

Abstract
The Heat shock proteins 90 family plays pivotal roles in root growth and plant development. Its isoforms have been identified in several plant species with transcripts presents in almost all stages of plant growth. However, its functional relevance has not been completely established. Therefore, in this study, we provide evidence about the role of SlHSP90.2 in tomato (Solanum lycopersicum) root development. Using tomato cultivars with differing root phenotypes, we have shown that SlHSP90.2 transcripts are in accordance with root architecture, i.e. high rooting cultivars had more expression of SlHSP90.2 as compared to low rooting cultivars. Moreover, overexpression of SlHSP90.2 gene in transgenic tomato plants showed significant increase in root biomass and architecture, as evident from the analysis of fresh and dry weights of root and shoot samples, primary root length and length and number of lateral roots. The transgenic lines are also more tolerant to salinity and drought stresses. The results of the present study suggest that genetic manipulation of HSP90.2 homologs in other crops can offer promising leads to develop plant with better root biomass and architecture and improved agronomics traits, like better water and mineral absorption, salinity and drought tolerance potential.
Keywords: Overexpression, qRT-PCR, Root growth, SlHSP90.2, Tomato, Transgenic
Introduction
The 90-kDa heat shock protein (HSP90) is an essential molecular chaperon playing key roles in eukaryotic cells. It is highly abundant in physiological conditions comprising of about 1–2% of cellular proteins in most plant tissues (Ticha et al. 2020). HSP90 play a vital role in signal transduction, protein folding, growth and development (Krishna and Gloor 2001; Young et al. 2001; Riggs et al. 2004). Environmental and genetic perturbation affects both the HSP90s and their interactors, leading to alternation of intricate molecular networks vital for broad phenotypic plasticity (Zabinsky et al. 2019). Although HSP90-regulated interaction networks have been thoroughly studied in animals, yet involvement in plant developmental processes is just starting to be understood (Ticha et al. 2020).
Different isoforms of HSP90 have been identified in plants. For instance, tomato (Zai et al. 2015) and Arabidopsis (Milioni et al. 1997) have seven members each, rice has nine (Wang et al. 2014), tobacco eleven (Song et al. 2019) and wheat has only three isoforms (Wang et al. 2011). Among these, the Arabidopsis isoforms have been extensively utilized for a better understanding of the role of HSP90 family in plants. AtHSP90.1 was found to be expressed in early stage of embryo development which increased significantly just before embryo maturation, with most prominent expression occurring in cotyledons. Similarly, AtHSP90.2 and AtHSP90.3 are highly expressed in tips of embryo, cotyledons and in the elongation zone of root apical meristem (Yabe et al. 1994; Prasinos et al. 2005). Furthermore, being a member of the HSP family, AtHS90.1 transcripts are also induced by heat shock. The rice HSP90 genes are expressed more strongly in stem and root than leaf, however, the expression pattern changes during stress conditions (Liu et al. 2006). Wheat HSP90 gene, TaHsp90.1 has more transcript levels in the stamen, while TaHsp90.2 and TaHsp90.3 are more abundant in the vegetative and reproductive organs (Wang et al. 2011). Soybean, an important oilseed crop, also possesses HSP90 isoforms. GmHsp90B1 and GmHsp90B2 expressed more in leaves and GmHSP90C1 in flowers. Interestingly, GmHsp90A3, GmHsp90A6 and GmHsp90B are expressed more in roots as compared to the other HSP90 isoforms in soybean. In addition, GmHsp90 gene were also induced by heat, salt and other osmotic stresses (Xu et al. 2013). In tobacco, out of eleven NtHSP90 members reported, the transcripts of six members –NtHSP90.4, NtHSP90.5, NtHSP90.8, NtHSP90.9, NtHSP90.10 and NtHSP90.11- were notably upregulated under cold and heat stresses (Song et al. 2019). In Potato, members of HSP90 showed differential expression in plant tissues, like flower and tuber (mature) and are also induced in response to heat, salinity and drought stresses (Li et al. 2020).
Despite the presence of HSP90 members in the different plant species, their functional relevance in plant development has not been fully established. HSP90 was found to affect hypocotyls, root elongation and primary root development (Queitsch et al. 2002; Sangster et al. 2008). Overexpressing OsHSP90 in tobacco showed more elongated roots as compared to untransformed controls (Liu et al. 2006). It has been observed that the role of HSP90 in root development is associated with cell proliferation in the apical meristem zone and cell elongation in the elongation zone through their interaction with their co-chaperones and interactors.
Tomato (Solanum lycopersicum) is being used as a model crop for genomics studies because of its small genome size and well-established transformation techniques. Among tomato HSP90 isoforms, SlHSP90.1 is highly expressed in mature fruit (Zai et al. 2015). SlHSP90.3 and SlHSP90.4 genes exhibit higher transcript levels in different tissues like, blossom buds, young leaves, and fruit of tomato. SlHSP90.5 was high, while SlHSP90.6 was low in different tissue tested. The expression level of SlHSP90.7 was low in the leaves, but high in the remaining tissues. SlHSP90 members are also upregulated under stress condition (Zai et al. 2015). In a previous work by our group five expressed sequence tag (EST) libraries were constructed in tomato to identify root-specific genes, genes involved in temporal (early and late) root development and genes involved in lateral and profuse branching. SlHSP90.2 was identified as one of the candidates and its expression in tomato roots was validated by semi quantitative RT-PCR (Kalidhasan et al. 2015). In this study we report that (i) SlHSP90.2 is expressed differentially in genotypes with differing root phenotypes (ii) its overexpression in transgenic lines resulted in enhanced root biomass and altered root architecture and (iii) the transgenic lines were more resilient to salinity and drought stress conditions.
Materials and methods
Plant material and growth conditions
Tomato (Solanum lycopersicum cv. Pusa Ruby) seeds were obtained from the National Seed Corporation, New Delhi, India. Seeds were surface sterilized, rinsed in sterile water, and sown in Magenta boxes containing full strength Murashige and Skoog medium (Himedia). Additionally, 3% (w/v) sucrose was added and after pH adjustment to 5.8, 0.8% (w/v) agar was added. Tomato plants were grown in temperature-controlled growth room with 14 h light and 10 h dark at 25 °C. Four-week-old seedlings were transferred to greenhouse conditions with natural day length (14 h light in summer and 10 h light in winter) and controlled temperature (25 °C during day and 18 °C during night) and watering (12 h watering cycle) conditions.
Comparison of morphogenetic traits of different tomato cultivars
Eight different genotypes of tomato with contrasting root development attributes Arka Ashish, Arka Meghali, IIHR-2624, IIHR-2617, IIHR-2615, IIHR-2613, Arka Saurabh and Arka Aahuti were obtained from IIHR (ICAR-Indian Institute of Horticultural Research), Bengaluru, Karnataka, India. Plants were sampled at seedling, vegetative, and mature stages. For examining vegetative and mature stages, plants were uprooted and soil clumps stuck to the tomato roots were cleaned carefully to ensure that the roots are intact and undamaged. Subsequently, the plants were photographed and the total root length was measured using a ruler. Simultaneously, the fresh root weight was also measured. Three plants were used per cultivar per replicate. The entire experiment was repeated twice with consistent results.
RNA extraction and quantitative real-time PCR (qRT-PCR)
Total RNA was isolated from either 8- or 40-day-old shoots or roots samples using the NucleoSpin® RNA Plant kit (MACHEREY–NAGEL/TaKaRa) according to the manufacturer’s protocol and quantified by nanodrop. Isolated RNA was used for cDNA synthesis using Verso cDNA synthesis kit. The full-length cDNA sequences of SlHSP90.2 were obtained from Solanaceae Genomics Network and used for designing primers for the gene (Table 1). The specificities of the primers were confirmed by running the RT-PCR product on agarose gel and subsequently the primers were used for qRT-PCR analysis. The amplicon size of each of them was less than 200 bp. qRT- PCR was performed in a Roche Light Cycler® 96 System (Roche) using SYBR Green master mix (DyNAmo Color Flash SYBR Green qPCR kit F-416L, Thermo scientific). The thermal cycling conditions were 95 °C for 60 s followed by 40 cycles of 95 °C for 10 s, 55 °C for 10 s, 72 °C for 10 s, and melting 95 °C for 5 s, 65 °C for 60 s, 97 °C for 1 s. Relative gene expression was calculated according to the delta-delta Ct method of the system. Tomato Actin gene was used as internal reference for all the qRT–PCR analysis. The expression analyses data represent means of three independent biological replicates each with three technical replicates.
Table 1.
Primers used in this study
| S. no. | Name of primer | Primer sequence 5′-3′ |
|---|---|---|
| 1 | SlHSP90.2 |
FP: 5’-GGGCCCATGTCGGACGTAGAGACGTTTG-3′ RP: 5′-CCCGGGTTAATCAACCTCCTCCATCTTGCT-3′ |
| 2 | qRT-SlHSP90.2 |
FP: 5′-GCTTCTCAGTCTTATCATCAAC-3′ RP: 5′-GCTTGCTCTTGTCGGTTA-3’ |
| 3 | Sl-Actin |
FP: 5′-TTATCACCATTGGTGCTGAG-3′ RP: 5′-CGATGTTTCCATACAGATCCTT-3’ |
| 4 | CaMV35S | 5′-GATTTCAGCGTGTCCTCTCC-3’ |
| 5 | HSP RT | 5′- GCTTGCTCTTGTCGGTTA-3’ |
| 6 | HSP TC | 5′-GCACAGGCACTTAGGGACTC-3’ |
| 7 | NOSA | 5′-TTTGCGCGCTATATTTTGTTT-3’ |
SlHSP90.2 isolation, plasmid construction and Agrobacterium-mediated tomato transformation
The full-length coding sequence of SlHSP90.2 was amplified from 8-day-old root cDNA of Pusa Ruby. The amplified SlHSP90.2 gene was cloned in pJET1.2 vector and sequenced.100% similarity was observed when the obtained sequence was aligned with native SlHSP90.2 gene from Pusa Ruby. To overexpress SlHSP90.2 in tomato it was subcloned into the binary vector pCAMBIA2301 under the control of the CaMV35S promoter with ApaI, SmaI and PstI restriction sites. The constructed plasmid was transformed into Agrobacterium tumefaciens strain AGL1, which was subsequently used for tomato transformation following the protocol of Sharma et al. (2009).
The integration of transgene in different tomato transgenic lines was confirmed by genotyping PCR and further validation was done using GUS reporter assay. The expression of the transgene was analyzed in three independent transgenic lines using qRT-PCR. The primers used for genotyping and qRT-PCR (Table 1). The phenotypic characterization of the transgenic lines was done as described earlier.
Leaf disc assay
Leaves from transgenic and control plants were cut into two pieces and floated on salt solutions with different concentration, viz.0, 200, 400, 600, and 800 mM NaCl solution for 72 h. At the end of 72 h chlorophyll retention in each sample was estimated using the protocol as in Porra (2002). The total chlorophyll content was calculated using the following formula.
[Chla] = 12.00 × A663.8–3.11 × A646.8; [Chlb] = 20.78 × 646.8–4.88 × A663.8; [Chla + b] = 17.67 × A646.8 + 7.12 × A663.8.
Analysis for drought tolerance
The drought tolerance assay was performed by placing surface-sterilized seeds of Pusa Ruby (control) and transgenic lines of SlHSP90.2 on cotton beds saturated with 200 mM mannitol in glass bottles. The germination percentage, root and shoot lengths and fresh weights were analyzed after 8 days of incubation at 25 °C. The treated seedlings were transferred back to water and grown for another 8 days for checking the recovery by measuring root length and fresh and dry weight of roots.
Results and discussion
SlHSP90.2 is expressed more in roots than in shoots of tomato
A previous study from our group has identified a pool of 36 ESTs from root specific, temporal, lateral rooting and profuse root libraries and validated their expression in roots by RT-PCR analysis. For this study, we selected SlHSP90.2 from root specific library to examine its role in root growth and development (Kalidhasan et al. 2015). In silico analysis of 1.0 kb genomic sequence upstream to the transcriptional start site of SlHSP90.2 genes using PlantCARE database revealed the presence of root specific cis-acting regulatory element TGCACG (data not shown). The identified cis-element is conserved with the promoters of other genes exhibiting root-preferred expression like, AtEXPA7, AtLRX1, TaEXPA7, ZmEXPA7 (Kim et al. 2006).
We, subsequently, compared the expression of SlHSP90.2 in Pusa Ruby in 8- and 40-day-old root and shoot cDNA samples by qRT-PCR. The 8- and 40-day-old stages were selected because we wanted to study the involvement of SlHSP90.2 in temporal regulation of root development, i.e. early as well as late phases of root development. It was observed that SlHSP90.2 had ~ fivefold and ~ threefold higher expression in 8-day-old and 40- day-old roots, respectively, as compared to their shoot counterparts (Fig. 1). This substantiated our previous observation where we validated its expression in roots by semi quantitative RT-PCR analysis (Kalidhasan et al. 2015). Besides, the data is also concurrent with other published reports showing the overexpression of HSP90.2 in roots of Arabidopsis, rice and tobacco (Liu et al. 2006; Yabe et al. 1994).
Fig. 1.
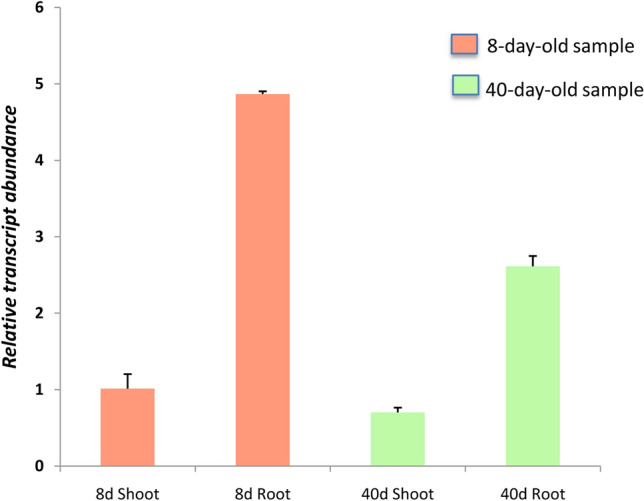
SlHSP90.2 shows higher expression in roots than shoots. Relative expression of SlHSP90.2 in 8- and 40-day-old root and shoot tissues of Pusa Ruby tomato plants were analyzed by qRT PCR analysis. The data presented are means ± SD from three independent biological replicates each with three technical replicates. 8-day-old shoot sample was used as the reference and the data was normalized against SlACTIN
Phenotypic analysis of tomato cultivars for root development
To assess the involvement of SlHSP90.2 gene expression in root development, we first compared the root phenotype of eight tomato cultivars obtained from IIHR, Bengaluru at 40-day-stage. Among the eight cultivars, Arka Meghali and IIHR-2624 showed longest roots, while IIHR-2615 and IIHR-2613 were found to have the shortest roots. Therefore, we selected Arka Meghali and IIHR-2624 as representatives of high rooting cultivars and IIHR-2615 and IIHR-2613 as representatives of low rooting cultivars. Pusa Ruby was intermediate between the differential rooting cultivars and hence, was also selected.
For detailed analysis, we compared the root system architecture (RSA) of the selected cultivars at different developmental stages (seedling, vegetative and mature stage). None of the selected cultivars showed any developmental aberrations at seedling stage (8 days after germination). However, Arka Meghali and IIHR-2624 showed significantly higher root biomass as compared to Pusa Ruby, IIHR-2615 and IIHR-2613, both at vegetative (40-day-old) and mature stages (90-day-old) (Fig. 2a–f).
Fig. 2.

A comparative study of attributes of Arka Meghali, IIHR-2624, Pusa Ruby, IIHR-2615 and IIHR-2613 tomato cultivars at 40- and 90-day-old stages. a A snapshot of the RSA of the different genotypes at 40-day-old stage. b A representation of RSA of different rooting cultivars of tomato at 90-day-old stage. c fresh root weight of different tomato cultivars at 40-day-old stage. d Fresh root weight of different tomato cultivars at 90-day-old stage. e Quantification of root length of the different genotypes at 40-day-old stage. The data represented mean ± SD of three samples per cultivar per growth stage
SlHSP90.2 is differentially expressed in cultivars with differing root phenotypes
To further validate SlHSP90.2 link with root biomass development, the expression of SlHSP90.2 was examined in these tomato cultivars with differing root phenotypes. Interestingly, SlHSP90.2 was up- regulated in the tomato cultivars showing high root biomass i.e. Arka Meghali and IIHR-2624 and was significantly down-regulated in low rooting cultivars, IIHR-2615 and IIHR-2613, as compared to Pusa ruby (Fig. 3a&b). These results highlight the involvement of SlHSP90.2 gene in tomato root development. The significantly high expression of SlHSP90.2 in high-rooting cultivars compared to low-rooting cultivars (Fig. 2) offers a strong piece of evidence suggesting a direct correlation of the expression of the gene with tomato root development. The adaptation of root system architecture would require transcriptional regulation of gene(s) involved in key root developmental processes, thereby validating the differential expression of SlHSP90.2 in the different tomato cultivars.
Fig. 3.
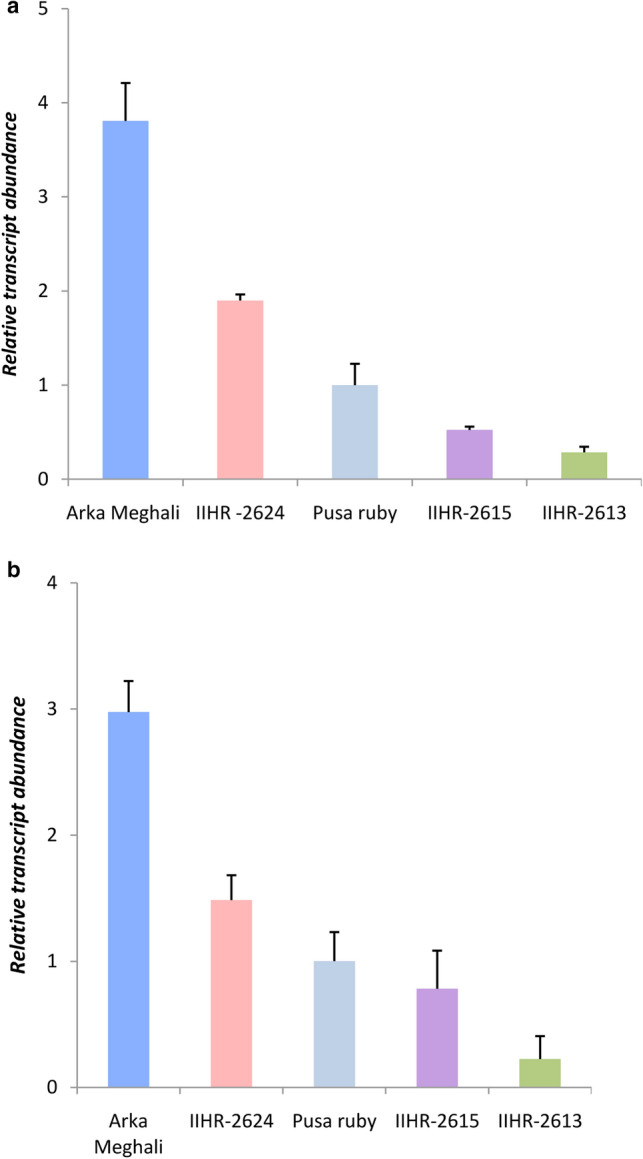
SlHSP90.2 is differentially expressed in tomato cultivars with different root phenotypes. Relative expression of SlHSP90.2 were analyzed in a 40-day-old and b 90-day-old root samples of the selected cultivars of tomato using qRT PCR analysis. The data presented are means ± SD from three independent biological replicates each with three technical replicates
Generation and analysis of transgenic tomato plants overexpressing SlHSP90.2
To functionally characterize the role of SlHSP90.2 in tomato root development, we generated transgenic lines in Pusa Ruby background. This was done by introducing 35S::SlHSP90.2 construct in Pusa Ruby cultivar of tomato (Fig. 4a) using Agrobacterium-mediated plant transformation. An additional transgenic line was generated using empty vector as a control. The transgenic lines including the control lines were screened by PCR using genomic DNA of tomato leaves as template and gene specific primers (data not shown). Further, all the three transgenic lines and control (empty vector) showed characteristic blue colour indicative of GUS activity (Fig. 4b). Significantly higher levels of SlHSP90.2 transcripts in all the three transgenic lines (L1, L2 and L3) tested as compared to controls (Pusa Ruby wild-type and transgenic line containing empty vector) (Fig. 4c) confirmed the integration and expression of the transgene.
Fig. 4.

Transgenic lines overexpressing SlHSP90.2 expresses more SlHSP90.2 than control plants. a A schematic representation of pCAMBIA2301 transformation vector showing SlHSP90.2 and GUS (β-glucuronidase) reporter genes under the control of 35S promoter (35SP) and NosA terminator. b Analysis of GUS expression in the roots of three independent transgenic lines. Untransformed Pusa Ruby and an empty vector transformed transgenic line were used as controls. c Relative expression of SlHSP90.2 in the 35S::SlHSP90.2 transgenic lines was analyzed using qRT-PCR. The data indicate means ± SD from three independent biological replicates each with three technical replicates. Normalization was done using SlACTIN
SlHSP90.2 transgenic lines, showed better lateral roots at seedling stage
To understand the involvement of SlHSP90.2 in tomato lateral root formation, we analyzed root morphology of transgenic plants overexpressing SlHSP90.2 and compared it with the control plants at 15-day stage. The overexpressing lines showed enhanced primary root length, number of lateral roots and lateral root length when compared to control. Apparently, transgenic lines had 35–38% more lateral roots and length as compared to controls (Fig. 5a–d). The results indicated that overexpression SlHSP90.2 had a positive effect on the development of root architecture in tomato.
Fig. 5.

SlHSP90.2 transgenic lines showed increased primary root length and enhanced lateral root length and number. a A snapshot of 35S::SlHSP90.2 seedlings at 15-day-old stage. 35S::SlHSP90.2 transgenic lines showed b increased primary root length, c more number of lateral roots per plant and d increased lateral root length as compared to the control plants. The data represented means ± SD of three samples
SlHSP90.2 transgenic lines showed enhanced root biomass
Having observed the involvement of SlHSP90.2 in lateral root formation of tomato, we further analyzed the roots of transgenic lines harboring SlHSP90.2 for differences in root biomass. For this, we investigated the shoot length, primary root length, fresh and dry weight of shoot and root at the 35-day-old stage. Compared to control, 35S::SlHSP90.2 transgenic lines showed improved phenotypes, with significant increase in primary roots length (Fig. 6a–e) and root dry weight (Fig. 6f&g). Moreover, higher expression of SlHSP90.2 gene in the transgenic lines significantly increased the root length and biomass as compared to Arka Meghali, a high rooting cultivar (Table 2). This is concurrent with rice rHSP90 gene as the overexpression of rHSP90 also showed more elongated roots as compared to wild type (Liu et al. 2020) indicating a direct involvement of HSP90 gene in root development.
Fig. 6.

SlHSP90.2 transgenic lines showed better shoot growth and enhanced root architecture than the control plants. a A photograph of three independent transgenic lines of 35S::SlHSP90.2 and controls at 35-day-old stage. SlHSP90.2 transgenic lines showed increased, b shoot length, c fresh shoot weight, d dry shoot weight, e primary root length, f fresh root weight and g dry root weight as compared to the control plants. The data represented means ± SD of three samples
Table 2.
Relative Fold change in different root growth parameters in Arka Meghali and Transgenic lines in comparison to Pusa Ruby
| Plant | Growth parameter | ||
|---|---|---|---|
| Root length (fold change) | Root biomass (fold change) | Real time expressionof SlHSP90.2 (fold change) | |
| ArkaMeghali/Pusa Ruby | 0.45 | 1.44 | 2.8 |
| Transgenics/Pusa Ruby | 2.03 | 1.97 | 4.67 |
Tolerance of transgenic lines to excess salinity and drought
Increased root biomass is often associated with an enhanced tolerance to salinity and drought. For example, the overexpression of rHSP90 in tobacco has shown more tolerance when exposed to 200 mM NaCl and the roots of the transgenics grew faster than the wild-type (Liu et al. 2006). This prompted us to check the salinity response of SlHSP90.2 transgenic lines. We tested the transgenic lines for their salinity tolerance in different concentrations of NaCl using leaf disc assay for 72 h and estimated chlorophyll retention using Porra et al. 2002. The leaf segments of transgenic lines could retain ~ 89, ~ 84, ~ 78, ~ 73% of chlorophyll at 200, 400, 600 and 800 mM NaCl, respectively, in contrast to the leaf segments of control plants, which could retain only ~ 79, ~ 55, ~ 29 and 18% of chlorophyll at the different NaCl concentrations (Fig. 7a&b). This confirmed that salinity-induced loss of chlorophyll was lower in SlHSP90.2 overexpressing lines compared to control plants, thereby further aligning with the rHSP90 gene.
Fig. 7.

SlHSP90.2 transgenic lines showed increased salt tolerance as compared to control plants. a The leaf pieces from control plants and SlHSP90.2 transgenic plants were floated on 0 (control), 200, 400, 600 and 800 mM NaCl salt solutions. b Quantitative analysis of % chlorophyll retention in the different genotypes subjected to salt stress at the end of 72 h on the different NaCl concentrations tested. The error bars in the graph represent the standard deviation of the values taken from all the three transgenic lines
In another set of experiment, the drought stress response in SlHSP90.2 transgenic lines were also tested For this, seeds of wild-type and transgenic lines were germinated in presence of 200 mM mannitol to mimic drought stress. The transgenic lines showed (i) more (90%) seed germination (Fig. 8a, b) and (ii) increased fresh weight and root length (Fig. 8c&d) as compared to control at the 8th day of germination in mannitol. We further tested the post-stress recovery response in transgenic lines. For this, seedlings were first grown in presence of mannitol for 8 days and subsequently, they were washed and transferred to water for another 8 days. The recovery response was examined by measuring fresh and dry root weight and root length of the seedlings. The transgenic lines recovered better as they showed more fresh and dry root weight and more root length as compared to control (Fig. 9a–c). Taken together, the data strongly suggested that SlHSP90.2 play a vital role in improving the salinity and drought tolerance of tomato plants by way of enhancing the root biomass and architecture and also helps in the recovery process after stress conditions are reversed.
Fig. 8.

SlHSP90.2 transgenic lines are more tolerant to drought stress. Seeds of control plants and SlHSP90.2 transgenic lines were examined for germination on 200 mM mannitol and subsequently their fresh weight and root lengths were quantified at 8-day stage. a Photograph of representative seedlings of Pusa Ruby control and tomato transgenic lines taken after 8-days of germination at 200 mM mannitol. b Seeds were germinated on 200 mM mannitol and percentage germination was calculated on each day upto the 8th day. c Fresh weight and d root length of 8-day-old seedlings germinated on mannitol. Data presented are means ± SD of five seedlings
Fig. 9.

The recovery response of transgenic plants after drought stress. The 8-day-old seedlings of Pusa Ruby control and SlHSP90.2 transgenic lines obtained after germination on 200 mM mannitol (in Fig. 8) were allowed to recover by placing them in water. a Photograph of seedlings representing control and tomato transgenic lines after 8 days of recovery process. b Quantification of root length and c fresh and dry weight after recovery period. Data presented are means ± SD of five seedlings
Conclusion
In conclusion, it may be suggested that (i) the expression of SlHSP90.2 has a direct relationship with root phenotypes in different genotypes, i.e. higher expression in cultivars with greater root biomass, (ii) overexpression of SlHSP90.2 in transgenic lines showed an increase in lateral root number, primary root length and root biomass and (iii) increased root biomass resulted in enhanced tolerance to salinity and drought stresses. The study opens up avenues for developing climate resilient cultivars of tomato with better root biomass and architecture and also offers a strategy for improving agronomic trait in other crop plants.
Authors’ contributions
AKG and VV conceived and designed the experiments. PKY performed majority of the experiments and supported by NG. PKY, NG, VV and AKG analyzed the data. PKY, VV and AKG wrote the manuscript. All authors read and approved the manuscript.
Funding
The research work was supported by a Department of Biotechnology, Government of India grant BT/PR6959/PBD/16/1010/2012. PKY was supported by a research scholarship from Central University of Rajasthan, Rajasthan, India. NG was a D.S. Kothari fellow. VV was supported by Ramalingaswami Re-entry fellowship program under Department of Biotechnology, Government of India (BT/RLF/Re-entry/22/2017). We also thank Indian Institute of Horticultural Research, Bengaluru, India for providing the seeds of tomato cultivars.
Compliance with ethical standards
Conflict of interest
The authors declare no conflict of interest.
Availability of data and material
All the data supporting the findings are available with the corresponding author and can be provided upon request.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Kalidhasan N, Joshi D, Bhatt TK, Gupta AK. Identification of key genes involved in root development of tomato using expressed sequence tag analysis. Physiol Mol Biol Plants. 2015;21:491–503. doi: 10.1007/s12298-015-0304-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim DW, Lee SH, Choi SB, Won SK, Heo YK. Functional conservation of a root hair cell-specific cis-element in angiosperms with different root hair distribution patterns. Plant Cell. 2006;18:2958–2970. doi: 10.1105/tpc.106.045229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krishna P, Gloor G. The Hsp90 family of proteins in Arabidopsis thaliana. Cell Stress Chaperones. 2001;6(3):238–246. doi: 10.1379/1466-1268(2001)006<0238:thfopi>2.0.co;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li W, Chen Y, Ye M, Wang D, Chen Q. Evolutionary history of the heat shock protein 90 (Hsp90) family of 43 plants and characterization of Hsp90s in Solanum tuberosum. Mol Biol Rep. 2020;47:6679–6691. doi: 10.1007/s11033-020-05722-x. [DOI] [PubMed] [Google Scholar]
- Liu D, Zhang X, Cheng Y, Takano T, Liu S. rHsp90 gene expression in response to several environmental stresses in rice (Oryza sativa L.) Plant Physiol Biochem. 2006;44:380–386. doi: 10.1016/j.plaphy.2006.06.011. [DOI] [PubMed] [Google Scholar]
- Liu M, Wang J, Gou J, Wang X, Li Z, Yang X, Sun S. Overexpression of NtSnRK2.2 enhances salt tolerance in Nicotiana tabacum by regulating carbohydrate metabolism and lateral root development. Funct Plant Biol. 2020;47:537–543. doi: 10.1071/FP19299. [DOI] [PubMed] [Google Scholar]
- Milioni D, Hatzopoulos P. Genomic organization of hsp90 gene family in Arabidopsis. Plant Mol Biol. 1997;35:955–961. doi: 10.1023/A:1005874521528. [DOI] [PubMed] [Google Scholar]
- Porra RJ. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynth Res. 2002;43:149–156. doi: 10.1023/A:1020470224740. [DOI] [PubMed] [Google Scholar]
- Prasinos C, Krampis K, Samakovli D, Hatzopoulos P. Tight regulation of expression of two Arabidopsis cytosolic Hsp90 genes during embryo development. J Exp Bot. 2005;6:633–644. doi: 10.1093/jxb/eri035. [DOI] [PubMed] [Google Scholar]
- Queitsch C, Sangster TA, Lindquist S. Hsp90 as a capacitor of phenotypic variation. Nat. 2002;417:618–624. doi: 10.1038/nature749. [DOI] [PubMed] [Google Scholar]
- Riggs DL, Cox MB, Cheung-Flynn J, Prapapanich V, Carrigan PE. Functional specificity of co-chaperone interactions with Hsp90 client proteins. Crit Rev Biochem Mol. 2004;39:279–295. doi: 10.1080/10409230490892513. [DOI] [PubMed] [Google Scholar]
- Sangster TA, Salathia N, Undurraga S, Milo R, Schellenberg K. HSP90 affects the expression of genetic variation and developmental stability in quantitative traits. Proc Natl Acad Sci. 2008;105:2963–2968. doi: 10.1073/pnas.0712200105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma MK, Solanke AU, Jani D, Singh Y, Sharma AK. A simple and efficient Agrobacterium-mediated procedure for transformation of tomato. J Biosci. 2009;34:423–433. doi: 10.1007/s12038-009-0049-8. [DOI] [PubMed] [Google Scholar]
- Song Z, Pan F, Yang C, Jia H, Jiang H. Genome-wide identification and expression analysis of HSP90 gene family in Nicotiana tabacum. BMC Genet. 2019;20:1–12. doi: 10.1186/s12863-019-0738-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tichá T, Samakovli D, Kuchařová A, Vavrdová T, Šamaj J. Multifaceted roles of HEAT SHOCK PROTEIN 90 molecular chaperones in plant development. J Exp Bot. 2020;71:3966–3985. doi: 10.1093/jxb/eraa177. [DOI] [PubMed] [Google Scholar]
- Wang GF, Wei X, Fan R, Zhou H, Wang X. Molecular analysis of common wheat genes encoding three types of cytosolic heat shock protein 90 (Hsp90): functional involvement of cytosolic Hsp90s in the control of wheat seedling growth and disease resistance. New Phytol. 2011;191:418–431. doi: 10.1111/j.1469-8137.2011.03715.x. [DOI] [PubMed] [Google Scholar]
- Wang Y, Lin S, Song Q, Li K, Tao H. Genome-wide identification of heat shock proteins (Hsps) and Hsp interactors in rice: Hsp70s as a case study. BMC Genom. 2014;15:315–344. doi: 10.1186/1471-2164-15-315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu J, Xue C, Xue D, Zhao J, Gai J. Overexpression of GmHsp90s, a heat shock protein 90 (Hsp90) gene family cloning from soybean, decrease damage of abiotic stresses in Arabidopsis thaliana. PLoS ONE. 2013;8:69810. doi: 10.1371/journal.pone.0069810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yabe N, Takahashi T, Komeda Y. Analysis of tissue-specific expression of Arabidopsis thaliana HSP90-family gene HSP81. Plant Cell Physiol. 1994;35:1207–1219. doi: 10.1093/oxfordjournals.pcp.a078715. [DOI] [PubMed] [Google Scholar]
- Young JC, Moarefi I, Hartl FU. Hsp90: a specialized but essential protein-folding tool. J Cell Biol. 2001;154:267–274. doi: 10.1083/jcb.200104079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zabinsky RA, Mason GA, Queitsch C, Jarosz DF. It’s not magic–Hsp90 and its effects on genetic and epigenetic variation. Semin Cell Dev Biol. 2019;88:21–35. doi: 10.1016/j.semcdb.2018.05.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zai WS, Miao LX, Xiong ZL, Zhang HL, Ma YR. Comprehensive identification and expression analysis of Hsp90s gene family in Solanum lycopersicum. Genet Mol Biol. 2015;14:7811–7820. doi: 10.4238/2015.July.14.7. [DOI] [PubMed] [Google Scholar]