

Abstract
Gamma oscillations have been argued to support visual perception by synchronizing the processing and transfer of information within and across areas of visual cortex. Here, we highlight recent findings implicating the influence of color on visual gamma oscillations and how these observations may relate to local cortical tuning and organization.
Visually induced gamma oscillations are high-frequency (>30 Hz) fluctuations in electrical brain activity that have been proposed to support perception by synchronizing neural firing [1]. Here, we highlight recent explorations into the influence of stimulus color on gamma oscillations. Together with a novel discovery of visual circuit organization, they advance our understanding of the neural processes underlying visual gamma oscillations and their role in perception.
Historically, gamma oscillations have been studied by recording local field potentials (LFP) in early visual cortex in response to simple grating stimuli. Over the years, the properties of induced gamma oscillations have been shown to be dependent on low-level stimulus attributes. For example, large diameter and high-contrast grating stimuli increase the amplitude and peak frequency of induced gamma oscillations [2]. Prior work has linked the influence of grating size to surround-suppression phenomena, such that large-scale gamma oscillations occur under conditions where local inhibition is strong [3]. Indeed, computational models of gamma genesis are based on the interplay between excitation and inhibition [1], for which specific inter-neurons are critical in generating oscillatory population dynamics. Therefore, it is important to understand how these stimulus–response properties are engaged during natural vision, particularly because evidence for visual gamma oscillations in response to more complex image stimuli differs between studies and has been an area of debate [4,5].
Why are grating stimuli particularly effective at inducing gamma oscillations? Intuitively, this could be related to the well-known orientation tuning of cells in primary visual cortex (V1). However, effects seen at the single-cell level do not necessarily capture the full range of responses observed at the population LFP level. Recently, Hermes and colleagues [6] used a simple model to demonstrate that low variability of oriented edges falling within the receptive field predicts the occurrence of gamma oscillations in visual cortex to both gratings and natural images. These findings highlight the sensitivity of gamma oscillations to the spatial structure of visual stimuli, in particular spatially homogenous oriented edges, such as gratings, which recruit inhibitory recurrent networks. However, more recent empirical work has also highlighted the importance of another stimulus dimension, orthogonal to spatial structure, color.
Recording from macaque V1, Shirhatti and Ray [7] recently reported a striking modulation in the amplitude of gamma oscillations in response to full-field uniform color stimuli. Remarkably, gamma amplitude varied as a function of stimulus hue, such that red stimuli elicited the strongest response, even greater than that observed for traditionally used achromatic gratings. This striking color-dependent modulation of gamma amplitude was further investigated by Peter and colleagues [8], who also quantified the influence of stimulus size, regularity across the receptive field, and color-opponency mechanisms. Importantly, they showed that, while the largest gamma oscillations were induced by red hues under many viewing conditions, an opponent color in the surround (green) or the adaptation to different background colors modulated this effect. Therefore, the strength of color tuning of gamma oscillations is also modulated by center-surround effects (increasing inhibition) and color-opponency mechanisms [8]. Most recently, Bartoli and colleagues [9] examined gamma oscillations in human visual cortex, using achromatic gratings, large-field uniform color stimuli, and natural images. Consistent with the non-human primate findings, the amplitude of gamma oscillations was strongest for achromatic gratings and red hues. Crucially, gamma amplitude was significantly enhanced when the same natural image was presented in red color versus gray-scale. Altogether, these studies highlight how color, color opponency, and the spatial structure of the stimulus can greatly influence the magnitude of visual gamma oscillations.
How do spatially uniform colors drive gamma oscillations similar to those evoked by gratings? Gamma oscillations appear to emerge when visual inputs contain features that match the tuning of local cortical neurons, and when these features are presented in a spatially coherent manner across the receptive field and its surround (Figure 1). The presence of these highly represented visual features in the stimulus creates a strong excitatory drive, with their high spatial homogeneity leading in parallel to increased inhibition. The origin of the inhibitory drive is an open question for future studies, as it may be linked to center-surround effects [3,8] or other mechanisms for homeostatic gain control, such as divisive normalization. It may seem surprising that stimulus orientation and color would engage in similar neural dynamics (i.e., gamma oscillations) as these two orthogonal classes of stimuli are traditionally conceptualized as being processed by separate neural pathways. However, the existence of neurons exhibiting both orientation tuning and color responsivity is suggestive of tight interconnections between these features in V1 [10]. Recently, work by Garg et al. provided a critical update to our view of orientation and color processing in primate V1 [11], showing that the spatial arrangement of cells sensitive to these features form partially overlapping circuits and domains. Using two-photon imaging to evaluate the response of macaque V1 to achromatic and chromatic gratings, Garg et al. revealed that many neurons display both color and orientation tuning. This finding should prompt further consideration for how the organization of neural tuning properties impacts population-level dynamics.
Figure 1. Schematic of Stimulus Features Influencing Gamma Generation in V1.
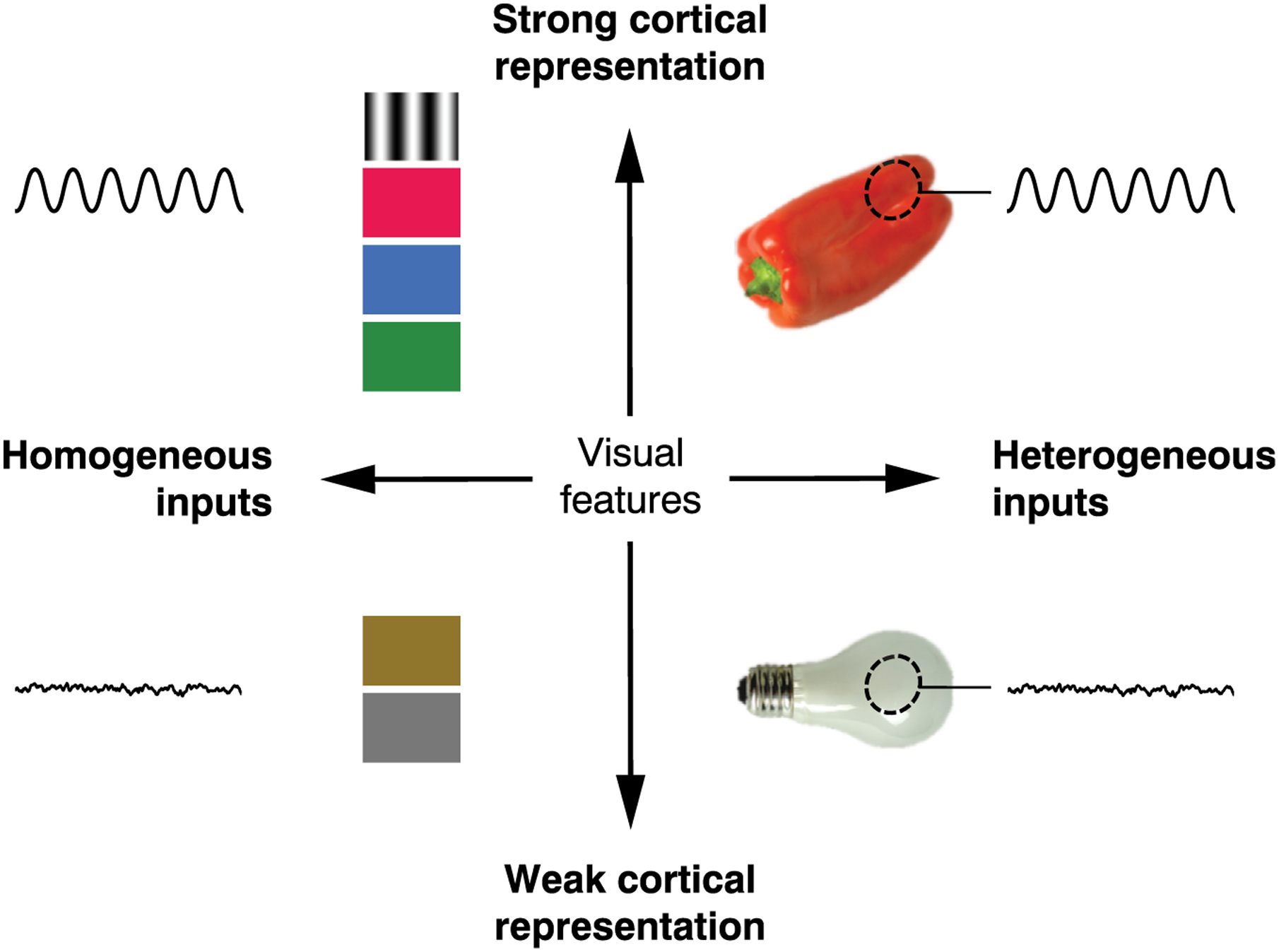
Visual stimuli and corresponding neural responses arranged by homogeneity of the stimulus (horizontal axis) and strength of the cortical representation for features contained in the stimulus (vertical axis). Gamma oscillations occur in response to visual features that are strongly represented in primary visual cortex and that are either homogeneous across the entire visual field (upper left) or homogeneous within the population receptive field (broken ellipse, upper right). Gamma oscillations are less reliably observed in response to stimuli that contain a heterogeneous mix of features (bottom right) or for stimuli containing features that are weakly represented in V1 (bottom left).
Specifically, the distribution and interactions of cells, responsive to different visual features, are fundamental in driving population tuning properties of gamma oscillations.
As noted earlier, recent electrophysiological studies evaluating the color tuning of gamma oscillations observed the strongest response to red hues under most visual conditions. Why would a uniform red stimulus induce stronger oscillations? There is currently no explanation for this puzzling result, only a handful of clues. Regarding color preference, Garg et al. found more cells preferring blue, followed by red, but few for green. However, red-selective cells displayed the highest proportion of joint color/orientation tuning, as well as the strongest orientation tuning [11]. Therefore, the unequal representation of cells with different joint-tuning preferences could underlie the observed bias toward red stimuli. However, it is still unclear why uniform red colors would be driving strong population responses. Color-opponency mechanisms may unevenly integrate cone signals in the retina and lateral geniculate nucleus [12], potentially contributing to the bias. Within V1, strong excitatory inputs would result in recurrent population dynamics (gamma oscillations) due to the spatial homogeneity of the stimulus across visual space, similar to gratings (Figure 2). Other circuit properties could also favor a stronger recurrent network activity to red: color channels may adapt differently [8] or V1 may exhibit a bias toward ecologically relevant color contrasts, perhaps driven by evolutionary needs. Future studies will be able to test these hypotheses and shed light on the complexity of color representation and its population-level encoding. Overall, these findings challenge future researchers to carefully control the color content of visual stimuli when investigating gamma oscillations.
Figure 2. Schematic of Joint Tuning in V1 and Possible Links to Gamma Generation.
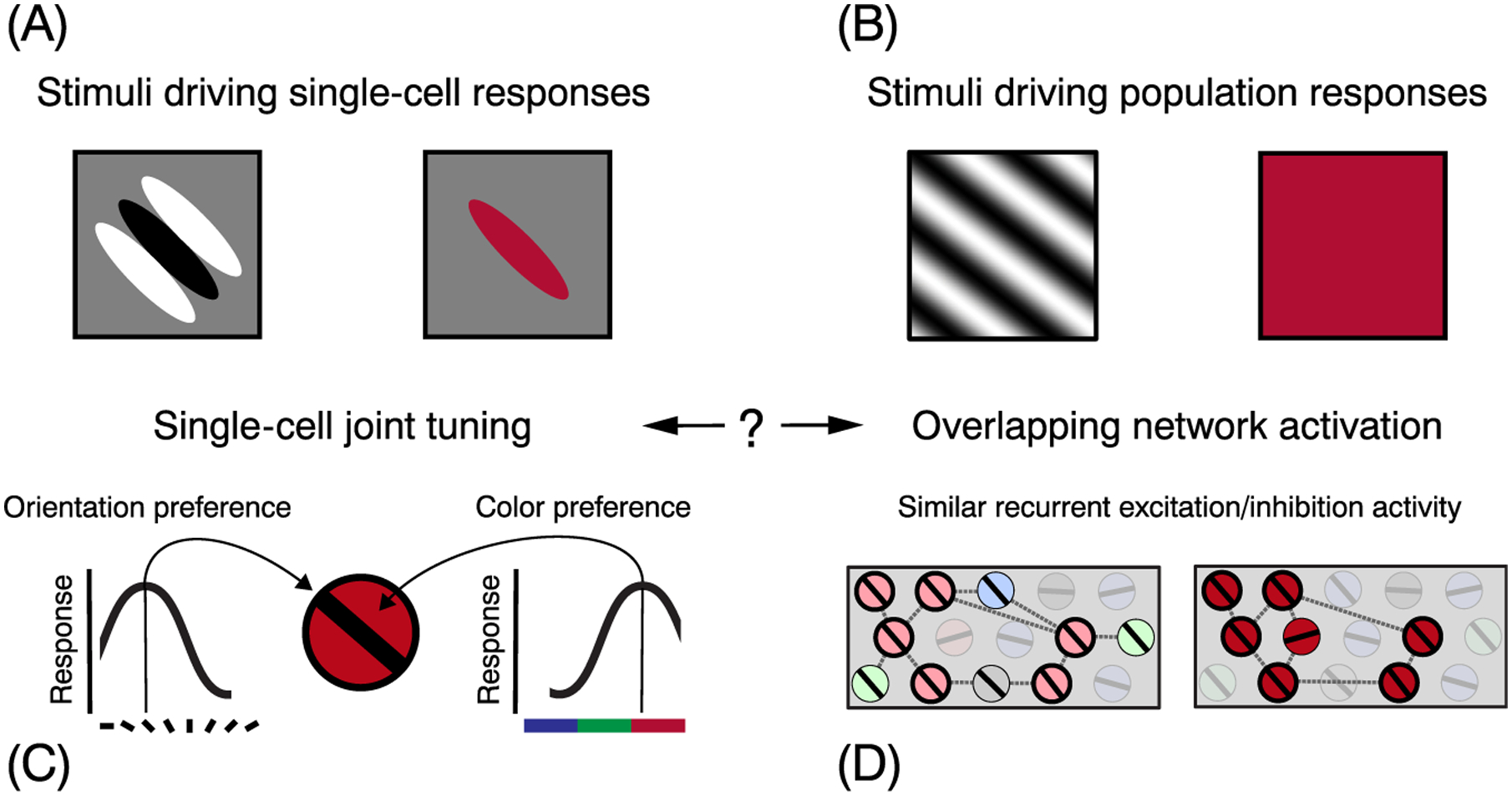
Different types of visual input drive responses at the single-cell (A) and population level (B). On the left, we schematize individual cells in V1 with joint tuning for orientation and color (C). On the right, we speculate on how large gratings and uniform red color stimuli could activate an overlapping subset of cells within the orientation and color feature maps [(D) highlighted cells], which are highly interconnected (broken lines) and recruit similar recurrent networks, leading to large gamma responses (one measure of population activity). Note that the cortical feature maps depicted here are highly schematized and do not accurately represent either the functional organization of feature maps or the nature of the excitatory and inhibitory interactions within those maps. How to reconcile single-cell and population-level stimulus–response mappings is a challenge for future studies.
We propose that cellular tuning properties and the spatial organization of stimulus feature maps have a crucial role in shaping the excitatory and inhibitory interactions that underlie gamma oscillations. In light of the recent findings summarized here, we suggest that gamma oscillations potentially constitute a population readout of local circuit tuning, carrying coarse information about visual input along primary stimulus dimensions. Therefore, visual gamma oscillations offer an opportunity to understand how population-level responses originate from single-cell properties and how similar principles may arise in other cortical regions. This not only provides an exciting testing ground for models seeking to link scales of brain organization, but also prompts a rethinking of the role of gamma oscillations in perception.
References
- 1.Buzsaki G and Wang XJ (2012) Mechanisms of gamma oscillations. Annu. Rev. Neurosci 35, 203–225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ray S and Maunsell JH (2010) Differences in gamma frequencies across visual cortex restrict their possible use in computation. Neuron 67, 885–896 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gieselmann MA and Thiele A (2008) Comparison of spatial integration and surround suppression characteristics in spiking activity and the local field potential in macaque V1. Eur. J. Neurosci 28, 447–459 [DOI] [PubMed] [Google Scholar]
- 4.Hermes D et al. (2015) Gamma oscillations in visual cortex: the stimulus matters. Trends Cogn. Sci 19, 57–58 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Brunet N et al. (2014) Gamma or no gamma, that is the question. Trends Cogn. Sci 18, 507–509 [DOI] [PubMed] [Google Scholar]
- 6.Hermes D et al. (2019) An image-computable model for the stimulus selectivity of gamma oscillations. eLife 8, e47035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Shirhatti V and Ray S (2018) Long-wavelength (reddish) hues induce unusually large gamma oscillations in the primate primary visual cortex. Proc. Natl. Acad. Sci. U. S. A 115, 4489–4494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Peter A et al. (2019) Surface color and predictability determine contextual modulation of V1 firing and gamma oscillations. eLife 8, e42101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bartoli E et al. (2019) Functionally distinct gamma range activity revealed by stimulus tuning in human visual cortex. Curr. Biol 29, 3345–3358 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Shapley R and Hawken M (2002) Neural mechanisms for color perception in the primary visual cortex. Curr. Opin. Neurobiol 12, 426–432 [DOI] [PubMed] [Google Scholar]
- 11.Garg AK et al. (2019) Color and orientation are jointly coded and spatially organized in primate primary visual cortex. Science 364, 1275–1279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.De Valois RL and De Valois KK (1993) A multi-stage color model. Vis. Res 33, 1053–1065 [DOI] [PubMed] [Google Scholar]