

Abstract
Rab35 is a small GTPase that regulates plasma membrane to early endosome vesicular trafficking and mediates actin remodeling to form actin-rich cellular structures. While the function of Rab35 in the cellular context has been examined, its role during development has not been well studied. In this study, we take advantage of the sea urchin’s high fecundity, external fertilization, and transparent embryos to determine the function of Rab35 during development. We found that loss of function of Rab35 results in defects in skeletogenesis and gastrulation, which were rescued by co-injection of sea urchin Rab35. The loss of Rab35’s function results in decreased endocytosis and impaired exocytosis, which may be important for skeletogenesis and gastrulation. Skeletal spicules of Rab35 knockdown embryos have reduced organized actin compared to the control, supporting the notion that Rab35 regulates actin dynamics. In addition, the skeletal and gastrulation defects induced by Rab35 knockdown were rescued by co-injection with Fascin, an actin-bundling protein, indicating that proper actin dynamics play a critical role for both skeletogenesis and gastrulation. Overall, results indicate that through its role in mediating vesicular trafficking and actin remodeling, Rab35 is an important regulator of embryonic structure formation in early development.
Keywords: Fascin, Sea urchin, Apical constriction, Gut, Primary mesenchyme cells
1. Introduction
Rab35 is a member of the Rab family of small GTPases that plays essential roles in vesicular trafficking between the plasma membrane and endosomes and actin remodeling in eukaryotic cells (Chaineau et al., 2013; Klinkert and Echard, 2016). Rab35 is conserved throughout the metazoans (Pereira-Leal and Seabra, 2001), and was initially found to be an essential regulator of fast endocytic recycling (Kouranti et al., 2006). As a small GTPase, it cycles between an active GTP-bound form and an inactive GDP-bound form mediated by multiple guanine exchange factors (GEFs) and GTPase activating proteins (GAPs), respectively (Chaineau et al., 2013). In its active, GTP-bound form, it localizes to the plasma membrane of the cell, as well as early endosomes (Kouranti et al., 2006; Sato et al., 2008). Through its vesicular trafficking role, Rab35 is known to regulate several essential processes, such as apico-basal polarity (Klinkert et al., 2016), cytokinesis (Kouranti et al., 2006), and cell-surface receptor trafficking (Allaire et al., 2010, 2013; Charrasse et al., 2013), that are important in development.
Cell surface proteins endocytosed from the plasma membrane are generally sorted to either the trans-Golgi network, lysosomes for degradation, or recycled back to the plasma membrane (Grant and Donaldson, 2009). Rab35 has been found to regulate the recycling of cadherins and integrins, cell surface proteins that promote cell-cell adhesion and cell-extracellular matrix adhesion (ECM), respectively (Allaire et al., 2013; Charrasse et al., 2013; Grant and Donaldson, 2009). As these characteristics of decreased cell-cell adhesion and increased cell-ECM interaction are often seen in metastases, decreased Rab35 expression correlates with high grade gliomas, and malignant breast and squamous tumor cells (Allaire et al., 2013). Additionally, regulation of cell-cell and cell-ECM interaction is critical during development, as cells must move in a coordinated fashion in order to form structured tissue and organs (Kominami and Takata, 2004).
In addition to its role in vesicular trafficking, Rab35 also remodels actin through recruitment of known actin mediators Cdc42 and Rac1 to the plasma membrane (Chua et al., 2010; Shim et al., 2010), and direct interaction with an actin-bundling protein, Fascin (Zhang et al., 2009). One of Rab35’s GEFs, Connecdenn3/DENN1C, also directly interacts with actin, thereby localizing actin to newly activated Rab35 (Marat et al., 2012). In addition, Rab35 localizes Fascin to the leading edge of the cell membrane, where Fascin bundles actin in order to facilitate cell motility (Zhang et al., 2009). The interaction of Fascin and Rab35 has also been found to regulate vesicular trafficking. Disruption of the function of Fascin results in decreased endocytosis, as well as decreased release of extracellular vesicles (Beghein et al., 2018; Clancy et al., 2018; Nozumi et al., 2017).
The role of Rab35 in development has been examined in only a few contexts. Rab35 is known to play a key role in planar cell polarity in Drosophila germ band extension (GBE) (Jewett et al., 2017). It is localized to the anterior-posterior surface of the epithelium, where it is essential for the uptake of excess membrane to maintain apical constriction during GBE (Jewett et al., 2017). Additionally, Rab35 helps to initiate the formation of apico-basal polarity during tubulogenesis (Klinkert et al., 2016). As Rab35 localizes to the cleavage furrow of dividing cells (Chesneau et al., 2012; Klinkert et al., 2016; Kouranti et al., 2006), it directly binds to the cytoplasmic tail of an anti-adhesive trans-membrane protein (PODXL) (Klinkert et al., 2016). As the cells divide, the repulsive forces between the extra-cellular portion of PODXL on the daughter cells result in the formation of a lumen between them (Klinkert et al., 2016). It has also been observed that in Drosophila, Rab35 knockdown (KD) using RNAi results in bent and gnarled bristles, but overexpression of Fascin in the same tissue restores normal bristle morphology, indicating Fascin is a major downstream effector of Rab35 (Zhang et al., 2009).
While the function of Rab35 has not been examined in sea urchin development, other small GTPases have been identified as being involved, such as RhoA during primary invagination of gastrulation, and Arf6 and Cdc42 during skeletogenesis and gastrulation (Beane et al., 2006; Sepúlveda-Ramírez et al., 2018; Stepicheva et al., 2017). As Arf6 and Cdc42 have been shown to interact with Rab35 (Allaire et al., 2013; Chesneau et al., 2012; Shim et al., 2010), and actin remodeling and vesicular trafficking of cell surface proteins regulate skeletogenesis and gastrulation, these developmental processes may also be dependent upon Rab35 (Annunziata et al., 2014; Duloquin et al., 2007; Kimberly and Hardin, 1998).
Skeletogenesis in the echinoid sea urchins is the result of a series of well characterized morphogenetic changes in response to chemotactic cues from the ectoderm and activation of biomineralization genes (Decker and Lennarz, 1988). During blastulation, the primary mesenchymal cells (PMCs) that form the larval skeleton undergo an epithelial to mesenchymal transition (EMT) to ingress into the blastocoel (Wu et al., 2007), where they endocytose cadherin from the plasma membrane to detach from the epithelial layer and its neighboring cells (Miller and McClay, 1997). The PMCs also undergo apical constriction to form ‘bottle cells’ which requires actin to constrict the apical surface of the cell (Anstrom, 1989, 1992). Presumably, in response to the vascular endothelial growth factor (Vegf) signaling pathway, the PMCs begin to migrate anteriorly, extending filopodia to create a shell for the deposition of the mineral component of the skeleton (Duloquin et al., 2007; Ettensohn, 1990). As PMCs undergo these morphogenic movements, the transcription factors Alx1 and Ets1 activate the biomineralization genes required for skeletal construction (Rafiq et al., 2012). The formation of the larval skeleton is therefore dependent upon PMCs undergoing EMT, responding to chemotactic signals, regulating cytoskeletal elements, and initiating a biomineralization program.
Rab35 may also regulate gastrulation, a crucial step during embryogenesis. During this time, the embryonic gut is formed and all three germ layers are specified (Annunziata et al., 2014; McClay et al., 2020). Gastrulation occurs in distinct phases, known as primary and secondary gastrulation (Kominami and Takata, 2004). Primary gastrulation begins with the invagination of the posterior end of the embryo (Beane et al., 2006). Primary invagination is followed by an extension and narrowing of the gut tube (Kominami and Takata, 2004). The final step in gastrulation is the differentiation into the fore-, mid-, and hindgut segments (Annunziata et al., 2014). It is worth noting that several of the mechanisms proposed for primary invagination are also mechanisms that could play a role in vertebrate neurulation (Davidson et al., 1995).
While many of the cellular functions of Rab35 are known, its function in the context of development remains unclear. To elucidate the function of Rab35 in early development, we used the sea urchin model, because of its high fecundity, ability to undergo external fertilization, and transparent embryos, to allow us to observe cellular processes critical for key developmental processes, such as skeletogenesis and gastrulation (McClay, 2011). As previous studies have indicated, G-proteins that interact with Rab35 play important roles in sea urchin embryonic structure formation (Sepúlveda-Ramírez et al., 2018; Stepicheva et al., 2017). In this study, we examined the function of Rab35 in early development and defined its molecular mechanism in mediating proper skeletogenesis and gastrulation. In Rab35 KD embryos, we observed shortened dorsoventral connecting rods (DVCs), mispatterned PMCs, as well as defects in gastrulation. Results indicated that Rab35 KD embryos have defects in vesicular trafficking and perturbed actin polymerization that impact skeletogenesis and gastrulation. Rab35 KD phenotypes can be completely rescued by the sea urchin Rab35 and mostly by the human Rab35, indicating specificity of the knockdown and functional conservation of the sea urchin and human Rab35. We also found Fascin overexpression to restore Rab35 KD-induced defects in skeletogenesis and gastrulation. Overall, this study indicates that the small GTPase Rab35 mediates important cellular processes that are critical for the proper formation of embryonic structures.
2. Results
2.1. Rab35 is expressed at low levels during development
The spatial and temporal expression of Rab35 was assayed at various developmental stages, ranging from egg, early cleavage, early blastula, blastula, gastrula, and larval stages (Fig. 1). Using whole mount fluorescence RNA in situ hybridization (WMFISH), we observed Rab35 to be expressed at very low levels, with its expression detectable at the early blastula stage and becomes undetectable in later stages of development.
Fig. 1.
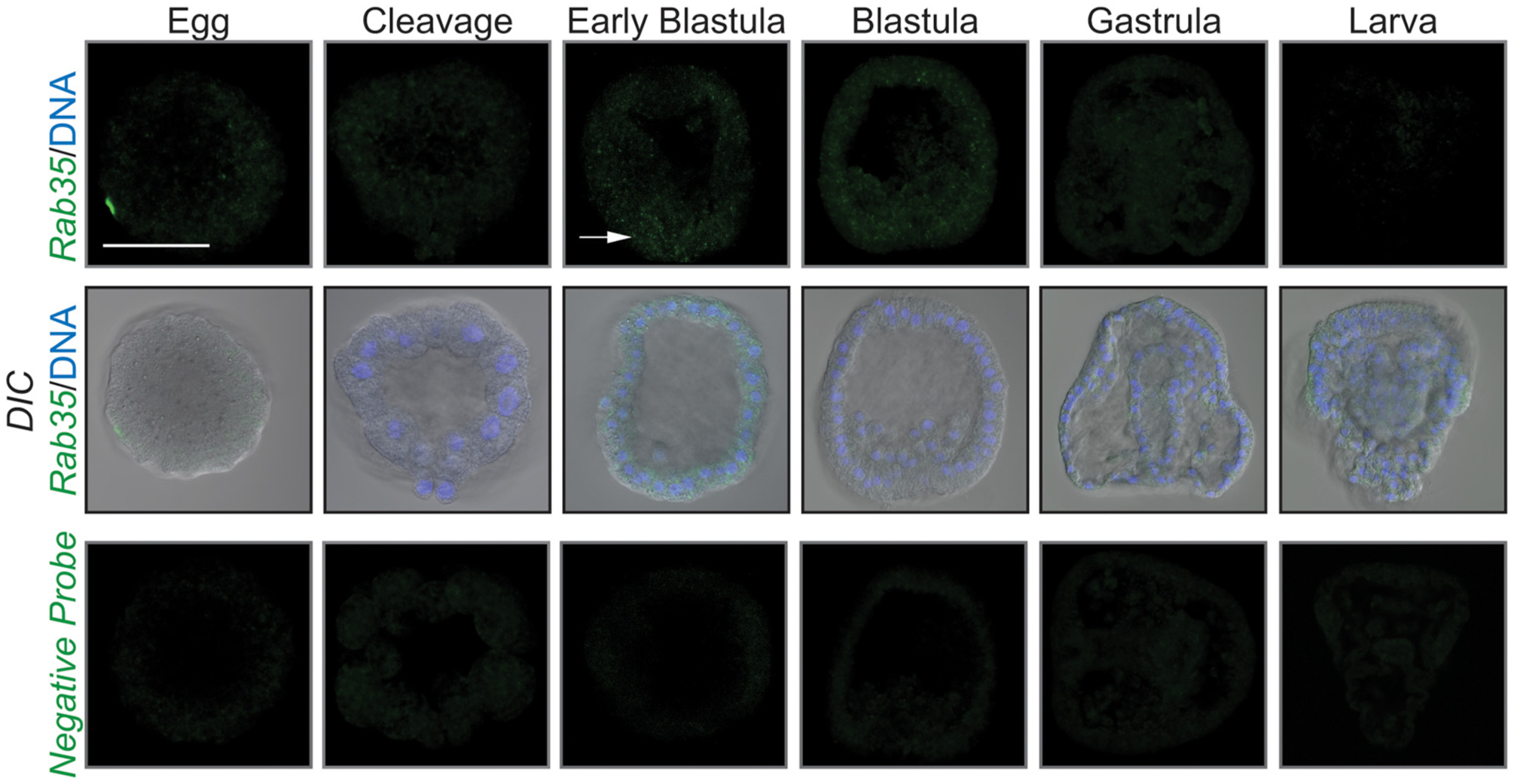
Rab35 is expressed at low levels during early development. Rab35 is expressed at a low level during early development. White arrow indicates area of expression in early blastula. Maximum intensity projections of confocal images obtained after WMFISH are shown. Scale bar is 50 μm. 3 biological replicates.
2.2. Rab35 is essential for gastrulation and skeletogenesis
In order to determine the function of Rab35 during development, a translation-blocking morpholino (MASO) was used to knock down Rab35 in the embryo. Results indicate a dose-dependent severity of shortened DVCs and gastrulation defects (Fig. 2). The defects in gastrulation ranged from a gut that does not extend to the anterior end of the embryo, a wider gut, to exogastrulation (Fig. 2B). These spicule and gastrulation defects are rescued by co-injection of the Rab35 MASO with SpRab35, suggesting that these defects are specific to Rab35 KD (Fig. 3A). However, the HsRab35 does not rescue the spicule defect. A bioinformatic analysis of human and sea urchin Rab35 protein sequences indicate that 81% of the residues are identical and 88% of the residues contain similar functional groups (Fig. S1). In order to determine whether the shortened DVCs were due to a defect in the patterning of the PMCs, the only cells that make the larval skeleton, control and Rab35 KD embryos were immunolabeled with an antibody (1D5) that recognizes PMCs and skeletal elements (McClay et al., 1983). Rab35 KD embryos exhibit clustered PMCs that do not migrate anteriorly along the embryo (Fig. 3C). Additionally, the Rab35 KD embryos lack intense labeling of the cytoplasmic sheaths that line the skeletal spicule (Fig. 3C) This PMC patterning and labeling defect is rescued by co-injection with either sea urchin or human Rab35 (Fig. 3C). HsRab35 co-injection with Rab35 MASO rescued the gastrulation and PMC patterning defects, indicating a conservation of function of Rab35 between the human and sea urchin Rab35 (Fig. 3).
Fig. 2.
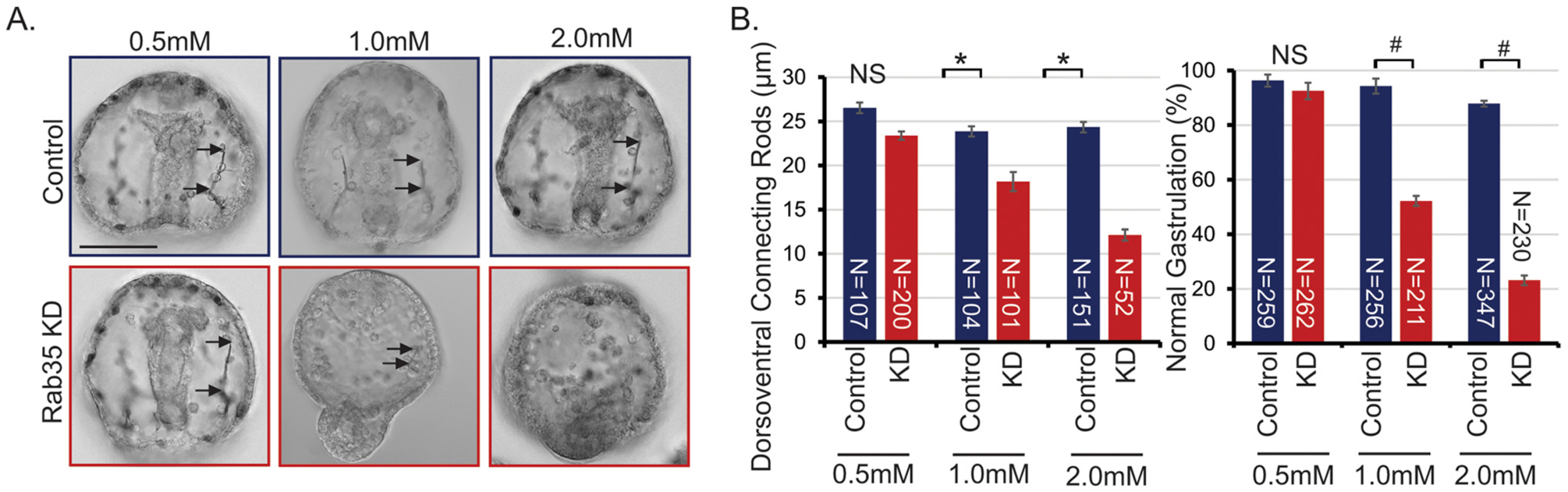
Rab35 KD results in defects in DVC and gastrulation in a dose-dependent manner. (A) Rab35 KD results in dorsoventral connecting rods (DVC) and gastrulation defects in a dose dependent manner. Images are single slices of DIC images. Arrows depict the length of DVCs. Scale bar is 50 μm. (B) Quantification of Rab35 KD defects with different doses of control or Rab35 MASO. 3–5 biological replicates; *p<0.005 using Student’s t-test; #p<0.01 using Cochran-Mantel-Haenszel test; N = total number of embryos examined. All error bars represent standard error of the mean (SEM).
Fig. 3.
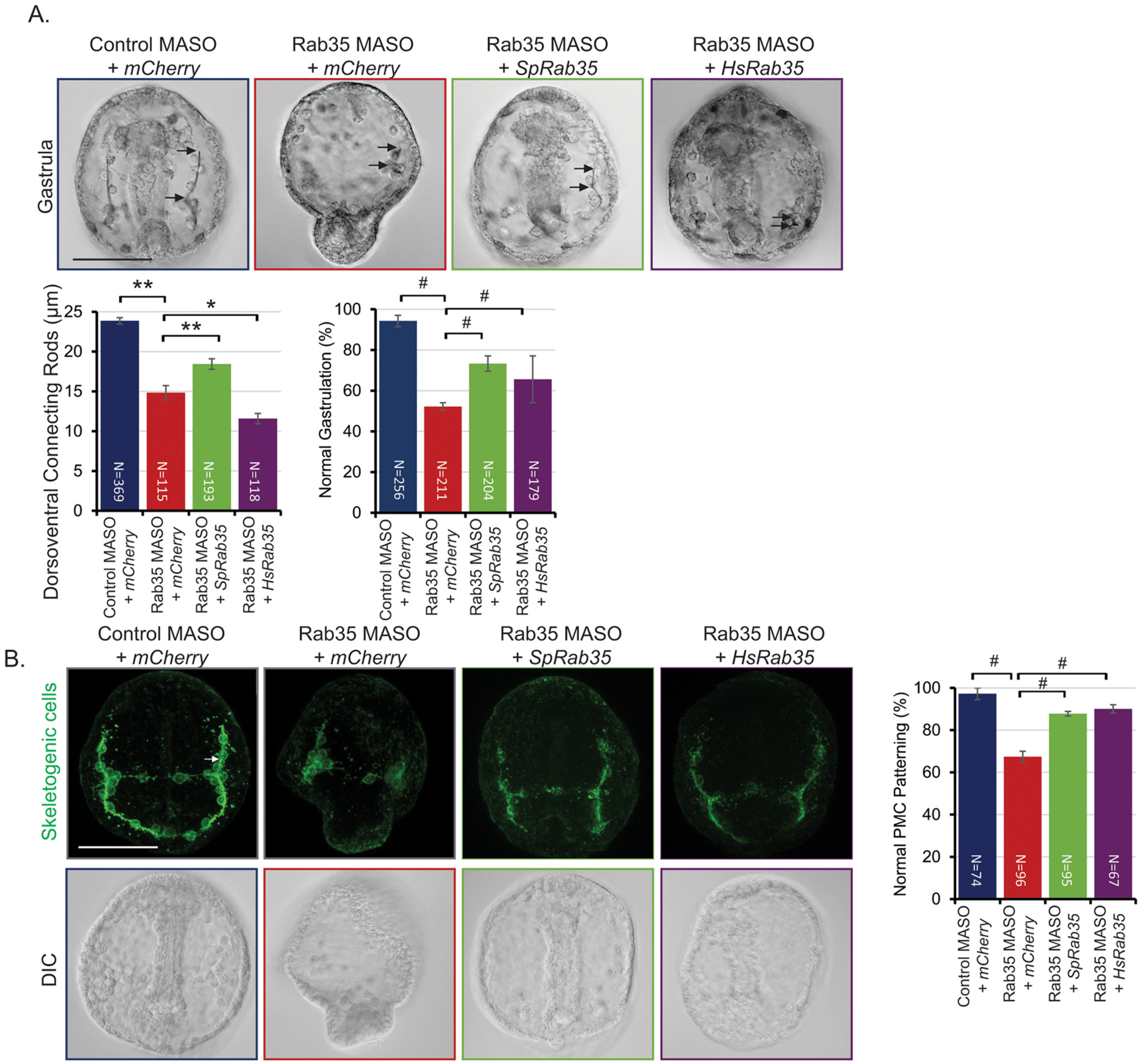
Rab35 KD defects can be rescued by both sea urchin (Sp) and human (Hs) Rab35. (A) Rab35 KD DVC defects are rescued by co-injection with SpRab35, but not HsRab35. However, Rab35 KD gastrulation defects are rescued by co-injection with either SpRab35 or HsRab35. Embryos were injected with 1 mM control or 1 mM Rab35 MASO as depicted; 3 biological replicates; *p<0.001, **p<0.005 using Student’s t-test; #p<0.01 using Cochran-Mantel-Haenszel test; arrows depict the length of DVCs. (B) Rab35 KD results in defects in patterning of PMCs, which is rescued with co-injection of either SpRab35 or HsRab35. Control embryos display PMCs organized in a subequatorial ring around the gut and PMCs migrated anteriorly along the embryo. On the contrary, Rab35 KD embryos exhibit PMCs that remain clustered in the ventrolateral region. Arrows indicate cytoplasmic cables; embryos were immunolabeled with an antibody (1D5) that recognizes PMCs and skeletal elements; #p<0.01 using Cochran-Mantel-Haenszel test. All error bars represent SEM. The scale bar is 50 μm.
2.3. Rab35 mediates vesicular trafficking in the early embryo
One of the important functions of Rab35 is to mediate plasma membrane to early endosome protein trafficking (Klinkert and Echard, 2016). In order to test Rab35’s role in vesicular trafficking during early development, we used FM1–43 to visualize vesicular trafficking. FM1–43 is an amphiphilic molecule consisting of a lipophilic tail linked to a positively charged head through a double bond bridge (Brumback et al., 2004). It contains a polar dicationic head, preventing it from diffusing across the membrane, so any cytoplasmic fluorescence is due to the presence of membrane bound vesicles (Brumback et al., 2004). Control and Rab35 KD embryos were exposed to FM1–43 for 20 min and imaged in its presence. The control embryos have a higher mean fluorescence of FM1–43 than the Rab35 KD blastulae (Fig. 4A). Numerous small vesicles are observed in control blastulae, but these are substantially reduced in Rab35 KD blastulae (Fig. 4A). In addition, both the control and Rab35 KD embryos contain multiple larger vesicles; however, the Rab35 KD embryos have significantly fewer of these larger vesicles (Fig. 4A). These results suggest that Rab35 regulates vesicular trafficking during early development.
Fig. 4.

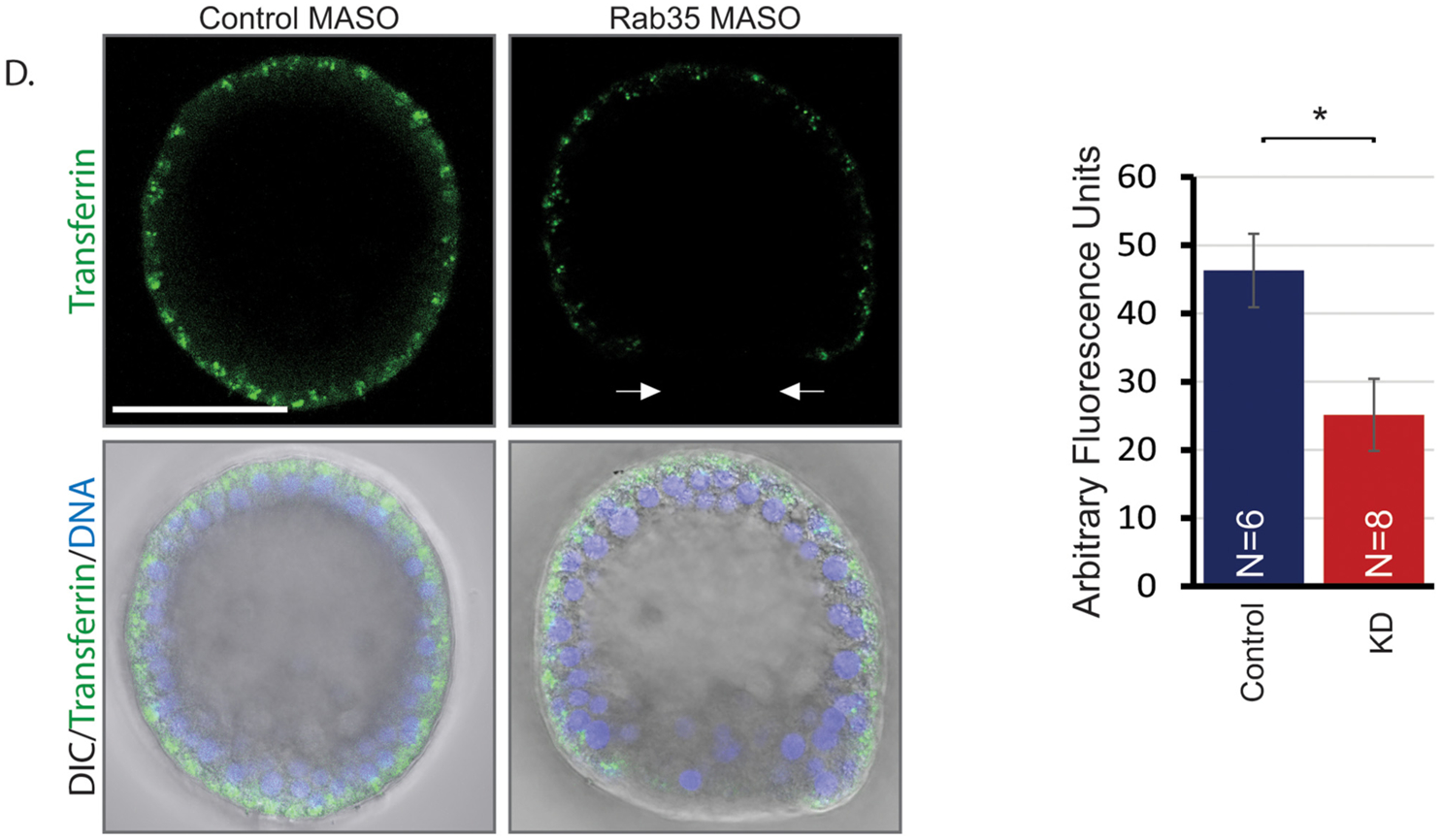
Rab35 mediates vesicular trafficking during early development. (A) Zygotes were injected with mCherry-LifeAct (channel not shown) and incubated with FM1–43. Control blastula stage embryos (24 hpf) contain a higher number of large vesicles and more FM1–43 fluorescence after incubation with FM1–43 than Rab35 KD embryos. Images in top panel are maximum intensity projections of 5 slices of confocal images. Images in bottom panel are enlarged maximum intensity projections of 5 slices that span 7 μm to capture overall vesicular content within a single cell (inset of the top panel). White arrow indicates large vesicle, and the red arrow indicates small vesicle. Scale bar is 50 μm. N = total number of embryos measured. *p<0.05 using Student’s t-test. 2 biological replicates; 5 embryos/replicate. (B) Fluorescence in control blastula stage embryos is lost more quickly, while fluorescence in Rab35 KD embryos is retained for longer amount of time. t = 0 indicates time immediately after FM1–43 wash out. 2 biological replicates; 1–2 embryos/condition. Images are single slices of confocal images at the same focal plane. *Indicates one of the several ingressed PMCs in the blastocoel. Scale bar is 50 μm. (C) Graph represents fluorescence retained normalized to initial fluorescence at t = 0; fluorescence measured at different optical planes throughout the embryo and averaged. Error bars represent SEM. (D) After 15 min of incubation with Alexa 488-conjugated transferrin, control embryos had a higher level of fluorescence than Rab35 KD embryos. Images are single slices of confocal images at the same focal plane. Arrows indicate area of lack of transferrin in Rab35 KD embryos; *p<0.05 using Student’s t-test. N=total number of embryos examined. Error bars represent SEM. Scale bar is 50 μm.
To determine if endocytosis and exocytosis are regulated by Rab35, control and Rab35 KD blastulae were exposed to FM1–43 for 3–4 h to saturate their membrane, followed by FM1–43 washout and immediate imaging (Fig. 4B). Results indicate that the control embryos retain 55% of their total fluorescence, while the Rab35 KD embryos retain 75% of their total fluorescence after 34 min of washout (Fig. 4C). Throughout the washout time course, the Rab35 KD embryos consistently retained significantly more total fluorescence compared to the control. These results suggest that Rab35 regulates exocytosis or vesicular recycling during early development.
As FM1–43 reports all types of endocytosis, we used Alexa 488-conjugated transferrin to examine the clathrin-mediated endocytic pathway, which Rab35 is known to regulate (Dikshit et al., 2015; Dutta and Donaldson, 2015). After incubation with transferrin for 15 min, embryos were placed in FSW for immediate imaging. Results indicate that control embryos have significantly higher fluorescence than Rab35 KD embryos, indicating that Rab35 mediates the clathrin-mediated endocytosis pathway (Fig. 4D). Additionally, in Rab35 KD blastula, the posterior end of the embryo, where invagination occurs, appears to lack transferrin.
2.4. Rab35 remodels actin to mediate gastrulation
Since Rab35 has actin remodeling function, we examined how Rab35 KD would affect actin distribution. In order to determine the role of actin localization mediated by Rab35 during gastrulation, embryos were injected with control or Rab35 MASO with LifeAct-mCherry, which encodes a 17 amino acid peptide that binds to F-actin (Riedl et al., 2008). Live imaging was conducted at early gastrula and gastrula stages (Fig. 5A). In control early gastrulae, the F-actin is observed along the apical surface of the entire embryo, while in the Rab35 KD embryos, the actin coalesces into foci at the basal surface of cells in the posterior end of the embryo, where gut invagination initiates during gastrulation (Fig. 5A). This suggests that the actin remodeling function of Rab35 may be important to localize actin at the apical surface of posterior cells to provide the mechanical strength for initial gut invagination. As the embryo continues the process of gastrulation, actin in control gastrulae becomes intensely localized to the apical surface of the gut epithelial cells that are undergoing gastrulation (Fig. 5A). In contrast, Rab35 KD embryos fail to gastrulate and lack this actin localization; instead, they display a more diffuse organization of actin. Additionally, actin is enriched in the skeletal spicule of control gastrula; while in Rab35 KD gastrula, the spicule shows little to no F-actin (Fig. 5B). The size and shape of the cells are more variable in the Rab35 KD embryos, compared to the control embryos (Fig. 5A), particularly in the posterior end of the embryo, indicating that Rab35 may regulate cell shape and size. Overall, these results indicate that actin remodeling, specifically at the site of invagination and the skeletal spicule, mediated by Rab35 is important for the initiation and maintenance of gastrulation, as well as skeletogenesis.
Fig. 5.
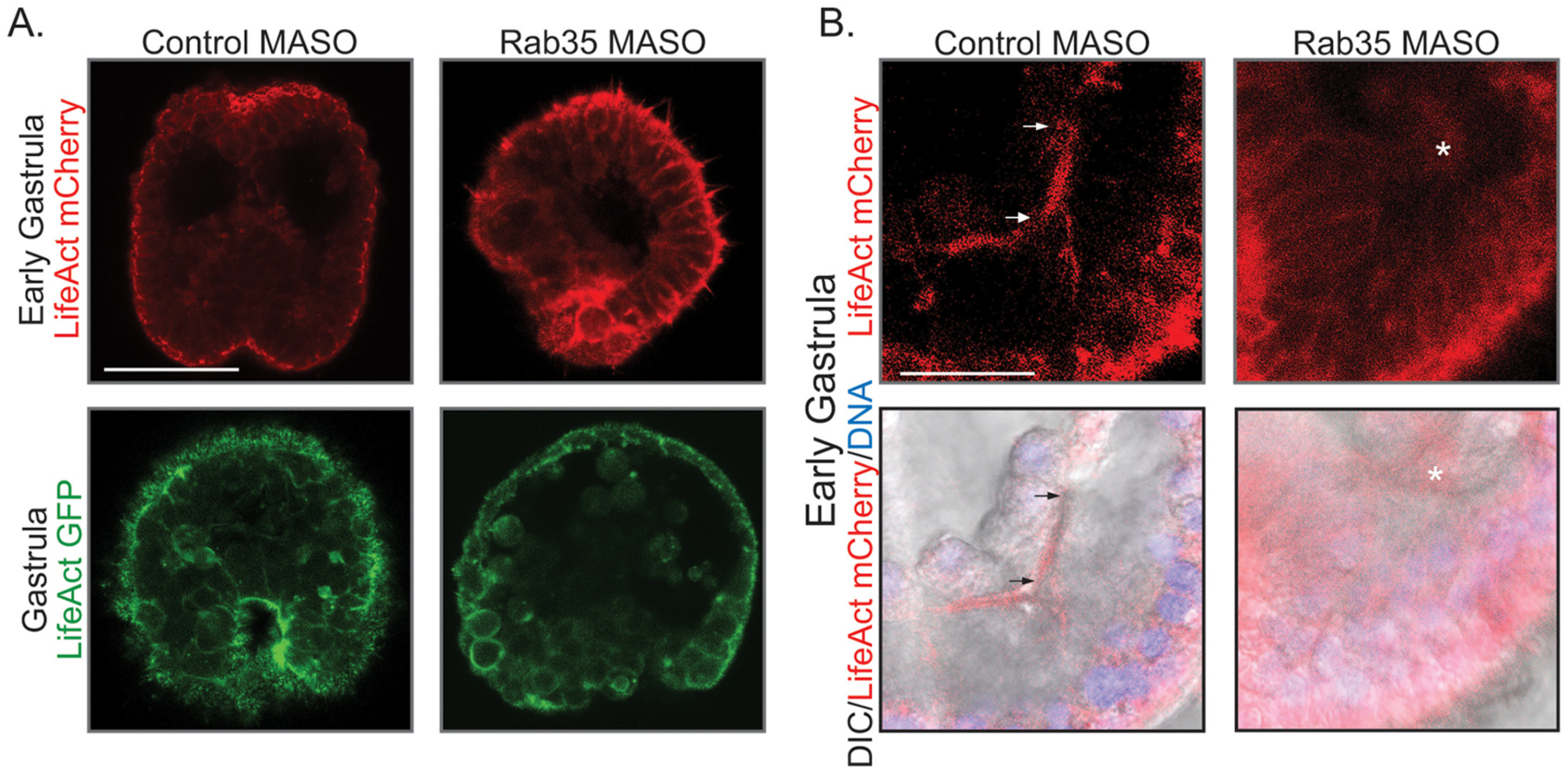
Rab35 may mediate gastrulation through its actin remodeling function. (A) Control early gastrula stage embryos (24 hpf) exhibit enriched actin at the apical plasma membrane, including at the site of invagination. Rab35 KD embryos develop punctate foci of actin at the site of gastrulation. 2 biological replicates; 4–5 embryos/replicate; Images are maximum intensity projections of 4 slices of confocal images. Scale bar is 50 μm. Control gastrula stage embryos (48 hpf) exhibit intense actin localization on the apical surface of the invaginating epithelial endodermal cells. Rab35 KD embryos have a more diffuse actin localization. 4–5 embryos in one replicate. Images are single slices of confocal images. Scale bar is 50 μm. (B) Control embryos exhibit ordered actin in the skeletal spicule, while Rab35 KD embryos lack actin in the skeletal spicules. Arrows indicate skeletal spicule. No skeletal spicule was observed in the Rab35 KD embryo, asterisk indicates LifeAct within a presumed PMC. Images are single slices of confocal images. Scale bar is 25 μm.
2.5. Rab35 interacts with Fascin to mediate skeletogenesis and gastrulation
In Drosophila, Rab35 interacts with actin bundling protein Fascin to mediate bristle formation (Zhang et al., 2009), and Rab35 RNAi or expression of DN-Rab35 led to kinked bristles which were rescued by Fascin overexpression. Therefore, we reasoned that the skeletogenesis and gastrulation defects caused by Rab35 KD is at least in part due to its interaction with Fascin. To test this hypothesis, newly fertilized eggs were co-injected with Rab35 MASO and SpFascin. Results indicate that Fascin is able to rescue the shortened DVC, PMC mispatterning, and gastrulation defects (Fig. 6).
Fig. 6.
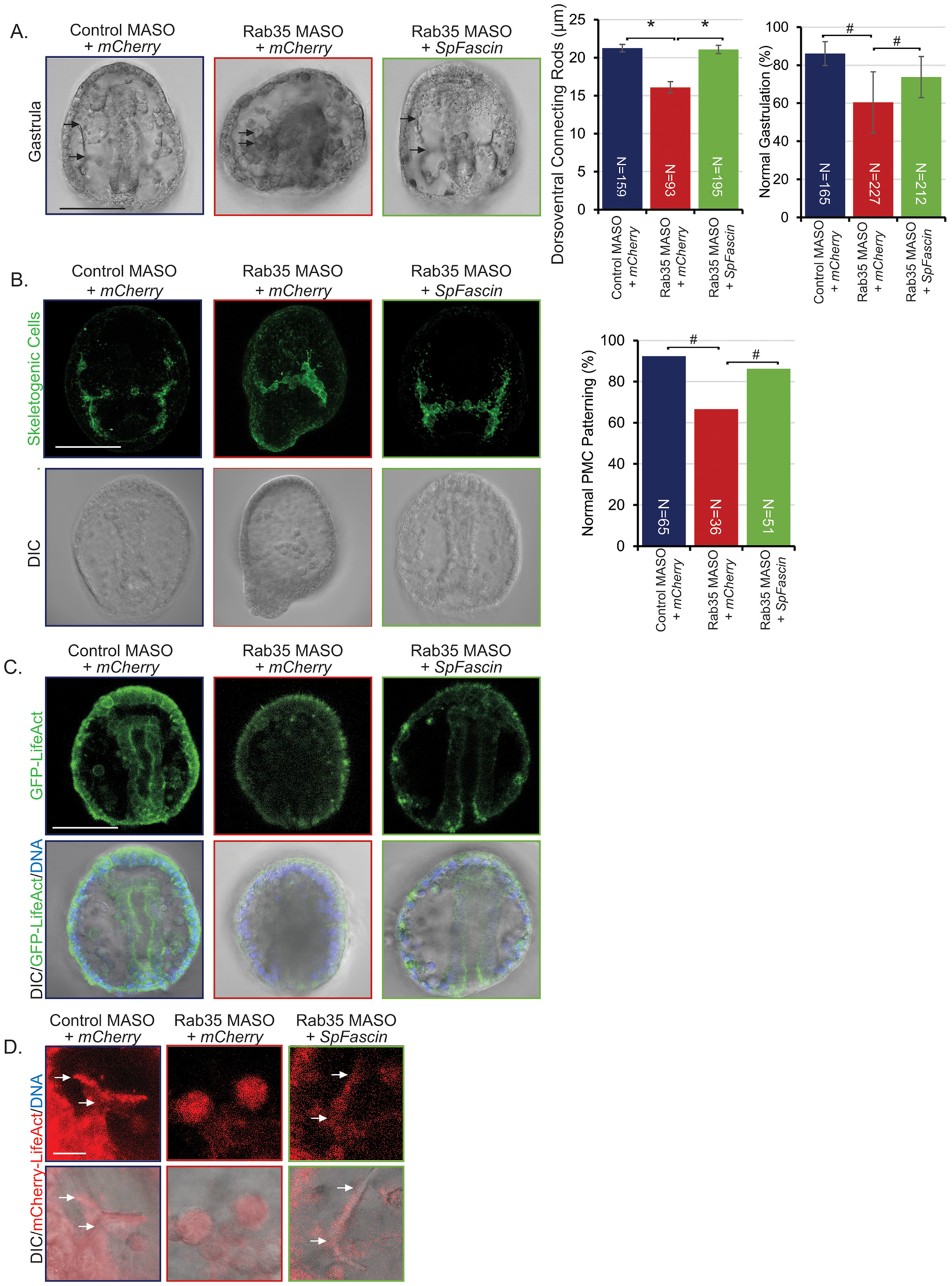
Rab35 KD is rescued by SpFascin. (A) Shortened DVC and gastrulation defects are rescued by co-injection with SpFascin. Embryos were injected with 1 mM control MASO or 1 mM Rab35 MASO as depicted; 3 biological replicates *p<0.001, **p<0.005 using Student’s t-test; error bars represent SEM. #p<0.001 using Cochran-Mantel-Haenszel test. Error bars represent SEM. (B) PMC patterning defects are rescued by co-injection with SpFascin. 6 embryos in one replicate. #p<0.001 using Fisher’s exact test. Error bars represent SEM. (C) Rab35 KD induced actin remodeling defects are rescued by co-injection with SpFascin. Scale bar is 50 μm. (D) Rab35 KD induced actin remodeling defects in skeletal spicules are rescued by co-injection with SpFascin. Arrows indicate skeletal spicule. No skeletal spicule was observed in the Rab35 KD embryo. Image of Rab35 KD embryo depicts where the presumptive skeletal spicule would be located. Fluorescent images are maximum intensity projections of 3 slices of confocal images. Scale bar is 10 μm.
To examine the distribution of F-actin, newly fertilized embryos were injected with control or Rab35 MASO and LifeAct-GFP with or without SpFascin. Results indicate that SpFascin is able to rescue actin localization at the site of invagination, which is important to provide the enriched actin necessary for successful gastrulation (Fig. 6C). Interestingly, SpFascin is able to rescue actin localization to the site of the skeletal spicule as well, although it does not seem to have the same linear actin organization as observed in the control (Fig. 6D). Results from this set of experiments indicate a functional interaction between Rab35 and Fascin in mediating actin localization and distribution, critical for skeletogenesis and gastrulation.
3. Discussion
Although Rab35 is not highly expressed during development, it is a critical regulator of skeletogenesis and gastrulation (Figs. 1, 2). Through its regulation of vesicular trafficking and actin dynamics (Figs. 4–6), Rab35 mediates cell shape change and motility, which are essential in the formation of embryonic structures, including the larval skeleton and gut. Importantly, many of the defects induced by Rab35 KD can be rescued by human Rab35 or overexpression of SpFascin, indicating that the function of Rab35 and its interaction with Fascin are evolutionarily conserved (Figs. 3, 6) (Zhang et al., 2009).
Although previous developmental transcriptomes based on RNA-Seq result have shown that Rab35 has the highest expression at the early cleavage stage 10 hpf (Tu et al., 2014), our FISH data indicate that, while not abundant, its expression peaks at early blastula stage (Fig. 1). This discrepancy may be due to the lower sensitivity and need for transcripts to be locally concentrated to be visualized in FISH compared to RNA-Seq.
The shortened DVCs observed in Rab35 KD embryos are likely due to perturbed patterning of the PMCs and Rab35’s potential role in biomineralization (Fig. 3B). The PMC patterning defects are rescued by co-injection with either sea urchin or human Rab35, indicating that Rab35 is functionally conserved between the sea urchin and human (Fig. 3B). However, the shortened DVCs observed in Rab35 KD embryos are not rescued by co-injection with HsRab35. This result of normal patterning of the PMCs, but shorter crystalline spicules has also been observed previously in the context of perturbation of P58A, P58B, and P16, which encode transmembrane proteins that are involved in biomineralization (Adomako-Ankomah and Ettensohn, 2011; Alvares et al., 2016; Cheers and Ettensohn, 2005). This suggests that sea urchin Rab35 may be responsible for biomineralization of the skeletal spicules that the human Rab35 cannot perform. The calcium that forms the spicule ultimately comes from the seawater and enters the PMCs both through calcium channels, as well as endocytosis (Morgulis et al., 2019; Stumpp et al., 2012; Vidavsky et al., 2016). It is possible that sea urchin Rab35 effectors responsible for calcium endocytosis are unable to bind to human Rab35. Most effectors bind to Rab35 at the ‘Switch’ and “Interswitch” regions. The ‘Interswitch’ region within the sea urchin Rab35 contains a charged lysine, whereas the human Rab35 contains a non-charged asparagine that could affect protein-protein interactions (Kuo and Zhang, 2019) (Fig. S1).
The shortened DVCs appear to be worsened by co-injection of HsRab35 with Rab35 MASO. If human Rab35 is unable to interact with sea urchin Rab35 effectors, the abundance of human Rab35 in the rescue embryos may compete for and sequester the endogenous sea urchin Rab35 GEFs and GAPs, preventing residual sea urchin Rab35 from inducing biomineralization. In essence, human Rab35 could be acting as a dominant-negative form of the protein, resulting in less biomineralization.
As a small GTPase, Rab35 plays a role in vesicular trafficking (Klinkert and Echard, 2016). We tested vesicular trafficking by assaying for FM1–43, which reports all types of endocytosis. Our results indicate that Rab35 KD embryos have significantly fewer large vesicles than control embryos (Fig. 4A). This is consistent with previous research in Drosophila, where a mutation in a Rab35 GAP, which promotes conversion of active to inactive Rab35, resulted in a higher number of large FM1–43 positive vesicles at the neuromuscular junction (Uytterhoeven et al., 2011). We also observed less FM1–43 mean fluorescence in the Rab35 KD embryos compared to the control, although this was not significant. This is likely due to the majority of the FM1–43 residing in the cell membranes, not in the vesicles themselves, leading to an under-representation of the true difference between control and KD embryos.
Our results indicate that Rab35 KD embryos retain fluorescent FM1–43 at a higher level than that of control embryos. This is consistent with previous research in which loss of function of Rab35 results in decreased exocytosis (Biesemann et al., 2017). It is possible that the increased retention of fluorescence is due to impaired vesicular recycling or it could be due to perturbed exocytosis, both of which would result in increased retention of fluorescence. The patterning of PMCs is thought to be dependent upon secretion of Vegf3 ligand from the ectoderm (Duloquin et al., 2007; Ettensohn and Adomako-Ankomah, 2019). Rab35 may regulate exocytosis of Vegf3 from the ectodermal cells, resulting in defects in PMC patterning (Fig. 3B). While very little is known about the secretion of Vegf ligands, the secretion of one isoform from COS-7 and HUVEC cell lines is dependent upon the presence of the phosphoinosotide PI(4,5)P2, a lipid which interacts with Rab35 (Guzmán-Hernández et al., 2014; Klinkert and Echard, 2016).
Rab35 is known to play a key role in actin localization (Chaineau et al., 2013; Chua et al., 2010; Klinkert and Echard, 2016; Zhang et al., 2009). Rab35 localizes actin remodeling proteins, such as Rac1 and Cdc42, which have been shown to be important in the sea urchin for skeletogenesis (Sepúlveda-Ramírez et al., 2018; Shim et al., 2010). The defects observed in DVCs and PMC patterning after Rab35 KD may be due to the inability of Rac1 and Cdc42 to properly localize actin, resulting in lack of cytoplasmic cables and improper cell motility (Fig. 3).
The role of vesicular trafficking during gastrulation is not fully understood. One model for gastrulation suggests that apical constriction of the cells at the vegetal plate results in bending forces that may cause primary invagination (Kominami and Takata, 2004; McClay et al., 2020; Takata and Kominami, 2004). Germ band extension in Drosophila embryos, analogous to gastrulation, involves contraction of acto-myosin filaments at the apical surface of the cell that is regulated by Rab35-mediated endocytosis (Jewett et al., 2017). Rab35 KD embryos exhibit fewer endocytic vesicles (Fig. 4A, B), and the transferrin uptake (indicative of clathrin-mediated endocytosis) is much less prominent in the posterior end of the embryo (Fig. 4D). Therefore, the lack of endocytosis may result in the failure of apical constriction in the posterior end of the embryo, ultimately resulting in the inability of Rab35 KD embryos to gastrulate properly. In addition, the cells of Rab35 KD embryos appear to have more variable size and shape, particularly in the posterior end of the embryo, where cell shape change is critical in the formation of the embryonic gut (Fig. 5A).
Actin dynamics also play a role during initiation of gastrulation; actin-mediated cell shape changes are required for primary invagination (Nakajima and Burke, 1996). This apical constriction may be due to RhoA, a small GTPase, which is known to regulate cytoskeletal rearrangements (Beane et al., 2006). Previously, actin has been shown to be present on the basal surface of the cells of the embryo and becomes enriched on the apical surface of the cell during PMC ingression (Anstrom, 1992). Actin also resides just below the apical surface of the cell that form actin-based microvilli (Bonder and Fishkind, 1996). Using LifeAct to examine actin in live embryos, we observed that in control embryos, actin is enriched on the apical surface of the cell, as well as on the apical surface of the endoderm lining the gut (Fig. 5A). This lack of actin enrichment in the Rab35 KD gastrulating embryos may result in lack of mechanical stiffness at the posterior end of the embryo where invagination occurs, potentially leading to the exogastrulation phenotype. Overall, our results have shown that Rab35 is critical for proper gastrulation through localization of actin to the posterior end of the embryo and enrichment on the apical surface of the cell. We have also shown that Rab35’s role in vesicular trafficking may regulate gastrulation, by ensuring uptake of plasma membrane to mediate apical constriction.
We observed that overexpression of Fascin after Rab35 KD results in rescue of the skeletogenesis and gastrulation defects (Fig. 6). Fascin acts by binding two F-actin molecules, to form linear actin bundles, such as actin-rich filopodia (Hashimoto et al., 2011; Jayo and Parsons, 2010; Zhang et al., 2009). Additionally, Fascin has been found to play a role in vesicular trafficking (Beghein et al., 2018; Clancy et al., 2018; Nozumi et al., 2017). Inhibition of Fascin’s function results in decreased endocytosis, as well as decreased extracellular vesicle release (Beghein et al., 2018; Clancy et al., 2018). We observed that overexpression of Fascin after Rab35 KD results in more actin localizing to the spicules, but the actin appears less distinct (Fig. 6D). Fascin overexpression is able not only to rescue PMC patterning (Fig. 6B) but is also able to rescue skeletal spicule length (Fig. 6A), suggesting it is rescuing cell motility as well as biomineralization. Rab35 has been shown to functionally interact with Fascin and actin, by localizing both proteins to the plasma membrane and allowing actin to bundle and polymerize to mediate cell motility (Zhang et al., 2009). The PMC patterning defects observed in Rab35 KD embryos may be rescued by the ability of Fascin to localize and bundle actin properly (Zhang et al., 2009). Additionally, as vesicular trafficking is also regulated by interaction between Fascin and Rab35 (Clancy et al., 2018), defects in biomineralization due to defects in endocytosis of calcium in the absence of Rab35 may be rescued by overexpression of Fascin. Fascin may rescue these Rab35 KD developmental defects, by stochastically allowing Fascin to be localized to the proper place at the time of skeletogenesis, rescuing defects in actin dynamics and vesicular trafficking.
We also observed Fascin to rescue gastrulation defects induced by Rab35 KD. Specifically, live imaging with LifeAct reveals rescue of actin localization to the site of invagination during gastrulation in these embryos (Fig. 6C). This suggests that overexpressed Fascin can localize and bundle actin in the absence of Rab35, consistent with previous study that showed Rab35 KD in Drosophila results in abnormal bristle morphology which can be rescued by overexpression of Fascin (Zhang et al., 2009). Fascin overexpression may also regulate changes in cell shape mediated by vesicular trafficking during gastrulation. Rab35 has been shown to mediate endocytosis of plasma membrane that has infolded into vesicles, resulting in apical constriction during GBE in Drosophila (Jewett et al., 2017). Since Fascin and Rab35 interact to mediate vesicular trafficking (Clancy et al., 2018), overexpression of Fascin in Rab35 KD embryos may regulate uptake of infolded plasma membrane, resulting in apical constriction during gastrulation. Previous studies and our results strongly indicate that the interaction between Rab35 and Fascin is evolutionarily conserved.
4. Conclusions
Overall, this study demonstrates that the small GTPase Rab35 plays a key role in skeletogenesis and gastrulation in sea urchin development. Through regulating actin polymerization spatially and temporally, as well as mediating vesicular trafficking, Rab35 controls important cellular processes such as cell motility and apical constriction (Fig. 7). These processes that are regulated by Rab35 are important for the formation of the larval skeleton and gut. We have also shown that Rab35’s function in these processes is evolutionarily conserved, as the human Rab35 can functionally rescue most defects in Rab35 KD embryos. The interaction between Rab35 and Fascin is also evolutionarily conserved. This study revealed that Rab35 is an essential regulator of key developmental processes, providing us a deeper understanding of how vital embryonic structures are formed.
Fig. 7.
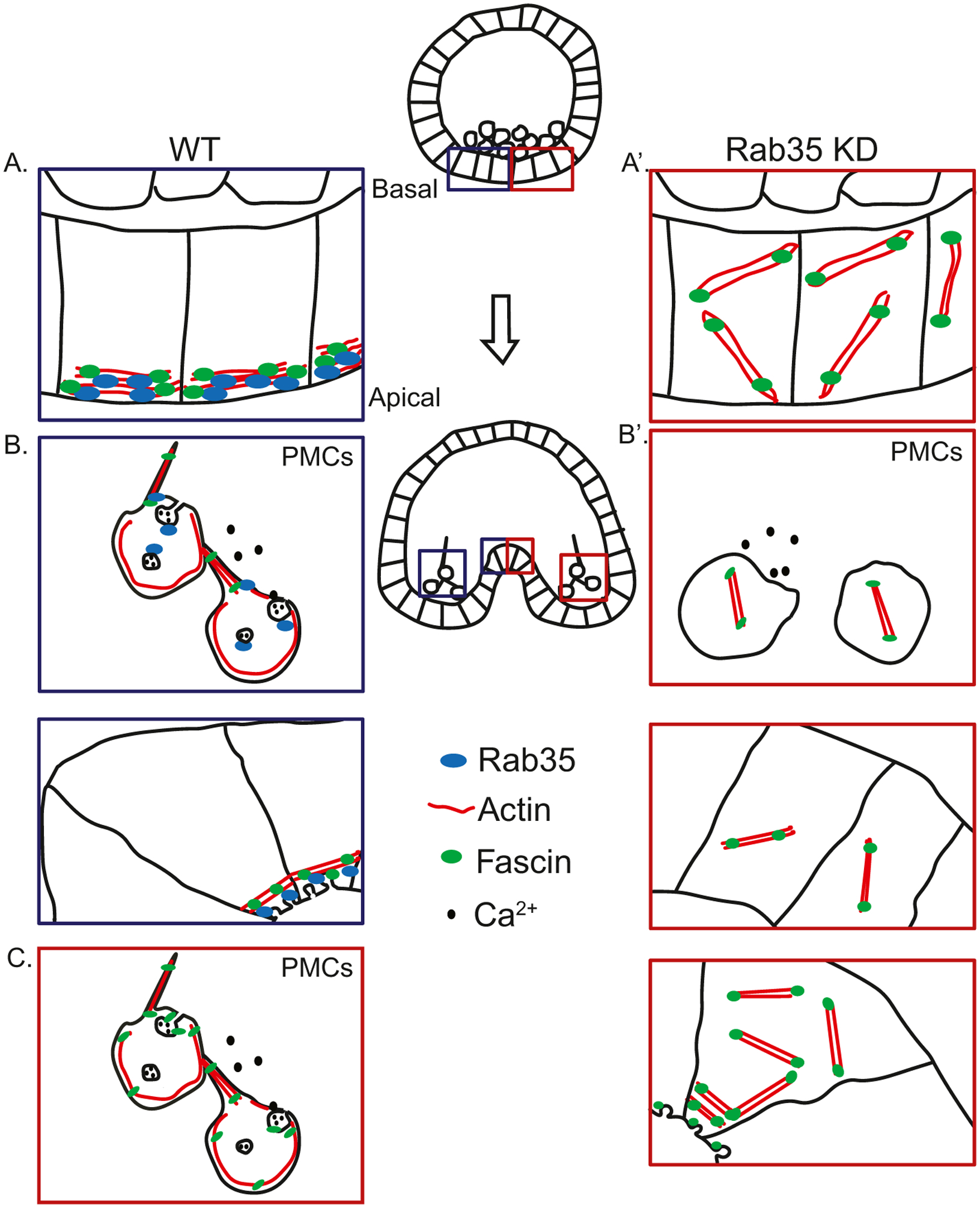
Working of model of the function of Rab35 in skeletogenesis and gastrulation. (A) During early gastrulation, Rab35 localizes Fascin and actin to the apical surface of the cell membrane. (A′) Rab35 KD embryos exhibit disorganized actin due to lack of Rab35 to localize actin. (B) In the PMCs, Rab35 localizes Fascin and actin to the filopodia, and mediates vesicular uptake of calcium to build the mineralized skeleton in PMCs. As invagination occurs, the localized actin constricts, and Rab35 and Fascin mediate uptake of excess apical plasma membrane into vesicles, causing apical constriction. (B′) In Rab35 KD embryos, the PMCs lack actin at the filopodia, due to the failure of available Rab35 to organize actin filaments, and cell motility is diminished, resulting in a lack of anterior migration of the PMCs, while the lack of endocytosis results in defects in biomineralization. At the site of gut invagination, the lack of Rab35 results in failure of apical constriction, leading to gastrulation defects. (C) Overexpression of Fascin (Fascin OE) in a Rab35 KD embryo results in stochastic bundling of actin at the plasma membrane and formation of filopodia to rescue the phenotype. Additionally, excess Fascin induces endocytosis of calcium in PMCs to allow biomineralization to occur, and uptake infolded membrane at the site of invagination to cause apical constriction, resulting in gastrulation.
5. Materials and methods
5.1. Animals
Adult sea urchins, Strongylocentrotus purpuratus, were obtained from Point Loma Marine Invertebrate lab (Lakeside, CA) and were maintained at 12 °C in artificial sea water (ASW) made from distilled water and Instant Ocean©. Adults were induced to shed either through shaking or intracoelomic injection of 0.5 M KCl. Embryos were cultured at 12 °C in filtered sea water (FSW) obtained from the Indian River Inlet (University of Delaware).
5.2. Whole mount fluorescence RNA in situ hybridization
To examine the localization of Rab35 transcripts, we used WMFISH. Rab35 was amplified from sea urchin cDNA using Phusion High Fidelity DNA polymerase (ThermoScientific, Waltham, MA) using primers in Table 1, and then cloned into ZeroBlunt (ThermoScientific, Waltham, MA). After linearizing the plasmid with BamHI (ThermoScientific, Waltham, MA), digoxigenin labeled RNA in situ probes were transcribed using the Digoxigenin RNA labeling kit (MilliporeSigma, St. Louis, MO) with Sp6 polymerase. WMFISH was performed as previously described (Sethi et al., 2014), using 0.3 ng/μL probe in hybridization buffer, without the use of yeast tRNA. The probe was detected using anti-digoxigenin-POD antibody (MilliporeSigma, St. Louis, MO) and exposed to TSA Plus amplification system (PerkinElmer, Waltham, MA) for 6 min. Embryos were counterstained with Hoechst 33342 (Lonza. Basel, Switzerland) at 1:1000 dilution in MOPS buffer (0.1M MOPS pH 7, 0.5 M sodium chloride, 0.1% Tween 20, and DEPC-treated water), then imaged using Zen software on a Zeiss LSM 880 confocal microscope and (Zeiss, White Plains, NY).
Table 1.
PCR primers.
| Sequence (5’ to 3’) | |
|---|---|
| Rab35 in situ forward | ATGGCGAGGGAATACGATCA |
| Rab35 in situ reverse (Sp6 binding site) | TATTTAGGTGACACTATAGTCATTTACCGCAGCATTTTC |
| SpRab35 CDS forward (Sp6 binding site) | ATTTAGGTGACACTATAGATGAATTTAAAATACAAATTTGG |
| SpRab35 CDS reverse | TTAGAATTCCCACAGTGTG |
| HsRab35 CDS forward (Sp6 binding site) | ATTTAGGTGACACTATAGATGGCCCGGGACTAC |
| HsRab35 CDS reverse | TTAGCAGCAGCGTTTCTTTCGTTA |
5.3. Constructs
Strongylocentrotus purpuratus (Sp) Rab35 and Fascin coding sequence (CDS) were determined from information available in the sea urchin genome database, version 4.2 (Echinobase.org) (Cameron et al., 2009; Kudtarkar and Cameron, 2017). PCR primers were designed to amplify the CDS from sea urchin cDNA (Table 1). The PCR amplicons were ligated into the ZeroBlunt vector (ThermoFisher, Waltham,MA), and DNA sequenced (Genewiz, South Plainfield, NJ). The Homo sapiens Rab35-GFP was obtained as a kind gift from Dr. Ramona Neunuebel (University of Delaware). Primers were designed to amplify the human Rab35 (HsRab35) without the GFP for the Rab35 KD rescue experiments (Table 1).
The SpRab35, HsRab35, and SpFascin constructs were linearized with XbaI, BamHI, and XbaI (ThermoScientific, Waltham, MA), respectively. After gel purification (Macherey-Nagel, Bethlehem, PA), the HsRab35 and SpRab35 constructs were in vitro transcribed with T7 mMESSAGE mMachine kit (ThermoFisher, Waltham, MA). The SpFascin construct was in vitro transcribed with Sp6 mMESSAGE mMachine kit (ThermoFisher, Waltham, MA). The mCherry-LifeAct and GFP-LifeAct constructs (a kind gift from Dr. Charles Shuster, New Mexico State University) (Sepúlveda-Ramírez et al., 2018) were linearized with NotI and in vitro transcribed with Sp6 mMESSAGE mMachine kit (ThermoFisher, Waltham, MA). mRNA was purified using NucleoSpin RNA clean up kit (Macherey-Nagel, Bethlehem, PA), and passed through a Millipore Ultrafree 0.22 μm centrifugal filter (MilliporeSigma, St. Louis, MO) prior to microinjections. SpRab35 and HsRab35 were injected at a final concentration of 1.2 μg/μL, and SpFascin was injected at a final concentration of 1.5 μg/μL. A control mCherry RNA was used at equivalent concentrations (Yajima and Wessel, 2012).
5.4. Microinjections
Microinjections were performed as described previously (Cheers and Ettensohn, 2004; Stepicheva and Song, 2014). All injection solutions contained 20% sterile glycerol and 2 mg/ml 10,000 MW neutral Texas Red or FITC anionic dextran (ThermoFisher, Waltham, MA) to mark the microinjected embryos.
Rab35 or control morpholino (MASO) were obtained from Gene-Tools (Philomath, OR). The Rab35 MASO (5′ CAGGTGATCGTATTCCCTCGCCATG 3′) was designed to be complementary to the 5′ region of the Rab35 mRNA spanning the start codon. The sequence was determined to be unique to SpRab35 by BLASTN of the sea urchin genome database. The control MASO (5′ CCTCTTACCTCAGTTACAATTTATA 3′) targets a human β-globin gene and is not complementary to the sea urchin genome (as determined by BLASTN against the sea urchin genome database). The Rab35 and control MASOs were injected at concentrations from 0.1mM-1mM. After it was determined that injection of 1mM Rab35 MASO resulted in approximately 50% normal gastrulation, that dose was used for the rest of the experiments.
When injecting mRNA, the appropriate amount of in vitro transcribed mRNA was lyophilized, then resuspended in 20% sterile glycerol, 2 mg/mL 10,000 MW FITC lysine-charged dextran (ThermoFisher, Waltham, MA), the appropriate amount of MASO, and RNase-free water (ThermoFisher, Waltham, MA). After microinjection, embryos were cultured to the gastrula stage (48 hpf), then fixed in 4% paraformaldehyde (20% paraformaldehyde; Electron Microscopy Sciences, Hatfield, PA) in filtered artificial seawater (Instant Ocean). Embryos were then imaged using AxioVision software and a Zeiss ObserverZ1 with an AxioCam MR3 camera (Zeiss, White Plains, NY).
5.5. Immunolabeling
To examine the patterning of PMCs, embryos were immunolabeled using the 1D5 antibody (a kind gift from Dr. David McClay, Duke University) as previously described (McClay et al., 1983) with minor modifications. After microinjection, embryos were fixed overnight at 4 °C in 4% paraformaldehyde (Electron Microscopy Sciences, Hatfield, PA) in filtered ASW. After washing for 10 min three times in PBS-Tween (PBS containing 0.05% Tween-20 [Bio-Rad, Hercules, CA]), embryos were blocked in 4% sheep serum in PBS-Tween for 1 h at room temperature. Embryos were incubated in 1D5 antibody at 1:100 dilution in 4% sheep serum in PBS-Tween at 4 °C for 36–48 h. They were then washed three times in PBS-Tween and incubated in Alexa 488 conjugated goat-anti-mouse antibody (ThermoFisher, Waltham, MA) at a 1:300 dilution in 4% sheep serum in PBS-Tween for 1 h at room temperature. After washing three times in PBS-Tween, they were counterstained with Hoechst 33342 (Lonza, Basel, Switzerland) at 1:1000 dilution in PBS-Tween for 5 min and washed three more times with PBS-Tween. Embryos were then imaged using Zen software with a Zeiss LSM 880 confocal microscope, or AxioVision software with a Zeiss Observer Z1 fluorescent microscope (Zeiss, White Plains, NY).
5.6. FM1–43 membrane labeling
To examine vesicular trafficking, embryos were injected with 800 ng/μL mCherry LifeAct mRNA with either 1 mM control or Rab35 MASO (GeneTools, Philomath, OR). They were incubated in FSW at 12 °C until early gastrula stage (28 hpf). They were then incubated in 2 μM FM1–43 (ThermoFisher, Waltham, MA), by diluting 1:1000 from 2 mM FM1–43 in ethanol and Hoechst 33342 (Lonza, Basel, Switzerland) at 1:1000 in FSW for 20 min at room temperature and immediately imaged in the presence of FM1–43 (Terasaki and Jaffe, 2004). Images were acquired using Zen software with a Zeiss LSM 880 confocal microscope (Zeiss, White Plains, NY). After imaging, maximum intensity projections were obtained and then ImageJ (Schneider et al., 2012) was used to determine the mean fluorescence of each embryo, as well as used to count the number of large vesicles (>400nm in diameter) in each embryo using the slice closest to the lens that showed distinct cell boundaries.
To examine the dynamics of exocytosis, we first incubated embryos with FM1–43, followed by washing out FM1–43. After microinjections, embryos were incubated in FSW at 12 °C until early blastula stage (20 hpf). They were then incubated in 2 μM FM1–43 (ThermoFisher, Waltham, MA) in FSW for 3 h, and NucBlue Live Cell ReadyProbe (ThermoFisher, Waltham, MA) to label DNA for 30 min (Terasaki and Jaffe, 2004). Embryos were deciliated by incubating in 2X ASW for 2 min, then washed in 2 μM FM1–43 in FSW for 5 min. Embryos were then placed on an imaging dish (MatTek, Ashland, MA) containing FSW and imaged using Zen software with a Zeiss LSM 710 confocal microscope (Zeiss, White Plains, NY). Images were obtained every 4 min for 1 h. The laser power and gain were unchanged throughout the experiment to ensure the fluorescence could be quantified. After imaging, ImageJ was used to quantify the total fluorescence per optical section at each time point. Optical sections were chosen as the top, bottom and middle of each embryo. To calculate the normalized fluorescence, after subtracting the background fluorescence, normalized fluorescence was calculated by dividing the net fluorescence by the net fluorescence at time t = 0.
5.7. Transferrin visualization
To examine clathrin-mediated endocytosis, we used Alexa 488-conjugated transferrin (ThermoFisher, Waltham, MA). After microinjections, embryos were incubated in FSW at 12 °C until blastula stage (26 hpf). The embryos were then incubated in Alexa 488-conjugated transferrin at a working concentration of 25 μg/mL in FSW and NucBlue Live Cell ReadyProbe (ThermoFisher, Waltham, MA) to label DNA for 15 min, according to manufacturer’s instructions. Embryos were then deciliated by incubating in 2X ASW and transferrin for 2 min to immobilize embryos for imaging, then washed in 25 μg/mL Alexa 488-conjugated transferrin in FSW for 5 min. Embryos were then placed on an imaging dish (MatTek, Ashland, MA) containing FSW and imaged with Zen software using a Zeiss LSM 710 confocal microscope (Zeiss, White Plains, NY). The laser power and gain were unchanged throughout the experiment.
Supplementary Material
Acknowledgements
The authors would like to thank David McClay (Duke University) for his kind gift of 1D5 antibody, Charles Shuster (New Mexico State University) for his kind gift of mCherry-LifeAct and GFP-LifeAct plasmids, and Ramona Neunuebel (University of Delaware) for her kind gift of human Rab35 plasmid, as well as her helpful advice and comments on the manuscript. We also thank the two anonymous reviewers for their valuable feedback.
Funding
This work is funded by NSF CAREER (IOS 1553338) to JLS, NIH NIGMS P20GM103446, and Townsend Biotechnology Fellowship to CR.
Footnotes
Supplementary data to this article can be found online at https://doi.org/10.1016/j.cdev.2021.203660.
References
- Adomako-Ankomah A, Ettensohn CA, 2011. P58-A and P58-B: novel proteins that mediate skeletogenesis in the sea urchin embryo. Dev. Biol 353, 81–93. 10.1016/j.ydbio.2011.02.021. [DOI] [PubMed] [Google Scholar]
- Allaire PD, Marat AL, Dall’Armi C, Di Paolo G, McPherson PS, Ritter B, 2010. The Connecdenn DENN domain: a GEF for Rab35 mediating cargo-specific exit from early endosomes. Mol. Cell 37, 370–382. 10.1016/j.molcel.2009.12.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allaire PD, Seyed Sadr M, Chaineau M, Seyed Sadr E, Konefal S, Fotouhi M, Maret D, Ritter B, Del Maestro RF, McPherson PS, 2013. Interplay between Rab35 and Arf6 controls cargo recycling to coordinate cell adhesion and migration. J. Cell Sci 126, 722–731. 10.1242/jcs.112375. [DOI] [PubMed] [Google Scholar]
- Alvares K, Ren Y, Feng JQ, Veis A, 2016. Expression of the invertebrate sea urchin P16 protein into mammalian MC3T3 osteoblasts transforms and reprograms them into “osteocyte-like” cells. J. Exp. Zool. Part B Mol. Dev. Evol 326, 38–46. 10.1002/jez.b.22663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Annunziata R, Perillo M, Andrikou C, Cole AG, Martinez P, Arnone MI, 2014. Pattern and process during sea urchin gut morphogenesis: the regulatory landscape. Genesis 52, 251–268. 10.1002/dvg.22738. [DOI] [PubMed] [Google Scholar]
- Anstrom JA, 1989. Sea urchin primary mesenchyme cells: ingression occurs independent of microtubules. Dev. Biol 131, 269–275. 10.1016/s0012-1606(89)80058-2. [DOI] [PubMed] [Google Scholar]
- Anstrom JA, 1992. Microfilaments, cell shape changes, and the formation of primary mesenchyme in sea urchin embryos. J. Exp. Zool 264, 312–322. 10.1002/jez.1402640310. [DOI] [PubMed] [Google Scholar]
- Beane WS, Gross JM, McClay DR, 2006. RhoA regulates initiation of invagination, but not convergent extension, during sea urchin gastrulation. Dev. Biol 292, 213–225. 10.1016/j.ydbio.2005.12.031. [DOI] [PubMed] [Google Scholar]
- Beghein E, Devriese D, Van Hoey E, Gettemans J, 2018. Cortactin and fascin-1 regulate extracellular vesicle release by controlling endosomal trafficking or invadopodia formation and function. Sci. Rep 8 10.1038/s41598-018-33868-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biesemann A, Gorontzi A, Barr F, Gerke V, 2017. Rab35 protein regulates evoked exocytosis of endothelial Weibel-Palade bodies. J. Biol. Chem 292, 11631–11640. 10.1074/jbc.M116.773333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonder EM, Fishkind DJ, 1996. Actin-membrane cytoskeletal dynamics in early sea urchin development. Curr. Top. Dev. Biol 10.1016/s0070-2153(08)60225-2. [DOI] [PubMed] [Google Scholar]
- Brumback A, Lieber JL, Angleson JK, Betz WJ, 2004. Using FM1–43 to study neuropeptide granule dynamics and exocytosis. Methods 33, 287–294. 10.1016/j.ymeth.2004.01.002. [DOI] [PubMed] [Google Scholar]
- Cameron RA, Samanta M, Yuan A, He D, Davidson E, 2009. SpBase: the sea urchin genome database and web site. Nucleic Acids Res 37, D750–D754. 10.1093/nar/gkn887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaineau M, Ioannou MS, McPherson PS, 2013. Rab35: GEFs, GAPs and Effectors. Traffic. 10.1111/tra.12096 n/a–n/a. [DOI] [PubMed] [Google Scholar]
- Charrasse S, Comunale F, De Rossi S, Echard A, Gauthier-Rouvière C, 2013. Rab35 regulates cadherin-mediated adherens junction formation and myoblast fusion. Mol. Biol. Cell 24, 234–245. 10.1091/mbc.e12-02-0167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheers MS, Ettensohn CA, 2004. Rapid microinjection of fertilized eggs. Methods Cell Biol 74, 287–310. [DOI] [PubMed] [Google Scholar]
- Cheers MS, Ettensohn CA, 2005. P16 is an essential regulator of skeletogenesis in the sea urchin embryo. Dev. Biol 283, 384–396. 10.1016/j.ydbio.2005.02.037. [DOI] [PubMed] [Google Scholar]
- Chesneau L, Dambournet D, Machicoane M, Kouranti I, Fukuda M, Goud B, Echard A, 2012. An ARF6/Rab35 GTPase Cascade for Endocytic Recycling and Successful Cytokinesis. Curr. Biol 22, 147–153. 10.1016/J.CUB.2011.11.058. [DOI] [PubMed] [Google Scholar]
- Chua CEL, Lim YS, Tang BL, 2010. Rab35 - a vesicular traffic-regulating small GTPase with actin modulating roles. FEBS Lett 584, 1–6. 10.1016/j.febslet.2009.11.051. [DOI] [PubMed] [Google Scholar]
- Clancy JW, Tricarico CJ, Marous DR, D’Souza-Schorey C, 2018. Coordinated regulation of intracellular fascin distribution governs tumor microvesicle release and invasive cell capacity. Mol. Cell. Biol 39 10.1128/MCB.00264-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davidson LA, Koehl MA, Keller R, Oster GF, 1995. How do sea urchins invaginate? Using biomechanics to distinguish between mechanisms of primary invagination. Development 121, 2005–2018. [DOI] [PubMed] [Google Scholar]
- Decker GL, Lennarz WJ, 1988. Skeletogenesis in the sea urchin embryo. Development 103, 231–247. [DOI] [PubMed] [Google Scholar]
- Di Tommaso P, Moretti S, Xenarios I, Orobitg M, Montanyola A, Chang J-M, Taly J-F, Notredame C, 2011. T-Coffee: a web server for the multiple sequence alignment of protein and RNA sequences using structural information and homology extension. Nucleic Acids Res 39, W13–W17. 10.1093/nar/gkr245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dikshit N, Bist P, Fenlon SN, Pulloor NK, Chua CEL, Scidmore MA, Carlyon JA, Tang BL, Chen SL, Sukumaran B, 2015. Intracellular uropathogenic E. coli exploits host Rab35 for iron acquisition and survival within urinary bladder cells. PLOS Pathog 11, e1005083. 10.1371/journal.ppat.1005083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duloquin L, Lhomond G, Gache C, 2007. Localized VEGF signaling from ectoderm to mesenchyme cells controls morphogenesis of the sea urchin embryo skeleton. Development 134, 2293–2302. 10.1242/dev.005108. [DOI] [PubMed] [Google Scholar]
- Dutta D, Donaldson JG, 2015. Sorting of clathrin-independent cargo proteins depends on Rab35 delivered by clathrin-mediated endocytosis. Traffic 16, 994–1009. 10.1111/tra.12302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ettensohn CA, 1990. The regulation of primary mesenchyme cell patterning. Dev. Biol 140, 261–271. 10.1016/0012-1606(90)90076-u. [DOI] [PubMed] [Google Scholar]
- Ettensohn CA, Adomako-Ankomah A, 2019. The evolution of a new cell type was associated with competition for a signaling ligand. PLoS Biol 17, e3000460. 10.1371/journal.pbio.3000460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grant BD, Donaldson JG, 2009. Pathways and mechanisms of endocytic recycling. Nat. Rev. Mol. Cell Biol 10, 597–608. 10.1038/nrm2755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guzmán-Hernández M, Potter G, Egervári K, Kiss J, Balla T, 2014. Secretion of VEGF-165 has unique characteristics, including shedding from the plasma membrane. Mol. Biol. Cell 25. 10.1091/MBC.E13-07-0418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashimoto Y, Kim DJ, Adams JC, 2011. The roles of fascins in health and disease. J. Pathol 224, 289–300. 10.1002/path.2894. [DOI] [PubMed] [Google Scholar]
- Jayo A, Parsons M, 2010. Fascin: a key regulator of cytoskeletal dynamics. Int. J. Biochem. Cell Biol 42, 1614–1617. 10.1016/j.biocel.2010.06.019. [DOI] [PubMed] [Google Scholar]
- Jewett CE, Vanderleest TE, Miao H, Xie Y, Madhu R, Loerke D, Blankenship JT, 2017. Planar polarized Rab35 functions as an oscillatory ratchet during cell intercalation in the Drosophila epithelium. Nat. Commun 8, 476. 10.1038/s41467-017-00553-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimberly EL, Hardin J, 1998. Bottle cells are required for the initiation of primary invagination in the sea urchin embryo. Dev. Biol 204, 235–250. 10.1006/DBIO.1998.9075. [DOI] [PubMed] [Google Scholar]
- Klinkert K, Echard A, 2016. Rab35 GTPase: a central regulator of phosphoinositides and F-actin in endocytic recycling and beyond. Traffic 17, 1063–1077. 10.1111/tra.12422. [DOI] [PubMed] [Google Scholar]
- Klinkert K, Rocancourt M, Houdusse A, Echard A, 2016. Rab35 GTPase couples cell division with initiation of epithelial apico-basal polarity and lumen opening. Nat. Commun 7, 11166 10.1038/ncomms11166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kominami T, Takata H, 2004. Gastrulation in the sea urchin embryo: a model system for analyzing the morphogenesis of a monolayered epithelium. Dev. Growth Differ 46, 309–326. 10.1111/j.1440-169x.2004.00755.x. [DOI] [PubMed] [Google Scholar]
- Kouranti I, Sachse M, Arouche N, Goud B, Echard A, 2006. Rab35 regulates an endocytic recycling pathway essential for the terminal steps of cytokinesis. Curr. Biol 16, 1719–1725. 10.1016/j.cub.2006.07.020. [DOI] [PubMed] [Google Scholar]
- Kudtarkar P, Cameron RA, 2017. Echinobase: an expanding resource for echinoderm genomic information. Database 2017. 10.1093/database/bax074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuo Y-C, Zhang X, 2019. Effector specificity mechanisms of Rab35 GTPase. Structure 27, 725–727. 10.1016/j.str.2019.04.011. [DOI] [PubMed] [Google Scholar]
- Marat AL, Ioannou MS, McPherson PS, 2012. Connecdenn 3/DENND1C binds actin linking Rab35 activation to the actin cytoskeleton. Mol. Biol. Cell 23, 163–175. 10.1091/mbc.e11-05-0474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McClay DR, 2011. Evolutionary crossroads in developmental biology: sea urchins. Development 138, 2639–2648. 10.1242/dev.048967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McClay D, Cannon G, Wessel G, Fink R, 1983. Time, Space and Pattern in Embryonic Development.
- McClay DR, Warner J, Martik M, Miranda E, Slota L, 2020. Gastrulation in the sea urchin. Curr. Top. Dev. Biol 136, 195–218. 10.1016/bs.ctdb.2019.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller JR, McClay DR, 1997. Characterization of the role of cadherin in regulating cell adhesion during sea urchin development. Dev. Biol 192, 323–339. 10.1006/dbio.1997.8740. [DOI] [PubMed] [Google Scholar]
- Morgulis M, Gildor T, Roopin M, Sher N, Malik A, Lalzar M, Dines M, Ben-Tabou de-Leon Shlomo, Khalaily L, Ben-Tabou de-Leon Smadar, 2019. Possible cooption of a VEGF-driven tubulogenesis program for biomineralization in echinoderms. Proc. Natl. Acad. Sci 116, 12353–12362. 10.1073/pnas.1902126116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakajima Y, Burke RD, 1996. The initial phase of gastrulation in sea urchins is accompanied by the formation of bottle cells. Dev. Biol 179, 436–446. 10.1006/dbio.1996.0273. [DOI] [PubMed] [Google Scholar]
- Nozumi M, Nakatsu F, Katoh K, Igarashi M, 2017. Coordinated movement of vesicles and actin bundles during nerve growth revealed by superresolution microscopy. Cell Rep 18, 2203–2216. 10.1016/j.celrep.2017.02.008. [DOI] [PubMed] [Google Scholar]
- Pereira-Leal JB, Seabra MC, 2001. Evolution of the rab family of small GTP-binding proteins. J. Mol. Biol 313, 889–901. 10.1006/jmbi.2001.5072. [DOI] [PubMed] [Google Scholar]
- Rafiq K, Cheers MS, Ettensohn CA, 2012. The genomic regulatory control of skeletal morphogenesis in the sea urchin. Development 139. 10.1242/DEV.073049. [DOI] [PubMed] [Google Scholar]
- Riedl J, Crevenna AH, Kessenbrock K, Yu JH, Neukirchen D, Bista M, Bradke F, Jenne D, Holak TA, Werb Z, Sixt M, Wedlich-Soldner R, 2008. Lifeact: a versatile marker to visualize F-actin. Nat. Methods 5, 605–607. 10.1038/nmeth.1220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato M, Sato K, Liou W, Pant S, Harada A, Grant BD, 2008. Regulation of endocytic recycling by C. elegans Rab35 and its regulator RME-4, a coated-pit protein. EMBO J 27, 1183–1196. 10.1038/emboj.2008.54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider CA, Rasband WS, Eliceiri KW, 2012. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sepúlveda-Ramírez SP, Toledo-Jacobo L, Henson JH, Shuster CB, 2018. Cdc42 controls primary mesenchyme cell morphogenesis in the sea urchin embryo. Dev. Biol 437, 140–151. 10.1016/j.ydbio.2018.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sethi AJ, Angerer RC, Angerer LM, 2014. Multicolor labeling in developmental gene regulatory network analysis. Methods Mol. Biol 1128, 249–262. 10.1007/978-1-62703-974-1_17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shim J, Lee S-M, Lee MS, Yoon J, Kweon H-S, Kim Y-J, 2010. Rab35 mediates transport of Cdc42 and Rac1 to the plasma membrane during phagocytosis. Mol. Cell. Biol 30, 1421–1433. 10.1128/MCB.01463-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stepicheva NA, Song JL, 2014. High throughput microinjections of sea urchin zygotes. J. Vis. Exp. JoVE, e50841. 10.3791/50841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stepicheva NA, Dumas M, Kobi P, Donaldson JG, Song JL, 2017. The small GTPase Arf6 regulates sea urchin morphogenesis. Differentiation 95, 31–43. 10.1016/j.diff.2017.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stumpp M, Hu MY, Melzner F, Gutowska MA, Dorey N, Himmerkus N, Holtmann WC, Dupont ST, Thorndyke MC, Bleich M, 2012. Acidified seawater impacts sea urchin larvae pH regulatory systems relevant for calcification. Proc. Natl. Acad. Sci 109, 18192–18197. 10.1073/pnas.1209174109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takata H, Kominami T, 2004. Pigment cells trigger the onset of gastrulation in tropical sea urchin Echinometra mathaei. Dev. Growth Differ 46, 23–35. [DOI] [PubMed] [Google Scholar]
- Terasaki M, Jaffe L, 2004. Labeling of cell membranes and compartments for live cell fluorescence microscopy. Methods Cell Biol 74. 10.1016/S0091-679X(04)74019-4. [DOI] [PubMed] [Google Scholar]
- Tu Q, Cameron RA, Davidson EH, 2014. Quantitative developmental transcriptomes of the sea urchin Strongylocentrotus purpuratus. Dev. Biol 385, 160–167. 10.1016/j.ydbio.2013.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uytterhoeven V, Kuenen S, Kasprowicz J, Miskiewicz K, Verstreken P, 2011. Loss of skywalker reveals synaptic endosomes as sorting stations for synaptic vesicle proteins. Cell 145, 117–132. 10.1016/j.cell.2011.02.039. [DOI] [PubMed] [Google Scholar]
- Vidavsky N, Addadi S, Schertel A, Ben-Ezra D, Shpigel M, Addadi L, Weiner S, 2016. Calcium transport into the cells of the sea urchin larva in relation to spicule formation. Proc. Natl. Acad. Sci 113, 12637–12642. 10.1073/pnas.1612017113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu S-Y, Ferkowicz M, McClay DR, 2007. Ingression of primary mesenchyme cells of the sea urchin embryo: a precisely timed epithelial mesenchymal transition. Birth Defects Res. Part C Embryo Today Rev 81, 241–252. 10.1002/bdrc.20113. [DOI] [PubMed] [Google Scholar]
- Yajima M, Wessel G, 2012. Autonomy in Specification of Primordial Germ Cells and Their Passive Translocation in the Sea Urchin. Development 139. 10.1242/DEV.082230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Fonovic M, Suyama K, Bogyo M, Scott MP, 2009. Rab35 controls actin bundling by recruiting fascin as an effector protein. Science (80-.). 325, 1250–1254. 10.1126/science.1174921. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.