

Abstract
Aims
Lipid metabolism might be compromised in type 1 diabetes, and the understanding of lipid physiology is critically important. This study aimed to compare the change in plasma lipid concentrations during carbohydrate dietary changes in individuals with type 1 diabetes and identify links to early‐stage dyslipidaemia. We hypothesized that (1) the lipidomic profiles after ingesting low or high carbohydrate diet for 12 weeks would be different; and (2) specific annotated lipid species could have significant associations with metabolic outcomes.
Methods
Ten adults with type 1 diabetes (mean ± SD: age 43.6 ± 13.8 years, diabetes duration 24.5 ± 13.4 years, BMI 24.9 ± 2.1 kg/m2, HbA1c 57.6 ± 2.6 mmol/mol) using insulin pumps participated in a randomized 2‐period crossover study with a 12‐week intervention period of low carbohydrate diet (< 100 g carbohydrates/day) or high carbohydrate diet (> 250 g carbohydrates/day), respectively, separated by a 12‐week washout period. A large‐scale non‐targeted lipidomics was performed with mass spectrometry in fasting plasma samples obtained before and after each diet intervention. Longitudinal lipid levels were analysed using linear mixed‐effects models.
Results
In total, 289 lipid species were identified from 14 major lipid classes. Comparing the two diets, 11 lipid species belonging to sphingomyelins, phosphatidylcholines and LPC(O‐16:0) were changed. All the 11 lipid species were significantly elevated during low carbohydrate diet. Two lipid species were most differentiated between diets, namely SM(d36:1) (β ± SE: 1.44 ± 0.28, FDR = 0.010) and PC(P‐36:4)/PC(O‐36:5) (β ± SE: 1.34 ± 0.25, FDR = 0.009) species. Polyunsaturated PC(35:4) was inversely associated with BMI and positively associated with HDL cholesterol (p < .001).
Conclusion
Lipidome‐wide outcome analysis of a randomized crossover trial of individuals with type 1 diabetes following a low carbohydrate diet showed an increase in sphingomyelins and phosphatidylcholines which are thought to reduce dyslipidaemia. The polyunsaturated phosphatidylcholine 35:4 was inversely associated with BMI and positively associated with HDL cholesterol (p < .001). Results from this study warrant for more investigation on the long‐term effect of single lipid species in type 1 diabetes.
Keywords: cardiovascular disease, dyslipidaemia, lipidomics, low carbohydrate diet, randomized trial, type 1 diabetes
This study demonstrates that unsaturated phosphatidylcholines and monounsaturated sphingomyelins were elevated with diet in individuals with type 1 diabetes following low carbohydrate diet.
![]()
1. INTRODUCTION
It is now recognized that type 1 diabetes is a major risk factor for developing cardiovascular events. 1 , 2 , 3 Dyslipidaemia is a modifiable risk factor for cardiovascular disease and highly prevalent in type 1 diabetes. 4 , 5 , 6 The present approach to diagnosing dyslipidaemia is based on the clinical measurement of the three main types of serum lipids, namely HDL cholesterol, LDL cholesterol and triglycerides. 7 However, since HDL‐cholesterol metabolism might be compromised in type 1 diabetes, the identification of alternative biomarkers is critically important. 3 , 4 , 5 , 6
Lipidomics studies suggest lipids as indicators or predictors for dyslipidaemia and cardiovascular disease. 8 , 9 , 10 , 11 , 12 , 13 , 14 However, to date, the plasma lipidome from individuals with type 1 diabetes has not been interrogated to understand diet‐dependent changes in a clinical setting.
Lipidomics expands the lipid information of the three main traditional mentioned lipids, since there are many more lipid species that can be measured with modern lipidomics methods. 15 , 16 , 17 Lipidomics provides a profile of hundreds of individual lipid species and on low‐abundance lipid species from a biological sample. Therefore, there is the potential to uncover novel insights into the pathophysiology in individuals with type 1 diabetes. The composition of the lipidome can reveal a fingerprint in relation to dyslipidaemia and cardiovascular disease in type 1 diabetes; thus, it can be an important tool for early screening.
In this lipidomics analysis of a randomized crossover trial, we compared the plasma lipids concentration from individuals with type 1 diabetes before and after ingesting isocaloric low carbohydrate diet (LCD) and high carbohydrate diet (HCD) for 12 weeks (<100 g vs. >250 g) separated by a 12‐week washout period obtained from a previous published study. 18 We hypothesized that (1) the lipidomics profiles before and after ingesting low or high carbohydrate diet for 12 weeks will be different; and (2) specific annotated lipid species have significant associations to metabolic characteristics.
2. METHODS
2.1. Clinical trial
Fasting plasma samples for lipidomics analysis were collected from a previously published study. 18 Participants were adults with insulin pump‐treated type 1 diabetes (mean ± SD: age 43.6 ± 13.8 years, diabetes duration 24.5 ± 13.4 years, BMI 24.9 ± 2.1 kg/m2, HbA1c 57.6 ± 2.6 mmol/mol). They underwent two 12‐week diet interventions separated by a 12‐week washout. Fasting plasma samples were collected before and after each intervention.
Main study outcomes from the previous article for the 14 study participants are given in Table 1. In brief, the study showed that glycaemic variability and time spent in hypoglycaemia were lower during LCD than during HCD. Mean glucose, however, was the same. Total daily bolus insulin doses were lower during LCD reflecting the lower carbohydrate intake. Finally, weight decreased during LCD, whereas it increased during HCD with a significant difference in change between groups. Systolic and diastolic BP increased during HCD (not significant); however, the between‐group differences were insignificant. The between‐group difference in HDL‐cholesterol levels was significant (p = .005) with the LCD showing an increase (LCD: 0.06 mmol/L; HCD: −0.03 mmol/L). No changes in fasting LDL cholesterol and triglyceride were detected. 18 In the present study, we therefore investigated whether the diet‐induced changes in relevant clinical variables (BMI, BP and HDL cholesterol) were associated with specific lipid species.
Table 1.
Metabolic characteristic measures before and after 12 weeks of LCD and HCD from the original study. 18 Data are mean (SD)
| Variable | LCD | HCD | LCD vs. HCD (p‐value) | ||||
|---|---|---|---|---|---|---|---|
| (Pre‐LCD) (N = 14) | (Post‐LCD) (N = 13) | (p‐value) | (Pre‐HCD) (N = 12) | (Post‐HCD) (N = 10) | (p‐value) | ||
| Cholesterol (mmol/L) | 4.5 (0.7) | 4.9 (0.7) | .284 | 4.8 (0.9) | 4.6 (0.6) | .271 | .084 |
| HDL cholesterol (mmol/L) | 1.97 (0.40) | 2.03 (0.40) | .500 | 2.09 (0.43) | 2.06 (0.48) | .084 | .005 |
| LDL cholesterol (mmol/L) | 2.3 (0.7) | 2.5 (0.7) | .238 | 2.5 (0.7) | 2.3 (0.5) | .301 | .095 |
| Triglyceride (mmol/L) | 0.6 (0.3) | 0.6 (0.2) | .999 | 0.6 (0.3) | 0.7 (0.3) | .363 | .452 |
| Systolic BP (mm Hg) | 134 (15) | 130 (14) | .485 | 128 (14) | 140 (9) | .007 | .091 |
| Diastolic BP (mm Hg) | 79 (13) | 77 (9) | .678 | 75 (13) | 82 (9) | .068 | .087 |
All participants were given personalized advice by a dietitian focused on eating strategies for meeting the carbohydrate criteria. LCDs contained less than 100 g carbohydrates per day, and HCDs contained a minimum of 250 g carbohydrates per day. Suggestions for a healthy composition of carbohydrates, fat and protein sources were provided; however, only the amount of carbohydrates was fixed. Carbohydrates intake was recorded in grams in the participants’ insulin pumps on a meal‐by‐meal basis, but there was no registration of actual food intake. 18
In this study, only samples from the 10 participants who completed both intervention periods are included.
2.2. Lipidomics analysis
A modified Folch lipid extraction method was used to analyse the total lipids from plasma samples. 19 Briefly, plasma samples were randomized and lipids were extracted from 10 µL plasma using chloroform:methanol (2:1 v/v) method following addition of nine different internal standards (stable isotope labelled and non‐physiological lipid species). A detailed name list of the used internal standards is available in the Methods S1. Samples were analysed in random order in positive and negative electrospray ionization modes using ultra‐high‐performance liquid chromatography‐quadrupole time‐of‐flight mass spectrometry (UHPLC‐Q‐TOF‐MS). The UHPLC system was from Agilent Technologies (Santa Clara, CA, USA) and was used as previously described. 20 , 21
2.3. Data pre‐processing
The lipidomics mass spectrometry data were pre‐processed in MZmine 2.18.2. 22 The workflow includes raw data import, filtering, peaks detection, chromatogram building, chromatogram deconvolution, peak list de‐isotoping, peak list alignment, gap filling and finally, peak annotation was carried out by combining MS and retention time information with in‐house lipid library with an m/z tolerance of 0.006 m/z and RT tolerance of 0.2 min. Lipidomics data post‐processing and analysis were performed in R programming language for statistical computing (https://www.r‐project.org/). 23 The pre‐processed data were normalized to internal standards. The missing values in the lipidomics data set were imputed with the k‐nearest neighbour algorithm. 24 To achieve normal distribution, data were log‐transformed. Coefficient of variation (Relative Standard Deviation; %RSD) for peak areas and retention times of lipid‐class specific internal standards were calculated. The measurement of 14 lipid classes was calculated by summing the individual lipid species within each class.
2.4. Statistical analysis
Data were analysed and visualized in R. Lipid species‐wise mixed‐effect models were used to consider the four repeated measures from each participant in the crossover trial. All statistical tests were corrected for multiple testing using the Benjamini‐Hochberg method. 25 Tests were corrected for multiple comparisons with false discovery rate (FDR).
First, differences between the two diets were modelled with lipid species‐wise mixed‐effect models, with time, diet and time‐diet interaction as fixed effects and the participant ID as random effects. We further conducted the lipid concentration changes found from the mentioned analysis within groups.
Second, associations between key lipid levels and the clinical covariates of interest were assessed: BMI, HDL cholesterol, systolic BP and diastolic BP. In this analysis, only lipid species with significant difference between groups from the first analyses were included. Association was tested by adding one clinical covariate at a time as an independent variable to the lipid‐wise mixed‐effect model detailed in the first step. A detailed description about the data analysis plan is available in the Methods S2. Finally, lipids of interest with significant association to clinical variables were semi‐quantified (relative quantified) and visualized in box plots grouped by diets and scatterplots. To support further research on the topic, the power to replicating the main finding in an identical study was estimated. This was done with a post hoc calculation of the power to finding an identical‐sized effect in an identical‐sized crossover study at an alpha level of 0.05.
3. RESULTS
3.1. Annotation of lipid species in plasma from type 1 diabetes
From the large‐scale untargeted lipidomics analysis, lipidome‐wide outcomes of the randomized trial resulted in annotation of 298 individual lipid species from 14 major lipid classes, including triacylglycerides (TGs), phosphatidylcholines (PCs), phosphatidylethanolamines (PEs), hexosyl‐ceramides (HexCer), sphingomyelins (SMs), lyso‐phosphatidylcholines (LPCs), ceramides (Cers), lactosyl‐ceramides (LactCers), free fatty acids (FAs), phosphatidylinositols (PIs), phosphatidylglycerols (PGs), lyso‐phosphatidylethanolamines (LPEs), phoshtatidylserines (PSs) and sulfatides (SHexCer). PCs and TGs dominated the data with 83 and 72 lipid species each followed by SMs and PEs with 30 identified lipid species each. PGs, LacCer, SHexCer, PSs and HexCers, on the other hand, showed only few identified lipid species each (1, 2, 3, 4 and 4). Additionally, Cers, LPCs, PIs, LPEs and FFAs were detected with 17, 16, 11, 8 and 8 identified lipid species, respectively. The dominance of identified lipid species within their respective lipid classes is shown in Figure S1. The coefficient of variation (%RSD) of peak areas was on average 17.41% for internal standards (Table S1).
3.2. Difference in the outcome between the diets
In total, 11 lipid species from phosphatidylcholine and sphingomyelin lipid classes and LPC(O‐16:0) had different outcome between groups (p < .05). In total, six out of the 83 annotated PCs, four out of the 30 annotated SMs, and LPC(O‐16:0) had different outcomes. Lipid names, 95% CI, FDR p‐values and the interaction slopes are given in Table 2. The strongest differences in the diet‐outcomes were monounsaturated SM(d36:1): (β ± SE: 1.44 ± 0.28, p = .01) and polyunsaturated PC(P‐36:4)/PC(O‐36:5): (β ± SE: 1.34 ± 0.24, p = .01). All the aforementioned lipid species were present in significantly higher amounts after LCD than after HCD (see the “Slope‐LCD” and “Slope‐HCD” columns in Table 2).
Table 2.
Results of the mixed‐effects model
| Name | Slope | L95 | U95 | SE | FDR | Slope‐LCD | Slope‐HCD |
|---|---|---|---|---|---|---|---|
| PC(P‐36:4)/PC(O‐36:5) | 1.34 | 0.83 | 1.85 | 0.24 | 0.009 | 0.63 | −0.70 |
| SM(d36:1) | 1.44 | 0.85 | 2.03 | 0.28 | 0.010 | 1.06 | −0.37 |
| PC(P‐38:5)/PC(O‐38:6) | 1.04 | 0.53 | 1.56 | 0.24 | 0.029 | 0.67 | −0.37 |
| SM(d36:2) | 0.76 | 0.38 | 1.13 | 0.18 | 0.029 | 0.65 | −0.10 |
| PC(P‐36:2)/PC(O‐36:3) | 1.59 | 0.77 | 2.42 | 0.39 | 0.036 | 0.89 | −0.70 |
| SM(d34:2) | 1.39 | 0.65 | 2.15 | 0.36 | 0.042 | 0.80 | −0.59 |
| PC(31:0) | 1.52 | 0.67 | 2.36 | 0.40 | 0.043 | 1.06 | −0.46 |
| PC(P‐38:4)/PC(O‐38:5) | 1.32 | 0.57 | 2.07 | 0.35 | 0.043 | 1.04 | −0.28 |
| PC(35:4) | 1.04 | 0.45 | 1.64 | 0.28 | 0.043 | 0.78 | −0.26 |
| LPC(O‐16:0) | 1.28 | 0.55 | 2.01 | 0.35 | 0.043 | 0.32 | −0.95 |
| SM(d38:2) | 0.89 | 0.38 | 1.39 | 0.24 | 0.043 | 0.64 | −0.25 |
Shown in the table are (Name) of individual lipid species, interaction slope (Slope), its lower and upper confidence intervals (L95, U95), standard error, p‐value of the slope (FDR) after correction for multiple testing, and slopes for LCD and HCD models (Slope‐LCD, Slope‐HCD) .
3.3. Association between lipid species and clinical variables
Finally, we investigated how the 11 lipid species, which responded to the two diets in different ways, were associated with metabolic characteristics reported previously. In the linear mixed‐effects model, there was an inverse association with the BMI and the lipid species PC(35:4): (β ± SE: −0.24 ± 0.05, p = .0005) and PC(31:0): (β ± SE: −0.21 ± 0.06, p = .003). The same trend of inverse association was also observed in diastolic BP, although not to statistical significance after correcting for multiple testing. PC(35:4) and PC(31:0) lipid species were elevated during the LCD intervention (see column “Slope‐LCD” and “Slope‐HCD” in Table 2), inferring a link between diet, circulating lipid concentrations, BMI and BP. There were no associations with the systolic BP that could be detected with statistical significance before and after correcting for multiple testing in this small study.
There was a positive association with the HDL cholesterol and three polyunsaturated phosphatidylcholine species (PC(35:4), PC(P‐36:4)/PC(O‐36:5), PC(P‐38:4)/PC(O‐38:5)) and two polyunsaturated sphingomyelin species (SM(d34:2), SM(d36:2)). Lipid names, 95% CI, FDR p‐values and slopes are given in Table S2.
One of the 11 significantly changed lipid species was associated both with BMI, diastolic BP and HDL, namely the polyunsaturated phosphatidylcholine 35:4. Figure 1A shows box plot of semi‐quantified pre‐post concentration comparisons of PC(35:4) in each diet intervention. Figure 1B and C shows scatter plots of PC(35:4) associations with BMI and HDL cholesterol. A post hoc power calculation of the discovered effect size in PC(35:4) revealed a 94% power to replicating the finding in a crossover study of same sample size.
Figure 1.
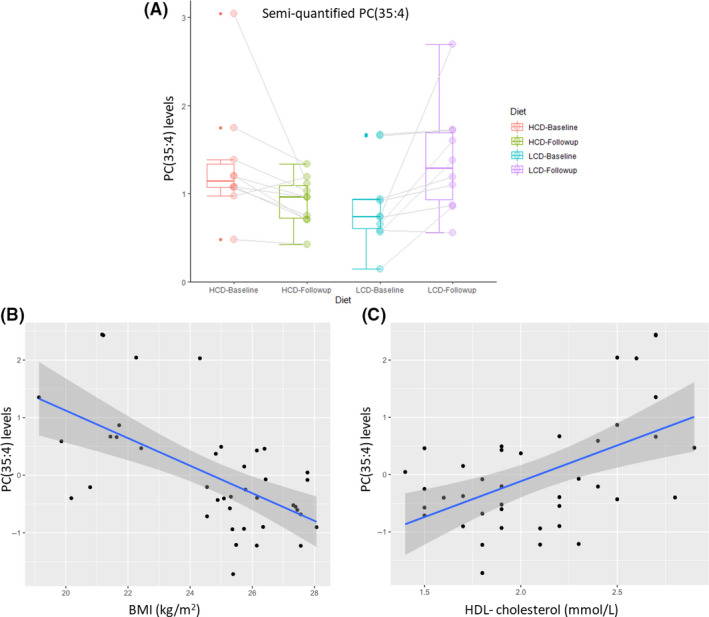
Jittered box plots of pre‐post lipid levels comparisons of PC(35:4) in each diet intervention and association with BMI and HDL cholesterol. PC(35:4): (β ± SE: 1.04 ± 0.28, p = .043) was elevated during LCD (a) and inversely associated with BMI (b) and positively associated with HDL cholesterol (c). HB: High carbohydrate diet at baseline, HF: High carbohydrate diet at follow‐up, LB: Low carbohydrate diet at baseline, LF: Low carbohydrate diet at follow‐up
4. DISCUSSION
To our knowledge, this is the first study comparing the lipidome in individuals with type 1 diabetes before and after low carbohydrate diet and high carbohydrate diet. Lipids are biomolecules with important functions, and their amount in the diet can modulate the blood lipidome in individuals with type 1 diabetes. In this study, we performed lipidomics in a randomized crossover trial and we compared the fasting plasma lipid profiles from individuals with type 1 diabetes before and after ingesting isocaloric LCD and HCD.
The main result of this study was that when both LDC and HDC were compared, 11 lipid species belonging to sphingomyelin and phosphatidylcholine were significantly changed in outcome between the diets (Table 2). Additionally, and most importantly, when we studied each trial arm separately, we observed that all the 11 significantly changed lipid species in the outcome were in greater concentration in the LCD arm, but none were altered significantly in the HCD arm. This was an unexpected result since total cholesterol, LDL and triglycerides had not changed significantly in this trial and only HDL cholesterol was significantly elevated in the LCD arm, suggesting a moderate lipid improvement with LCD. Also this suggests that a prolonged LCD could help with adjusting lipid classes since there is existing evidence that individuals with type 1 diabetes have decreased levels of blood sphingolipids and phosphatidylcholines, abundant in HDL. 26
Phosphatidylcholines and sphingomyelins are two important classes of phospholipids essential for cell membrane function and major components of HDL cholesterol specific those containing polyunsaturated fatty acids. 14 , 27 , 28 Interestingly, a noteworthy result from our study is that the LCD arm showed that HDL cholesterol was increased and positively associated with polyunsaturated sphingomyelins and phosphatidylcholines (Table S2). In rodents, sphingomyelin supplementation has been shown to help reduce the intestinal absorption of serum lipids such as cholesterol and triglycerides. 12 Sphingomyelin levels have also been associated with mild‐to‐moderate hypertension. 29 Based on these findings, supplementation could be an avenue to explore in the context of dyslipidaemia in type 1 diabetes. The findings for this study were in line with previous work on LCD in which similarly HDL cholesterol and diastolic BP were increased and decreased, respectively, with less carbohydrate intake, in a cross‐sectional carbohydrate intake study (carbohydrate intake of < 130 g/day and > 253 g/day) in type 1 diabetes. 2 Analogous results have been reported in individuals with type 2 diabetes, in which LCD resulted in a significant increase in HDL cholesterol and decrease in high BP. 30 , 31
Previous studies have demonstrated that abnormal sphingolipids levels are found at the onset of type 1 diabetes in regulating beta cell biology and inflammation. 26 Fenofibrate, a lipid lowering medication, is known to regulate sphingolipid metabolism and has been suggested as an important treatment in the management of dyslipidaemia. 32 It has been shown that very‐long‐chain sphingomyelin were increased in three weeks fenofibrate‐treated NOD mice and this had a beneficial effect on blood glucose homeostasis. 32 Interestingly, the main study outcome from Schmidt et al 18 showed that glycaemic variability and time spent in hypoglycaemia were lower during LCD than during HCD.
As the total amounts of cholesterol did not change with LCD, this was a positive result since hypercholesterolaemia is a major risk factor for developing cardiovascular disease. 33 , 34 , 35 The presence of CVD is related to disturbances in lipoprotein metabolism and dyslipidaemia. 36 Dyslipidaemia in individuals with diabetes not only fuels the reduction of HDL‐cholesterol concentration, but also it modifies the composition of lipoprotein. 3 , 4 , 5 , 6 Increasing evidence suggests that in patients with chronic inflammatory disorders, HDL cholesterol may lose important antiatherosclerosis properties and become dysfunctional. 36 So far, no therapeutic strategy to raise HDL cholesterol has been successful in reducing CVD. 36 In this study, we found three phosphatidylcholine lipid species (PC‐(O‐36:5), PC‐(O‐38:5) and PC(35:4)) and two sphingomyelin lipid species (SM(d36:2) and SM(d34:2)) associated with HDL cholesterol (Table S2). Interestingly, all these lipid species were increased during the LCD arm suggesting that LCD led to better functionality and composition of HDL‐cholesterol particles as they are enriched in plasma phosphatidylcholine (see the “Slope‐LCD” column in Table 2).
Remarkably, a previous lipidomics study in 3779 type 2 diabetes‐based cohort researchers found that PC(35:4) lipid species out of the 7 novel lipid species were associated with CVD. 37 PC(35:4) was suggested to improve prediction of CVD mortality. Remarkable result to emerge from our data is that PC(35:4) was the strongest positively associated lipid species with HDL cholesterol (Table S2) and increased during the LCD arm. Importantly, PC(35:4) was also inversely associated with BMI (Figure 1B) and diastolic BP. This and our investigation demonstrate the potential of lipid species as biomarkers for CVD risk in type 1 diabetes.
4.1. Strengths and limitations
There are certain limitations in the present study: Exact food intake for each study participant was not recorded. The diet plans were not conducted in a controlled environment. This means that it is difficult to know, what proportion of other energy sources—proteins or fats—the carbohydrates were replaced with in the LCD. While this varies by participant, we now know that the replacement resulted in the elevation of the concentrations of sphingomyelins and phosphatidylcholines in LCD. Another limitation is the small sample size of 10 participants, which was mitigated by the crossover study design. We view the randomized crossover study design and the high level of adherence to the carbohydrates criteria during LCD and HCD as two core strengths of this study. Further, the comprehensive lipidomic profiling is a strength of this study in type 1 diabetes.
In conclusion, our novel data now provide the foundation for future work aimed at diet lipid modulation in type 1 diabetes. We have demonstrated that low carbohydrate diet elevates blood sphingomyelin and phosphatidylcholine lipid species in type 1 diabetes, which are thought to reduce dyslipidaemia. Further, one single lipid species, polyunsaturated phosphatidylcholine 35:4, was inversely associated with BMI and positively associated with HDL cholesterol. Therapeutic development for diabetic complications such as dyslipidaemia and cardiovascular disease will require a better understanding of the crosstalk between diet and lipid metabolism in individuals with type 1 diabetes. This study provides support for several existing paradigms of dyslipidaemia and suggests new avenues of prevention of dyslipidaemia in type 1 diabetes. Confirming our findings, dietary lipid species, such as PC(35:4) should be tested in a larger cohort to fully understand the therapeutic opportunities in type 1 diabetes. Based on our estimate, the 94% probability of replicating the finding already in an identical‐sized study is considerably higher than the power of 80% that is typically used as a starting point at planning a study. For a deeper understanding of the mechanisms behind these lipidomic responses, however, a larger study is warranted.
CONFLICT OF INTEREST
None of the investigators has personal interests in the conduct or the outcomes of the study.
AUTHOR CONTRIBUTION
C.L‐Q., KN and FP designed and supervised the study. SS, AR and MBC undertook the clinical study. NA performed lipidomics analysis, statistical analysis and wrote the manuscript. TS and MK planned and supervised the statistical analysis. All authors critically reviewed and approved the final manuscript.
Supporting information
Supplementary Material
ACKNOWLEDGEMENTS
The authors thank the study participants.
Al‐Sari N, Schmidt S, Suvitaival T, et al. Changes in the lipidome in type 1 diabetes following low carbohydrate diet: Post‐hoc analysis of a randomized crossover trial. Endocrinol Diab Metab. 2021;4:e00213. 10.1002/edm2.213
Trial registration Clinicaltrials.gov NCT02888691
Funding Information
The lipidomics study did not receive any specific grant from funding agencies in the public, commercial or not‐for‐profit sectors. The original trial was funded by a research grant from the Danish Diabetes Academy supported by the Novo Nordisk Foundation, the Danish Diabetes Association, Vissing Fonden and The AP Møller Foundation.
Contributor Information
Kirsten Nørgaard, Email: kirsten.noergaard@regionh.dk.
Cristina Legido‐Quigley, Email: cristina.legido.quigley@regionh.dk.
DATA AVAILABILITY STATEMENT
Data sets may be requested from the corresponding author.
REFERENCES
- 1. Melendez‐Ramirez LY, Richards RJ, Cefalu WT. Complications of type 1 diabetes. Endocrinol Metab Clin North Am. 2010;39(3):625‐640. [DOI] [PubMed] [Google Scholar]
- 2. Ahola AJ, Forsblom C, Harjutsalo V, Groop P. Dietary carbohydrate intake and cardio‐metabolic risk factors in type 1 diabetes. Diabetes Res Clin Pract. 2019;155(2019):107818. [DOI] [PubMed] [Google Scholar]
- 3. Schofield JD, Liu Y, Rao‐Balakrishna P, Mlik RA, Soran H. diabetes dyslipidemia. Diabetes Ther. 2016;7(2):203‐219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Lopes SA, Fumani C, Jornayvaz FR, Philippe J, Gastaldi G. (2018) Dyslipidemia management in patients with type 1 diabetes. Rev Med Suisse 14(609), 1118‐1122. [PubMed] [Google Scholar]
- 5. Vergès B. Dyslipidemia in type 1 diabetes: a masked danger. Trends Endocrinol Metab. 2020;31(6):422‐434. [DOI] [PubMed] [Google Scholar]
- 6. Homma TK, Endo CM, Saruhashi T, et al. Dyslipidemia in young patients with type 1 diabetes mellitus. Arch Endocrinol Metab. 2015;215‐219. [DOI] [PubMed] [Google Scholar]
- 7. Wu L, Parhofer KG. Diabetic dyslipidemia. Metabolism. 2014;63(12):1469‐1479. [DOI] [PubMed] [Google Scholar]
- 8. Schofield JD, Steiner R, Liu Y, et al. Ceramides, sphingolipids and deoxysphingolipids: Derailed carbohydrate and fatty acid metabolism in diabetes. Diabet Med. 2017;34(1):9. [Google Scholar]
- 9. Alewijnse AE, Peters SLM. Sphingolipid signalling in the cardiovascular system: Good, bad or both? Eur J Pharmacol. 2008;585(2–3):292‐302. [DOI] [PubMed] [Google Scholar]
- 10. Jensen PN, Fretts AM, Hoofnagle AN, et al. Plasma ceramides and sphingomyelins in relation to atrial fibrillation risk: the cardiovascular health study. Am Heart J. 2020;9(4):1–13. 10.1161/JAHA.119.012853 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Yeboah J, McNamara C, Jiang XC, et al. Association of plasma sphingomyelin levels and incident coronary heart disease events in an adult population: Multi‐ethnic study of atherosclerosis. Arterioscler Thromb Vasc Biol. 2010;30(3):628‐633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Norris GH, Blesso CN. Dietary sphingolipids : potential for management of dyslipidemia and nonalcoholic fatty liver disease. Nutr Res Rev. 2017;75(4):274‐285. [DOI] [PubMed] [Google Scholar]
- 13. Stegemann C, Pechlaner R, Willeit P. Lipidomics profiling and risk of cardiovascular disease in the prospective population‐based bruneck study. J. Vasc Surg. 2014;60(2):532. [DOI] [PubMed] [Google Scholar]
- 14. Meikle PJ, Christopher MJ. Lipidomics is providing new insight into the metabolic syndrome and its sequelae. Curr Opin Lipidol. 2011;22(3):210‐215. [DOI] [PubMed] [Google Scholar]
- 15. Yang K, Han X. Lipidomics: Techniques, applications, and outcomes related to biomedical sciences. Trends Biochem Sci. 2017;41(11):954‐969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Loizides‐mangold U. On the future of mass‐spectrometry‐based lipidomics. FEBS J. 2013;280(12):2817‐2829. [DOI] [PubMed] [Google Scholar]
- 17. Ståhlman M, Pham HT, Adiels M, et al. Clinical dyslipidaemia is associated with changes in the lipid composition and inflammatory properties of apoprotein‐B‐containing lipoproteinjs from women with type 2 diabetes. Diabetologia. 2012;55:1156‐1166. [DOI] [PubMed] [Google Scholar]
- 18. Schmidt S, Christensen MB, Serifovski N, et al. Low versus high carbohydrate diet in type 1 diabetes : A 12‐week randomized open‐label crossover study. Diabetes Obes Metab. 2019;21(7):1680‐1688. [DOI] [PubMed] [Google Scholar]
- 19. Folch J, Lees M, Sloane GHS. A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem. 1957;226:497‐509. [PubMed] [Google Scholar]
- 20. Tofte N, Suvitaival T, Ahonen L, et al. Lipidomic analysis reveals sphingomyelin and phosphatidylcholine species associated with renal impairment and all‐cause mortality in type 1 diabetes. Sci Rep. 2019;79:16398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Bowden JA, Heckert A, Ulmer CZ, et al. Harmonizing lipidomics: NIST interlaboratory comparison exercise for lipidomics using SRM 1950‐Metabolites in Frozen Human Plasma. J Lipid Res. 2017;58:2275‐2288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Pluskal T, Castillo S, Villar‐Briones A, Orešič M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry‐based molecular profile data. BMC Bioinformatics. 2010;11(1):395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. The R Project for Statistical Computing . Available at: https://www.r‐project.org/. Accessed: 1 Aug 2019
- 24. Sas KM, Karnovsky A, Michailidis G, Pennathur S. Metabolomics and diabetes : analytical and computational approaches. Diabetes. 2015;64(3):718‐732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Storey JD, Tibshirani R. Statistical significance for genomewide studies. Proc Natl Acad Sci USA. 2003;100(16):9440‐9445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Lamichhane S, Ahonen L, Dyrlund TS, Siljander H. Data Descriptor : A longitudinal plasma lipidomics dataset from children who developed islet autoimmunity and type 1 diabetes. Sci Data. 2018;5:1‐9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Ohvo‐Rekilä H, Ramstedt B, Leppimäki P, Slotte JP. Cholesterol interactions with phospholipids in membranes. Prog Lipid Res. 2002;41(1):66‐97. [DOI] [PubMed] [Google Scholar]
- 28. Cohn JS, Kamili A, Wat E, Chung RWS, Tandy S. Dietary phospholipids and intestinal cholesterol absorption. Nutrients. 2010;2(2):116‐127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Shahin MH, Gong Y, Frye RF. Sphingolipid metabolic pathway impacts thiazide diuretics blood pressure response: Insights from genomics, metabolomics, and lipidomics. Am Heart J. 2018;7(1). 10.1161/JAHA.117.006656.1–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Feinman RD, Pogozelski WK, Astrup A, et al. Dietary carbohydrate restriction as the first approach in diabetes management: Critical review and evidence base. Nutrition. 2015;31(1):1‐13. [DOI] [PubMed] [Google Scholar]
- 31. Fechner E, Smeets ETHC, Schrauwen P, Mensink R. The effect of different degrees of carbohydrate restriction and carbohydrate replacement on cardiometabolic risk markers in humans‐A systematic review and meta‐analysis. Nutrition. 2020;12(4):991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Holm LJ, Haupt‐Jorgensen M, Giacobini JD, et al. Fenofibrate increases very‐long‐chain sphingolipids and improves blood glucose homeostasis in NOD mice. Diabetologia. 2019;62:2262‐2272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Paquette M, Baass A. Predicting cardiovascular disease in familial hypercholesterolemia. Curr Opin Lipidol. 2018;29(4):299‐306. [DOI] [PubMed] [Google Scholar]
- 34. Sanin V, Koenig W. Therapy of hypercholesterolemia in primary prevention. Dtsch Med Wochenschr. 2019;144(5):322‐328. [DOI] [PubMed] [Google Scholar]
- 35. Illingworth DR. Management of hypercholesterolemia. Med Clin North Am. 2000;84(1):23‐42. [DOI] [PubMed] [Google Scholar]
- 36. Rysz J, Gluba‐Brzózka A, Rysz‐Górzynska M, Franczyk B. The role and function of HDL in patients with chronic kidney disease and the risk of cardiovascular disease. Int J Mol Sci. 2020;21:601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Alshehry ZH, Mundra PA, Barlow CK, et al. Plasma lipidomics profiles improve on traditional risk factors for the prediction of cardiovascular events in type 2 diabetes mellitus. Circulation. 2016;134:1637‐1650. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Material
Data Availability Statement
Data sets may be requested from the corresponding author.