

Abstract
Magnetoencephalography (MEG) is a noninvasive, silent, and totally passive neurophysiological imaging method with excellent temporal resolution (~1 ms) and good spatial precision (~3–5 mm). While MEG studies of neuroHIV remain relatively rare, the number of studies per year has sharply increased recently and this trend will likely continue into the foreseeable future. The current in-depth review focuses on the studies that have been conducted to date, which include investigations of somatosensory and visual modalities, resting-state, as well as motor control and higher-level functions such as working memory and visual attention. The review begins with an introduction to the principles and methods of MEG, and then transitions to a review of each of the empirical studies that have been conducted to date, separated by sensory modality for the basic studies and cognitive domain for the higher-level investigations. As such, this review attempts to be exhaustive in its coverage of empirical MEG studies of neuroHIV. Across studies major themes emerge including aberrant neural oscillatory activity in HIV-infected adults, both in primary sensory regions of the brain and higher-order executive regions. Many studies have also connected the amplitude of neural oscillations to behavioral and/or neuropsychological function in the study population, making a vital connection to performance and improving the veracity of the findings. One conspicuous emerging area is the use of MEG to distinguish cognitively-impaired from unimpaired HIV-infected adults, with major success reported and future studies sure to come. The review concludes with a summary of findings and suggested focus areas for future studies.
Keywords: HAND, oscillation, HIV-infected, alpha, theta, gamma, beta, aging, motor, attention, visual, connectivity
Introduction
With the availability of potent combination antiretroviral therapy (cART), the nature of HIV-infection dramatically shifted from a terminal illness to a chronic manageable condition, with a life expectancy that approaches that of seronegative persons.1–2 Despite these advances, HIV-infected patients remain at a significantly increased risk of developing HIV-associated neurocognitive disorders (HAND),3–8 and recent studies have shown that 35–70% of all HIV-infected patients are at least subtly impaired on neuropsychological tests of cognitive function.1–7 This persistence is at least partially driven by people simply living longer with milder neurocognitive disorders, although the trend also reflects that cART is far more effective in the periphery than in the brain. Overall, the prevalence of HIV-associated dementia (HAD) has sharply decreased in the cART era,3–4,9–12 while that of milder forms of HAND has remained largely the same or slightly increased.3–11 Patients with HAND are more likely to be unemployed or under employed, have greater problems with medication adherence, and have lower quality of life.13–15 Thus, the economic and societal burdens of HIV/AIDS are accentuated in cases of HAND.
Unfortunately, despite major scientific efforts, the neurobiological underpinnings of HAND remain incompletely understood. Numerous studies using structural and functional neuroimaging in humans have successfully identified the brain regions and networks most affected by the disease, but we still have a limited understanding of the inherent physiology in these brain regions, and how it is different in patients with HAND. There are also no current biomarkers of HAND, and hence diagnoses are based on standard neuropsychological testing and the exclusion of other contributing factors (e.g., comorbid conditions). Thus, there is a clear need for additional research using state-of-the-art tools to advance the field. One such tool that has seen increasing use in neuroHIV is magnetoencephalography (MEG). Compared to the other neuroimaging methods discussed in the accompanying chapters, MEG is unique in that it provides precise spatiotemporal data on the neural dynamics that serve cognitive processing. In other words, MEG is capable of characterizing neuronal activity on a millisecond temporal scale, which is critical given the extremely rapid time scale of neuronal traffic and circuit-level neural activity in the human brain.
In the current review, we start by providing some background information on MEG, including its physical and physiological origins, basic experimental design, terminology, and output variables. We then transition to empirical studies that have used MEG in the context of neuroHIV, which we subdivide into sections based on the cognitive and/or sensory process of interest. We then close by highlighting general conclusions and areas of consensus, pointing out areas in need of further study, and recommending future directions.
Magnetoencephalographic (MEG) Imaging: A Primer
MEG is a noninvasive human neurophysiological recording method that is primarily utilized in studies of systems-level brain function. The physical signal digitized in MEG is comprised of summated magnetic fields produced by ionic currents within active neural populations. Just as electric current within power lines produces magnetic fields, ionic currents within active neurons also produce magnetic fields according to the same physical principles (e.g., “the right-hand rule” of magnetism).16 These so-called neuromagnetic fields are ultra-minute (10−15 T) relative to other magnetic fields in the environment (e.g., the earth’s static field is about 10−5 T), and thus MEG signals are normally recorded in magnetically-shielded rooms using special sensors and/or noise-cancellation software.16–21 In regard to the physiological basis of these ionic currents, MEG is most sensitive to dendritic currents in large populations of cortical pyramidal cells, as opposed to their action potentials.16,22 However, other cortical neurons also contribute to the signal, as do cells in subcortical and cerebellar regions to a lesser extent.22 The depth (i.e., distance from the MEG sensors), orientation, and synchronicity of the neural populations also affect the strength of MEG signals, although modern MEG systems are generally sensitive to responses across the entire brain.23 Importantly, signal measurement in MEG occurs at the sensor-level and state-of-the-art systems are equipped with a large number of sensors (e.g., ~300) arranged in a helmet-like array that surrounds the participants head (Figure 1). Consequently, it is necessary to project these sensor measurements into source space (i.e., anatomical coordinates), which is generally accomplished using volume conductor models and optimization algorithms such as minimum-norm estimation, beamforming, and/or other related source estimation techniques.16–21 This process of projecting the MEG signals into the brain is often termed MEG source reconstruction.
Figure 1: Modern Magnetoencephalography (MEG) System.
A whole-head MEG system equipped with 306 cortical sensors is shown. MEG instruments are generally housed in magnetically-shielded rooms to reduce interference from magnetic fields in the local environment.
Although the first MEG measurements occurred over 50 years ago,24 the analytical sophistication and instrumentation of the field have dramatically increased over the past decade.17–21 MEG is currently the only noninvasive high-resolution imaging technique that measures the neurophysiology directly (i.e., does not rely on vascular responses) and the only functional brain imaging method to possess both high spatial (2–5 mm) and temporal resolution (i.e., millisecond level).20–21 Another unique advantage of MEG over other electrophysiological and noninvasive imaging methods is that it does not require a reference electrode, which allows local spontaneous activity to be accurately quantified, greatly simplifies the interpretation of responses, and facilitates network modeling and connectivity analyses.
As mentioned above, this review focuses on the use of MEG in people living with HIV-infection. The main applications in this population include the identification of potential biomarkers of cognitive impairment secondary to HIV-infection (i.e., HAND), and the generation of basic knowledge concerning the neural origin and mechanisms of HAND. Of note, this review attempts to be exhaustive in its coverage of MEG in neuroHIV, as the first papers on this topic appeared in 2012 and to date there are only 13 published articles listed in PubMed (according to a search of “HIV and magnetoencephalography” and “HIV and MEG”). Of these 13 papers, two are review articles that include a MEG/neuroHIV component, one is a case study, and the other 10 are empirical studies. Interestingly, of the 10 empirical papers, three were published in 2018 (30%) so the frequency of study appears to be increasing.
Before beginning, a few words about the interpretation and nature of MEG results are in order. First, with the exception of resting-state studies, MEG experiments almost always use event-related designs and are laid out very similarly to invasive electrophysiological investigations in monkeys and rats. Briefly, there is a baseline period with no stimulation, followed by the occurrence of an event or stimulus at time 0.0 seconds (s), which leads to an “active” period when the brain’s response to the stimulus occurs. This series of events constitutes a single trial, and the typical MEG experiment consists of 80–100 trials per condition, per participant.17 The MEG signals are recorded throughout the experiment and the individual trials are aggregated (offline) in each participant to increase the signal-to-noise ratio of the primary brain responses. In interpreting the signals, it is generally the relative response (active compared with baseline) that is of paramount interest, as neural cells spontaneously fire and cortical regions are constantly undergoing local electrical fluctuations that are seemingly unrelated to the sensory and cognitive processes of interest. Second, as mentioned above, there is no reference electrode in MEG. Thus, unlike other electrophysiological methods, the signal amplitude is not a difference measure between two electrodes or the average across multiple electrodes. The amplitude is specific to the location and not relative to electrical activity at any other location. Third, over the past decade, neurophysiological studies have increasingly focused on population-level neural oscillatory activity.25–26 In studies focusing on oscillatory analyses, rhythmic neural activity in discrete frequency bands, including delta (1–4 Hz), theta (4–7 Hz), alpha (8–14 Hz), beta (14–26 Hz), gamma (30–50 Hz), and high gamma (> 50 Hz) are examined separately to quantify rhythmic changes in the underlying neural population during cognitive processing.25 Note that MEG studies have generally focused on one or two frequency bands, based on their hypotheses, and that the precise definition of a band (e.g., alpha) is frequently determined statistically for a given study; for example, the definition of alpha may vary from 8–12 Hz to 8–14 Hz in different studies and this range is commonly determined using a data-driven approach. Probing such frequency bands separately enhances the signal-to-noise ratio of each, and enables identification of high-frequency (e.g., gamma range) neural responses that are often of lower amplitude. These high-frequency responses are widely thought to be critical for regional interactions across active brain networks,26 and thus characterizing them is of major importance for understanding overall brain function. Of note, oscillatory analyses largely rely on the same event-related experimental designs, except that each trial is generally longer to provide an adequately ‘silent’ baseline, with the relative responses again being of primary interest (i.e., active period compared with pre-stimulus baseline). As will become apparent from the studies discussed in this review, the application of oscillatory analysis methods to MEG signals and subsequent source reconstruction is a powerful approach for identifying the dynamics of brain function, and has facilitated major discoveries in several distinct areas.
Finally, a succinct overview of MEG terminology and metrics will promote understanding of the critical aspects of the studies discussed below, as some terms in the MEG field are somewhat unintuitive. Critically, the most common metric is the amplitude or power of the neural signal, and this can expressed for a particular voxel in the brain, a whole brain region, or at the MEG sensor-level. Pseudo-t is another common MEG unit; in functional maps using this metric, each pseudo-t value reflects the noise-normalized differential power between an active and a baseline period for a specific voxel. Other common units include the phase of the signal, based on a sensor- or voxel-level time series, as well as an array of functional connectivity measures that are typically computed using the voxel time series data. Such functional connectivity measures are thought to reflect inter-regional interactions, and common metrics include coherence, phase-locking factor, phase-lag index, and many others. Lastly, for oscillatory approaches, two common terms are event-related desynchronization (ERD) and event-related synchronization (ERS). An ERD refers to a decrease in band-limited power or amplitude (relative to a baseline) in response to a specific stimulus event or movement, whereas an ERS refers to an increase in band-limited power or amplitude (relative to baseline) in response to a specific event. ERD and ERS can be computed at the sensor- or voxel-level and are typically expressed in percentage units, reflecting the percent increase or decrease relative to the baseline period.
NeuroHIV in the cART Era
As mentioned above, the advent of combination antiretroviral therapy shifted the nature of HIV-infection from a terminal illness to a chronic manageable condition, but HIV-infected patients remain at a significantly elevated risk of developing cognitive impairments, with 35–70% of all patients exhibiting at least subtle deficits on tests of neuropsychological function.1–7 Numerous neuropsychological studies have documented aberrations across a broad range of functional domains (e.g., attention, fine motor), but the basic pathophysiology remains largely unresolved. Below, we review the MEG studies of neuroHIV that have been conducted to date. Each subsection focuses on studies that targeted a distinct cognitive construct and/or sensory modality, which we hope improves clarity and helps to illuminate the specific areas most in need of further research.
Resting State & Reliability studies
Consistent with the functional MRI (fMRI) literature, the most active area of MEG research in neuroHIV has been resting-state studies. Of course, resting-state studies have the advantage that they are easier to implement and that there is no chance of the imaging results being biased by differences in behavioral performance. Given these and other benefits, resting-state investigations have dominated the search for biomarkers across many different disease states, and this is certainly true for HAND and other neural alterations associated with neuroHIV.
One of the earliest studies in this field focused on the test-retest reliability of resting-state MEG signals in HIV-infected adults and matched healthy controls. All participants completed two five minute sessions (i.e., eyes-open and eyes-closed) of resting-state MEG, as well as neuropsychological testing, and then returned 24 weeks later for a follow-up MEG session that was identical to the first.28 For analysis, the authors divided the sensor-level MEG signals into eight sub-regions and five canonical frequency bands (e.g., left occipital alpha) and then focused on the power in each region. The study’s primary findings were that the MEG measurements were highly stable in both groups, as determined by the intra-class correlation (ICC) coefficient. In fact, the median stability was r = 0.80 and r = 0.72 during the eyes-closed and eyes-open conditions, respectively, with the highest values in the temporal lobes and the lowest in the frontal lobes.28 Spectrally, the gamma frequency range was the least reliable across both conditions, while the other canonical bands were far more reliable, although there was some variation by region.28 Regardless, median ICC values of 0.80 and 0.72 indicate that the measurements are highly reliable and bode well for the promise of MEG as a marker of disease. Beyond the reliability analysis, this study also showed that resting gamma power in the right occipital and right frontal region was stronger in uninfected controls relative to HIV-infected patients, and that individuals with high theta-to-gamma ratios tended to have lower cognitive test performance, regardless of serostatus.28 Overall, this study provided clear evidence that MEG is reliable, as well as the first indication that MEG signals may be capable of distinguishing adults with and without HIV-infection.
A later study by the same group went a step further and looked at functional connectivity during the resting state.29 Specifically, the authors collected five minutes of MEG data during eyes-closed rest and computed the sensor-by-sensor mutual information, which is a metric thought to reflect functional connectivity. Critically, the authors identified a group of MEG sensor pairs where the mutual information metric distinguished HIV-infected and uninfected participants.29 These sensor pairs were generally between left occipital and right frontal areas of the MEG helmet, and across these groupings the mutual information value was higher in the uninfected controls compared to the HIV-infected patients. Furthermore, the mean mutual information value averaged across all sensor pairs successfully discriminated the HIV-infected patients from the uninfected controls with only one exception, which suggests that measures of functional connectivity may have high sensitivity and specificity for identifying HIV-related neural alterations.29
Finally, a more recent MEG study focused on older HIV-infected participants and included a relatively larger sample than the previous resting-state investigations. In this study,30 older participants (half infected with HIV) underwent six-minutes of eyes-open rest during MEG, and the resulting data were projected into the brain and regional time series were extracted for 29 distinct brain regions.30 These time series were then subject to spectral analyses to quantify power in five canonical frequency bands (i.e., delta, theta, alpha, beta, and gamma). The key findings where that resting-state beta activity was significantly reduced in bilateral regions of the supplementary motor area, paracentral lobule, posterior cingulate, and the superior parietal lobule of older HIV-infected participants compared with demographically-matched uninfected controls. Further, the authors found that beta activity in the posterior cingulate, a key region in the default-mode network, was significantly correlated with scores on the Hopkins Verbal Learning Test-Revised, such that higher (better) scores were associated with stronger beta activity.30 Unlike previous studies,28 the authors did not find group differences in gamma activity or any other band besides beta. However, this disparity could reflect that the more recent study30 focused on older participants, and/or differences in the methodological approach adopted by the authors (e.g., sensor- vs. source-level analyses). Regardless, these studies have advanced understanding of resting-state neurophysiological alterations in neuroHIV, and illuminated key questions that remain unanswered; thus, future work is certainly needed.
Motor Control & Processing
While neuropsychological testing has documented abnormalities across a broad range of functional domains, deficits in fine motor control and reduced psychomotor speed have been some of the most common findings in HAND.9,31–34 In fact, this was partially reflected in the original nomenclature, which included diagnostic categories for HAD (3 subtypes) and a less severe condition termed minor cognitive motor disorder (MCMD).35 The MCMD category was phased out in 2007 with the adoption of the current classification system, which includes HAD, mild neurocognitive impairment, and asymptomatic cognitive impairment.10 Nonetheless, motor-related deficits remain a critical component of neuroHIV and are an essential feature of HAND assessment.36 Furthermore, despite the frequency of motor control and psychomotor speed impairments in neuroHIV, very few studies have attempted to identify the neural bases of such deficits, and in fact only one such MEG study of motor control has been published to date. This is problematic as non-motor issues like abnormal executive functioning can sharply decrease performance on measures of motor coordination and psychomotor speed, and ultimately masquerade as pure motor deficits in the absence of extensive neuropsychological testing.
The one existing neuroHIV motor study focused on the oscillatory dynamics of motor planning operations using a finger-tapping task during MEG. Briefly, recent neurophysiological studies of motor planning in humans have largely focused on an event-related desynchronization (ERD) in the 14–28 Hz beta range, which has been termed the peri-movement beta ERD.37–45 This response has been by far the most widely studied of the three main motor oscillatory responses (see below), and typically begins about 500 ms before movement onset and persists through the duration of the movement, with the strongest response peak in the motor hand knob region of the precentral gyrus contralateral to movement.43–45 Other regions that generate a peri-movement beta ERD, especially during complex movements, include the supplementary motor area (SMA), premotor cortices, parietal cortices, and cerebellum.37,39–40,43–46 Many independent studies have probed the functional role of the peri-movement beta ERD, and these studies have overwhelmingly supported the notion that this response is crucial to motor planning and movement selection. For example, the response is known to be modulated by the complexity of the movement to be executed,39 the certainty of the pending movement direction,47 the similarity between potential movement plans,48 and other cue-related factors.49 Beyond the beta ERD, there is a resynchronization in the beta band that follows movement termination, which has been termed the post-movement beta rebound (PMBR),37–38,42–45,50 as well as a high-frequency gamma response that coincides with movement onset and has been linked to the motor execution signal.44,51–56 However, neither of these responses have been examined in neuroHIV using MEG or any other modality, and thus we focus our background section on the beta ERD.
To investigate motor planning in neuroHIV, one study examined 24 participants (half with HIV – all virally suppressed) using MEG.57 All participants completed an externally-paced finger tapping task using their right index finger during MEG, and the resulting magnetic signals were decomposed into the time-frequency domain and imaged using a beamformer. The key results indicated a robust peri-movement beta ERD from −350 to 250 ms (movement onset = 0 ms) in the bilateral precentral gyri of each group, with the strongest response being centered on the motor hand knob feature of the contralateral (left) precentral gyrus (Figure 2).57 As shown in Figure 2, a strong beta ERD was also detected in the SMA of uninfected participants, but not seropositive participants. In regard to group differences, beta ERD responses were significantly stronger in controls relative to HIV-infected participants in the bilateral precentral gyri and the SMA (Figure 3). Conversely, HIV-infected participants had stronger responses in the right dorsolateral prefrontal cortex, medial prefrontal cortices, and a small area of the left inferior frontal gyrus. Interestingly, the beta response amplitude in the left precentral gyrus and SMA was significantly correlated with performance on the Grooved Pegboard test (dominant hand), while performance on the Trailmaking Test A and B was correlated with response amplitude in multiple prefrontal regions of HIV-infected participants.57
Figure 2. Beta Event-Related Desynchronizations (ERD) in Each Group.
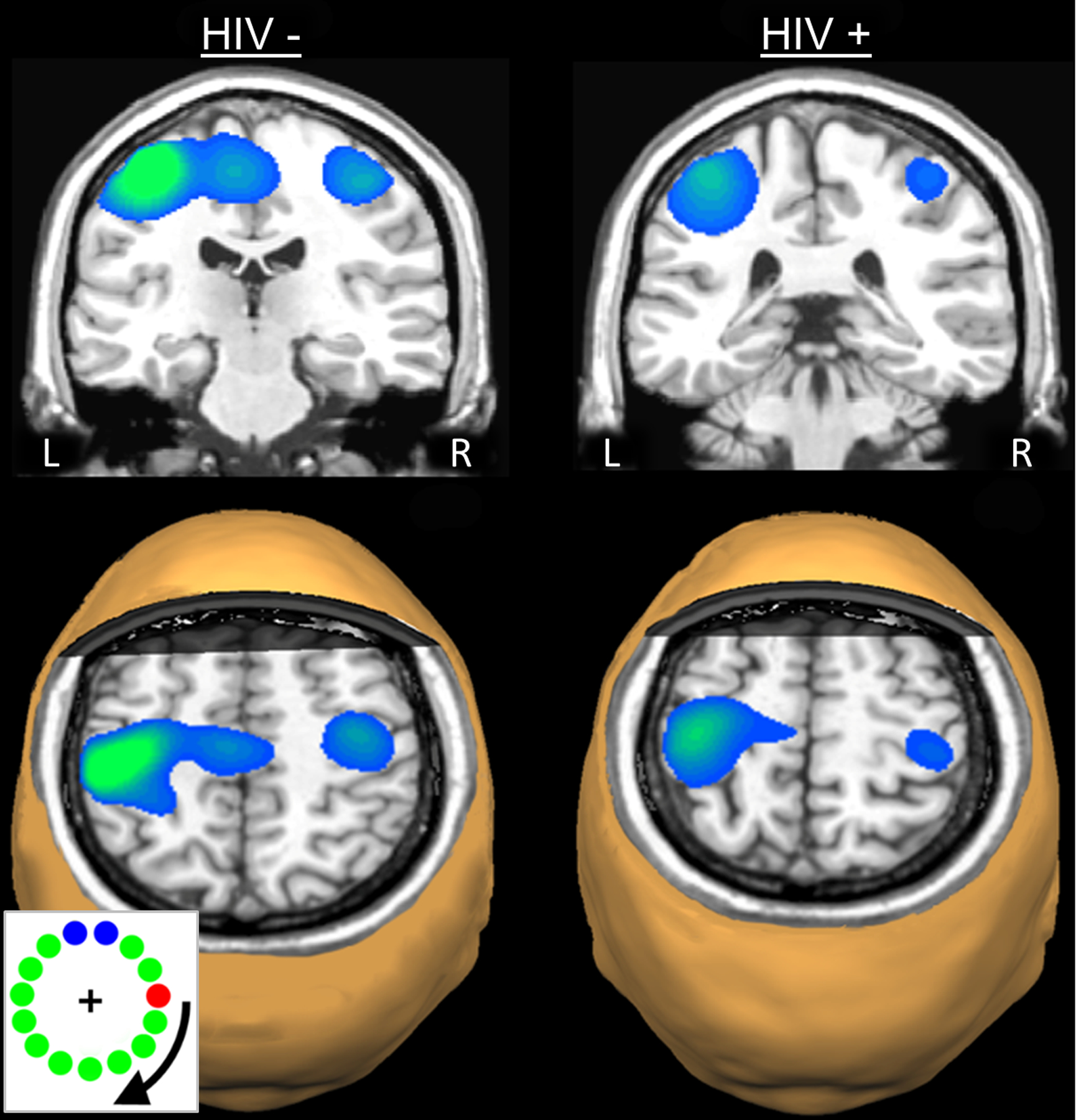
Strong beta ERD responses in the left precentral gyrus, extending posterior to include areas of the postcentral gyrus were discerned in the statistical parametric maps (SPMs) of each group. Strong ERD responses in the supplementary motor area (SMA) and the right precentral gyrus were also apparent in the uninfected controls. Images are shown in neurological convention (Left = Left) and have been thresholded at (p < 0.001, cluster-corrected). An image from the motor task is shown in the bottom left corner. Individuals made one movement when the red dot intersected area comprised of blue dots.
Figure 3. Group Differences in Beta ERD Activity.
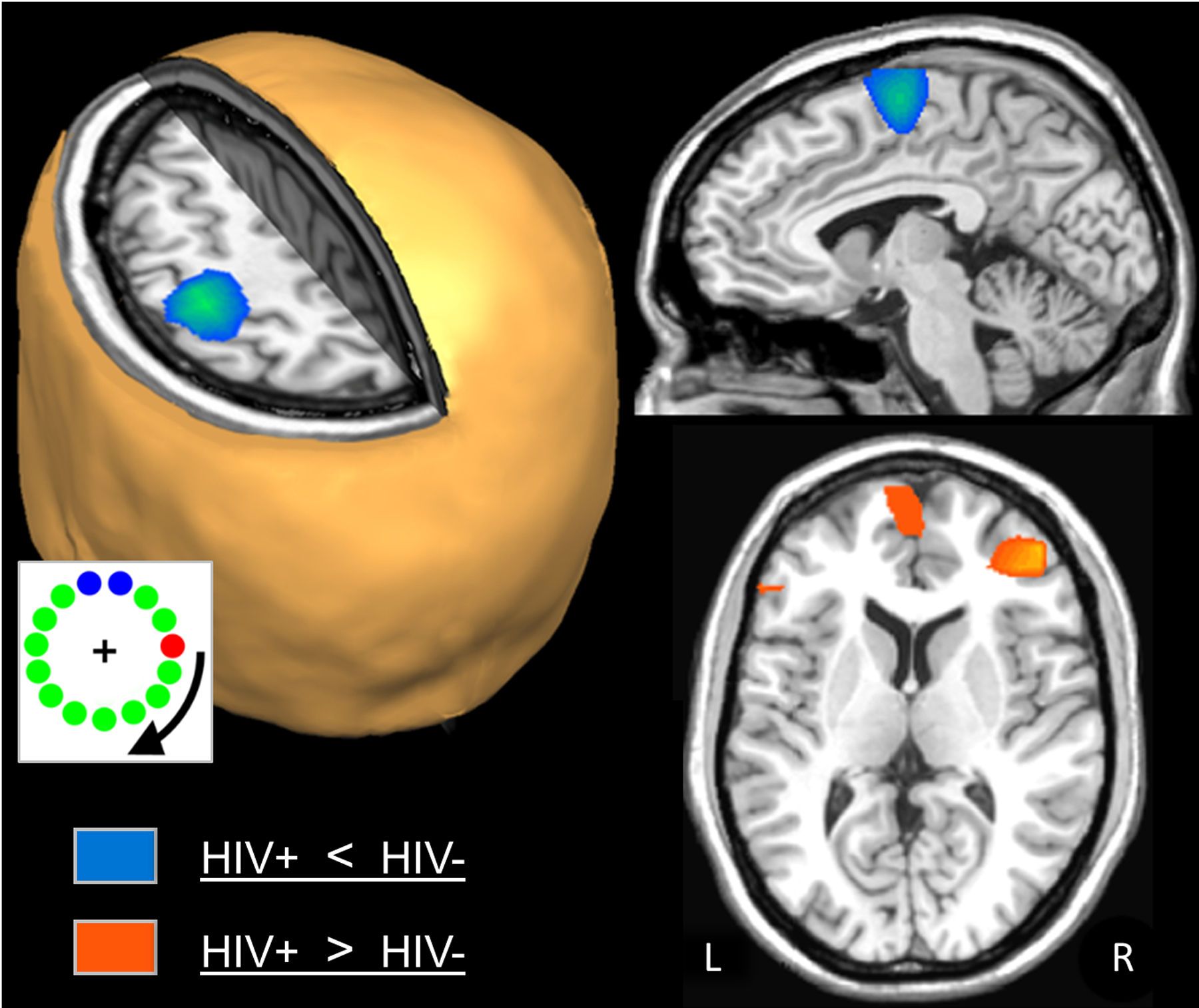
Uninfected adults exhibited significantly stronger beta ERD activity compared with HIV-infected adults in the left precentral gyrus, SMA, and a small area of the right precentral gyrus (blue areas; p < 0.01, cluster-corrected). In contrast, HIV-infected adults had significantly stronger beta ERD responses in the right (orange areas; p < 0.01, cluster-corrected). Again, an image of the task paradigm is shown in the lower left.
In summary, the one MEG study to date that has examined motor function found robust neural differences during planning operations across a distributed motor network. These differences included reduced beta responses in the primary motor cortices (i.e., hypoactivity) of HIV-infected participants, as well as stronger responses in prefrontal regions involved in executive function. These responses correlated with neuropsychological tests of motor function in the infected group, which supports the veracity of these neurophysiological findings. Clearly, many questions remain concerning neuroHIV and cortical motor function, and studies that consider the full gamut of oscillatory motor responses (e.g., post-movement beta rebound and motor gamma responses) are direly needed. Studies that use motor tasks capable of eliciting behavioral differences would be especially informative.
Somatosensory Processing
Deficits in motor control and processing generally need to be understood in the context of sensorimotor interactions and the degree of functioning within the somatosensory system. In other words, impaired somatosensory processing can have major negative consequences for motor control, and it is important to understand both systems to have a full grasp of motor function in any particular population. Thus far, there have been two MEG studies of somatosensory processing in neuroHIV. These studies were especially groundbreaking as there were no published fMRI or other functional imaging studies of somatosensation in HIV, although previous structural imaging studies had suggested widespread abnormalities in HIV-infected patients across the somatosensory strip and motor cortices.58 Thus, there was clearly circumstantial evidence for aberrations in the somatosensory system in neuroHIV.
In the first study, 17 virally-suppressed HIV-infected adults and 17 demographically-matched uninfected controls completed a mechanoreceptor stimulation paradigm using an air-puff paradigm during MEG, and underwent high-resolution structural MRI.59 The resulting MEG signals were transformed into the time-frequency domain, and time-frequency windows with significant responses (across all participants) relative to baseline were imaged using a beamformer. The whole-brain maps were then examined statistically for group differences and combined with structural gray matter maps derived from voxel-based morphometry using VBM8 and DARTEL implemented in the Statistical Parametric Mapping Software (SPM8; Wellcome Trust Centre for Neuroimaging, http://www.fil.ion.ucl.ac.uk).60 The key MEG findings were that all participants exhibited an increase (i.e., synchronization) in 4–8 Hz activity from 10 to 260 ms following stimulation onset, which mapped to the left postcentral gyrus directly posterior to the motor hand-knob feature. Interestingly, uninfected participants also exhibited a strong decrease in 8–14 Hz alpha activity during the same time period (Figure 4),59,61 which mapped to the left prefrontal cortex. In regard to group differences, the 4–8 Hz theta response was stronger in uninfected participants, which gave rise to a significant group difference in the left postcentral gyrus directly posterior to the hand-knob feature of the precentral gyrus. Likewise, the prefrontal alpha decrease was only observed in uninfected participants, which resulted in a relatively large group difference cluster centered on the left inferior frontal gyrus (Figure 4).59 The authors also extracted the peak group difference voxel from each of the significant clusters, and conducted a sensitivity/specificity analysis for both HIV-infection and cognitive impairment (9 of the 17 HIV-infected adults had HAND). This analysis revealed very high (>85%) sensitivity and specificity for identifying both HIV-infection and cognitive impairment based solely on these MEG metrics.59 Finally, the VBM results indicated reduced gray matter in HIV-infected adults across several brain regions, including the left parahippocampal and lingual gyri, left postcentral gyrus, right cerebellum, and several other regions. Of particular interest, the cluster of reduced gray matter in the left postcentral gyrus was anatomically coincident with the reduced MEG theta responses observed in the HIV-infected adults, which suggests that at least some functional differences in this population may be attributable to structural aberrations, although others (e.g., prefrontal alpha) appear to be more purely functional in nature.59
Figure 4. Significant Theta (4–8 Hz) and Alpha (8–14 Hz) Neuronal Activity Following Tactile Stimulation to the Right Hand.
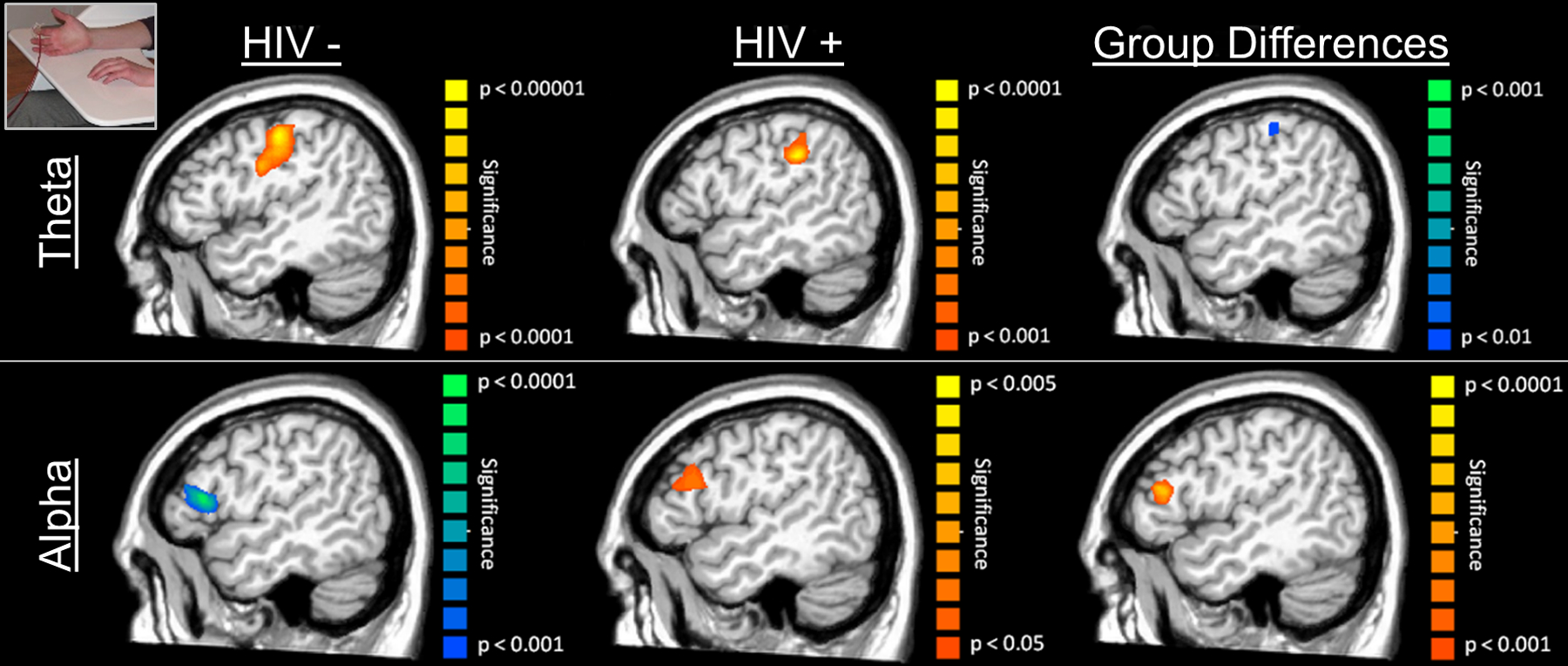
(Top): Uninfected controls (left) and HIV-infected adults (middle) exhibited theta responses in the left postcentral gyrus following mechanoreceptor stimulation, with the neural activity being stronger and more consistent in uninfected controls (note different P-value scales). The group effect (right; Controls > HIV-infected adults) was restricted to a small area of the left somatosensory cortex, and no brain areas had stronger 4–8 Hz activity in HIV-infected participants. Note the task design image in the top left corner. (Bottom): Uninfected controls exhibited a strong decrease in local alpha activity in the left prefrontal cortex (left) during the 0.01 – 0.26 s time window, whereas HIV-infected adults had a small increase in alpha activity in this same brain region (middle) and time window. Thus, the responses were in the opposite direction, which gave rise to a group effect of higher alpha activity in the HIV-infected participants (right panel). Again, note the different scales for P-values across images.
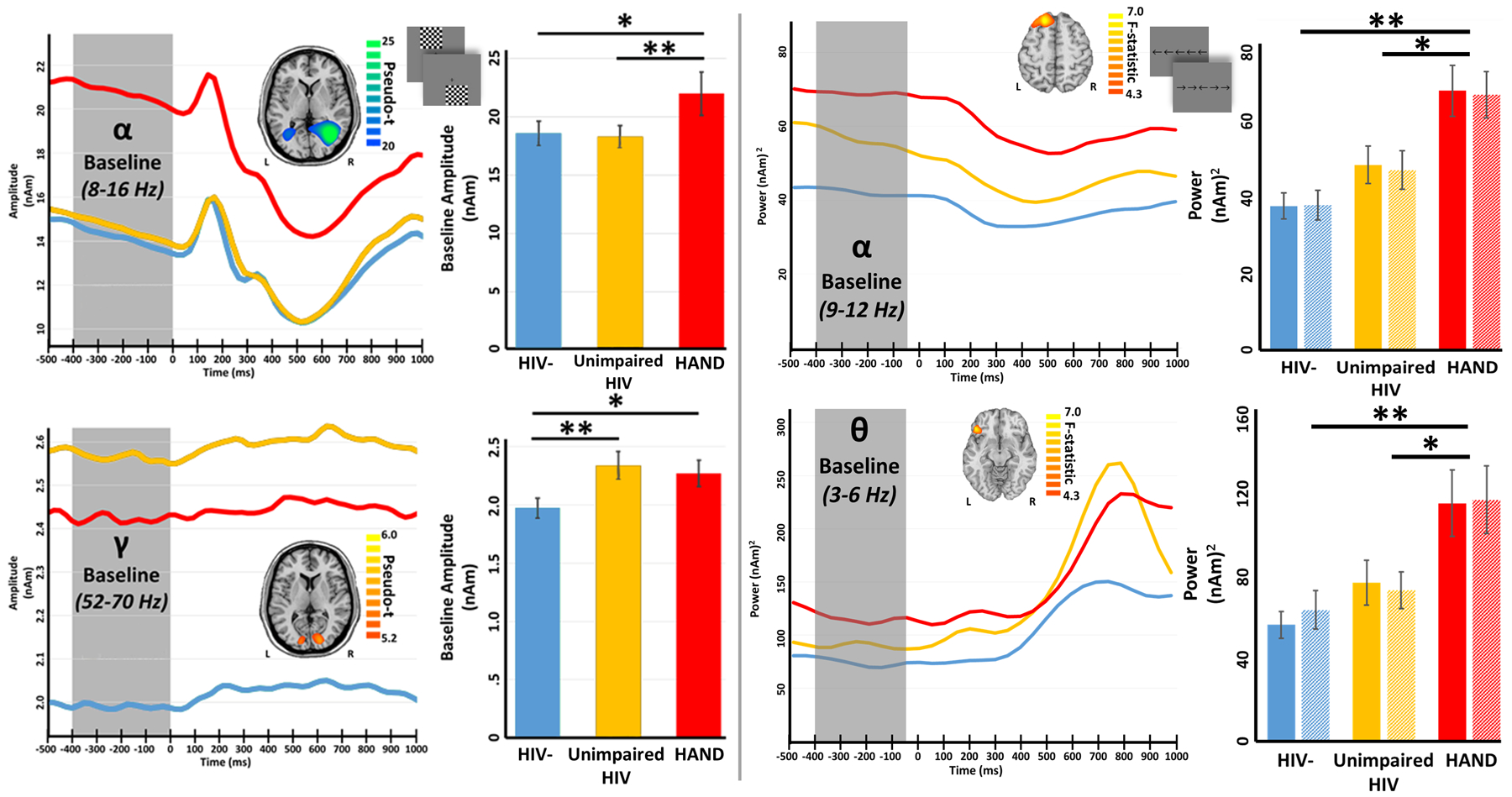
A larger and more recent study has also examined somatosensory processing in adults with HIV, although this study used paired-pulse electrical stimulation to elicit somatosensory cortical responses, which targets decidedly different experimental questions.62 Briefly, 71 participants (43 with HIV) completed a paired-pulse median nerve stimulation paradigm during MEG. The pulses were separated by 500 ms, which allowed the authors to look at sensory gating responses. Such gating is a neurophysiological phenomenon whereby the response to the second stimulus in a pair of identical stimuli is attenuated. This attenuation is thought to reflect the capacity of the CNS to filter less relevant information and preserve resources for behaviorally-relevant stimuli.63 Substantial evidence suggests that patients with cerebral palsy, schizophrenia, and even healthy older adults exhibit impaired gating of auditory and/or somatosensory stimuli, which is thought to reflect an impairment in bottom-up, pre-attentive inhibitory processing.63–66 Since attentive and/or top-down inhibitory processing is thought to be a critical deficit in neuroHIV (see later sections), such gating studies should provide key information in this population. In the study by Spooner and colleagues,62 the MEG signals were transformed into the time-frequency domain, revealing a robust response from 20–75 Hz in all participants during the 50 ms window following each stimulation (i.e., 0–50 ms and 500–550 ms). These responses were separately imaged in each participant and then averaged group-wise to reveal that the same region of primary somatosensory cortices generated the neural response in each group and to each stimulation. The authors then extracted the time series of the peak voxel in the grand averaged map for all subsequent analyses. The key findings indicated significantly reduced neural responses to both the first and second stimulation in HIV-infected participants relative to uninfected controls (Figure 5, top panel). In regard to gating, both groups exhibited significant sensory gating responses (i.e., response to the second stimulus was significantly weaker than the response to the first), but there was no group difference in the magnitude of the gating effect. The latter finding was surprising, as numerous studies63–64,66 have shown reduced gating in individuals with neurological and psychiatric disorders and such a reduction was hypothesized by the authors. Interestingly, follow-up analyses indicated that the peak gamma frequency of the second response relative to the first response was significantly higher in HIV-infected adults compared to controls (Figure 5, bottom left panel), and the authors argued that this alteration could be compensatory and underlie the preserved sensory gating function seen in these patients.62 Finally, since several recent studies have connected increased spontaneous activity during the baseline to aging,40,65 and given evidence of accelerated aging in HIV,67 the authors evaluated the power of spontaneous activity in the 20–75 Hz range in the same peak voxel as was used in their main analyses. This revealed that HIV-infected adults indeed had significantly elevated spontaneous activity relative to uninfected controls. Although future studies are needed to elucidate this effect, such elevated spontaneous gamma activity in HIV may underlie the reduced response amplitude observed in patients in the current study.68
Figure 5. Somatosensory Gamma (20–75 Hz) Activity Following Electrical Stimulation.
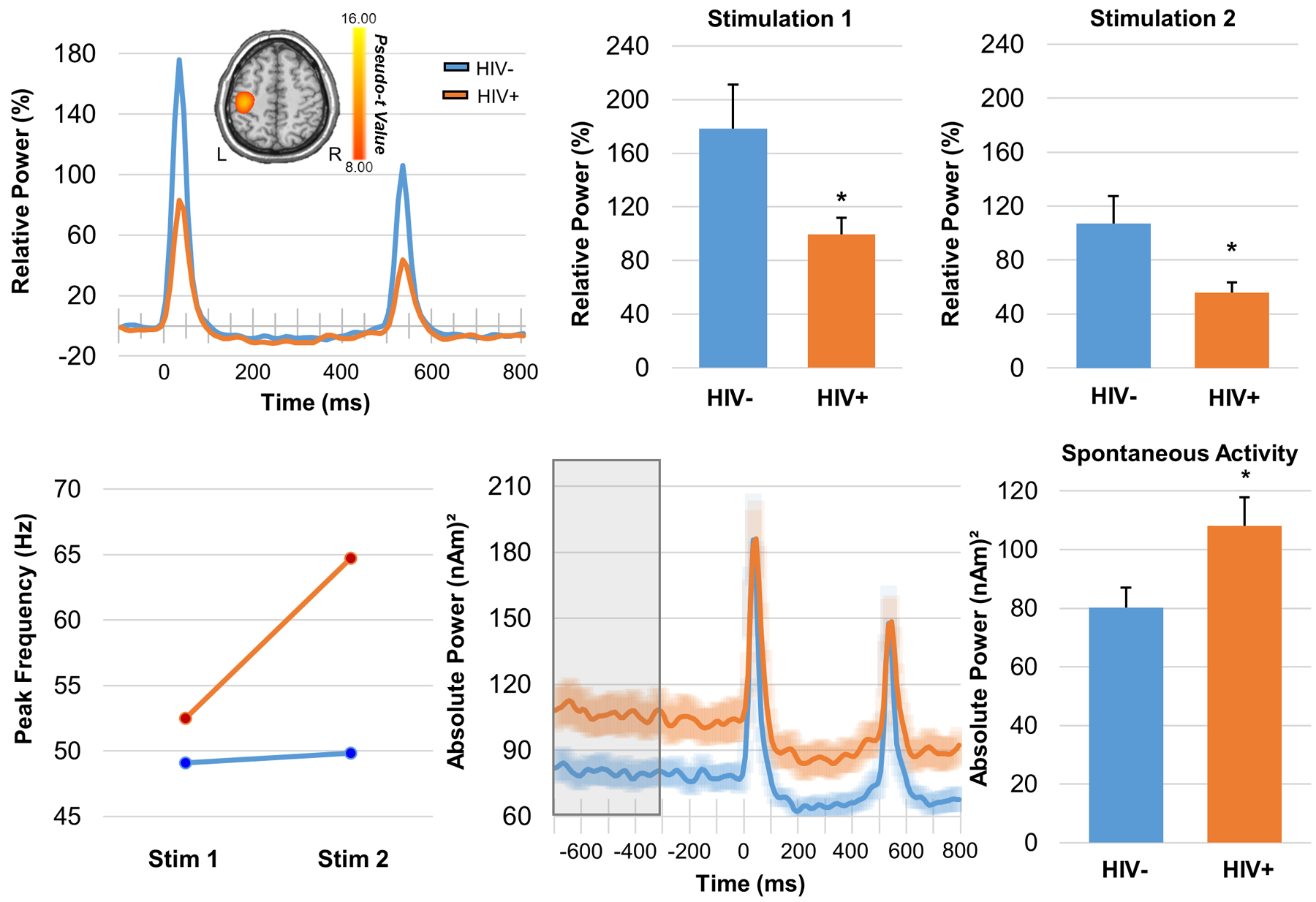
(Top): HIV-infected adults exhibited reduced response power to somatosensory stimulation. Disease-related power differences in response to each electrical stimulation were seen in the relative power envelope for the 20–75 Hz band (far left). Two-sample t-tests revealed reduced response power to stimulation 1 (middle) 2 (far right) in HIV-infected adults compared to controls (*p < 0.05). (Bottom): HIV-infection modulated the peak gamma frequency in the somatosensory cortex (far left); a mixed-model ANOVA revealed a significant stimulation-by-group interaction, which indicated that HIV-infected adults had a higher peak gamma frequency in response to the second stimulation relative to controls (p = 0.045). The time series extracted from the peak voxel and averaged across each group revealed elevated spontaneous neural activity immediately preceding stimulus onset in HIV-infected adults (middle). Two-sample t-tests revealed significantly elevated gamma activity during the baseline period (−700 to −300 ms) in adults with HIV relative to controls (*p < 0.05).
In summary, only two studies have examined somatosensory function in adults with HIV and both of these investigations focused on virally-suppressed populations and found widespread evidence for dysfunction in key somatosensory circuitry. These studies used different stimulation approaches and focused on different spectral windows (i.e., theta and gamma), but ultimately reached many of the same conclusions. The recent work showing preserved gating along with increased spontaneous activity and higher peak frequencies is especially worthy of follow-up, as these parameters could directly contribute to aberrant primary sensory responses. Finally, these findings have direct implications for understanding motor dysfunction in neuroHIV and future studies should evaluate these together within the same study, as abnormalities within the somatosensory system could impact motor function directly and lead to key impairments in daily activities.
Working Memory
Clinically-significant impairments in working memory are common in adults infected with HIV and are widely believed to contribute to other higher-level deficits in reasoning and decision making, which in-turn have a major impact on daily functioning and societal independence.9–10,69 These deficits are known to extend across both verbal and spatial domains, but their precise cognitive and neurological origins remain largely unknown. Several neuropsychological studies have suggested that impairments in working memory arise from encoding deficiencies, whereas others have indicated that central executive processes are likely the origin.70 Most functional neuroimaging studies have used fMRI and tended to support both views, but their conclusions remain at least partially tentative as aberrations in encoding stimuli would affect the maintenance of such memory representations, and these studies have not been able to temporally distinguish encoding and maintenance periods.69–72 More broadly, previous functional imaging studies in HIV-infected adults have often reported abnormalities in both primary sensory and higher-order frontal and prefrontal cortices, which again conflates the encoding and maintenance operations serving working memory function.71–76 Given these data, the precise timing of MEG could be especially valuable in identifying the cognitive (i.e., encoding vs. maintenance) and neural origins of working memory deficits in HIV, and to date only one such study has directly addressed these questions in the verbal domain of working memory.77
In this study, 36 older adults (half infected with HIV, all virally-suppressed) completed a verbal working memory task during MEG. Both groups were demographically-matched in regard to age, sex, ethnicity, and handedness, with the mean age across both groups being about 58 years at the time of scan.77 The task was specifically designed to distinguish encoding and maintenance operations. Briefly, participants were instructed to memorize six letters that appeared in a grid (encoding). After 2.0 s these letters disappeared from the grid (maintenance period) and 3.0 s later a probe letter appeared in the grid (retrieval period), with the participants being instructed to respond “yes” or “no” via button press as to whether the probe letter was included in the original six-letter encoding grid (Figure 6). This task had been validated by the investigators in many previous studies focuses on normative populations and those with other neurological and psychiatric diseases.78–85 The key findings were that the uninfected adults performed significantly better on the task (~82.5% accurate) than did the adults with HIV (~76% accurate). In regard to the MEG data, all participants exhibited a robust decrease in 9–16 Hz alpha/low-beta activity during the encoding period in left occipito-temporal sensors, as well as a robust increase in 9–12 Hz alpha activity in occipital cortices during maintenance.77 To identify the time course of these responses, the spectral windows were imaged using a beamformer in 400 ms windows starting 200 ms after the onset of the encoding grid and extending until the onset of the probe letter (5000 ms). The results indicated a strong decrease in alpha/low-beta activity that began in the posterior occipital cortices and spread anteriorly to include the left inferior and superior temporal cortices, left supramarginal gyrus, and left inferior frontal cortices (Figure 6). These dynamics appeared to be similar in HIV-infected and uninfected participants, especially during the encoding period as there were no group differences observed throughout this period.77 In contrast, during memory maintenance, HIV-infected adults exhibited significantly stronger responses (i.e., alpha decreases) in the left supramarginal gyrus during the earliest time period, and this was sustained through the majority of the maintenance period (Figure 7). Similar sustained group differences were also observed in the left inferior frontal gyrus and left cerebellum. In contrast, the opposite pattern was observed in the homologue right inferior frontal gyrus, as uninfected participants exhibited stronger alpha decreases relative to HIV-infected adults (Figure 7). Controls also exhibited a significantly stronger alpha synchronization (i.e., increase) in the occipital cortices throughout the majority of the maintenance period and the amplitude of this response was significantly correlated with accuracy on the working memory task in controls, but not HIV-infected adults. Lastly, this study also found that the strength of the alpha decreases in the left supramarginal gyrus during maintenance were significantly correlated with performance on the working memory task, as well as global neuropsychological function and the attention and memory composite scores in HIV-infected patients.77
Figure 6. Oscillatory Activity during Verbal Working Memory.
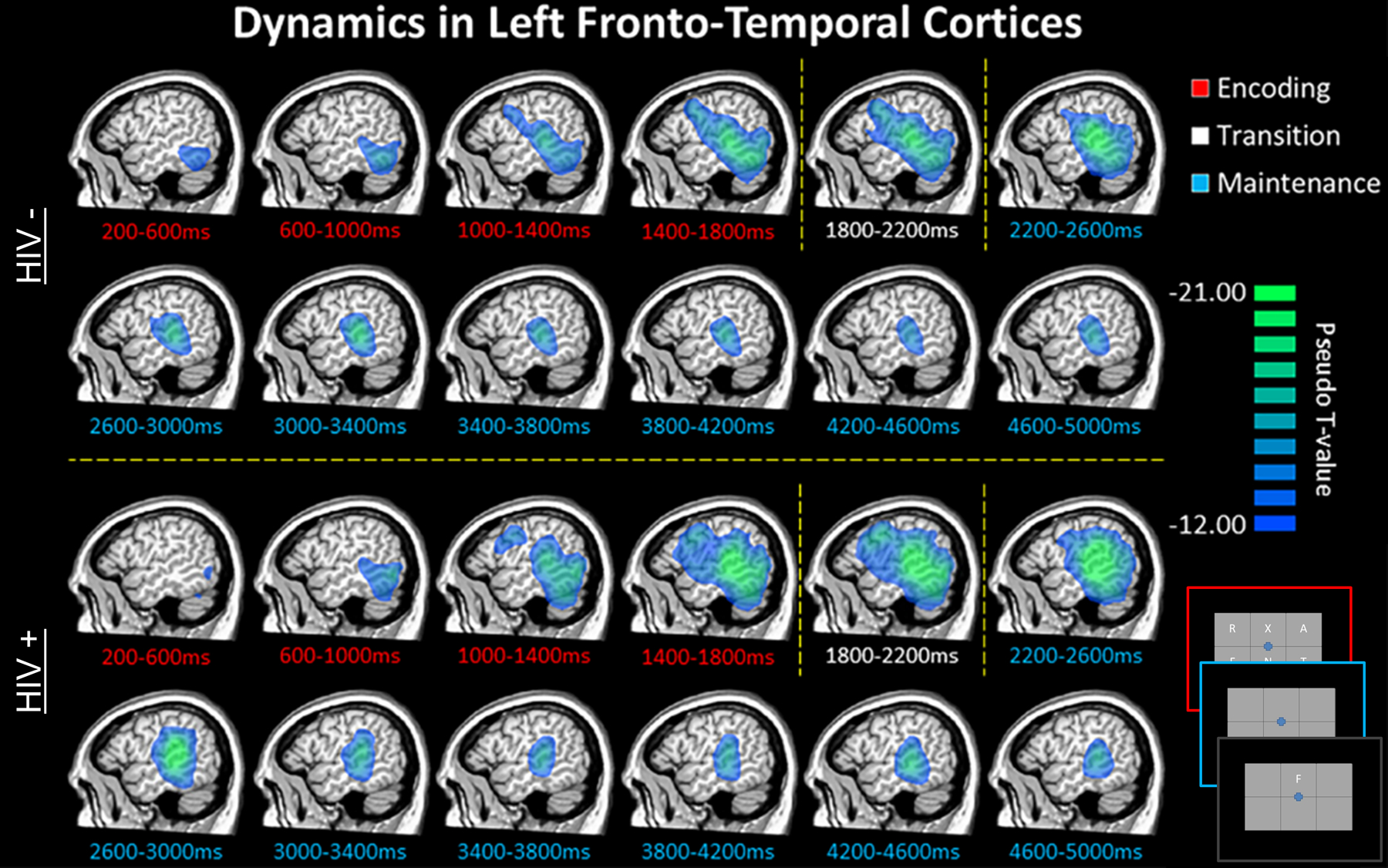
Group mean beamformer images for uninfected older adults (top) and demographically-matched HIV-infected adults (bottom) are displayed across all time bins spanning the encoding (red labels), transition (white labels), and maintenance (blue labels) phases. An image of the task design is shown in the bottom right corner. Across both groups, there was a sustained decrease in alpha/beta oscillatory activity in left fronto-temporal cortices throughout most of the encoding and maintenance periods. This decrease began in posterior regions early in the encoding phase, and then spread anterior and superior to include language-related cortices in the left supramarginal and prefrontal areas during the latter half of encoding and early maintenance period. These responses slowly dissipated throughout the remainder of the maintenance phase.
Figure 7. Group Oscillatory Differences during Working Memory Maintenance.
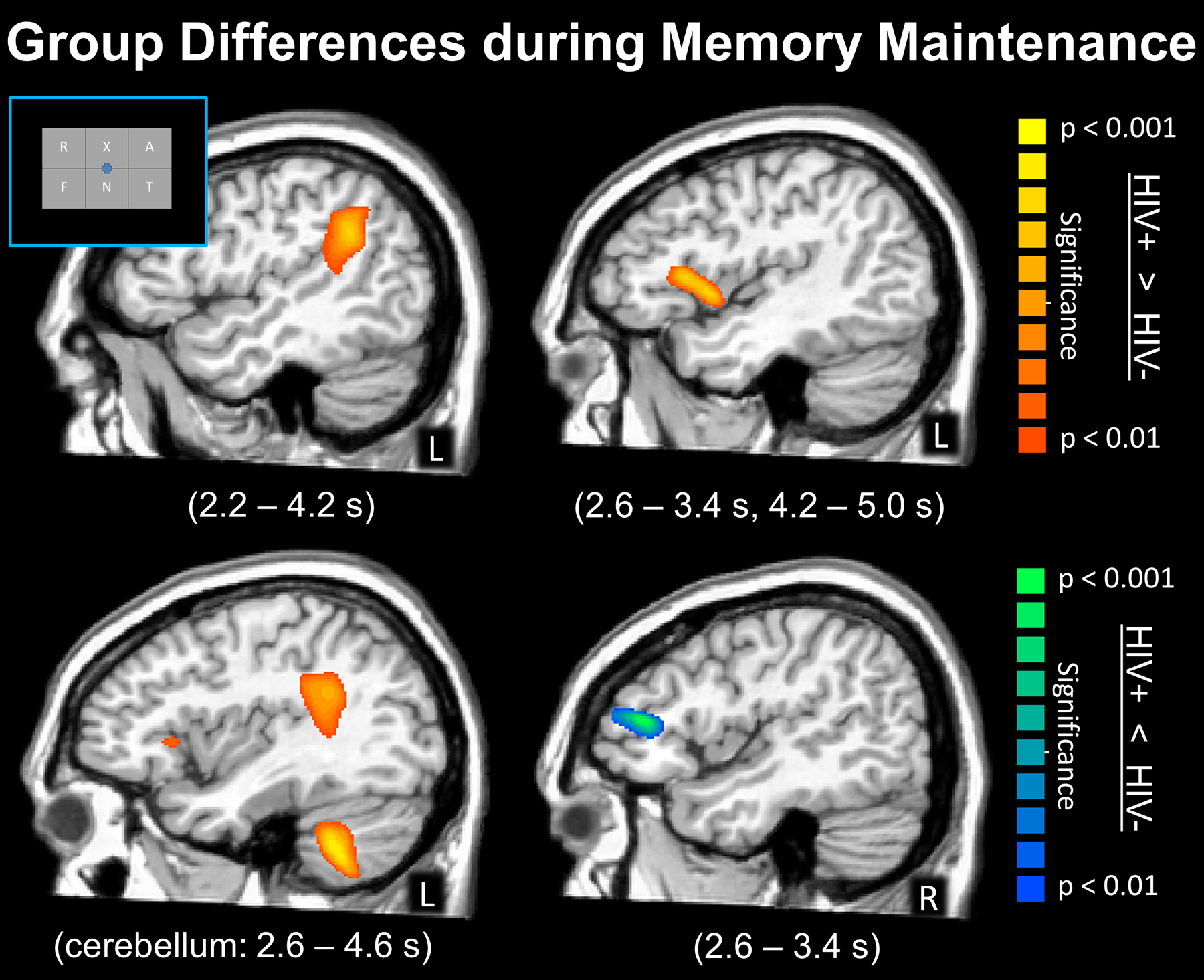
Statistical parametric maps showing group differences during the maintenance period, with the time windows containing significant effects listed in the parentheses under each image. (Top Left) HIV-infected adults had significantly stronger alpha decreases (hyper-activation) in the left supramarginal cortices from 2.2 to 4.2 s after the onset of the encoding grid, which corresponded to the majority of the maintenance period. (Top Right) Stronger alpha decreases (hyper-activation) in HIV-infected adults were also observed in the left inferior frontal gyrus from 2.6 to 3.4 s and from 4.2 to 5.0 s. (Bottom Left) Stronger alpha decreases in HIV-infected adults were also observed in the left cerebellum from 2.6 to 4.6 s; the group differences in the supramarginal and left inferior frontal gyrus can also be seen in this image. (Bottom Right) In contrast, uninfected adults exhibited stronger alpha decreases in the right IFG compared to HIV-infected adults, which is consistent with healthy aging studies showing recruitment of homologue cortices in the right hemisphere to aid in ongoing processing. All maps have been thresholded at (p < 0.01, corrected).
Given the widespread consequences of working memory dysfunction for other aspects of cognitive processing, as well as personal independence and quality of life, it is surprising that so few studies have examined this domain in HIV-infected adults. Beyond the limited number of fMRI studies, only one MEG study has been conducted to date. As mentioned above, this study clearly indicated that the impairments primarily reside in memory maintenance and not stimulus encoding, as there were no group differences during the 2.0 s encoding time window. Furthermore, during maintenance HIV-infected adults exhibited hyper-activations in the well-established left hemispheric verbal working memory networks, and did not widely utilize right hemispheric homologue regions to the same degree as age matched controls. The latter finding is particularly important, as studies have shown that healthy older adults more broadly utilize homologue regions during cognitively-demanding working memory paradigms relative to their younger peers.83 In addition, this study showed that the well-known occipital alpha synchronization during maintenance is abnormal in HIV-infected adults, and this almost certainly contributed to their poor performance on the working memory task.78–85 Obviously, much remains to be discovered in this area and future studies should focus on both spatial and verbal working memory to determine if both are affected equally.86–87 Likewise, therapeutic studies targeting memory maintenance operations could be helpful in reducing the burden of working memory deficits in this population.
Visual-Spatial Processing & Visual Attention
Visual processing and visual attention has been one of the most frequent areas of investigation in functional neuroimaging studies of neuroHIV. Early fMRI studies reported aberrations in visual-perceptual and visual attention networks in HIV-infected adults, and these findings have generally been confirmed and expanded in more recent work. For example, Chang and colleagues found that HIV-infected patients exhibit markedly increased fMRI activation in lateral occipital, posterior parietal, and prefrontal regions during attention tasks.88–89 More recent studies have focused more exclusively on visual cortex and found that HIV-infected adults have reduced activation in primary visual cortex during a visual stimulation task relative to healthy controls,76 as well as reduced cerebral-blood-flow to the same visual regions during the resting-state.75 Corruption of parietal- and fronto-visual functional connectivity in the resting state has also been reported in HIV-infected adults, and these fronto-visual attention networks align particularly closely with those that are compromised in advanced aging.90 In regard to MEG studies, there have been a total of three to date, with the earliest focused on basic visual processing,91 a second focused on visuo-spatial processing and attention,92 and a third that examined visual selective attention.93 We will review each of these studies in turn.
The first study included 23 participants (12 HIV-infected) who were group matched on major demographic factors. All participants completed a basic visual processing task during MEG, which involved fixating on a crosshair while a small checkboard box appeared in the top right visual field quadrant.91 All MEG data were transformed into the time-frequency domain, which revealed strong oscillations from 6–12 Hz during early stimulus processing (i.e., 50–225 ms, with 0 ms being stimulus onset). Source imaging of this time window revealed activity in the right dorsolateral prefrontal cortex, the posterior calcarine cortex, the posterior cingulate, and the frontal eye fields. In regard to group differences, the key findings were that controls showed greater 6–12 Hz power in the right frontal eye fields, right dorsolateral prefrontal cortex, and the posterior cingulate compared with HIV-infected adults.91 Interestingly, the authors also found that the amplitude of responses in both the right dorsolateral prefrontal cortices and right frontal eye fields of HIV-infected patients was positively correlated with the retention index of the Hopkins Verbal Learning Test Revised (HVLT-R), and this effect survived stringent Bonferroni correction. In other words, the more “control-like” the responses in these regions, the higher the retention index score was in the HIV-infected group, which supports the notion that these oscillatory differences are integral to cognitive dysfunction.91
While this early study provided significant insight, later MEG studies of visual-spatial processing and selective attention have added a critical new dimension. These studies have focused on the key question of whether MEG measures of visual attention/processing can be used to distinguish HIV-infected adults with HAND from those who are cognitively unimpaired. One study focused on neural oscillatory activity in visual occipital cortices and examined 67 participants (43 HIV-infected) who were divided into three demographically-matched groups, including uninfected controls, unimpaired HIV-infected adults, and adults with HAND.92 All participants underwent MEG during a visual-spatial processing task that required participants to respond (i.e., button press) regarding the spatial location of the visual stimulus. This task is known to elicit robust, multispectral oscillatory responses.94–96 The resulting MEG data were probed for significant oscillatory visual responses and, in agreement with previous studies using this task,94–96 the authors observed strong theta, alpha, and gamma responses in MEG sensors near occipital cortex during early stimulus processing. These three visual responses were imaged in each participant using a beamformer and the resulting images were averaged across groups to identify the peak voxels for each response. The time series for each of these peaks was then extracted for each person and used in the group-wise statistical analyses. The behavioral results indicated that uninfected controls responded significantly faster than both the unimpaired HIV-infected and HAND groups, and that the group with HAND also responded significantly slower than the unimpaired HIV-infected group (Figure 8, top panel).92 Adults with HAND were also significantly less accurate on the task compared with the other two groups. As per the MEG data, the key findings were that visual theta responses were significantly stronger in the uninfected group relative to both unimpaired HIV-infected and HAND groups, and that the relationship between accuracy on the task and the strength of occipital theta activity significantly differed between the groups (Figure 9). In regard to the alpha and gamma responses, no group differences were observed in regard to the strength of each oscillatory response. However, given recent findings in the aging and HIV literatures,40,62,65 the authors also examined the power of spontaneous neural activity in each of these spectral windows and their results were groundbreaking. Essentially, spontaneous alpha activity in visual occipital cortices was sharply elevated in the group with HAND, but not the control or the unimpaired HIV-infected group (Figure 10, top left panel). In contrast, spontaneous occipital gamma activity was significantly elevated in both unimpaired HIV-infected adults and those with HAND relative to uninfected controls (Figure 10, bottom left panel).92 In other words, based on the power of spontaneous activity during the pre-stimulus baseline, one could distinguish each of the three groups from each other. These findings were of major importance as they suggested that specific MEG indices may have the capacity to serve as diagnostic markers of cognitive impairment in neuroHIV, which would have a major impact as HAND diagnoses are currently based on extensive neuropsychological testing and the exclusion of opportunistic infections and other confounding factors. Further, the overall pattern of increased spontaneous activity is consistent with the notion of accelerated aging in HIV-infected adults,67 as multiple studies in healthy controls have supported the view that spontaneous cortical activity increases with age.40,65 In sum, this MEG study provided critical new data and was the first to suggest that MEG may have the capacity to discern those HIV-infected adults with HAND from those who are cognitively unimpaired.
Figure 8. Behavioral Results from Visuo-Spatial Processing and Selective Attention Tasks.
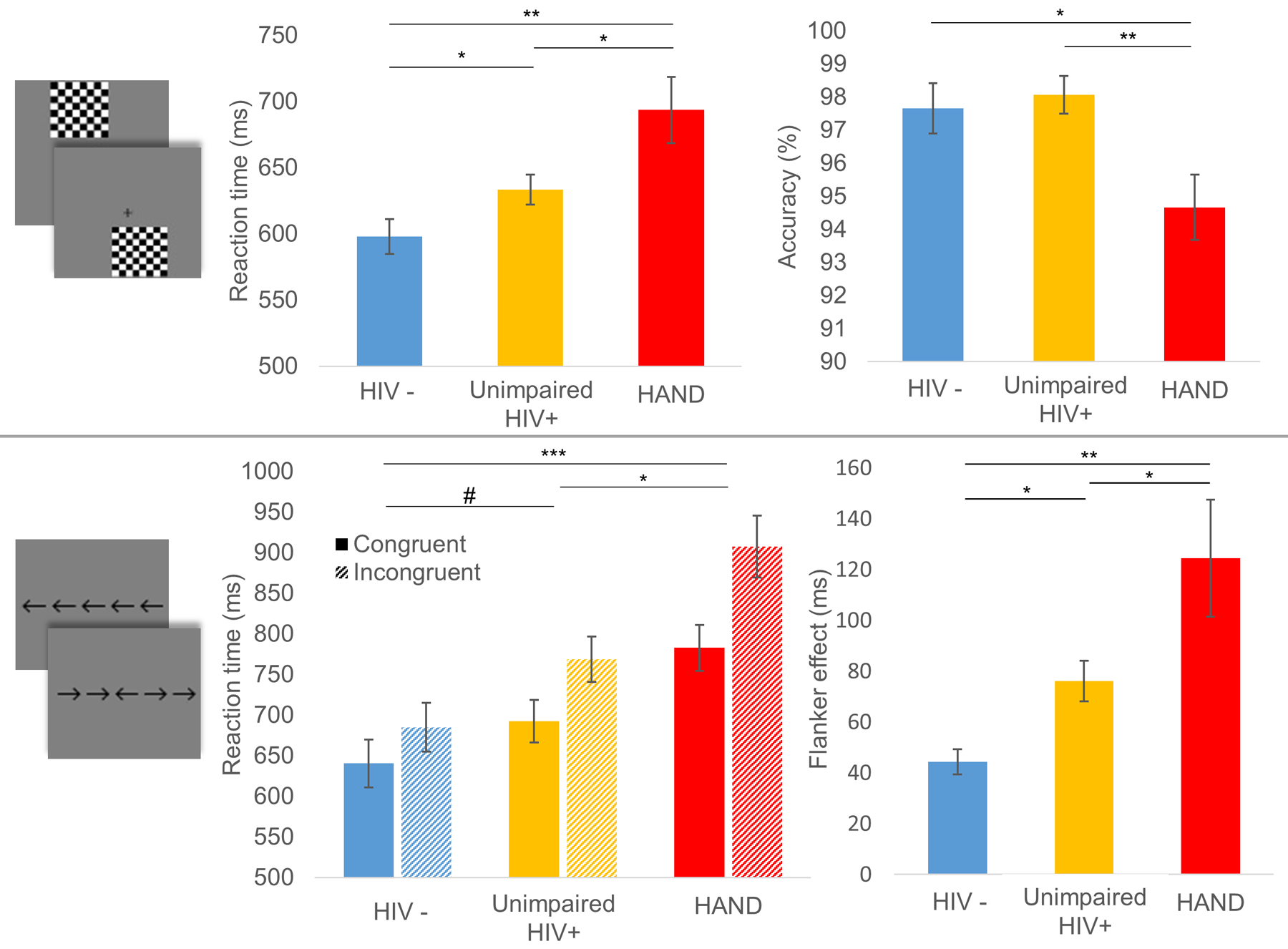
(Top): Images depicting the task design are shown to the far left, followed by reaction time (middle) and accuracy (right) results from the visuo-spatial task. In the bar graphs, participant group denoted on the x-axes. Reaction time (in milliseconds) is displayed on the y-axis of the graph in the middle, and accuracy (in % correct) is displayed on the y-axis of the graph to the right. Results indicated a clear stair-step effect whereby controls were significantly faster than unimpaired HIV-infected adults who were significantly faster than adults with HAND. As per accuracy, both controls and unimpaired HIV-infected adults were significantly more accurate than adults with HAND. (Bottom): As with the top, images depicting the task design are shown to the far left, with reaction time (middle) on the selective attention task in the middle. Data showing the flanker effect (Reaction time to Incongruent – Congruent trials) is shown to the far right. The results indicated that adults with HAND responded slower on both types of trials relative to both unimpaired HIV-infected adults and uninfected controls (middle). As per the flanker effect, a stair-step pattern was observed whereby controls were faster than both groups and unimpaired HIV-infected were faster than those with HAND (#p < .10, *p < .05, **p < .01).
Figure 9. Visual theta responses are reduced in HIV-infection and HAND.
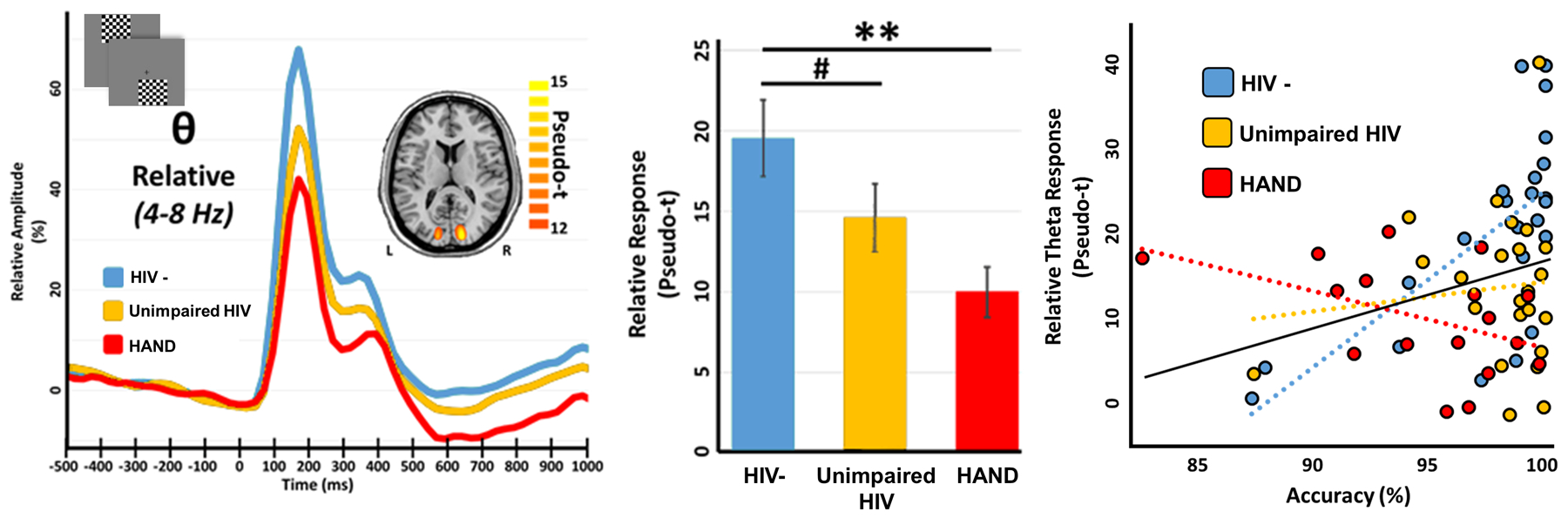
(Left): Peak voxel time series for each group; time (in millisecond) is denoted on the x-axis, with 0 ms defined as the onset of the visual stimulus, and relative amplitude (in % of baseline) denoted on the y-axis. Example task images are shown in the top left corner and the inlaid source map shows the peak-voxel locations used for pseudo-t extraction, along with its respective color scale legend. (Middle): Bar graph reveals extracted pseudo-t values averaged across participants per group, with significance levels of post-hoc statistical analyses indicated by asterisks above the bars (#p < .10, *p < .05, **p < .01). (Right): Group-wise relationships between oscillatory theta responses and task accuracy. Theta oscillatory activity (in pseudo-t) is shown on the y-axis and task accuracy (in % correct) on the x-axis. The three dashed lines of best-fit represent the group-wise relationships between these variables, with the color legend inlaid above. The solid black overlaid line of best-fit represents this relationship across all participants.
Figure 10. Spontaneous Neural Activity is Increased in HIV-infection and HAND.
(Left): Peak voxel time series during a visuo-spatial processing task is separated by group for alpha (top) and gamma (bottom) on the far left. Time (in milliseconds) is denoted on the x-axes, with 0 ms defined as the onset of the visual stimulus, and absolute amplitude (in nAm) denoted on the y-axes. The inlaid maps show the peak-voxel location where data were extracted, along with its respective color scale legend. The bar graphs to the right represent pre-stimulus amplitude values averaged across participants per group, with significance levels of post-hoc statistical analyses indicated by asterisks above the bars (*p < .05, **p < .01). Spontaneous alpha distinguished the HAND group from the other two groups, while spontaneous gamma during the baseline distinguished controls from the other two groups. (Right): Peak voxel time series during a visual selective attention task is separated by group for alpha (top) and theta (bottom) activity. All axes are the same as above, except that the y-axes show power (nAm2) instead of amplitude (nAm). As shown, alpha activity in the left superior frontal region differed between the HAND group and the other two groups, and the same was observed for theta activity in the left inferior frontal. Of note, spontaneous alpha in multiple regions distinguished all three groups (not shown).
The third MEG investigation in this area followed a similar model to the previous study in that there were three groups of participants (HAND, unimpaired HIV-infected, and uninfected controls), but this study focused on visual selective attention function and the impact of cognitive interference.93 The study included 20 participants per group and the three groups were closely matched on major demographic factors, including age, sex, ethnicity, education, and handedness. Again, all HIV-infected participants were virally-suppressed. Participants completed a visual flanker task during MEG, which required them to respond as to the direction (left or right) of a middle arrow that was presented alongside flanking arrows that were either pointing in the same direction (Congruent) or the opposite direction (Incongruent; see bottom panel of Figure 8). The classic finding in this task is that participants respond slower in the incongruent relative to the congruent condition due to the interference caused by the flanking arrows, and this difference in reaction time has been termed the flanker effect. Thus, the authors examined this behavioral effect as well as the associated neural oscillations using MEG. Their main behavioral findings were that adults with HAND were significantly slower than both unimpaired HIV-infected and control adults across both congruent and incongruent conditions, while the latter two groups marginally differed with controls being the fastest responders (Figure 8, bottom panel).93 In addition, all participants exhibited a significant flanker effect (i.e., responded slower in the incongruent condition) and the magnitude of this effect differed in a stepwise fashion across the three groups, being smallest in the uninfected controls and largest in the adults with HAND. As per the MEG signals, these data were decomposed into the time-frequency domain, which revealed strong responses in both the theta (3–6 Hz, 50–400 ms) and alpha (9–12 Hz, 250–650 ms) bands, in agreement with previous MEG studies using the flanker task.97–98 The authors then imaged these responses separately for the congruent and incongruent condition, and subtracted the resulting congruent maps from the incongruent maps to derive a whole brain map of so-called flanker interference activity. An ANOVA of these maps revealed group differences in theta inference activity in the left inferior frontal cortices, left dorsolateral prefrontal cortices, left temporoparietal junction, and left inferior parietal regions (Figure 11).93 Likewise, group differences in alpha interference activity were found in the left superior frontal gyrus, middle frontal gyrus, supramarginal gyrus, and left angular gyrus, as well as the right dorsolateral prefrontal cortices (Figure 11). Importantly, the authors found that left inferior frontal theta activity was a significant positive predictor of executive composite scores, while the left dorsolateral prefrontal cortex theta response was a significant positive predictor of attention composite scores (Figure 12). Further, group-wise post-hoc difference testing of these regions found that theta interference activity in both areas was significantly reduced in the group with HAND relative to the other two groups, and did not differ between controls and unimpaired HIV-infected adults.93 Finally, the authors also examined whether there were group differences in spontaneous activity in each of the brain regions identified in the whole-brain theta and alpha ANOVAs. These analyses revealed that adults with HAND had increased spontaneous theta activity relative to infected controls in most of the regions identified in the whole-brain theta ANOVA, as well as increased spontaneous activity relative to unimpaired HIV-infected participants in the left inferior frontal and left temporoparietal junction regions (Figure 10, right panel). In contrast, alpha activity was significantly elevated in the HAND group relative to both the unimpaired HIV-infected and uninfected controls groups in almost all regions identified in the whole-brain alpha ANOVA (Figure 10, right panel).93 Thus, in this study, the spontaneous alpha results alone (in several distinct brain regions) were enough to distinguish the three groups, and importantly made a clear distinction between the unimpaired HIV-infected adults and those with HAND. Furthermore, the behavioral task was very sensitive to HIV-related changes in cognitive performance, as the flanker effect revealed a unique stepwise pattern of decline in visual selective attention function among the HIV-infected patients with and without cognitive impairment.
Figure 11. Group Differences in Theta and Alpha Activity Related to Visual Selective Attention.
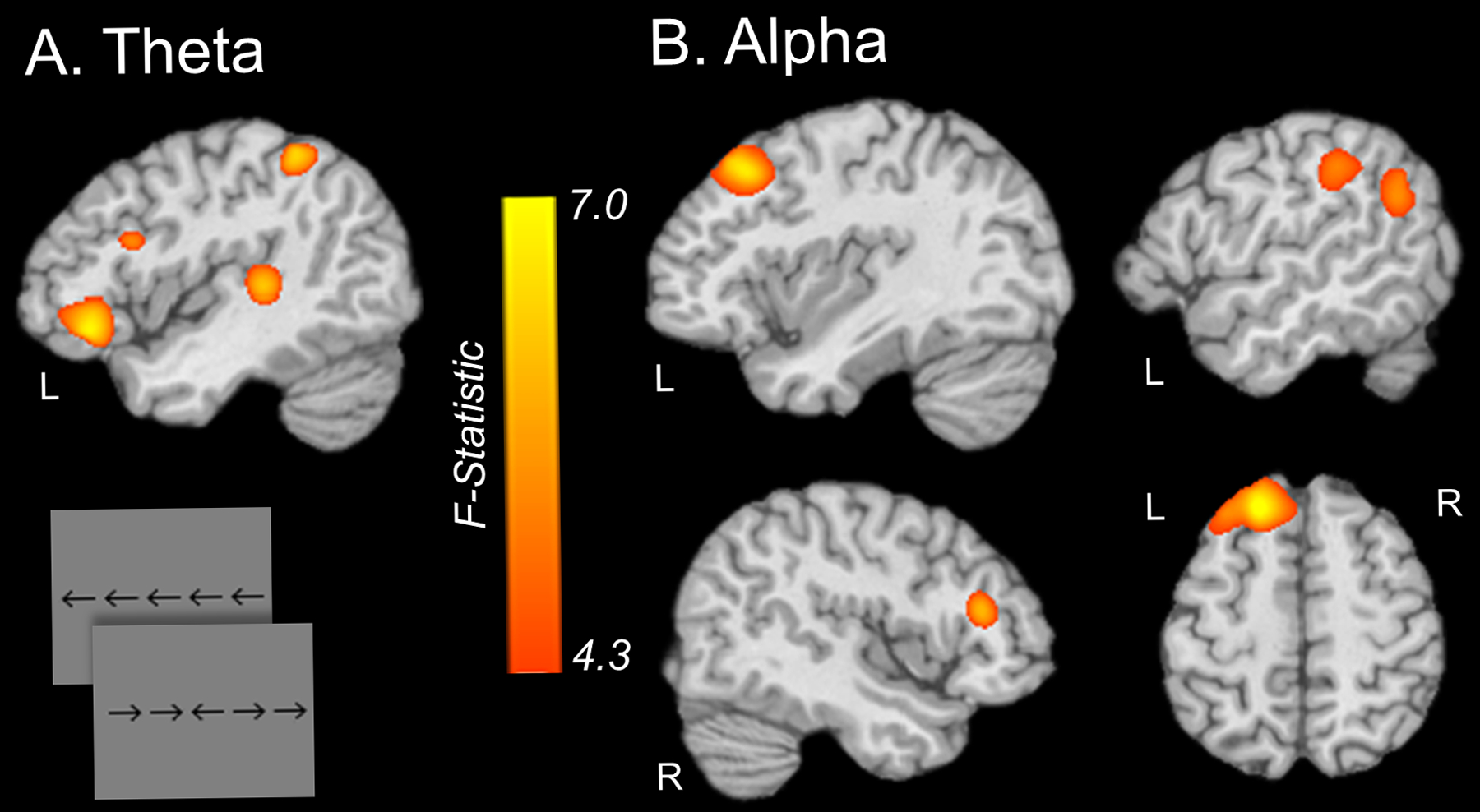
(A) Whole-brain ANOVA of flanker interference activity (maps of Incongruent – Congruent trials) in the theta range showed significant group differences in the left inferior frontal gyrus, left dorsolateral prefrontal cortex, left temporoparietal junction, and the left inferior parietal cortex among uninfected controls, unimpaired HIV-infected adults, and adults with HAND. (B) Whole-brain ANOVA of alpha flanker interference activity across the same three groups revealed significant differences in the left superior frontal gyrus, left middle frontal gyrus, left supramarginal gyrus, left angular gyrus, and right dorsolateral prefrontal cortex. Both maps are shown at an alpha level of .05 (F > 4.3).
Figure 12. Frontal Theta Activity Predicts Neuropsychological Performance.
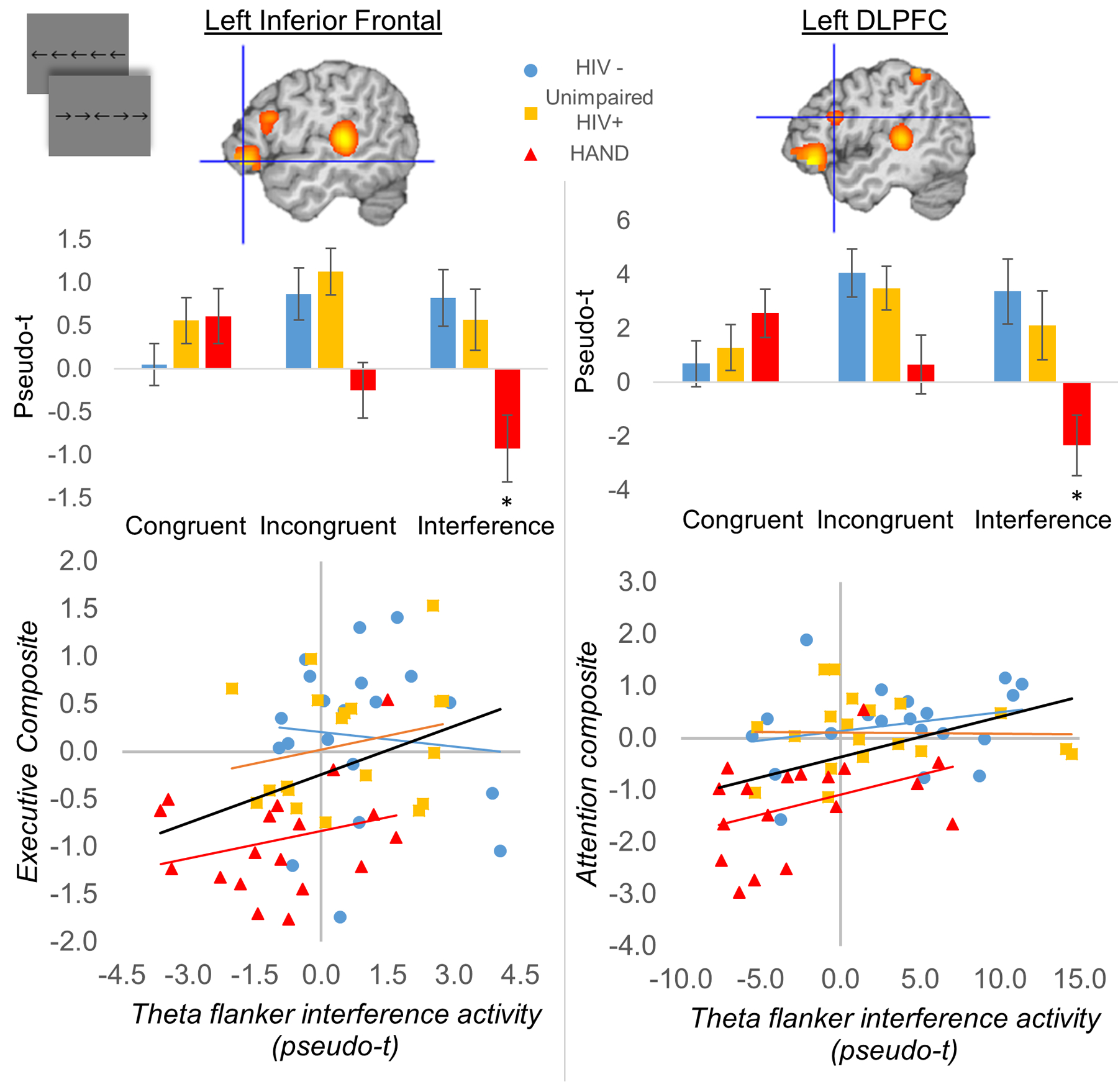
(Left) Flanker interference activity (Incongruent – Congruent activity) in the left inferior frontal gyrus predicted executive function composite scores (p < .05). (Right) Flanker interference activity in the left dorsolateral prefrontal cortex predicted attention composite scores (p < .05).
In conclusion, there have been three MEG studies of visual processing and/or visual attention function involving HIV-infected patients, and these studies have identified key aberrations across multiple nodes of the networks serving basic visual function, visuo-spatial processing, and executive function. Some of the most exciting findings have emerged in the past year with evidence that MEG markers of visual processing and visual selective attention may hold diagnostic value in distinguishing HIV-infected adults with and without cognitive impairment. The fact that these findings were observed in closely-matched, virally-suppressed individuals makes these data all the more exciting. Of course, further studies involving larger samples are necessary to confirm these findings, and to begin to move these data into prediction models for classifying groups. Regardless, visual processing studies in neuroHIV have provided critical new data in the past year, which have helped propel the functional neuroimaging of HIV community to new heights.
Conclusions & Future Directions
MEG is an ideal tool for examining the mechanisms of brain dysfunction in neuroHIV, as its spatiotemporal precision allows for accurate characterization of spectral alterations, as well as delays and slowing in specific neural systems. While most early studies focused on the resting-state, MEG studies of sensory processing in almost all modalities are now available and in the past two years multiple investigations of higher-order processes have now been published. Nonetheless, MEG studies of the neuronal aberrations associated with HIV-infection remain rare, with substantial work ongoing and many questions in need of further investigation.
Despite the sparsity of MEG investigations, several key conclusions can be drawn. First, aberrations in neural oscillatory activity stretch across all investigated sensory modalities, and in the same study are generally observed in both the primary sensory region (e.g., visual cortex) and higher-order brain areas such as the dorsolateral prefrontal cortex. These aberrations do not appear to be related to active viremia, as almost all MEG studies since 2013 have reported that their study populations were virally suppressed. Further, the notion that these abnormalities could be related to potential neurotoxic effects of certain antiretrovirals has not been supported in prior functional neuroimaging work.99 Demographic factors also appear unlikely as virtually all studies to date closely matched their patient and control groups on age, sex, ethnicity, and other important factors. Alterations in neurovascular coupling in HIV are also unlikely,100 as MEG is a direct measure of the physiology and should not be affected by such differences. This leads to the probable conclusion that such neural oscillatory aberrations are a key feature of the HIV-infected brain and not an artifact of inadequacies in study design or implementation.
Interestingly, the specific nature of these neural oscillatory abnormalities appears to differ by brain region and by cognitive/perceptual task. Some of these inter-regional disparities, especially the spectral findings, likely reflect low-level healthy differences in the underlying circuitry (e.g., healthy occipital neurons synchronize at the alpha rhythm when at rest), and thus the aberrations observed in HIV-infected adults likely reflect core dysfunction in these neural populations in regard to their specific abilities to compute, synchronize, and interact with neighboring and distant populations. Along these lines, it is interesting to note that HIV-related aberrations were often observed in the dominant spectral rhythm of the involved brain region. For example, widespread differences in beta activity were noted in the primary motor cortex of the only study to examine motor control in neuroHIV,57 and beta is widely acknowledged to be the dominant spectral rhythm of the motor system. Likewise, tasks with major visual components all reported differences in alpha activity,77,92–93 and differences in theta activity were often noted in the prefrontal cortex. Beyond these generalities, early MEG studies of the resting-state have highlighted deficits in fronto-visual circuits,29 which were later replicated in studies directly examining visual selective attention.93 Studies of somatosensory processing have universally shown deficits in the postcentral gyrus (primary somatosensory cortex) of HIV-infected adults.59,62 The one investigation of working memory processing focused on the verbal domain and showed behavioral deficits, as well as aberrations in memory maintenance operations across a broad network of left hemispheric language regions and the right inferior frontal gyrus (i.e., homologue of Broca’s area).77 Several studies of visual processing and visual attention have also been conducted and these have shown multispectral deficits in the occipital visual cortices, as well as parietal and prefrontal regions.91–93 Looking across studies it is interesting to note that studies of motor control,57 somatosensory processing,59 resting-state,29 working memory,77 visual processing,91 and visual selective attention93 all highlighted deficits in the prefrontal cortices of HIV-infected adults. Thus, conjunctive abnormalities in both modality-specific regions and prefrontal cortices appears to be a common thread across the majority of neuroHIV MEG studies to date. Finally, studies have shown that MEG measurements have high test-retest reliability in HIV-infected adults,28 which is clearly important for the development of MEG-based diagnostic markers of HAND and other diseases.
In regard to future studies, one of the most exciting areas has been the recent discovery that specific MEG metrics may be able to distinguish unimpaired HIV-infected individuals from those with HAND.92–93 This is a major breakthrough and future studies are needed to further refine the approach and test the predictive capacity of these markers in different HIV-infected populations (i.e., those not used in the development of the markers). The same and/or related MEG metrics may also be useful in monitoring HAND progression and establishing prognoses for HIV-infected individuals more broadly, but of course more work is needed. Regarding the latter, long-term longitudinal MEG studies would be the most informative route, so that the specific metric and clinical outcome could be continually monitored and compared. Concerning specific perceptual and cognitive domains of study, additional work is needed across the board. With no more than three studies in any specific area, there are numerous findings that need to be further confirmed and many lingering questions. Further, there are many domains that have yet to be investigated (e.g., auditory circuitry), and no studies to date have utilized executive function tasks, spatial working memory, nor multiple different forms of attention. Thus, many studies across many domains are required. The increasing availability of MEG should help facilitate this work over the next few years, as many new MEG sites are currently coming online. Finally, future studies should examine the impact of disease duration (time since diagnosis), CD4 nadir, substance abuse, and aging on HIV-related neural alterations, as these factors may be particularly important in understanding the pathophysiology and developing the most precise markers of HAND and its precursors.
Acknowledgements
This work was supported by NIH grants R01 MH103220 (TWW), R01 MH116782 (TWW), R01 MH118013 (TWW), R01 DA047828 (TWW), R03 DA041917 (TWW), P30 MH062261 (TWW), F31 AG055332 (AIW), and NSF grant #1539067 (TWW).
References
- 1.Nakagawa F, May M, Phillips A. (2013): Life expectancy living with HIV: recent estimates and future implications. Curr Opin Infect Dis 26(1):17–25. [DOI] [PubMed] [Google Scholar]
- 2.Antiretroviral Therapy Cohort Collaboration. (2008): Life expectancy of individuals on combination antiretroviral therapy in high-income countries: a collaborative analysis of 14 cohort studies. Lancet 372:293–299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Heaton RK, Clifford DB, Franklin DR Jr, et al. (2010): HIV-associated neurocognitive disorders persist in the era of potent antiretroviral therapy: CHARTER Study. Neurology 75:2087–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Heaton RK, Franklin DR, Ellis RJ, et al. (2011): HIV-associated neurocognitive disorders before and during the era of combination antiretroviral therapy: differences in rates, nature, and predictors. J Neurovirol 17:3–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gannon P, Khan MZ, Kolson DL. (2011): Current understanding of HIV-associated neurocognitive disorders pathogenesis. Curr Opin Neurol 24:275–83. Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Simioni S, Cavassini M, Annoni JM, et al. (2010): Cognitive dysfunction in HIV patients despite long-standing suppression of viremia. AIDS 24:1243–1250. [DOI] [PubMed] [Google Scholar]
- 7.Grant I, Franklin DR Jr, Deutsch R, et al. (2014): Asymptomatic HIV-associated neurocognitive impairment increases risk for symptomatic decline. Neurology 82:2055–2062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sacktor N, McDermott MP, Marder K, et al. (2002): HIV-associated cognitive impairment before and after the advent of combination therapy. J Neurovirol 8:136–42. [DOI] [PubMed] [Google Scholar]
- 9.Cysique LA, Brew BJ. (2009): Neuropsychological functioning and antiretroviral treatment in HIV/AIDS: A review. Neuropsychol Rev 19:169–85. Review. [DOI] [PubMed] [Google Scholar]
- 10.Antinori A, Arendt G, Becker JT, et al. (2007): Updated research nosology for HIV-associated neurocognitive disorders. Neurology 69:1789–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Robertson KR, Smurzynski M, Parsons TD, et al. (2007): The prevalence and incidence of neurocognitive impairment in the HAART era. AIDS 21:1915–1921. [DOI] [PubMed] [Google Scholar]
- 12.Dore GJ, McDonald A, Li Y, et al. (2003): Marked improvement in survival following AIDS dementia complex in the era of highly active antiretroviral therapy. AIDS 17:1539–1545. [DOI] [PubMed] [Google Scholar]
- 13.Heaton RK, Marcotte TD, Mindt MR, et al. (2004): The impact of HIV-associated neuropsychological impairment on everyday functioning. J Int Neuropsychol Soc 10:317–331. [DOI] [PubMed] [Google Scholar]
- 14.Albert SM, Marder K, Dooneief G, et al. (1995): Neuropsychologic impairment in early HIV infection. A risk factor for work disability. Arch Neurol 52:525–530. [DOI] [PubMed] [Google Scholar]
- 15.Marcotte TD, Wolfson T, Rosenthal TJ, et al. (2004): A multimodal assessment of driving performance in HIV infection. Neurology 63:1417–22. [DOI] [PubMed] [Google Scholar]
- 16.Hämäläinen M, Hari R, Ilmoniemi RJ, Knuutila J, Lounasmaa OV Magnetoencephalography -Theory, instrumentation, and applications to noninvasive studies of the working human brain. Rev Mod Phys. 1993;65(2):413–495. [Google Scholar]
- 17.Wilson TW (2014). Noninvasive neurophysiological imaging with magnetoencephalography. In Xiong H & Gendelman HE (Eds), Current laboratory methods in neuroscience research (pp.293–311). New York: Springer Press. [Google Scholar]
- 18.Supek S, Aine CJ (Eds). (2014). Magnetoencephalography: From signals to dynamic cortical networks. Verlag: Springer Press. [Google Scholar]
- 19.Hansen PC, Kringelbach ML, Salmelin R (Eds). (2010). MEG: An introduction to the methods. New York: Oxford University Press. [Google Scholar]
- 20.Hari R, Salmelin R Magnetoencephalography: From SQUIDs to neuroscience. Neuroimage 20th anniversary special edition. Neuroimage. 2012;61(2):386–96. [DOI] [PubMed] [Google Scholar]
- 21.Wilson TW, Heinrichs-Graham E, Proskovec AL, McDermott TJ. (2016): Neuroimaging with magnetoencephalography: A dynamic view of brain pathophysiology. Transl Res. 175:17–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Murakami S, Okada Y Contributions of principal neocortical neurons to magnetoencephalography and electroencephalography signals. J Physiol. 2006;575:925–936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hillebrand A, Barnes GR A quantitative assessment of the sensitivity of whole-head MEG to activity in the adult human cortex. Neuroimage. 2002;16:638.-. [DOI] [PubMed] [Google Scholar]
- 24.Cohen DS Magnetoencephalography: Evidence of magnetic fields produced by alpha rhythm currents. Science 1968;161:784.-. [DOI] [PubMed] [Google Scholar]
- 25.Jensen O, Spaak E, Zumer JM (2014). Human brain oscillations: From physiological mechanisms to analysis and cognition. In Supek S & Aine CJ (Eds), Magnetoencephalography: From signals to dynamic cortical networks (pp. 359–403). Verlag: Springer Press. [Google Scholar]
- 26.Uhlhaas PJ, Pipa G, Lima B, Melloni L, Neuenschwander S, Nikolić D, Singer W Neural synchrony in cortical networks: history, concept and current status. Front Integr Neurosci. 2009;3:17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.O’Neill GC, Tewarie P, Vidaurre D, Liuzzi L, Woolrich MW, Brookes MJ. Dynamics of large-scale electrophysiological networks: A technical review. Neuroimage 2018; 180:559–576. [DOI] [PubMed] [Google Scholar]
- 28.Becker JT, Fabrizio M, Sudre G, Haridis A, Ambrose T, Aizenstein HJ, Eddy W, Lopez OL, Wolk DA, Parkkonen L, Bagic A. (2012): Potential utility of resting-state magnetoencephalography as a biomarker of CNS abnormality in HIV disease. J Neurosci Methods 206(2):176–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Becker JT, Bajo R, Fabrizio M, Sudre G, Cuesta P, Aizenstein HJ, Lopez OL, Wolk D, Parkkonen L, Maestu F, Bagic A. (2012): Functional connectivity measured with magnetoencephalography identifies persons with HIV disease. Brain Imaging Behav 6(3):366–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Becker KM, Heinrichs-Graham E, Fox HS, Robertson KR, Sandkovsky U, O’Neill J, Swindells S, Wilson TW. (2013): Decreased MEG beta oscillations in HIV-infected older adults during the resting-state. J Neurovirol 19:586–594. PMC3913174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hardy DJ, Vance DE (2009): The neuropsychology of HIV/AIDS in older adults. Neuropsychol Rev 19:263–72. [DOI] [PubMed] [Google Scholar]
- 32.Joska JA, Gouse H, Paul RH, Stein DJ, Flisher AJ (2010) Does highly active antiretroviral therapy improve neurocognitive function? A systematic review. J Neurovirol 6:101–14. Review. [DOI] [PubMed] [Google Scholar]
- 33.Robertson K, Liner J, Heaton R (2009) Neuropsychological assessment of HIV-infected populations in international settings. Neuropsychol Rev 19:232–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Woods SP, Moore DJ, Weber E, Grant I (2009) Cognitive neuropsychology of HIV-associated neurocognitive disorders. Neuropsychol Rev 19:152–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Janssen RS, Cornblath DR, Epstein LG, McArthur J, Price RW (1991) Nomenclature and research case definitions for neurological manifestations of human immunodeficiency virus type-1 (HIV-1) infection. Report of a Working Group of the American Academy of Neurology AIDS Task Force. Neurology 41: 778–85. [DOI] [PubMed] [Google Scholar]
- 36.The Mind Exchange Working Group. Assessment, diagnosis, and treatment of HIV-associated neurocognitive disorder: A consensus report of the mind exchange program. Clin Infect Dis 2013. January 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Cheyne D, Bakhtazad L, Gaetz W, 2006. Spatiotemporal mapping of cortical activity accompanying voluntary movements using an event-related beamforming approach. Hum Brain Mapp 27, 213–229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gaetz W, Macdonald M, Cheyne D, Snead OC, 2010. Neuromagnetic imaging of movement-related cortical oscillations in children and adults: age predicts post-movement beta rebound. Neuroimage 51, 792–807. [DOI] [PubMed] [Google Scholar]
- 39.Heinrichs-Graham E, Wilson TW, 2015. Coding complexity in the human motor circuit. Hum Brain Mapp 36, 5155–5167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Heinrichs-Graham E, Wilson TW, 2016. Is an absolute level of cortical beta suppression required for proper movement? Magnetoencephalographic evidence from healthy aging. Neuroimage 134, 514–521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Heinrichs-Graham E, Wilson TW, Santamaria PM, Heithoff SK, Torres-Russotto D, Hutter-Saunders JA, Estes KA, Meza JL, Mosley RL, Gendelman HE, 2014. Neuromagnetic evidence of abnormal movement-related beta desynchronization in Parkinson’s disease. Cereb Cortex 24, 2669–2678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Jurkiewicz MT, Gaetz WC, Bostan AC, Cheyne D, 2006. Post-movement beta rebound is generated in motor cortex: evidence from neuromagnetic recordings. Neuroimage 32, 1281–1289. [DOI] [PubMed] [Google Scholar]
- 43.Wilson TW, Heinrichs-Graham E, Becker KM, 2014. Circadian modulation of motor-related beta oscillatory responses. Neuroimage 102 Pt 2, 531–539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wilson TW, Slason E, Asherin R, Kronberg E, Reite ML, Teale PD, Rojas DC, 2010. An extended motor network generates beta and gamma oscillatory perturbations during development. Brain Cogn 73, 75–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wilson TW, Slason E, Asherin R, Kronberg E, Teale PD, Reite ML, Rojas DC, 2011. Abnormal gamma and beta MEG activity during finger movements in early-onset psychosis. Dev Neuropsychol 36, 596–613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kurz MJ, Becker KM, Heinrichs-Graham E, Wilson TW, 2014. Neurophysiological abnormalities in the sensorimotor cortices during the motor planning and movement execution stages of children with cerebral palsy. Dev Med Child Neurol 56, 1072–1077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Tzagarakis C, Ince NF, Leuthold AC, Pellizzer G, 2010. Beta-band activity during motor planning reflects response uncertainty. J Neurosci 30, 11270–11277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Grent-’t-Jong T, Oostenveld R, Jensen O, Medendorp WP, Praamstra P, 2014. Competitive interactions in sensorimotor cortex: oscillations express separation between alternative movement targets. J Neurophysiol 112, 224–232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Heinrichs-Graham E, Arpin DJ, Wilson TW, 2016. Cue-related Temporal Factors Modulate Movement-related Beta Oscillatory Activity in the Human Motor Circuit. J Cogn Neurosci 28, 1039–1051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Heinrichs-Graham E, Kurz MJ, Gehringer JL, Wilson TW, 2017. The functional role of post-movement beta oscillations in movement termination. Brain Struct Funct. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Muthukumaraswamy SD, 2010. Functional properties of human primary motor cortex gamma oscillations. J Neurophysiol 104, 2873–2885. [DOI] [PubMed] [Google Scholar]
- 52.Grent-’t-Jong T, Oostenveld R, Jensen O, Medendorp WP, Praamstra P, 2013. Oscillatory dynamics of response competition in human sensorimotor cortex. Neuroimage 83, 27–34. [DOI] [PubMed] [Google Scholar]
- 53.Cheyne D, Bells S, Ferrari P, Gaetz W, Bostan AC, 2008. Self-paced movements induce high-frequency gamma oscillations in primary motor cortex. Neuroimage 42, 332–342. [DOI] [PubMed] [Google Scholar]
- 54.Gaetz W, Edgar JC, Wang DJ, Roberts TP, 2011. Relating MEG measured motor cortical oscillations to resting gamma-aminobutyric acid (GABA) concentration. Neuroimage 55, 616–621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Gaetz W, Liu C, Zhu H, Bloy L, Roberts TP, 2013. Evidence for a motor gamma-band network governing response interference. Neuroimage 74, 245–253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Heinrichs-Graham E, Hoburg JM, Wilson TW. The peak frequency of motor-related gamma oscillations is modulated by response competition. Neuroimage 2018. January 15;165:27–34. doi: 10.1016/j.neuroimage.2017.09.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Wilson TW, Heinrichs-Graham E, Robertson KR, et al. (2013): Functional brain abnormalities during finger-tapping in HIV-infected older adults: A magnetoencephalography study. J Neuroimmune Pharmacol 8(4):965–974. PMC3809128 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Thompson PM, Dutton RA, Hayashi KM, Toga AW, Lopez OL, Aizenstein HJ, et al. (2005): Thinning of the cerebral cortex visualized in HIV/AIDS reflects CD4+ T lymphocyte decline. Proc Natl Acad Sci U S A 102:15647–15652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wilson TW, Heinrichs-Graham E, Becker KM, Aloi J, Robertson KR, Sandkovsky U, White ML, O’Neill J, Knott NL, Fox HS, Swindells S. (2015): Multimodal neuroimaging evidence of alterations in cortical structure and function in HIV-infected older adults. Hum Brain Mapp 36:897–910. PMC4491915 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ashburner J (2007): A fast diffeomorphic image registration algorithm. Neuroimage 38:95–113. [DOI] [PubMed] [Google Scholar]
- 61.Yousry TA, Schmid UD, Alkadhi H, Schmidt D, Peraud A, Buettner A, et al. (1997): Localization of the motor hand area to a knob on the precentral gyrus: A new landmark. Brain120:141–157. [DOI] [PubMed] [Google Scholar]
- 62.Spooner RK, Wiesman AI, Mills MS, O’Neill J, Robertson KR, Fox HS, Swindells S, Wilson TW. Aberrant oscillatory dynamics during somatosensory processing in HIV-infected adults. Neuroimage Clin. 2018. July 10;20:85–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Cromwell HC, Mears RP, Wan L, Boutros NN, 2008. Sensory gating: a translational effort from basic to clinical science. Clin EEG Neurosci 39, 69–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kurz MJ, Wiesman AI, Coolidge NM, Wilson TW, 2017. Children with Cerebral Palsy Hyper-Gate Somatosensory Stimulations of the Foot. Cereb Cortex, 1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Spooner RK, Wiesman AI, Proskovec AL, Heinrichs-Graham E, Wilson TW, 2019. Rhythmic Spontaneous Activity Mediates the Age-Related Decline in Somatosensory Function. Cereb Cortex, 29, 680–688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Thoma RJ, Meier A, Houck J, Clark VP, Lewine JD, Turner J, Calhoun V, Stephen J, 2017. Diminished auditory sensory gating during active auditory verbal hallucinations. Schizophr Res. 188, 125–131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Gross AM, Jaeger PA, Kreisberg JF, Licon K, Jepsen KL, Khosroheidari M, Morsey BM, Swindells S, Shen H, Ng CT, Flagg K, Chen D, Zhang K, Fox HS, Ideker T. (2016): Methylome-wide Analysis of Chronic HIV Infection Reveals Five-Year Increase in Biological Age and Epigenetic Targeting of HLA. Mol Cell 62(2):157–168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Heinrichs-Graham E, McDermott TJ, Mills MS, Wiesman AI, Wang YP, Stephen JM, Calhoun VD, Wilson TW. The lifespan trajectory of neural oscillatory activity in the motor system. Dev Cogn Neurosci. 2018. April;30:159–168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sundermann EE, et al. Genetic predictor of working memory and prefrontal function in women with HIV. J. Neurovirol 21, 81–91 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Woods SP, Moore DJ, Weber E, Grant I Cognitive neuropsychology of HIV-associated neurocognitive disorders. Neuropsychol. Rev 19, 152–168 (2009). Review. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Plessis SD, et al. HIV infection and the fronto-striatal system: a systematic review and meta-analysis of fMRI studies. AIDS 28, 803–811 (2014). Review. [DOI] [PubMed] [Google Scholar]
- 72.Ernst T, Chang L, Jovicich J, Ames N, Arnold S Abnormal brain activation on functional MRI in cognitively asymptomatic HIV patients. Neurology 59, 1343–1349 (2002). [DOI] [PubMed] [Google Scholar]
- 73.Ernst T, et al. Declined neural efficiency in cognitively stable human immunodeficiency virus patients. Ann. Neurol 65, 316–325 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Chang L, Yakupov R, Nakama H, Stokes B, Ernst T Antiretroviral treatment is associated with increased attentional load-dependent brain activation in HIV patients. J. Neuroimmune Pharmacol 3, 95–104 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Ances BM, et al. Resting cerebral blood flow: a potential biomarker of the effects of HIV in the brain. Neurology 73, 702–708 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Ances BM, et al. HIV infection and aging independently affect brain function as measured by functional magnetic resonance imaging. J. Infect. Dis 201, 336–340 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Wilson TW, Proskovec AL, Heinrichs-Graham E, O’Neill J, Robertson KR, Fox HS, & Swindells S. (2017): Aberrant neuronal dynamics during working memory operations in the aging HIV-infected brain. Sci Rep 7:41568. PMC5290733 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Embury CM, Wiesman AI, Proskovec AL, Heinrichs-Graham E, McDermott TJ, Lord GH, Brau KL, Drincic AT, Desouza CV, Wilson TW. 2018. Altered Brain Dynamics in Patients With Type 1 Diabetes During Working Memory Processing. Diabetes. 67:1140–1148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Embury CM, Wiesman AI, Proskovec AL, Mills MS, Heinrichs-Graham E, Wang YP, Calhoun VD, Stephen JM, Wilson TW. 2018. Neural dynamics of verbal working memory processing in children and adolescents. Neuroimage. 185:191–197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Heinrichs-Graham E, Wilson TW. 2015. Spatiotemporal oscillatory dynamics during the encoding and maintenance phases of a visual working memory task. Cortex. 69:121–130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.McDermott TJ, Badura-Brack AS, Becker KM, Ryan TJ, Bar-Haim Y, Pine DS, Khanna MM, Heinrichs-Graham E, Wilson TW. 2016. Attention training improves aberrant neural dynamics during working memory processing in veterans with PTSD. Cogn Affect Behav Neurosci. 16:1140–1149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.McDermott TJ, Badura-Brack AS, Becker KM, Ryan TJ, Khanna MM, Heinrichs-Graham E, Wilson TW. 2016. Male veterans with PTSD exhibit aberrant neural dynamics during working memory processing: an MEG study. J Psychiatry Neurosci. 41:251–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Proskovec AL, Heinrichs-Graham E, Wilson TW. 2016. Aging modulates the oscillatory dynamics underlying successful working memory encoding and maintenance. Hum Brain Mapp. 37:2348–2361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Proskovec AL, Heinrichs-Graham E, Wilson TW. 2019. Load modulates the alpha and beta oscillatory dynamics serving verbal working memory. Neuroimage. 184:256–265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Wiesman AI, Heinrichs-Graham E, McDermott TJ, Santamaria PM, Gendelman HE, Wilson TW. 2016. Quiet connections: Reduced fronto-temporal connectivity in nondemented Parkinson’s Disease during working memory encoding. Hum Brain Mapp. 37:3224–3235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Proskovec AL, Wiesman AI, Heinrichs-Graham E, Wilson TW. 2018. Beta Oscillatory Dynamics in the Prefrontal and Superior Temporal Cortices Predict Spatial Working Memory Performance. Sci Rep 8:8488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Proskovec AL, Wiesman AI, Heinrichs-Graham E, Wilson TW. 2019. Load effects on spatial working memory performance are linked to distributed alpha and beta oscillations. Human Brain Mapping In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Chang L, Speck O, Miller EN, Braun J, Jovicich J, Koch C, et al. Neural correlates of attention and working memory deficits in HIV patients. Neurology 2001; 57(6): 1001–7. [DOI] [PubMed] [Google Scholar]
- 89.Chang L, Tomasi D, Yakupov R, Lozar C, Arnold S, Caparelli E, et al. Adaptation of the attention network in human immunodeficiency virus brain injury. Ann Neurol 2004; 56(2): 259–72. [DOI] [PubMed] [Google Scholar]
- 90.Thomas JB, Brier MR, Snyder AZ, Vaida FF, Ances BM. Pathways to neurodegeneration: effects of HIV and aging on resting-state functional connectivity. Neurology 2013; 80(13): 1186–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Wilson TW, Fox HS, Robertson KR, et al. (2013): Abnormal MEG oscillatory activity during visual processing in the prefrontal cortices and frontal eye-fields of the aging HIV brain. PLoS One, 8(6):e66241. PMC3688779 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Wiesman AI, O’Neill J, Mills MS, Robertson KR, Fox HS, Swindells S, Wilson TW. Aberrant occipital dynamics differentiate HIV-infected patients with and without cognitive impairment. Brain. 2018. June 1;141(6):1678–1690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Lew BJ, McDermott TJ, Wiesman AI, O’Neill J, Mills MS, Robertson KR, Fox HS, Swindells S, Wilson TW. Neural dynamics of selective attention deficits in HIV-associated neurocognitive disorder. Neurology. 2018. November 13;91(20):e1860–e1869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Wiesman AI, Wilson TW. The impact of age and sex on the oscillatory dynamics of visuospatial processing. Neuroimage. 2019. January 15;185:513–520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Wiesman AI, Mills MS, McDermott TJ, Spooner RK, Coolidge NM, Wilson TW. Polarity-dependent modulation of multi-spectral neuronal activity by transcranial direct current stimulation. Cortex. 2018. November;108:222–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Wiesman AI, Heinrichs-Graham E, Proskovec AL, McDermott TJ, Wilson TW. Oscillations during observations: Dynamic oscillatory networks serving visuospatial attention. Hum Brain Mapp. 2017. October;38(10):5128–5140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.McDermott TJ, Wiesman AI, Proskovec AL, Heinrichs-Graham E, Wilson TW. Spatiotemporal oscillatory dynamics of visual selective attention during a flanker task. Neuroimage. 2017. August 1;156:277–285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.McDermott TJ, Wiesman AI, Mills MS, Spooner RK, Coolidge NM, Proskovec AL, Heinrichs-Graham E, Wilson TW. tDCS modulates behavioral performance and the neural oscillatory dynamics serving visual selective attention. Hum Brain Mapp. 2019. February 15;40(3):729–740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Brier MR, Wu Q, Tanenbaum AB, Westerhaus ET, Kharasch ED, Ances BM. Effect of HAART on Brain Organization and Function in HIV-Negative Subjects. J Neuroimmune Pharmacol 2015. December;10(4):517–21. doi: 10.1007/s11481-015-9634-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Ances B, Vaida F, Ellis R, Buxton R. Test-retest stability of calibrated BOLD-fMRI in HIV- and HIV+ subjects. Neuroimage. 2011. February 1;54(3):2156–62. [DOI] [PMC free article] [PubMed] [Google Scholar]