

Abstract
Perceptual learning has been widely used to study the plasticity of the visual system in adults. Owing to the belief that practice makes perfect, perceptual learning protocols usually require subjects to practice a task thousands of times over days, even weeks. However, we know very little about the relationship between training amount and behavioral improvement. Here, four groups of subjects underwent motion direction discrimination training over 8 days with 40, 120, 360, or 1080 trials per day. Surprisingly, different daily training amounts induced similar improvement across the four groups, and the similarity lasted for at least 2 weeks. Moreover, the group with 40 training trials per day showed more learning transfer from the trained direction to the untrained directions than the group with 1080 training trials per day immediately after training and 2 weeks later. These findings suggest that perceptual learning of motion direction discrimination is not always dependent on the daily training amount and less training leads to more transfer.
Keywords: perceptual learning, training amount, motion direction discrimination, specificity, plasticity
Introduction
The human brain can achieve long-term improvement in perceptual sensitivity as a result of learning (Fahle & Poggio, 2002). A prevailing view on this improvement is that “practice makes perfect,” implying that tens of thousands of trials of training over days or weeks are necessary to induce substantial improvement in performance (Aberg et al., 2009; Banai & Lavner, 2014; Censor, Sagi, & Cohen, 2012; Chung, Levi, & Li, 2006; Husk, Bennett, & Sekuler, 2007; Sigman & Gilbert, 2000). However, several recent studies showed that a small amount of training was sufficient to induce perceptual learning (Amar-Halpert et al., 2017; Hussain, Sekuler, & Bennett, 2009; Molloy et al., 2012). Hussain et al. (2009) examined the amount of practice needed to improve performance on texture and face identification. In a texture identification task, they found that 105 trials of practice on the first day were required to enhance performance relative to the control group at the start of testing on the second day. In a face identification task, even only 21 trials of practice could enhance performance relative to the control group (Hussain et al., 2009). In a recent study on texture discrimination learning (Amar-Halpert et al., 2017), participants underwent a practice for 252 trials on the first day, and then they returned for 3 daily sessions with only five near-threshold trials per session. Discrimination thresholds were measured on the first day and the fifth day. Intriguingly, such short training resulted in a remarkable learning effect. Based on this finding, Amar-Halpert and colleagues proposed that learning was due to a memory reactivation mechanism.
It has been shown that training beyond a certain amount could not further benefit learning (Karni & Sagi, 1993; Savion-Lemieux, T., & Penhune, V. B., 2005). In a temporal–interval discrimination task, Wright and Sabin (2007) trained subjects for either 360 or 900 trials per day for 6 days. Significant learning occurred with both 360 and 900 training trials per day, and 900 training trials per day did not induce greater improvement relative to 360 training trials. Likewise, similar effects were also observed with a mirror-reading letter task (Ofen-Noy, Dudai, & Karni, 2003), a visual texture discrimination task (Karni & Sagi, 1993), and an auditory identification task (Roth, 2005). Notably, overtraining could even be detrimental to the learning effect already acquired (Ashley & Pearson, 2012; Censor, Karni, & Sagi, 2006; Mednick et al., 2002; Mednick, Arman, & Boynton, 2005; Ofen, Moran, & Sagi, 2007). Mednick et al. (2005) measured the performance on a texture discrimination task in three 1-hour sessions and found that the performance deteriorated steadily both within and across the first two sessions. Because repeated within-day testing led to a retinotopically specific decrease in performance, such perceptual deterioration is not simply due to general fatigue or boredom. Therefore, intensive training might lead to limited behavior improvement.
In this study, we aimed to investigate the relationship between daily training amount and behavioral improvement—how does the daily training amount modulate the magnitude and specificity of the perceptual learning effect with a motion direction discrimination task? We were also interested in how long the modulation effects could persist. Participants were trained for 40, 120, 360, or 1080 trials per day with a visual motion direction discrimination task. Threshold measurements were conducted before, one day after, and two weeks after eight training days at the trained direction and the untrained directions (30°, 60°, and 90° away from the trained direction).
Methods
Subjects
Fifty-nine subjects (21 males) participated in the study. Their ages ranged from 18 to 28. All subjects were naïve to the purpose of the study and had never participated in any perceptual learning experiment before. They were right-handed with reported normal or corrected-to-normal vision and had no known neurological or visual disorders. They gave written, informed consent in accordance with the procedures and protocols approved by the human subject review committee of Peking University. This study adhered to the Declaration of Helsinki.
Stimuli and apparatus
Similar to our previous study (Chen et al., 2015), visual stimuli were random-dot kinematograms (RDKs) with 100% coherence (Figure 1A). All dots in a RDK moved in the same direction (luminance: 3.76 cd/ m2; diameter: 0.1°; speed: 10°/sec). At any one moment, 400 dots were visible within an 8° circular aperture. The dots were presented against a gray background (luminance: 19.8 cd/m2). The visual stimuli were presented on an IIYAMA HM204DT 22-in monitor, with a spatial resolution of 1024 × 768 and a refresh rate of 60 Hz. Subjects viewed the stimuli from a distance of 60 cm. Their head was stabilized using a head and chin rest.
Figure 1.

Stimuli and experimental protocol. (A) Schematic description of a two-alternative forced-choice (2-AFC) trial in a QUEST staircase for measuring motion direction discrimination thresholds. (B) Experimental protocol. Four groups of subjects underwent motion direction discrimination training at a fixed motion direction over eight days with 1, 3, 9, or 27 QUEST staircases per day, respectively. They were tested at 0°, 30°, 60°, and 90° away from the trained direction on the days before (Pre), immediately after (Post1), and two weeks (Post2) after training.
Designs
Fifty-nine subjects were randomly assigned into four training groups (n = 12, 11, 12, and 12), respectively and a control group (n = 12). Four training groups underwent four phases (Figure 1B): pretraining test (Pre), motion direction discrimination training, post-training test 1 (Post1), and post-training test 2 (Post2). The control group only underwent Pre, Post1, and Post2. Pre and Post1 took place on the days immediately before and after training, and Post2 took place 2 weeks after training.
During the training phase, each subject underwent eight daily training sessions to perform a motion direction discrimination task at a direction of θ, which was chosen randomly from eight directions: 22.5°, 67.5°, 112.5°, 157.5°, 202.5°, 247.5°, 292.5°, and 337.5° (0° was the rightward direction) at the beginning and was fixed for all the sessions. For the four training groups, a daily training session consisted of 1, 3, 9, and 27 QUEST (Watson & Pelli, 1983) staircases of 40 trials, corresponding with 40, 120, 360, or 1080 trials, respectively. In a trial, two RDKs with motion directions of θ + Δθ/2 and θ – Δθ/2 were presented successively for 200 ms each and were separated by a 600-ms blank interval. The temporal order of these two RDKs was randomized. Subjects were asked to make a two-alternative forced-choice judgment of the direction of the second RDK relative to the first one (clockwise or counterclockwise). Informative feedback was provided after each response by brightening (correct response) or dimming (wrong response) the fixation point, which could facilitate learning (Goldhacker, Rosengarth, Plank, & Greenlee, 2014). The next trial began 1 second after feedback. The Δθ varied trial by trial and was controlled by the QUEST staircase to estimate subjects' discrimination thresholds at 75% correct. Except for the first staircase, each staircase started with the threshold derived from the preceding staircase.
During the three test phases, psychophysical tests were performed at four motion directions, which were 0°, 30°, 60°, and 90° deviated from the trained direction all either clockwise or counterclockwise (hereafter referred to as 0°, 30°, 60°, and 90°). The four directions were counterbalanced for individual subjects. Ten QUEST staircases (same as above) were completed for each direction. Discrimination thresholds from the 10 staircases for each direction were averaged as a measure of subjects’ discrimination performance. Subjects’ performance improvement at a direction was calculated as (pretraining threshold – post-training threshold)/pretraining threshold × 100%. Before the test phases, subjects practiced 10 trials per direction to get familiar with the stimuli and the experimental procedure. In our data analyses, Bonferroni correction was applied with t-tests and analysis of variance (ANOVA) involving multiple comparisons. We verified data distribution assumptions of normality and heteroscedasticity by Shapiro–Wilk and Levene's tests before performing ANOVAs.
Results
Learning effect was independent of daily training amount
To explore whether there was any difference between groups before learning, we applied ANOVAs to the thresholds measured before learning at the four directions (0°, 30°, 60°, and 90°) with training amount (1, 3, 9, and 27 QUEST staircases) as a between-subject factor. No difference was found at these directions, 0°: F (3, 43) = 0.207, p = 0.891; 30°: F (3, 43) = 1.045, p = 0.382; 60°: F (3, 43) = 0.860, p = 0.469; 90°: F (3, 43) = 2.088, p = 0.116. Therefore, there was no difference between groups that preceded the study. Throughout the training, motion direction discrimination thresholds of the four groups of subjects gradually decreased (Figure 2A). We compared the discrimination thresholds at the trained direction before and after learning across groups using a mixed-design ANOVA with test (Pre, Post1, and Post2) as a within-subject factor and training amount (1, 3, 9, and 27 QUEST staircases) as a between-subject factor. The main effect of test was significant, F (2, 42) = 100.359, p < 0.0001. The main effect of training amount was not significant, F (3, 43) = 0.212, p = 0.887. The interaction between test and training amount was not significant either, F (6, 86) = 0.539, p = 0.777. The thresholds measured at Pre were significantly higher than those at Post1, 1 QUEST staircase, t (11) = 8.835, p < 0.0001; 3 QUEST staircases, t (10) = 7.052, p < 0.001; 9 QUEST staircases, t (11) = 5.493, p < 0.001; 27 QUEST staircases, t (11) = 8.788, p < 0.0001, and at Post2, 1 QUEST staircase, t (11) = 6.714, p < 0.0001; 3 QUEST staircases, t (10) = 5.420, p < 0.001; 9 QUEST staircases, t (11) = 4.759, p < 0.01; 27 QUEST staircases, t (11) = 8.519, p < 0.0001 (Figure 2B). These finding suggest that our training procedure led to a significant learning effect and the learning effect might be independent of the training amount.
Figure 2.

Perceptual learning effects at the trained direction. (A) Learning curves with different training amounts. (Left) Normalized motion direction discrimination thresholds are plotted as a function of the training day and then fitted with a power function. Motion direction discrimination thresholds were normalized by dividing them by the thresholds measured on the first training day. (Right) Motion direction discrimination thresholds measured by the first QUEST staircase on each day are plotted as a function of the training day and then fitted with a power function. (B) Motion direction discrimination thresholds at the trained direction at Pre, Post1, and Post2 for the four training groups. Asterisks indicate significant difference between Pre and Post1, Post2 (***p < 0.001). (C) Percent improvements in motion direction discrimination performance at Post1 and Post2, relative to Pre. Error bars denote 1 SEM calculated across subjects.
We calculated the percent improvements in discrimination performance after training. The improvements at the trained direction were more than 40% at Post1 and more than 38% at Post2 in all the four groups. A mixed-design ANOVA was applied to the percent improvements with test (Post1 and Post2) as a within-subject factor, and training amount (1, 3, 9, and 27 QUEST staircases) as a between-subject factor. The main effects of training amount, F (3, 43) = 1.210, p = 0.318, and test, F (1, 43) = 1.880, p = 0.177 were not significant. The interaction between test and training amount was not significant either, F (3, 43) = 0.175, p = 0.913 (Figure 2C). To examine whether there was more learning in the 27 QUEST group than that in the 1 QUEST group, we performed a t-test on the percent improvements at the trained direction in Figure 2C. No significant difference was found between the two groups, at both Post1, t (22) = 1.344, p = 0.193, and Post2 t (22) = 0.851, p = 0.404. These results confirmed that the improvement at the training direction did not depend on the daily training amount.
Learning specificity was dependent on daily training amount
We further investigated the relationship between learning specificity and daily training amount. Figure 3 shows the percent improvements in motion direction discrimination performance at the untrained directions. A mixed-design ANOVA was applied to the percent improvements with test (Post1 and Post2) and motion direction (30°, 60°, and 90°) as within-subject factors and training amount (1, 3, 9, and 27 QUEST staircases) as a between-subject factor. The main effect of direction was not significant, F (2, 42) = 0.092, p = 0.912, and we did not find any significant interaction, all p > 0.247, suggesting little difference between the three untrained directions. The main effect of training amount was not significant. F (3, 43) = 1.500, p = 0.228. The percent improvements at the three untrained directions were significantly greater than zero at Post1 and Post2, all t (46) > 4.308, p < 0.001, but significantly lower than those at the trained direction at Post1 and Post2, all t (46) > 5.367, p < 0.001. Therefore, we quantified the learning specificity as the difference between the percent improvement at the trained direction and the average percent improvement at the three untrained directions.
Figure 3.

(A) Motion direction discrimination thresholds at the trained (0°) and untrained (30°, 60°, and 90°) directions at Pre (left), Post1 (middle), and Post2 (right) for the four training groups. (B) Percent improvements in motion direction discrimination performance at the trained and untrained directions for the four training groups at Post1 (left) and Post2 (right), relative to Pre. Error bars denote 1 SEM calculated across subjects.
Figure 4 shows the learning specificities in the four training groups. A mixed-design ANOVA was applied to the learning specificity with test (Post1 and Post2) as a within-subject factor and training amount (1, 3, 9, and 27 QUEST staircases) as a between-subject factor. The main effect of training amount was marginally significant, F (3, 43) = 2.383, p < 0.1. The main effect of test was not significant, F (1, 43) = 1.035, p = 0.315, and the interaction between test and training amount was not significant either, F (3, 43) = 0.242, p = 0.867. Then, we made comparisons between training amount conditions at Post1 and Post2. Planned t-tests showed that the group receiving 27 QUEST staircases training per day exhibited stronger specificity than the group receiving 1 QUEST staircase training per day at Post1, t (22) = –2.779, p < 0.01, and Post2, t (22) = –1.929, p < 0.05. Our results demonstrated that less training led to less specificity or more transfer, and the characteristic lasted for at least two weeks.
Figure 4.
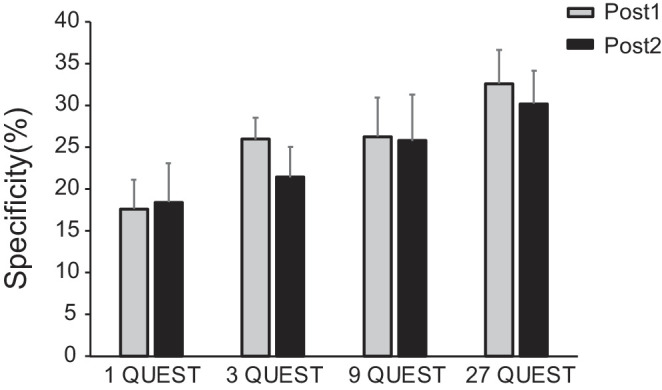
Learning specificity in the four training groups. Error bars denote 1 SEM calculated across subjects.
Test–retest effect
It remains unclear to what extent the performance improvements in the trained and untrained directions are due to a test-retest effect occurring at Pre, Post1, and Post2. To quantify the test–retest effect, we collected data from a control group, which only underwent Pre, Post1, and Post2. Relative to Pre, the percent improvements averaged across the four directions were 3.095%, one-sample t-test (47) = 0.656, p = 0.515, at Post1 and 18.116%, t (47) = 4.401, p < 0.001, at Post2. Notably, the improvements at the untrained direction at Post2 were largely due to the test-retest effect.
Discussion
In this study, we examined the relationship between daily training amount and two visual learning outcomes: the improvement at the trained feature, and the transfer effect to the untrained features. We found that (1) a small daily training amount of 40 trials was sufficient to induce a significant behavioral improvement; no further improvement was observed in groups with larger daily training amounts and (2) the group with the smallest daily training amount exhibited the largest transfer effect. These effects persisted up to 2 weeks after training. These findings shed light on determining the training amount in practical application and help to better understand the role of training amount in some key ideas such as consolidation–reactivation, transfer, and stabilization in learning.
Traditional perceptual learning studies have hundreds or even thousands of training trials per day. Here we show that only 40 trials of daily practice were enough to trigger an improvement comparable to 1080 trials of daily practice. This finding supports a memory-reactivation framework for perceptual learning. Throughout multiple training sessions, the learning effects gained from individual training sessions transform from short- to long-term memory via a process named consolidation (McGaugh, 2000; Wright & Sabin, 2007). After the initial memory consolidation has been established, brief reactivations may trigger reconsolidation-like processes to improve the existing perceptual memory (Amar-Halpert et al., 2017; Bang et al., 2018). Amar-Halpert et al. (2017) have shown that decreasing the standard training amount (from 252 trials to 5 trials) on day 2 to day 4 led to no change in the overall learning effect. However, further decreasing the training from a standard to a small amount on day 1 led to a significant decrease in the overall learning effect. In the present study, all the subjects underwent a pretest of 400 trials for each condition, which established the new memory. After that, 40 trials of daily training were sufficient to reactivate the memory for reconsolidation. Our results indicate that motion perceptual learning, as a specific kind of procedural memory, might function via a consolidation-reactivation mechanism.
In contrast, overtraining might be detrimental to perceptual learning, which was referred to as perceptual deterioration (Mednick et al., 2002, 2005). Induced by too much training, perceptual deterioration is possibly due to sensory adaptation (Censor et al., 2006), strengthening less efficient neuronal connections and accumulating noise in the brain network (Censor & Sagi, 2008), or changes in the ability for attention to selectively enhance the responses of low-level sensory neurons (Mednick et al., 2005). In our study, perceptual deterioration might counteract the learning effect after a certain amount of practice, leading to saturated overall learning effects. The saturation of learning has also been observed in other visual, auditory, and motor learning studies (Amar-Halpert et al., 2017; Fox et al., 2016; Molloy et al., 2012; Wright & Sabin, 2007). These evidence suggests that independence of overall learning effect on daily training beyond a certain amount may be a general principle underlying skill learning.
Interestingly, we found that an increase in the daily training amount increased specificity. In other words, prolonged training decrease transfer. This effect may seem counterintuitive. As subjects become more proficient at a practiced task, we logically expect more transfer. However, many visual perceptual learning studies have reported that prolonged training increased the specificity of learning, with less transfer to untrained visual orientation or retinal locations (Hung & Seitz, 2014; Hussain et al., 2012; Jeter et al., 2010). The daily training amount may determine the degree of transfer together with another key factor, namely, stimuli variation. According to an “overfitting” account, increasing the training amount increases the chance of overfitting the neural system, which limits transfer. This phenomenon is likely to occur when the training data has a small variation, which may not well represent the feature space. In contrast, for data with a large variation that represents a broader feature space, increasing the training trials may not lead to overfitting (Lengyel & Fiser, 2019). In the present study, stimuli variation was introduced at the pretest and post-test stages, and was kept constant across the groups. This test gave subjects a sufficient amount of training (400 trials for each direction) over a relatively broad feature space (four motion direction with 0°, 30°, 60°, and 90° offset from the trained direction). During 8 training days, subjects received training on a specified motion direction with a near-threshold variation. Note that we used continuous staircases for each training day; except for the first staircase, each staircase started with the threshold derived from the preceding staircase. Our training protocol resembles the single prolonged staircase used in Hung and Seitz (2014) and other perceptual learning studies (Jehee et al., 2012; Schoups et al., 1995). Therefore, by increasing the training amount, we increased the number of near-threshold trials. Because such training over-represents a particular feature in the space, increasing the daily training amount leads to overfitting and greater specificity. Consistent with Hung and Seitz (2014), our results showed that prolonged training at threshold affects transfer in perceptual learning. It is worth mentioning that the account of stimuli variation and specificity in perceptual learning is reminiscent of Eleanor Gibson and James Gibson's ecological approach to perception, which suggested more variability led to a more general learning result (Gibson & Gibson, 1955). The 1 QUEST group might undergo a larger variation, therefore showing more transfer than the 27 QUEST group.
Perceptual learning with fine feature discrimination tasks usually results in high specificity and less transfer (e.g., Liu, 1999; Shiu & Pashler, 1992). Liu (1999) reported that, although learning in a motion discrimination task with a 3° directional difference was strongly specific to the training direction, learning transferred to new motion directions with an 8° directional difference. The idea that training precision modulates the degree of transfer in perceptual learning has been suggested in earlier psychophysical studies (Ahissar & Hochstein, 1997; Jeter et al., 2009) and is recently modeled using a deep neural network (Wenliang & Seitz, 2018).
In addition, our present findings provided the first piece of evidence for the long-term modulation effect of training amount on specificity, which persisted for at least 2 weeks after training. Future studies are needed to evaluate how the degree of transfer was modulated under different manipulations of the stimuli variations, such as changing the range of stimuli in the feature space, changing the probability distribution of stimuli (e.g., the ratio between the training amount of the trained and untrained features), and changing the time point the variation is presented (e.g., early, middle, or late training phase).
Training with a small daily amount provides a promising alternative protocol for perceptual learning studies in the future. When deciding on the training amount in practice, the following factors should be taken into consideration. (1) Generalization. Based on the current and previous learning studies with a motion or orientation discrimination paradigm, a larger daily training amount leads to less transfer to the untrained feature or spatial location. If one aims to induce a learning effect highly specific to the trained feature for a baseline control, a classical training paradigm with hundreds or thousands of daily training trials would be required. Otherwise, fewer trials (e.g., 40 trials or 5 trials in the middle phase of learning in Amar-Halpert et al. [2017]) in a daily session may be a choice for efficiency. (2) Stability. According to the hyperstabilizes account of overlearning, the learning effect becomes less susceptible to interference with an increasing daily training amount. Therefore, if subjects are trained with multiple tasks in the same session, hundreds of trials (e.g., more than 600 trials in Shibata et al., 2017) may still be needed. Finally, training with a small daily amount may have values in clinical applications, including the therapy of myopia (Yan et al., 2015), presbyopia (Polat, 2009; Sterkin et al., 2018), and low vision (Liu et al., 2007; Yu et al., 2010).
Recent advances in magnetic resonance spectroscopy techniques have pointed to a common neurochemical substrate mediating changes in stability and transfer in skill learning: γ-aminobutyric acid (GABA), a primary neurotransmitter of inhibitory processing (Bang et al., 2018; Robertson, 2018; Shibata et al., 2017). Prolonged training increased the GABAergic inhibitory process, which may facilitate learning through retuning the neurons to better signal the feature difference (Frangou et al., 2019) and stabilize the learned skill to protect it from interference (Shibata et al., 2017). A motor learning study with direct current stimulation showed that increasing the concentration of GABA in the motor cortex impairs transfer while decreasing the concentration of GABA enhances transfer (Waters-Metenier et al., 2014). These findings suggest a close link between stabilization and transfer at behavioral and neurochemical levels, indicating that prolonged practice may be responsible for stabilizing the memory and for reducing transfer (Robertson, 2018).
In sum, using a visual motion direction discrimination task, we showed that the learning effect is independent of daily training amount, whereas the transfer effect depends on the daily training amount. These findings not only help us to develop efficient protocols for research and clinical application, but also shed light on essential questions such as generalization in skill learning and stabilization in memory. In the future, neuroimaging and neuromodulation techniques are needed to further uncover the mechanisms underlying the relationship between training amount, transfer, and stabilization in learning and memory.
Acknowledgments
Supported by the National Natural Science Foundation of China (31930053, 31671168, 31421003, and 31971031), Beijing Municipal Science and Technology Commission (Z181100001518002), and Beijing Academy of Artificial Intelligence (BAAI).
Commercial relationships: none.
Corresponding authors: Fang Fang and Nihong Chen.
Email: ffang@pku.edu.cn, nihongch@tsinghua.edu.cn.
Address: School of Psychological and Cognitive Sciences, Peking University, Beijing 100871, People's Republic of China.
References
- Aberg, K. C., Tartaglia, E. M., & Herzog, M. H. (2009). Perceptual learning with Chevrons requires a minimal number of trials, transfers to untrained directions, but does not require sleep. Vision Research, 49(16), 2087–2094. [DOI] [PubMed] [Google Scholar]
- Ahissar, M., & Hochstein, S. (1997). Task difficulty and the specificity of perceptual learning. Nature, 387(6631), 401–406. [DOI] [PubMed] [Google Scholar]
- Amar-Halpert, R., Laor-Maayany, R., Nemni, S., Rosenblatt, J. D., & Censor, N. (2017). Memory reactivation improves visual perception. Nature Neuroscience, 20(10), 1325–1328. [DOI] [PubMed] [Google Scholar]
- Ashley, S., & Pearson, J. (2012). When more equals less: Overtraining inhibits perceptual learning owing to lack of wakeful consolidation. Proceedings of the Royal Society B: Biological Sciences, 279(1745), 4143–4147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banai, K., & Lavner, Y. (2014). The effects of training length on the perceptual learning of time-compressed speech and its generalization. Journal of the Acoustical Society of America, 136(4), 1908–1917. [DOI] [PubMed] [Google Scholar]
- Bang, J. W., Shibata, K., Frank, S. M., Walsh, E. G., Greenlee, M. W., Watanabe, T., & Sasaki, Y. (2018). Consolidation and reconsolidation share behavioural and neurochemical mechanisms. Nature Human Behaviour, 2(7), 507–513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Censor, N., Karni, A., & Sagi, D. (2006). A link between perceptual learning, adaptation and sleep. Vision Research, 46(23), 4071–4074. [DOI] [PubMed] [Google Scholar]
- Censor, N., & Sagi, D. (2008). Benefits of efficient consolidation: Short training enables long-term resistance to perceptual adaptation induced by intensive testing. Vision Research, 48(7), 970–977. [DOI] [PubMed] [Google Scholar]
- Censor, N., Sagi, D., & Cohen, L. G. (2012). Common mechanisms of human perceptual and motor learning. Nature Reviews Neuroscience, 13(9), 658–664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, N., Bi, T., Zhou, T., Li, S., Liu, Z., & Fang, F. (2015). Sharpened cortical tuning and enhanced cortico-cortical communication contribute to the long-term neural mechanisms of visual motion perceptual learning. NeuroImage, 115, 17–29. [DOI] [PubMed] [Google Scholar]
- Chung, S. T. L., Levi, D. M., & Li, R. W. (2006). Learning to identify contrast-defined letters in peripheral vision. Vision Research, 46(6–7), 1038–1047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fahle, M., & Poggio, T. (Eds.). (2002). Perceptual Learning. Cambridge, MA: MIT Press. [Google Scholar]
- Fox, O., Karni, A., & Adi-Japha, E. (2016). The consolidation of a motor skill in young adults with ADHD: Shorter practice can be better. Research in Developmental Disabilities, 51–52, 135–144. [DOI] [PubMed] [Google Scholar]
- Frangou, P., Emir, U. E., Karlaftis, V. M., Nettekoven, C., Hinson, E. L., Larcombe, S., & Kourtzi, Z. (2019). Learning to optimize perceptual decisions through suppressive interactions in the human brain. Nature Communications, 10(1), 474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibson, J. J., & Gibson, E. J. (1955). Perceptual learning: Differentiation or enrichment? Psychological Review, 62(1), 32–41. [DOI] [PubMed] [Google Scholar]
- Goldhacker, M., Rosengarth, K., Plank, T., & Greenlee, M. W. (2014). The effect of feedback on performance and brain activation during perceptual learning. Vision Research, 99, 99–110. [DOI] [PubMed] [Google Scholar]
- Hung, S.-C., & Seitz, A. R. (2014). Prolonged Training at Threshold Promotes Robust Retinotopic Specificity in Perceptual Learning. Journal of Neuroscience, 34(25), 8423–8431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Husk, J. S., Bennett, P. J., & Sekuler, A. B. (2007). Inverting houses and textures: Investigating the characteristics of learned inversion effects. Vision Research, 47(27), 3350–3359. [DOI] [PubMed] [Google Scholar]
- Hussain, Z., Sekuler, A. B., & Bennett, P. J. (2009). How much practice is needed to produce perceptual learning? Vision Research, 49(21), 2624–2634. [DOI] [PubMed] [Google Scholar]
- Hussain, Z., Bennett, P. J., & Sekuler, A. B. (2012). Versatile perceptual learning of textures after variable exposures. Vision Research, 61, 89–94. [DOI] [PubMed] [Google Scholar]
- Jehee, J. F. M., Ling, S., Swisher, J. D., van Bergen, R. S., & Tong, F. (2012). Perceptual learning selectively refines orientation representations in early visual cortex. Journal of Neuroscience, 32 (47), 16747–16753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeter, P. E., Dosher, B. A., Petrov, A., & Lu, Z. L. (2009). Task precision at transfer determines specificity of perceptual learning. Journal of Vision, 9(3), 1–1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeter, Pamela E., Dosher, B. A., Liu, S.-H., & Lu, Z.-L. (2010). Specificity of perceptual learning increases with increased training. Vision Research, 50(19), 1928–1940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karni, A., & Sagi, D. (1993). The time course of learning a visual skill. Nature, 365(6443), 250–252. [DOI] [PubMed] [Google Scholar]
- Lengyel, G., & Fiser, J. (2019). The relationship between initial threshold, learning, and generalization in perceptual learning. Journal of Vision, 19(4), 28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, Z. (1999). Perceptual learning in motion discrimination that generalizes across motion directions. Proceedings of the National Academy of Sciences of the United States of America, 96(24), 14085–14087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, L., Kuyk, T., & Fuhr, P. (2007). Visual search training in subjects with severe to profound low vision. Vision Research, 47, 2627–2636. [DOI] [PubMed] [Google Scholar]
- McGaugh, J. L. (2000). Memory—A century of consolidation. Science, 287(5451), 248–251. [DOI] [PubMed] [Google Scholar]
- Mednick, S. C., Arman, A. C., & Boynton, G. M. (2005). The time course and specificity of perceptual deterioration. Proceedings of the National Academy of Sciences of the United States of American, 102(10), 3881–3885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mednick, Sara C., Nakayama, K., Cantero, J. L., Atienza, M., Levin, A. A., Pathak, N., & Stickgold, R. (2002). The restorative effect of naps on perceptual deterioration. Nature Neuroscience, 5(7), 677–681. [DOI] [PubMed] [Google Scholar]
- Molloy, K., Moore, D. R., Sohoglu, E., & Amitay, S. (2012). Less is more: Latent learning is maximized by shorter training sessions in auditory perceptual learning. PLoS One, 7(5), e36929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ofen, N., Moran, A., & Sagi, D. (2007). Effects of trial repetition in texture discrimination. Vision Research, 47(8), 1094–1102. [DOI] [PubMed] [Google Scholar]
- Ofen-Noy, N., Dudai, Y., & Karni, A. (2003). Skill learning in mirror reading: How repetition determines acquisition. Cognitive Brain Research, 17(2), 507–521. [DOI] [PubMed] [Google Scholar]
- Polat, U. (2009). Making perceptual learning practical to improve visual functions. Vision Research, 49(21), 2566–2573. [DOI] [PubMed] [Google Scholar]
- Robertson, E. M. (2018). Memory instability as a gateway to generalization. PLoS Biology, 16(3), e2004633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roth, D. A.-E. (2005). A latent consolidation phase in auditory identification learning: Time in the awake state is sufficient. Learning & Memory, 12(2), 159–164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savion-Lemieux, T., & Penhune, V. B. (2005). The effects of practice and delay on motor skill learning and retention. Experimental Brain Research, 161(4), 423–431. [DOI] [PubMed] [Google Scholar]
- Schoups, A. A., Vogels, R., & Orban, G. A. (1995). Human perceptual learning in identifying the oblique orientation: Retinotopy, orientation specificity and monocularity. Journal of Physiology, 483(3), 797–810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shibata, K., Sasaki, Y., Bang, J. W., Walsh, E. G., Machizawa, M. G., Tamaki, M., . . . Watanabe, T. (2017). Overlearning hyperstabilizes a skill by rapidly making neurochemical processing inhibitory-dominant. Nature Neuroscience, 20(3), 470–475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shiu, L.-P., & Pashler, H. (1992). Improvement in line orientation discrimination is retinally local but dependent on cognitive set. Perception & Psychophysics, 52(5), 582–588. [DOI] [PubMed] [Google Scholar]
- Sigman, M., & Gilbert, C. D. (2000). Learning to find a shape. Nature Neuroscience, 3(3), 264–269. [DOI] [PubMed] [Google Scholar]
- Sterkin, A., Levy, Y., Pokroy, R., Lev, M., Levian, L., Doron, R., & Polat, U. (2018). Vision improvement in pilots with presbyopia following perceptual learning. Vision Research, 152 (2017), 61–73. [DOI] [PubMed] [Google Scholar]
- Waters-Metenier, S., Husain, M., Wiestler, T., & Diedrichsen, J. (2014). Bihemispheric transcranial direct current stimulation enhances effector-independent representations of motor synergy and sequence learning. Journal of Neuroscience, 34(3), 1037–1050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watson, A. B., & Pelli, D. G. (1983). Quest: A Bayesian adaptive psychometric method. Perception & Psychophysics, 33(2), 113–120. [DOI] [PubMed] [Google Scholar]
- Wenliang, L. K., & Seitz, A. R. (2018). Deep neural networks for modeling visual perceptual learning. Journal of Neuroscience, 38(27), 6028–6044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright, B. A., & Sabin, A. T. (2007). Perceptual learning: How much daily training is enough? Experimental Brain Research, 180(4), 727–736. [DOI] [PubMed] [Google Scholar]
- Yan, F.-F., Zhou, J., Zhao, W., Li, M., Xi, J., Lu, Z.-L., & Huang, C.-B. (2015). Perceptual learning improves neural processing in myopic vision. Journal of Vision, 15(10), 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu, D., Cheung, S., Legge, G. E., & Chung, S. T. L. (2010). Reading speed in the peripheral visual field of older adults: Does it benefit from perceptual learning? Vision Research, 50(9), 860–869. [DOI] [PMC free article] [PubMed] [Google Scholar]