

Abstract
The review discusses a new approach to the prevention and treatment of viral infections based on the use of pine needles polyprenyl phosphate (PPP) and associated with the infringement of prenylation process—the attachment of farnesol or geranyl geraniol to the viral protein. Currently, prenylation has been detected in type 1 adenovirus, hepatitis C virus, several herpes viruses, influenza virus, HIV. However, this list is far from complete, given that prenylated proteins play an extremely important role in the activity of the virus. We assume that the interferon produced in response to PPP may suppress expression of the SREBP2 transcription factor. As a result, the mevalonic acid pathway is violated and, as a result, the formation of early polyprenols precursors (geraniol, geranyl geraniol, farnesol), which are necessary for the prenylation of viral proteins, is blocked and the formation of mature, virulent virus particles is broken. As a consequence, the maturation of viral particles is inhibited, and defective particles are formed. Polyprenol was extracted from greenery (pine, fir and spruce needles, mulberry leaves, etc.), purified by chromatography, phosphorylated and identified by HPLC and NMR. Obtained PPP was used as antiviral in some experimental models in vitro and in vivo. During numerous studies, it was found that PPP manifested versatile antiviral effects, both in vitro and in vivo. The maximum effect was observed with viruses in which the presence of prenylated proteins was established, namely influenza A virus, HIV-1, tick-borne encephalitis virus, hepatitis A and C viruses, herpes simplex viruses type 1 and 2, some coronavirus. The available data obtained both in the experimental conditions and during clinical trials allow us to regard PPPs as safe and effective medicine for prevention and treatment of viral diseases.
Keywords: Plant polyprenols, Prenylation, Viral infection, Interferons, SREBP2, Coronavirus
Introduction
About 20% of population mortality is associated with infectious diseases, in which significant place is occupied by viral infections. According to the World Health Organization (WHO), 37 million of people in the world get sick with HIV every year, 257 million–with hepatitis B virus (HBV), 70 million–with hepatitis C virus (HCV; viral hepatitis) and 3–5 million–with flu (Naesens et al. 2016; WHO 2007–2017). Vaccination is considered as the best way to protect population against viral infections. However, vaccines may often lose effectiveness in the world of rapidly evolving pathogens. Or they have not yet been developed, as has recently happened with such emerging infections as Ebola, Zika or SARS-Cov-2. That is why scientists have been searching for effective antiviral medicines for more than 50 years. To date, the number of officially registered antiviral drugs counts more than one hundred and their number continues to increase at an average rate of 4–6 new drugs per year. Almost half of the medicines are used to treat HIV infection, one fifth–HCV, 9%–HBV, 12% are specific for herpes virus infections, 5%–for influenza virus, and the remaining infections account for 6% (Chaudhuri et al. 2018; Ianevski et al. 2018).
Unfortunately, as antiviral drugs are used, resistance begins to develop against them. One way to reduce the rate of appearance of the resistant pathogens is the use of various combinations of antiviral agents, and the trend towards the use of combination therapy has intensified in recent years (Chaudhuri et al. 2018). With this aim, a constant search of new targets is carried to inhibit the reproduction of viruses (or to affect the life cycle of the virus). Of the antiviral agents that exist today, 38% act on the polymerase, 24%–on viral proteases, 8%–on non-structural HCV protein 5A, in the treatment of which outstanding results have been achieved in recent years, 4% are aimed against integrase, while 14% have the M2 ion channel, neuraminidase, hemagglutinin and influenza nucleoprotein as their targets. The remaining 12% of the drugs affect the host, not the viruses. This group includes mainly interferons (IFNs) and their inducers (Chaudhuri et al. 2018; Ershov and Narovlyansky 2015; Kanda et al. 2017). This review focuses on the group of preventive and therapeutic medicines, the main active substance of which are polyprenols.
Prenols and Post-Translational Protein Modification
Prenols, and more generally polyisoprenoids, are present in all living organisms and consist of repeating isoprene units. They are synthesized in the mevalonic acid pathway (Fig. 1).
Fig. 1.

Mevalonate pathway and stages of isoprenoid synthesis. OPP oxygen-pyrophosphate, HMG-CoA 3-hydroxy-3-methylglutaryl-CoA
This pathway begins with hydroxymethylglutaryl coenzyme A, which is converted by hydroxymethylglutaryl coenzyme A reductase to mevalonate, from which isopentinyl pyrophosphate (IPP) is then formed and isomerized to dimethylallyl pyrophosphate. Then, a number of new IPP links sequentially begin to attach to the latter, as a result of which geranyl PP and farnesyl PP are formed. Two farnesol molecules, connected head to head, form squalene, from which cholesterol is synthesized. If IPP units continue to link with the farnesyl PP, according to “the head-to-tail” principle, then the polyprenyl pyrophosphate is formed as a result, and dolichol and dolichyl PP are formed from it. In addition, such a polyprenol chain serves as the basis for the synthesis of ubiquinone (Jones et al. 2009; Morita et al. 2007; Zhang et al. 2015).
Free polyprenols are metabolically inactive, while phosphorylated polyprenols serve as intermediate sugar acceptors for protein glycosylation (Hartley and Imperiali 2012; Welti 2013; Zhang et al. 2015).
Farnesil PP, geranyl geranyl PP and geranyl PP are used to prenylate small ATPases (R proteins) and early viral proteins. The process consists of three stages: first, the farnesyl or geranyl geranyl group is attached to the 4th cysteine residue from the C-terminus (Fig. 2).
Fig. 2.

Posttranslational prenylation of proteins
Fig. 2–A is any aliphatic amino acid, and X–any amino acid.
Then, RCE1 protease cleaves the last three amino acids and finally methyltransferase methylates cysteine. Currently, this process has been detected in such viruses as type 1 adenovirus, HCVs, several herpes viruses, influenza virus, HIV (Table 1) (Marakasova et al. 2017).
Table 1.
Viruses with prenylated proteins or sensitivity to statins
| Viruses | Prenylated proteins | Inhibition of a replication with statins |
|---|---|---|
| Human adenovirus 1 | 6 kDa protein/E1A | |
| Hepatitis delta virus | Large delta antigen | |
| Hepatitis C virus | Non-structural 5A protein | + |
| Human herpesvirus 5 (CMV) | Uncharacterized protein IRL9 | + |
| Influenza A virus | PB2 protein | + |
| Human parainfluenza virus 3 | Hemagglutinin-neuraminidase | |
| Molluscum contagiosum virus subtype 1 | MC155R | |
| Human immunodeficiency virus 1 | Nef protein | + |
| Human herpesvirus (HSV) 1, 2 | UL32 protein | |
| Pseudorabies virus (Aujeszky’s disease) | Us2 | + |
| Respiratory syncytial virus | Host RhoA | + |
| Rotavirus | Unknown | + |
| Dengue virus (DENV), japanese encephalitis virus (JEV), West Nile virus (WNV), tick-borne encephalitis virus (TBEV) | Unknown | + |
However, this list is far from complete, given the role that prenylated proteins play in the life cycle of the virus: binding to the cell, penetration into the cell and into the nucleus, replication of the viral genome, as well as transcription processes, using either the RNA matrix or the DNA matrix. Finally, this also includes functions related to the suppression of host defense systems (Einav and Glenn 2003; Marakasova et al. 2017; Palsuledesai and Distefano 2015).
Antiviral Activity of Polyprenyl Phosphates
During numerous studies, it was found that polyprenyl phosphate (PPP) obtained from coniferous trees needles with the general formula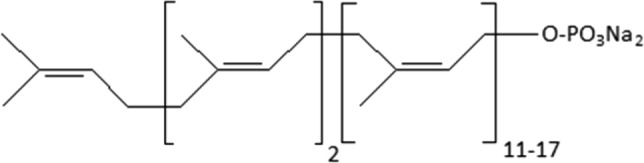 exhibited broad antiviral activity, both in vitro and in vivo (Narovlyansky et al. 2012, 2014, 2018). If we arrange all studied viral infections by their sensitivity to PPP, taking as a basis the efficiency coefficient (in case of in vivo experiments, the latter is estimated by the difference in mortality in the control and experimental groups, divided by the mortality in the control group, and for in vitro experiments–by the difference in virus titers in control and experimental groups, divided by the titer of the virus in the control), then the following pattern may be observed (Table 2).
exhibited broad antiviral activity, both in vitro and in vivo (Narovlyansky et al. 2012, 2014, 2018). If we arrange all studied viral infections by their sensitivity to PPP, taking as a basis the efficiency coefficient (in case of in vivo experiments, the latter is estimated by the difference in mortality in the control and experimental groups, divided by the mortality in the control group, and for in vitro experiments–by the difference in virus titers in control and experimental groups, divided by the titer of the virus in the control), then the following pattern may be observed (Table 2).
Table 2.
Efficiency of PPP antiviral action
| Viruses | Concentration/dosage of the PPP | Route of administration | Efficiency coefficient (Ec)* |
||
|---|---|---|---|---|---|
| Family | Genus | Species | |||
| Paramixoviridae | Morbillivirus | Canine distemper virus | 5–20 mcg/mouse | In vitro | 1 |
| Orthomyxoviridae | Orthomyxovirus | Influenza A virus | 200 mcg/ml | In vitro | 1 |
| Retroviridae | Oncovirus type C | HIV-1 | 500 mcg/ml | in vitro | 0.98 |
| Togaviridae | Flavivirus | Tick born encephalitis virus | 500 mcg | i/c | 0.98 |
| Picornaviridae | Enterovirus | Hepatitis A virus | 4 mcg/ml | In vitro | 0.91 |
| Herpesviridae | Varicellovirus | Aujeszky’s disease virus | 10–20 mg/rabbit | s/c, per os | 0.9 |
| Herpesviridae | Simplexvirus | Herpes simplex virus type 1 | 100 mcg/mouse | i/m | 0.81 |
| Coronaviridae | Coronavirus | Mouse hepatitis virus | 200 mcg/mouse | Per os | 0.75 |
| Orthomyxoviridae | Orthomyxovirus | Influenza A virus | 5 mcg/mouse | i/m | 0.67 |
| Togaviridae | Alphavirus | Western equine encephalomyelitis virus | 100 mcg /mouse | s/c | 0.63 |
| Rhabdoviridae | Lissavirus | Rabies virus | 200 mcg /mouse | i/c | 0.6 |
| Togaviridae | Flavivirus | Yellow fever virus | 200 mcg /ml | In vitro | 0.6 |
| Retroviridae | Oncovirus type 1 | Bovine leukemia virus | 200 mcg /ml | In vitro | 0.563 |
| Adenoviridae | Mastadenovirus | Canine adenovirus type I | 200 mcg /ml | In vitro | 0.471 |
| Paramyxoviridae | Paramyxovirus | Mumps virus | 200 mcg /ml | in vitro | 0.22 |
| Paramyxoviridae | Morbillivirus | Measles virus | 200 mcg /ml | In vitro | 0.14 |
*Ec = (A–B)/A, where A means virus titer (lethality) in control groups and B means virus titer (lethality) in groups treated with PPP. Viruses with prenylated proteins are highlighted in grey
s/c subcutaneous, i/m intramuscular, i/c intracutaneous
Viruses for which prenylated proteins are known are highlighted in grey. Obviously, PPP exerts the most antiviral activity towards viruses belonging to this group (the only exception is canine distemper virus, for which existence of prenylated proteins is not shown).
Result of PPP interaction with the tick-borne encephalitis virus of the Sofjin strain is shown in the Fig. 3.
Fig. 3.

Formation of defective viral particles after exposure to PPP. Tick-borne encephalitis virions, 2 h after the addition of a placebo (bottom photo) or PPP (top photo)
Here, electronic microphotographs of the virus suspension are presented. Most virions treated with PPP had a loose or damaged envelope. There were also small particles with a diameter of 20 nm (possibly virions devoid of envelope). Such particles are not able to cause infection in animals. In control samples treated with a placebo solution, such virions were absent. So, after treatment of infected cell cultures with PPP, the synthesis of early viral proteins is suppressed and defective interfering virus particles are formed (Ozherelkov et al. 2000).
All these data served as the basis for the use of PPP as an antiviral drug. In Russia, the first PPP medicine was approved for use in veterinary medicine in 1996 (Danilov et al. 1996). Later, in the early 2000s, a similar technology of producing a veterinary drug was developed using mulberry leaves polyprenols (Legendre et al. 2009, 2017a; b; Kuritz 2009).
Possible Mechanisms of PPP Antiviral Activity
It was shown that the administration of PPP in vivo stimulated production of type 1 and type 2 interferons (Pronin et al. 2000, 2002) and launched IFN-mediated suppression of isoprenoid metabolites synthesis in the mevalonate pathway (Blanc et al. 2011). Also, PPP was shown to inhibit production of the sterol regulatory element-binding protein-2 (SREBP2) transcription factor in mice (Fig. 4).
Fig. 4.

Content of SREBP2 in the liver lysate of mice after PPP administration
It is known that following interaction with IFNR1 receptors IFN also causes the suppression of SREBP2 (Blanc et al. 2011)–the main regulator of the mevalonic acid pathway, and a key transcription factor regulating the production of the earliest polyprenols precursors–geraniol, farnesol and geranyl geraniol. SREBP2 controls synthesis of hydroxymethylglutaryl coenzyme A synthase and reductase, as well as farnesyl synthase. Squalene, which serves as the main precursor of cholesterol, is then formed from farnesol, as mentioned above, and the blockade of SREBP2 ultimately leads to suppression of cholesterol synthesis. The earliest precursors of polyprenols—geraniol, farnesol, and to a lesser extent geranyl geraniol—are also necessary for the prenylation of viral proteins. Without them, the normal assembly of the viral capsid is violated and as a result leading to the formation of defective, non-virulent particles (Sato 2010).
At least for some of the PPP-susceptible viruses, a good therapeutic effect is achieved after using of statins (Table 1), the main therapeutic drugs used for dyslipidemia correction in case of increased blood cholesterol levels. The target of statins is hydroxymethylglutaryl coenzyme A reductase, a key enzyme of the mevalonate pathway involved in the synthesis of not only cholesterol, but also the early prenols, farnesol and geranyl geraniol involved in the prenylation of viral proteins (Bogovič et al. 2018). In some cases (respiratory syncytial virus), the therapeutic effect of statins is at least partly mediated through the blocking of prenylation of P proteins which are also involved in the proper assembly of virions (Gower and Graham 2001; Martínez-Gutierrez et al. 2011).
Experiments performed on hyperlipidemic mice after administration of Poloxamer 407 showed (Pronin et al. 2014a) that PPP combined with beta-sitosterol, another polyisoprenoid that blocks intestinal cholesterol absorption, reduced blood cholesterol and triglycerides levels. This effect was comparable to the action of atorvastatin. A similar effect was obtained in dogs with type 1 diabetes (Annikov et al. 2019).
As shown in Fig. 5, another mechanism of PPP action is associated with a direct dose-dependent inhibitory effect on the activity of 5- and 15-lipoxygenases (Sanin et al. 2011a).
Fig. 5.

Effect of PPP on lipoxygenase-5 activity
Soybean lipoxygenase-5 activity was assayed with linoleic acid as the substrate at a room temperature, and oxygen concentration was monitored polarigraphically with Clark oxygen electrode (Pronin et al. 2002).
Lipoxygenase-5 is known to catalyze the formation of leukotriene B4 from arachidonic acid, which, after interacting with its receptor, inhibits the activity of phospholipase D2, which ensures the formation of phosphatidyl acid from phosphatidylserine. Phosphatidyl acid, in turn, activates NADPH oxidase, the main inducer of the oxidative burst and the associated manifestations of the inflammatory process (Bonnans and Levy 2007).
Since PPP in some respects behaves like a lectin specific for glycoproteins containing mannose (Sobolev et al. 2010), it may inhibit binding of the viruses to their receptors. Following direct treatment with PPP, the viral envelope is prone to disintegrate. PPP also induces a calcium signal, NF-κB activation, production of type 1 and type 2 interferons and tumor necrosis factor α (Danilov et al. 2002; Kuritz 2009; Pronin et al. 2014b). In addition, it is assumed that PPP directly inhibits the synthesis of prenols necessary for viral replication (Narovlyansky et al. 2018), and its excess leads to the formation of dolichyl phosphates (an important product of human cells necessary for glycosylation of proteins and maintaining the structure of cell membranes). Dolichyl phosphates, in their turn, induce additional synthesis of interferons, which lead to a “cyclization” of the process and to an increase of the antiviral effect.
The Effectiveness of PPP in the Treatment of Viral Infections in Animals
A far from complete assessment of the effectiveness of PPP in the treatment of domestic animals is shown in Table 3.
Table 3.
Efficiency of PPP in the treatment of viral infections in dogs and cats
| Pets | Diseases | Total | Cured | % |
|---|---|---|---|---|
| Dogs | Canine distemper | 300 | 140 | 70 |
| Parvovirus enteritis | 156 | 140 | 90 | |
| Infectious hepatitis and adenovirosis | 96 | 91 | 95 | |
| Viral papillomatosis | 25 | 24 | 96 | |
| Cats | Panleukopenia | 240 | 204 | 85 |
| Calicivirosis | 190 | 186 | 98 | |
| Infectious rhinotracheitis | 280 | 277 | 99 | |
| Feline infectious peritonitis | 18 | 7 | 39 |
The drug has shown a good results in the treatment of canine distemper, parvoviral enteritis, infectious hepatitis, adenovirosis and papillomatosis in dogs (Ozherelkov et al. 2001; Narovlyansky et al. 2018), as well as panleukopenia, calicivirosis, herpes viral rhinotracheitis and coronaviral infectious peritonitis in cats. With a fatal wet form of feline infectious peritonitis induced by coronavirus in cats, complete remission was observed after the use of PPP in several animals. Some other coronaviral infections of animals are also treated with PPP (viral gastroenteritis in pigs, coronaviral enteritis in dogs, infectious bronchitis in chickens).
In several model systems, PPP has been shown to be effective against herpes viruses of types 1 and 2 (Fig. 6):
In vitro, the drug inhibited by 80–96% symplast formation in the culture of Vero cells infected with herpes simplex virus (HSV) type 1 (Vasil’ev et al. 2008).
In vivo in mice with meningoencephalitis caused by HSV type 1, PPP inoculation led to decrease in mortality by 47–50% (Narovlyansky et al. 2015a, 2018).
In rabbits infected with the Aujeszky disease virus, mortality decreased by 90% (in the prophylactic treatment protocol) or by 47% (in the therapeutic treatment protocol) (Sanin et al. 2011b).
In male guinea pigs infected with HSV type 2 (experimental genital herpes model), the total symptom severity index lessened by 13 points, the duration of the disease decreased by 4.3 days (Narovlyansky 2015a, d).
Experimental infection of HSV-2 in female guinea pigs during puberty is manifested by an irreversible violation of ovogenesis in reproductive age–a progressive decrease in the total population of follicles and oocytes, incomplete ovo-folliculogenesis, anovulation and infertility (Fig. 7) (Borovaya et al. 2015).
Fig. 6.

PPP anti-herpetic activity. a treatment of herpetic meningoencephalitis (HSV type 1) in mice; b treatment of Aujeszky’s disease in rabbits; c treatment of experimental genital herpes infection (HSV type 2) in guinea pigs. The following signs of the disease were recorded: edema and hyperemia of the urethra and external genital organs; vesicular rash, exanthema, papules, pustules; erosive and erosive-ulcerative lesions, herpetic crusts; vaginal discharge; violation of the resorption of the vaginal contents of the vaginal mucosa. These symptoms were scored in the form of both individual symptoms and the symptom complexes, where 1 symptom is equal to 1 point. The effectiveness of the treatment was evaluated by changing the specific manifestations of the clinical signs of the disease localized on the female genitalia. TDSI means the total disease severity index, TEI means the therapeutic effect index, ADD means the average duration of the disease. d Inhibition of the symplast formation in Vero cells infected with HSV type 1. Mean inhibition percent 87 ± 1.81 (M ± m), p < 0.001
Fig. 7.

Follicles of the ovary of a guinea pig infected with herpes simplex virus type 2 (a) and after treatment with PPP (b). Hematoxylin–eosin staining
The injection of PPP during the experimental infection with HSV-2 prevented subsequent spontaneous exacerbations, helped to preserve the population of germ cells in the ovaries, as well as a full-fledged process of ovo-folliculogenesis and sexual cycles (Fig. 7).
PPP as a Prospective Antiherpetic Remedy in Humane Medicine
The obtained data justified the development of an anti-herpetic drug based on PPP. Three phases of clinical trials were carried out. The drug showed an acceptable safety profile and a good level of tolerance. The study was carried out on 62 patients with genital herpes who underwent the standard protocol of therapy with acyclovir following which they were given either three PPP injections, or three injection of placebo, after which they were observed for 90 days (Narovlyansky et al. 2015b, 2015c; Ershov et al. 2017; Pronin et al. 2016; Sedov et al. 2018).
The results obtained showed that (Table 4):
The duration of the inter-relapse period (remission period) in patients who were treated with PPP statistically significantly increased from 29.36 ± 2.16 days (three months before the start of the study) to 42.98 ± 3.29 days after treatment (p = 0.0002).
The relapse rate of genital herpes throughout the study period (three months) significantly decreased in the group of patients treated with PPP and amounted to 1.94 ± 0.19, while the average number of relapses before treatment was 3.03 ± 0.02 (p < 0.05).
The total score of relapse according to local (itching, burning, soreness, hyperemia), general (weakness, headache, chills, neuralgia, myalgia) signs of relapse, body temperature, number of lesions, area of lesions, the number of vesicular erosive elements in the lesion in the treatment group decreased from 7.45 ± 0.36 points at the first visit (screening) before treatment to 4.79 ± 0.55 points during treatment (p = 0.0002).
An increase in the duration of the inter-relapse period and a decrease in the frequency of relapses of herpes simplex in patients of the treatment group compared with the control group (placebo) were accompanied by a change in the level of leukocyte virus-induced interferon (LVI-IFN). In the treatment group, along with a decrease in the frequency of relapses of genital herpes in 70% of patients, the level of IFI-LVI remained unchanged or increased in 64.4% of patients. In the control group, with a decrease in the relapse rate in only 55% of patients, the level of LVI-IFN remained unchanged or increased only in 36.8%.
Table 4.
Therapeutic efficacy of PPP in the treatment of genital herpes
| Parameter studied | Control group | Treatment group |
|---|---|---|
| Remission period (days ± m) | 29.36 ± 2.16 | 42.98 ± 3.29; p = 0.0002 |
| Relapse rate (M ± m) | 3.03 ± 0.02 | 1,94 ± 0.19; p < 0.05 |
| The total score of relapse (points ± m) | 7.45 ± 0.36 | 4.79 ± 0.55; p = 0.0002 |
| Patients with increased or unchanged level of leukocyte virus-induced IFN | 36.8 | 64.4 |
M mean value, m standard error
Conclusion
Phosphorylated polyprenols from coniferous trees needles inhibit the transcription factor SREBP2, which plays a key role in the synthesis of early prenols (farnesol and geranyl geraniol) necessary for post-transcriptional modification of viral proteins. As a result, inoculation of PPP violates the correct assembly of viral particles.
Phosphorylated polyprenols have a wide spectrum of antiviral activity, which is more expressed against viruses with prenylated proteins.
The available data obtained both in the experimental conditions and during clinical trials allow us to regard PPPs as safe and effective medicine for prevention and treatment of viral diseases, as well as for correction of dyslipidemia (Table 5).
Table 5.
Possible benefits and drawbacks of the potential use of PPP in humans and animals
| Advantages | Disadvantages |
|---|---|
| Susceptibility of many viruses to the medicine | Maximum effect early in the infection |
| Resistance to the medicine is not developed | Possible short-term temperature rise |
| Low toxicity (LD 50 12.5 g / kg) | Poor water solubility |
| Poor antigenicity | Stability of an aqueous solution in the alkaline zone (pH 8.5–10.5) |
| Anti-inflammatory activity | Possible local reactions when injected (soreness, redness) |
| Innate immunity activation |
Acknowledgements
Part of the work was carried out with the financial support from the Ministry of Industry and Trade and the Ministry of Education and Science of the Russian Federation under the Federal Target Program “Development of the Pharmaceutical and Medical Industry of the Russian Federation for the Period Until 2020 and Beyond” and the financial support from Micro-Plus CJSC and GamaVetPharm LLC.
Abbreviations
- IFN
Interferon
- HPLC
High-Performance Liquid Chromatography
- HSV
Herpes Simplex Virus
- NMR
Nuclear Magnetic Resonance
- PPP
Polyprenyl Phosphate
- SREBP2
Sterol Regulatory Element-Binding Protein-2
- IPP
Isopentinyl Pyrophosphate
Declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Annikov VV, Narovyansky AN, Sanin AV, et al. Correction of dyslipidemia with combined use of beta-sitosterol and polyprenyl phosphates in dogs with type 1 diabetes mellitus. Russian Veterinary J. 2019;5:12–15. doi: 10.32416/article_5d935e17e234b1.31437966. [DOI] [Google Scholar]
- Blanc M, Hsieh WY, Robertson KA. Host defense against viral infection involves interferon mediated down-regulation of sterol biosynthesis. PLoS Biol. 2011;9:e1000598. doi: 10.1371/journal.pbio.1000598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bogovič P, Lusa L, Stupica D, et al. Impact of pre-existing treatment with statins on the course and outcome of tick-borne encephalitis. PLoS ONE. 2018;13:e0204773. doi: 10.1371/journal.pone.0204773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonnans C, Levy BD. Lipid mediators as agonists for the resolution of acute lung inflammation and injury. Am J Respir Cell Mol Biol. 2007;36:201–205. doi: 10.1165/rcmb.2006-0269TR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borovaya TG, Didenko LV, Narovlyansky AN, et al. Histological characteristics of rete ovarii in acute period of genital herpes virus infection. Morphol Newsl. 2015;2:15–20 . doi: 10.20340/mv-mn.2015.0(2):15-20. [DOI] [Google Scholar]
- Chaudhuri S, Symons JA, Deval J. Innovation and trends in the development and approval of antiviral medicines: 1987–2017 and beyond. Antiviral Res. 2018;155:76–88. doi: 10.1016/j.antiviral.2018.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Danilov LL, Maltsev SD, Deyeva AV, et al. Phosprenyl: a novel drug with antiviral and immune modulating activity. Arch Immunol Ther Exp. 1996;44:395–400. [PubMed] [Google Scholar]
- Danilov LL, Deeva AV, Maltsev SD et al (2002) Remedy for the prevention and treatment of infectious diseases and the correction of pathological conditions of a living organism. RF Patent No. 2177788, 01/10/2002
- Einav S, Glenn JS. Prenylation inhibitors: a novel class of antiviral agents. J Antimicrob Chemother. 2003;52:883–886. doi: 10.1093/jac/dkg490. [DOI] [PubMed] [Google Scholar]
- Ershov FI, Narovlyansky AN. Usage of interferon inducers during viral infections. Vopr Virusol. 2015;60:5–10. [PubMed] [Google Scholar]
- Ershov FI, Narovlyansky AN, Pronin AV et al (2017) Genital Herpes in Russia: the Scope of the Problem and Treatment Prospects. In Berhardt LV (ed) Advances in Medicine and Biology, Nova Science Publishers, Inc., New York, vol 124, chapter 6, pp 121–174
- Gower TL, Graham BS. Antiviral activity of lovastatin against respiratory syncytial virus in vivo and in vitro. Antimicrob Agents Chemother. 2001;45:1231–1237. doi: 10.1128/AAC.45.4.1231-1237.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartley MD, Imperiali B. At the membrane frontier: a prospectus on the remarkable evolutionary conservation of polyprenols and polyprenyl-phosphates. Arch Biochem Biophys. 2012;517:83–97. doi: 10.1016/j.abb.2011.10.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ianevski A, Zusinaite E, Kuivanen S, et al. Novel activities of safe-in-human broad-spectrum antiviral agents. Antiviral Res. 2018;154:174–182. doi: 10.1016/j.antiviral.2018.04.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones MB, Rosenberg JN, Betenbaugh MJ, et al. Structure and synthesis of polyisoprenoids used in N-glycosylation across the three domains of life. Biochim Biophys Acta. 2009;1790:485–494. doi: 10.1016/j.bbagen.2009.03.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanda T, Nirei K, Matsumoto N, et al. Retreatment of patients with treatment failure of direct-acting antivirals: focus on hepatitis C virus genotype 1b. World J Gastroenterol. 2017;23:8120–8127. doi: 10.3748/wjg.v23.i46.8120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuritz T (2009) Methods and compositions for modulation of innate immunity, US Patent 0306017
- Legendre AM, Bartges JW. Effect of Polyprenyl immunostimulant on the survival times of three cats with the dry form of feline infectious peritonitis. J Feline Med Surg. 2009;11:624–626. doi: 10.1016/j.jfms.2008.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Legendre AM, Kuritz T, Galyon G, et al. Polyprenyl immunostimulant treatment of cats with presumptive non-effusive feline infectious peritonitis in a field study. Front Vet Sci. 2017;4:7. doi: 10.3389/fvets.2017.00007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Legendre AM, Kuritz T, Heidel RE, et al. Polyprenyl immunostimulant in feline rhinotracheitis: randomized placebo-controlled experimental and field safety studies. Front Vet Sci. 2017;4:24. doi: 10.3389/fvets.2017.00024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marakasova ES, Eisenhaber B, Maurer-Stroh S, et al. Prenylation of viral proteins by enzymes of the host: virus-driven rationale for therapy with statins and FT/GGT1 inhibitors. BioEssays. 2017;39(10):1700014. doi: 10.1002/bies.201700014. [DOI] [PubMed] [Google Scholar]
- Martínez-Gutierrez M, Castellanos JE, Gallego-Gómez JC. Statins reduce dengue virus production via decreased virion assembly. Intervirology. 2011;54:202–216. doi: 10.1159/000321892. [DOI] [PubMed] [Google Scholar]
- Morita CT, Jin C, Sarikonda G, et al. Nonpeptide antigens, presentation mechanisms, and immunological memory of human Vγ2Vδ2 T cells: discriminating friend from foe through the recognition of prenyl pyrophosphate antigens. Immunol Rev. 2007;215:59–76. doi: 10.1111/j.1600-065X.2006.00479.x. [DOI] [PubMed] [Google Scholar]
- Naesens L, Stevaert A, Vanderlinden E. Antiviral therapies on the horizon for influenza. Curr Opin Pharmacol. 2016;30:106–115. doi: 10.1016/j.coph.2016.08.003. [DOI] [PubMed] [Google Scholar]
- Narovlyansky AN, Deriabin PG, Sedov AM, et al. Antiviral activity of polyprenyl phosphates in an experimental infection caused by the hepatitis C virus in vitro. Zh J Microbiol Epidemiol Immunobiol. 2012;5:81–84. [PubMed] [Google Scholar]
- Narovlyansky AN, Ozherelkov SV, Sanin AV, et al. Antiviral activity and possible mechanisms of action of moraprenyl phosphates in experimental infection caused by the herpes simplex virus type 1. Zh J Microbiol Epidemiol Immunobiol. 2014;5:54–60. [PubMed] [Google Scholar]
- Narovlyansky AN, Sedov AM, Pronin AV, et al. Treatment of patients with chronic recurrent herpesvirus infection of genital localization: a clinical study of Fortepren preparation. Zh J Microbiol Epidemiol Immunobiol. 2015;4:112–118. [PubMed] [Google Scholar]
- Narovlyansky AN, Ivanova AM, Shevlyagina NV, et al. Efficiency of polyprenyl phosphates in experimental model of genital herpes. Vopr Virusol. 2015;60:9–13. [PubMed] [Google Scholar]
- Narovlyansky AN, Pronin AV, Sanin AV, et al. ID: 85: the cytokine-regulating activity of polyprenyl phosphates: clinical studies of ForteprenR. Cytokine. 2015;76:80–81. doi: 10.1016/j.cyto.2015.08.112. [DOI] [Google Scholar]
- Narovlyansky AN, Ivanova AM, Shevlyagina NV, et al. Effectiveness of the use of polyprenyl phosphates in the experimental model of genital herpes. Vopr Virusol. 2015;60:9–13. [PubMed] [Google Scholar]
- Narovlyansky AN, Pronin AV, Sanin AV, et al. Isoprenoids: polyprenols and plyprenyl phosphates as physiologically important metabolic regulators. New York: Nova Science Publishers Inc; 2018. p. 183. [Google Scholar]
- Ozherelkov SV, Timofeev AV, Novikova GP, et al. Protective effect of the new antiviral drug Phosprenyl with experimental tick-borne encephalitis. Vopr Virusol. 2000;45:33–37. [PubMed] [Google Scholar]
- Ozherelkov SV, Belousova RV, Danilov LL, et al. Phosprenyl inhibits the proliferation of cattle diarrhea virus and bovine infectious rhinotracheitis virus in sensitive cell cultures. Vopr Vir. 2001;5:43–45. [PubMed] [Google Scholar]
- Palsuledesai CC, Distefano MD. Protein prenylation: enzymes, therapeutics, and biotechnology applications. ACS Chem Biol. 2015;10:51–62 . doi: 10.1021/cb500791f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pronin AV, Ozerelkov SV, Narovlyansky AN, et al. Role of cytokines in immunomodulatory effects of polyprenylphosphate: new generation of antiviral drug. Russ J Immunol. 2000;5:156–164. [PubMed] [Google Scholar]
- Pronin AV, Grigorieva EA, Sanin AV, et al. Polyprenols as possible factors that determine an instructive role of the innate immunity in the acquired immune response. Russ J Immunol. 2002;7:135–142. [PubMed] [Google Scholar]
- Pronin AV, Danilov LL, Narovlyansky AN, et al. Plant polyisoprenoids and control of cholesterol level. Arch Immunol Ther Exp. 2014;62:31–39. doi: 10.1007/s00005-013-0253-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pronin AV, Ozherelkov SV, Deeva AV, et al. Polyprenyl phosphates as adjuvants polarizing the immune response towards Th1. Russian J Infect Immun. 2014;2:645. doi: 10.15789/2220-7619-2012-3-645-650. [DOI] [Google Scholar]
- Pronin AV, Narovlyansky AN, Shulzhenko AE, et al. New polyprenyl phosphate based preparation Fortepren as promising cytokine regulating antiviral remedy. Cytokine Growth Factor Rev. 2016;30:119–126. doi: 10.1016/j.cytogfr.2016.04.001. [DOI] [PubMed] [Google Scholar]
- Sanin AV, Ganshina IV, Sudyina GF, et al. Phosphorylated polyprenols are a new class of compounds with anti-inflammatory and bronchodilator activity. Russian J Infect Immun. 2011;4:355–360. [Google Scholar]
- Sanin AV, Narovlyansky AN, Pronin AV (2011b) Immunomodulators in agriculture: a tribute to fashion or necessity? Russian Veterinary J 1:37–40. https://cyberleninka.ru/article/n/immunomodulyatory-v-selskom-hozyaystve-dan-mode-ili-neobhodimost/viewer
- Sato R. Sterol metabolism and SREBP activation. Arch Biochem Biophys. 2010;501:177–181. doi: 10.1016/j.abb.2010.06.004. [DOI] [PubMed] [Google Scholar]
- Sedov AM, Narovlyansky AN, Pronin AV, et al. Mechanism of antiviral action and evaluation of the effectiveness of the new drug Fortepren® in the treatment of chronic recurrent herpes virus infection of genital localization. J Microbiol Epidemiol Immunobiol. 2018;5:81–87. doi: 10.36233/0372-9311-2018-5-81-87. [DOI] [Google Scholar]
- Sobolev SM, NikolaevaTN GEA, et al. The role of lectin-substrate recognition in the immunoregulatory interaction of interleukin-2 and IgG. Med Immunol. 2010;12:13–20 . doi: 10.15789/1563-0625-2010-1-2-13-20. [DOI] [Google Scholar]
- Vasil’evOzherelkovKozlov ANSVVV, et al. Antiviral and immunomodulating activity of polyprenyl phosphates in viral infections. Antibiot Khimioter. 2008;53:3–8. [PubMed] [Google Scholar]
- Welti M. Regulation of dolichol-linked glycosylation. Glycoconj J. 2013;30:51–56. doi: 10.1007/s10719-012-9417-y. [DOI] [PubMed] [Google Scholar]
- WHO (2007–2017) Viral hepatitis: a secret killer becomes apparent. https://www.who.int/publications/10-year-review/hepatitis/en/
- Zhang Q, Huang L, Zhang C, et al. Synthesis and biological activity of polyprenols. Fitotherapia. 2015;106:184–193. doi: 10.1016/j.fitote.2015.09.008. [DOI] [PubMed] [Google Scholar]