

Abstract
The G protein‐coupled free fatty acid receptor 2 (FFA2R) is highly expressed on neutrophils and was previously described to regulate neutrophil activation. Allosteric targeting of G protein‐coupled receptors (GPCRs) is increasingly explored to create distinct pharmacology compared to endogenous, orthosteric ligands. The consequence of allosteric versus orthosteric FFA2R activation for neutrophil response, however, is currently largely elusive. Here, different FFA2R desensitization profiles in human neutrophils following allosteric or orthosteric activation are reported. Using a set of neutrophil functional assays to measure calcium flux, pERK1/2, chemotaxis, cellular degranulation, and oxidative burst together with holistic and pathway‐unbiased whole cell sensing based on dynamic mass redistribution, it is found that the synthetic positive allosteric modulator agonist 4‐CMTB potently activates neutrophils and simultaneously alters FFA2R responsiveness toward the endogenous, orthosteric agonist propionic acid (C3) after homologous and heterologous receptor desensitization. Stimulation with C3 or the hierarchically superior chemokine receptor activator IL‐8 led to strong FFA2R desensitization and rendered neutrophils unresponsive toward repeated stimulation with C3. In contrast, stimulation with allosteric 4‐CMTB engaged a distinct composition of signaling pathways as compared to orthosteric receptor activation and was able to activate neutrophils that underwent homologous and heterologous desensitization with C3 and IL‐8, respectively. Moreover, allosteric FFA2R activation could re‐sensitize FFA2 toward the endogenous agonist C3 after homologous and heterologous desensitization. Given the fact that receptor desensitization is critical in neutrophils to sense and adapt to their current environment, these findings are expected to be useful for the discovery of novel pharmacological mechanisms to modulate neutrophil responsiveness therapeutically.
Keywords: DMR, GPCR, GPR43, neutrophils, PAM, rescue
Graphical Abstract
The selective FFA2 receptor ago‐PAM 4‐CMTB re‐sensitizes human neutrophils toward orthosteric FFA2 activation.
Abbreviations
- C3
propionic acid
- DAMP
danger‐associated molecular pattern
- DMR
dynamic mass redistribution
- FFA2R
free fatty acid receptor 2
- GPCR
G protein‐coupled receptor
- HDAC
histone deacetylase
- PAM
positive allosteric modulator
- PAMP
pathogen‐associated molecular pattern
- PTX
pertussis toxin
- ROS
reactive oxygen species
- SCFA
short‐chain fatty acids
1. INTRODUCTION
Neutrophilic granulocytes are the most abundant leukocytes in circulation and belong to the primal armamentarium against both septic and aseptic infections in humans. They are guided to sites of inflammation or infection by sensing their environment through cell surface receptors, which aid in controlling neutrophil activity in situ to respond to local stimuli. Activated neutrophils clear the affected tissue from invading microbes by either phagocytic activity, release of toxic agents, such as reactive oxygen species (ROS) or by coordinating with other immune cells. 1 , 2 Thus, together with monocytes, neutrophilic granulocytes are key actors in inflammation resolution. An imbalance in neutrophil response results in either insufficient tissue clearing of the causative source or exaggerated immune reaction eventually leading to detrimental unresolved inflammation. 1 , 3
Neutrophil recruitment and activation are subjected to complex biochemical mechanisms involving release and recognition of cytokines as well as danger‐ and pathogen‐associated molecular patterns (DAMPS and PAMPs, respectively). 4 In an inflammatory surrounding, a fine‐tuned regulation of immune cell receptor responsiveness and receptor signaling is particularly important. G protein‐coupled receptors (GPCRs) are essential neutrophil cell surface receptors that process environmental cues to allow leukocyte recruitment and activation. 4 Beside the well‐established neutrophil GPCRs (formyl peptide receptors [FPRs], chemokine receptors [CXCRs], classical chemoattractant receptors [complement receptor C5a, leukotriene receptor LTB4, platelet‐activating factor receptor PAFR]), also the free fatty acid receptor FFA2 (formerly GPR43) is highly expressed on neutrophils and is reported to be involved in inflammatory responses. 5 , 6
Short‐chain fatty acids (SCFA, such as acetate, propionate, and butyrate) mainly derived from the gut microbiota through digestion of dietary fiber are the endogenous ligands of the FFA2 receptor acting as orthosteric agonists with micromolar potency. 5 , 6 SCFA can reach excessively high amounts at the site of generation in the gut but are also found in the systemic circulation at micromolar concentrations. 7 The exact role of the FFA2 receptor in the regulation of neutrophil response especially in context of other neutrophil stimuli is so far only insufficiently understood in part due to a lack of FFA2 receptor specific ligands. The tool box to study FFA2R functionality in neutrophils in more detail was broadened by the discovery of the FFA2R‐specific allosteric agonist 4‐CMTB 8 and the FFA2R‐selective orthosteric antagonist CATPB, 9 , 10 enabling the experimental exploration of site‐specific FFA2 receptor signaling. Whether allosteric as compared to orthosteric FFA2R activation has distinct consequences for neutrophil response is, however, currently still unknown. We here investigated FFA2R signaling downstream of allosteric versus orthosteric activation in calcium release assays as well as in phenotypic label‐free measurements of neutrophil cytoskeletal rearrangement, ERK1/2 phosphorylation, receptor internalization, and immune response assays, determining neutrophil migratory and adhesion properties as well as degranulation and ROS production. We found the regulation and signaling fingerprint of FFA2R activation in neutrophils to be dependent on the ligand mode, that is, orthosteric or allosteric targeting, which led to adapted neutrophil immunological functions and modulation of neutrophil responsiveness after desensitization.
2. MATERIALS AND METHODS
2.1. Chemicals and reagents
All chemicals and reagents were purchased from Sigma–Aldrich, unless specified otherwise. Assay buffer for functional neutrophil assays was made from Dulbecco`s modified PBS (with 0.9 mmol/L Ca2+ and 0.5 mmol/L Mg2+; 0.1% BSA, 10 mmol/L HEPES, and 10 mmol/L glucose, pH 7.4). HBSS buffer for DMR measurements was from ThermoFisher. Anti‐CD16‐PE‐Cy5(3G8), anti‐CD63‐FITC(H5C6) and anti‐CD11b‐PE(ICRF44) were from Becton Dickinson (Vienna, Austria). CellFix and FACS‐Flow were from Becton Dickinson (Vienna, Austria). PVP‐free polycarbonate filters were from NeuroProbe (Gaithersburg, USA). Fluo‐3‐AM was supplied from Life Sciences (Vienna, Austria). Fixative solution was prepared by adding 9 ml of distilled water and 30 ml of FACS‐Flow to 1 ml of CellFix. DMR experiments were performed using the Epic® System (Corning, USA) in fibronectin‐coated cell‐based biosensor plates (Corning, USA). FR900359 was a kind gift of Evi Kostenis and Gabriele König (University of Bonn, Germany).
2.2. Preparation of human neutrophils
Blood was taken from healthy volunteers according to a protocol approved by the Ethics Committee of the Medical University of Graz and processed as described previously. 11 In short, polymorphonuclear leukocytes (PMNL) were prepared by means of dextran sedimentation of erythrocytes followed by centrifugation on a Histopaque‐1077 layer.
2.3. Chemotaxis assay
PMNL were pretreated as indicated, placed into the top wells of a 5 µm pore‐sized 48‐well micro‐Boyden chamber and were allowed to migrate toward the indicated chemoattractant for 1 h at 37°C. Migrated cells in the bottom wells were enumerated by flow cytometry and neutrophils were identified from eosinophils according to their lower auto‐fluorescence in the FL‐2 (585/42 nm) channel.
2.4. Calcium measurements
PMNL (106 cells/ml) were treated with 2 µM of Fluo‐3‐AM in the presence of 0.02% pluronic F‐127 for 1 h at room temperature. Cells were pretreated as indicated and stimulated with the respective agonist. Samples were stained with anti‐CD16‐PE‐Cy5 and neutrophils were distinguished from eosinophils as CD16+ cells. Changes in intracellular Ca2+ were detected as fluorescence increase of the Ca2+ sensitive dye Fluo‐3 in the FL1‐(530/30 nm) channel. For antagonist measurements, cells were pre‐incubated with CATPB for 30 min before agonist addition.
2.5. ERK1/2 phosphorylation assay
PMNL (106 cells/ml) were treated with either vehicle or FFA2 agonist for 5 min at room temperature. Neutrophils were subsequently stimulated for another 5 min with the indicated compounds at room temperature and immediately stopped by addition of 4× volume of ice‐cold PBS. The cells were centrifuged at 400 × g at 4°C for 5 min and suspended in 300 µl of lysis buffer (CellSignaling #9803) supplemented with protease and phosphatase inhibitors. Samples were then centrifuged at 16,000 × g at 4°C for 20 min. Supernatant was used in an ELISA detecting total (Cell Signaling #7050) and phosphor‐ERK1/2 (Thr202/Tyr204) (Cell Signaling #7177) according to manufacturer's instructions.
2.6. CD11b measurements
PMNL were pretreated for 5 min at room temperature as indicated and incubated with agonists for 30 min at 37°C. Samples were stained with anti‐CD16‐PE‐Cy5 and anti‐CD11b‐PE (ICRF44) Ab and neutrophils were distinguished from eosinophils as CD16+ cells. CD11b up‐regulation was analyzed as fluorescence increase in the FL‐2 (585/42 nm) channel by flow cytometry and expressed as percent of the vehicle response.
2.7. Respiratory burst
PMNL were stained with anti‐CD16‐PE‐Cy5 pretreated as indicated and cells were stimulated with serial dilutions of the respective agonist in the presence of 50 µM DCFDA for 20 min at 37°C. Respiratory burst was immediately quantified by flow cytometry as the increase in the FL‐1 (530/30 nm) channel due to the conversion of DCFDA into DCF by reactive oxygen. Neutrophils were identified as CD‐16+ cells and responses were expressed as percent of the vehicle response.
2.8. CD63 measurements
PMNL were pretreated for 5 min at RT as indicated and labeled with anti‐CD63‐FITC Ab to assess degranulation and anti‐CD16‐PE‐Cy5 to identify neutrophils. Samples were treated with cytochalasin B (5 µg/ml) for 5 min at 37°C and degranulation was induced with serial dilutions of C3 or 4‐CMTB for 20 min at 37°C. CD63 expression was detected in the FL‐1 (530/30 nm) channel by flow cytometry and expressed as percent of the respective control response.
2.9. Dynamic mass redistribution measurements
Dynamic mass redistribution (DMR) measurements were performed in suspension mode as previously described in detail. 12 Briefly, 60,000 freshly isolated neutrophils per well were seeded into a 384‐well biosensor plate in HBSS buffer containing 20 mM HEPES, centrifuged for 30 s at 150 × g and incubated at 28°C for 2 h in the DMR reader before the measurement. Compounds were diluted in HBSS (+20 mM HEPES), added to the biosensor plate after 3 min of baseline read with a liquid handling system under continuous DMR recording. Where indicated, cells were preincubated with G protein inhibitors for 1 h prior to DMR measurements.
2.10. Receptor internalization
A total of 2 × 104 PMNLs/well were seeded in 50 µl RPMI medium in a 96‐well CellBind plate (Corning) and let adhere for 30 min at 37°C. Cells were incubated for 5 min (+5 min pre‐incubation) with compounds diluted in RPMI containing 0.1% BSA at 37°C. Reaction was stopped by addition of 100 µl ice‐cold PBS. Supernatant was carefully discarded and wells were washed with 200 µl PBS/well for 5 min under slight shaking. Cells were fixed with 4% PFA for 15 min and subsequently washed twice with 200 µl PBS/well for 5 min. Each well was incubated for 20 min at room temperature with 100 µl of quenching solution (1% H2O2 in PBS). After 2 washing steps (200 µl PBS, 5 min), blocking buffer (10% FBS in PBS) was added, incubated for 1 h at room temperature and washed again once with 200 µl PBS for 5 min. Cells were incubated with anti‐human FFA2R antibody (abcam #124272) at a dilution of 1:1000 overnight at 4°C in 50 µl blocking buffer. After 3 washing steps with 200 µl PBS for 5 min, secondary Ab (Jackson #111‐035‐003) at a dilution of 1:1000 in 50 µl blocking buffer was added and incubated for 60 min at room temperature under constant shaking. Subsequently, cells were washed 3 times (200 µl PBS, 5 min) and incubated with 100 µl TMB substrat (Raybiotech) for 20 min and signal was measured at 450 nm at a Tecan multimode reader (Tecan infinite 200).
2.11. Data analysis and statistics
All graphs were plotted and statistically analyzed as indicated using GraphPad Prism 8 (GraphPad, USA).
3. RESULTS
3.1. FFA2R activation leads to calcium response in human neutrophils
To study the consequences of orthosteric versus allosteric FFA2R activation in neutrophilic granulocytes, we monitored calcium release in isolated human neutrophils after stimulation with the orthosteric agonist propionic acid (C3) or the FFA2R selective allosteric agonist 2‐(4‐chlorophenyl)‐3‐methyl‐N‐(thiazol‐2‐yl)butanamide (4‐CMTB). 8 Regardless of the different activation mode, both compounds led to a robust and dose‐dependent intracellular calcium response. 4‐CMTB, however, displayed higher potency and efficacy as compared to C3 (Fig. 1A and B). Receptor signal origin of C3 and 4‐CMTB from the orthosteric and an allosteric site, respectively, was confirmed by use of the selective orthosteric FFAR2 antagonist CATPB, which completely abolished C3, but not 4‐CMTB‐induced calcium release (Fig. 1C and D). In contrast to what has been found in overexpressing HEK293 cells, these data indicate that 4‐CMTB acts as a non‐orthosteric, purely allosteric FFA2R activator in primary human neutrophils, which endogenously express FFA2R. 13
FIGURE 1.
Orthosteric and allosteric FFA2R activation on neutrophils. Calcium measurements in human neutrophils upon stimulation with increasing concentrations of C3 (A) or 4‐CMTB (B). Concentration‐response‐curves from peak calcium flux induced by C3 (C) or 4‐CMTB (D) in the presence of increasing concentrations of the orthosteric FFA2 antagonist CATPB. pEC50 (C3): 2.9 ± 0.1; pEC50 (4‐CMTB): 4.47 ± 0.04. Shown are mean values + sem, measured in triplicates (A and B) and mean ± sem of 3 independent experiments (C and D)
3.2. Orthosteric and allosteric FFA2R activation result in different homologous receptor desensitization in neutrophils
To ensure a tailored cell response depending on the current biological need during the complex situation of inflammation and in the presence of abundant chemokines, immune cells developed a hierarchical system of receptor responsiveness toward various stimuli. Receptor desensitization is a key regulatory pathway that cells employ to rapidly adapt to their biochemical environment. Homologous as well as heterologous receptor (cross‐)desensitization and leukocyte priming are manifestations of such regulation. 14 To test whether neutrophils control FFA2 signaling accordingly, we activated the receptor repeatedly with equipotent concentrations of either orthosteric or allosteric FFA2R agonist and measured the ability to induce a cytosolic calcium increase. We found that pre‐stimulation with the orthosteric agonist C3 abolished responsiveness to repeated stimulation with C3 indicating strong homologous desensitization of the FFA2 receptor toward orthosteric activation, whereas allosteric pretreatment with 4‐CMTB only partially diminished the calcium response to a repeated allosteric activation (Fig. 2A and B). Moreover, neutrophils remained sensitive toward orthosteric stimulation if preceded by allosteric activation (Fig. 2C). To check whether additional pathways previously shown to be activated downstream of FFA2R in neutrophils are likewise differentially affected, we assessed ERK1/2 phosphorylation as a readout of the MAPK signaling pathway upon orthosteric and allosteric FFA2R activation. Analogous to the calcium flux, we found a similar pERK1/2 activation pattern after orthosteric and allosteric pre‐stimulation. Pre‐incubation with C3 completely abolished the response toward orthosteric activation and allosteric pre‐stimulation was largely without effect on repeated activation with 4‐CMTB. Allosteric pre‐stimulation with 4‐CMTB, however, led to a strong response toward orthosteric FFA2 activation by C3 (Fig. 2D). These data suggest that orthosteric and allosteric FFA2 activation use different mechanisms to regulate receptor responsiveness and control signaling pathways associated with the FFA2 receptor in neutrophils.
FIGURE 2.
FFA2R desensitization upon orthosteric and allosteric receptor activation. Homologous FFA2 receptor desensitization experiments in human neutrophils. Real time calcium measurements under repeated stimulation with either C3 (1 mM; A), 4‐CMTB (30 µM; B), or pre‐stimulation with 4‐CMTB (30 µM) and final stimulation with C3 (1 mM) (C). (D) ERK1/2 phosphorylation under repeated stimulation with either vehicle, C3 (1 mM) or 4‐CMTB (30 µM). Ratios of phosphorylated and total ERK1/2 level are depicted. Plotted are mean values + sem, measured in triplicates. Arrows indicate compound addition. **/**** P < 0.005/0.0001; 1‐way ANOVA with Dunnett's multiple comparison test
3.3. Allosteric activation resensitizes heterologously and homologously desensitized FFA2 receptor responsiveness toward orthosteric activation
The different mechanisms of orthosteric versus allosteric FFA2R activation in the homologous desensitization experiments were also recapitulated in receptor cross‐desensitization experiments comprising C3 and 4‐CMTB as well as the hierarchically superior CXCR1/2 receptor stimulus IL‐8. Stimulation with IL‐8 resulted in a strong calcium increase, upon which the orthosteric FFA2R agonist C3 was impotent to evoke a cell response, indicative of heterologous receptor desensitization (Fig. 3A). In contrast, 4‐CMTB generated an unabated calcium signal after IL‐8 stimulation (Fig. 3B). FFA2 stimulation with either C3 or 4‐CMTB was, however, without effect on the ability of neutrophils to respond to IL‐8 treatment, indicating that heterologous receptor desensitization occurs from CXCR1/2 to FFA2 receptors but not vice versa and corroborating a hierarchy of CXCR1/2 receptors being superior to FFA2 receptors (Fig. 3A and B, inserts).
FIGURE 3.
Rescue of orthosteric FFA2R neutrophil response by 4‐CMTB. Heterologous and homologous receptor cross‐de/resensitization experiments in human neutrophils using IL‐8 (3 nM), C3 (1 mM), and 4‐CMTB (30 µM). Real‐time calcium measurements upon repeated stimulation with different combinations of IL‐8/C3 (A), IL‐8/4‐CMTB (B), 4‐CMTB/IL‐8/C3 (C), and C3/4‐CMTB/C3 (D). Assessment of FFA2 receptor internalization after stimulation with C3 (1 mM), 4‐CMTB (30 µM), or IL‐8 (3 nM) (E) and after pre‐stimulation with orthosteric (1 mM C3) or allosteric (30µM 4‐CMTB) FFA2 agonists (F). Shown are mean values + sem, measured in triplicates. Arrows indicate compound addition. ***/**** P < 0.0005/0.0001; 1‐way ANOVA with Dunnett's multiple comparison test
We then conducted a triple stimulation experiment comprising allosteric and orthosteric FFA2R activation as well as IL‐8 priming. Intriguingly, after neutrophil pretreatment with 4‐CMTB followed by IL‐8 stimulation, that was previously sufficient to block orthosteric FFA2 activation, C3 regained the capacity to evoke a solid calcium response (Fig. 3C). Moreover, this observation was also evident after homologous orthosteric desensitization, since FFA2 receptors regained responsiveness toward C3 stimulation after desensitization with C3 and subsequent 4‐CMTB treatment (Fig. 3D). This indicates that allosteric 4‐CMTB receptor activation can function as a resensitizer for orthosteric FFA2R‐mediated neutrophil activation. To further investigate how the FFA2 receptor is regulated on neutrophils we quantified receptor internalization upon different stimuli. While FFA2R cell surface abundance was unaffected by treatment with the CXCR1/2 agonist IL‐8, both orthosteric and allosteric FFA2R activation led to marked reduction of FFA2 receptors on the plasma membrane as compared to vehicle‐treated neutrophils (Fig. 3E). Intriguingly, neutrophil pretreatment with both C3 and 4‐CMTB was not able to re‐establish initial FFA2 receptor surface expression level regardless of the second stimulus (Fig. 3F). This indicates that the observed (cross‐)desensitization as well as the resensitization of FFA2 receptors by allosteric targeting is not mediated by regulation of FFA2 receptor presence on the plasma membrane of neutrophils but rather suggests that allosteric receptor activation ensues molecular changes at the receptor or post‐receptor level to modify FFA2R sensitivity to differentially respond to ligand stimulation.
3.4. C3 and 4‐CMTB differentially engage G proteins to mediate calcium response in neutrophils
To gain a deeper understanding of the underlying mechanisms of orthosteric versus allosteric FFA2 receptor activation in human neutrophils, we inspected neutrophil calcium response under two aspects: (1) to detect a possible extracellular calcium component, we used the cation chelating agent EGTA and (2) to decipher the input of Gαi/o (hereafter Gαi) and Gαq/11/14 (hereafter Gαq) proteins, to the signaling effects downstream of the FFA2 receptor, we used the selective G protein inhibitors PTX to block Gαi, and FR900359 (FR) 15 to inhibit Gαq signaling, respectively. Both the initial peak as well as the following plateau phase of FFA2R‐mediated neutrophil calcium flux was largely unaffected by the presence of EGTA indicating no impact of extracellular calcium stores to the signal generated by C3 and 4‐CMTB (Fig. 4A). G protein pathway analysis of calcium release in neutrophils revealed that C3‐mediated response was sensitive to PTX, but not to FR pretreatment if used in isolation. The signal decrease under a combined treatment with PTX and FR was, however, significantly more pronounced compared to single Gαi protein inhibition and revealed a modest PTX‐ and FR‐insensitive calcium signal (Fig. 4B). These data indicate a primary Gαi input for orthosteric FFA2 receptor calcium signaling with subordinate input from Gαq proteins as well as a Gαi/q‐independent component. In contrast, 4‐CMTB‐induced calcium response was insensitive to either PTX, FR, or a combination of both, suggesting a substantial Gαi/q‐independent signaling mechanism downstream of allosteric FFA2 receptor activation.
FIGURE 4.
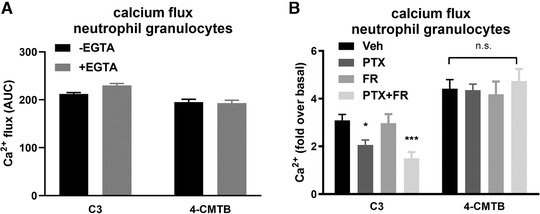
Analysis of calcium release in neutrophils by C3 and 4‐CMTB mediated FFA2R activation. Calcium release in human neutrophils in absence or presence of EGTA stimulated with C3 (3 mM) or 4‐CMTB (50 µM) (A). Quantification of calcium flux induced by C3 (3 mM) or 4‐CMTB (50 µM) in human neutrophil pretreated for 1 h with vehicle, PTX (10 µg/ml), FR (1 µM), or a combination of PTX and FR (C). Depicted are mean values + sem, measured in triplicates. */*** P < 0.05/0.001, 1‐Way‐ANOVA with Dunnett's multiple comparison test
3.5. Label‐free whole cell sensing of human neutrophils reveals distinct signaling fingerprints downstream of orthosteric and allosteric FFA2R activation
The different signal composition downstream of orthosteric and allosteric FFA2 receptor stimulation and its significance for neutrophil cell response was further analyzed in a pathway unbiased holistic cell sensor assay based on dynamic mass redistribution (DMR). The label‐free DMR assay measures cytoskeletal rearrangements in living cells in real time and thus allows the simultaneous monitoring of consequences of different signaling pathways. 16 Positive DMR signals reflect an increase in cellular mass density above an optical biosensor surface, while negative DMR signals indicate a decrease in cell bulk near the sensor. The unperturbed whole cell responses of human neutrophils stimulated with increasing concentration of C3 or 4‐CMTB indicate an extensive apparent difference between orthosteric and allosteric FFA2 receptor activation. While C3 induced a strong positive DMR signal, 4‐CMTB appeared as a weak activator of total DMR in neutrophils by showing only minor changes in relation to baseline and a tendency toward negative DMR signatures indicative for an opposite direction of net mass movement as compared to C3‐induced positive DMR (Fig. 5A and B). Application of signaling pathway inhibitors substantiated the findings from the calcium assay and revealed a dual input of Gαi and Gαq to C3‐mediated responses with a minor Gαi/q‐independent component (Fig. 5C). 4‐CMTB‐induced neutrophil mass movement comprise a substantial Gαq input to cell shape change, because FR pretreatment significantly altered the cytoskeleton rearrangement and unmasked a strong DMR response of 4‐CMTB (Fig. 5D, compare black with purple trace). PTX pre‐incubation revealed only a minor Gαi input to the overall allosteric FFA2R signaling because the respective 4‐CMTB traces showed only slight changes (Fig. 5D, compare black with green trace). A positive DMR response persisted under combined pretreatment with PTX and FR, reflecting a strong Gαi/q‐independent component of 4‐CMTB signaling in neutrophils (Fig. 5D, brown trace).
FIGURE 5.
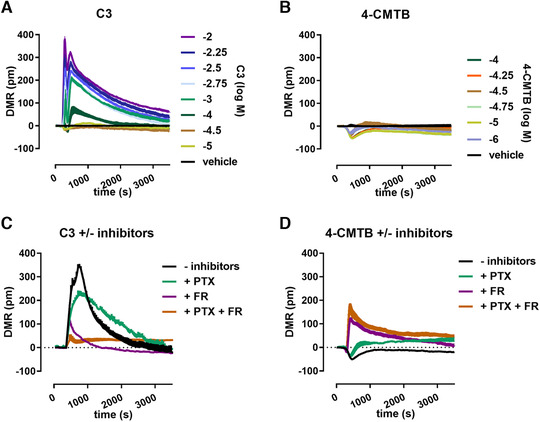
Holistic neutrophil response to FFA2R activation and analysis of underlying pathway composition. DMR recordings of human neutrophils stimulated with increasing concentrations of C3 (A) or 4‐CMTB (B). Cell response of neutrophil pretreated for 1 h with or without G protein inhibitors (3 µg/ml PTX, 1µM FR) and stimulated with 3 mM C3 (C) or 100 µM 4‐CMTB (D). Traces are depicted as mean + sem measured in triplicates, DMR signatures are corrected for response toward vehicle addition
Taken together, the data indicate that orthosteric and allosteric FFA2R activation involves Gαi and Gαq proteins as well as Gαi/q‐independent mechanisms, although to different extents. Moreover, the assembly of the individual signaling inputs to the overall activation structure in neutrophils differs between orthosteric and allosteric FFA2R activation. These differences eventually translate into the distinct composition of (1) a common signaling event, such as calcium flux, and of (2) a discrete phenotypic cell response, such as change in neutrophil morphology (DMR).
3.6. FFA2R stimulation on neutrophils does not lead to strong chemotactic responses
Based on these data, we hypothesized that the different activation mechanisms at the FFA2 receptor by C3 and 4‐CMTB translate into differences in neutrophil phenotype, namely chemotaxis, adhesion, degranulation, and oxidative burst capability. Because previous reports suggest chemotaxis as main effect of FFA2R activation on neutrophils, we assessed the migratory phenotype of neutrophils after orthosteric or allosteric FFA2 receptor activation. In line with literature data that stem from human neutrophils and FFA2R agonists, we detected maximum chemotactic indices that were comparable between C3 and 4‐CMTB and that were following a similar bell‐shaped dose‐response relationship (Fig. 6A and B). 5 , 17 The magnitude of the migratory response, however, was relatively low compared to IL‐8, which might insinuate a subordinate role of FFA2R signaling in neutrophil chemotaxis in an inflammatory surrounding. 18 To investigate whether allosteric priming affects chemotactic properties, neutrophils were prestimulated with either C3 or 4‐CMTB before monitoring of migration toward FFA2R agonists. Priming could not enhance chemotaxis toward FFA2R stimulation (1) indicating that allosteric FFA2 receptor sensitization does not affect all pathways downstream of FFA2R in neutrophils equally but differentially modulates neutrophil phenotypes and (2) strengthening the conclusion that FFA2R plays a minor role as a significant chemotaxis receptor in neutrophils.
FIGURE 6.
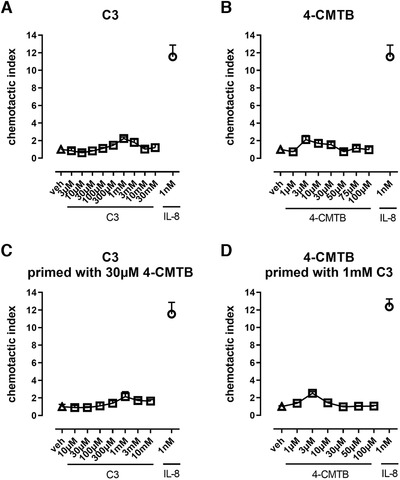
Chemotaxis of human neutrophils toward FFA2R activation. Chemotactic response of human neutrophils toward increasing concentrations of the orthosteric FFA2R agonist C3 (A) or the allosteric FFA2R agonist 4‐CMTB (B). Chemotaxis toward C3 stimulation after priming with 30 µM 4‐CMTB (C) or toward 4‐CMTB stimulation after priming with 1 mM C3 (D). IL‐8 was used for comparison as a standard neutrophil chemoattractant at a concentration of 1 nM (open circles). Depicted are mean values + sem of 3 independent experiments
3.7. CD11b expression is differentially regulated by C3 and 4‐CMTB stimulation
Neutrophil granule mobilization and adhesion is a hallmark in the process of neutrophil activation to defend the host against pathogens. Cell surface expression of CD11b (integrin α‐M, ITGAM) serves herein as a measure of leukocyte adhesion as well as mobilization of secretory vesicles and transport of cargo carrying granules. 19 We found this marker to be differentially regulated by FFA2R activation. While C3 only marginally increased CD11b cell surface expression (Fig. 7A), 4‐CMTB strongly up‐regulated CD11b surface presence on neutrophils (Fig. 7B). Despite lacking evidence for a significant role of FFA2 on neutrophil chemotaxis these data indicate a receptor site‐specific FFA2 functionality in the leukocyte activation process. Priming neutrophils with 4‐CMTB before stimulating with C3 showed that neutrophils were sensitized by 4‐CMTB to further enhance C3‐mediated CD11b surface abundance globally over the concentration range (Fig. 7A), while priming with C3 rather shifted the concentration response curve of 4‐CMTB to the left but could not further increase maximum responses (Fig. 7B).
FIGURE 7.
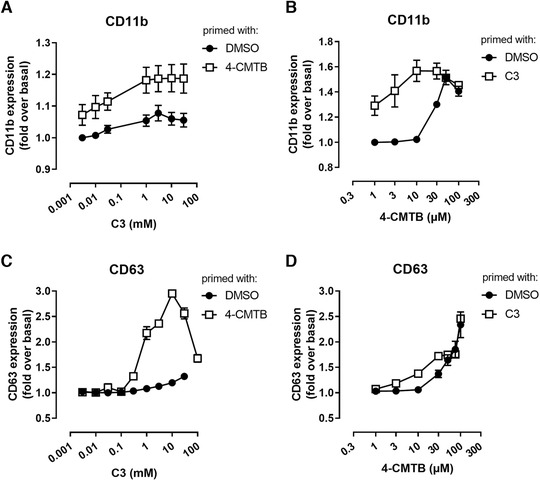
Analysis of neutrophil activation marker upon orthosteric and allosteric FFA2R activation. CD11b expression on human neutrophils stimulated with increasing concentrations of C3 (A) or 4‐CMTB (B) after priming with 30 µM 4‐CMTB or 1 mM C3, respectively. Measurement of CD63 expression on human neutrophils after stimulation with increasing concentrations of C3 (C) or 4‐CMTB (D) after neutrophil priming with 30 µM 4‐CMTB or 1 mM C3, respectively. Depicted are mean values as fold over basal ± sem, n = 5. Where not shown, error bars are within symbol size
3.8. Allosteric FFA2 receptor targeting rescues orthosteric receptor signaling to induce neutrophil activation under desensitized conditions
We further extended our investigations on the expression of CD63 (cluster of differentiation protein 63), which serves as a marker for azurophilic (primary) degranulation and neutrophil activation mediating their degranulation at sites of inflammation. 19 Orthosteric activation by C3 could merely induce CD63 expression at concentrations that fully activate FFA2 receptor signaling (Fig. 7C). 4‐CMTB, however, led to robust CD63 level in a dose‐dependent manner (Fig. 7D). Intriguingly, priming with 4‐CMTB turned C3 into a highly effective neutrophil activator with strong enhancement of maximal activation level (Fig. 7C). Priming with C3 and subsequent stimulation with 4‐CMTB, however, left maximal CD63 level largely unaffected compared to stimulation with 4‐CMTB alone and only slightly shifted the 4‐CMTB concentration response curve to the left (Fig. 7D). This emphasizes—in line with results from calcium measurements—the conclusion that orthosteric FFA2R signaling through C3, when silent on its own, can be re‐enabled by pre‐stimulation with the allosteric agonist 4‐CMTB.
As a complementary view on leukocyte activation, we assessed the capability of neutrophils to produce reactive oxygen species (ROS) after FFA2 receptor activation. ROS production by immune cells is an effective tool to destroy infectious agents and constitute an important means in first line defense of neutrophils. 1 , 2 We found that C3 failed to increase ROS production while 4‐CMTB showed only marginally increased ROS levels in human neutrophils (Fig. 8A and B). Neutrophils primed with C3 and stimulated with 4‐CMTB subsequently produced slightly elevated ROS levels, whereas priming with the allosteric activator promoted massive ROS production upon subsequent C3 stimulation (Fig. 8C and D).
FIGURE 8.
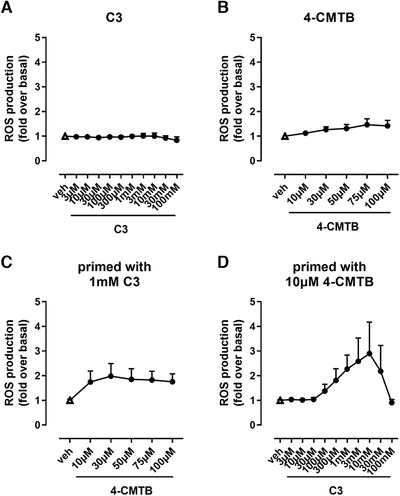
Reactive oxygen species production by human neutrophils upon orthosteric and allosteric FFA2R activation. ROS production in human neutrophils after stimulation with C3 (A), 4‐CMTB (B), or after priming with 1 mM C3 and subsequent stimulation with increasing concentrations of 4‐CMTB (C) or after priming with 10 µM 4‐CMTB and subsequent stimulation with increasing concentrations of C3 (D). Depicted are mean values + sem, n = 3. Where not shown, error bars are within symbol size
As paralleled in the preceding complementary analyses, these phenotypic neutrophil readouts underline that orthosteric and allosteric FFA2R signaling differentially translate into neutrophil response. Moreover, orthosteric FFA2R signaling can be restored by pre‐stimulation of the receptor with the allosteric agonist 4‐CMTB, thus substantiating the hypothesis that allosteric FFA2 receptor engagement may play a decisive role in FFA2R functionality in neutrophils.
4. DISCUSSION
Allosteric modulation is a promising approach in drug discovery and development. Numerous advantages are postulated to be associated with allostery, for example, (1) the spatio‐temporal preservation of physiological tone if allosteric ligands function as positive or negative allosteric modulators without intrinsic activity, (2) the chance to access targets previously marked as "undruggable" or to design drugs with higher target specificity as allosteric binding sites are commonly less conserved among structurally related proteins, (3) a wider safety window due to a "ceiling effect" of allosteric modulation, or (4) the possibility to generate new pharmacology, such as change in the signaling downstream of a target depending on the activation mode, a phenomenon referred to as signaling bias. 20 , 21 It was previously shown that the allosteric agonist 4‐CMTB sequentially activates the FFA2 receptor to induce a different activation pattern compared to the endogenous orthosteric agonist C3 in the recombinant HEK293 cell system. 13 Whether allosteric receptor activation in human neutrophils endogenously expressing FFA2R entails distinct physiological consequences compared to orthosteric FFA2R activation remained yet unexplored. We here provide evidence for a distinct role of allosteric versus orthosteric FFA2 receptor activation in primary human neutrophils. 4‐CMTB and C3 induced a different signaling fingerprint in neutrophils and led to distinct whole cell responses, such as cytoskeletal rearrangement, measures of microbicidal activity, and adhesion properties.
In agreement with previous reports, we detected low chemotactic activity toward FFA2 stimulation as compared to migration toward chemokine receptor activation (here toward IL‐8 or described for the chemotactic peptide (fMLP), respectively). 18 A negligible role of the FFA2 receptor for neutrophil response under inflammatory conditions, in which cytokine level can increase tremendously, is further fueled by the finding that either orthosteric FFA2 agonism or fMLP effectively silenced subsequent orthosteric FFA2‐induced neutrophil activation. This indicates that the FFA2 receptor underwent strong homologous and heterologous receptor desensitization. 18 We here corroborate homologous and heterologous FFA2 receptor desensitization and found allosteric targeting to re‐sensitize the FFA2 receptor toward orthosteric stimulation. Apparent GPCR desensitization could be linked to various mechanisms. These include (1) receptor phosphorylation, (2) β‐arrestin recruitment to the receptor, (3) receptor internalization, (4) receptor trafficking and potentially signaling from intracellular locations, (5) regulation of receptor movement by interweaving with the actin cytoskeletal, or (6) downstream signaling bias to feedback on upstream receptor sensitivity. A comprehensive analysis of all potential mechanisms is beyond the scope of this work and hence the exact mechanisms of FFA2 receptor de/resentization in neutrophils still remain to be elucidated. Nevertheless, the hitherto reported data on FFA2R regulation of neutrophils narrow down possible underlying mechanisms. β‐Arrestin binding was reported to both mediate FFA2 receptor internalization as well as to shape anti‐inflammatory FFA2 signaling in neutrophils. 22 However, C3 and 4 CMTB have been shown to induce comparable level of β‐arrestin recruitment contradicting a differential recruitment of β‐arrestin to orthosteric versus allosteric activated FFA2 receptors as the underlying decisive mechanism. 23 Likewise, regulation of receptor mobility involving the actin cytoskeleton can mediate both GPCR desensitization and reactivation, but disrupting the actin cytoskeletal apparatus was shown to be insufficient to regulate FFA2 receptor responsiveness on neutrophils. 18 Finally, the here presented data disprove the hypothesis that receptor neutrophil membrane abundance plays a key role in the observed phenomenon of FFA2 receptor resensitization since (1) orthosteric (C3) and allosteric (4‐CMTB) FFA2R agonists did not differ in their capability to internalize FFA2 receptors from the membrane, while they evidently induced different level of receptor desensitization on neutrophils, (2) IL‐8 did not affect FFA2 receptor surface level, although leading to substantial FFA2R desensitization, and (3) allosteric resensitization of the FFA2 receptor by 4‐CMTB did not correlate with restoration of initial membrane receptor level. Hence, other mechanisms are likely to drive the observed allosteric FFA2R receptor resensitization. It is tempting to speculate that the different signaling pathway composition, that is, signaling bias, downstream of allosteric versus orthosteric FFA2 receptor activation eventually lead to differential regulation of FFA2R responsiveness. However, hypotheses involving signaling bias, FFA2R phosphorylation, and/or receptor trafficking as determining processes in FFA2 receptor de/resensitization require future experimental evaluation.
Several studies suggest a relevance of the FFA2 receptor in preclinical in vitro and in vivo disease models ranging from asthma and colitis to diabetes and cancer, all of which are driven by inflammatory processes to a different degree. 17 , 24 , 25 , 26 , 27 , 28 , 29 , 30 , 31 , 32 , 33 Free fatty acid receptor signaling is furthermore likely also underlying the observation that a high‐fiber diet is proven to be associated with an amelioration or prevention of immunological or neoplastic disorders but the exact mechanism of FFA receptors in these processes is only partially understood. 24 , 25 , 34 , 35 An increase of SCFAs by the metabolizing microbiota and subsequent activation of anti‐inflammatory and immune‐regulatory pathways involving SCFA receptors such as FFA2R is thought to be responsible for these effects. 24 , 25 However, whether activation or inactivation of the FFA2 receptor is eventually the goal of a pharmacological therapy is still a matter of debate. The discussion might be complicated by the fact that SCFA activity is not only mediated by GPCRs, but also non‐receptor mediated signaling pathways (HDAC inhibition, metabolic effects), via different cell types (hematopoietic and non‐hematopoietic cells) entailing different physiological functions and displaying species‐specific differences. 25 , 29 , 36 Some studies support an anti‐inflammatory role of FFA2 receptor activation by controlling the release of inflammatory mediators and regulation of immune cell infiltration in atherosclerosis, colitis, asthma, and arthritis models. 17 , 28 , 37 Others demonstrated a more complex and bifunctional role of the receptor with FFA2R−/− mice showing decreased leukocyte response in an acute colitis model and reduced tissue damage in a chronic colitis model. 26 Furthermore, M1 and M2 Mϕs in adipose tissue were differentially regulated by FFA2R to ensure tissue homeostasis confirming the notion that FFA2R conveys both pro‐inflammatory and anti‐inflammatory actions. 38 Our finding that allosteric targeting of the FFA2 receptor allows neutrophils to remain responsive under desensitizing conditions adds, however, another twist to the interpretation of FFA2R relevance in a disease setting.
A balanced response of neutrophils becomes particularly important in the state of exuberant inflammation during sepsis, which is characterized by a dysfunction of the immune response resulting in both high patient morbidity as well as mortality. 39 Neutrophils are key players in the first line of host defense and failure of neutrophil function is a hallmark in the pathophysiology of sepsis. 3 Intensified neutrophil response to an infection was correlated to higher survival rates of septic patients. 40 Furthermore, FFA2R signaling has been demonstrated to be beneficial in an acute setting mimicked by a mouse model of bacterial lung infection with K. pneumoniae while FFA2R−/− mice had a defect in the clearance of a C. rodentium infection. 27 , 41 These findings underline the importance of FFA2R‐mediated acute neutrophil response as the initial immune defense mechanism. In line with this, a recent study found that a higher FFA2 receptor expression level was associated with an increase in the 30‐day survival rate of sepsis patients corroborating an integral role of FFA2 during and after sepsis. 42 Because neutrophil responsiveness is tightly regulated by receptor desensitization mechanisms, it is tempting to speculate that an increase in FFA2 receptor sensitivity, for example, by allosteric priming, toward the endogenous ligand might be of benefit under septic conditions during massive local cytokine release to increase neutrophil response. A previous study showed that allosteric priming of the FFA2 receptor by another allosteric ligand (Cmp 58) augmented both potency and efficacy of the orthosteric agonist acetate in human neutrophil NADPH‐oxidase response. 43 Lind et al. observed this phenomenon also between FFA2 and another receptor, P2Y2, and suggest an allosteric mechanism underlying the inter‐receptor cross‐talk. 44 Although these studies use structurally slightly different ligands than in the here presented data, the general finding of allosteric FFA2 receptor priming corroborates our observations of an allosteric mechanism impacting the signaling repertoire of the endogenous receptor activator. Beyond that, our finding of a distinct allosteric FFA2 receptor activation to resensitize neutrophils toward the endogenous receptor activator could provide an intricate way to safeguard FFA2R functionality and to preserve neutrophil responsiveness toward SCFA.
While desensitization of neutrophil receptors is crucial in the coordination of immune response, our knowledge about this complex regulation is only rudimentary. It is therefore fundamental to develop a growing understanding of the underlying molecular mechanisms for future treatments of inflammatory diseases. A mechanism to reorganize the hierarchy of neutrophil stimuli by resensitization of one receptor with an allosteric agonist toward the orthosteric ligand bodes a new pharmacological approach to enhance neutrophil functionality under desensitizing conditions. Future studies will have to explore if the mechanism found here ex vivo is of relevance in an in vivo setting.
AUTHORSHIP
R.F., J.N., U.H., S.H., and J.S. performed experiments, analyzed data, and reviewed the manuscript. F.E. reviewed and revised the manuscript. M.G. conceived the study, performed experiments, analyzed data, and wrote the manuscript.
DISCLOSURE
J.N., U.H., M.G. and F.E. are employees of Bayer AG. The other authors declare no conflict of interests.
ACKNOWLEDGMENTS
The authors thank Evi Kostenis for providing infrastructure and support to this project and Kathrin Rohrer for technical support.
Frei R, Nordlohne J, Hüser U, et al. Allosteric targeting of the FFA2 receptor (GPR43) restores responsiveness of desensitized human neutrophils. J Leukoc Biol. 2021;109:741–751. 10.1002/JLB.2A0720-432R
REFERENCES
- 1. Bardoel BW, Kenny EF, Sollberger G, Zychlinsky A. The balancing act of neutrophils. Cell Host Microbe. 2014;15:526‐536. [DOI] [PubMed] [Google Scholar]
- 2. Mócsai A. Diverse novel functions of neutrophils in immunity, inflammation, and beyond. J Exp Med. 2013;210:1283‐1299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Shen XF, Cao K, Jiang JP, Guan WX, Du JF. Neutrophil dysregulation during sepsis: an overview and update. J Cell Mol Med. 2017;21:1687‐1697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Futosi K, Fodor S, Mócsai A. Reprint of neutrophil cell surface receptors and their intracellular signal transduction pathways. Int Immunopharmacol. 2013;17:1185‐1197. [DOI] [PubMed] [Google Scholar]
- 5. Le Poul E, Loison C, Struyf S, et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cells activation. J Biol Chem. 2003. [DOI] [PubMed] [Google Scholar]
- 6. Brown AJ, Goldsworthy SM, Barnes AA, et al. The orphan G protein‐coupled receptors GPR41 and GPR43 are activated by propionate and other short‐chain carboxylic acids. J Biol Chem. 2002. [DOI] [PubMed] [Google Scholar]
- 7. Cummings J, Pomare E, Branch W, Naylor C, Macfarlane G. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut. 1987;28:1221‐1227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Lee T, Schwandner R, Swaminath G, et al. Identification and functional characterization of allosteric agonists for the G protein‐coupled receptor FFA2. Mol Pharmacol. 2008. [DOI] [PubMed] [Google Scholar]
- 9. Brantis C, Ooms F, Bernard J. Novel amino acid derivatives and their use as GPR43 receptor modulators. PCT Int Appl. 2011. WO_2011_092284_A1. [Google Scholar]
- 10. Hudson BD, Tikhonova IG, Pandey SK, Ulven T, Milligan G. Extracellular ionic locks determine variation in constitutive activity and ligand potency between species orthologs of the free fatty acid receptors FFA2 and FFA3. J Biol Chem. 2012:396259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Schratl P, Sturm EM, Royer JF, et al. Hierarchy of eosinophil chemoattractants: role of p38 mitogen‐activated protein kinase. Eur J Immunol. 2006;36:2401‐2409. [DOI] [PubMed] [Google Scholar]
- 12. Grundmann M. Label‐free dynamic mass redistribution and bio‐impedance methods for drug discovery. Curr Protoc Pharmacol. 2017;77:9.24. 1‐9.24. 21. [DOI] [PubMed] [Google Scholar]
- 13. Grundmann M, Tikhonova IG, Hudson BD, et al. A molecular mechanism for sequential activation of a G protein‐coupled receptor. Cell Chem Biol. 2016;23:392‐403. [DOI] [PubMed] [Google Scholar]
- 14. Ali H, Richardson RM, Haribabu B, Snyderman R. Chemoattractant receptor cross‐desensitization. J Biol Chem. 1999;274:6027‐6030. [DOI] [PubMed] [Google Scholar]
- 15. Schrage R, Schmitz A‐L, Gaffal E, et al. The experimental power of FR900359 to study Gq‐regulated biological processes. Nat Commun. 2015;6:10156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Schröder R, Janssen N, Schmidt J, et al. Deconvolution of complex G protein‐coupled receptor signaling in live cells using dynamic mass redistribution measurements. Nat Biotechnol. 2010;28:943‐949. [DOI] [PubMed] [Google Scholar]
- 17. Maslowski KM, Vieira AT, Ng A, et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature. 2009;461:1282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Björkman L, Mårtensson J, Winther M, et al. The neutrophil response induced by an agonist for the free fatty acid receptor 2 (FFA2R/GPR43) is primed by TNF‐α and by receptor uncoupling from the cytoskeleton but attenuated by tissue recruitment. Mol Cell Biol. 2016;36:2583‐2595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Kuijpers T, Tool A, Van der Schoot C, et al. Membrane surface antigen expression on neutrophils: a reappraisal of the use of surface markers for neutrophil activation. Blood. 1991;78:1105‐1111. [PubMed] [Google Scholar]
- 20. Conn PJ, Christopoulos A, Lindsley CW. Allosteric modulators of GPCRs: a novel approach for the treatment of CNS disorders. Nat Rev Drug Discovery. 2009;8:41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Wenthur CJ, Gentry PR, Mathews TP, Lindsley CW. Drugs for allosteric sites on receptors. Annu Rev Pharmacol Toxicol. 2014;54:165‐184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Lee SU, In HJ, Kwon MS, et al. β‐Arrestin 2 mediates G protein‐coupled receptor 43 signals to nuclear factor‐κB. Biol Pharm Bull. 2013:b13‐00312. [DOI] [PubMed] [Google Scholar]
- 23. Hudson BD, Due‐Hansen ME, Christiansen E, et al. Defining the molecular basis for the first potent and selective orthosteric agonists of the FFA2 free fatty acid receptor. J Biol Chem. 2013;288:17296‐17312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Sivaprakasam S, Gurav A, Paschall A, et al. An essential role of Ffar2 (Gpr43) in dietary fibre‐mediated promotion of healthy composition of gut microbiota and suppression of intestinal carcinogenesis. Oncogenesis. 2016;5:e238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Macia L, Tan J, Vieira AT, et al. Metabolite‐sensing receptors GPR43 and GPR109A facilitate dietary fibre‐induced gut homeostasis through regulation of the inflammasome. Nat Commun. 2015;6:7734. [DOI] [PubMed] [Google Scholar]
- 26. Sina C, Gavrilova O, Förster M, et al. G protein‐coupled receptor 43 is essential for neutrophil recruitment during intestinal inflammation. J Immunol. 2009;183:7514‐7522. [DOI] [PubMed] [Google Scholar]
- 27. Kim MH, Kang SG, Park JH, Yanagisawa M, Kim CH. Short‐chain fatty acids activate GPR41 and GPR43 on intestinal epithelial cells to promote inflammatory responses in mice. Gastroenterology. 2013;145:396‐406.e10. [DOI] [PubMed] [Google Scholar]
- 28. Kamp ME, Shim R, Nicholls AJ, et al. G protein‐coupled receptor 43 modulates neutrophil recruitment during acute inflammation. PLoS One. 2016;11:e0163750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Ang Z, Er JZ, Tan NS, et al. Human and mouse monocytes display distinct signalling and cytokine profiles upon stimulation with FFAR2/FFAR3 short‐chain fatty acid receptor agonists. Sci Rep. 2016;6:34145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Tang C, Ahmed K, Gille A, et al. Loss of FFA2 and FFA3 increases insulin secretion and improves glucose tolerance in type 2 diabetes. Nat Med. 2015;21:173. [DOI] [PubMed] [Google Scholar]
- 31. Kimura I, Ozawa K, Inoue D, et al. The gut microbiota suppresses insulin‐mediated fat accumulation via the short‐chain fatty acid receptor GPR43. Nat Commun. 2013;4:1829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Vieira AT, Macia L, Galvão I, et al. A role for gut microbiota and the metabolite‐sensing receptor GPR43 in a murine model of gout. Arthritis Rheumatol. 2015;67:1646‐1656. [DOI] [PubMed] [Google Scholar]
- 33. Tolhurst G, Heffron H, Lam YS, et al. Short‐chain fatty acids stimulate glucagon‐like peptide‐1 secretion via the G‐protein–coupled receptor FFAR2. Diabetes. 2011:DB_111019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Park Y, Subar AF, Hollenbeck A, Schatzkin A. Dietary fiber intake and mortality in the NIH‐AARP diet and health study. Arch Intern Med. 2011;171:1061‐1068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Anderson JW, Baird P, Davis RH, et al. Health benefits of dietary fiber. Nutr Rev. 2009;67:188‐205. [DOI] [PubMed] [Google Scholar]
- 36. Li M, van Esch BC, Wagenaar GT, Garssen J, Folkerts G, Henricks PA. Pro‐and anti‐inflammatory effects of short chain fatty acids on immune and endothelial cells. Eur J Pharmacol. 2018;831:52‐59. [DOI] [PubMed] [Google Scholar]
- 37. Li M, Van Esch BEC, Henricks PA, Garssen J, Folkerts G. The anti‐inflammatory effects of short chain fatty acids on lipopolysaccharide‐or tumor necrosis factor α‐stimulated endothelial cells via activation of GPR41/43 and inhibition of HDACs. Front Pharmacol. 2018;9:533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Nakajima A, Nakatani A, Hasegawa S, et al. The short chain fatty acid receptor GPR43 regulates inflammatory signals in adipose tissue M2‐type macrophages. PLoS One. 2017;12:e0179696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Angus DC, Van der Poll T. Severe sepsis and septic shock. N Engl J Med. 2013;369:840‐851. [DOI] [PubMed] [Google Scholar]
- 40. Tavares‐Murta BM, Zaparoli M, Ferreira RB, et al. Failure of neutrophil chemotactic function in septic patients. Crit Care Med. 2002;30:1056‐1061. [DOI] [PubMed] [Google Scholar]
- 41. Galvão I, Tavares LP, Corrêa RO, et al. The metabolic sensor gPr43 receptor plays a role in the control of Klebsiella pneumoniae infection in the lung. Front Immunol. 2018;9:142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Carr ZJ, Louw A, Fehr G, Li JD, Kunselman A, Ruiz‐Velasco V. Increased whole blood FFA2/GPR43 receptor expression is associated with increased 30‐day survival in patients with sepsis. BMC Res Notes. 2018;11:41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Mårtensson J, Holdfeldt A, Sundqvist M, et al. Neutrophil priming that turns natural FFA2R agonists into potent activators of the superoxide generating NADPH‐oxidase. J Leukoc Biol. 2018;104:1117‐1132. [DOI] [PubMed] [Google Scholar]
- 44. Lind S, Holdfeldt A, Mårtensson J, et al. Functional selective ATP receptor signaling controlled by the free fatty acid receptor 2 through a novel allosteric modulation mechanism. FASEB J. 2019;33:6887‐6903. [DOI] [PubMed] [Google Scholar]