

Abstract
General anesthesia is characterized by loss of consciousness, amnesia, analgesia, and immobility. Important molecular targets of general anesthetics have been identified, but the neural circuits underlying the discrete endpoints of general anesthesia remain incompletely understood. General anesthesia and natural sleep share the common feature of reversible unconsciousness, and recent developments in neuroscience have enabled elegant studies that investigate the brain nuclei and neural circuits underlying this important endpoint.
A common approach to measure cortical activity across the brain is electroencephalography (EEG), which can reflect local neuronal activity as well as connectivity among brain regions. The EEG oscillations observed during general anesthesia depend greatly on the anesthetic agent as well as dosing, and only some resemble those observed during sleep. For example, the EEG oscillations during dexmedetomidine sedation are similar to those of stage 2 non-rapid eye movement (NREM) sleep, but high doses of propofol and ether anesthetics produce burst suppression, a pattern that is never observed during natural sleep.
Sleep is primarily driven by withdrawal of subcortical excitation to the cortex, but anesthetics can directly act at both subcortical and cortical targets. While some anesthetics appear to activate specific sleep-active regions to induce unconsciousness, not all sleep-active regions play a significant role in anesthesia. Anesthetics also inhibit cortical neurons, and it is likely that each class of anesthetic drugs produces a distinct combination of subcortical and cortical effects that lead to unconsciousness. Conversely, arousal circuits that promote wakefulness are involved in anesthetic emergence and activating them can induce emergence and accelerate recovery of consciousness.
Modern neuroscience techniques that enable the manipulation of specific neural circuits have led to new insights into the neural circuitry underlying general anesthesia and sleep. In the coming years, we will continue to better understand the mechanisms that generate these distinct states of reversible unconsciousness.
Introduction:
General anesthetics produce a reversible state of unconsciousness, amnesia, analgesia, and immobility in response to pain. Since the introduction of general anesthesia into clinical practice, research in anesthetic mechanisms has been shaped by several influential theories. Claude Bernard proposed that anesthetics share a single, unified mechanism of action.1 This hypothesis was strongly reinforced by the findings of Meyer and Overton, who independently discovered that anesthetic potency correlated strongly with lipophilicity.2 Thus the lipid theory—stating that anesthetics act by disrupting the lipid bilayer of neuronal cell membranes—dominated thinking in the field for nearly a century.
In the 1980s, it was discovered that general anesthetics interact with hydrophobic sites on proteins,3 commencing a new era of identifying relevant protein targets for anesthetics. It is now evident that anesthetics act at specific molecular sites on ionotropic receptors, voltage-gated ion channels, and metabotropic receptors to produce neuronal inhibition.4 Substantial work has focused on anesthetic potentiation of γ-aminobutyric acid (GABA) receptors, which are responsible for the majority of inhibitory neurotransmission in the brain.5
Despite important advances in the molecular pharmacology of general anesthetics, the neuroanatomic sites of anesthetic action were largely unknown until it was reported that the spinal cord—not the brain—is the predominant site where anesthetics act to block motor responses to painful stimuli.6,7 Another seminal finding was that the microinjection of barbiturates into the upper brainstem of rodents produces general anesthesia.8 These important findings demonstrated that discrete neuroanatomic sites underlie different endpoints of anesthesia such as immobility and unconsciousness.
In recent years, there has been growing interest in studying how anesthetic drugs converge onto neural pathways of natural sleep to produce unconsciousness.9 Sleep serves as a useful metaphor for anesthesiologists to explain general anesthesia to patients, as the two share the behavioral trait of reversible unconsciousness.10 However, the states are distinct and clarity of terminology should be a goal in clinical and scientific discourse. In this review, the states of general anesthesia and sleep will be compared, highlighting important similarities and differences in neurophysiology and the neural circuit mechanisms underlying unconsciousness.
Comparison of Anesthesia and Sleep States
Important characteristics of general anesthesia and natural sleep are summarized in Table 1. Reversible loss of consciousness is a common feature, but the depth of unconsciousness differs markedly. During natural sleep, responsiveness to loud verbal or strong tactile stimulation remains intact, as well as the ability to maintain or restore a patent airway with adequate spontaneous ventilation. Based on the degree of unresponsiveness and airway patency, natural sleep is akin to the American Society of Anesthesiologists’ definition of “moderate sedation.”11 This is in contrast to general anesthesia, which is defined by behavioral unresponsiveness, even to painful stimuli. It is worth noting, however, that individuals with anatomically compromised airways may be vulnerable to reduced muscle activation and ventilatory drive during unconsciousness, in the setting of both general anesthesia and sleep (for a comprehensive review of this topic, please see the review by Hillman and Chung12). Individuals with obstructive sleep apnea are particularly vulnerable to developing a compromised upper airway when administered sedatives, analgesics and muscle relaxants. Preoperative screening tools to identify individuals with obstructive sleep apnea can mitigate vulnerabilities and other comorbidities that normally increase perioperative risk of hypoventilation and asphyxia during general anesthesia.13
Table 1.
A comparison of general anesthesia and sleep:
| Natural Sleep | General Anesthesia | |
|---|---|---|
| Loss of consciousness? | Yes | Yes |
| Reversible? | Yes | Yes |
| Arousable? | Yes- with sufficient external stimuli | No |
| Cyclical/Ultradian rhythm? | Yes | No |
| Susceptible to homeostatic pressures? | Yes | Maybe |
| Can relieve sleep pressure? | Yes | Maybe |
| Memory formation? | Important for memory processing and consolidation | No |
| Sense of time passing? | Yes | No |
Sleep is a natural process that occurs rhythmically, alternating between non-rapid eye movement (NREM) sleep and rapid eye movement (REM) sleep in 90–120 minute cycles throughout the night.14–16 In humans, NREM sleep is classified into three stages (N1-N3) from lighter to deeper sleep.16 In contrast, during REM sleep the cerebral cortex is active, more closely resembling the awake state.15
The two main mechanisms that affect the timing and duration of sleep are homeostatic mechanisms (i.e. the pressure to sleep that increases with prolonged wakefulness) and circadian rhythms.17 In animals, prolonged propofol exposure does not result in sleep deprivation, suggesting that propofol sedation satisfies the homeostatic need for sleep.18 However, in sleep-deprived animals sevoflurane exposure allows for selective recovery of NREM sleep,19 while REM sleep debt continues to accumulate.20 These studies suggest that different anesthetics may satisfy the homeostatic need for specific components of natural sleep. On the other hand, the NMDA receptor antagonist ketamine has been shown to inhibit light-entrainment to the suprachiasmatic nucleus clock (the process by which the circadian clock is synchronized to light cues), and extended exposure to GABAergic anesthetics is predicted to induce temporary phase-shifts in the sleep-wake cycle.21 Phase-shift effects could alter the trajectory of post-operative recovery, and clinical research on this topic is ongoing.21 Molecular studies have shown that exposure to sevoflurane, dexmedetomidine and propofol can each inhibit the expression of key circadian clock genes, though these effects were more notable when anesthetics were administered during the active phase (dark-phase for rodents) than during the inactive phase (light-phase for rodents).21 Overall, the accrued sleep debt is usually not considered when clinicians administer anesthetics for surgery, but ongoing research has the potential to improve postoperative care by examining postoperative sleep disturbances.
The effects of sleep and anesthesia on memory reveal fundamental differences between the two states. People awaken from sleep with the sense that time has passed, and sleep plays an important role in memory processing and consolidation.22,23 Upon emergence from general anesthesia, patients often describe feeling like no time had passed since induction, even after many hours of surgery. It has been reported that formation of new memories during emergence does not occur until patients can maintain sustained wakefulness.24 Anesthetics administered after a learning task impair memory consolidation and disrupt hippocampal activity.25–28 It is evident that general anesthetics hamper learning and memory, whereas sleep has beneficial effects on memory formation and consolidation.
EEG Findings in Sleep and General Anesthesia
A comparison of the frontal electroencephalogram (EEG) during natural sleep and general anesthesia reveals some shared features, but also many state- and drug-specific differences. Upon sleep onset, there is an increase in frontal slow (<1 Hz) and delta (1–4 Hz) oscillations, with frontal areas synchronizing earlier than other cortical areas.29 The three stages of human NREM sleep are further characterized based on unique EEG features such as high amplitude k-complexes, and sleep spindles in the 12–16 Hz range (Fig. 1A).17,30 The most prominent EEG feature shared between natural sleep and general anesthesia is the presence of large amplitude slow and delta oscillations.30,31 Slow-delta oscillations can be induced by dexmedetomidine, propofol, and ether anesthetics,32–35 and partly originate from a loss of excitatory inputs from the brainstem arousal nuclei to the cortex.30
Figure 1.
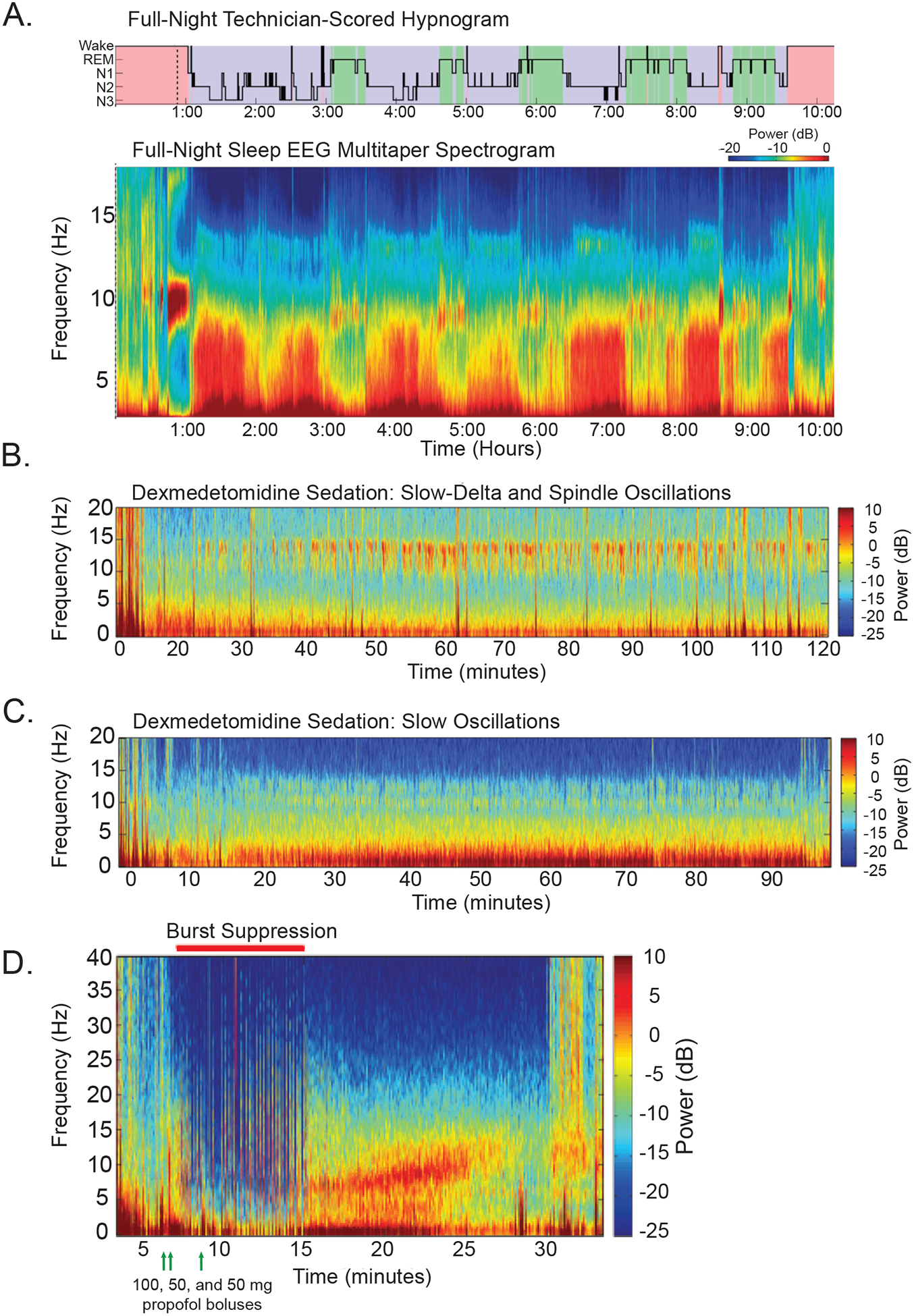
Electroencephalographic features distinct to general anesthesia and sleep. A comparison of frontal EEG recordings from humans during general anesthesia and sleep reveal similarities and differences in the neural oscillations observed. A) A full night’s natural sleep is shown as a spectrogram (lower panel) across hours of the night with the corresponding hypnogram (upper panel) identifying the stages of wake, rapid eye movement (REM) sleep, and non-rapid eye movement (NREM) stages 1–3 sleep (N1–3). The pattern of progressing sleep stages can be seen as cyclic increases in power (warmer red colors) for frequencies <10 Hz. Some sleep spindles can be seen at 12–15 Hz during the N1-N2 stages of sleep. B) An EEG of a patient receiving a continuous infusion of low dose dexmedetomidine (0.65 μh kg−1 h−1) shows mild sedation in which spindles (9–15 Hz) and increased power of slow-delta (0.1–4 Hz) oscillations can be seen. The presence of spindles and slow-delta activity is similar to those features seen during NREM stage 2 sleep. C) An EEG of a patient receiving a continuous infusion of higher dose dexmedetomidine (0.85 μg kg−1 h−1) shows moderate to deep sedation in which slow-delta oscillations are primarily increased. D) A bolus dose of propofol (100 mg, first green arrow) at minute 3 causes the EEG to transition to increased slow oscillation power. Two additional propofol boluses (50 mg each, green arrows) cause the EEG to shift to a burst suppression pattern from minutes 8 to 17. Afterward, the EEG shifts again to increased slow-delta and alpha (9–12 Hz) oscillation power (minute 17–25), until the patient begins to emerge at 24 minutes and the slow-delta and alpha oscillation power begins to decrease. Part A is adapted with permission from Prerau MJ, Brown RE, Bianchi MT, Ellenbogen JM, Purdon PL. Sleep Neurophysiological Dynamics Through the Lens of Multitaper Spectral Analysis. Physiology. 2017;32(1):60–92. Parts B-D are adapted with permission from Purdon PL, Sampson A, Pavone KJ, Brown EN. Clinical Electroencephalography for Anesthesiologists: Part I: Background and Basic Signatures. Anesthesiology. 2015;123(4):937–960. Abbreviations: Electroencephalogram (EEG), non-rapid eye movement (NREM), and rapid eye movement (REM).
Dexmedetomidine, a selective α2-adrenergic receptor agonist, produces EEG findings most closely resembling NREM sleep (Fig. 1B–C). Spindles, which are commonly observed in NREM Stage 2, are also present in the frontal EEG during dexmedetomidine sedation, with peak power around 13 Hz.34,36,37 At higher doses that produce deeper sedation, spindles disappear and slow-delta oscillations dominate the EEG, similar to NREM Stage 3 sleep.36,38
Propofol enhances inhibitory neurotransmission by potentiating the activity of GABA at synaptic and extrasynaptic GABAA receptors.9 Compared to dexmedetomidine, propofol produces larger amplitude slow oscillations representative of longer periods of neuronal silence and enhanced disconnection from the environment.33,34 Upon loss of consciousness with propofol, there is an anteriorization of alpha power from the occipital cortex to the frontal cortex that is reversed upon return of consciousness.39–41 Unlike sleep spindles that are intermittent and brief, propofol induces continuous alpha (8–12 Hz) oscillations.39,42 Propofol also increases functional connectivity between the anterior and posterior cingulate cortices.33 At very high doses, propofol produces burst suppression, a pattern of intermittent bursts of activity punctuated by electrical silence, a pattern that is not observed during natural sleep (Fig. 1D).
Ether anesthetics such as isoflurane and sevoflurane are known to modulate several important molecular targets to produce neuronal inhibition, including GABAA receptors, glutamate receptors, and two-pore domain potassium channels.43 Like propofol, sevoflurane increases slow-delta and alpha oscillations. The latter are focused in the frontal cortex and have a peak around 10 Hz during unconsciousness.35,44 However, at higher doses sevoflurane also increases theta (4–8 Hz) oscillations that are not observed during propofol anesthesia.35 At very high doses, ether anesthetics also produce burst suppression like propofol.
Ketamine is a dissociative anesthetic that is thought to act primarily by blocking excitatory NMDA-type glutamate receptors.4 At high doses that induce unconsciousness, ketamine produces a unique “gamma burst” pattern consisting of gamma (30–70 Hz) oscillations alternating with slow-delta oscillations.45,46 Although gamma oscillations can be associated with wakeful consciousness, those observed during ketamine anesthesia are distinct and may be the result of ketamine preferentially inhibiting cortical interneurons, resulting in increased pyramidal neuronal activity.45 In addition, ketamine-induced EEG patterns may originate from decreased brainstem inputs to the thalamus and cortex or from the direct inhibitory actions of ketamine on the thalamus.45
In summary, while many anesthetics produce slow, delta, and alpha oscillations that resemble certain EEG features of NREM sleep, each drug class produces a unique constellation of neurophysiological signatures that are dose- and state-specific. Compared to other anesthetics and sedatives, dexmedetomidine produces EEG patterns that are most similar to NREM sleep, suggesting that the states may share similar underlying mechanisms at the level of neural circuits. Whereas, high doses propofol and ether anesthetics produce burst suppression, and ketamine produces “gamma burst” activity, none of which are observed during natural sleep.
Cortical Effects of Anesthesia and Sleep
During sleep, unconsciousness is primarily driven by sleep-active, subcortical areas that inhibit ascending arousal nuclei.47,48 This inhibition decreases the release of excitatory neurotransmitters—such as acetylcholine, histamine, and norepinephrine—and reduces cortical activation.17 In contrast, anesthetic agents can act directly on a variety of cortical receptors, decreasing the spontaneous activity of cortical neurons49 and the sensitivity of those neurons to subcortical inputs.50
Many recent studies have revealed that the direct actions of anesthetics on cortical neurons—or “top-down” mechanisms—are important for anesthetic-induced unconsciousness.51 Anesthetics may induce unconsciousness by direct actions on cortical neurons, which have been demonstrated in cortical slice preparations. Isoflurane, enflurane, halothane, propofol, pentobarbital, and ketamine have all been shown to inhibit spontaneous action potentials in cortical neurons.49,52 Isoflurane decreases the amplitude of excitatory synaptic potentials,50,53 while ketamine preferentially inhibits NMDA receptors on GABAergic interneurons, leading to disinhibition of pyramidal neurons.54–56
Activity of genetically distinct neuronal types is also affected in ways that causally influence the cortical EEG. For instance, parvalbumin-positive and pyramidal neurons fire most actively during the peak of the slow oscillation during both sleep and chloral hydrate anesthesia.57,58 Such parallels between sleep and anesthesia may underlie the similar slow and delta oscillations observed in both states that are traditionally thought to underlie loss of consciousness.59 The cortical actions of anesthetics may point to a shared mechanism that produces anesthetic-induced unconsciousness: despite affecting diverse molecular targets, propofol, ketamine, and sevoflurane each reduce frontal-to-parietal brain communication.60
Anesthetics likely induce unconsciousness at both the subcortical and cortical levels, acting on subcortical areas to decrease arousal while simultaneously affecting the cortex to degrade the contents of consciousness.61 In support of this hypothesis, the delivery of carbachol to the prefrontal cortex of rats mimics the stimulation of the subcortical ascending cholinergic inputs to the cortex and restores wakefulness during sevoflurane anesthesia.62 However, carbachol delivery does not restore high gamma (85–155 Hz) connectivity between cortical areas, suggesting that the contents of consciousness remain disrupted.63 Additionally, anesthetics modulate information exchanges between auditory stimuli and the primary auditory cortex action potentials that are not linked to the transition to loss of consciousness.64 Together, these results suggest that direct actions on the cortex play a critical role in anesthetic-induced unconsciousness.
Subcortical Circuits Underlying Anesthesia and Sleep
Based on their profound inhibitory effects on the brain, systemically administered general anesthetics were historically thought to function by widespread neural suppression. This notion was challenged by a study that identified a focal zone in the rat midbrain that produced rapid and reversable loss of consciousness after microinjections of pentobarbital.8 Referred to as the mesopontine tegmentum anesthetic locus (MPTA), this region (Fig. 2) has since been implicated in sleep65 and described as a consciousness “switch” for anesthesia.66 While not all anesthetics are equally impacted by MPTA lesions,67 there is substantial evidence supporting the “shared circuits hypothesis” of anesthesia and sleep.
Figure 2.
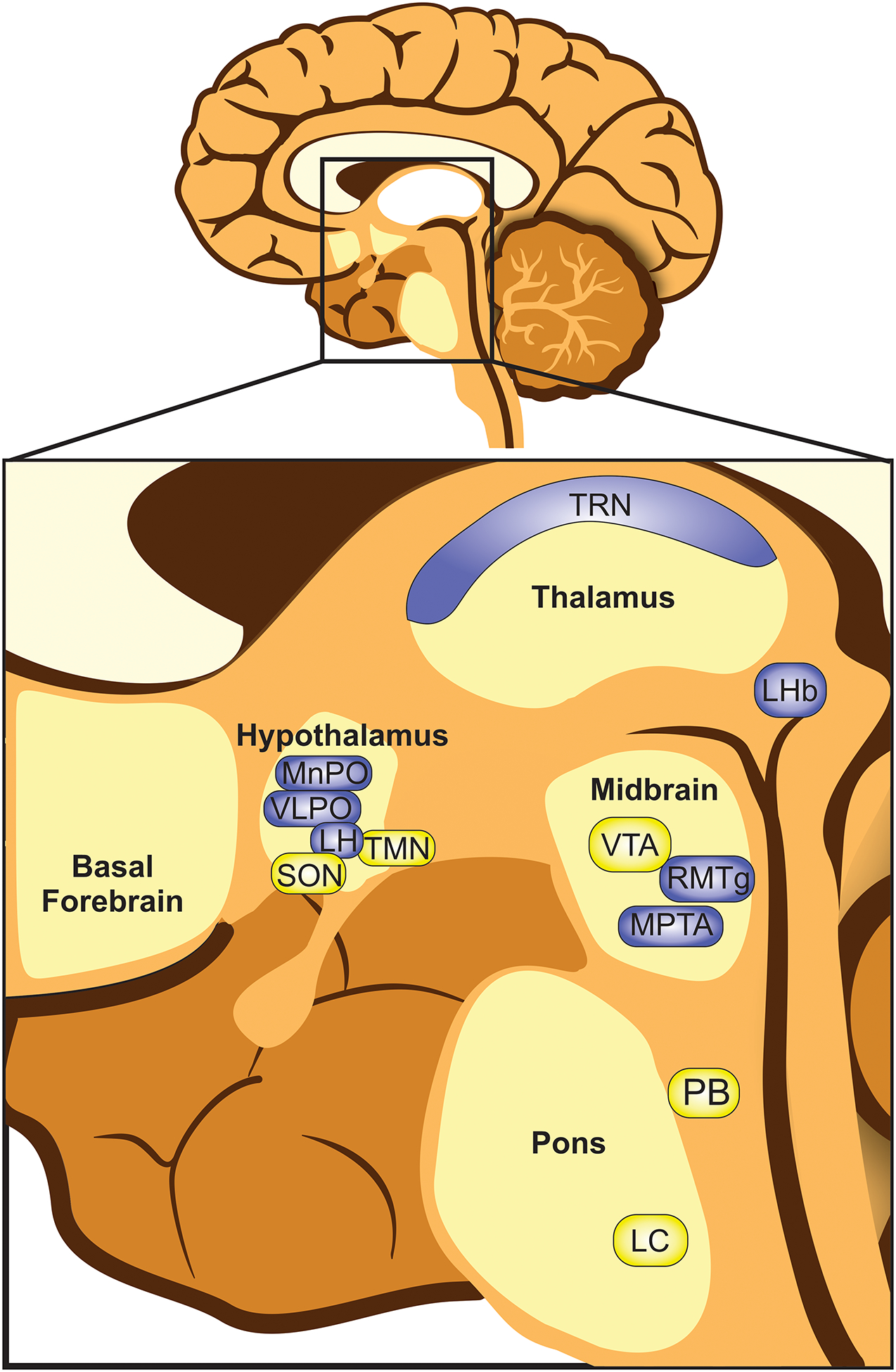
Subcortical areas implicated in the regulation of sleep and anesthesia-induced loss of consciousness. Subcortical areas are shown in the magnified inset of a human brain. Loss of consciousness generally involves silencing wake-active nuclei (yellow) and stimulating sleep-active nuclei (blue). Hypothalamic nuclei: lateral hypothalamus (LH), ventrolateral preoptic area (VLPO), median preoptic area (MnPO), supraoptic nucleus (SON) and the tuberomammillary nucleus (TMN). Thalamic nuclei: thalamic reticular nucleus (TRN). Lateral habenula (LHb). Midbrain: ventral tegmental nucleus (VTA), rostromedial tegmental nucleus (RMTg), and mesopontine tegmental anesthesia area (MPTA). Pons: locus coeruleus (LC) and parabrachial nucleus (PB). Note: not all structures are located midline. For a comprehensive review of the putative sites of action of distinct anesthetics on the brain, please see the review by Brown, et al.107
The role of the ventrolateral preoptic nucleus (VLPO, Fig. 2) in promoting sleep has long been supported by early investigations of neural activity across arousal states, and from lesion studies.67,68 Advanced optogenetic, pharmacogenetic, and RNA sequencing techniques have further characterized the sleep-promoting neurons in the VLPO, which send inhibitory projections to arousal circuits to both induce and maintain sleep.69–71 While GABAergic anesthetics have been shown to activate sleep-promoting VLPO neurons,72,73 lesion studies have yielded conflicting results. Ablating the VLPO attenuates loss of righting induced by propofol,74 but effects on isoflurane are inconsistent with reports showing both increased and decreased sensitivity, possibly due to the long-term sleep disruptions caused by VLPO lesions.73,75 More recently, selective targeting of GABAergic and glutamatergic VLPO neurons using designer receptors has confirmed that isoflurane does not rely on the VLPO to induce loss of consciousness, in contrast to natural sleep.71 Similarly, the median preoptic nucleus (MnPO), a region both functionally and structurally connected to the VLPO,76 initiates and maintains NREM sleep but not isoflurane-induced unconsciousness.71
Another inhibitory brain region that may be involved in both general anesthesia and sleep is the thalamic reticular nucleus (TRN), which is involved in NREM sleep68 and generates spindle oscillations.77 TRN stimulation further suppresses cortical activity during isoflurane anesthesia,68 and increases sensitivity to propofol through GABAB receptors.78,79 A recent study found a novel group of GABAergic and glutamatergic neurons in the supraoptic nucleus (SON) of the hypothalamus that was activated by multiple anesthetics and that promoted NREM sleep.80 Using modern genetic techniques, the group also identified neurons which serve as a common substrate for both anesthesia and analgesia in the central amygdala,81 illustrating the potential for new tools to advance our understanding of the neural circuitry underlying anesthesia and sleep.82
Investigations of the lateral habenula (LHb), a region that modulates midbrain dopaminergic neurotransmission, have demonstrated its involvement in facilitating NREM sleep,100,101 as well as maintaining circadian rhythms,83–85 homeostatic sleep regulation,86 and NREM sleep stability.87 A variety of general anesthetics activate LHb neurons, including propofol, ketamine, pentobarbital, and chloral hydrate,72,88 and inhibiting LHb glutamate release reduces sensitivity to propofol. These findings implicate brain areas downstream from the LHb in anesthetic induction.87 The LHb sends projections to multiple regions including the GABAergic rostromedial tegmental nucleus (RMTg), which in turn sends inhibitory projections to dopamine neurons in the ventral tegmental area (VTA).89,90 Although studies of the RMTg have been limited in the context of sleep and anesthesia, it appears to play a role in NREM sleep.90 Furthermore, inhibition of its downstream targets, the dopaminergic VTA and substantia nigra pars compacta, promotes sleep,90–92 and selectively activating a subpopulation of GABAergic VTA projections to the lateral hypothalamus increases anesthetic depth.93 Another target of the VTA implicated in sleep is the core of the nucleus accumbens (NAc),94 which regulates NREM sleep.94,95 Interestingly, although GABAA agonists infused into the NAc do not induce unconsciousness, they do increase sensitivity to halothane anesthesia.96
Additionally, general anesthetics directly inhibit wake-active nuclei, such as the tuberomammilary nucleus (TMN) and locus coeruleus (LC). The TMN is the only brain site containing histaminergic neurons,97 which are active during waking periods and quiescent in sleep. Both GABAA receptor activation98 and NMDA receptor antagonism99 in the TMN promote NREM sleep. Likewise, GABAergic anesthetics inhibit TMN neural activity.72 Although TMN lesions increase sensitivity to isoflurane, they do not affect pentobarbital or propofol anesthesia.100
The LC is comprised of arousal-promoting noradrenergic neurons, which are most active during wake, less active during NREM sleep, and silent during REM sleep.101 Optogenetic studies have demonstrated that reduced LC firing promotes memory consolidation and decreases sensory-evoked awakenings during REM sleep.102,103 Both propofol and isoflurane inhibit LC neural firing.104 However, while stimulating LC neurons impairs isoflurane-induced unconsciousness,105 LC inhibition is not required for propofol’s effects.74 Moreover, halothane has no impact on LC firing rate.104
Lastly, although cholinergic neurons in the laterodorsal and peduculopontine tegmentum (LDT, PPT) drive the transition from NREM to REM sleep,106 they are not activated by a broad range of anesthetics, including isoflurane, pentobarbital, and ketamine.72 Overall, while anesthesia and sleep share some overlapping subcortical mechanisms (Fig. 2), there are many differences that are often anesthetic-specific. For a comprehensive review of the putative sites of action of distinct anesthetics on the brain, please see the review by Brown, et al.107
Neural Circuits of Arousal and Anesthetic Emergence
Recently, there has been considerable interest in investigating the role of subcortical arousal circuits in anesthetic emergence.108 The central medial thalamus, part of the ascending reticular activating system, has been studied widely in the context of arousal and anesthetic emergence. Neurons in the central medial thalamus are inhibited by anesthetics.109–112 Microinjection studies of the central medial thalamus have shown that activating nicotinic acetylcholine receptors or blocking Kv1 potassium channels increases neural activity and reverses sevoflurane-induced unconsciousness.110,113,114 Microinjections of norepinephrine into the central medial thalamus during propofol anesthesia lead to EEG signs of cortical arousal and accelerates time to emergence.115 These findings suggest that activating neurons in the central medial thalamus during anesthesia can shorten time to emergence and, in some cases, reverse anesthetic-induced unconsciousness.
The arousal-promoting orexinergic system also has notable effects on emergence. Suppressing orexinergic signaling delays emergence from ether anesthetics, but not the alkane anesthetic halothane.104,116–119 Plasma orexin-A levels increase in humans upon emergence from propofol and sevoflurane anesthesia, consistent with the notion that the orexinergic system is involved in emergence.120,121
The cholinergic system, which is critical for promoting arousal, also modulates emergence from anesthesia. Intracerebroventricular microinfusions of drugs promoting cholinergic neurotransmission during continuous isoflurane anesthesia lead to EEG and behavioral signs of arousal, although return of righting reflex was not observed.122 Microinjecting the cholinergic agonist, carbachol, into the prefrontal cortex restores the righting reflex and other conscious behaviors in rats during sevoflurane anesthesia,62 suggesting that cholinergic neurotransmission plays a critical role in emergence.
Enhancing arousal-promoting dopaminergic neurotransmission from the ventral tegmental area (VTA)—particularly via activation of dopamine type 1 (D1) receptors—has been shown to restore consciousness in rodents anesthetized with propofol and ether anesthetics.123–129 Of the multiple dopaminergic projections from the VTA, inputs to the nucleus accumbens provide the most potent arousal stimulus from natural sleep.92
The parabrachial nucleus is a glutamatergic brainstem arousal nucleus involved in wakefulness and anesthetic emergence, with projections to the basal forebrain, thalamus, and arousal-promoting orexinergic neurons of the lateral hypothalamus.130 Parabrachial neurons are suppressed by anesthetics and activated during emergence.130,131 Optogenetic, chemogenetic, and electrical stimulation of parabrachial neurons generates EEG and behavioral evidence of arousal during sleep and general anesthesia by propofol, isoflurane and sevoflurane.130–132
Neuromodulators have also been found to play a role in emergence. Studies have shown that adenosine antagonists, including caffeine, can facilitate emergence from isoflurane and propofol anesthesia in humans and rodents.133–135 More recently, a study in humans found that intravenous caffeine accelerated time to emergence from isoflurane anesthesia.136 Finally, the benzodiazepine antagonist flumazenil has been shown to have modest effects on accelerating emergence time from general anesthesia in rodents and humans.137–139
Discussion
General anesthetics are a diverse class of drugs that appear to produce unconsciousness by targeting numerous neural circuits in cortical and subcortical regions. Each anesthetic produces distinct cortical EEG signatures that are dose-dependent, which suggests that multiple neural circuits contribute to producing unconsciousness. This is evident when comparing dissimilar agents such as the GABAergic anesthetic propofol and the NMDA receptor antagonist ketamine. However, the existence of similar neural oscillations during general anesthesia and sleep (e.g. slow-delta oscillations) suggests that some general anesthetics converge onto sleep-active neural circuits to induce unconsciousness. However, general anesthetics also directly inhibit cortical neurons, as well as subcortical arousal-promoting neurons. All these actions likely contribute to suppress wakefulness. In addition, activating arousal nuclei accelerates emergence and restores consciousness in anesthetized subjects.
Recent advances in neuroscience such as optogenetics and chemogenetics have enabled detailed studies of the neural circuit mechanisms underlying anesthesia and sleep.82 Recently, a new genetic technique of tracking, targeting, and stimulating specific neural populations has allowed the targeting of specific behavior-associated brain loci, including “anesthesia-activated neurons” (AANs) located in the supraoptic nucleus of the hypothalamus.140 These AAN neurons were not only activated by a range of anesthetics, but their activation also promoted slow-wave sleep, illustrating the potential of these emerging techniques to further advance our knowledge of the neural circuitry underlying complex phenomena. Gaining insights into the neural circuitry underlying general anesthesia and sleep will likely lead to improved methods for controlling arousal states in the perioperative setting.
Acknowledgements:
This research was supported in part by grants R01-GM126155, P01- GM118629, and F32-GM137491 from the National Institutes of Health (Bethesda, Maryland), and by Scholar Award 220020406 from the James S. McDonnell Foundation (Saint Louis, Missouri). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. Authors OAM, ERZ, KFV, RK, EDM and CJN declare no conflicts of interest. Author KS is a consultant to Takeda Pharmaceuticals. Authors OAM, ERZ, KFV, RK and EDM helped to write and edit the manuscript. Author CJN helped to conceptually design and edit the manuscript. Author KS helped to conceptually design, write, and edit the manuscript.
Glossary of Terms:
- AANs
Anesthesia-activated neurons
- D1
Dopamine type 1
- EEG
Electroencephalogram
- Kv1 potassium channels
Voltage-gated potassium channel family 1
- LHb
Lateral habenula
- LH
Lateral hypothalamus
- LDT, PPT
Laterodorsal and peduculopontine tegmentum
- LC
Locus coeruleus
- MPTA
Mesopontine tegmentum anesthetic locus
- NMDA
N-methyl-D-aspartate
- NREM
Non-rapid eye movement
- NAc
Nucleus accumbens
- PB
Parabrachial nucleus
- REM
Rapid eye movement
- RMTg
Rostromedial tegmental nucleus
- SON
Supraoptic nucleus
- TRN
Thalamic reticular nucleus
- TMN
Tuberomammilary nucleus
- VTA
Ventral tegmental area
- VLPO
Ventrolateral preoptic nucleus
- GABAA receptors
γ-aminobutyric acid type A receptors
- GABAB receptors
γ-aminobutyric acid type B receptors
Footnotes
Conflicts of Interest: Authors OAM, ERZ, KFV, RK, EDM and CJN declare no conflicts of interest. Author KS is a consultant to Takeda Pharmaceuticals.
References:
- 1.Perouansky M The Quest for a Unified Model of Anesthetic Action: A Century in Claude Bernard’s Shadow. Anesthesiology. 2012;117(3):465–474. [DOI] [PubMed] [Google Scholar]
- 2.Meyer H Zur Theorie der Alkoholnarkose. Archiv für experimentelle Pathologie und Pharmakologie. 1899;42(2):109–118. [Google Scholar]
- 3.Franks NP, Lieb WR. Do general anaesthetics act by competitive binding to specific receptors? Nature. 1984;310(5978):599–601. [DOI] [PubMed] [Google Scholar]
- 4.Solt K, Forman SA. Correlating the clinical actions and molecular mechanisms of general anesthetics. Current opinion in anaesthesiology. 2007;20(4):300–306. [DOI] [PubMed] [Google Scholar]
- 5.Franks NP. General anaesthesia: from molecular targets to neuronal pathways of sleep and arousal. Nature reviews. 2008;9(5):370–386. [DOI] [PubMed] [Google Scholar]
- 6.Rampil IJ. Anesthetic potency is not altered after hypothermic spinal cord transection in rats. Anesthesiology. 1994;80(3):606–610. [DOI] [PubMed] [Google Scholar]
- 7.Antognini JF, Schwartz K. Exaggerated anesthetic requirements in the preferentially anesthetized brain. Anesthesiology. 1993;79(6):1244–1249. [DOI] [PubMed] [Google Scholar]
- 8.Devor M, Zalkind V. Reversible analgesia, atonia, and loss of consciousness on bilateral intracerebral microinjection of pentobarbital. Pain. 2001;94(1):101–112. [DOI] [PubMed] [Google Scholar]
- 9.Hemmings HC Jr., Riegelhaupt PM, Kelz MB, et al. Towards a Comprehensive Understanding of Anesthetic Mechanisms of Action: A Decade of Discovery. Trends Pharmacol Sci. 2019;40(7):464–481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mashour GA, Pal D. Interfaces of sleep and anesthesia. Anesthesiol Clin. 2012;30(2):385–398. [DOI] [PubMed] [Google Scholar]
- 11.Committee on Quality M, Departmental A. Continuum of Depth of Sedation: Definition of General Anesthesia and Levels of Sedation/Analgesia. In: American Society of Anesthesiologists; 2019. [Google Scholar]
- 12.Hillman DR, Chung F. Anaesthetic management of sleep-disordered breathing in adults. Respirology. 2017;22(2):230–239. [DOI] [PubMed] [Google Scholar]
- 13.Nagappa M, Wong J, Singh M, Wong DT, Chung F. An update on the various practical applications of the STOP-Bang questionnaire in anesthesia, surgery, and perioperative medicine. Current opinion in anaesthesiology. 2017;30(1):118–125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Miyazaki S, Liu C-Y, Hayashi Y. Sleep in vertebrate and invertebrate animals, and insights into the function and evolution of sleep. Neurosci Res. 2017;118:3–12. [DOI] [PubMed] [Google Scholar]
- 15.Peever J, Fuller PM. The Biology of REM Sleep. Curr Biol. 2017;27(22):R1237–R1248. [DOI] [PubMed] [Google Scholar]
- 16.Le Bon O Relationships between REM and NREM in the NREM-REM sleep cycle: a review on competing concepts. Sleep Med. 2020;70:6–16. [DOI] [PubMed] [Google Scholar]
- 17.Brown RE, Basheer R, McKenna JT, Strecker RE, McCarley RW. Control of sleep and wakefulness. Physiol Rev. 2012;92(3):1087–1187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tung A, Lynch JP, Mendelson WB. Prolonged sedation with propofol in the rat does not result in sleep deprivation. Anesth Analg. 2001;92(5):1232–1236. [DOI] [PubMed] [Google Scholar]
- 19.Pal D, Lipinski WJ, Walker AJ, Turner AM, Mashour GA. State-specific effects of sevoflurane anesthesia on sleep homeostasis: selective recovery of slow wave but not rapid eye movement sleep. Anesthesiology. 2011;114(2):302–310. [DOI] [PubMed] [Google Scholar]
- 20.Pick J, Chen Y, Moore JT, et al. Rapid eye movement sleep debt accrues in mice exposed to volatile anesthetics. Anesthesiology. 2011;115(4):702–712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Poulsen RC, Warman GR, Sleigh J, Ludin NM, Cheeseman JF. How does general anaesthesia affect the circadian clock? Sleep Med Rev. 2018;37:35–44. [DOI] [PubMed] [Google Scholar]
- 22.Ahuja S, Chen RK, Kam K, Pettibone WD, Osorio RS, Varga AW. Role of normal sleep and sleep apnea in human memory processing. Nat Sci Sleep. 2018;10:255–269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Vertes RP, Linley SB. No cognitive processing in the unconscious, Anesthetic-Like, state of sleep. J Comp Neurol. 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dutton RC, Smith WD, Smith NT. Wakeful response to command indicates memory potential during emergence from general anesthesia. J Clin Monit. 1995;11(1):35–40. [DOI] [PubMed] [Google Scholar]
- 25.Zurek AA, Yu J, Wang D-S, et al. Sustained increase in α5GABAA receptor function impairs memory after anesthesia. J Clin Invest. 2014;124(12):5437–5441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zhang J, Zhang X, Jiang W. Propofol impairs spatial memory consolidation and prevents learning-induced increase in hippocampal matrix metalloproteinase-9 levels in rat. Neuroreport. 2013;24(15):831–836. [DOI] [PubMed] [Google Scholar]
- 27.Moon DU, Esfahani-Bayerl N, Finke C, et al. Propofol modulates early memory consolidation in humans. eNeuro. 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Dai S, Perouansky M, Pearce RA. Isoflurane enhances both fast and slow synaptic inhibition in the hippocampus at amnestic concentrations. Anesthesiology. 2012;116(4):816–823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Marzano C, Moroni F, Gorgoni M, Nobili L, Ferrara M, De Gennaro L. How we fall asleep: regional and temporal differences in electroencephalographic synchronization at sleep onset. Sleep Med. 2013;14(11):1112–1122. [DOI] [PubMed] [Google Scholar]
- 30.Akeju O, Brown EN. Neural oscillations demonstrate that general anesthesia and sedative states are neurophysiologically distinct from sleep. Curr Opin Neurobiol. 2017;44:178–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mölle M, Marshall L, Gais S, Born J. Grouping of spindle activity during slow oscillations in human non-rapid eye movement sleep. J Neurosci. 2002;22(24):10941–10947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Akeju O, Hamilos AE, Song AH, Pavone KJ, Purdon PL, Brown EN. GABAA circuit mechanisms are associated with ether anesthesia-induced unconsciousness. Clin Neurophysiol. 2016;127(6):2472–2481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Murphy M, Bruno MA, Riedner BA, et al. Propofol anesthesia and sleep: a high-density EEG study. Sleep. 2011;34(3):283–291a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Akeju O, Pavone KJ, Westover MB, et al. A comparison of propofol- and dexmedetomidine-induced electroencephalogram dynamics using spectral and coherence analysis. Anesthesiology. 2014;121(5):978–989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Akeju O, Westover MB, Pavone KJ, et al. Effects of sevoflurane and propofol on frontal electroencephalogram power and coherence. Anesthesiology. 2014;121(5):990–998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Akeju O, Kim SE, Vazquez R, et al. Spatiotemporal Dynamics of Dexmedetomidine-Induced Electroencephalogram Oscillations. PLoS One. 2016;11(10):e0163431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Huupponen E, Maksimow A, Lapinlampi P, et al. Electroencephalogram spindle activity during dexmedetomidine sedation and physiological sleep. Acta Anaesthesiol Scand. 2008;52(2):289–294. [DOI] [PubMed] [Google Scholar]
- 38.Scheinin A, Kallionpää RE, Li D, et al. Differentiating Drug-related and State-related Effects of Dexmedetomidine and Propofol on the Electroencephalogram. Anesthesiology. 2018;129(1):22–36. [DOI] [PubMed] [Google Scholar]
- 39.Purdon PL, Pierce ET, Mukamel EA, et al. Electroencephalogram signatures of loss and recovery of consciousness from propofol. Proc Natl Acad Sci U S A. 2013;110(12):E1142–1151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ching S, Cimenser A, Purdon PL, Brown EN, Kopell NJ. Thalamocortical model for a propofol-induced alpha-rhythm associated with loss of consciousness. Proc Natl Acad Sci U S A. 2010;107(52):22665–22670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Supp GG, Siegel M, Hipp JF, Engel AK. Cortical hypersynchrony predicts breakdown of sensory processing during loss of consciousness. Curr Biol. 2011;21(23):1988–1993. [DOI] [PubMed] [Google Scholar]
- 42.Steriade M, McCormick DA, Sejnowski TJ. Thalamocortical oscillations in the sleeping and aroused brain. Science. 1993;262(5134):679–685. [DOI] [PubMed] [Google Scholar]
- 43.Franks NP. General anaesthesia: from molecular targets to neuronal pathways of sleep and arousal. Nat Rev Neurosci. 2008;9(5):370–386. [DOI] [PubMed] [Google Scholar]
- 44.Gugino LD, Chabot RJ, Prichep LS, John ER, Formanek V, Aglio LS. Quantitative EEG changes associated with loss and return of consciousness in healthy adult volunteers anaesthetized with propofol or sevoflurane. Br J Anaesth. 2001;87(3):421–428. [DOI] [PubMed] [Google Scholar]
- 45.Akeju O, Song AH, Hamilos AE, et al. Electroencephalogram signatures of ketamine anesthesia-induced unconsciousness. Clin Neurophysiol. 2016;127(6):2414–2422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Maksimow A, Särkelä M, Långsjö JW, et al. Increase in high frequency EEG activity explains the poor performance of EEG spectral entropy monitor during S-ketamine anesthesia. Clin Neurophysiol. 2006;117(8):1660–1668. [DOI] [PubMed] [Google Scholar]
- 47.Saper CB, Scammell TE, Lu J. Hypothalamic regulation of sleep and circadian rhythms. Nature. 2005;437(7063):1257–1263. [DOI] [PubMed] [Google Scholar]
- 48.Schwartz MD, Kilduff TS. The Neurobiology of Sleep and Wakefulness. Psychiatric Clinics of North America. 2015;38(4):615–644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Antkowiak B Different Actions of General Anesthetics on the Firing Patters of Neocortical Neurons Mediated by the GABAA Receptor. Anesthesiology. 1999;91:500–511. [DOI] [PubMed] [Google Scholar]
- 50.Puil E, El-Beheiry H. Anaesthetic suppression of transmitter actions in neocortex. British Journal of Pharmacology. 1990;101(1):61–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mashour GA. Top-down mechanisms of anesthetic-induced unconsciousness. Frontiers in Systems Neuroscience. 2014;8(June):1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Antkowiak B, Helfrich-Forster C. Effects of small concentrations of volatile anesthetics on action potential firing of neocortical neurons in vitro. In. Vol 881998:1592–1605. [DOI] [PubMed] [Google Scholar]
- 53.El-Beheiry H, Puil E. Anaesthetic depression of excitatory synaptic transmission in neocortex. Experimental Brain Research. 1989;77(1):87–93. [DOI] [PubMed] [Google Scholar]
- 54.Ali F, Gerhard D, Sweasy K, et al. Ketamine disinhibits dendrites and enhances calcium signals in prefrontal dendritic spines. Nature Communications. 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Olney JW, Newcomer JW, Farber NB. NMDA receptor hypofunction model of schizophrenia. Journal of Psychiatric Research. 1999;33:523–533. [DOI] [PubMed] [Google Scholar]
- 56.Shen G, Han F, Shi WX. Effects of Low Doses of Ketamine on Pyramidal Neurons in Rat Prefrontal Cortex. Neuroscience. 2018;384:178–187. [DOI] [PubMed] [Google Scholar]
- 57.Niethard N, Ngo H-VVV, Ehrlich I, Born J. Cortical circuit activity underlying sleep slow oscillations and spindles. Proceedings of the National Academy of Sciences of the United States of America. 2018;115(39):E9220–E9229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ushimaru M, Kawaguchi Y. Temporal structure of neuronal activity among cortical neuron subtypes during slow oscillations in anesthetized rats. Journal of Neuroscience. 2015;35(34):11988–12001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Gemignani A, Menicucci D, Laurino M, et al. Linking sleep slow oscillations with consciousness theories: New vistas on slow wave sleep unconsciousness. Archives Italiennes de Biologie. 2015;153(2–3):135–143. [DOI] [PubMed] [Google Scholar]
- 60.Lee U, Ku S, Noh G, Baek S, Choi B, Mashour GA. Disruption of Frontal-Parietal Communication by Ketamine, Propofol, and Sevoflurane. Anesthesiology. 2013;118(6). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Mashour GA, Hudetz AG. Bottom-Up and Top-Down Mechanisms of General Anesthetics Modulate Different Dimensions of Consciousness. Frontiers in Neural Circuits. 2017;11(June):1–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Pal D, Dean JG, Liu T, et al. Differential Role of Prefrontal and Parietal Cortices in Controlling Level of Consciousness. Curr Biol. 2018;28(13):2145–2152 e2145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Pal D, Li D, Dean JG, et al. Level of Consciousness Is Dissociable from Electroencephalographic Measures of Cortical Connectivity, Slow Oscillations, and Complexity. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2020;40(3):605–618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Banks MI, Moran NS, Krause BM, Grady SM, Uhlrich DJ, Manning KA. Altered stimulus representation in rat auditory cortex is not causal for loss of consciousness under general anaesthesia. British Journal of Anaesthesia. 2018;121(3):605–615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lanir-Azaria S, Meiri G, Avigdor T, Minert A, Devor M. Enhanced wakefulness following lesions of a mesopontine locus essential for the induction of general anesthesia. Behav Brain Res. 2018;341:198–211. [DOI] [PubMed] [Google Scholar]
- 66.Sukhotinsky I, Minert A, Soja P, Devor M. Mesopontine Switch for the Induction of General Anesthesia by Dedicated Neural Pathways. Anesth Analg. 2016;123(5):1274–1285. [DOI] [PubMed] [Google Scholar]
- 67.Minert A, Baron M, Devor M. Reduced Sensitivity to Anesthetic Agents upon Lesioning the Mesopontine Tegmental Anesthesia Area in Rats Depends on Anesthetic Type. Anesthesiology. 2020;132(3):535–550. [DOI] [PubMed] [Google Scholar]
- 68.Lewis LD, Voigts J, Flores FJ, et al. Thalamic reticular nucleus induces fast and local modulation of arousal state. Elife. 2015;4:e08760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Chung S, Weber F, Zhong P, et al. Identification of preoptic sleep neurons using retrograde labelling and gene profiling. Nature. 2017;545(7655):477–481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Saito YC, Tsujino N, Hasegawa E, et al. GABAergic neurons in the preoptic area send direct inhibitory projections to orexin neurons. Front Neural Circuits. 2013;7:192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Vanini G, Bassana M, Mast M, et al. Activation of Preoptic GABAergic or Glutamatergic Neurons Modulates Sleep-Wake Architecture, but Not Anesthetic State Transitions. Curr Biol. 2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Lu J, Nelson LE, Franks N, Maze M, Chamberlin NL, Saper CB. Role of endogenous sleep-wake and analgesic systems in anesthesia. J Comp Neurol. 2008;508(4):648–662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Moore JT, Chen J, Han B, et al. Direct activation of sleep-promoting VLPO neurons by volatile anesthetics contributes to anesthetic hypnosis. Curr Biol. 2012;22(21):2008–2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zhang Y, Yu T, Yuan J, Yu B-W. The ventrolateral preoptic nucleus is required for propofol-induced inhibition of locus coeruleus neuronal activity. Neurol Sci. 2015;36(12):2177–2184. [DOI] [PubMed] [Google Scholar]
- 75.Eikermann M, Vetrivelan R, Grosse-Sundrup M, et al. The ventrolateral preoptic nucleus is not required for isoflurane general anesthesia. Brain Res. 2011;1426:30–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Walter A, van der Spek L, Hardy E, Bemelmans AP, Rouach N, Rancillac A. Structural and functional connections between the median and the ventrolateral preoptic nucleus. Brain Struct Funct. 2019;224(9):3045–3057. [DOI] [PubMed] [Google Scholar]
- 77.Halassa MM, Siegle JH, Ritt JT, Ting JT, Feng G, Moore CI. Selective optical drive of thalamic reticular nucleus generates thalamic bursts and cortical spindles. Nat Neurosci. 2011;14(9):1118–1120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Zhang Y, Fu B, Liu C, et al. Activation of noradrenergic terminals in the reticular thalamus delays arousal from propofol anesthesia in mice. FASEB J. 2019;33(6):7252–7260. [DOI] [PubMed] [Google Scholar]
- 79.Zhang Y, Liu C, Zhang L, et al. Effects of Propofol on Electrical Synaptic Strength in Coupling Reticular Thalamic GABAergic Parvalbumin-Expressing Neurons. Front Neurosci. 2020;14:364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.A Common Neuroendocrine Substrate for Diverse General Anesthetics and Sleep 2019. [DOI] [PMC free article] [PubMed]
- 81.Hua T, Chen B, Lu D, et al. General anesthetics activate a potent central pain-suppression circuit in the amygdala. Nat Neurosci. 2020;23(7):854–868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Melonakos ED, Moody OA, Nikolaeva K, Kato R, Nehs CJ, Solt K. Manipulating Neural Circuits in Anesthesia Research. Anesthesiology: The Journal of the American Society of Anesthesiologists. 2020;133(1):19–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Goldstein R A GABAergic habenulo-raphe pathway mediation of the hypnogenic effects of vasotocin in cat. Neuroscience. 1983;10(3):941–945. [DOI] [PubMed] [Google Scholar]
- 84.Bourdy R, Barrot M. A new control center for dopaminergic systems: pulling the VTA by the tail. Trends Neurosci. 2012;35(11):681–690. [DOI] [PubMed] [Google Scholar]
- 85.Zhao H, Rusak B. Circadian firing-rate rhythms and light responses of rat habenular nucleus neurons in vivo and in vitro. Neuroscience. 2005;132(2):519–528. [DOI] [PubMed] [Google Scholar]
- 86.Zhang B, Gao Y, Li Y, Yang J, Zhao H. Sleep Deprivation Influences Circadian Gene Expression in the Lateral Habenula. Behav Neurol. 2016;2016:7919534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Gelegen C, Miracca G, Ran MZ, et al. Excitatory Pathways from the Lateral Habenula Enable Propofol-Induced Sedation. Curr Biol. 2018;28(4):580–587.e585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Abulafia R, Zalkind V, Devor M. Cerebral activity during the anesthesia-like state induced by mesopontine microinjection of pentobarbital. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2009;29(21):7053–7064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Jhou TC, Geisler S, Marinelli M, Degarmo BA, Zahm DS. The mesopontine rostromedial tegmental nucleus: A structure targeted by the lateral habenula that projects to the ventral tegmental area of Tsai and substantia nigra compacta. J Comp Neurol. 2009;513(6):566–596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Yang S-R, Hu Z-Z, Luo Y-J, et al. The rostromedial tegmental nucleus is essential for non-rapid eye movement sleep. PLoS Biol. 2018;16(4):e2002909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Sun H-X, Wang D-R, Ye C-B, et al. Activation of the ventral tegmental area increased wakefulness in mice. Sleep Biol Rhythms. 2017;15(2):107–115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Eban-Rothschild A, Rothschild G, Giardino WJ, Jones JR, de Lecea L. VTA dopaminergic neurons regulate ethologically relevant sleep-wake behaviors. Nature neuroscience. 2016;19(10):1356–1366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Yin L, Li L, Deng J, et al. Optogenetic/Chemogenetic Activation of GABAergic Neurons in the Ventral Tegmental Area Facilitates General Anesthesia via Projections to the Lateral Hypothalamus in Mice. Front Neural Circuits. 2019;13:73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Qiu MH, Liu W, Qu WM, Urade Y, Lu J, Huang ZL. The role of nucleus accumbens core/shell in sleep-wake regulation and their involvement in modafinil-induced arousal. PLoS One. 2012;7(9):e45471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Oishi Y, Xu Q, Wang L, et al. Slow-wave sleep is controlled by a subset of nucleus accumbens core neurons in mice. Nat Commun. 2017;8(1):734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Ma J, Leung LS. Limbic system participates in mediating the effects of general anesthetics. Neuropsychopharmacology. 2006;31(6):1177–1192. [DOI] [PubMed] [Google Scholar]
- 97.Panula P, Pirvola U, Auvinen S, Airaksinen MS. Histamine-immunoreactive nerve fibers in the rat brain. Neuroscience. 1989;28(3):585–610. [DOI] [PubMed] [Google Scholar]
- 98.Xie JF, Fan K, Wang C, et al. Inactivation of the Tuberomammillary Nucleus by GABAA Receptor Agonist Promotes Slow Wave Sleep in Freely Moving Rats and Histamine-Treated Rats. Neurochem Res. 2017;42(8):2314–2325. [DOI] [PubMed] [Google Scholar]
- 99.Yin D, Dong H, Wang TX, et al. Glutamate Activates the Histaminergic Tuberomammillary Nucleus and Increases Wakefulness in Rats. Neuroscience. 2019;413:86–98. [DOI] [PubMed] [Google Scholar]
- 100.Luo T, Leung LS. Involvement of tuberomamillary histaminergic neurons in isoflurane anesthesia. Anesthesiology. 2011;115(1):36–43. [DOI] [PubMed] [Google Scholar]
- 101.Aston-Jones G, Bloom FE. Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J Neurosci. 1981;1(8):876–886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Hayat H, Regev N, Matosevich N, et al. Locus coeruleus norepinephrine activity mediates sensory-evoked awakenings from sleep. Sci Adv. 2020;6(15):eaaz4232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Swift KM, Gross BA, Frazer MA, et al. Abnormal Locus Coeruleus Sleep Activity Alters Sleep Signatures of Memory Consolidation and Impairs Place Cell Stability and Spatial Memory. Curr Biol. 2018;28(22):3599–3609 e3594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Gompf H, Chen J, Sun Y, Yanagisawa M, Aston-Jones G, Kelz MB. Halothane-induced hypnosis is not accompanied by inactivation of orexinergic output in rodents. Anesthesiology. 2009;111(5):1001–1009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Vazey EM, Aston-Jones G. Designer receptor manipulations reveal a role of the locus coeruleus noradrenergic system in isoflurane general anesthesia. Proc Natl Acad Sci USA. 2014;111(10):3859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Van Dort CJ, Zachs DP, Kenny JD, et al. Optogenetic activation of cholinergic neurons in the PPT or LDT induces REM sleep. Proc Natl Acad Sci U S A. 2015;112(2):584–589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Brown EN, Purdon PL, Van Dort CJ. General anesthesia and altered states of arousal: a systems neuroscience analysis. Annu Rev Neurosci. 2011;34:601–628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Kelz MB, García PS, Mashour GA, Solt K. Escape From Oblivion: Neural Mechanisms of Emergence From General Anesthesia. Anesth Analg. 2019;128(4):726–736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Flood P, Ramirez-Latorre J, Role L. Alpha 4 beta 2 neuronal nicotinic acetylcholine receptors in the central nervous system are inhibited by isoflurane and propofol, but alpha 7-type nicotinic acetylcholine receptors are unaffected. Anesthesiology. 1997;86(4):859–865. [DOI] [PubMed] [Google Scholar]
- 110.Lioudyno MI, Birch AM, Tanaka BS, et al. Shaker-related potassium channels in the central medial nucleus of the thalamus are important molecular targets for arousal suppression by volatile general anesthetics. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2013;33(41):16310–16322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Vertes RP, Hoover WB, Rodriguez JJ. Projections of the central medial nucleus of the thalamus in the rat: Node in cortical, striatal and limbic forebrain circuitry. Neuroscience. 2012;219:120–136. [DOI] [PubMed] [Google Scholar]
- 112.Violet JM, Downie DL, Nakisa RC, Lieb WR, Franks NP. Differential sensitivities of mammalian neuronal and muscle nicotinic acetylcholine receptors to general anesthetics. Anesthesiology. 1997;86(4):866–874. [DOI] [PubMed] [Google Scholar]
- 113.Alkire MT, Asher CD, Franciscus AM, Hahn EL. Thalamic microinfusion of antibody to a voltage-gated potassium channel restores consciousness during anesthesia. Anesthesiology. 2009;110(4):766–773. [DOI] [PubMed] [Google Scholar]
- 114.Alkire MT, McReynolds JR, Hahn EL, Trivedi AN. Thalamic microinjection of nicotine reverses sevoflurane-induced loss of righting reflex in the rat. Anesthesiology. 2007;107(2):264–272. [DOI] [PubMed] [Google Scholar]
- 115.Fu B, Yu T, Yuan J, Gong X, Zhang M. Noradrenergic transmission in the central medial thalamic nucleus modulates the electroencephalographic activity and emergence from propofol anesthesia in rats. J Neurochem. 2017;140(6):862–873. [DOI] [PubMed] [Google Scholar]
- 116.Dong H-L, Fukuda S, Murata E, Higuchi T. Excitatory and inhibitory actions of isoflurane on the cholinergic ascending arousal system of the rat. Anesthesiology. 2006;104(1):122–133. [DOI] [PubMed] [Google Scholar]
- 117.Kelz MB, Sun Y, Chen J, et al. An essential role for orexins in emergence from general anesthesia. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(4):1309–1314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Wasilczuk AZ, Maier KL, Kelz MB. The Mouse as a Model Organism for Assessing Anesthetic Sensitivity. Methods Enzymol. 2018;602:211–228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Zhang L-N, Yang C, Ouyang P-R, et al. Orexin-A facilitates emergence of the rat from isoflurane anesthesia via mediation of the basal forebrain. Neuropeptides. 2016;58:7–14. [DOI] [PubMed] [Google Scholar]
- 120.Kushikata T, Yoshida H, Kudo M, Kudo T, Hirota K. Changes in plasma orexin A during propofol-fentanyl anaesthesia in patients undergoing eye surgery. British journal of anaesthesia. 2010;104(6):723–727. [DOI] [PubMed] [Google Scholar]
- 121.Kushikata T, Yoshida H, Kudo M, Kudo T, Hirota K. Plasma orexin A increases at emergence from sevoflurane-fentanyl anesthesia in patients undergoing ophthalmologic surgery. Neurosci Lett. 2010;482(3):212–215. [DOI] [PubMed] [Google Scholar]
- 122.Hudetz AG, Wood JD, Kampine JP. Cholinergic reversal of isoflurane anesthesia in rats as measured by cross-approximate entropy of the electroencephalogram. Anesthesiology. 2003;99(5):1125–1131. [DOI] [PubMed] [Google Scholar]
- 123.Kenny JD, Taylor NE, Brown EN, Solt K. Dextroamphetamine (but Not Atomoxetine) Induces Reanimation from General Anesthesia: Implications for the Roles of Dopamine and Norepinephrine in Active Emergence. PloS one. 2015;10(7):e0131914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Taylor NE, Chemali JJ, Brown EN, Solt K. Activation of D1 dopamine receptors induces emergence from isoflurane general anesthesia. Anesthesiology. 2013;118(1):30–39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Solt K, Cotten JF, Cimenser A, Wong KFK, Chemali JJ, Brown EN. Methylphenidate actively induces emergence from general anesthesia. Anesthesiology. 2011;115(4):791–803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Chemali JJ, Van Dort CJ, Brown EN, Solt K. Active emergence from propofol general anesthesia is induced by methylphenidate. Anesthesiology. 2012;116(5):998–1005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Solt K, Van Dort CJ, Chemali JJ, Taylor NE, Kenny JD, Brown EN. Electrical stimulation of the ventral tegmental area induces reanimation from general anesthesia. Anesthesiology. 2014;121(2):311–319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Taylor NE, Van Dort CJ, Kenny JD, et al. Optogenetic activation of dopamine neurons in the ventral tegmental area induces reanimation from general anesthesia. Proceedings of the National Academy of Sciences of the United States of America. 2016;113(45):12826–12831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Oishi Y, Suzuki Y, Takahashi K, et al. Activation of ventral tegmental area dopamine neurons produces wakefulness through dopamine D(2)-like receptors in mice. Brain structure & function. 2017;222(6):2907–2915. [DOI] [PubMed] [Google Scholar]
- 130.Muindi F, Kenny JD, Taylor NE, et al. Electrical stimulation of the parabrachial nucleus induces reanimation from isoflurane general anesthesia. Behavioural brain research. 2016;306:20–25. [DOI] [PubMed] [Google Scholar]
- 131.Luo T, Yu S, Cai S, et al. Parabrachial Neurons Promote Behavior and Electroencephalographic Arousal From General Anesthesia. Front Mol Neurosci. 2018;11:420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Wang T-X, Xiong B, Xu W, et al. Activation of Parabrachial Nucleus Glutamatergic Neurons Accelerates Reanimation from Sevoflurane Anesthesia in Mice. Anesthesiology. 2019;130(1):106–118. [DOI] [PubMed] [Google Scholar]
- 133.Fong R, Khokhar S, Chowdhury AN, et al. Caffeine accelerates recovery from general anesthesia via multiple pathways. J Neurophysiol. 2017;118(3):1591–1597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Van Dort CJ, Baghdoyan HA, Lydic R. Adenosine A(1) and A(2A) receptors in mouse prefrontal cortex modulate acetylcholine release and behavioral arousal. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2009;29(3):871–881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Wang Q, Fong R, Mason P, Fox AP, Xie Z. Caffeine accelerates recovery from general anesthesia. J Neurophysiol. 2014;111(6):1331–1340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Fong R, Wang L, Zacny JP, et al. Caffeine Accelerates Emergence from Isoflurane Anesthesia in Humans: A Randomized, Double-blind, Crossover Study. Anesthesiology. 2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Dahaba AA, Bornemann H, Rehak PH, Wang G, Wu XM, Metzler H. Effect of flumazenil on bispectral index monitoring in unpremedicated patients. Anesthesiology. 2009;110(5):1036–1040. [DOI] [PubMed] [Google Scholar]
- 138.Weinbroum AA, Geller E. Flumazenil improves cognitive and neuromotor emergence and attenuates shivering after halothane-, enflurane- and isoflurane-based anesthesia. Can J Anaesth. 2001;48(10):963–972. [DOI] [PubMed] [Google Scholar]
- 139.Safavynia SA, Keating G, Speigel I, et al. Effects of gamma-Aminobutyric Acid Type A Receptor Modulation by Flumazenil on Emergence from General Anesthesia. Anesthesiology. 2016;125(1):147–158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Jiang-Xie LF, Yin L, Zhao S, et al. A Common Neuroendocrine Substrate for Diverse General Anesthetics and Sleep. Neuron. 2019;102(5):1053–1065 e1054. [DOI] [PMC free article] [PubMed] [Google Scholar]