-2017-82185-R to M.L.-M., supported by Spanish Ministry for Economy and Competitiveness PRE2018-085233.
The authors declare no competing financial interests.
References
- Akers KG, Martinez-Canabal A, Restivo L, Yiu AP, De Cristofaro A, Hsiang HL, Wheeler AL, Guskjolen A, Niibori Y, Shoji H, Ohira K, Richards BA, Miyakawa T, Josselyn SA, Frankland PW (2014) Hippocampal neurogenesis regulates forgetting during adulthood and infancy. Science 344:598–602. 10.1126/science.1248903 [DOI] [PubMed] [Google Scholar]
- Allen KM, Fung SJ, Weickert CS (2016) Cell proliferation is reduced in the hippocampus in schizophrenia. Aust N Z J Psychiatry 50:473–480. 10.1177/0004867415589793 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altman J (1963) Autoradiographic investigation of cell proliferation in the brains of rats and cats. Anat Rec 145:573–591. 10.1002/ar.1091450409 [DOI] [PubMed] [Google Scholar]
- Altman J, Das GD (1965) Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J Comp Neurol 124:319–335. 10.1002/cne.901240303 [DOI] [PubMed] [Google Scholar]
- Alvarez-Buylla A, Theelen M, Nottebohm F (1988) Birth of projection neurons in the higher vocal center of the canary forebrain before, during, and after song learning. Proc Natl Acad Sci USA 85:8722–8726. 10.1073/pnas.85.22.8722 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amrein I, Dechmann DK, Winter Y, Lipp HP (2007) Absent or low rate of adult neurogenesis in the hippocampus of bats (Chiroptera). PLoS One 2:e455. 10.1371/journal.pone.0000455 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anacker C, Hen R (2017) Adult hippocampal neurogenesis and cognitive flexibility: linking memory and mood. Nat Rev Neurosci 18:335–346. 10.1038/nrn.2017.45 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anacker C, Luna VM, Stevens GS, Millette A, Shores R, Jimenez JC, Chen B, Hen R (2018) Hippocampal neurogenesis confers stress resilience by inhibiting the ventral dentate gyrus. Nature 559:98–102. 10.1038/s41586-018-0262-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayer R, Franke H, Ficker C, Richter M, Lessig R, Büttner A, Weber M (2015) Alterations of neuronal precursor cells in stages of human adult neurogenesis in heroin addicts. Drug Alcohol Depend 156:139–149. 10.1016/j.drugalcdep.2015.09.005 [DOI] [PubMed] [Google Scholar]
- Bernstein EE, McNally RJ (2019) Examining the effects of exercise on pattern separation and the moderating effects of mood symptoms. Behav Ther 50:582–593. 10.1016/j.beth.2018.09.007 [DOI] [PubMed] [Google Scholar]
- Bischofberger J (2007) Young and excitable: new neurons in memory networks. Nat Neurosci 10:273–275. 10.1038/nn0307-273 [DOI] [PubMed] [Google Scholar]
- Blümcke I, Schewe JC, Normann S, Brüstle O, Schramm J, Elger CE, Wiestler OD (2001) Increase of nestin-immunoreactive neural precursor cells in the dentate gyrus of pediatric patients with early-onset temporal lobe epilepsy. Hippocampus 11:311–321. 10.1002/hipo.1045 [DOI] [PubMed] [Google Scholar]
- Boekhoorn K, Joels M, Lucassen PJ (2006) Increased proliferation reflects glial and vascular-associated changes, but not neurogenesis in the presenile Alzheimer hippocampus. Neurobiol Dis 24:1–14. 10.1016/j.nbd.2006.04.017 [DOI] [PubMed] [Google Scholar]
- Boldrini M, Underwood MD, Hen R, Rosoklija GB, Dwork AJ, Mann JJ, Arango V (2009) Antidepressants increase neural progenitor cells in the human hippocampus. Neuropsychopharmacology 34:2376–2389. 10.1038/npp.2009.75 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boldrini M, Hen R, Underwood MD, Rosoklija GB, Dwork AJ, Mann JJ, Arango V (2012) Hippocampal angiogenesis and progenitor cell proliferation are increased with antidepressant use in major depression. Biol Psychiatry 72:562–571. 10.1016/j.biopsych.2012.04.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boldrini M, Fulmore CA, Tartt AN, Simeon LR, Pavlova I, Poposka V, Rosoklija GB, Stankov A, Arango V, Dwork AJ, Hen R, Mann JJ (2018) Human hippocampal neurogenesis persists throughout aging. Cell Stem Cell 22:589–599.e585. 10.1016/j.stem.2018.03.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brandt MD, Jessberger S, Steiner B, Kronenberg G, Reuter K, Bick-Sander A, von der Behrens W, Kempermann G (2003) Transient calretinin expression defines early postmitotic step of neuronal differentiation in adult hippocampal neurogenesis of mice. Mol Cell Neurosci 24:603–613. 10.1016/s1044-7431(03)00207-0 [DOI] [PubMed] [Google Scholar]
- Brown J, Cooper-Kuhn CM, Kempermann G, Van Praag H, Winkler J, Gage FH, Kuhn HG (2003) Enriched environment and physical activity stimulate hippocampal but not olfactory bulb neurogenesis. Eur J Neurosci 17:2042–2046. 10.1046/j.1460-9568.2003.02647.x [DOI] [PubMed] [Google Scholar]
- Cameron HA, Gould E (1994) Adult neurogenesis is regulated by adrenal steroids in the dentate gyrus. Neuroscience 61:203–209. 10.1016/0306-4522(94)90224-0 [DOI] [PubMed] [Google Scholar]
- Cipriani S, Ferrer I, Aronica E, Kovacs GG, Verney C, Nardelli J, Khung S, Delezoide AL, Milenkovic I, Rasika S, Manivet P, Benifla JL, Deriot N, Gressens P, Adle-Biassette H (2018) Hippocampal radial glial subtypes and their neurogenic potential in human fetuses and healthy and Alzheimer's disease adults. Cereb Cortex 28:2458–2478. 10.1093/cercor/bhy096 [DOI] [PubMed] [Google Scholar]
- Couillard-Despres S, Winner B, Schaubeck S, Aigner R, Vroemen M, Weidner N, Bogdahn U, Winkler J, Kuhn HG, Aigner L (2005) Doublecortin expression levels in adult brain reflect neurogenesis. Eur J Neurosci 21:1–14. 10.1111/j.1460-9568.2004.03813.x [DOI] [PubMed] [Google Scholar]
- Crews L, Adame A, Patrick C, Delaney A, Pham E, Rockenstein E, Hansen L, Masliah E (2010) Increased BMP6 levels in the brains of Alzheimer's disease patients and APP transgenic mice are accompanied by impaired neurogenesis. J Neurosci 30:12252–12262. 10.1523/JNEUROSCI.1305-10.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chawana R, Alagaili A, Patzke N, Spocter MA, Mohammed OB, Kaswera C, Gilissen E, Bennett NC, Ihunwo AO, Manger PR (2014) Microbats appear to have adult hippocampal neurogenesis, but post-capture stress causes a rapid decline in the number of neurons expressing doublecortin. Neuroscience 277:724–733. 10.1016/j.neuroscience.2014.07.063 [DOI] [PubMed] [Google Scholar]
- D'Alessio L, Konopka H, López EM, Seoane E, Consalvo D, Oddo S, Kochen S, López-Costa JJ (2010) Doublecortin (DCX) immunoreactivity in hippocampus of chronic refractory temporal lobe epilepsy patients with hippocampal sclerosis. Seizure 19:567–572. 10.1016/j.seizure.2010.09.004 [DOI] [PubMed] [Google Scholar]
- D'Alessio L, Konopka H, Escobar E, Acuña A, Oddo S, Solís P, Seoane E, Kochen S (2015) Dentate gyrus expression of nestin-immunoreactivity in patients with drug-resistant temporal lobe epilepsy and hippocampal sclerosis. Seizure 27:75–79. 10.1016/j.seizure.2015.02.008 [DOI] [PubMed] [Google Scholar]
- Dennis CV, Suh LS, Rodriguez ML, Kril JJ, Sutherland GT (2016) Human adult neurogenesis across the ages: an immunohistochemical study. Neuropathol Appl Neurobiol 42:621–638. 10.1111/nan.12337 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doetsch F, Caille I, Lim DA, Garcia-Verdugo JM, Alvarez-Buylla A (1999) Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell 97:703–716. 10.1016/s0092-8674(00)80783-7 [DOI] [PubMed] [Google Scholar]
- Epp JR, Beasley CL, Galea LA (2013) Increased hippocampal neurogenesis and p21 expression in depression: dependent on antidepressants, sex, age, and antipsychotic exposure. Neuropsychopharmacology 38:2297–2306. 10.1038/npp.2013.132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH (1998) Neurogenesis in the adult human hippocampus. Nat Med 4:1313–1317. 10.1038/3305 [DOI] [PubMed] [Google Scholar]
- Ernst A, Alkass K, Bernard S, Salehpour M, Perl S, Tisdale J, Possnert G, Druid H, Frisen J (2014) Neurogenesis in the striatum of the adult human brain. Cell 156:1072–1083. 10.1016/j.cell.2014.01.044 [DOI] [PubMed] [Google Scholar]
- Fasemore TM, Patzke N, Kaswera-Kyamakya C, Gilissen E, Manger PR, Ihunwo AO (2018) The distribution of Ki-67 and doublecortin-immunopositive cells in the brains of three Strepsirrhine primates: Galago demidoff, Perodicticus potto, and Lemur catta. Neuroscience 372:46–57. 10.1016/j.neuroscience.2017.12.037 [DOI] [PubMed] [Google Scholar]
- Filippov V, Kronenberg G, Pivneva T, Reuter K, Steiner B, Wang LP, Yamaguchi M, Kettenmann H, Kempermann G (2003) Subpopulation of nestin-expressing progenitor cells in the adult murine hippocampus shows electrophysiological and morphological characteristics of astrocytes. Mol Cell Neurosci 23:373–382. 10.1016/s1044-7431(03)00060-5 [DOI] [PubMed] [Google Scholar]
- Flor-García M, Terreros-Roncal J, Moreno-Jiménez EP, Ávila J, Rábano A, Llorens-Martín M (2020) Unraveling human adult hippocampal neurogenesis. Nat Protoc 15:668–693. 10.1038/s41596-019-0267-y [DOI] [PubMed] [Google Scholar]
- Gatome CW, Mwangi DK, Lipp HP, Amrein I (2010) Hippocampal neurogenesis and cortical cellular plasticity in Wahlberg's epauletted fruit bat: a qualitative and quantitative study. Brain Behav Evol 76:116–127. 10.1159/000320210 [DOI] [PubMed] [Google Scholar]
- Ge S, Yang CH, Hsu KS, Ming GL, Song H (2007) A critical period for enhanced synaptic plasticity in newly generated neurons of the adult brain. Neuron 54:559–566. 10.1016/j.neuron.2007.05.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geha S, Pallud J, Junier MP, Devaux B, Leonard N, Chassoux F, Chneiweiss H, Daumas-Duport C, Varlet P (2010) NG2+/Olig2+ cells are the major cycle-related cell population of the adult human normal brain. Brain Pathology 20:399–411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez-Nicola D, Suzzi S, Vargas-Caballero M, Fransen NL, Al-Malki H, Cebrian-Silla A, Garcia-Verdugo JM, Riecken K, Fehse B, Perry VH (2014) Temporal dynamics of hippocampal neurogenesis in chronic neurodegeneration. Brain 137:2312–2328. 10.1093/brain/awu155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gould E, Daniels DC, Cameron HA, McEwen BS (1992) Expression of adrenal steroid receptors by newly born cells and pyknotic cells in the dentate gyrus of the postnatal rat. Mol Cell Neurosci 3:44–48. 10.1016/1044-7431(92)90007-o [DOI] [PubMed] [Google Scholar]
- Gould E, Reeves AJ, Fallah M, Tanapat P, Gross CG, Fuchs E (1999) Hippocampal neurogenesis in adult Old World primates. Proc Natl Acad Sci USA 96:5263–5267. 10.1073/pnas.96.9.5263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu Y, Arruda-Carvalho M, Wang J, Janoschka SR, Josselyn SA, Frankland PW, Ge S (2012) Optical controlling reveals time-dependent roles for adult-born dentate granule cells. Nat Neurosci 15:1700–1706. 10.1038/nn.3260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heigele S, Sultan S, Toni N, Bischofberger J (2016) Bidirectional GABAergic control of action potential firing in newborn hippocampal granule cells. Nat Neurosci 19:263–270. 10.1038/nn.4218 [DOI] [PubMed] [Google Scholar]
- Hill AS, Sahay A, Hen R (2015) Increasing adult hippocampal neurogenesis is sufficient to reduce anxiety and depression-like behaviors. Neuropsychopharmacology 40:2368–2378. 10.1038/npp.2015.85 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishikawa R, Fukushima H, Frankland PW, Kida S (2016) Hippocampal neurogenesis enhancers promote forgetting of remote fear memory after hippocampal reactivation by retrieval. Elife 5:e17464. 10.7554/eLife.17464 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jabes A, Lavenex PB, Amaral DG, Lavenex P (2010) Quantitative analysis of postnatal neurogenesis and neuron number in the macaque monkey dentate gyrus. Eur J Neurosci 31:273–285. 10.1111/j.1460-9568.2009.07061.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jessberger S, Romer B, Babu H, Kempermann G (2005) Seizures induce proliferation and dispersion of doublecortin-positive hippocampal progenitor cells. Exp Neurol 196:342–351. 10.1016/j.expneurol.2005.08.010 [DOI] [PubMed] [Google Scholar]
- Jin K, Peel AL, Mao XO, Xie L, Cottrell BA, Henshall DC, Greenberg DA (2004) Increased hippocampal neurogenesis in Alzheimer's disease. Proc Natl Acad Sci U S A 101:343–347. 10.1073/pnas.2634794100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson M, Ekonomou A, Hobbs C, Ballard CG, Perry RH, Perry EK (2011) Neurogenic marker abnormalities in the hippocampus in dementia with Lewy bodies. Hippocampus 21:1126–1136. 10.1002/hipo.20826 [DOI] [PubMed] [Google Scholar]
- Kaplan MS, Hinds JW (1977) Neurogenesis in the adult rat: electron microscopic analysis of light radioautographs. Science 197:1092–1094. 10.1126/science.887941 [DOI] [PubMed] [Google Scholar]
- Kelsch W, Lin CW, Lois C (2008) Sequential development of synapses in dendritic domains during adult neurogenesis. Proc Natl Acad Sci USA 105:16803–16808. 10.1073/pnas.0807970105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kempermann G, Kuhn HG, Gage FH (1997) More hippocampal neurons in adult mice living in an enriched environment. Nature 386:493–495. 10.1038/386493a0 [DOI] [PubMed] [Google Scholar]
- Kempermann G, van Praag H, Gage FH (2000) Activity-dependent regulation of neuronal plasticity and self repair. Prog Brain Res 127:35–48. 10.1016/s0079-6123(00)27004-0 [DOI] [PubMed] [Google Scholar]
- Kempermann G, Gast D, Gage FH (2002) Neuroplasticity in old age: sustained fivefold induction of hippocampal neurogenesis by long-term environmental enrichment. Ann Neurol 52:135–143. 10.1002/ana.10262 [DOI] [PubMed] [Google Scholar]
- Kempermann G, Gast D, Kronenberg G, Yamaguchi M, Gage FH (2003) Early determination and long-term persistence of adult-generated new neurons in the hippocampus of mice. Development 130:391–399. 10.1242/dev.00203 [DOI] [PubMed] [Google Scholar]
- Kempermann G, Jessberger S, Steiner B, Kronenberg G (2004) Milestones of neuronal development in the adult hippocampus. Trends Neurosci 27:447–452. 10.1016/j.tins.2004.05.013 [DOI] [PubMed] [Google Scholar]
- Kempermann G, Gage FH, Aigner L, Song H, Curtis MA, Thuret S, Kuhn HG, Jessberger S, Frankland PW, Cameron HA, Gould E, Hen R, Abrous DN, Toni N, Schinder AF, Zhao X, Lucassen PJ, Frisen J (2018) Human adult neurogenesis: evidence and remaining questions. Cell Stem Cell 23:25–30. 10.1016/j.stem.2018.04.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knoth R, Singec I, Ditter M, Pantazis G, Capetian P, Meyer RP, Horvat V, Volk B, Kempermann G (2010) Murine features of neurogenesis in the human hippocampus across the lifespan from 0 to 100 yrs. PLoS One 5:e8809. 10.1371/journal.pone.0008809 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kohler SJ, Williams NI, Stanton GB, Cameron JL, Greenough WT (2011) Maturation time of new granule cells in the dentate gyrus of adult macaque monkeys exceeds six months. Proc Natl Acad Sci USA 108:10326–10331. 10.1073/pnas.1017099108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kordower JH, Chen EY, Morrison JH (2010) Long-term gonadal hormone treatment and endogenous neurogenesis in the dentate gyrus of the adult female monkey. Exp Neurol 224:252–257. 10.1016/j.expneurol.2010.03.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kornack DR, Rakic P (1999) Continuation of neurogenesis in the hippocampus of the adult macaque monkey. Proc Natl Acad Sci USA 96:5768–5773. 10.1073/pnas.96.10.5768 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuhn HG, Dickinson-Anson H, Gage FH (1996) Neurogenesis in the dentate gyrus of the adult rat: age-related decrease of neuronal progenitor proliferation. J Neurosci 16:2027–2033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laplagne DA, Esposito MS, Piatti VC, Morgenstern NA, Zhao C, van Praag H, Gage FH, Schinder AF (2006) Functional convergence of neurons generated in the developing and adult hippocampus. PLoS Biol 4:e409. 10.1371/journal.pbio.0040409 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le Maître TW, Dhanabalan G, Bogdanovic N, Alkass K, Druid H (2018) Effects of alcohol abuse on proliferating cells, stem/progenitor cells, and immature neurons in the adult human hippocampus. Neuropsychopharmacology 43:690–699. 10.1038/npp.2017.251 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Sultan S, Heigele S, Schmidt-Salzmann C, Toni N, Bischofberger J (2017) Silent synapses generate sparse and orthogonal action potential firing in adult-born hippocampal granule cells. Elife 6:e23612. 10.7554/eLife.23612 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu YWJ, Curtis MA, Gibbons HM, Mee EW, Bergin PS, Teoh HH, Connor B, Dragunow M, Faull RLM (2008) Doublecortin expression in the normal and epileptic adult human brain. Eur J Neurosci 28:2254–2265. 10.1111/j.1460-9568.2008.06518.x [DOI] [PubMed] [Google Scholar]
- Liu J, Reeves C, Jacques T, McEvoy M, Miserocchi A, Thompson P, Sisodiya S, Thom M (2018) Nestin-expressing cell types in the temporal lobe and hippocampus: Morphology, differentiation, and proliferative capacity. Glia 66:62–77. 10.1002/glia.23211 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llorens-Martín M, Torres-Alemán I, Trejo JL (2006) Pronounced individual variation in the response to the stimulatory action of exercise on immature hippocampal neurons. Hippocampus 16:480–490. 10.1002/hipo.20175 [DOI] [PubMed] [Google Scholar]
- Lois C, Alvarez-Buylla A (1994) Long-distance neuronal migration in the adult mammalian brain. Science 264:1145–1148. 10.1126/science.8178174 [DOI] [PubMed] [Google Scholar]
- Lois C, Garcia-Verdugo JM, Alvarez-Buylla A (1996) Chain migration of neuronal precursors. Science 271:978–981. 10.1126/science.271.5251.978 [DOI] [PubMed] [Google Scholar]
- Lucassen PJ, Stumpel MW, Wang Q, Aronica E (2010) Decreased numbers of progenitor cells but no response to antidepressant drugs in the hippocampus of elderly depressed patients. Neuropharmacology 58:940–949. 10.1016/j.neuropharm.2010.01.012 [DOI] [PubMed] [Google Scholar]
- Lucassen PJ, Toni N, Kempermann G, Frisen J, Gage FH, Swaab DF (2020) Limits to human neurogenesis: really? Mol Psychiatry 25:2207–2209. 10.1038/s41380-018-0337-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luna VM, Anacker C, Burghardt NS, Khandaker H, Andreu V, Millette A, Leary P, Ravenelle R, Jimenez JC, Mastrodonato A, Denny CA, Fenton AA, Scharfman HE, Hen R (2019) Adult-born hippocampal neurons bidirectionally modulate entorhinal inputs into the dentate gyrus. Science 364:578–583. 10.1126/science.aat8789 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luskin MB, Pearlman AL, Sanes JR (1988) Cell lineage in the cerebral cortex of the mouse studied in vivo and in vitro with a recombinant retrovirus. Neuron 1:635–647. 10.1016/0896-6273(88)90163-8 [DOI] [PubMed] [Google Scholar]
- Manganas LN, Zhang X, Li Y, Hazel RD, Smith SD, Wagshul ME, Henn F, Benveniste H, Djuric PM, Enikolopov G, Maletic-Savatic M (2007) Magnetic resonance spectroscopy identifies neural progenitor cells in the live human brain. Science 318:980–985. 10.1126/science.1147851 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marin-Burgin A, Mongiat LA, Pardi MB, Schinder AF (2012) Unique processing during a period of high excitation/inhibition balance in adult-born neurons. Science 335:1238–1242. 10.1126/science.1214956 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez-Martin P, Avila J, AD Research Unit Investigators (2010) Alzheimer Center Reina Sofia Foundation: fighting the disease and providing overall solutions. J Alzheimers Dis 21:337–348. [DOI] [PubMed] [Google Scholar]
- Mattiesen WC, Tauber SC, Gerber J, Bunkowski S, Brück W, Nau R (2009) Increased neurogenesis after hypoxic-ischemic encephalopathy in humans is age related. Acta Neuropathol 117:525–534. 10.1007/s00401-009-0509-0 [DOI] [PubMed] [Google Scholar]
- Mathews KJ, Allen KM, Boerrigter D, Ball H, Weickert CS, Double KL (2017) Evidence for reduced neurogenesis in the aging human hippocampus despite stable stem cell markers. Aging Cell 16:1195–1199. 10.1111/acel.12641 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McAvoy KM, Scobie KN, Berger S, Russo C, Guo N, Decharatanachart P, Vega-Ramirez H, Miake-Lye S, Whalen M, Nelson M, Bergami M, Bartsch D, Hen R, Berninger B, Sahay A (2016) Modulating neuronal competition dynamics in the dentate gyrus to rejuvenate aging memory circuits. Neuron 91:1356–1373. 10.1016/j.neuron.2016.08.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller JA, Nathanson J, Franjic D, Shim S, Dalley RA, Shapouri S, Smith KA, Sunkin SM, Bernard A, Bennett JL, Lee CK, Hawrylycz MJ, Jones AR, Amaral DG, Sestan N, Gage FH, Lein ES (2013) Conserved molecular signatures of neurogenesis in the hippocampal subgranular zone of rodents and primates. Development 140:4633–4644. 10.1242/dev.097212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monje ML, Vogel H, Masek M, Ligon KL, Fisher PG, Palmer TD (2007) Impaired human hippocampal neurogenesis after treatment for central nervous system malignancies. Ann Neurol 62:515–520. 10.1002/ana.21214 [DOI] [PubMed] [Google Scholar]
- Moreno-Jiménez EP, Flor-García M, Terreros-Roncal J, Rábano A, Cafini F, Pallas-Bazarra N, Ávila J, Llorens-Martín M (2019) Adult hippocampal neurogenesis is abundant in neurologically healthy subjects and drops sharply in patients with Alzheimer's disease. Nat Med 25:554–560. 10.1038/s41591-019-0375-9 [DOI] [PubMed] [Google Scholar]
- Nakashiba T, Cushman JD, Pelkey KA, Renaudineau S, Buhl DL, McHugh TJ, Rodriguez Barrera V, Chittajallu R, Iwamoto KS, McBain CJ, Fanselow MS, Tonegawa S (2012) Young dentate granule cells mediate pattern separation, whereas old granule cells facilitate pattern completion. Cell 149:188–201. 10.1016/j.cell.2012.01.046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ngwenya LB, Heyworth NC, Shwe Y, Moore TL, Rosene DL (2015) Age-related changes in dentate gyrus cell numbers, neurogenesis, and associations with cognitive impairments in the rhesus monkey. Front Syst Neurosci 9:102. 10.3389/fnsys.2015.00102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nottebohm F (1981) A brain for all seasons: cyclical anatomical changes in song control nuclei of the canary brain. Science 214:1368–1370. 10.1126/science.7313697 [DOI] [PubMed] [Google Scholar]
- Nowakowski RS, Lewin SB, Miller MW (1989) Bromodeoxyuridine immunohistochemical determination of the lengths of the cell cycle and the DNA-synthetic phase for an anatomically defined population. J Neurocytol 18:311–318. 10.1007/BF01190834 [DOI] [PubMed] [Google Scholar]
- Overstreet Wadiche L, Bromberg DA, Bensen AL, Westbrook GL (2005) GABAergic signaling to newborn neurons in dentate gyrus. J Neurophysiol 94:4528–4532. 10.1152/jn.00633.2005 [DOI] [PubMed] [Google Scholar]
- Overstreet-Wadiche LS, Bromberg DA, Bensen AL, Westbrook GL (2006) Seizures accelerate functional integration of adult-generated granule cells. J Neurosci 26:4095–4103. 10.1523/JNEUROSCI.5508-05.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paredes MF, Sorrells SF, Cebrian-Silla A, Sandoval K, Qi D, Kelley KW, James D, Mayer S, Chang J, Auguste KI, Chang EF, Gutierrez Martin AJ, Kriegstein AR, Mathern GW, Oldham MC, Huang EJ, Garcia-Verdugo JM, Yang Z, Alvarez-Buylla A (2018) Does adult neurogenesis persist in the human hippocampus? Cell Stem Cell 23:780–781. 10.1016/j.stem.2018.11.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parent JM, Yu TW, Leibowitz RT, Geschwind DH, Sloviter RS, Lowenstein DH (1997) Dentate granule cell neurogenesis is increased by seizures and contributes to aberrant network reorganization in the adult rat hippocampus. J Neurosci 17:3727–3738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paton JA, Nottebohm FN (1984) Neurons generated in the adult brain are recruited into functional circuits. Science 225:1046–1048. 10.1126/science.6474166 [DOI] [PubMed] [Google Scholar]
- Perera TD, Coplan JD, Lisanby SH, Lipira CM, Arif M, Carpio C, Spitzer G, Santarelli L, Scharf B, Hen R, Rosoklija G, Sackeim HA, Dwork AJ (2007) Antidepressant-induced neurogenesis in the hippocampus of adult nonhuman primates. J Neurosci 27:4894–4901. 10.1523/JNEUROSCI.0237-07.2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perry EK, Johnson M, Ekonomou A, Perry RH, Ballard C, Attems J (2012) Neurogenic abnormalities in Alzheimer's disease differ between stages of neurogenesis and are partly related to cholinergic pathology. Neurobiol Dis 47:155–162. 10.1016/j.nbd.2012.03.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piatti VC, Davies-Sala MG, Esposito MS, Mongiat LA, Trinchero MF, Schinder AF (2011) The timing for neuronal maturation in the adult hippocampus is modulated by local network activity. J Neurosci 31:7715–7728. 10.1523/JNEUROSCI.1380-11.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao MS, Shetty AK (2004) Efficacy of doublecortin as a marker to analyse the absolute number and dendritic growth of newly generated neurons in the adult dentate gyrus. Eur J Neurosci 19:234–246. 10.1111/j.0953-816x.2003.03123.x [DOI] [PubMed] [Google Scholar]
- Revest JM, Dupret D, Koehl M, Funk-Reiter C, Grosjean N, Piazza PV, Abrous DN (2009) Adult hippocampal neurogenesis is involved in anxiety-related behaviors. Mol Psychiatry 14:959–967. 10.1038/mp.2009.15 [DOI] [PubMed] [Google Scholar]
- Rhodes JS, van Praag H, Jeffrey S, Girard I, Mitchell GS, Garland T Jr, Gage FH (2003) Exercise increases hippocampal neurogenesis to high levels but does not improve spatial learning in mice bred for increased voluntary wheel running. Behav Neurosci 117:1006–1016. 10.1037/0735-7044.117.5.1006 [DOI] [PubMed] [Google Scholar]
- Riphagen JM, Schmiedek L, Gronenschild E, Yassa MA, Priovoulos N, Sack AT, Verhey FR, Jacobs HI (2020) Associations between pattern separation and hippocampal subfield structure and function vary along the lifespan: a 7 T imaging study. Sci Rep 10:7572. 10.1038/s41598-020-64595-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sahay A, Scobie KN, Hill AS, O'Carroll CM, Kheirbek MA, Burghardt NS, Fenton AA, Dranovsky A, Hen R (2011) Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature 472:466–470. 10.1038/nature09817 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S, Weisstaub N, Lee J, Duman R, Arancio O, Belzung C, Hen R (2003) Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 301:805–809. 10.1126/science.1083328 [DOI] [PubMed] [Google Scholar]
- Schinder AF, Gage FH (2004) A hypothesis about the role of adult neurogenesis in hippocampal function. Physiology (Bethesda) 19:253–261. 10.1152/physiol.00012.2004 [DOI] [PubMed] [Google Scholar]
- Schmidt-Hieber C, Jonas P, Bischofberger J (2004) Enhanced synaptic plasticity in newly generated granule cells of the adult hippocampus. Nature 429:184–187. 10.1038/nature02553 [DOI] [PubMed] [Google Scholar]
- Seki T, Hori T, Miyata H, Maehara M, Namba T (2019) Analysis of proliferating neuronal progenitors and immature neurons in the human hippocampus surgically removed from control and epileptic patients. Sci Rep 9:18194. 10.1038/s41598-019-54684-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seri B, García-Verdugo JM, McEwen BS, Alvarez-Buylla A (2001) Astrocytes give rise to new neurons in the adult mammalian hippocampus. J Neurosci 21:7153–7160. 10.1523/JNEUROSCI.21-18-07153.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sierra A, Encinas JM, Maletic-Savatic M (2011) Adult human neurogenesis: from microscopy to magnetic resonance imaging. Front Neurosci 5:47. 10.3389/fnins.2011.00047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snyder JS (2019) Recalibrating the relevance of adult neurogenesis. Trends Neurosci 42:164–178. 10.1016/j.tins.2018.12.001 [DOI] [PubMed] [Google Scholar]
- Snyder JS, Soumier A, Brewer M, Pickel J, Cameron HA (2011) Adult hippocampal neurogenesis buffers stress responses and depressive behaviour. Nature 476:458–461. 10.1038/nature10287 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sorrells SF, Paredes MF, Cebrian-Silla A, Sandoval K, Qi D, Kelley KW, James D, Mayer S, Chang J, Auguste KI, Chang EF, Gutierrez AJ, Kriegstein AR, Mathern GW, Oldham MC, Huang EJ, Garcia-Verdugo JM, Yang Z, Alvarez-Buylla A (2018) Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature 555:377–381. 10.1038/nature25975 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spalding KL, Bergmann O, Alkass K, Bernard S, Salehpour M, Huttner HB, Bostrom E, Westerlund I, Vial C, Buchholz BA, Possnert G, Mash DC, Druid H, Frisen J (2013) Dynamics of hippocampal neurogenesis in adult humans. Cell 153:1219–1227. 10.1016/j.cell.2013.05.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stanfield BB, Trice JE (1988) Evidence that granule cells generated in the dentate gyrus of adult rats extend axonal projections. Exp Brain Res 72:399–406. 10.1007/BF00250261 [DOI] [PubMed] [Google Scholar]
- Stȩpień T, Tarka S, Chutorański D, Felczak P, Acewicz A, Wierzba-Bobrowicz T (2018) Neurogenesis in adult human brain after hemorrhage and ischemic stroke. Folia Neuropathol 56:293–300. 10.5114/fn.2018.80862 [DOI] [PubMed] [Google Scholar]
- Suwabe K, Byun K, Hyodo K, Reagh ZM, Roberts JM, Matsushita A, Saotome K, Ochi G, Fukuie T, Suzuki K, Sankai Y, Yassa MA, Soya H (2018) Rapid stimulation of human dentate gyrus function with acute mild exercise. Proc Natl Acad Sci USA 115:10487–10492. 10.1073/pnas.1805668115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tartt AN, Fulmore CA, Liu Y, Rosoklija GB, Dwork AJ, Arango V, Hen R, Mann JJ, Boldrini M (2018) Considerations for assessing the extent of hippocampal neurogenesis in the adult and aging human brain. Cell Stem Cell 23:782–783. 10.1016/j.stem.2018.10.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Temprana SG, Mongiat LA, Yang SM, Trinchero MF, Alvarez DD, Kropff E, Giacomini D, Beltramone N, Lanuza GM, Schinder AF (2015) Delayed coupling to feedback inhibition during a critical period for the integration of adult-born granule cells. Neuron 85:116–130. 10.1016/j.neuron.2014.11.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tobin MK, Musaraca K, Disouky A, Shetti A, Bheri A, Honer WG, Kim N, Dawe RJ, Bennett DA, Arfanakis K, Lazarov O (2019) Human hippocampal neurogenesis persists in aged adults and Alzheimer's disease patients. Cell Stem Cell 24:974–982.e973. 10.1016/j.stem.2019.05.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toni N, Teng EM, Bushong EA, Aimone JB, Zhao C, Consiglio A, van Praag H, Martone ME, Ellisman MH, Gage FH (2007) Synapse formation on neurons born in the adult hippocampus. Nat Neurosci 10:727–734. 10.1038/nn1908 [DOI] [PubMed] [Google Scholar]
- Toni N, Laplagne DA, Zhao C, Lombardi G, Ribak CE, Gage FH, Schinder AF (2008) Neurons born in the adult dentate gyrus form functional synapses with target cells. Nat Neurosci 11:901–907. 10.1038/nn.2156 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trinchero MF, Buttner KA, Sulkes Cuevas JN, Temprana SG, Fontanet PA, Monzon-Salinas MC, Ledda F, Paratcha G, Schinder AF (2017) High plasticity of new granule cells in the aging hippocampus. Cell Rep 21:1129–1139. 10.1016/j.celrep.2017.09.064 [DOI] [PubMed] [Google Scholar]
- Tronel S, Charrier V, Sage C, Maitre M, Leste-Lasserre T, Abrous DN (2015) Adult-born dentate neurons are recruited in both spatial memory encoding and retrieval. Hippocampus 25:1472–1479. 10.1002/hipo.22468 [DOI] [PubMed] [Google Scholar]
- van Praag H, Kempermann G, Gage FH (1999) Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat Neurosci 2:266–270. 10.1038/6368 [DOI] [PubMed] [Google Scholar]
- van Praag H, Kempermann G, Gage FH (2000) Neural consequences of environmental enrichment. Nat Rev Neurosci 1:191–198. 10.1038/35044558 [DOI] [PubMed] [Google Scholar]
- van Praag H, Schinder AF, Christie BR, Toni N, Palmer TD, Gage FH (2002) Functional neurogenesis in the adult hippocampus. Nature 415:1030–1034. 10.1038/4151030a [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waldvogel H, Curtis M, Baer K, Rees M, Faull R (2006) Immunohistochemical staining of post-mortem adult human brain sections. Nat Protoc 1:2719–2732. [DOI] [PubMed] [Google Scholar]
- Wiget F, van Dijk RM, Louet ER, Slomianka L, Amrein I (2017) Effects of strain and species on the septo-temporal distribution of adult neurogenesis in rodents. Front Neurosci 11:719. 10.3389/fnins.2017.00719 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan TF, Li J, Ding F, Arias-Carrion O (2014) Evidence of adult neurogenesis in non-human primates and human. Cell Tissue Res 358:17–23. 10.1007/s00441-014-1980-z [DOI] [PubMed] [Google Scholar]
- Zhao C, Teng EM, Summers RG Jr, Ming GL, Gage FH (2006) Distinct morphological stages of dentate granule neuron maturation in the adult mouse hippocampus. J Neurosci 26:3–11. 10.1523/JNEUROSCI.3648-05.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]