

Abstract
Background:
Accumulation of fatty acids in liver causes lipotoxicity which is followed by nonalcoholic fatty liver disease. The association between intakes of trans-fatty acids with metabolic diseases is still controversial. Accordingly, the objective of this study was to investigate the in vitro effects of trans-palmitoleic acid (tPA) and palmitic acid (PA) on lipid accumulation in hepatocytes, focusing on the gene expression of sirtuin 1 (SIRT1) as well as the transcriptional activity of peroxisome proliferator-activated receptor alpha (PPARα).
Materials and Methods:
In this experimental study, hepatocellular carcinoma (HepG2) cells were cultured and treated with various concentrations of tPA and PA (C16:0). The accumulation of triglyceride in the cells was measured by enzymatic method. Gene expression was evaluated by real-time polymerase chain reaction. The activity of PPARα was assessed by luciferase reporter assay after transfection of human embryonic kidney 293T cells by a vector containing the PPAR response element.
Results:
While concentration >1 mM for PA and cis-PA (cPA) reduced the viability of hepatocytes, tPA revealed an opposite effect and increased cell survival. Lipid accumulation in HepG2 cells after treatment with tPA was significantly lower than that in cells treated with PA. In addition, tPA at physiological concentration had no effect on the expression of SIRT1 while at high concentration significantly augmented its expression. There was a modest increase in PPARα activity at low concentration of tPA.
Conclusion:
tPA causes less lipid accumulation in hepatocytes with no detrimental effect on cell viability and might be beneficial for liver cells by the activation of SIRT1 and induction of PPARα activity.
Keywords: Gene expression, liver, palmitoleic acid, peroxisome proliferator-activated receptor alpha, Sirtuin 1
INTRODUCTION
Nonalcoholic fatty liver disease (NAFLD) is a disease characterized by the accumulation of triglycerides (TG) in liver, which results from an imbalance between the synthesis and oxidation of fatty acids. Lifestyle and genetic factors play an important role in the pathogenesis of this disease.[1] Dietary fatty acids play a major role in the early stages of NAFLD through impaired insulin signaling and fat accumulation in the liver.[2]
Trans fatty acids (TFAs) are unsaturated fatty acids, which contain at least one double bound in the form of trans configuration. TFAs are formed synthetically as a byproduct in the hydrogenation process of unsaturated fatty acids of vegetable oils and naturally by intestinal bacteria in ruminant animals.[3,4] Therefore, foods used with hydrogenated oils, meat, and milk of ruminants contain TFA.
Trans-palmitoleic acid (tPA),16:1t9, is one of the TFAs that is naturally found in dairy products and meat fat.[5] Although cis-fatty acids have beneficial effects for human health, there is a debate about the positive or negative effects of TFAs on cardiovascular and metabolic diseases.[6] Some epidemiological studies suggest that the excessive consumption of TFAs can lead to dyslipidemia, endothelial dysfunction, inflammation, and ultimately cardiovascular disease,[3] while some other studies have reported beneficial effects of naturally occurring TFAs such as tPA and conjugated linoleic acid (CLA) in metabolic diseases including diabetes, insulin resistance, and NAFLD.[7,8] No adverse effect on cholesterol and LDL has also been reported after consumption of dairy TFAs in a randomized clinical trial.[9] Promising health benefits have been reported for dietary tPA; and thus, dietary consumption of this fatty acid might be beneficial for well-being. However, the effects of this fatty acid on lipid accumulation in hepatocytes and its potential in amelioration of NAFLD have not yet been examined. In case of beneficial effects of tPA are established, it could be considered for dietary interventions for the management of NAFLD and similar disorders.
Peroxisome proliferator-activated receptors (PPARs), which have three isoforms including alpha, delta, and gamma, are considered the master regulators of lipid metabolism.[10] Among PPAR isoforms, PPARα regulates the transcription of genes associated with lipid homeostasis, decreases circulating TG, and increases high-density lipoprotein levels. PPARα is abundantly expressed in the liver and dietary fatty acids can activate this nuclear receptor.[11] Sirtuin 1 (SIRT1) is an efficient partner of PPARα and is required for the activation of PPARα coactivator.[12] SIRT1 is a nicotinamide adenine dinucleotide-dependent deacetylase that regulates lipid metabolism and is involved in fatty acid synthesis, oxidation, and adipocyte differentiation.[13] Hepatic SIRT1 regulates lipid homeostasis by positively regulating PPARα.[12] SIRT1 also exerts its protective effect against metabolic dysregulation in cardiomyocytes through PPARα.[14] The diminished SIRT1 expression plays an important role in the pathogenesis of hepatic steatosis, hepatic inflammation, metabolic syndrome, and type 2 diabetes.[15]
The objective of this study was to investigate in vitro effects of physiological and supraphysiological doses of tPA, on hepatic SIRT1 gene expression as one of the major regulators of lipid metabolism and compare the results with those of palmitic acid (PA). The effect of tPA on PPARα activity in human embryonic kidney (HEK-293) cells was also examined.
MATERIALS AND METHODS
Cell culture and fatty acids treatment
In this experimental study, human hepatocellular carcinoma (HepG2) cells and human embryonic kidney (HEK-293) cells were purchased from the Iranian Biological Resource Center (IBRC, Tehran, Iran). The cells were cultured in Dulbecco Minimal Eagle's medium (DMEM, Gibco) supplemented with 10% fetal bovine serum (FBS) (Gibco, Germany) and streptomycin-penicillin (100 μg/ml) in a humidified atmosphere of 5% CO2. The cells were plated in cell culture flasks at least 24 h before treatment. PA (C16:0), and tPA (trans-9-C16:1) as well as cPA (cis-9-C16:1) (Sigma, USA) were conjugated with 1% bovine serum albumin (BSA) (Sigma, USA). For this purpose, PA was first dissolved in 50% ethanol at 54°C. PA solution was then mixed with DMEM containing 1% BSA and incubated at 37°C for 2 h while shaking and subsequently added to cultured cells after filtration. Medium containing 1% BSA together with the corresponding amount of alcohol but lacking the fatty acid was used as the vehicle control. The 0.1–1.5 mM concentrations of tPA were used for gene expression and PPARα activity assay and for co-treatment experiments 0.1–1.5 mM concentrations of tPA together with 1 mM PA were applied to cultured cells. The concentrations of fatty acids used for cell viability assay are mentioned in the following section.
Cell viability assay
The effect of t-PA, cis-palmitoleic aid and PA on cell viability were determined using 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay. HepG2 cells (8 × 103/well) were seeded in 96-well plates and treated with 0, 0.25, 0.5, 0.75, 1, 1.5, 2, 2.5 mM of PA, cis-PA (cPA) or tPA or with a mixture of PA (1 mM) and tPA (0.1, 0.75, 1, and 1.5 mM) for 24 and 48 h. Then, MTT (Sigma, USA) solution was added to each well at the final concentration of 0.5 mg/ml. After 4 h, the fromazan crystals were dissolved in dimethyl sulfoxide and the absorbance was measured by plate reader at 570 nm.
Real-time polymerase chain reaction
To evaluate the effect of fatty acids on gene expression of SIRT1, HepG2 cells were treated with different concentrations of tPA or combination of tPA and PA for 24 h in 6-well plates. Then, cells were collected and the total RNA was extracted with GeneAll Ribospin™ total RNA purification kit (GeneAll Biotechnology, Seoul, South Korea). The synthesis of complementary DNA (cDNA) was carried out by RevertAidFirst Strand cDNA Synthesis Kit (Thermo Fisher, UK). The mRNA levels corresponding to SIRT1, PPARα, and glyceraldehyde phosphate dehydrogenase (GAPDH) (as the normalizer) were assessed by real-time polymerase chain reaction using SYBR Green Real Q Plus Master Mix (Ampliqon, Denmark) on Corbett Rotor Gene 6000 Light Cycler (Qiagen, Hilden, Germany). Specific primers were used as follows: SIRT1 forward: 5'-TGCGGGAATCCAAAGGATAA-3' and reverse: 5'-CAGGCAAGATGCTGTTGCA-3', GAPDH forward: 5'-GGGAAGGTGAAGGTCGGAGT-3' and reverse: 5'-TCCACTTTACCAGAGTTAAAAGCAGG-3', PPARα forward: 5'-CCGTTATCTGAAGAGTTCCTGC-3' and reverse: 5'-GTTGTGTGACATCCCGACAG-3'.
Serial dilutions of a pooled sample cDNA were used for standard curve generation. The data were normalized against GAPDH transcript level. The ΔΔCT method was used to calculate the relative gene expression.
Measurement of intracellular triglyceride
HepG2 cells were seeded at an initial density of 3 × 105 cells/well in a 6-well plate and treated with various concentrations of fatty acids for 48 h. In order to extract intracellular lipids, cells were homogenized in NP-40 detergent followed by heating at 100°C for 5 min and then cooling down to room temperature.[16] This procedure was performed twice to completely solubilize the intracellular lipids. The extracted TG was measured by enzymatic method using TG assay kit (Pars Azmoon, Iran) and normalized to the total protein concentration which was measured by bicinchoninic acid protein quantitative kit (Thermo Fisher Scientific, USA), using BSA as the standard.
The peroxisome proliferator-activated receptor alpha transcriptional activity assay
To examine the PPARα transcriptional activity, reporter assay was performed with PPARα-responsive firefly luciferase reporter and constitutively expressing Renilla construct (Cignal reporter assay kit, Qiagen, USA). A mixture of noninducible firefly luciferase reporter and constitutively expressing Renilla construct was used as the negative control. Constitutively expressing green fluorescent protein, firefly and Renilla luciferase construct was also used as the positive control.
HEK-293T cells were seeded at the density of 5 × 104 cells/well in a 24-well poly D-lysine-coated plates and were kept in serum-and antibiotic-free culture medium for 24 h. Subsequently, a mixture of polyethyleneimine (PEI) and the above vectors in Opti-MEM medium (Gibco, Germany) was added to the cells. After 4 h, the medium was replaced with the fresh one containing FBS (10%) and the cells were maintained for 48 h at normal culture conditions.
The activity of both firefly and Renilla luciferase enzymes was detected 48 h post-transfection by a dual luciferase assay kit following the instructions provided with the kit (Promega, USA). The activity of Renilla luciferase was recorded and used for the normalization of firefly luciferase activity.
Statistical analysis
All data were presented as mean ± standard deviation. The data were analyzed using one-way analysis of variance test using Dunnett's post-hoc test for comparing the differences between groups. Each experiment was performed as duplicate measurements, repeated independently at least three times and the mean values were analyzed. Values of P < 0.05 were considered statistically significant. Analyses of the resultant data were conducted by GraphPad Prism software (USA, San Diego).
RESULTS
Effect of fatty acids on the viability of hepatocellular carcinoma cells
The effects of tPA, cPA, and PA on the viability of HepG2 cells are shown in Figure 1. Results showed that not only tPA had no cytotoxic effect on the cells but also significantly increased cell viability at a concentration above 1.5 mM at 24 h (mean difference = −43.65, 95% confidence interval [CI]: −78.57–8.72) (P < 0.001) and 0.75 mM at 48 h (mean difference = −17.5, 95% CI: −51.51–16.51) (P < 0.001). On the contrary, PA significantly decreased cell viability from the 1.5 mM concentration at 24 h (mean difference = 36.42, 95% CI: 15.84–56.99) (P < 0.001). To evaluate whether the presence of double bond per se is responsible for the increased cell viability, cPA was also used for viability assessment and it was observed that cPA, similar to PA, decreased cell survival (mean difference = 25.97, 95% CI: 1.38–40.56) (P < 0.001).
Figure 1.
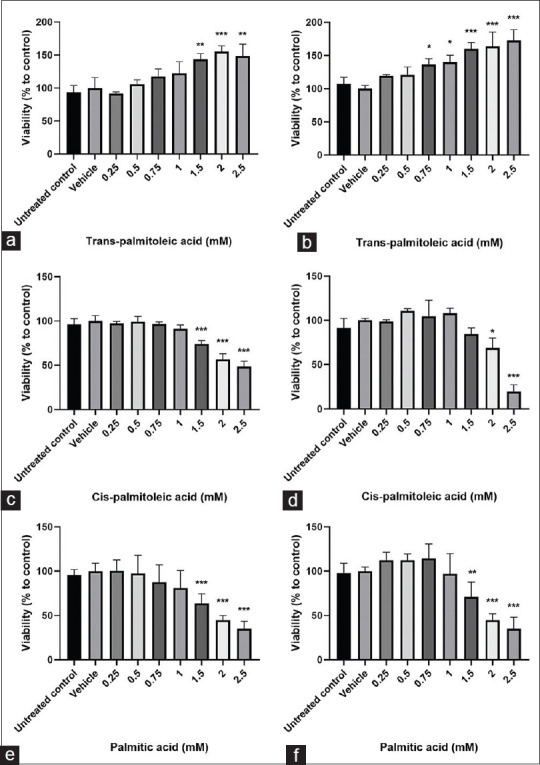
The effect of trans-palmitoleic acid (tPA), cis-palmitoleic acid and palmitic acid on cell viability in HepG2 cells. Cells were treated with different concentrations of trans-palmitoleic acid for 24 h (a) and 48 h (b), cis-palmitoleic acid for 24 h (c) and 48 h (d) and palmitic acid for 24 h (e) and 48 h (f). Results are expressed as mean ± standard deviation of three separate experiments, each group containing 6 replicates. *P < 0.05, **P < 0.01, ***P < 0.001 compared to untreated control
In the next step, the effect of co-treatment of tPA and PA was investigated. According to the results, concentrations of 0.75, 1, 1.5 mM for tPA were selected for subsequent experiments. To investigate the effects of tPA at the physiological concentration, which is more achievable and can be obtained by diet, a 0.1 mM concentration was also chosen based on previous studies.[17] The effects of co-treatment of PA (1 mM) with the aforementioned concentrations of tPA on cell viability were evaluated. As shown in Figure 2, when the cells were co-treated with tPA and PA, the cells were protected against the cytotoxic effects of PA and cell viability was significantly augmented at concentrations of 1 and 1.5 mM of tPA (mean difference = −39.28, 95% CI: −58.71 to −19.85 and mean difference = –50.59, 95% CI: –70.02 to –31.15, respectively) (P < 0.001).
Figure 2.
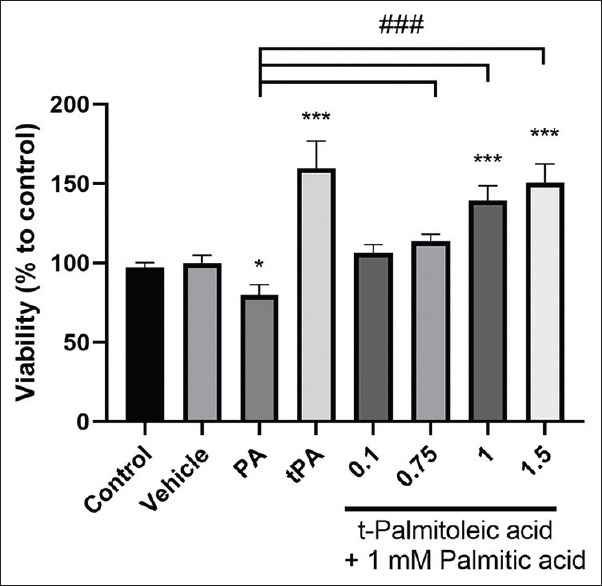
The effect of co-treatment of palmitic acid (PA) and trans-palmitoleic acid (tPA) on cell viability in HepG2 cells. Cells were treated with PA (1 mM), tPA (1.5 mM) or co-treated with both PA (1 mM) and tPA (0.1, 0.75, 1 and 1.5 mM) for 24 h. Results are expressed as mean ± standard deviation. Treated cells were compared with vehicle as negative control; *P < 0.05, **P < 0.01, ***P < 0.001. ###P < 0.001 compared to palmitic acid group
The effect of trans-palmitoleic acid on sirtuin 1 gene expression in HepG2 cells
The effect of tPA on SIRT1 gene expression is shown in Figure 3. Various concentrations of tPA were examined on SIRT1 gene expression and the results showed that tPA at the concentration of 1.5 mM significantly increased the mRNA expression of SIRT1 (mean difference = –0.57, 95% CI: –1.13 to –0.006) (P < 0.05). However, lower concentrations of tPA had no effect on the expression of SIRT1 gene. PA was not able to induce the expression of SIRT1. Nevertheless, the induction of SIRT1 gene expression was also observed in the cells co-treated with both tPA and PA (mean difference = –0.82, 95% CI: –1.39 to –0.24) (P < 0.05) [Figure 3].
Figure 3.
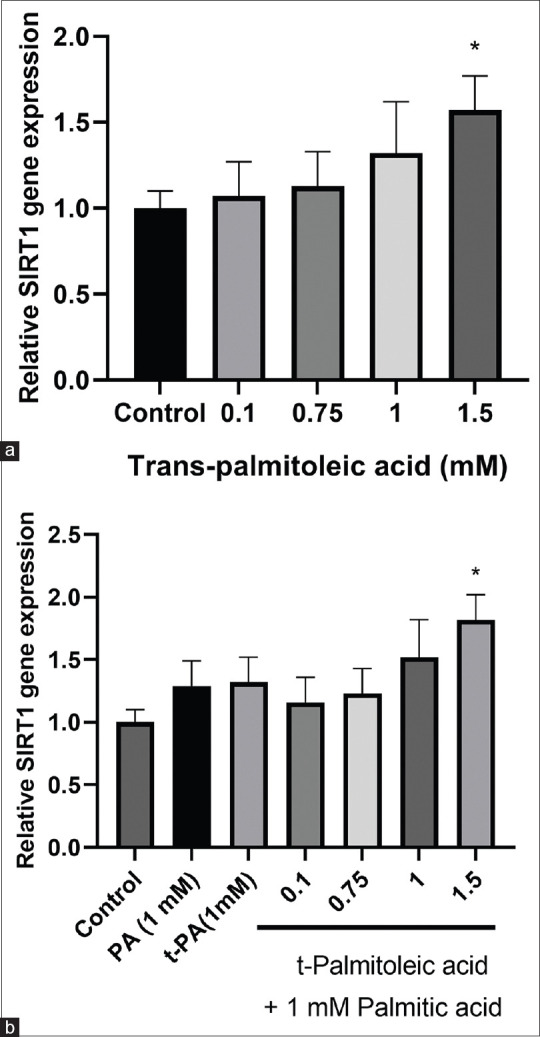
(a) The effect of various concentrations of trans-palmitoleic acid (tPA) on sirtuin 1 gene expression in HepG2 cells. (b) The effect of tPA either alone or together with 1 mM concentration of palmitic acid (PA) on sirtuin 1 gene expression in hepatocellular carcinoma cells. The results are presented as mean ± standard deviation *P < 0.05
The effect of trans-palmitoleic acid on triglycerides level in HepG2 cells
The intracellular TG level was measured after HepG2 cells were treated with PA, tPA or the combination of PA and tPA for 24 h [Figure 4]. As expected, both PA and tPA significantly increased lipid accumulation in HepG2 cells; however, the effect of tPA on TG accumulation was significantly lower than that of PA at similar concentrations. When combined with PA, tPA effectively ameliorated the steatotic effect of PA, especially at lower concentrations (P < 0.001, compared to PA alone) (mean difference = 73.17, 95% CI: 61.65–84.7).
Figure 4.
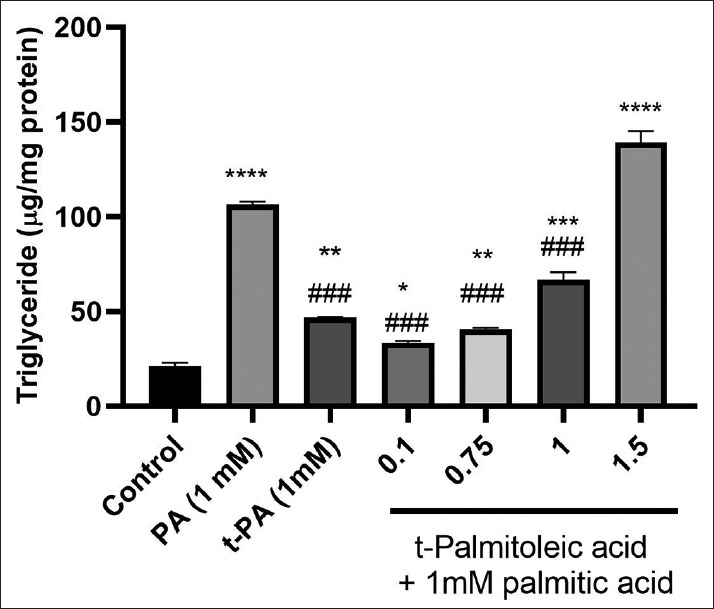
The effects of palmitic acid (PA) and trans-palmitoleic acid (tPA) separately or combined together on intracellular triglyceride levels in HepG2 cells. The data are mean ± standard deviation of the results. *Significantly different compared to control group; *P < 0.05, **P < 0.01, ***P < 0.001. #Significantly different from palmitic acid; ###P < 0.001
Effect of trans-palmitoleic acid on PPARα transcriptional activity in HEK293T cells
Since fatty acids are known as ligands and activators of PPARα, the transcriptional activity of PPARα was examined in response to the studied fatty acids. First, the PPARα expression was assessed and it was found that the expression of PPARα was low in HepG2 cells. Therefore, the expression of PPARα was measured in HEK293T cells and because of high expression of PPARα in these cells; they were used for the evaluation of PPARα transcriptional activity in response to tPA. As shown in Figure 5, the activity of luciferase which was indicative of transcriptional activity of PPARα, was significantly increased by the physiological concentration of tPA (0.1 mM) (mean difference = –0.87, 95% CI: –1.61 to –0.12) (P < 0.05); however, at higher concentrations no significant effect was detected. One millimolar concentration of PA was unable to induce any significant change in the activity of luciferase. Although co-treatment with both PA and low concentration of tPA caused a modest increase in the transcriptional activity of PPARα, this effect was not statistically significant.
Figure 5.
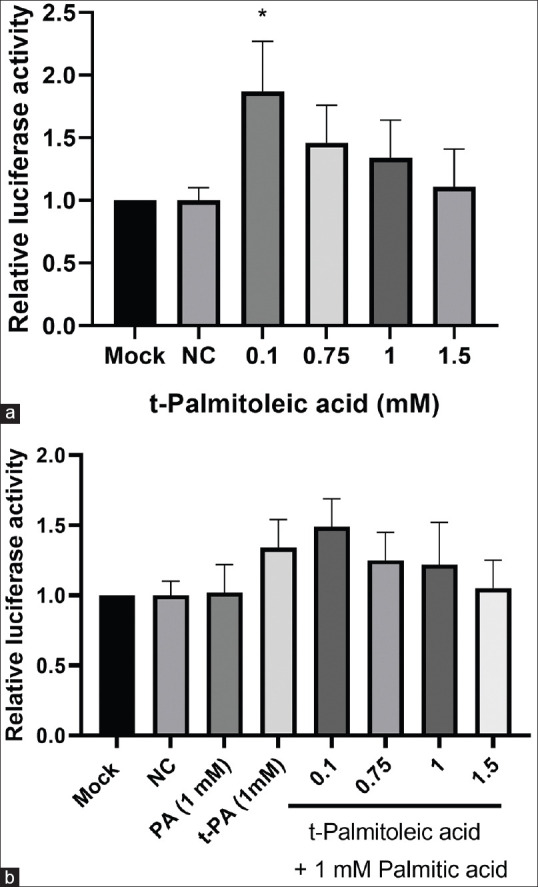
(a) The effect of different concentrations of trans-palmitoleic acid (tPA) on transcriptional activity of peroxisome proliferator-activated receptors alpha in HEK-293T cells, measured after transfection of cells with the vector containing the PPARα response element. Firefly luciferase activity was normalized to that of Renilla and the results were presented relative to the mock control. (b) The effect of different concentrations of tPA, combined with 1 mM concentration of palmitic acid (PA) on the transcriptional activity of PPARα. The data are presented as mean ± standard deviation *P < 0.05 compared to mock control. NC: Negative control
DISCUSSION
Numerous epidemiological and experimental studies have investigated the relationship of TFA consumption with cardiovascular and liver diseases. In this study, we first examined the effect of tPA compared with PA on HepG2 cell viability. The results showed that tPA, unlike PA, not only did not decrease the viability of HepG2 cells but also remarkably increased their survival. Cis-palmitoleic acid contains a double bond similar to tPA, but acted alike PA and caused a decline in cell survival, pointing out that the trans configuration of the double bond might be responsible for the beneficial effect of tPA. Interestingly, co-treatment of tPA with PA reduced the adverse effects of PA on cell viability.
This is the first study to reveal the protective effect of tPA on hepatocytes and against PA-induced cell toxicity; however, discrepancy in the effects of various fatty acids such as oleic acid on cell survival has been previously reported. Zeng et al. showed that oleate, unlike PA, did not deteriorate the survival of HepG2 cells and the combination of oleate and PA efficiently restored the cell viability to its normal level.[18] Nemecz et al. also investigated the effect of oleate and PA on pancreatic β-cells and showed that, in contrast to PA, oleate exerted a protective effect on β-cells through amelioration of oxidative and endoplasmic reticulum stress and decline of inflammatory cytokines.[19] Oleic acid has also been shown to be capable of increasing proliferation of mammary epithelial cells through the modulation of intracellular signaling pathways such as PI3K/Akt pathway.[20] Another naturally occurring TFA, conjugated linoleic acid (CLA), has also been reported to exert beneficial effects on liver cells. CLA have proved to be a potent inducer for fibroblast growth factor 21 (FGF21) production[21] and on the other hand, FGF21 is a known enhancer of hepatocyte regeneration and proliferation[22] which may mediate the protective effects of CLA on hepatocytes.
SIRT1 has long been considered a longevity factor that promotes cell survival and its inhibition leads to reduced hepatic cell viability and induction of apoptosis.[23,24,25] Reduced SIRT1 expression has been previously reported in obese subjects.[26]
In this study, we assessed the expression of SIRT1 in response to tPA and PA and found that tPA could significantly induce the gene expression of SIRT1. This effect was in line with the enhancement of cell viability observed for tPA and indicates that the up-regulation of SIRT1 might be at least in part responsible for the increased cell viability by tPA. According to our review of literature, this is the first study to report the inducing effect of tPA on SIRT1 expression, Nevertheless, some other natural cis or TFAs have been shown as effective positive regulators of SIRT1. CLA is among these fatty acids and can potently activate SIRT1 in myotubes and adipocytes.[27,28] Lim et al. also showed stimulatory effect of oleic acid on SIRT1 activity. Consistent with our findings, they did not observe any positive effect on SIRT1 for PA.[29] Protective effect of omega-3 fatty acids on hepatocytes has also been attributed to upregulation of SIRT1 and modulation of its downstream targets by these fatty acids.[30]
Another finding of the current study was the differential TG accumulation in hepatocytes in response to tPA and PA. Although both fatty acids augmented lipid deposition in HepG2 cells, the effect was comparatively lower for tPA, especially at physiological concentrations which only marginally increased intracellular TG levels compared to control cells. Consistently, Mozaffarian et al. showed that increased circulating tPA is associated with lower TG, fasting insulin, blood pressure, and incident of diabetes in human subjects.[8]
Fatty acid regulation of hepatic gene transcription is preformed through the modulation of several transcription factors that have been identified as targets of various fatty acid. PPARα activation increases lipid oxidation and decreases TG accumulation.[31,32] Here, we investigated the PPARα activity in response to fatty acids. The limitation of our study was the low expression of PPARα in HepG2 cells and therefore we assessed the activity of this nuclear receptor in a different cell line with high transfection efficiency. We showed that physiological concentration of tPA significantly induced PPARα transcriptional activity assessed by luciferase reporter assay. Thus, low TG accumulation in response to the physiological concentration of tPA might be attributed to the corresponding induction of PPARα activity.
Fatty acids are among the ligands of PPARα; however, previous studies have shown that different fatty acids act contrarily toward PPARs. PA and stearic acid inhibited while palmitoleic acid and oleic acid that are their monounsaturated counterparts, enhanced PPARα transactivation. Even among the unsaturated fatty acids, linoleic and γ-linolenic acid have proved to be ineffective in the activation of PPARα; whereas, α-linolenic acid strongly amplified transactivation. Low doses of arachidonic, eicosapentaenoic, and docosahexaenoic acid activated while their high dose suppressed the activity of PPARα.[33] Cruz et al. reported that palmitoleic acid was able to increase fatty acid β-oxidation in adipocytes.[34] Based on our findings, the previously reported stimulation of β-oxidation might be attributed to its positive effect on PPARα transcriptional activity.
Investigating the effect of tPA on lipid accumulation in liver and the metabolic pathways related to lipid metabolism in animal studies or randomized clinical trials might shed more light on the value of tPA and is therefore suggested for future studies to evaluate the effectiveness of this fatty acid for dietary interventions.
CONCLUSION
In summary, we have demonstrated that tPA, in contrast to PA, increased hepatic cell viability and up-regulated SIRT1 at high concentrations while enhanced PPARα at low concentrations. Thus, both low and high concentrations of tPA are beneficial and in contrast to PA, are not devastating for hepatocytes. Further human and animal studies might provide evidence for the possibility and benefit of long-term consumption of this fatty acid in health and disease.
Financial support and sponsorship
This study was approved and financially supported by grant number 943776 from the National Institute for Medical Research Development, Tehran, Iran, and also registered and approved in Science and Research Branch, Islamic Azad University, Tehran, Iran.
Conflicts of interest
There are no conflicts of interest.
REFERENCES
- 1.Solga S, Alkhuraishe AR, Clark JM, Torbenson M, Greenwald A, Diehl AM, et al. Dietary composition and nonalcoholic fatty liver disease. Dig Dis Sci. 2004;49:1578–83. doi: 10.1023/b:ddas.0000043367.69470.b7. [DOI] [PubMed] [Google Scholar]
- 2.Tamer F, Ulug E, Akyol A, Nergiz-Unal R. The potential efficacy of dietary fatty acids and fructose induced inflammation and oxidative stress on the insulin signaling and fat accumulation in mice. Food Chem Toxicol. 2020;135:110914. doi: 10.1016/j.fct.2019.110914. [DOI] [PubMed] [Google Scholar]
- 3.Hunter JE. Dietary trans fatty acids: Review of recent human studies and food industry responses. Lipids. 2006;41:967–92. doi: 10.1007/s11745-006-5049-y. [DOI] [PubMed] [Google Scholar]
- 4.McKain N, Shingfield KJ, Wallace RJ. Metabolism of conjugated linoleic acids and 18: 1 fatty acids by ruminal bacteria: Products and mechanisms. Microbiology. 2010;156:579–88. doi: 10.1099/mic.0.036442-0. [DOI] [PubMed] [Google Scholar]
- 5.Destaillats F, Wolff RL, Precht D, Molkentin J. Study of individual trans-and cis-1:1 isomers incow, goat, and ewe cheese fats by gas-liquid chromatography with emphasis on the trans-Δ3 isomer. Lipids. 2000;35:1027–32. doi: 10.1007/s11745-000-0614-y. [DOI] [PubMed] [Google Scholar]
- 6.Brouwer I, Wanders A, Katan M. Trans fatty acids and cardiovascular health: Research completed? Eur J Clin Nutr. 2013;67:541. doi: 10.1038/ejcn.2013.43. [DOI] [PubMed] [Google Scholar]
- 7.Nagao K, Inoue N, Wang YM, Shirouchi B, Yanagita T. Dietary conjugated linoleic acid alleviates nonalcoholic fatty liver disease in Zucker (fa/fa) rats. J Nutr. 2005;135:9–13. doi: 10.1093/jn/135.1.9. [DOI] [PubMed] [Google Scholar]
- 8.Mozaffarian D, de Oliveira Otto MC, Lemaitre RN, Fretts AM, Hotamisligil G, Tsai MY, et al. trans-Palmitoleic acid, other dairy fat biomarkers, and incident diabetes: The Multi-Ethnic Study of Atherosclerosis (MESA) Am J Clin Nutr. 2013;97:854–61. doi: 10.3945/ajcn.112.045468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lacroix E, Charest A, Cyr A, Baril-Gravel L, Lebeuf Y, Paquin P, et al. Randomized controlled study of the effect of a butter naturally enriched in trans fatty acids on blood lipids in healthy women. Am J Clin Nutr. 2011;95:318–25. doi: 10.3945/ajcn.111.023408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chinetti G, Fruchart JC, Staels B. Peroxisome proliferator-activated receptors and inflammation: From basic science to clinical applications. Int J Obes Relat Metab Disord. 2003;27(Suppl 3):S41–5. doi: 10.1038/sj.ijo.0802499. [DOI] [PubMed] [Google Scholar]
- 11.Berger JP, Akiyama TE, Meinke PT. PPARs: Therapeutic targets for metabolic disease. Trends Pharmacol Sci. 2005;26:244–51. doi: 10.1016/j.tips.2005.03.003. [DOI] [PubMed] [Google Scholar]
- 12.Purushotham A, Schug TT, Xu Q, Surapureddi S, Guo X, Li X. Hepatocyte-specific deletion of SIRT1 alters fatty acid metabolism and results in hepatic steatosis and inflammation. Cell Metab. 2009;9:327–38. doi: 10.1016/j.cmet.2009.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Guarente L. Sirtuins as potential targets for metabolic syndrome. Nature. 2006;444:868–74. doi: 10.1038/nature05486. [DOI] [PubMed] [Google Scholar]
- 14.Planavila A, Iglesias R, Giralt M, Villarroya F. Sirt1 acts in association with PPARα to protect the heart from hypertrophy, metabolic dysregulation, and inflammation. Cardiovasc Res. 2011;90:276–84. doi: 10.1093/cvr/cvq376. [DOI] [PubMed] [Google Scholar]
- 15.de Kreutzenberg SV, Ceolotto G, Papparella I, Bortoluzzi A, Semplicini A, Dalla Man C, et al. Downregulation of the longevity-associated protein sirtuin 1 in insulin resistance and metabolic syndrome: Potential biochemical mechanisms. Diabetes. 2010;59:1006–15. doi: 10.2337/db09-1187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lanaspa MA, Sanchez-Lozada LG, Cicerchi C, Li N, Roncal-Jimenez CA, Ishimoto T, et al. Uric acid stimulates fructokinase and accelerates fructose metabolism in the development of fatty liver. PLoS One. 2012;7:e47948. doi: 10.1371/journal.pone.0047948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Da Silva MS, Julien P, Bilodeau J-F, Barbier O, Rudkowska I. Trans fatty acids suppress TNF-α-induced inflammatory gene expression in endothelial (HUVEC) and hepatocellular carcinoma (HepG2) cells. Lipids. 2017;52:315–25. doi: 10.1007/s11745-017-4243-4. [DOI] [PubMed] [Google Scholar]
- 18.Zeng X, Liu X, Luo R, Chen Y. Oleic acid ameliorates palmitic acid induced pyroptosis by attenuating endoplasmic reticulum stress in HepG2 cells. FASEB J. 2019;33(1 Suppl):48727. [Google Scholar]
- 19.Nemecz M, Constantin A, Dumitrescu M, Alexandru N, Filippi A, Tanko G, et al. The distinct effects of palmitic and oleic acid on pancreatic beta cell function: The elucidation of associated mechanisms and effector molecules. Front Pharmacol. 2018;9:1554. doi: 10.3389/fphar.2018.01554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Meng Y, Zhang J, Yuan C, Zhang F, Fu Q, Su H, et al. Oleic acid stimulates HC11 mammary epithelial cells proliferation and mammary gland development in peripubertal mice through activation of CD36-Ca2+and PI3K/Akt signaling pathway. Oncotarget. 2018;9:12982–94. doi: 10.18632/oncotarget.24204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yu J, Yu B, Jiang H, Chen D. Conjugated linoleic acid induces hepatic expression of fibroblast growth factor 21 through PPAR-α. Br J Nutr. 2012;107:461–5. doi: 10.1017/S0007114511003205. [DOI] [PubMed] [Google Scholar]
- 22.Schumacher JD, Guo GL. Regulation of hepatic stellate cells and fibrogenesis by fibroblast growth factors. BioMed Res Int. 2016;2016:8323747. doi: 10.1155/2016/8323747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cohen HY, Miller C, Bitterman KJ, Wall NR, Hekking B, Kessler B, et al. Calorie restriction promotes mammalian cell survival by inducing the SIRT1 deacetylase. Science. 2004;305:390–2. doi: 10.1126/science.1099196. [DOI] [PubMed] [Google Scholar]
- 24.Borji M, Nourbakhsh M, Shafiee SM, Owji AA, Abdolvahabi Z, Hesari Z, et al. Down-Regulation of SIRT1 Expression by mir-23b Contributes to Lipid Accumulation in HepG2 Cells. Biochem Genet. 2019;57:507–21. doi: 10.1007/s10528-019-09905-5. [DOI] [PubMed] [Google Scholar]
- 25.Yousefi Z, Nourbakhsh M, Abdolvahabi Z, Ghorbanhosseini SS, Hesari Z, Yarahmadi S, et al. microRNA-141 is associated with hepatic steatosis by downregulating the sirtuin1/AMP-activated protein kinase pathway in hepatocytes. J Cell Physiol. 2020;235:880–90. doi: 10.1002/jcp.29002. [DOI] [PubMed] [Google Scholar]
- 26.Arab Sadeghabadi Z, Nourbakhsh M, Pasalar P, Emamgholipour S, Golestani A, Larijani B, et al. Reduced gene expression of sirtuins and active AMPK levels in children and adolescents with obesity and insulin resistance. Obes Res Clin Pract. 2018;12:167–73. doi: 10.1016/j.orcp.2017.10.004. [DOI] [PubMed] [Google Scholar]
- 27.Kim YO, Park Y. Conjugated linoleic acid (CLA) activates PGC-1alpha via AMPK and SIRT1 in C2C12 myotubes. FASEB J. 2013;27(1 Suppl):63725. [Google Scholar]
- 28.Jiang S, Wang W, Miner J, Fromm M. Cross regulation of sirtuin 1, AMPK, and PPARγ in conjugated linoleic acid treated adipocytes. PLoS One. 2012;7:e48874. doi: 10.1371/journal.pone.0048874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lim JH, Gerhart-Hines Z, Dominy JE, Lee Y, Kim S, Tabata M, et al. Oleic acid stimulates complete oxidation of fatty acids through protein kinase A-dependent activation of SIRT1-PGC1α complex. J Biol Chem. 2013;288:7117–26. doi: 10.1074/jbc.M112.415729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wang T, Yang B, Ji R, Xu W, Mai K, Ai Q. Omega-3 polyunsaturated fatty acids alleviate hepatic steatosis-induced inflammation through Sirt1-mediated nuclear translocation of NF-κB p65 subunit in hepatocytes of large yellow croaker (Larmichthys crocea) Fish Shellfish Immunol. 2017;71:76–82. doi: 10.1016/j.fsi.2017.09.064. [DOI] [PubMed] [Google Scholar]
- 31.Djouadi F, Aubey F, Schlemmer D, Bastin J. Peroxisome proliferator activated receptor δ (PPARδ) agonist but not PPARα corrects carnitine palmitoyl transferase 2 deficiency in human muscle cells. J Clin Endocrinol Metab. 2005;90:1791–97. doi: 10.1210/jc.2004-1936. [DOI] [PubMed] [Google Scholar]
- 32.Muoio DM, Way JM, Tanner CJ, Winegar DA, Kliewer SA, Houmard JA, et al. Peroxisome proliferator-activated receptor-α regulates fatty acid utilization in primary human skeletal muscle cells. Diabetes. 2002;51:901–9. doi: 10.2337/diabetes.51.4.901. [DOI] [PubMed] [Google Scholar]
- 33.Popeijus H, van Otterdijk S, Krieken S, Konings M, Serbonij K, Plat J, et al. Fatty acid chain length and saturation influences PPAR transcriptional activation and repression in HepG2 cells. Molecular Nutr Food Res. 2014;58:2342–9. doi: 10.1002/mnfr.201400314. [DOI] [PubMed] [Google Scholar]
- 34.Cruz MM, Lopes AB, Crisma AR, de Sá RCC, Kuwabara WMT, Curi R, et al. Palmitoleic acid (16:1n7) increases oxygen consumption, fatty acid oxidation and ATP content in white adipocytes. Lipids in health and disease. 2018;17:55. doi: 10.1186/s12944-018-0710-z. [DOI] [PMC free article] [PubMed] [Google Scholar]