

Abstract
COVID-19 (Coronavirus Disease 2019) is a highly contagious infection and associated with high mortality rates, primarily in elderly; patients with heart failure; high blood pressure; diabetes mellitus; and those who are smokers. These conditions are associated to increase in the level of the pulmonary epithelium expression of angiotensin-converting enzyme 2 (ACE-2), which is a recognized receptor of the S protein of the causative agent SARS-CoV-2 (Severe Acute Respiratory Syndrome Coronavirus 2). Severe cases are manifested by parenchymal lung involvement with a significant inflammatory response and the development of microvascular thrombosis. Several factors have been involved in developing this prothrombotic state, including the inflammatory reaction itself with the participation of proinflammatory cytokines, endothelial dysfunction/endotheliitis, the presence of antiphospholipid antibodies, and possibly the tissue factor (TF) overexpression. ARS-Cov-19 ACE-2 down-regulation has been associated with an increase in angiotensin 2 (AT2). The action of proinflammatory cytokines, the increase in AT2 and the presence of antiphospholipid antibodies are known factors for TF activation and overexpression. It is very likely that the overexpression of TF in COVID-19 may be related to the pathogenesis of the disease, hence the importance of knowing the aspects related to this protein and the therapeutic strategies that can be derived. Different therapeutic strategies are being built to curb the expression of TF as a therapeutic target for various prothrombotic events; therefore, analyzing this treatment strategy for COVID-19-associated coagulopathy is rational. Medications such as celecoxib, cyclosporine or colchicine can impact on COVID-19, in addition to its anti-inflammatory effect, through inhibition of TF.
Keywords: COVID-19, SARS-CoV-2, coagulopathy, Covid-2-associated coagulopathy, thrombosis, tissue factor
Introduction
In December 2019, there was a new highly contagious infectious disease outbreak in Wuhan, China. Some infected patients developed severe acute respiratory syndrome (SARS) and a systemic inflammatory response syndrome (SIRS) associated with a high mortality rate. This disease then rapidly spread throughout the world and a pandemic was declared in March 2020.1 The causative agent of the novel disease known as COVID-19 (Coronavirus Disease 2019) was isolated and identified as a new coronavirus, known as SARS-CoV-2 (Severe Acute Respiratory Syndrome Coronavirus 2).2 Studies report that S protein from the virion surface binds to the angiotensin-converting enzyme 2 (ACE-2) expressed in respiratory epithelial cells, thus triggering mechanisms that cause the virus enter the cell where it manages to generate its replicants.3 At least 2 types of biological response have been identified and are related to the pathogenesis of the disease: the induction of an inflammatory response and the generation of a procoagulant state.
The immune/inflammatory response can be mild and rapidly self-limiting. Where the immune/inflammatory response is mild and rapidly self-limiting, patients will present as asymptomatic or with only mild symptoms. Patients with less severe disease symptoms achieve, as far as is known, both cellular and humoral immunity for protection against future virus exposures.4 This immune response may be poorly regulated in other infected individuals, progressing to SARS and SIRS, both of which are associated with high mortality rates.5 Moreover, many of these patients develop thrombotic coagulopathy,6 which is clinically manifested as cerebrovascular accidents,7 acroischemia,8 or pulmonary thromboembolism.9 The presence of thrombi in pulmonary microcirculation is the most common result when performing an autopsy (27%).10 The most common abnormalities in hemostatic screening tests are prolonged prothrombin times (PT) and partial thromboplastin times (PTT), elevated D-dimer serum levels, and reduced fibrinogen and platelet count levels.11 Thrombin, tissue plasminogen activator (tPA), tissue factor pathway inhibitor (TFPI) and vascular endothelial growth factor (VEGF) significantly more increased in the critical versus noncritical patients.12
COVID-19 severity has been associated with advanced age and comorbidities such as chronic obstructive pulmonary disease (COPD), heart failure, hypertension (HTN), diabetes mellitus, and smoking.13 These comorbidities have been associated with pulmonary ACE-2 overexpression; moreover, this group of viruses causes ACE-2 down-regulation, resulting in increased angiotensin 2 (AT2).14 This acute increase in levels of AT2 may have direct implications for the immune, vascular endothelial and coagulation responses.15,16
Accumulation of AT-II secondary to ACE2 downregulation may promote clot formation via interactions with endothelial cells and platelets17; TF may be involved in this process.18 These findings may also be associated with additional prothrombotic events that have been identified in patients diagnosed with COVID-19, including increases in serum IL-6 which has characterized activities with respect to coagulation factors,19 and platelets.20 IL-6 can activate directly TF.21 T lymphocytes as well as the cytokines IL-6, and TNF-α may have direct roles in promoting microvascular damage associated with AT-II.22–24
Neutrophil extracellular traps (NETs) are extracellular webs of chromatin, microbicidal proteins, and oxidant enzymes that are released by neutrophils to contain infections, and have potential to propagate inflammation and microvascular thrombosis. Sera from individuals with COVID-19 triggered NET release from control neutrophils in vitro.25 Interestingly IL-1β mediates arterial thrombus formation via NET-associated TF.26
It’s important to put attention on generation of proinflammatory cytokines and vascular inflammation are mediated through a number of pattern recognition receptors, known as toll-like receptors (TLRs) and nod-like receptors (NLRs).27
Activation of TLRs through oxidized phospholipids or damage associated molecular pattern (DAMP) can activate the production of TNFα, IL6, and other cytokines that are responsible for cytokine storm in COVID-19.28 Interestingly, one in silico interaction study hypothesized that TLR4 may be involved in recognizing molecular pattern from spike protein of coronavirus. Further studies will be required to understand the roles of TLRs in COVID-19.29
Both an acute inflammatory response30 and increased AT231 are known causes for TF overexpression and the activation of the blood clotting cascade, an event that has been associated with COVID-19 pathogenesis. TF upregulation may possibly participate in the pathogenesis of the disease32-34 as part of a chain of events where endothelial dysfunction/endotheliitis are involved,35 Willebrand factor and soluble thrombomodulin overexpression36 and ADAMTS13 deficiency (a disintegrin and metalloproteinase with a thrombospondin type 1 motif, member 13).37,38 Unmasking these pathways in COVID-19 would have a key role in designing therapeutic strategies. On the other hand, the increase in AT2 can cause vasoconstriction with increased blood pressure, which can lead to hemorrhagic stroke.39 In some patients with COVID-19, significant deposits of terminal complement components C5b-9 (membrane attack complex), C4d, and mannose binding lectin (MBL)-associated serine protease 2 (MASP-2) have been found in the microvasculature of different organs.40
This accumulated observation of the activation of the complement system in COVID-19, it is quite likely to contribute in developing thrombotic processes,41–43 as previously assessed from the physiological interplay of complement and blood clotting cascades.44 The role of TF activation induced by activated complement factors such as C5a through the effect of protein disulfide isomerase (PDI)45 should be examined in COVID-19, which could be the basis for developing therapeutic proposals.
Protein C, a natural anti-coagulant is reduced in patient with COVID-19.46 Activated protein C is a prominent anti-coagulant, anti-inflammatory molecule that regulate endothelial dysfunction and TF expression in monocyte could be useful therapeutically.47
Figure 1 summary as immunological dysfunction, ACE2 downregulation, and AT2 and TF upregulation would be involved in the thrombosis pathogenesis in COVID-19.
Figure 1.
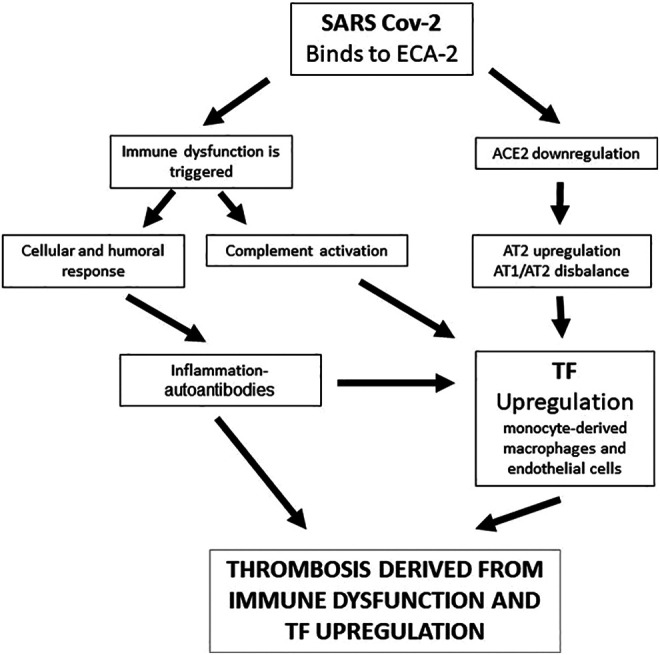
Summary of the possible mechanisms associated with thrombosis in COVID-19 where immunological dysfunction, angiotensin-converting enzyme 2 (ACE2) downregulation, and angiotensin 2 (AT2) and tissue factor (TF) upregulation would be involved.
Tissue Factor (TF)
TF was discovered in 1904 as a protein that converted prothrombin to thrombin at the plasma level.48 Being a transmembrane protein with a high affinity for Factor VII and Factor VIIa, it is known as thromboplastin, coagulation factor III or CD 142; moreover, it acts as an initiator of the blood clotting cascade by activating Factors IX49 and X.50 It first appeared in animals 450 million years ago in the evolutionary path of agnate fish.51 The gene is found on chromosome 1 and has 6 exons.52
It occurs dimerically, with 2 extracellular domains belonging to the immunoglobulin superfamily, the intracellular part is bound to intracellular Jak-STAT signaling structures, being classified within the group of type II cytokine receptors.53,54 It emits intracellular responses that lead to compartment-specific changes in calcium,55 with biological cell migration and proliferation effects,56 as well as angiogenic effects at embryonic stages.57
TF contains an allosteric disulfide bond that stabilizes the carboxyl-terminal domain involved in ligand interactions with coagulation factors VIIa and X. PDI is involved in modifying allosteric disulfide by reduction to achieve the activation thereof (change from an encrypted to a decrypted conformation of TF).58,59
TF is mainly expressed in endothelial cells,60 platelets,61 polymorphonuclear cells,62 T lymphocytes,63 and fibroblasts.64 It is located mainly in lipid rafts65 and closely linked to β1 integrins.66
The main physiologic trigger for TF activation (transition from an inactive encrypted to a decrypted conformation) is tissue injury that results in hemostasis. This activation primarily occurs in the endothelium, platelets, and perivascular cells.67 Under pathological conditions, it is released into the blood as vesicles or microparticles. The release of microparticles has been observed in conditions where there are thrombotic events, from fractured atheromatous plaque (at the level of Foam cells), monocytes in septic states, or neoplastic cells.68–70
Different TF gene agonists in endothelial cells have been studied, including tumor necrosis factor alpha (TNFα),71 interleukin-1β (IL-1β),72 CD40 L,73 vascular endothelial growth factor (VEGF),74 serotonin,75 histamine,76 and C-reactive protein.77 Stimulated P-selectin increases monocyte TF expression.78 The induction of intracellular signaling is essential in the TF activation process.79–81
Anti-cardiolipin antibodies and anti-β2-glycoprotein 1 increase TF expression in monocytes in antiphospholipid syndrome.82,83 Vitamin D can modulate TF expression in these patients.84
TF Expression Regulation
The TF activation process is regulated at both transcriptional and post-transcriptional levels.85 The positions of various membrane phospholipids that generate different polarities,86 the PDI effect on disulfide bonds,87 and the TFPI (tissue factor pathway inhibitor) effect of the inhibitory protein88 are all involved. Hemostasis requires a balance between TF and TFPI.
These pathways converge at a common point in the complement system. When activated, factors such as C5a can activate the TF by modifying its sulfhydryl radicals via PDI mediation.89
Anti-inflammatory cytokines such as IL-4, IL-10 and IL-13,90–93 as well as prostaglandin (PG) E1 and PGI2, can suppress TF in monocytes and macrophages.94
TF in Viral Infections
The coagulation system can be activated via TF during viral infections such as the Herpes simplex virus,95 the human immunodeficiency virus (HIV),96 the Coxsackievirus B3,97 dengue,98 or Ebola.99 This response most likely evolved as a host defense system to prevent the virus from spreading. However, acute viremia can lead to a type of consumptive coagulopathy, thus contributing to multiorgan failure and death.100 The activation of toll-like receptor 3 (TLR3) in the presence of double-stranded RNA viruses can induce TF expression in cultured endothelial cells and activate the coagulation system in mice.101 Note that the inhibition of the factor VIIa-tissue factor complex reduced cytokine storm and mortality in a Rhesus macaque model of Ebola hemorrhagic fever.102
TF in COVID-19
Three recent reviews speculate that induction of TF expression may play a role in COVID-19-related thrombosis.32–34 The correlation between TF expression and the severity of COVID-19 has been studied both its expression in monocytes103 and in extracellular vesicles.104 Platelets from severe COVID-19 patients were able to induce TF expression ex vivo in monocytes from healthy volunteers.105 TF can increase its expression during hypoxia and generate a thrombotic tendency; phenomenon that is mediated through extracellular RNA activated Toll-like receptor 3-activated protein 1 signaling.106
It is too early to conclude at which point in the cascade of biological events triggered in COVID-19 this TF overexpression is found; later studies will surely clarify it for us.
TF-Specific Inhibitors
There are vast therapeutic strategies focused on reducing the blockade of TF expression in prothrombotic conditions and it is the subject of various studies.107
There are 5 types of TF-specific inhibitors that are currently being examined to be medically administered as antithrombotic agents: monoclonal antibodies against TF,108 recombinant TFPI, Ixolaris (protein identified in the tick saliva with 2 Kunitz-like domains),109,110 molecules that interfere with the TF binding site with factor VIIa111 and anticoagulant nematode protein C2.112
Other Medications That Can Modulate TF Expression
Various drugs for regular use in different fields of medicine have been studied for their potential inhibitory role in TF expression. Published studies have been of a different nature, including in vitro, in vivo, and ex vivo studies, and usually report the intracellular mechanisms of action involved. Some of the drugs studied are angiotensin-converting enzyme 1 (ACE-1) inhibitors,113 antioxidants,114 clopidogrel,115 salicylates (acetylsalicylic acid, ibuprofen, indomethacin),116 specific COX-2-specific inhibitors (celecoxib),71 cyclosporine A,117 hydroxyurea,118 vitamin D,119 pentoxifylline,120 and Omega-3.121 Interestingly, glucocorticoids such as dexamethasone increase TF induction in LPS-stimulated monocytes have little effect on endothelial cells.122
Another source of TF activation inhibition may be through the suppression of the complement pathway, as could eculizumab, a C5-specific inhibitor, on which studies for COVID-19 are even being conducted.123 Other drugs such as various flavonoids 19,124 bacitracin125 or selenium,126 by means of inhibition of protein disulfide isomerase, are other potential possibilities.
TF Inhibition in COVID-19
As previously commented, it is too early to know the true role of the over-expression of TF in COVID-19; likewise, the possible therapies derived from these findings are just beginning to be formulated. Inhibition of TF could be a therapeutic strategy in patients with COVID-19 who are initiating thrombotic coagulopathy, whose criteria would be of a clinical nature associated with an increase in D-dimer. ACE-I drugs due to their mechanism of action have been the object of study during the pandemic; although it was initially thought that they could be beneficial,127 it has been shown that they do not intervene with the course of the disease, suggesting users of these medications should not discontinue or change their treatment.128 Antithrombotic therapy in COVID-19 is based in low molecular weight heparins and aspirin; intermediate-dose anticoagulation and aspirin were each associated with a lower cumulative incidence of in-hospital death.129 The impact of salicylate therapies on overexpressed TF in patients with COVID-19 remains to be demonstrated. Celecoxib, is a candidate for treatment of COVID-19, taking into account its effect as an anti-inflammatory medication130; it would also be necessary to study the effect as an inhibitor of TF. Cyclosporine A might be a candidate in severe COVID-19 to prevent the cytokine storm (or hyperinflammation) and inactivate viral replication (117) disregarding its potential role in inhibiting TF and possible antiviral effect.131 Pentoxifylline has also been proposed for the treatment of COVID-19 as a drug with antiviral, anti-inflammatory and bronchodilatory effects.132 Colchicine recently associated with impact of on mortality in patients with COVID-19133,134; In addition to its anti-inflammatory effect, it may also exhibit an inhibition of TF135,136 and has a possible antiviral effect.137
Vitamin D deficiency has been associated with risk for the development of COVID-19 and its deficiency must be corrected138; its effect on immune dynamics is postulated,139 however its effect as a TF inhibitor has yet to be demonstrated. Recent reviews speculate on the usefulness of antioxidants140,141 and selenium.126
Conclusion
The coagulopathy associated with COVID-19 appears to be multifactorial, involving inflammatory processes; endothelial and platelet dysfunction; the presence of antiphospholipid antibodies; and, because of the increase in AT2 in response to ACE-2 down-regulation, quite possibly TF overexpression. Different therapeutic targets have been derived from TF expression inhibition, which should be targeted by possible therapeutic candidates for the coagulopathy associated with COVID-19.
Acknowledgment
The authors thank Dr. David Ascher for the valuable review of article.
Footnotes
Author Contributions: All authors contributed to the preparation of the work and writing of the manuscript.
Declaration of Conflicting Interests: The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding: The author(s) received no financial support for the research, authorship, and/or publication of this article.
ORCID iD: Carlos A. Cañas  https://orcid.org/0000-0002-6879-3700
https://orcid.org/0000-0002-6879-3700
References
- 1. Zhu N, Zhang D, Wang W, et al. A novel coronavirus from patients with pneumonia in China, 2019. N Engl J Med. 2020;382(8):727–733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Zhou F, Yu T, Du R, et al. Clinical course and risk factors for mortality of adult in patients with COVID-19 in Wuhan, China: a retrospective cohort study. Lancet. 2020;395(10229):1054–1062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Yan R, Zhang Y, Li Y, Xia L, Guo Y, Zhou Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science. 2020;367(6485):1444–1448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Wang D, Hu B, Hu C, et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus-infected pneumonia in Wuhan, China. JAMA. 2020;323(11):1061–1069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Guan WJ, Ni ZY, Hu Y, et al. Clinical characteristics of coronavirus disease 2019 in China. N Engl J Med. 2020;382(18):1708–1720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Becker RC. COVID-19 update: Covid-19-associated coagulopathy. J Thromb Thrombolysis. 2020;50(1):54–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Jin Y, Yang H, Ji W, et al. Virology, epidemiology, pathogenesis, and control of COVID-19. Viruses. 2020;12(4):372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Zhang Y, Cao W, Xiao M, et al. Clinical and coagulation characteristics of 7 patients with critical COVID-2019 pneumonia and acro-ischemia. Zhonghua Xue Ye Xue Za Zhi. 2020;41(0):E006. [DOI] [PubMed] [Google Scholar]
- 9. Danzi GB, Lo M, Galeazzi G, Gherbesi E. Acute pulmonary embolism and COVID-19 pneumonia: a random association? Eur Heart J. 2020;41(19):1858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Klok FA, Kruip MJHA, van der Meer NJM, et al. Incidence of thrombotic complications in critically ill ICU patients with COVID-19. Thromb Res. 2020;191:145–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Tang N, Li D, Wang X, Sun Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J Thromb Haemost. 2020;18(4):844–847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. White D, MacDonald S, Edwards T, et al. Evaluation of COVID-19 coagulopathy; laboratory characterization using thrombin generation and nonconventional haemostasis assays. Int J Lab Hematol. 2021;43(2):123–130. [DOI] [PubMed] [Google Scholar]
- 13. Hamming I, Cooper ME, Haagmans BL, et al. The emerging role of ACE2 in physiology and disease. J Pathol. 2007;212(1):1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Li W, Moore MJ, Vasilieva N, et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature. 2003;426(6965):450–454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Harrison DG, Guzik TJ, Lob HE, et al. Inflammation, immunity, and hypertension. Hypertension. 2011;57(2):132–140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Senchenkova EY, Russell J, Kurmaeva E, et al. Role of T lymphocytes in angiotensin II-mediated microvascular thrombosis. Hypertension. 2011;58(5):959–965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Dmitrieva NI, Burg MB. Elevated sodium and dehydration stimulate inflammatory signaling in endothelial cells and promote atherosclerosis. PLoS One. 2015;10(6):e0128870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Brown NJ, Vaughan DE. Prothrombotic effects of angiotensin. Adv Intern Med. 2000;45:419–429. [PubMed] [Google Scholar]
- 19. Hou T, Tieu B, Ray S, et al. Roles of IL-6-gp130 signaling in vascular inflammation. Curr Cardiol Rev. 2008;4(3):179–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Ishibashi T, Kimura H, Shikama Y, et al. Interleukin-6 is a potent thrombopoietic factor in vivo in mice. Blood. 1989;74(4):1241–1244. [PubMed] [Google Scholar]
- 21. Tang XL, Jiang ZY, Dong J, et al. Expression of tissue factor induced by IL-6 in HUVEC. Sichuan Da Xue Xue Bao Yi Xue Ban. 2006;37(2):234–237. [PubMed] [Google Scholar]
- 22. Guzik TJ, Hoch NE, Brown KA, et al. Role of the T cell in the genesis of angiotensin II induced hypertension and vascular dysfunction. J Exp Med. 2007;204(10):2449–2460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Sun J, Axelsson J, Machowska A, et al. Biomarkers of cardiovascular disease and mortality risk in patients with advanced CKD. Clin J Am Soc Nephrol. 2016;11(7):1163–1172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Bautista LE, Vera LM, Arenas IA, Gamarra G. Independent association between inflammatory markers (C-reactive protein, interleukin-6, and TNF-α) and essential hypertension. J Hum Hypertens. 2005;19(2):149–154. [DOI] [PubMed] [Google Scholar]
- 25. Zuo Y, Yalavarthi S, Shi H, et al. Neutrophil extracellular traps in COVID-19. JCI Insight. 2020;5(11):e138999. doi:10.1172/jci.insight.138999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Liberale L, Holy EW, Akhmedov A, et al. Interleukin-1β mediates arterial thrombus formation via NET-associated tissue factor. J Clin Med. 2019;8(12):E2072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. O’Neill LA, Bowie AG. The family of five: TIR-domain-containing adaptors in Toll-like receptor signaling. Nat Rev Immunol. 2007;7(5):353–364. [DOI] [PubMed] [Google Scholar]
- 28. Choudhury A, Mukherjee S. In silico studies on the comparative characterization of the interactions of SARS-CoV-2 spike glycoprotein with ACE-2 receptor homologs and human TLRs. J Med Virol. 2020;92(10):2105–2113. doi:10.1002/jmv.25987 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Biswas I, Khan GA. Coagulation disorders in COVID-19: role of toll-like receptors. J Inflamm Res. 2020;13:823–828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Mackman N. Regulation of the tissue factor gene. FASEB J. 1995;9(10):883–889. [DOI] [PubMed] [Google Scholar]
- 31. Lösche W. Platelets and tissue factor. Platelets. 2005;16(6):313–319. [DOI] [PubMed] [Google Scholar]
- 32. Bautista-Vargas M, Bonilla-Abadía F, Cañas CA. Potential role for tissue factor in the pathogenesis of hypercoagulability associated with in COVID-19. J Thromb Thrombolysis. 2020;50(3):1–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. DiNicolantonio JJ, McCarty M. Thrombotic complications of COVID-19 may reflect an upregulation of endothelial tissue factor expression that is contingent on activation of endosomal NADPH oxidase. Open Heart. 2020;7(1):e001337. doi:10.1136/openhrt-2020-001337 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Mackman N, Antoniak S, Wolberg AS, Kasthuri R, Key NS. Coagulation abnormalities and thrombosis in patients infected with SARS-CoV-2 and other pandemic viruses. Arterioscler Thromb Vasc Biol. 2020;40(9):2033–2044. doi:10.1161/ATVBAHA.120.314514 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Ackermann M, Verleden SE, Kuehnel M, Haverich A, Welte T. Pulmonary vascular endothelialitis, thrombosis, and angiogenesis in Covid-19. N Engl J Med. 2020;383(2). 120–128. doi:10.1056/NEJMoa2015432 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Goshua G, Pine AB, Meizlish ML, et al. Endotheliopathy in COVID-19-associated coagulopathy: evidence from a single-centre, cross-sectional study. Lancet Haematol. 2020;7(8):e575–e582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Schwameis M, Schörgenhofer C, Assinger A, Steiner MM, Jilma B. VWF excess and ADAMTS13 deficiency: a unifying pathomechanism linking inflammation to thrombosis in DIC, malaria, and TTP. Thromb Haemost. 2015;113(4):708–718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Bray MA, Sartain SA, Gollamudi J, Rumbaut RE. Microvascular thrombosis: experimental and clinical implications. Transl Res. 2020;23;S1931-5244(20)30108-30100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Wang Z, Yang Y, Liang X, et al. COVID-19 associated ischemic stroke and hemorrhagic stroke: incidence, potential pathological mechanism, and management. Front Neurol. 2020;11:571996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Magro C, Mulvey JJ, Berlin D, et al. Complement associated microvascular injury and thrombosis in the pathogenesis of severe COVID-19 infection: a report of five cases. Transl Res. 2020;220:1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Noris M, Benigni A, Remuzzi G. The case of complement activation in COVID-19 multiorgan impact. Kidney Int. 2020;98(2);314–322. S0085-2538(20)30556-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Fletcher-Sandersjöö A, Bellander BM. Is COVID-19 associated thrombosis caused by overactivation of the complement cascade? A literature review. Thromb Res. 2020;194:36–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Chauhan AJ, Wiffen LJ, Brown TP. COVID-19: a collision of complement, coagulation and inflammatory pathways. J Thromb Haemost. 2020. doi:10.1111/jth.14981 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Subramaniam S, Jurk K, Hobohm L, et al. Distinct contributions of complement factors to platelet activation and fibrin formation in venous thrombus development. Blood. 2017;129(1):2291–2302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Ritis K, Doumas M, Mastellos D, et al. A novel C5a receptor-tissue factor cross-talk in neutrophils links innate immunity to coagulation pathways. J Immunol. 2006;177:4794–4802. [DOI] [PubMed] [Google Scholar]
- 46. Calderon-Lopez MY, Garcia-Leon N, Gomez-Arevalillo S, Martin-Serrano P, Matilla-Garcia A. Coronavirus disease 2019 and coagulopathy: other prothrombotic coagulation factors. Blood Coagul Fibrinolysis. 2021;32:44–49. [DOI] [PubMed] [Google Scholar]
- 47. Griffin JH, Lyden P. COVID-19 hypothesis: Activated protein C for therapy of virus-induced pathologic thromboinflammation. Res Pract Thromb Haemost. 2020;4:506–509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Morawitz P. Beitrage zur kenntnis der blutgerinnung. Beitr Chem Physio. Pathol. 1904;5:133–141. [Google Scholar]
- 49. Lu G, Broze GJ, Jr, Krishnaswamy S. Formation of factors IXa and Xa by the extrinsic pathway: differential regulation by tissue factor pathway inhibitor and antithrombin III. J Biol Chem. 2004;279(17):17241–17249. [DOI] [PubMed] [Google Scholar]
- 50. Yamamoto M, Nakagaki T, Kisiel W. Tissue factor-dependent autoactivation of human blood coagulation factor VII. J Biol Chem. 1992;267:19089–19094. [PubMed] [Google Scholar]
- 51. Davidson CJ, Tuddenham EG, McVey JH. 450 million years of hemostasis. J Thromb Haemost. 2003;1(7):1487–1494. [DOI] [PubMed] [Google Scholar]
- 52. Mackman N, Morrissey JH, Fowler B, Edgington TS. Complete sequence of the human tissue factor gene, a highly regulated cellular receptor that initiates the coagulation protease cascade. Biochemistry. 1989;28(4):1755–1762. [DOI] [PubMed] [Google Scholar]
- 53. Bazan JF. Structural design and molecular evolution of a cytokine receptor superfamily. Proc Natl Acad Sci U S A. 1990;87(18):6934–6938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Harlos K, Martin DM, O’Brien DP, et al. Crystal structure of the extracellular region of human tissue factor. Nature. 1994;370(6491):662–666. [DOI] [PubMed] [Google Scholar]
- 55. Coughlin SR. Sol Sherry lecture in thrombosis: how thrombin ‘talks’ to cells: molecular mechanisms and roles in vivo. Arterioscler Thromb Vasc Biol. 1998;18(4):514–518. [DOI] [PubMed] [Google Scholar]
- 56. Pyo RT, Sato Y, Mackman N, Taubman MB. Mice deficient in tissue factor demonstrate attenuated intimal hyperplasia in response to vascular injury and decreased smooth muscle cell migration. Thromb Haemost. 2004;92(3):451–458. [DOI] [PubMed] [Google Scholar]
- 57. Carmeliet P, Mackman N, Moons L, et al. Role of tissue factor in embryonic blood vessel development. Nature. 1996;383(6595):73–75. [DOI] [PubMed] [Google Scholar]
- 58. Ahamed J, Versteeg HH, Kerver M, Chen VM, Mueller BM. Disulfide isomerization switches tissue factor from coagulation to cell signaling. Proc Natl Acad Sci USA. 2006;103(38):13932–13937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Versteeg HH, Ruf W. Thiol pathways in the regulation of tissue factor prothrombotic activity. Curr Opin Hematol. 2011;18(5):343–348. [DOI] [PubMed] [Google Scholar]
- 60. Parry GC, Mackman N. Transcriptional regulation of tissue factor expression in human endothelial cells. Arterioscler Thromb Vasc Biol. 1995;15(5):612–621. [DOI] [PubMed] [Google Scholar]
- 61. Schwertz H, Tolley ND, Foulks JM, et al. Signal-dependent splicing of tissue factor pre-mRNA modulates the thrombogenicity of human platelets. J Exp Med. 2006;203(11):2433–2440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Ritis K, Doumas M, Mastellos D, et al. A novel C5a receptor-tissue factor cross-talk in neutrophils links innate immunity to coagulation pathways. J Immunol. 2006;177(7):4794–4802. [DOI] [PubMed] [Google Scholar]
- 63. De Palma R, Cirillo P, Ciccarelli G, et al. Expression of functional tissue factor in activated T-lymphocytes in vitro and in vivo: a possible contribution of immunity to thrombosis? Int J Cardiol. 2016;218:188–195. [DOI] [PubMed] [Google Scholar]
- 64. Mandal SK, Iakhiaev A, Pendurthi UR, Rao LV. Acute cholesterol depletion impairs functional expression of tissue factor in fibroblasts: modulation of tissue factor activity by membrane cholesterol. Blood. 2005;105(1):153–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Pendurthi UR, Rao LV. Role of tissue factor disulfides and lipid rafts in signaling. Thromb Res. 2008;122(suppl 1):S14–S18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Versteeg HH, Schaffner F, Kerver M, et al. Inhibition of tissue factor signaling suppresses tumor growth. Blood. 2008;111(1):190–199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Scholz T, Temmler U, Krause S, Heptinstall S, Losche W. Transfer of tissue factor from platelets to monocytes: role of platelet-derived microvesicles and CD62P. Thromb Haemost. 2002;88(6):1033–1038. [PubMed] [Google Scholar]
- 68. Mackman N, Luther T. Platelet tissue factor: to be or not to be. Thromb Res. 2013;132(1):3–5. [DOI] [PubMed] [Google Scholar]
- 69. Grover SP, Mackman N. Tissue factor an essential mediator of hemostasis and trigger of thrombosis. Arterioscler Thromb Vasc Biol. 2018;38(4):709–725. [DOI] [PubMed] [Google Scholar]
- 70. Hoffman M, Whinna HC, Monroe DM. Circulating tissue factor accumulates in thrombi, but not in hemostatic plugs. J Thromb Haemost. 2006;4(9):2092–2093. [DOI] [PubMed] [Google Scholar]
- 71. Steffel J, Hermann M, Greutert H, et al. Celecoxib decreases endothelial tissue factor expression through inhibition of c-Jun terminal NH2 kinase phosphorylation. Circulation. 2005;111(13):1685–1689. [DOI] [PubMed] [Google Scholar]
- 72. Napoleone E, Di Santo A, Lorenzet R. Monocytes upregulate endothelial cell expression of tissue factor: a role for cell-cell contact and cross-talk. Blood. 1997;89(2):541–549. [PubMed] [Google Scholar]
- 73. Bavendiek U, Libby P, Kilbride M, Reynolds R, Mackman N, Schonbeck U. Induction of tissue factor expression in human endothelial cells by CD40 ligand is mediated via activator protein 1, nuclear factor kappa B, and Egr-1. J Biol Chem. 2002;277(28):25032–25039. [DOI] [PubMed] [Google Scholar]
- 74. Camera M, Giesen PL, Fallon J, et al. Cooperation between VEGF and TNF-alpha is necessary for exposure of active tissue factor on the surface of human endothelial cells. Arterioscler Thromb Vasc Biol. 1999;19(3):531–537. [DOI] [PubMed] [Google Scholar]
- 75. Kawano H, Tsuji H, Nishimura H, et al. Serotonin induces the expression of tissue factor and plasminogen activator inhibitor-1 in cultured rat aortic endothelial cells. Blood. 2001;97(6):1697–1702. [DOI] [PubMed] [Google Scholar]
- 76. Steffel J, Akhmedov A, Greutert H, Luscher TF, Tanner FC. Histamine induces tissue factor expression: implications for acute coronary syndromes. Circulation. 2005;112(3):341–349. [DOI] [PubMed] [Google Scholar]
- 77. Cermak J, Key NS, Bach RR, Balla J, Jacob HS, Vercellotti GM. C-reactive protein induces human peripheral blood monocytes to synthesize tissue factor. Blood. 1993;82(2):513–520. [PubMed] [Google Scholar]
- 78. Celi A, Pellegrini G, Lorenzet R, et al. P-selectin induces the expression of tissue factor on monocytes. Proc Natl Acad Sci USA. 1994;91(19):8767–8771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Hölschermann H, Dürfeld F, Maus U, et al. Cyclosporine an inhibits tissue factor expression in monocytes/macrophages. Blood. 1996;88(10):3837–3845. [PubMed] [Google Scholar]
- 80. Hoffman M, Monroe DM III. A cell-based model of hemostasis. Thromb Haemost. 2001;85(6):958–965. [PubMed] [Google Scholar]
- 81. Schabbauer G, Schweighofer B, Mechtcheriakova D, Lucerna M, Binder BR, Hofer E. Nuclear factor of activated T cells and early growth response-1 cooperate to mediate tissue factor gene induction by vascular endothelial growth factor in endothelial cells. Thromb Haemost. 2007;97(6):988–997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Reverter JC, Tàssies D, Font J, et al. Hypercoagulable state in patients with antiphospholipid syndrome is related to high induced tissue factor expression on monocytes and to low free protein S. Arterioscler Thromb Vasc Biol. 1996;16(11):1319–1326. [DOI] [PubMed] [Google Scholar]
- 83. Cuadrado MJ, López-Pedrera C, Khamashta MA, et al. Thrombosis in primary antiphospholipid syndrome: a pivotal role for monocyte tissue factor expression. Arthritis Rheum. 1997;40(5):834–841. [DOI] [PubMed] [Google Scholar]
- 84. Agmon-Levin N, Blank M, Zandman-Goddard G, et al. Vitamin D: an instrumental factor in the anti-phospholipid syndrome by inhibition of tissue factor expression. Ann Rheum Dis. 2011;70(1):145–150. [DOI] [PubMed] [Google Scholar]
- 85. Rao LV, Pendurthi UR. Regulation of tissue factor coagulant activity on cell surfaces. J Thromb Haemost. 2012;10(11):2242–2253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Shaw AW, Pureza VS, Sligar SG, Morrissey JH. The local phospholipid environment modulates the activation of blood clotting. J Biol Chem. 2007;282(9):6556–6563. [DOI] [PubMed] [Google Scholar]
- 87. Wolberg AS, Monroe DM, Roberts HR, Hoffman MR. Tissue factor de-encryption: ionophore treatment induces changes in tissue factor activity by phosphatidylserine-dependent and -independent mechanisms. Blood Coagul Fibrinolysis. 1999;10(4):201–210. [PubMed] [Google Scholar]
- 88. Bach R, Rifkin DB. Expression of tissue factor procoagulant activity: regulation by cytosolic calcium. Proc Natl Acad Sci USA. 1990;87(18):6995–6999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Ikeda K, Nagasawa K, Horiuchi T, Tsuru T, Nishizaka Niho Y. C5a induces tissue factor activity on endothelial cells. Thromb Haemost. 1997;77(2):394–398. [PubMed] [Google Scholar]
- 90. Lindmarck E, Tenno T, Chen J, Siegbahn A. IL-10 inhibits LPS-induced human monocyte tissue factor expression in whole blood. Br J Haematol. 1998;102(2):597–604. [DOI] [PubMed] [Google Scholar]
- 91. Herbert JM, Savi P, Laplace MC, et al. IL-4 and IL-13 exhibit comparable abilities to reduce pyrogen-induced expression of procoagulant activity in endothelial cells and monocytes. FEBS Lett. 1993;328(3):268–270. [DOI] [PubMed] [Google Scholar]
- 92. Osnes LT, Westvik AB, Joo GB, Okkenhaug C, Kierulf P. Inhibition of IL-1 induced tissue factor (TF) synthesis and procoagulant activity (PCA) in purified human monocytes by IL-4, IL-10 and IL-13. Cytokine. 1996;8(11):822–827. [DOI] [PubMed] [Google Scholar]
- 93. Ernofsson M, Tenno T, Siegbahn A. Inhibition of tissue factor surface expression in human peripheral blood monocytes exposed to cytokines. Br J Haematol. 1996:95(2):249–257. [DOI] [PubMed] [Google Scholar]
- 94. Crutchley DJ, Hirsh MJ. The stable prostacyclin analog, iloprost and prostaglandin E1 inhibit monocyte procoagulant activity in vitro. Blood. 1991;78(2):382–386. [PubMed] [Google Scholar]
- 95. Key NS, Vercellotti GM, Winkelmann JC, et al. Infection of vascular endothelial cells with herpes simplex virus enhances tissue factor activity and reduces thrombomodulin expression. Proc Natl Acad Sci USA. 1990;87(18):7095–7099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Funderburg NT, Mayne E, Sieg SF, et al. Increased tissue factor expression on circulating monocytes in chronic HIV infection: relationship to in vivo coagulation and immune activation. Blood. 2010;115(2):161–167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Antoniak S, Boltzen U, Riad A, et al. Viral myocarditis and coagulopathy: increased tissue factor expression and plasma thrombogenicity. J Mol Cell Cardiol. 2008;45(1):118–126. [DOI] [PubMed] [Google Scholar]
- 98. Huerta-Zepeda A, Cabello-Gutierrez C, Cime-Castillo J, et al. Crosstalk between coagulation and inflammation during dengue virus infection. Thromb Haemost. 2008;99(5):936–943. [DOI] [PubMed] [Google Scholar]
- 99. Geisbert TW, Young HA, Jahrling PB, Davis KJ, Kagan E, Hensley LE. Mechanisms underlying coagulation abnormalities in Ebola hemorrhagic fever: overexpression of tissue factor in primate monocytes/macrophages is a key event. J Infect Dis. 2003;188(11):1618–1629. [DOI] [PubMed] [Google Scholar]
- 100. Goeijenbier M, van Wissen M, van de Weg C, et al. Review: viral infections and mechanisms of thrombosis and bleeding. J Med Virol. 2012;84(10):1680–1696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Shibamiya A, Hersemeyer K, Schmidt Woll T, et al. A key role for Toll-like receptor-3 in disrupting the hemostasis balance on endothelial cells. Blood. 2009;113(3):714–722. [DOI] [PubMed] [Google Scholar]
- 102. Geisbert TW, Hensley LE, Jahrling PB, et al. Treatment of Ebola virus infection with a recombinant inhibitor of factor VIIa/tissue factor: a study in Rhesus monkeys. Lancet. 2003;362(9400):1953–1958. [DOI] [PubMed] [Google Scholar]
- 103. Hottz ED, Azevedo-Quintanilha IG, Palhinha L, et al. Platelet activation and platelet-monocyte aggregate formation trigger tissue factor expression in patients with severe COVID-19. Blood. 2020;136(11):1330–1341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Rosell A, Havervall S, von Meijenfeldt F, et al. Patients with COVID-19 have elevated levels of circulating extracellular vesicle tissue factor activity that is associated with severity and mortality. Arterioscler Thromb Vasc Biol. 2020;41(2):ATVBAHA120315547. doi:10.1161/ATVBAHA.120.315547 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Houston DS. Tissue factor—a therapeutic target for thrombotic disorders. Expert Opin Ther Targets. 2002;6(2):159–174. [DOI] [PubMed] [Google Scholar]
- 106. Bhagat S, Biswas I, Ahmed R, Khan GA. Hypoxia induced up-regulation of tissue factor is mediated through extracellular RNA activated Toll-like receptor 3-activated protein 1 signalling. Blood Cells Mol Dis. 2020. September;84:102459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Golino P. The inhibitors of the tissue factor: factor VII pathway. Thromb Res. 2002;106(3):V257–V265. [DOI] [PubMed] [Google Scholar]
- 108. Francischetti IM, Valenzuela JG, Andersen JF, Mather TN, Ribeiro JM. Ixolaris, a novel recombinant tissue factor pathway inhibitor (TFPI) from the salivary gland of the tick, Ixodes scapularis: identification of factor X and factor Xa as scaffolds for the inhibition of factor VIIa/tissue factor complex. Blood. 2002;99(10):3602–3612. [DOI] [PubMed] [Google Scholar]
- 109. Corral-Rodríguez MA, Macedo-Ribeiro S, Barbosa Pereira PJ, Fuentes-Prior P. Tick-derived Kunitz-type inhibitors as antihemostatic factors. Insect Biochem Mol Biol. 2009;39(9):579–595. [DOI] [PubMed] [Google Scholar]
- 110. Rao LV, Ezban M. Active site-blocked activated factor VII as an effective antithrombotic agent: mechanism of action. Blood Coagul Fibrinolysis. 2000;11(suppl. 1):S135–S143. [DOI] [PubMed] [Google Scholar]
- 111. Lee AY, Vlasuk GP. Recombinant nematode anticoagulant protein c2 and other inhibitors targeting blood coagulation factor VIIa/tissue factor. J Intern Med. 2003;254(4):313–321. [DOI] [PubMed] [Google Scholar]
- 112. Giugliano RP, Wiviott SD, Stone PH, et al. ANTHEM-TIMI-32 Investigators. Recombinant nematode anticoagulant protein c2 in patients with non-ST-segment elevation acute coronary syndrome: the ANTHEM-TIMI-32 trial. J Am Coll Cardiol. 2007;49(25):2398–2407. [DOI] [PubMed] [Google Scholar]
- 113. Brambilla M, Gelosa P, Rossetti L, et al. Impact of angiotensin-converting enzyme inhibition on platelet tissue factor expression in stroke-prone rats. J Hypertens. 2018;36(6):1360–1371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114. Francischetti IM, Gordon E, Bizzarro B, et al. Tempol, an intracellular antioxidant, inhibits tissue factor expression, attenuates dendritic cell function, and is partially protective in a murine model of cerebral malaria. PLoS One. 2014;9(2):e87140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Savi P, Bernat A, Dumas A, Aït-Chek L, Herbert JM. Effect of aspirin and clopidogrel on platelet-dependent tissue factor expression in endothelial cells. Thromb Res. 1994;73(2):117–124. [DOI] [PubMed] [Google Scholar]
- 116. Oeth P, Mackman N. Salicylates inhibit lipopolysaccharide-induced transcriptional activation of the tissue factor gene in human monocytic cells. Blood. 1995;86(11):4144–4152. [PubMed] [Google Scholar]
- 117. Pathania YS. Cyclosporine: hope for severe COVID-19? BMJ Support Palliat Care. 2021;bmjspcare–2020-002681. doi:10.1136/bmjspcare-2020-002681 [DOI] [PubMed] [Google Scholar]
- 118. Maugeri N, Giordano G, Petrilli MP, et al. Inhibition of tissue factor expression by hydroxyurea in polymorphonuclear leukocytes from patients with myeloproliferative disorders: a new effect for an old drug? J Thromb Haemost. 2006;4(12):2593–2598. [DOI] [PubMed] [Google Scholar]
- 119. Chung J, Koyama T, Ohsawa M, Shibamiya A, Hoshi A, Hirosawa S. 1,25(OH)(2)D(3) blocks TNF-induced monocytic tissue factor expression by inhibition of transcription factors AP-1 and NF-kappaB. Lab Invest. 2007;87(6):540–547. [DOI] [PubMed] [Google Scholar]
- 120. De Prost D, Ollivier V, Hakim J. Pentoxifylline inhibition of procoagulant activity generated by activated mononuclear phagocytes. Mol Pharmacol. 1990;38(4):562–566. [PubMed] [Google Scholar]
- 121. Tremoli E, Eligini S, Colli S, et al. Effects of omega 3 fatty acid ethyl esters on monocyte tissue factor expression. World Rev Nutr Diet. 1994;76:55–59. [DOI] [PubMed] [Google Scholar]
- 122. Bottles KD, Morrissey JH. Dexamethasone enhances agonist induction of tissue factor in monocytes but not in endothelial cells. Blood Coagul Fibrinolysis. 1993;4(3):405–414. [DOI] [PubMed] [Google Scholar]
- 123. COVID-19 Clinical Trials. 2020. (NCT04288713).
- 124. Wang L, Essex DW. A new antithrombotic strategy: inhibition of the c-terminal active site of protein disulfide isomerase. J Thromb Haemost. 2017;15(4):770–773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Karala AR, Ruddock LW. Bacitracin is not a specific inhibitor of protein disulfide isomerase. FEBS J. 2010;277(11):2454–2462. [DOI] [PubMed] [Google Scholar]
- 126. Kieliszek M, Lipinski B. Selenium supplementation in the prevention of coronavirus infections (COVID-19). Med Hypotheses. 2020;143:109878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Şenkal N, Meral R, Medetalibeyoğlu A, Konyaoğlu H, Kose M, Tukek T. Association between chronic ACE inhibitor exposure and decreased odds of severe disease in patients with COVID-19. Anatol J Cardiol. 2020;24(1):21–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Morales DR, Conover MM, You SC, et al. Renin-angiotensin system blockers and susceptibility to COVID-19: an international, open science, cohort analysis. Lancet Digit Health. 2020;3(2):e98–e114. S2589-7500(20)30289-2. doi:10.1016/S2589-7500(20)30289-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Meizlish ML, Goshua G, Liu Y, et al. Intermediate-dose anticoagulation, aspirin, and in-hospital mortality in COVID-19: a propensity score-matched analysis. Am J Hematol. 2021;96(4):471–479. doi:10.1002/ajh.26102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Baghaki S, Yalcin CE, Baghaki HS, Aydin SY, Daghan B, Yavuz E. COX2 inhibition in the treatment of COVID-19: review of literature to propose repositioning of celecoxib for randomized controlled studies. Int J Infect Dis. 2020;101:29–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Glowacka P, Rudnicka L, Warszawik-Hendzel O, et al. The antiviral properties of cyclosporine. focus on coronavirus, hepatitis C virus, influenza virus, and human immunodeficiency virus infections. Biology (Basel). 2020;9(8):192. doi:10.3390/biology9080192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132. Ghasemnejad-Berenji M, Pashapour S, Sadeghpour S. Pentoxifylline, a drug with antiviral, anti-inflammatory and bronchodilatory effects may be a possible drug candidate for SARS-CoV-2. Med Princ Pract. 2020;30(1):98–100. doi:10.1159/000512234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. Vrachatis DA, Giannopoulos GV, Giotaki SG, et al. Impact of colchicine on mortality in patients with COVID-19. A meta-analysis. Hellenic J Cardiol. 2021;S1109-9666(20):30285–30292. doi:10.1016/j.hjc.2020.11.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Pinzón MA, Arango DC, Betancur JF, et al. Outcome of patients with COVID-19 pneumonia treated with corticosteroids and colchicine in Colombia. Ann Clin Microbiol Antimicrob. 2020;2020. 10.21203/rs.3.rs-94922/v [DOI] [PMC free article] [PubMed]
- 135. Cimmino G, Conte S, Pellegrino G, Ciccarelli G, Barra G. Colchicine inhibits tissue factor expression in activated Tlymphocytes via NFkB. Eur Heart J. 2016;37. Abstract Supplement: 587:P2852. [Google Scholar]
- 136. Cimmino G, Conte S, Morello A, et al. Colchicine inhibits the prothrombotic effects of oxLDL in human endothelial cells. Vascul Pharmacol. 2021. April;137:106822. doi:10.1016/j.vph.2020.106822 [DOI] [PubMed] [Google Scholar]
- 137. Richter M, Boldescu V, Graf D, et al. Synthesis, biological evaluation, and molecular docking of combretastatin and colchicine derivatives and their hce1-activated prodrugs as antiviral agents. ChemMedChem. 2019;14(4):469–483. doi:10.1002/cmdc.201800641 [DOI] [PubMed] [Google Scholar]
- 138. Katz J, Yue S, Xue W. Increased risk for COVID-19 in patients with vitamin D deficiency. Nutrition. 2020;84:111106. doi:10.1016/j.nut.2020.111106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Kalia V, Studzinski GP, Sarkar S. Role of vitamin D in regulating COVID-19 severity—an immunological perspective. J Leukoc Biol. 2021. doi:10.1002/JLB.4COVR1020-698 R [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Kassi EN, Papavassiliou KA, Papavassiliou AG. Defective anti-oxidant system: an aggravating factor for COVID-19 patients outcome? Arch Med Res. 2020;51(7):726–727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Qin M, Cao Z, Wen J, et al. An antioxidant enzyme therapeutic for COVID-19. Adv Mater. 2020;32(43):e2004901. doi:10.1002/adma.202004901 [DOI] [PubMed] [Google Scholar]