

Abstract
Licochalcone A (LA), a flavonoid found in licorice, has anticancer, antioxidant, anti-inflammatory, and neuroprotective properties. Here, we explored the effect of injecting LA into the tail vein of middle-aged C57BL/6 mice on their cognitive ability as measured by the Morris water maze (MWM) test and cerebral blood flow (CBF). The related mechanisms were assessed via RNA-seq, and T (CD3e+) and B (CD45R/B220+) cells in the spleen and whole blood were quantified via flow cytometry. LA improved the cognitive ability, according to the MWM test results, and upregulated the CBF level of treated mice. The RNA-seq results indicate that LA affected the interleukin (IL)-17 signaling pathway, which is related to T- and B-cell proliferation, and the flow cytometry data suggest that LA promoted T- and B-cell proliferation in the spleen and whole blood. We also performed immune reconstruction via a tail vein injection of lymphocytes into B-NDG (NOD-PrkdcscidIl2rgtm1/Bcge) mice before treating them with LA. We tested cognitive ability by subjecting these animals to new object recognition tests and quantified the splenic and whole blood T and B cells. Cognitive ability improved after immune reconstruction and LA treatment, and LA promoted T- and B-cell proliferation in the spleen and whole blood. This study demonstrates that LA, by activating the IL-17 signaling pathway, promotes T- and B-cell proliferation in the spleen and whole blood of mice and improves cognitive ability. Thus, LA may have immune-modulating therapeutic potential for improving cognition.
Keywords: licochalcone A, cognitive ability, T cells, B cells
INTRODUCTION
Cognitive decline is a characteristic of human aging, but age-related learning and memory deterioration also occur in rats and mice [1–3]. Aging is regulated by a variety of systems in the body, such as the nervous system [4], immune system [5], and metabolic system.
The immune system modulates the population sizes and functions of various cell subsets; this system can also increase inflammation and contribute to immune senescence [6]. T cells play a key role in the immune system. For example, regulatory T cells (Tregs) regulate oligodendrocyte differentiation, and central nervous system function is also affected by Tregs [7, 8]. Whether changes in the immune system can affect cognition still needs further research.
Licochalcone A (LA) is a flavonoid found in licorice (structure shown in Figure 1) that has been shown to have various clinically interesting pharmacologic effects, including antioxidant [9], anti-inflammatory [10], anticancer [11], and neuroprotective effects [12]. However, the mechanisms by which LA affects the immune system and the cognitive consequences of these effects are unknown. Here, to determine if LA has potential for development as a therapeutic treatment to improve cognitive ability, we first tested whether LA could improve the cognitive abilities of mice and investigated the underlying mechanisms by conducting RNA-seq of the hippocampus and assessing the proliferation of lymphocytes, such as T cells, B cells, and natural killer (NK) cells in the spleen and whole blood. We also treated immune-reconstructed B-NDG (NOD-PrkdcscidIl2rgtm1/Bcge) mice with LA, then subjected the animals to new object recognition tests and measured the T- and B-cell levels in the spleen and whole blood.
Figure 1.
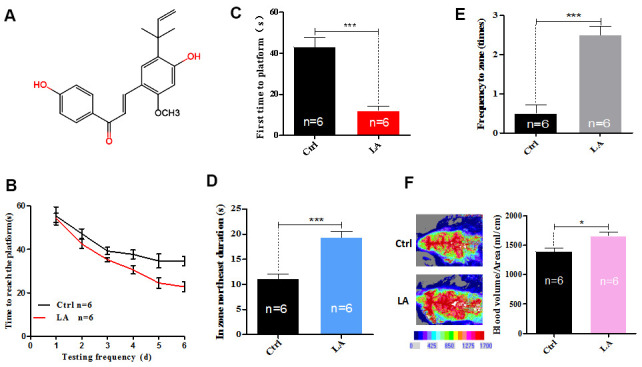
Effect of LA treatment on the cognitive ability of middle-aged C57BL/6 mice. (A) The chemical structure of LA. (B) Time taken by LA-treated or control (Ctrl)-treated mice to reach the platform in the MWM test over a 6-day experiment. (C) First latency to the platform in the MWM test. (D) In zone target duration in the MWM test. (E) Frequency in zone in the MWM test. (F) Cerebral blood flow level in LA- or Ctrl-treated mice. The data are presented as the mean±SD of three independent experiments. Statistical significance was determined using unpaired t-tests. *p < 0.05, **p < 0.01 compared with the Ctrl group; “n” indicates the number of animals in each experimental group.
RESULTS
LA improves the cognitive ability of middle-aged C57BL/6 mice
Cognitive ability declines with age [13, 14], and the Morris water maze (MWM) test [15] is commonly used to assess the spatial memory and cognitive ability of aging mice. In the present study, middle-aged mice were injected with LA into the tail vein every other day for a month and then subjected to the MWM test. The results show that, over the course of 6 days, the time that it took for the LA-treated mice to find the platform gradually decreased, unlike that in the control-treated mice (Figure 1B). The lower amounts of time required for the LA-treated mice to find the platform suggest that treatment with LA improved the spatial memory and cognitive ability of middle-aged C57BL/6 mice. To further evaluate the spatial memory and cognitive ability of the mice, analysis software was used to analyze their swimming tracks over 1 min, and the following three indicators were included: first time to platform (Figure 1C), in zone target duration (Figure 1D), and frequency in zone (Figure 1E). The MWM test results show that LA treatment significantly improved all measured indices related to cognitive ability in middle-aged C57BL/6 mice compared with control treatment.
Cerebral blood flow (CBF) is a key factor related to cognitive ability [16], and when the level of CBF is depressed, it can lead to cognitive dysfunction [17]. Changes in CBF are associated with various brain disorders [18]. CBF in the occipital lobe is important for brain functioning and regulates multiple neurological processes [19]. Because changes in the CBF are related to cognitive impairment [20], we measured the CBF in the LA-treated and control-treated mice. The results show that the CBF level in middle-aged C57BL/6 mice was higher for LA-treated animals than it was for control-treated animals (Figure 1F). The MWM test and CBF results indicate that LA treatment improves the cognitive ability of middle-aged C57BL/6 mice.
LA affects the immune system by regulating the interleukin (IL)-17 signaling pathway
Because the behavioral experiment results suggested that LA treatment improves cognitive ability, we next investigated the associated mechanism by performing RNA-seq on the hippocampus of C57BL/6 mice. The resulting Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway classification map, shown in Figure 2A, detected that 52 signaling pathways were found to be associated with the immune system. As seen in the bubble diagram shown in Figure 2B, LA was found to regulate the interleukin (IL)-17 signaling pathway, which is important in immune system function [21, 22]. Furthermore, as seen in the heatmap of differentially expressed genes related to the IL-17 signaling pathway, 11 of the detected genes are related to the proliferation of lymphocytes, including T cells, B cells, and NK cells (Figure 2C and Table 1) [23–45]. The RNA-seq analysis results suggest that LA treatment affects the proliferation of lymphocytes, including T cells and B cells, by regulating the IL-17 signaling pathway.
Figure 2.

Analysis of the mechanism by which LA regulates the immune system conducted using RNA-seq in the hippocampus. (A) Kegg pathway classification map. (B) Bubble diagram showing the pathways regulated by LA. (C) Heatmap showing the differentially expressed genes related to the IL-17 signaling pathway. The data are presented as the mean±SD of three independent experiments. Statistical significance was determined using unpaired t-tests. *p < 0.05, **p < 0.01 compared with the Ctrl group.
Table 1. Table for genes in heatmap which regulate lymphocytes proliferation.
| Genes regulated by IL-17 signaling pathway which related to lymphocytes | |||
| Genes | Lymphocytes | Genes | Lymphocytes |
| Omd | T cell [23], B cell [24], NK cell [25] | cd79a | T cell [36], B cell [37] |
| Slc6a12 | T cell [26], B cell [27], NK cell [28] | Wnt6 | T cell [38], B cell [39] |
| Slc47a1 | T cell [29], B cell [30], NK cell [31] | Foxc2 | T cell [40], B cell [41] |
| Jaml | T cell [32] | Foxd1 | T cell [42], B cell [43] |
| Myzap | T cell and B cell [33] | Cdh1 | T cell [44], B cell [45] |
| Spdef | T cell [34], NK cell [35] | ||
LA promotes T- and B-cell proliferation in middle-aged C57BL/6 mice
To determine whether LA affects lymphocyte proliferation, the percentages of T cells, effector T cells, B cells, effector B cells, and NK cells among the lymphocytes in the spleen and whole blood of LA- or control-treated middle-aged C57BL/6 mice were measured by flow cytometry. In the spleen, the percentages of B cells (CD45R/B220+), T cells (CD3e+), and NK cells (CD49b+) were all significantly higher in the LA-treated mice compared with the control-treated mice (Figure 3A, 3B). Similarly, the percentage of effector T cells (CD44+) in the spleen was significantly higher in the LA-treated mice than in the control-treated mice (Supplementary Figure 1A, 1C); however, the percentage of effector B cells (CD27+) was not different between the two treatment groups (Supplementary Figure 1B, 1D).
Figure 3.
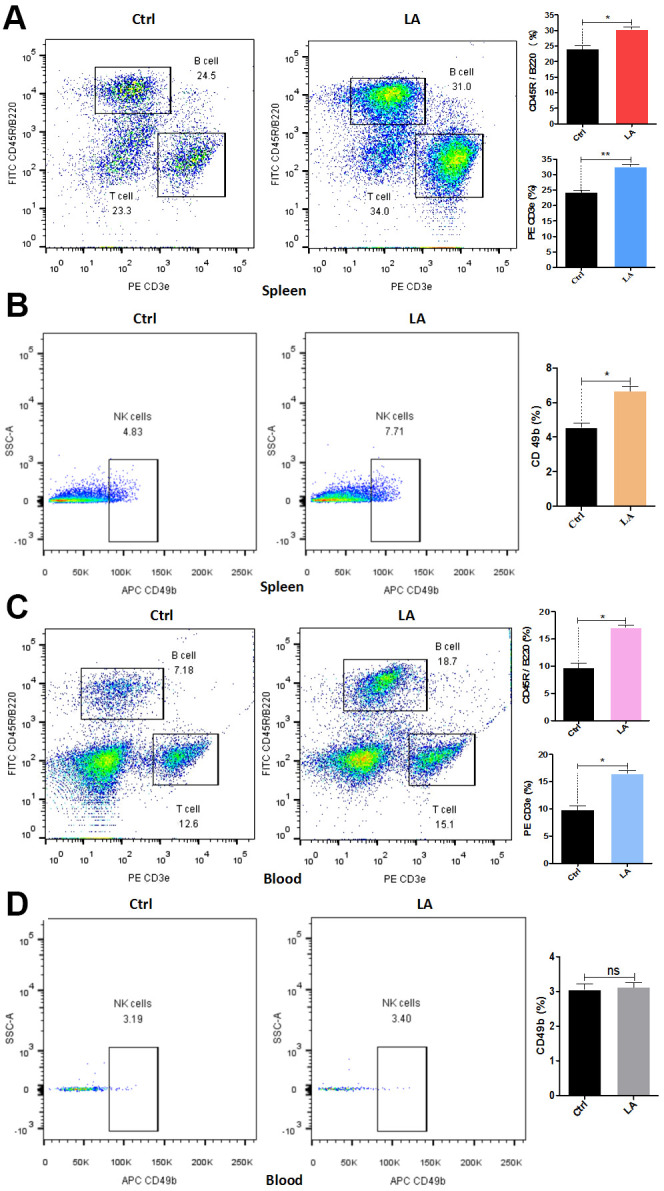
LA effect on lymphocyte proliferation in C57BL/6 mice. (A–D) T-cell and B-cell (A, C) or NK-cell (B, D) proliferation in the spleen (A, B) or whole blood (C, D) of LA- or control (Ctrl)-treated mice. The data are presented as the mean±SD of three independent experiments. Statistical significance was determined using unpaired t-tests. *p < 0.05, **p < 0.01 compared with the Ctrl group.
As in the spleen, the percentages of B cells, T cells, and effector T cells in whole blood were all significantly higher for the LA-treated mice than for the control-treated mice (Figure 3C and Supplementary Figure 1A), whereas the percentage of effector B cells was not different between the two groups. However, unlike in the spleen, the percentage of NK cells also did not differ between the two treatment groups (Figure 2D).
In vitro experiments were conducted to evaluate the effect of LA on T-cell and B-cell proliferation. The results show that immune cells from the spleens of LA-treated mice promoted significantly more T-cell and B-cell proliferation compared with those from control-treated mice (Supplementary Figure 2A). The flow cytometry results reveal that immune cells from the blood of LA-treated mice also promoted T-cell and B-cell proliferation significantly more compared with blood immune cells from control-treated mice (Supplementary Figure 2B). The relative mRNA expression results show that the mRNA expression of T cells (CD3e+) and B cells (CD45R/B220+) was increased by LA treatment as compared with control treatment (Supplementary Figure 2C).
The flow cytometry results from in vitro experiments suggest that LA treatment promoted the proliferation of T and B cells. The expression of homeobox B (HOXB) cluster genes was evaluated as a measure of activated lymphocytes [46] because HOXB-3 plays an essential role in regulating B lymphopoiesis in mice [47] and HOXB-3 overexpression promotes the proliferation and differentiation processes of lymphocytes [48], while HOXB-5 is dispensable for T-cell proliferation [49]. The runt-related transcription factor (RUNX) family is involved in the development of T and B lymphocytes [50]; RUNX-1 functions in T-cell population maintenance [51], and RUNX-2 can promote B-cell proliferation [52]. Additionally, T and B cells are also influenced by RUNX-3 [53]. To learn about the mechanism of how LA activates the immune system in vitro, we examined the mRNA expression of in vitro spleen cells. The results show that in LA-treated cells, the mRNA expression levels of HOXB-3, RUNX-2, and RUNX-3 are all significantly higher than those in control-treated cells (Supplementary Figure 2D). Thus, the mechanism by which LA activates the immune system, at least in vitro, involves the activation of HOXB-3, RUNX-2, and RUNX-3 expression to promote the proliferation of T and B cells.
To further investigate the effect of LA on neurons, we examined expression of the neural stem cell marker SOX2 [54, 55] in the hippocampus and subventricular zone (SVZ) of LA-treated mice and control mice. The results show that SOX2 expression in the hippocampus was significantly higher for the LA-treated aging mice than for the control-treated aging mice (Supplementary Figure 2E, 2F). The result also indicates that LA treatment improved the cognition of middle-aged C57BL/6 mice.
In conclusion, LA promotes the proliferation of T and B cells in middle-aged C57BL/6 mice by activating the IL-17 signaling pathway, and the results of in vitro work suggest that LA treatment improves the percentage of T and B cells via activating the expression of HOXB-3, RUNX-2, and RUNX-3.
LA treatment can promote T- and B-cell proliferation in B-NDG mice
The experiments conducted in C57BL/6 mice described above revealed that LA treatment improved the T- and B-cell proliferation in the whole blood and spleen. To further investigate the relationship between LA treatment and the immune system, we next conducted experiments in B-NDG mice, which have a severe immune defect phenotype [56]. The B-NDG mice underwent immune reconstitution (IR) via a tail vein injection of lymphocytes, after which they were treated with LA for 30 days.
Previous work found that meningeal T-cell composition is coupled to the central nervous system draining deep cervical lymph nodes, that the normal flow of meningeal T cells regulates cognition [57], and that an increase in the percentage of circulating B and T cells ameliorated brain cognition [58]. Because cognitive ability has been reported to be partially regulated by the immune system [59, 60], we tested the effect of LA treatment on the cognitive ability of B-NDG mice by using the new object recognition test. The new object-touch time for LA-treated IR mice was significantly higher than that for PBS-treated IR mice and PBS-treated non-IR mice (Figure 4A, 4B). These results suggest that LA treatment improved the memory of B-NDG mice. Prior research suggested that T lymphocytes contribute to an increase in hippocampal neurogenesis and working memory [61] and that B lymphocytes may represent a therapeutic option for the treatment of cerebral contusion [62]. These findings indicate that memory improvement is closely related to the regulation of T and B cells.
Figure 4.

Effect of LA on T- and B-cell proliferation in B-NDG mice. (A) New object recognition results for the IR+LA, IR and non-IR groups. (B) Statistical analysis of the results shown in (A). (C, D) T- and B-cell proliferation in the spleen (C) and whole blood (D). The data are presented as the mean±SD of three independent experiments. Statistical significance was determined using unpaired t-tests. *p < 0.05, **p < 0.01 compared with the non-IR group; “n” indicates the number of animals for each experimental group.
Because our above results suggested that LA treatment promotes T- and B-cell proliferation, the improved memory of LA-treated IR B-NDG mice may be due to an LA treatment-induced increase in T and B cells. To further investigate this possibility, we examined the lymphocyte populations in B-NDG mice. The percentages of T cells and B cells in the spleen were significantly higher in LA-treated IR mice than in PBS-treated IR mice or PBS-treated non-IR mice (Figure 4C). However, in whole blood, the percentage of T cells was significantly higher in LA-treated IR mice than in PBS-treated IR mice or PBS-treated non-IR mice, and the percentage of B cells in these mice was significantly higher compared with that in the PBS-treated IR mice or PBS-treated non-IR mice (Figure 4D). These results suggest that LA treatment in B-NDG mice promotes the production of lymphocytes, specifically T cells and B cells.
Because the behavioral experiment and flow cytometry results suggested that LA treatment improves the cognitive ability of IR mice, we next investigated the associated mechanism by performing RNA-seq on the hippocampus of B-NDG mice. The differentially expressed genes between the PBS-treated IR mice and PBS-treated non-IR mice, PBS-treated IR mice and LA-treated IR mice, and PBS-treated non-IR mice and LA-treated mice are listed in Supplementary Figure 3A. In the LA-treated mice, 126 of the differentially expressed genes were upregulated and 26 of these genes were downregulated. As shown by the bubble diagram in Supplementary Figure 3B, LA was found to regulate the Th1- and Th2-cell differentiation signaling pathway, which controls Th1- and Th2-cell differentiation during the T-cell activation process [63]; notably, Th1 and Th2 are involved in the pathogenesis of many immune-mediated diseases [64]. Furthermore, as seen in the heatmap shown in Supplementary Figure 3C, among the differentially expressed genes related to the Th1- and Th2-cell differentiation signaling pathway, the H2-Eb1, H2-Ab1, H2-Aa, CD74 genes were upregulated in the LA-treated IR mice as compared with their levels in the PBS-treated IR mice or PBS-treated non-IR mice. Previous work suggested that the activation of H2-Eb1, H2-Ab1, H2-Aa, and CD74 expression promotes T-cell synapse formation [65]. Additionally, another study reported that T cells were significantly higher among autoantibodies against CD74 [66], and anti-CD74 monoclonal antibody may provide a basis for novel therapeutic approaches to treating B-cell malignancies [67]. Furthermore, we conducted a gene set enrichment analysis of these differentially expressed genes, which similarly revealed that LA treatment activated the Th1- and Th2-cell differentiation signaling pathway (Supplementary Figure 3D). The RNA-seq analysis results suggest that LA treatment affects the proliferation of lymphocytes, including T cells and B cells, by activating the Th1- and Th2-cell differentiation signaling pathway.
In vitro experiments were conducted to evaluate the effect of LA on the proliferation of T or B cells. The results show that immune cells from the spleens of IR mice treated with LA promoted the proliferation of T and B cells significantly more as compared with splenic immune cells from PBS-treated IR mice (Supplementary Figure 4A). The flow cytometry results reveal that immune cells from the blood of LA-treated IR mice also promoted the proliferation of T cell significantly more as compared with blood immune cells from PBS-treated IR mice, whereas the percentage of B cells did not differ significantly between the two groups (Supplementary Figure 4B). The relative mRNA expression results show that the mRNA expression of T cells (CD3e+) and B cells (CD45R/B220+) is increased by LA treatment (Supplementary Figure 4C).
As shown above, the in vitro flow cytometry results indicated that LA treatment promotes T-cell and B-cell proliferation. To learn about the mechanism by which LA activates the immune system in vitro, we next examined the mRNA expression of in vitro spleen cells. The mRNA expression levels of HOXB-2, HOXB-3, and RUNX-2 were significantly higher in LA-treated IR cells than in PBS-treated IR cells (Supplementary Figure 4D). Thus, the mechanism by which LA activates the immune system in vitro involves the activation of HOXB-2, HOXB-3, and RUNX-2 expression, which promote the proliferation of T and B cells.
In conclusion, LA can activate the immune reconstitution of B-NDG mice by promoting the proliferation of T and B cells, and in vitro experiments further revealed that LA promotes T-cell proliferation by activating the expression of HOXB-2, HOXB-3, and RUNX-2.
DISCUSSION
Here, to investigate whether LA can improve cognitive ability, LA was injected into the tail veins of middle-aged mice, and the animals were assessed by using the MWM test, which measures cognitive ability [68], and measuring their CBF levels, which are a key factor for neuroprotection [69]. The MWM test results indicate that LA improved the memory of treated mice, and the CBF results show that LA treatment upregulates the CBF level.
RNA-seq was performed on hippocampus samples; the results show that LA treatment affected the IL-17 signaling pathway and that the signal pathway changes were related to the proliferation of lymphocyte subsets, including T cells, B cells, and NK cells, through the regulation of immune system gene expression. IL-17 is a proinflammatory cytokine that is crucial for a variety of processes, including host defense and inflammatory disease pathogenesis [70]. Besides having proinflammatory roles, IL-17 is also a neuromodulator [71], and the IL-17 signaling pathway regulates multiple phenotypes, including learning, immunity, and longevity [72].
Our flow cytometry results indicate that LA treatment significantly promoted the proliferation of T cells and B cells in the spleen and whole blood. Previous work found that IL-17 can regulate T-cell proliferation in the peripheral blood [73]. Additionally, IL-17 can activate B cells, which leads to an inflammatory immune reaction [74], and can promote T cell–B cell interactions that induce B-cell expansion and antibody production [75].
Together, our findings in middle-aged C57BL/6 mice indicate that LA promotes T- and B-cell proliferation in the spleen and whole blood by regulating the IL-17 signaling pathway. Our in vitro results further suggest that LA treatment promotes the proliferation of T and B cells by activating the expression of HOXB-3, RUNX-2 and RUNX-3.
The results of our new object recognition experiment in LA-treated IR B-NDG mice provide further evidence that LA treatment improves the cognitive ability of mice. Additionally, the flow cytometry results show that LA promoted T- and B-cell proliferation in the spleen and promoted T-cell proliferation in the blood of these animals. Thus, LA may improve the cognitive ability of mice by stimulating the proliferation of lymphocytes, such as T cells and B cells. Other research in B-NDG mice, which are severely immunodeficient mice that do not have lymphocytes such as mature T or B cells, found that treatment targeting chimeric antigen receptor (CAR)-modified T cells promotes T-cell proliferation [76], which further supports our finding that LA treatment improves T- and B-cell proliferation. The results of our in vitro experiments also suggest that LA treatment promotes the proliferation of T cells by activating the expression of HOXB-2, HOXB-3, and RUNX-2 genes.
Our results indicate that LA treatment promotes T- and B-cell proliferation and improves the cognitive ability of middle-aged C57BL/6 mice by activating the IL-17 signaling pathway. Previous research in rats revealed that increasing the Th1/Th2 cytokine balance in the hippocampus reduced cognitive deficits by improving memory [77]. Lymphocyte activation has a clinical impact, as indicated by the reported correlation between activated T-cell levels and the pathology of Alzheimer’s disease [78], and B-cell diapedesis, which mediated memory recovery, occurred in areas remote to the infarction [79]. Thus, T- and B-cell proliferation play an important role in improving memory by regulating related signaling pathways.
LA, a major flavonoid in Glycyrrhiza inflata, has multiple known pharmacological effects [80]; here, we explored the influence of LA on cognitive ability from the perspective of the immune system. G. inflata is an important Chinese medicinal plant and is considered to have potential for future drug development as an anti-coronavirus 19 agent [81, 82]. There are many molecules known to be capable of promoting lymphocyte proliferation. For example, icariin exerts neuroprotective effects via modulating the CD4+ T lymphocyte-related immunoinflammatory responses in APP/PS1 mice and may be a promising drug against Alzheimer’s disease progression [83]. Additionally, glycyrrhizic acid also improves cognition in aging mice through promoting T- and B-cell proliferation [84], and resveratrol significantly improves the growth of T cells in the human circulating immune system [85]. Furthermore, ginsenoside Rg1 ameliorates aging through upregulating T-cell proliferation [86]. Thus, exploring the effects of active ingredients in medicinal plants can reveal clinically useful findings.
In conclusion, our data suggest that LA can improve cognitive ability by inducing T-cell proliferation in the spleen and whole blood. This is the first demonstration of LA affecting the immune function of middle-aged C57BL/6 mice and B-NDG mice. Additionally, LA was found to affect the IL-17 signaling pathway by regulating T- and B-cell proliferation in middle-aged C57BL/6 mice. Furthermore, LA was found to affect the Th1- and Th2-cell differentiation signaling pathway and promote IR by regulating T- and B-cell proliferation in B-NDG mice. These findings suggest that it may be possible to improve cognitive ability by activating the immune system via the use of a small molecular compound.
MATERIALS AND METHODS
Drug
LA (C21H2O4; MW: 338; purity ≥98%, HPLC grade) was purchased from Chengdu Push Biotechnology Co. Ltd. (Sichuan, China) and stored at 2-8° C in a dark, dry place. The stock solution had a concentration of 15 mg/L in phosphate-buffered saline (PBS).
Animals
This study was approved by the Animal Research Committee of Tongji University School of Medicine and was conducted in accordance with institutional guidelines. For treatment, C57BL/6 mice (12 months old, female) purchased from Shanghai SLAC Laboratory Animal Co. were injected with LA via the tail vein every other day for a month. B-NDG mice (8 weeks old, female) purchased from Jiangsu Biocytogen Laboratory Animal Co. (BCM002F). All mice were maintained on a 12-h light/dark cycle with free access to food and water. Each experimental group was composed of six mice.
MWM test
For the MWM test, a cylindrical tank with a diameter of 120 cm was filled with water. The water temperature was maintained at room temperature, and a platform was placed 1cm below the water level for use as a marker of mouse cognition. The spatial positions corresponding to the southeast (SE), east (E), southwest (SW), and west (W) directions of the platform were marked with different colors to provide a reference for spatial memory in the cognitive process of the mice.
Our experiment was divided into two stages. In the first stage, each mouse was required to swim for 1 min in the four directions corresponding to the platform in the pool. In the second stage, the platform was removed from the pool on day 6, and the mice were left to swim for 1 min at the farthest point from the platform. The swimming track of each mouse was recorded by a computerized tracking/image analyzer system [87].
CBF detection
The brains of anesthetized mice were exposed, and images were acquired with a laser speckle contrast imager (PeriCam PSI System, Perimed, Stockholm, Sweden). The PeriCam PSI HD system was used to calculate an arbitrary index of cerebral blood flow (perfusion units) in the ipsilateral hemisphere.
RNA-seq analysis
RNA-seq was performed independently and uniformly on the total RNA extracted from hippocampus samples from LA- or control-treated mice. The clean reads were aligned to the reference gene sequence using bowtie-2, and the gene expression levels of each sample were calculated. Differential gene detection was conducted by applying the DEGseq method [88]. The statistical results are based on the ma-plot method, in which the number of reads for specific genes obtained from the sample were randomly sampled, and then p-values were calculated according to the normal distribution and corrected to q-values. To improve the accuracy of identified differentially expressed genes (DEGs), genes with a difference multiple of more than double the lower value and a q-value of ≤0.001 were defined and screened as significant DEGs. RNA sequencing data were deposited at GSE144123.
Flow cytometry
Single-cell suspensions were generated from the spleen and blood collected from experimental mice. After red blood cells were eliminated by using the Red Cell Lysis buffer solution (BD Biosciences, San Jose, CA, USA), lymphocytes were filtered through a 70-μm cell strainer (Jet Biofil, Guangzhou, China) and diluted in 1× PBS (Thermo Fisher Scientific, Waltham, MA, USA). T cells (CD3e+), B cells (CD3e−CD45R/B220+), NK cells (CD3e−CD49b+), effector T cells (CD45+CD3e+CD44+), and effector B cells (CD45+CD3e−, CD45R/B220+CD138+CD27+) in the spleen and blood were directly quantified using flow cytometry with monoclonal antibodies (1 μg/ml) directed against the noted specific human and mouse antigens (Table 2; all antibodies purchased from BD Pharmingen, San Diego, CA, USA). The stained cells were detected using a flow cytometer (Beckman FC-500, Miami, FL, USA) and analyzed with FlowJo 7.6.1. software.
Table 2. Antibodies for examining lymphocytes.
| Antibodies | Source | Article number |
| CD45R/B220 | BD Biosciences | Cat# 553087 |
| CD3e | BD Biosciences | Cat# 553063 |
| CD27 | BD Biosciences | Cat# 558754 |
| CD138 | BD Biosciences | Cat# 558626 |
| CD44 | BD Biosciences | Cat# 553134 |
| CD45 | BD Biosciences | Cat# 566439 |
| CD49b | BD Biosciences | Cat# 558295 |
Immune reconstitution (IR)
Lymphocytes were harvested from middle-aged C57BL/6 mouse spleens by removing red blood cells with lysis buffer (BD Biosciences, San Jose, CA, USA) and filtering the cells through a 70-μm cell strainer (Jet Biofil, Guangzhou, China) using 1×PBS (Thermo Fisher Scientific, Waltham, MA, USA). The lymphocytes were kept alive by storing them on ice. These cells (3×105 per animal) were then injected into the tail veins of 8-week-old B-NDG (NOD-PrkdcscidIl2rgtm1/Bcge) mice, after which the animals were treated with LA for 30 days.
New object recognition test
The new object recognition test was employed to examine the memory ability of B-NDG mice [89]. This model evaluates the memory ability of the tested mice according to the length of exploration time spent on familiar objects (ones they have seen previously) and on new objects (ones they have not seen before). When the tested mice have not forgotten the familiar objects, they will spend more time exploring the new objects. However, when the tested mice have forgotten the familiar objects, they will spent roughly the same time exploring the new objects as they do exploring the familiar objects. VisuTrack of Animal Behavior software was used for the analysis.
Cell culture
LA (C21H22O4; MW: 338; purity >98%, HPLC grade) was purchased from Chengdu Push Biotechnology Co. Ltd. (Sichuan, China) and stored at 2-8° C in a dark, dry place. The concentration of the stock solution was 100 mM in dimethyl sulfoxide (DMSO). The final DMSO concentration did not exceed 0.1% in the culture medium. Lymphocytes obtained from spleen and blood were cultured in a humidified 5% CO2 atmosphere at 37° C in DMEM/F12 (Thermo Fisher Scientific, Waltham, MA, USA) medium supplemented with 10% FBS and 100 U/mL penicillin and streptomycin.
qRT-PCR analysis
Total RNA was isolated using TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA), and cDNA was prepared using the Prime Script™ RT Master Mix (Perfect Real Time) (Takara, Dalian, China) according to the manufacturer’s protocols. The qRT-PCR reactions were performed using SYBR green fluorescent dye (BioRad). Primer sequences are listed in the Table 3.
Table 3. Primer sequences used in this study.
| Genes | Forward primers (5’-3’) | Reverse primers (5’-3’) |
| CD3e | CTGCTACACACCAGCCTCAA | GTAATAAATGACCATCAGCAAGC |
| CD45R | CGGAAGTTCCTGGAGCACCTCTC | AAGTACACCTTGGCCCCCACGTA |
| HOXB-2 | GATGGCCTGAACCTCATCGA | AGTTCGGTCCGGTTCCAGAT |
| HOXB-3 | AGATATTCCCCTGGATGAAAGA | GAACTCCTTCTCCAG CTCCAC |
| HOXB-5 | GTGCCAATGTTGTGTGTTGC | TCAGGTAGCTTGTTCCTTGG |
| RUNX-1 | TGGCAGGCAACGATGAAAAC | CGCTCGGAAAAGGACAAACTC |
| RUNX-2 | CAGGAAGAGCGGCAAGTATTA | AAGGTCCGAAGTTGAGGGAAA |
| RUNX-3 | TTGCCAAGCCTTATCGGAA | CAGGGGAGAAATCGATGACA |
| Actin | TATTGGCAACGAGCGGTTC | ATGCCACAGGATTCCATACCC |
Immunofluorescence assay
From our experiment, mice from each of the LA group and control group were randomly selected. Following anesthesia and killing, half of the brain from each mouse was fixed with 4% paraformaldehyde (PFA) overnight at 4° C using 15% and 30% sucrose gradient dehydration. Next, optimal cutting temperature compound (OCT) (Tissue-Tek, USA) was used to coat the brain hemispheres at 4° C for 2 h to discharge air bubbles. Then the brains were placed in an embedding box in absolute ethanol at -80° C to allow slow solidification. Serial sections (sagittal section, 10 μm) were chosen per mouse by freezing microtome. The tissue slices were treated with methanol or 4% PFA for 10 min at -20° C. After three washes, all slides were incubated in blocking buffer (3% BSA 0.3% triton X-100 in PBS) for 1 h at room temperature. Slides were immersed in 200 μL primary anti-SOX2 antibody (1:500) per slide for overnight incubation at 4° C, followed by secondary antibody anti-rabbit for 1h in the dark at room temperature. DAPI reagent was used as a counter staining medium. The slides were examined using an Olympus BX53 microscope.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 5.0, and the data are expressed as the mean ± standard error of the mean (mean ± SD). Statistically significant differences between two groups were assessed by using unpaired t-tests. Statistical comparisons of more than two groups were performed through an analysis of variance (ANOVA). A two-tailed p-value of <0.05 was considered statistically significant.
Supplementary Material
Abbreviations
- LA
licochalcone A
- IL-17
interleukin
- MWM
morris water maze
- CBF
cerebral blood flow
- T cells
CD3e+
- B cells
CD45R/B220+
- NK cell
natural killer (CD49b) cells
- effector T cells
CD44+
- effector B cells
CD27+
- CNS
central nervous system
- RNA-Seq
RNA sequencing
- DEGs
differentially expressed genes
- BND-G
mice NOD-PrkdcscidIl2rgtm1/Bcge mice
- Ctrl
control
Footnotes
AUTHOR CONTRIBUTIONS: W.Y.T. performed experiments. W.Y.T. and L.H.L. analyzed data. L.H.L. supervised the project. Z.J.B., L.H.F., and L.H.L designed the project. W.Y.T. wrote the paper; L.H.L. revised the manuscript, and all authors approved the final version of the manuscript.
CONFLICTS OF INTEREST: The authors declare that they have no conflicts of interest.
FUNDING: This work was supported by grants from the National Key Research and Development Program (2020YFC2002800), National Natural Science Foundation of China (Grant No 31671539), and Major Program of Development Fund for Shanghai Zhangjiang National Innovation Demonstration Zone<Stem Cell Strategic Biobank and Stem Cell Clinical Technology Transformation Platform> (ZJ2018-ZD-004).
REFERENCES
- 1.Frick KM, Burlingame LA, Arters JA, Berger-Sweeney J. Reference memory, anxiety and estrous cyclicity in C57BL/6NIA mice are affected by age and sex. Neuroscience. 2000; 95:293–307. 10.1016/s0306-4522(99)00418-2 [DOI] [PubMed] [Google Scholar]
- 2.Shoji H, Takao K, Hattori S, Miyakawa T. Age-related changes in behavior in C57BL/6J mice from young adulthood to middle age. Mol Brain. 2016; 9:11. 10.1186/s13041-016-0191-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bach ME, Barad M, Son H, Zhuo M, Lu YF, Shih R, Mansuy I, Hawkins RD, Kandel ER. Age-related defects in spatial memory are correlated with defects in the late phase of hippocampal long-term potentiation in vitro and are attenuated by drugs that enhance the cAMP signaling pathway. Proc Natl Acad Sci USA. 1999; 96:5280–85. 10.1073/pnas.96.9.5280 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Liu J, Zhang B, Lei H, Feng Z, Liu J, Hsu AL, Xu XZ. Functional aging in the nervous system contributes to age-dependent motor activity decline in C. elegans. Cell Metab. 2013; 18:392–402. 10.1016/j.cmet.2013.08.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nikolich-Žugich J. The twilight of immunity: emerging concepts in aging of the immune system. Nat Immunol. 2018; 19:10–19. 10.1038/s41590-017-0006-x [DOI] [PubMed] [Google Scholar]
- 6.Pansarasa O, Pistono C, Davin A, Bordoni M, Mimmi MC, Guaita A, Cereda C. Altered immune system in frailty: genetics and diet may influence inflammation. Ageing Res Rev. 2019; 54:100935. 10.1016/j.arr.2019.100935 [DOI] [PubMed] [Google Scholar]
- 7.Dauer DJ, Huang Z, Ha GK, Kim J, Khosrowzadeh D, Petitto JM. Age and facial nerve axotomy-induced T cell trafficking: relation to microglial and motor neuron status. Brain Behav Immun. 2011; 25:77–82. 10.1016/j.bbi.2010.08.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yshii L, Gebauer C, Bernard-Valnet R, Liblau R. Neurons and T cells: understanding this interaction for inflammatory neurological diseases. Eur J Immunol. 2015; 45:2712–20. 10.1002/eji.201545759 [DOI] [PubMed] [Google Scholar]
- 9.Liang M, Li X, Ouyang X, Xie H, Chen D. Antioxidant mechanisms of echinatin and licochalcone A. Molecules. 2018; 24:3. 10.3390/molecules24010003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yang G, Lee HE, Yeon SH, Kang HC, Cho YY, Lee HS, Zouboulis CC, Han SH, Lee JH, Lee JY. Licochalcone A attenuates acne symptoms mediated by suppression of NLRP3 inflammasome. Phytother Res. 2018; 32:2551–59. 10.1002/ptr.6195 [DOI] [PubMed] [Google Scholar]
- 11.Zeng G, Shen H, Yang Y, Cai X, Xun W. Licochalcone A as a potent antitumor agent suppresses growth of human oral cancer SCC-25 cells in vitro via caspase-3 dependent pathways. Tumour Biol. 2014; 35:6549–55. 10.1007/s13277-014-1877-1 [DOI] [PubMed] [Google Scholar]
- 12.Huang B, Liu J, Ju C, Yang D, Chen G, Xu S, Zeng Y, Yan X, Wang W, Liu D, Fu S. Licochalcone A prevents the loss of dopaminergic neurons by inhibiting microglial activation in lipopolysaccharide (LPS)-induced Parkinson’s disease models. Int J Mol Sci. 2017; 18:2043. 10.3390/ijms18102043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Atienza M, Ziontz J, Cantero JL. Low-grade inflammation in the relationship between sleep disruption, dysfunctional adiposity, and cognitive decline in aging. Sleep Med Rev. 2018; 42:171–83. 10.1016/j.smrv.2018.08.002 [DOI] [PubMed] [Google Scholar]
- 14.Wang C, Zhang X, Teng Z, Zhang T, Li Y. Downregulation of PI3K/Akt/mTOR signaling pathway in curcumin-induced autophagy in APP/PS1 double transgenic mice. Eur J Pharmacol. 2014; 740:312–20. 10.1016/j.ejphar.2014.06.051 [DOI] [PubMed] [Google Scholar]
- 15.Wesseling H, Xu B, Want EJ, Holmes E, Guest PC, Karayiorgou M, Gogos JA, Bahn S. System-based proteomic and metabonomic analysis of the Df(16)A+/- mouse identifies potential miR-185 targets and molecular pathway alterations. Mol Psychiatry. 2017; 22:384–95. 10.1038/mp.2016.27 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hoffmann A, Montoro CI, Reyes Del Paso GA, Duschek S. Cerebral blood flow modulations during cognitive control in major depressive disorder. J Affect Disord. 2018; 237:118–25. 10.1016/j.jad.2018.05.011 [DOI] [PubMed] [Google Scholar]
- 17.Ogoh S, Yoo JK, Badrov MB, Parker RS, Anderson EH, Wiblin JL, North CS, Suris A, Fu Q. Cerebral blood flow regulation and cognitive function in women with posttraumatic stress disorder. J Appl Physiol (1985). 2018; 125:1627–35. 10.1152/japplphysiol.00502.2018 [DOI] [PubMed] [Google Scholar]
- 18.Han H, Zhang R, Liu G, Qiao H, Chen Z, Liu Y, Chen X, Li D, Wang Y, Zhao X. Reduction of cerebral blood flow in community-based adults with subclinical cerebrovascular atherosclerosis: a 3.0T magnetic resonance imaging study. Neuroimage. 2019; 188:302–08. 10.1016/j.neuroimage.2018.12.021 [DOI] [PubMed] [Google Scholar]
- 19.Ikram MA, Zonneveld HI, Roshchupkin G, Smith AV, Franco OH, Sigurdsson S, van Duijn C, Uitterlinden AG, Launer LJ, Vernooij MW, Gudnason V, Adams HH. Heritability and genome-wide associations studies of cerebral blood flow in the general population. J Cereb Blood Flow Metab. 2018; 38:1598–608. 10.1177/0271678X17715861 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jefferson AL, Cambronero FE, Liu D, Moore EE, Neal JE, Terry JG, Nair S, Pechman KR, Rane S, Davis LT, Gifford KA, Hohman TJ, Bell SP, et al. Higher Aortic Stiffness Is Related to Lower Cerebral Blood Flow and Preserved Cerebrovascular Reactivity in Older Adults. Circulation. 2018; 138:1951–62. 10.1161/CIRCULATIONAHA.118.032410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Komiyama Y, Nakae S, Matsuki T, Nambu A, Ishigame H, Kakuta S, Sudo K, Iwakura Y. IL-17 plays an important role in the development of experimental autoimmune encephalomyelitis. J Immunol. 2006; 177:566–73. 10.4049/jimmunol.177.1.566 [DOI] [PubMed] [Google Scholar]
- 22.Flynn SM, Chen C, Artan M, Barratt S, Crisp A, Nelson GM, Peak-Chew SY, Begum F, Skehel M, de Bono M. MALT-1 mediates IL-17 neural signaling to regulate C. elegans behavior, immunity and longevity. Nat Commun. 2020; 11:2099. 10.1038/s41467-020-15872-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Upadhyay R, Dua B, Sharma B, Natrajan M, Jain AK, Kithiganahalli Narayanaswamy B, Joshi B. Transcription factors STAT-4, STAT-6 and CREB regulate Th1/Th2 response in leprosy patients: effect of M. leprae antigens. BMC Infect Dis. 2019; 19:52. 10.1186/s12879-018-3601-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Macor S, Porrini AM, Giampietro A, Malatesta G, Gambi D. Multiple sclerosis: an immune system activation disease. Acta Neurol (Napoli). 1991; 13:590–96. [PubMed] [Google Scholar]
- 25.Kastrukoff LF, Morgan NG, Aziz TM, Zecchini D, Berkowitz J, Paty DW. Natural killer (NK) cells in chronic progressive multiple sclerosis patients treated with lymphoblastoid interferon. J Neuroimmunol. 1988; 20:15–23. 10.1016/0165-5728(88)90109-9 [DOI] [PubMed] [Google Scholar]
- 26.Camarillo GF, Goyon EI, Zuñiga RB, Salas LAS, Escárcega AEP, Yamamoto-Furusho JK. Gene Expression Profiling of Mediators Associated with the Inflammatory Pathways in the Intestinal Tissue from Patients with Ulcerative Colitis. Mediators Inflamm. 2020; 2020:9238970. 10.1155/2020/9238970 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Riley RL, Khomtchouk K, Blomberg BB. Inflammatory immune cells may impair the preBCR checkpoint, reduce new B cell production, and alter the antibody repertoire in old age. Exp Gerontol. 2018; 105:87–93. 10.1016/j.exger.2018.01.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Maghazachi AA. Role of chemokines in the biology of natural killer cells. Curr Top Microbiol Immunol. 2010; 341:37–58. 10.1007/82_2010_20 [DOI] [PubMed] [Google Scholar]
- 29.Xue G, Cheng Y, Ran F, Li X, Huang T, Yang Y, Zhang Y. SLC gene-modified dendritic cells mediate T cell-dependent anti-gastric cancer immune responses in vitro. Oncol Rep. 2013; 29:595–604. 10.3892/or.2012.2154 [DOI] [PubMed] [Google Scholar]
- 30.Tsuneto M, Tokoyoda K, Kajikhina E, Hauser AE, Hara T, Tani-Ichi S, Ikuta K, Melchers F. B-cell progenitors and precursors change their microenvironment in fetal liver during early development. Stem Cells. 2013; 31:2800–12. 10.1002/stem.1421 [DOI] [PubMed] [Google Scholar]
- 31.Robertson MJ, Williams BT, Christopherson K 2nd, Brahmi Z, Hromas R. Regulation of human natural killer cell migration and proliferation by the exodus subfamily of CC chemokines. Cell Immunol. 2000; 199:8–14. 10.1006/cimm.1999.1601 [DOI] [PubMed] [Google Scholar]
- 32.Witherden DA, Verdino P, Rieder SE, Garijo O, Mills RE, Teyton L, Fischer WH, Wilson IA, Havran WL. The junctional adhesion molecule JAML is a costimulatory receptor for epithelial gammadelta T cell activation. Science. 2010; 329:1205–10. 10.1126/science.1192698 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Abreu-Velez AM, Valencia-Yepes CA, Upegui-Zapata YA, Upegui-Quiceno E, Mesa-Herrera NR, Velazquez-Velez JE, Howard MS. Patients with a new variant of endemic pemphigus foliaceus have autoantibodies against arrector pili muscle, colocalizing with MYZAP, p0071, desmoplakins 1 and 2 and ARVCF. Clin Exp Dermatol. 2017; 42:874–80. 10.1111/ced.13214 [DOI] [PubMed] [Google Scholar]
- 34.Rajavelu P, Chen G, Xu Y, Kitzmiller JA, Korfhagen TR, Whitsett JA. Airway epithelial SPDEF integrates goblet cell differentiation and pulmonary Th2 inflammation. J Clin Invest. 2015; 125:2021–31. 10.1172/JCI79422 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Maeda Y, Chen G, Xu Y, Haitchi HM, Du L, Keiser AR, Howarth PH, Davies DE, Holgate ST, Whitsett JA. Airway epithelial transcription factor NK2 homeobox 1 inhibits mucous cell metaplasia and Th2 inflammation. Am J Respir Crit Care Med. 2011; 184:421–29. 10.1164/rccm.201101-0106OC [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bo W, Mei K, Zhaoming W, Hongtian Y, Yanfeng B, Xiaoying N. CD79a positive T cell lymphoma with bone marrow involvement. Pathology. 2011; 43:737–40. 10.1097/PAT.0b013e32834c726f [DOI] [PubMed] [Google Scholar]
- 37.Liu X, Li YS, Shinton SA, Rhodes J, Tang L, Feng H, Jette CA, Look AT, Hayakawa K, Hardy RR. Zebrafish B cell development without a pre-B cell stage, revealed by CD79 fluorescence reporter transgenes. J Immunol. 2017; 199:1706–15. 10.4049/jimmunol.1700552 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hwang JT, Kelly GM. GATA6 and FOXA2 regulate Wnt6 expression during extraembryonic endoderm formation. Stem Cells Dev. 2012; 21:3220–32. 10.1089/scd.2011.0492 [DOI] [PubMed] [Google Scholar]
- 39.Zheng XL, Yu HG. Wnt6 contributes tumorigenesis and development of colon cancer via its effects on cell proliferation, apoptosis, cell-cycle and migration. Oncol Lett. 2018; 16:1163–72. 10.3892/ol.2018.8729 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Itoh M, Kawagoe S, Okano HJ, Nakagawa H. Integration-free T cell-derived human induced pluripotent stem cells (iPSCs) from a patient with lymphedema-distichiasis syndrome (LDS) carrying an insertion-deletion complex mutation in the FOXC2 gene. Stem Cell Res. 2016; 16:611–3. 10.1016/j.scr.2016.03.004 [DOI] [PubMed] [Google Scholar]
- 41.Yang C, Cui X, Dai X, Liao W. Downregulation of Foxc2 enhances apoptosis induced by 5-fluorouracil through activation of MAPK and AKT pathways in colorectal cancer. Oncol Lett. 2016; 11:1549–54. 10.3892/ol.2016.4097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lin L, Peng SL. Coordination of NF-kappaB and NFAT antagonism by the forkhead transcription factor Foxd1. J Immunol. 2006; 176:4793–803. 10.4049/jimmunol.176.8.4793 [DOI] [PubMed] [Google Scholar]
- 43.Nagel S, Meyer C, Kaufmann M, Drexler HG, MacLeod RA. Deregulated FOX genes in Hodgkin lymphoma. Genes Chromosomes Cancer. 2014; 53:917–33. 10.1002/gcc.22204 [DOI] [PubMed] [Google Scholar]
- 44.Xu B, Ma R, Russell L, Yoo JY, Han J, Cui H, Yi P, Zhang J, Nakashima H, Dai H, Chiocca EA, Kaur B, Caligiuri MA, Yu J. An oncolytic herpesvirus expressing E-cadherin improves survival in mouse models of glioblastoma. Nat Biotechnol. 2019; 37:45–54. 10.1038/nbt.4302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Jacobs G, Hellmig S, Huse K, Titz A, Franke A, Kwiatkowski R, Ott S, Kosmahl M, Fischbach W, Lucius R, Klapper W, Fölsch UR, Hampe J, et al. Polymorphisms in the 3'-untranslated region of the CDH1 gene are a risk factor for primary gastric diffuse large B-cell lymphoma. Haematologica. 2011; 96:987–95. 10.3324/haematol.2010.033126 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Carè A, Testa U, Bassani A, Tritarelli E, Montesoro E, Samoggia P, Cianetti L, Peschle C. Coordinate expression and proliferative role of HOXB genes in activated adult T lymphocytes. Mol Cell Biol. 1994; 14:4872–77. 10.1128/mcb.14.7.4872 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ko KH, Lam QL, Zhang M, Wong CK, Lo CK, Kahmeyer-Gabbe M, Tsang WH, Tsang SL, Chan LC, Sham MH, Lu L. Hoxb3 deficiency impairs B lymphopoiesis in mouse bone marrow. Exp Hematol. 2007; 35:465–75. 10.1016/j.exphem.2006.10.014 [DOI] [PubMed] [Google Scholar]
- 48.Sauvageau G, Thorsteinsdottir U, Hough MR, Hugo P, Lawrence HJ, Largman C, Humphries RK. Overexpression of HOXB3 in hematopoietic cells causes defective lymphoid development and progressive myeloproliferation. Immunity. 1997; 6:13–22. 10.1016/s1074-7613(00)80238-1 [DOI] [PubMed] [Google Scholar]
- 49.Stik G, Graf T. Hoxb5, a Trojan horse to generate T cells. Nat Immunol. 2018; 19:210–12. 10.1038/s41590-018-0053-y [DOI] [PubMed] [Google Scholar]
- 50.Ebihara T, Seo W, Taniuchi I. Roles of RUNX complexes in immune cell development. Adv Exp Med Biol. 2017; 962:395–413. 10.1007/978-981-10-3233-2_24 [DOI] [PubMed] [Google Scholar]
- 51.Mizutani S, Yoshida T, Zhao X, Nimer SD, Taniwaki M, Okuda T. Loss of RUNX1/AML1 arginine-methylation impairs peripheral T cell homeostasis. Br J Haematol. 2015; 170:859–73. 10.1111/bjh.13499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhang PP, Wang YC, Cheng C, Zhang F, Ding DZ, Chen DK. Runt-related transcription factor 2 influences cell adhesion-mediated drug resistance and cell proliferation in B-cell non-Hodgkin’s lymphoma and multiple myeloma. Leuk Res. 2020; 92:106340. 10.1016/j.leukres.2020.106340 [DOI] [PubMed] [Google Scholar]
- 53.Boto P, Csuth TI, Szatmari I. RUNX3-mediated immune cell development and maturation. Crit Rev Immunol. 2018; 38:63–78. 10.1615/CritRevImmunol.2018025488 [DOI] [PubMed] [Google Scholar]
- 54.Zhou C, Yang X, Sun Y, Yu H, Zhang Y, Jin Y. Comprehensive profiling reveals mechanisms of SOX2-mediated cell fate specification in human ESCs and NPCs. Cell Res. 2016; 26:171–89. 10.1038/cr.2016.15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Toda T, Hsu JY, Linker SB, Hu L, Schafer ST, Mertens J, Jacinto FV, Hetzer MW, Gage FH. Nup153 interacts with Sox2 to enable bimodal gene regulation and maintenance of neural progenitor cells. Cell Stem Cell. 2017; 21:618–34.e7. 10.1016/j.stem.2017.08.012 [DOI] [PubMed] [Google Scholar]
- 56.Wege AK, Schmidt M, Ueberham E, Ponnath M, Ortmann O, Brockhoff G, Lehmann J. Co-transplantation of human hematopoietic stem cells and human breast cancer cells in NSG mice: a novel approach to generate tumor cell specific human antibodies. MAbs. 2014; 6:968–77. 10.4161/mabs.29111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Radjavi A, Smirnov I, Derecki N, Kipnis J. Dynamics of the meningeal CD4(+) T-cell repertoire are defined by the cervical lymph nodes and facilitate cognitive task performance in mice. Mol Psychiatry. 2014; 19:531–33. 10.1038/mp.2013.79 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Poinsatte K, Smith EE, Torres VO, Ortega SB, Huebinger RM, Cullum CM, Monson NL, Zhang R, Stowe AM. T and B cell subsets differentially correlate with amyloid deposition and neurocognitive function in patients with amnestic mild cognitive impairment after one year of physical activity. Exerc Immunol Rev. 2019; 25:34–49. [PMC free article] [PubMed] [Google Scholar]
- 59.Ji M, Xie XX, Liu DQ, Yu XL, Zhang Y, Zhang LX, Wang SW, Huang YR, Liu RT. Hepatitis B core VLP-based mis-disordered tau vaccine elicits strong immune response and alleviates cognitive deficits and neuropathology progression in Tau.P301S mouse model of Alzheimer's disease and frontotemporal dementia. Alzheimers Res Ther. 2018; 10:55. 10.1186/s13195-018-0378-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mouro FM, Ribeiro JA, Sebastião AM, Dawson N. Chronic, intermittent treatment with a cannabinoid receptor agonist impairs recognition memory and brain network functional connectivity. J Neurochem. 2018; 147:71–83. 10.1111/jnc.14549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Qi F, Yang J, Xia Y, Yuan Q, Guo K, Zou J, Yao Z. A(H1N1) vaccination recruits T lymphocytes to the choroid plexus for the promotion of hippocampal neurogenesis and working memory in pregnant mice. Brain Behav Immun. 2016; 53:72–83. 10.1016/j.bbi.2015.11.006 [DOI] [PubMed] [Google Scholar]
- 62.Sîrbulescu RF, Chung JY, Edmiston WJ 3rd, Poznansky SA, Poznansky MC, Whalen MJ. Intraparenchymal application of mature B lymphocytes improves structural and functional outcome after contusion traumatic brain injury. J Neurotrauma. 2019; 36:2579–89. 10.1089/neu.2018.6368 [DOI] [PubMed] [Google Scholar]
- 63.Zhang Y, Zhang Y, Gu W, Sun B. TH1/TH2 cell differentiation and molecular signals. Adv Exp Med Biol. 2014; 841:15–44. 10.1007/978-94-017-9487-9_2 [DOI] [PubMed] [Google Scholar]
- 64.Lund RJ, Löytömäki M, Naumanen T, Dixon C, Chen Z, Ahlfors H, Tuomela S, Tahvanainen J, Scheinin J, Henttinen T, Rasool O, Lahesmaa R. Genome-wide identification of novel genes involved in early Th1 and Th2 cell differentiation. J Immunol. 2007; 178:3648–60. 10.4049/jimmunol.178.6.3648 [DOI] [PubMed] [Google Scholar]
- 65.Stables MJ, Shah S, Camon EB, Lovering RC, Newson J, Bystrom J, Farrow S, Gilroy DW. Transcriptomic analyses of murine resolution-phase macrophages. Blood. 2011; 118:e192–208. 10.1182/blood-2011-04-345330 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Sogkas G, Klose K, Baerlecken N, Schweikhard E, Matthias T, Kniesch K, Schmidt RE, Witte T. CD74 is a T cell antigen in spondyloarthritis. Clin Exp Rheumatol. 2020; 38:195–202. [PubMed] [Google Scholar]
- 67.Stein R, Mattes MJ, Cardillo TM, Hansen HJ, Chang CH, Burton J, Govindan S, Goldenberg DM. CD74: a new candidate target for the immunotherapy of B-cell neoplasms. Clin Cancer Res. 2007; 13:5556s–63s. 10.1158/1078-0432.CCR-07-1167 [DOI] [PubMed] [Google Scholar]
- 68.Wei JP, Wen W, Dai Y, Qin LX, Wen YQ, Duan DD, Xu SJ. Drinking water temperature affects cognitive function and progression of Alzheimer’s disease in a mouse model. Acta Pharmacol Sin. 2021; 42:45–54. 10.1038/s41401-020-0407-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Fogel MA, Li C, Elci OU, Pawlowski T, Schwab PJ, Wilson F, Nicolson SC, Montenegro LM, Diaz L, Spray TL, Gaynor JW, Fuller S, Mascio C, et al. Neurological injury and cerebral blood flow in single ventricles throughout staged surgical reconstruction. Circulation. 2017; 135:671–82. 10.1161/CIRCULATIONAHA.116.021724 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Li X, Bechara R, Zhao J, McGeachy MJ, Gaffen SL. IL-17 receptor-based signaling and implications for disease. Nat Immunol. 2019; 20:1594–602. 10.1038/s41590-019-0514-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Li T, Zhang Y, Han D, Hua R, Guo B, Hu S, Yan X, Xu T. Involvement of IL-17 in secodary brain injury after a traumatic brain injury in rats. Neuromolecular Med. 2017; 19:541–54. 10.1007/s2017-017-8468-4 [DOI] [PubMed] [Google Scholar]
- 72.Chen C, Itakura E, Nelson GM, Sheng M, Laurent P, Fenk LA, Butcher RA, Hegde RS, de Bono M. IL-17 is a neuromodulator of caenorhabditis elegans sensory responses. Nature. 2017; 542:43–48. 10.1038/nature20818 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Song L, Ma S, Chen L, Miao L, Tao M, Liu H. Long-term prognostic significance of interleukin-17-producing T cells in patients with non-small cell lung cancer. Cancer Sci. 2019; 110:2100–09. 10.1111/cas.14068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Park JS, Kim NR, Lim MA, Kim SM, Hwang SH, Jung KA, Choi J, Park SH, Cho ML. Deficiency of IL-1 receptor antagonist suppresses IL-10-producing B cells in autoimmune arthritis in an IL-17/Th17-dependent manner. Immunol Lett. 2018; 199:44–52. 10.1016/j.imlet.2018.05.003 [DOI] [PubMed] [Google Scholar]
- 75.Zhang CJ, Wang C, Jiang M, Gu C, Xiao J, Chen X, Martin BN, Tang F, Yamamoto E, Xian Y, Wang H, Li F, Sartor RB, et al. Act1 is a negative regulator in T and B cells via direct inhibition of STAT3. Nat Commun. 2018; 9:2745. 10.1038/s41467-018-04974-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Chen R, Wang M, Liu Q, Wu J, Huang W, Li X, Du B, Xu Q, Duan J, Jiao S, Lee HS, Jung NC, Lee JH, et al. Sequential treatment with aT19 cells generates memory CAR-T cells and prolongs the lifespan of Raji-B-NDG mice. Cancer Lett. 2020; 469:162–72. 10.1016/j.canlet.2019.10.022 [DOI] [PubMed] [Google Scholar]
- 77.Chen L, Yao Y, Wei C, Sun Y, Ma X, Zhang R, Xu X, Hao J. T cell immunity to glatiramer acetate ameliorates cognitive deficits induced by chronic cerebral hypoperfusion by modulating the microenvironment. Sci Rep. 2015; 5:14308. 10.1038/srep14308 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lueg G, Gross CC, Lohmann H, Johnen A, Kemmling A, Deppe M, Groger J, Minnerup J, Wiendl H, Meuth SG, Duning T. Clinical relevance of specific T-cell activation in the blood and cerebrospinal fluid of patients with mild Alzheimer’s disease. Neurobiol Aging. 2015; 36:81–89. 10.1016/j.neurobiolaging.2014.08.008 [DOI] [PubMed] [Google Scholar]
- 79.Ortega SB, Torres VO, Latchney SE, Whoolery CW, Noorbhai IZ, Poinsatte K, Selvaraj UM, Benson MA, Meeuwissen AJ, Plautz EJ, Kong X, Ramirez DM, Ajay AD, et al. B cells migrate into remote brain areas and support neurogenesis and functional recovery after focal stroke in mice. Proc Natl Acad Sci USA. 2020; 117:4983–93. 10.1073/pnas.1913292117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Zhao H, Jiang JT, Zheng QS. [Advance in studies on pharmacological effects of licochalcone A]. Zhongguo Zhong Yao Za Zhi. 2013; 38:3814–18. [PubMed] [Google Scholar]
- 81.Idrees M, Khan S, Memon NH, Zhang Z. Effect of the phytochemical agents against the SARS-CoV and selected some of them for application to COVID-19: a mini-review. Curr Pharm Biotechnol. 2020. [Epub ahead of print]. 10.2174/1389201021666200703201458 [DOI] [PubMed] [Google Scholar]
- 82.Fan T, Chen Y, Bai Y, Ma F, Wang H, Yang Y, Chen J, Lin Y. [Analysis of medication characteristics of traditional Chinese medicine in treating COVID-19 based on data mining]. Zhejiang Da Xue Xue Bao Yi Xue Ban. 2020; 49:260–69. 10.3785/j.issn.1008-9292.2020.03.14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Zhu T, Zhang F, Li H, He Y, Zhang G, Huang N, Guo M, Li X. Long-term icariin treatment ameliorates cognitive deficits via CD4+ T cell-mediated immuno-inflammatory responses in APP/PS1 mice. Clin Interv Aging. 2019; 14:817–26. 10.2147/CIA.S208068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Jiang R, Gao J, Shen J, Zhu X, Wang H, Feng S, Huang C, Shen H, Liu H. Glycyrrhizic acid improves cognitive levels of aging mice by regulating T/B cell proliferation. Front Aging Neurosci. 2020; 12:570116. 10.3389/fnagi.2020.570116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Espinoza JL, Trung LQ, Inaoka PT, Yamada K, An DT, Mizuno S, Nakao S, Takami A. The repeated administration of resveratrol has measurable effects on circulating T-cell subsets in humans. Oxid Med Cell Longev. 2017; 2017:6781872. 10.1155/2017/6781872 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.He J, Yu Q, Wu C, Sun Z, Wu X, Liu R, Zhang H. Acupuncture of the beishu acupoint participates in regulatory effects of ginsenoside Rg1 on T cell subsets of rats with chronic fatigue syndrome. Ann Palliat Med. 2020; 9:3436–46. 10.21037/apm-20-1714 [DOI] [PubMed] [Google Scholar]
- 87.Zhu X, Shen J, Feng S, Huang C, Liu Z, Sun YE, Liu H. Metformin improves cognition of aged mice by promoting cerebral angiogenesis and neurogenesis. Aging (Albany NY). 2020; 12:17845–62. 10.18632/aging.103693 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Wang K, Singh D, Zeng Z, Coleman SJ, Huang Y, Savich GL, He X, Mieczkowski P, Grimm SA, Perou CM, MacLeod JN, Chiang DY, Prins JF, Liu J. MapSplice: accurate mapping of RNA-seq reads for splice junction discovery. Nucleic Acids Res. 2010; 38:e178. 10.1093/nar/gkq622 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Ennaceur A, Delacour J. A new one-trial test for neurobiological studies of memory in rats. 1: behavioral data. Behav Brain Res. 1988; 31:47–59. 10.1016/0166-4328(88)90157-x [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.