

Abstract
Viral infections lead to the death of more than a million people each year around the world, both directly and indirectly. Viruses interfere with many cell functions, particularly critical pathways for cell death, by affecting various intracellular mediators. MicroRNAs (miRNAs) are a major example of these mediators because they are involved in many (if not most) cellular mechanisms. Virus-regulated miRNAs have been implicated in three cell death pathways, namely, apoptosis, autophagy, and anoikis. Several molecules (e.g., BECN1 and B cell lymphoma 2 [BCL2] family members) are involved in both apoptosis and autophagy, while activation of anoikis leads to cell death similar to apoptosis. These mechanistic similarities suggest that common regulators, including some miRNAs (e.g., miR-21 and miR-192), are involved in different cell death pathways. Because the balance between cell proliferation and cell death is pivotal to the homeostasis of the human body, miRNAs that regulate cell death pathways have drawn much attention from researchers. miR-21 is regulated by several viruses and can affect both apoptosis and anoikis via modulating various targets, such as PDCD4, PTEN, interleukin (IL)-12, Maspin, and Fas-L. miR-34 can be downregulated by viral infection and has different effects on apoptosis, depending on the type of virus and/or host cell. The present review summarizes the existing knowledge on virus-regulated miRNAs involved in the modulation of cell death pathways. Understanding the mechanisms for virus-mediated regulation of cell death pathways could provide valuable information to improve the diagnosis and treatment of many viral diseases.
Keywords: cell death, apoptosis, autophagy, anoikis, viruses, microRNAs
Graphical Abstract
The miRNAs that are involved in the modulation of cell death pathways after virus infection show a degree of complexity and a variety of functions. Hence, these molecules can be used for further development of diagnostic and therapeutic techniques.
Introduction
Virus infections have long posed a threat to human public health. Although virus-related deaths have sharply declined during the past century,1 the death toll is still relatively high. According to the World Health Organization (WHO), respiratory diseases linked to seasonal influenza lead to 650,000 deaths each year. Most recently, the ongoing pandemic of coronavirus disease 2019 (Covid-19) has officially been responsible for 1.6 million deaths so far (December 12, 2020). Additionally, viruses are indirectly involved in other lethal diseases; for instance, 15% of human cancer deaths are estimated to be associated with a viral infection.2
Over the whole range of viruses, from small RNA viruses (e.g., dengue virus [DENV]) to large DNA viruses (e.g., poxviruses), viruses uniquely manipulate the host cell machinery in order to facilitate the pathogenic process.3 Cell death pathways in particular have been shown to be strongly affected by viral infections. Because a marked imbalance between the rate of cell proliferation and cell death is associated with serious diseases such as cancer,4 dysregulation of cell death pathways, whether caused by a virus or other factors, is of great importance. Several factors associated with modulation of cell death pathways (e.g., autophagy-related genes5 and survivin6) have been shown to be dysregulated in virus-infected cells. One of the major molecular controllers of cell death is microRNAs (miRNAs), which can be affected by viral infections.
miRNAs are small non-coding RNAs that recognize and bind to the 3′ untranslated region (UTR) of messenger RNAs (mRNAs) that code for cellular proteins, in order to block the translation of the target protein or, to a lesser extent, degrade the mRNA.7, 8, 9, 10 miRNAs are key modulators of many fundamental cellular processes, including cellular metabolism, proliferation, and cell death in a variety of cell types, such as stem cells, immune cells, cancer cells, and especially virus-infected cells.7,11, 12, 13, 14 Hence, the virus-modulated expression of miRNAs that can result in the regulation of cell death pathways may explain the pathogenesis and lead to the possible treatment of viral diseases.
Viruses can affect cell death pathways by the deregulation of miRNAs, leading to interference with apoptotic and autophagic mechanisms, as shown in Figures 1 and 2. In the present review we summarize the effects of viruses on various miRNAs involved in the regulation of different cell death pathways, including apoptosis, autophagy, and anoikis.
Figure 1.

Indirect effects of viruses on the apoptotic pathway through deregulation of cellular miRNAs
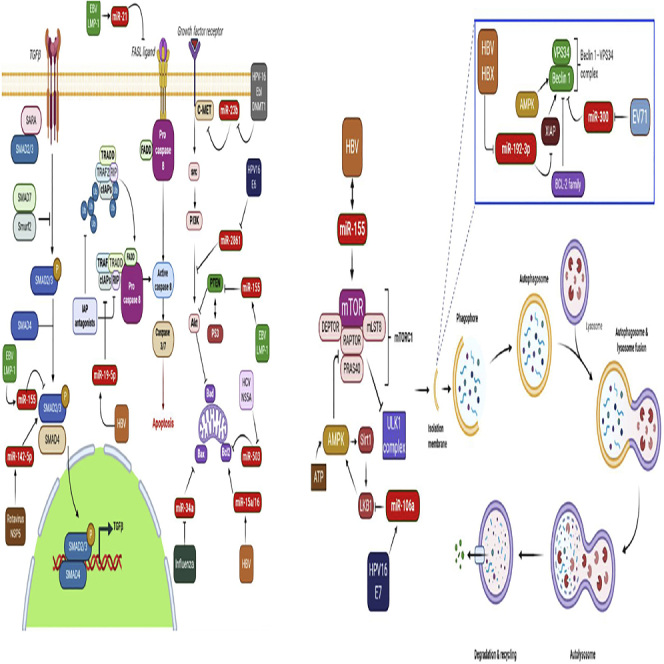
miR-2861 increased the apoptosis of cervical cancer cells through the PI3K/AKT pathway by targeting EGFR. HPV16 E6 is able to downregulate the miR-2861 expression level and can contribute to tumor development. Moreover, HPV-16 E6/DNMT1 can suppress miR-23b expression and contribute to apoptosis resistance through the blocking inhibitory effect of miR-23b on c-MET. EBV-LMP-1 protects B cell lymphoma from rituximab-induced apoptosis via miR-155-mediated Akt activation and upregulation of Mcl-1. LMP1 can increase miR-21 to promote the resistance of nasopharyngeal carcinoma cells to cisplatin-induced apoptosis by suppressing PDCD4 and Fas-L. HBV infection can also lead to the induction of apoptosis via the upregulation of miR-194-5p expression in vitro. miR-194-5p-mediated suppression of cFLIP expression strongly sensitizes HepG2 cells to undergo apoptosis in response to a physiological stimulus. The transforming growth factor β (TGF-β) signaling pathway, a major intercellular signaling pathway in mammalian cells, plays a key role in regulating many cellular processes, such as cell proliferation, differentiation, and apoptosis. TGF-β signaling is known to depend on the formation of Smad2/3-Smad4 transcription regulatory complexes. EBV LMP-2A inhibits the expression level of Smad2 through regulating miR-155-5p in gastric cancer cell lines and inhibits apoptosis. Rotavirus infection leads to up-regulation of TGF-β, which may lead to early apoptosis thus preventing virus progression. However, rotavirus counteracts this by upregulation of miR-142-5p. Rotavirus NSP5 upregulates miR-142-5p, which targets several components of the TGF-β signaling pathway. The mitochondrial pathway of apoptosis is dependent on the BCL-2 (B cell CLL/lymphoma 2) family of proteins for the efficient release of pro-apoptotic factors from the mitochondrial intermembrane space. The BCL-2 family is divided into three groups based on their primary function: (1) anti-apoptotic proteins (BCL-2, BCL-XL, BCL-W, MCL-1, BFL-1/A1); (2) pro-apoptotic pore formers (BAX, BAK, BOK); and (3) pro-apoptotic BH3-only proteins (e.g., BAD, BID, BIK, BIM). miR-503 and miR-15a/16 can target the 3′ UTR of Bcl-2 and inhibit hepatocellular carcinoma cell growth. However, HCV NS5A and HBV mRNA decrease miR-503 and miR-15a/16 expression, respectively, and increase Bcl-2 expression, which leads to a decrease in apoptosis.
Figure 2.

Indirect effects of viruses on the autophagy pathway through deregulation of cellular miRNAs
Autophagy involves the spatially and temporarily coordinated activation of multiple molecular components, including the ULK1 (UNC-51-like kinase 1), FIP200 (FAK family kinase-interacting protein of 200 kDa), and ATG13–ATG101 complex, which is functionally coupled to the negative autophagy regulator, mTOR complex 1, and initiates autophagy. The lipid kinase vacuolar protein sorting 34 (VPS34)-Beclin-1 complex is usually inactivated by anti-apoptotic proteins from the BCL-2 family, but when it is activated it drives the nucleation of the isolation membrane.15 The AMP-activated protein kinase (AMPK)-mammalian target of rapamycin (mTOR) signaling pathway is well known to be associated with autophagy, and AMPK can promote the initiation of autophagy. Activation of AMPK can result in the inhibition of mTOR, which is commonly activated in malignant cells.16 miR-106a is upregulated by the HPV-16 E7 oncogene and stimulates cell proliferation and suppresses autophagy by targeting LKB1 via the AMPK-mTOR pathway in HPV-16-associated cervical cancer. Autophagy contributes to HBV replication, as confirmed by observations that autophagy inhibition strongly impaired replication of HBV in liver cells. The expression level of miR-155 was upregulated in HBV-infected cells, and miR-155 reinforced HBV replication by affecting the SOCS1/Akt/mTOR-autophagy axis. HBV also promoted autophagy through the miR-192-3p-XIAP axis, which is important for HBV replication in vitro and in vivo. Beclin-1 is a key autophagy-promoting gene in the early phase of autophagosome formation. It has been shown that EV71 infection induced autophagosome formation through the reduction of cellular miR-30a, which led to the inhibition of Beclin-1.
Apoptosis
The apoptosis pathways differ between normal cells and cancer cells and even between different cancer cell lines, making the design of therapeutic approaches based on apoptosis induction somewhat challenging.17,18 Therefore, these studies, which were fully focused on apoptosis, have been recently expanded to cover other cell death pathways, i.e., autophagy and anoikis.19 Exploring the cell death pathways and their mechanistic regulation may allow advances to be made in the diagnosis and treatment of several diseases.
Derived from the Greek root “apo” plus “ptosis,” meaning “falling off” (such as leaves falling off a tree),20 the term apoptosis refers to programmed cell death type I that has been long regarded as the main mechanism of controlled cell death.21 Apoptotic cell death is characterized by certain morphological features and some enzymatic processes occurring within the cell, which are designed to allow the removal of unneeded or defective cells without any significant risks for the surrounding tissues.20
Researchers have divided apoptotic pathways into two broad classes, the intrinsic (mitochondrial) pathway and the extrinsic pathway (i.e., death receptor and the perforin/granzyme pathways) (Figure 3). Therefore, the extrinsic signaling pathway involves the tumor necrosis factor (TNF) receptor gene superfamily, and the intrinsic signaling pathway involves many components that can have a negative or positive effect on apoptosis. Apoptosis can be triggered by negative signals arising from the lack of certain growth factors, cytokines, and hormones. The mitochondrial membrane is influenced by such changes and thus the mitochondrial apoptosis pathway is activated. The control and regulation of mitochondrial apoptosis requires a balance between anti-apoptotic as well as pro-apoptotic members of the B cell lymphoma 2 (Bcl-2) protein family. Additionally, the inhibitor of apoptosis (IAP) family consisting of baculoviral IAP repeat-containing (BIRC) proteins has been found to be a major regulator of apoptosis, because these proteins affect both the intrinsic and extrinsic pathways.22
Figure 3.

Comparison of apoptosis and anoikis
The lack of ECM contact or the engagement with inappropriate ECM leads to the activation of anoikis either by death receptors (extrinsic pathway) or mitochondria (intrinsic pathway).
Both of these initial pathways (intrinsic and extrinsic) initiate the executioner (secondary) pathway of apoptosis through different, but related, routes. Both initial and executioner pathways of apoptosis are highly dependent on a family of cysteine-aspartic acid proteases called caspases. Caspases are divided into two groups, i.e., initiator or executioner, according to the pathway in which they participate.23,24 After detection of cell damage, procaspases are converted into the active initiator caspases (caspase-8 from the extrinsic pathway, and caspase-9 from the intrinsic pathway), which, in turn, activate the executioner caspases (caspase-3, -6, and -7). Once activated, the executioner caspases embark on a consecutive series of steps that culminate in DNA fragmentation. These steps include endonuclease activation, degradation of the nuclear proteins and cytoskeleton, protein cross-linking, the expression of ligands designed to attract phagocytes, and the formation of apoptotic bodies.24,25 The apoptotic bodies, which contain the potentially pathogenic remnants of cellular contents, will be further removed mainly by macrophages.20
Normal levels of apoptosis are required not only for cellular and organ homeostasis, but can also inhibit carcinogenesis.26,27 Apoptosis plays a major role in modulating multiple cellular processes such as the appropriate function and development of the immune system, hormone-regulated atrophy, embryonic development, cell recycling, and cell death in response to toxic stimuli.24 Activation of apoptosis irreversibly triggers efficient and rapid cell death through mitochondrial membrane permeabilization and caspase activation.28,29 The downregulation of apoptosis in cancer cells not only increases the proliferation rate, but it prolongs the survival time of the cancer cells through multiple pathways. Moreover, resistance of the malignant cells to apoptosis leads to several consequences, such as oncogene activation, metabolic stress, lack of blood supply, nutrient and growth factor depletion, tolerance to hypoxia, and failure of treatment interventions.28,30 In addition to cancer cells, virus-infected cells such as those infected with hepatitis B virus (HBV)31 or Epstein-Barr virus32 show resistance to apoptosis activation.
Autophagy
Autophagy is mainly a cytoprotective process designed to protect against starvation, in which cytoplasmic components undergo sequestration, autophagosomal transport to lysosomes, and subsequent degradation33, 34, 35 (Figure 4). It is notable that lysosomes have the ability to digest these components, which can then be recycled for the creation of new cellular proteins and new organelles or be additionally processed and used as a source of energy. Moreover, autophagy may be activated by diverse stresses, such as nutrient deprivation (caloric restriction), or may be caused by signals arising in the course of cellular differentiation or embryogenesis, and it can also be triggered by damaged intracellular organelles.36 In addition, a basal level of autophagy exists that results in the physiological turnover of damaged organelles, cytoplasmic contents, as well as longer-lived proteins.37
Figure 4.

Mechanism of the autophagy pathway
A series of stepwise processes, i.e., initiation, nucleation, elongation, maturation, and fusion with lysosomes, are involved in autophagy. Autophagy is induced at a basal level under normal conditions and is further stimulated by stress, for example, by nutrient deprivation
During the initial step of autophagy, a sensor called the mechanistic (previously mammalian) target of rapamycin (mTOR) is activated to form complex I (mTORCI), in response to several autophagy inducers. mTORCI can stimulate autophagy via initiating a signal transduction cascade by phosphorylating selected protein components of the autophagy machinery.38 mTORCI centrally contributes to the modulation of the ULK1-ATG13-FIP200 (FAK family-interacting protein of 200 kDa) complex, as shown by the inhibition of mTORCI with rapamycin increasing the kinase activity of ULK1.39 Researchers have divided phosphatidylinositol 3-kinase (PI3K) enzymes into three different isoforms (class I, II, and III). The mTOR signaling pathway is stimulated by class I PI3K, which also suppresses autophagy, whereas the class III PI3K isoform (Vps34) activates autophagy. There is not enough information about class II PI3K activity and autophagy.40 In addition, mitogen-activated protein kinase (MAPK) signaling may activate autophagy via activation of AMPK and consequent autophagy-associated gene translation. Therefore, PI3K-AKT signaling suppresses autophagy by activating mTOR and suppressing p53. Furthermore, the Vps34 complex combines with Beclin-1 to increase autophagic cell death. However, the binding of ATG5 to ATG12, as well as ATG16L, and phosphatidylethanolamine (PE) to the microtubule-related protein 1 light-chain 3 (LC3) is crucial to initiate and develop autophagy.41 Once initiated, the autophagy process will further progress to carry out its function.
Autophagy is designed either to enhance cell survival in physiologically stressed conditions, or to provoke an alternative programmed cell death pathway. In stressful conditions such as deprivation of growth factors or nutrients, autophagy ensures that cells receive enough vital metabolic substrates, thereby compensating for the metabolic stress imposed on the cells and enhancing cell survival.42, 43, 44, 45 Furthermore, autophagy has been found to be induced in cells that are detached from the extracellular matrix (ECM) in order to avoid anoikis cell death.46 While the main function of autophagy is the recycling of cellular contents, it can also play a role as a cell death pathway, especially to dispose of aging cells and neoplastic cells.36 Unlike apoptotic cell death, autophagy is a reversible mechanism, which can be reduced after elimination of the autophagy-stimulating stress.47 However, both apoptosis and autophagy can function either to regulate normal cell activities such as cellular homeostasis and physiological recycling of cell contents, or else to trigger cell death when necessary.
Anoikis
Originating from a Greek word meaning homelessness, anoikis is another type of cell death induced by the detachment of adherent cells from the ECM (Figure 3).48 It has also been physiologically associated with tissue homeostasis, embryonic development, and the pathogenesis of diseases.49, 50, 51
Anoikis can be considered to be a subset of the apoptosis mechanisms,52, 53, 54 and it can also be categorized into intrinsic and extrinsic pathways.55 As mentioned earlier, the intrinsic pathway requires caspase activation due to permeabilization of the outer mitochondrial membrane via pro-apoptotic protein members of the Bcl-2 family.56 Likewise, the extrinsic pathway involves death receptors like TNF superfamily receptors, and ultimate activation of caspase-8. The two pathways have no biological distinction because the intrinsic pathway may be induced by caspase-8, which is also activated by the extrinsic pathway.57,58
The initiation of anoikis depends on proteins mediating the cellular interaction with the ECM and with neighboring cells, such as E-cadherin and integrins. Integrins are cell membrane receptors consisting of two components, α and β subunits. When integrins are ligated to their ECM receptors, they activate pro-survival signaling pathways, such as the PI3K/Akt pathway.59 Alternatively, integrins trigger anoikis and apoptosis when they adopt an unligated conformation.58 The apoptotic response of integrins is mediated by recruiting caspase-8.60,61 E-cadherin is a transmembrane protein with a cytoplasmic domain that is bound to β-catenin, which then binds to α-catenin. Because α-catenin binds to actin filaments, this complex mediates the binding E-cadherin to the cytoskeleton, and it regulates the adhesion between neighboring cells.62,63 In addition, E-cadherin can trigger anoikis via the RAF/extracellular signal-regulated kinase (ERK) and PI3K/Akt pathways.64,65 The loss of E-cadherin contributes to anoikis evasion in hepatocyte spheroids in the absence of ECM.66 Hence, the loss of E-cadherin and the gain of N-cadherin are the hallmarks of the epithelial-to-mesenchymal transition (EMT). The EMT is characterized by the loss of anoikis, and it allows detached cancer cells to survive long enough to produce metastasis.67,68 As mentioned above, the main factor responsible for initiating anoikis is cell detachment from the surrounding ECM. The appropriate interactions between adjoining cells and between cells and the surrounding matrix are a key factor for the survival and differentiation of epithelial cells. Regarded as a critical point in carcinogenesis,69,70 anchorage-independent growth results from decreased cell-ECM interactions, and in normal cells the resultant integrin signaling triggers anoikis.55 Alternatively, the appropriate anchorage between cells and matrix downregulates anoikis in normal cells.49 However, one characteristic of tumor cells is that they can circumvent anoikis when they become detached from their ECM. Therefore, cancer cells can easily invade the surrounding tissues after becoming detached from their primary location.51,71, 72, 73 Some virus-infected cells have been also proven to be resistant to anoikis-mediated cell death.74,75 For instance, anchorage-independent high-risk human papillomavirus (hrHPV)-transformed cells avoid anoikis, probably by inactivation of tumor suppressor genes.76
miRNAs and cell death pathways
Recently, miRNAs have become popular targets for controlling and treating a variety of diseases. miRNAs have been applied in the clinical settings as diagnostic biomarkers for several diseases, and they are also being investigated in vaccine development.7
Although miRNAs are mostly considered to act as post-transcriptional regulators, their regulatory function has been also described at transcriptional levels. The effects of miRNAs on the target gene mRNA are largely dependent on the degree of complementarity between the target-gene transcriptional sequence and the sequence of the miRNA. There are two possible scenarios. (1) When the miRNA is fully complementary to the target gene mRNA, the function is the same as that of small interfering RNA (siRNA) and ultimately leads to cleavage of the target mRNA. (2) When the miRNA is not perfectly complementary to the target gene mRNA, it can still bind and thus inhibits translation by interfering with the mRNA stability.77
A wide variety of cellular processes have been reported to be regulated by miRNAs, the most important of which is cell death processes (apoptosis, autophagy, anoikis). This has been proposed to be a key mechanism for treating several serious conditions.
Defects in the regulation of apoptosis (either too little or too much) are involved in several human diseases, such as neurodegenerative conditions, ischemic tissue damage, autoimmune disorders, and in numerous types of cancer.24 Slattery et al.22 described the involvement of multiple miRNAs in the process of apoptosis, both in the intrinsic and extrinsic pathways through regulating the expression of specific genes (extrinsic pathway genes CASP7 and BIRC5, and intrinsic pathway genes CSF2RB and BCL2) in colorectal cancer. In another similar study, some apoptosis-associated genes, including BCL2 and PUMA (a Bcl-2 family member), were found to be regulated by various miRNAs, such as miR-148a, miR-125b, miR-143, and miR-203.78, 79, 80, 81 Furthermore, miRNAs have been reported to regulate other targets in apoptosis-related pathways, such as PEG10, BTG1, ID1, interleukin (IL)-32, and NCF2.82
miRNAs can target several autophagy-related complexes, thereby modulating autophagy at various stages, including initiation, vesicle nucleation, elongation, and completion.83 miRNAs can regulate autophagy by targeting autophagy proteins such as beclin (the first autophagy protein to be discovered), or autophagy-related signaling pathways such as AMPK-mTOR.41 For example, miR-30a can inhibit autophagy by downregulating beclin expression, and miR-138 can upregulate the AMPK-mTOR pathway.84,85
Because autophagy and apoptosis share some similar intracellular pathways involved in cellular homeostasis, they also share some upstream mediators, such as specific miRNAs.86,87 Studies have confirmed the ability of certain miRNAs to inhibit both processes. For instance, there is a physical interaction between BECN1 and the anti-apoptotic family members (BCL2, MCL1, and BCL2L1), which is crucial for connecting the two pathways. Under normal conditions, BECN1 and anti-apoptotic BCL2 family members are capable of binding to each other to maintain cellular homeostasis. However, in situations where the cells are subjected to stressful conditions, the complex between BECN1 and the BCL2 family members disassociates, increasing autophagy and suppressing apoptosis.88, 89, 90 Furthermore, MIR30A is capable of reducing the cytoplasmic level of BECN1, while numerous other miRNAs can decrease the expression of anti-apoptotic family members, such as BCL2 and MCL1.91,92 Therefore, miRNAs can have an active involvement in the regulation of autophagy and apoptosis by altering protein-protein interactions. In addition, SQSTM1 (sequestosome-1) has been proposed to be another mediator that is controlled by miRNAs. One study showed the ability of SQSTM1 to modulate the polyubiquitination and aggregation of CASP8 that is crucial for the extrinsic apoptosis pathway.93 Put differently, SQSTM1 is capable of negatively regulating the degradation of autophagic protein LC3 via the 20S proteasome.94 The mechanism underlying the interaction between SQSTM1 and the apoptosis and autophagic pathways is being further investigated.
Various genes simultaneously take part in the autophagy and apoptosis pathways, including factors such as DAPK1 (death-associated protein kinase 1), EEF2 (eukaryotic translation elongation factor 2), and some signal transduction mediators such as TP53 (p53) and ATG5.86,95,96 Some miRNAs can exert a regulatory effect on common targets in both apoptosis and autophagy pathways, thus regulating both of them at the same time. MIR101, for instance, regulates a gene shared between the two pathways, which in turn controls the expression of RAB5A, STMN1, and ATG4D as autophagic proteins and MCL1 as an anti-apoptotic protein.97,98 During hypoxia-reoxygenation endogenous MIR204 promoted the effect of chemotherapy to induce apoptosis via BCL2 downregulation, but it inhibited autophagy through LC3-II protein modulation.99,100
Besides apoptosis and autophagy, anoikis cell death is also affected by various miRNAs. For instance, hsa-miR-34c-3p/5p,101 hsa-miR-203,102 and hsa-mir-129-2/-137/-3663103 were all shown to be involved in modulating anchorage-independent growth and therefore anoikis. It was suggested that methylation-dependent downregulation of tumor suppressive miRNAs played a major role in anchorage-independent growth and in circumventing anoikis.76,103 Once the cancer cells or the virus-infected cells are fully anchorage-independent, they may bypass anoikis and further spread throughout the body, thereby disseminating the disease. Moreover, several miRNAs have been recently found to be directly associated with either the positive or negative regulation of anoikis.104 miRNA 204,105, 106, 107, 108 miR-200c,109, 110, 111 and miR-146a112, 113, 114, 115 can regulate anoikis in cancer cells, whereas some other miRNAs such as miR-10775 and miR-30a116 can perform a similar function in virus-infected cells. Hence, a broad spectrum of cellular mechanisms, especially cell death pathways, have been reported to be regulated by miRNAs in recent decades. In this review, we summarize the cell death-regulatory functions of miRNAs that can be dysregulated in virus infections (Table 1).
Table 1.
Virus-regulated miRNAs known to play roles in modulating cell death pathways
| Virus | Viral protein | miRNA | Expression of miRNA | Target of miRNA | Inhibition/ induction of cell death | Disease | Sample type | Ref. |
|---|---|---|---|---|---|---|---|---|
| HPV 16 | E7 | miR-27b | up | polo-like kinase2 | inhibition of apoptosis | cervical cancer |
in vitro (SiHa and CaSki) | 31 |
| HPV 16 | E7 | miR-21 | up | – | inhibition of apoptosis | cervical cancer |
in vitro (HeLa) | 117 |
| HPV 16 and 18 | E6 | miR-34a | down | – | inhibition of apoptosis | cervical cancer |
human (cervical cancer tissue samples) in vitro (SiHa, HeLa, CaSki) |
118 |
| HPV 16 | E6 | miR-2861 | down | CCND1, EGFR, AKT2 | inhibition of apoptosis | cervical cancer |
human (n = 57 cervical cancer tissue samples) in vitro (SiHa and CaSki) |
119 |
| HPV 16 | E6 | miR-23b | down | c-MET | inhibition of apoptosis | cervical cancer | in vitro (SiHa) | 120 |
| HPV 16 | E5 | miR-196a | down | HoxB8 | inhibition of apoptosis | cervical cancer |
in vitro (HeLa, SiHa) | 121 |
| HPV 16 and 18 | E6, E7 | miR-18a | up | STK4 | inhibition of apoptosis | cervical cancer |
human (cervical cancer tissue samples) in vitro (SiHa, HeLa, CaSki, SW756, C33A) |
122 |
| EBV | EBNA2 | miR-34a | down | PD-L1 | inhibition of apoptosis | Burkitt’s lymphoma | in vitro (Mutu I and Mutu III, Daudi, Jijoye, LCL, OMA4, U2932) | 123 |
| EBV | LMP2A | miR-155-5p | up | Smad2 | inhibition of apoptosis | gastric cancer | in vitro (GT39, SNU719, EBVnGC, HGC27, SGC7901) | 124 |
| EBV | LMP1 | miR-21 | up | PDCD4, Fas-L | inhibition of apoptosis | nasopharyngeal carcinoma | in vitro (C666-1 HONE1, CNE2) | 125 |
| EBV | LMP1 | miR-155 | up | UBQLN1 | inhibition of apoptosis | nasopharyngeal carcinoma | human (n = 8 radio-resistant tissue samples of NPC) in vitro (CNE-1, CNE-2) |
126 |
| EBV | – | miR-155 | up | – | inhibition of apoptosis | B cell lymphoma | in vitro (EF3D, SDLCL) | 32 |
| EBV | LMP1 | miR-155 | up | PTEN | inhibition of apoptosis | B cell lymphoma | in vitro (SKW6.4 rituximab resistance) | 127 |
| EBV | EBNA1 | miR34a | down | – | inhibition of apoptosis | gastric cancer | in vitro (SNU719) | 128 |
| EBV | – | miR-194 | down | IL-10 | inhibition of apoptosis | PTLD | human (n = 6 PBMC samples of PTLD) in vitro |
129 |
| HBV | HBx | miR-21 | up | IL-12 | inhibition of apoptosis | HCC | in vitro (HepG2.2.15) | 130 |
| HBV | HBx | miR-21 | up | PDCD4, PTEN |
inhibition of apoptosis | HCC | in vitro (HepG2, Huh7) | 131 |
| HBV | HBx | miR-181a | up | PTEN | inhibition of apoptosis | HCC | in vitro (HepG2) | 132 |
| HBV | HBx | miR-192-5p | up | BIM | inhibition of apoptosis | HCC | in vitro (HepG2) | 133 |
| HBV | HBx | miR-331-3p | up | ING5 | inhibition of apoptosis | HCC | human (HCC tissue samples) in vitro (HepG2.2.15, SMMC7721) |
134 |
| HBV | HBx | miR-602 | up | RASSF1A | inhibition of apoptosis | HCC | human (n = 21 HCC tissue samples) (HepG2.2.15, HepG2-HBX) |
135 |
| HBV | – | miR-15a | down | Smad7 | inhibition of apoptosis | HCC | human (n = 40 HCC tissue samples) in vitro (HepG2-4D14, HepG2-C5)/in vivo |
136 |
| HBV | – | miR-30e-5p | down | MAP4K4 | inhibition of apoptosis | HCC | human (n = 55 HCC tissue samples) in vitro (HepG2.2.15, Hep3B) |
137 |
| HBV | – | miR-15a/16 Cluster | down | Bcl-2 | inhibition of apoptosis | HCC | human (n = 40 HCC tissue samples) in vitro (HepG2.2.15)/in vivo |
138 |
| HBV | HBx | miR-1236 | down | alpha-fetoprotein (AFP) | inhibition of apoptosis | HCC | human (n = 97 tissue samples of HCC) in vitro/in vivo | 139 |
| HBV | HBx | miR-1270 | down | alpha-fetoprotein (AFP) | inhibition of apoptosis | HCC | human (n = 97 HCC tissue samples) in vitro/in vivo |
139 |
| HBV | – | miR-101-3p | down | Rab5a | inhibition of apoptosis | HCC | in vitro (SMMC-7721, HepG2.2.15) | 140 |
| HBV | HBx | miR-15a and 15b | down | CCND1 | inhibition of apoptosis | HCC | in vitro (SK-HEP-1, Huh7, HepG2) | 141 |
| HBV | HBx | miR-16 | down | CCND1 | inhibition of apoptosis | HCC | in vitro (SK-HEP-1, Huh7, HepG2) | 141 |
| HBV | HBx | let-7 | down | – | inhibition of apoptosis | HCC | human (n = 19 HCC tissue samples) in vitro (HepG2.2.15) |
142 |
| HBV | HBx | miR-193b | down | – | inhibition of apoptosis | HCC | human (HCC tissue samples) in vitro |
143 |
| HBV | HBx | miR-548p | down | HBXIP | inhibition of apoptosis | HCC | human (n = 21 HCC tissue samples) in vitro (Hep3B, HepG2.2.15) |
144 |
| HBV | HBx | miR-375 | down | AEG-1 | inhibition of apoptosis | HCC | in vitro (SMMC-7721, HepG2-HBx) | 145 |
| HBV | HBx | miR-136 | down | AEG-1 | inhibition of apoptosis | HCC | in vitro (SMMC-7721, HepG2-HBx | 145 |
| HBV | HBx | miR-192 | down | – | inhibition of apoptosis | HCC | in vitro (HepG2) | 146 |
| HBV | HBx | miRNA-145 | down | CUL5 | inhibition of apoptosis | HCC | human (n = 25 HCC tissue samples) in vitro (HepG2.2.15) |
147 |
| HBV | HBc, HBx |
miR-328-3p | up | FOXO4 | inhibition of apoptosis | – | in vitro (THLE-2) | 148 |
| HBV | – | miR-98-5p | up | NF-κB | induction of apoptosis | HCC | human (n = 30 HCC tissue samples) in vitro (MHCC97H, Huh7) |
149 |
| HCV | – | miR-155 | up | APC | inhibition of apoptosis | HCC | human (n = 10 HCC tissue samples) in vitro (Huh7)/In vivo |
150 |
| HCV | core | miR-345 | up | p21Waf1/Ci1 | inhibition of apoptosis | – | in vitro (Huh7) | 151 |
| HCV | core | miR-93 | up | – | inhibition of apoptosis | – | in vitro (Huh7) | 151 |
| HCV | core | miR-30c | down | – | inhibition of apoptosis | HCC | human (n = 152 HCC tissue samples) in vitro (HepG2) |
152 |
| HCV | core | miR-203a | down | – | inhibition of apoptosis | HCC | human (n = 152 HCC tissue samples) in vitro (HepG2) |
152 |
| HCV | NS5A | miR-503 | down | Bcl-2 | inhibition of apoptosis | – | in vitro (HepG2) | 153 |
| HCV | – | miR-193b | down | Mcl-1 | inhibition of apoptosis | HCC | in vitro (Hep-394) | 154 |
| HCV | – | miR-181c | down | ATM | inhibition of apoptosis | – | human (n = 8 tissue samples of chronically HCV-infected) in vivo (HepG2) |
155 |
| HTLV-1 | Tax | miR-155 | up | – | inhibition of apoptosis | ATLL | in vitro (MT-2, C5/MJ, ED-40515) | 156 |
| HTLV-1 | Tax | miR-130b | up | TP53INP1 | inhibition of apoptosis | ATLL | human (PBMC samples of ATLL) in vitro (MT-4) | 157 |
| HTLV-1 | – | miR-93 | up | TP53INP1 | inhibition of apoptosis | ATLL | human (PBMC samples of ATLL) in vitro (MT-4) | 157 |
| Influenza A | – | miR-29c | up | BCL2L2 | induction of apoptosis | influenza | in vitro (A549) | 158 |
| GaHV-2 | Meq | miR-21 | up | PDCD4 | inhibition of apoptosis | Marek’s disease, lymphoma | in vitro (MSB-1) | 159 |
| Rotavirus | NSP5 | miR-142-5p | up | SMAD3, TGFβR2 | inhibition of apoptosis | – | in vitro (HT29) | 160 |
| HSV-1 | – | miR-23a | up | IRF1 | inhibition of apoptosis | – | in vitro (HeLa) | 161 |
| HBV | HBx | miR-194-5p | up | cFLIP, SODD | induction of apoptosis | HCC | in vitro (HepG2) | 133 |
| HCV | – | miR-200c | up | FAP1 | induction of apoptosis | hepatic fibrosis | human (n = 10 tissue samples of HCV fibrosis) | 162 |
| Influenza A (H1N1) | – | miR-34a | down | Bax | induction of apoptosis | influenza | human (n = 10 influenza serum samples) in vitro (A549) | 163 |
| Enterovirus 71 | – | let-7b | up | CCND1 | induction of apoptosis | – | in vitro (SH-SY5Y) | 164 |
| West Nile virus | – | Hs_154 | up | ECOP, CTCF | induction of apoptosis | – | In vitro (SK-N-MC and HEK293) | 165 |
| RSV | NS1 | miR-24 | down | – | induction of apoptosis | – | In vitro (A549) | 166 |
| HBV | HBx | miR-192-3p | down | XIAP | induction of autophagy | hepatitis B | human (serum samples of hepatitis B) in vitro (HepG2.2.15) |
167 |
| HBV | HBsAg | miR-155 | up | SOCS1/Akt/mTOR | induction of autophagy | – | in vitro (HepG2, HepG2.2.15) | 168 |
| Enterovirus 71 | – | miR-30a | down | Beclin-1 | induction of autophagy | – | in vitro (Vero, Hep2) | 116 |
| HBV | – | miR-224 | up | Smad4 | inhibition of autophagy | HCC | human (HCC tissue samples) in vivo | 169 |
| HPV 16 | E7 | miR-106a | up | LKB1 | inhibition of autophagy | cervical cancer | human (n = 91 cervical cancer tissue samples) in vitro (SiHa, HeLa, CaSki, HEK293T) |
170 |
| HBV | HBx | miR-7 | up | maspin | inhibition of anoikis | HCC | human (n = 69 HCC tissue samples) in vitro (HepG2x, Hep3Bx, HepG2.2.15) |
75 |
| HBV | HBx | miR-107 | up | maspin | inhibition of anoikis | HCC | human (n = 69 HCC tissue samples) in vitro (HepG2x, Hep3Bx, HepG2) |
75 |
| HBV | HBx | miR-21 | up | maspin | inhibition of anoikis | HCC | human (n = 69 HCC tissue samples) in vitro (HepG2x, Hep3Bx, HepG2) |
75 |
PBMC, peripheral blood mononuclear cell.
RNA-binding proteins in virus-miRNA interactions
RNA-binding proteins are involved in the function of miRNAs in various conditions such as viral infections. For example, flaviviruses, which are single-stranded (ss)-positive RNA viruses, interfere with the miRNA processing machinery by allowing the accumulation of the non-coding (nc) subgenomic flavivirus RNA (sfRNA) that then carries out sequestration of the double-stranded RNA (dsRNA) binding proteins Dicer and AGO2.171 Dicer interacts with the transactivation response RNA-binding protein (TRBP) and the protein activator of protein kinase R (PKR) (PACT), two dsRNA-binding proteins that play roles in the efficiency and specificity of miRNA processing.172
Alternatively, some miRNAs such as miR-101 also downregulate the RNA-binding protein G-rich sequence factor 1 (GRSF1), whose binding to herpes simplex virus 1 (HSV-1) p40 mRNA typically increases its expression, facilitating viral replication. Therefore, induction of miR-101 expression during HSV-1 infection downregulates both ATP5B and GRSF1 expression, consequently attenuating viral replication and preventing lytic cell death.173
It was reported that PCBP2, a member of the heterogeneous nuclear ribonucleoprotein (hnRNP) E family of RNA-binding proteins, can interact with a sequence-specific motif of single-stranded poly(C) polynucleotides and is involved in the regulation of defense responses to viruses and the negative regulation of immune effector cells. PCBP2 could serve as a target gene of miR-HA-3p in H5N1 influenza virus-induced uncontrolled immune responses. In addition to regulating mRNA translation and maintaining mRNA stability, PCBP2 is essential for the prevention of excessive immune responses by serving as a pivotal negative regulator in the mitochondrial antiviral signaling protein (MAVS) signaling pathway.174
Furthermore, the miR-17/92 cluster and miR-20a have been reported to target the p300-cyclic AMP (cAMP)-responsive element binding protein (CREB)-binding protein-associated factor (PCAF) and render the cells less susceptible to HIV-1 infection. It is known that PCAF has a role in tat acetylation, and acetylated tat is transcriptionally more active.81 Alternatively, HIV-1 tends to suppress these miRNAs and upregulate PCAF expression, leading to a possible increase in viral infectivity.175,176
Viruses have had ample time to evolve in ways that prevent the binding of RNA-induced silencing complexes (RISCs) programmed by cellular miRNAs, leading to their destruction. The rapid loss of cognate RISC-binding sites has been observed when rapidly evolving viruses such as HIV-1 are grown in the presence of artificial siRNAs, which function in a similar manner to miRNAs when they bind to perfectly complementary RNA targets.177,178 Moreover, viral gene products that block RISC function are common in plants and insect viruses, where RNAi represents a major innate immune response to viral infection. It remains unclear whether viruses that infect mammals, which do not mount an innate RNAi response to viral infection of somatic cells, commonly encode inhibitors of miRNA biogenesis.179
Therapeutic potential of miRNAs in virus infections
miRNAs are involved in the regulation of almost all physiological processes, such as cell development, differentiation, proliferation, and apoptosis. In this context, miRNAs may have great potential as new biomarkers for diagnosis and in therapeutic approaches for viral infections. To date, more than 1,000 different human miRNAs have been identified, and it is predicted that miRNAs control the expression of approximately 60% of all human genes.180 Recently, new methods and tools have been developed for the detection and quantification of virus-regulated miRNAs. For example, the use of multiplexed qRT-PCR, microarrays, or next-generation sequencing (NGS)-based genome-wide approaches have helped to provide expression profiles of large numbers of miRNAs.181 Moreover, bioinformatics approaches and existing databases have enabled the discovery of thousands of miRNAs in viruses, e.g., Vir-Mir db, BiTargeting, and ViTa.182
The aberrant expression of miRNAs has been linked to the initiation and progression of virus infections. Some miRNAs have been demonstrated to be involved in the pathogenesis of viral diseases, e.g., mir-122 and mir-21 in hepatitis C virus (HCV), and mir-507 and mir-136 in influenza.183
Furthermore, the structure and quantity of miRNAs contained within exosomes secreted from virally infected and non-infected cells are different.184 Therefore, miRNAs could provide a new approach for better clinical decision-making. Moreover, miRNAs carry out potent and specific gene silencing, making them attractive therapeutic tools. To date, most efforts in this setting have been to explore the potential application of miRNA therapeutics for viral infections.185
However, some challenges faced by the use of miRNAs as therapeutic tools include: (1) degradation by nucleases after introduction into biological systems, (2) poor cell membrane penetration, (3) entrapment in endosomes, (4) insufficient binding affinity for complementary sequences, (5) inadequate delivery to desired target tissues, (6) off-target effects and unwanted toxicity, and (7) activation of innate immune responses.186 Despite these limitations, future research could improve the miRNA properties to overcome these challenges. For example, improvements in miRNA delivery are being addressed by chemical modifications and applications of nanotechnology.186
Indirect regulation of the apoptosis pathway in viral infections by cellular miRNAs
A vast array of virus-related miRNAs have been found to be involved in the modulation of cellular apoptosis.
HBV
Several miRNAs are known to participate in the life cycle of HBV infections.187 HBV causes acute and chronic hepatitis in humans, and it has become a serious health issue affecting 240 million individuals worldwide.188,189 Epigenetic modifications occurring in HBV infection increase the expression of miR-1 and miR-449a and in turn increase the expression of the nuclear receptor farnesoid X receptor α (FXRα), which stimulates the core HBV promoter function as well as HBV replication.190,191 Moreover, miR-373 and miR-372 are capable of upregulating HBV gene expression via the nuclear factor (NF)-κI/B axis.192 It was found that the miR-122 levels were negatively correlated with intrahepatic viral load, hepatic necrosis, and inflammation. The depletion of endogenous miR-122 by its antisense inhibitor led to enhanced HBV replication, whereas overexpression of miR-122 by transfection using a mimic or an expression vector inhibited viral production. miR-122 can have an indirect effect on HBV replication via repression of the expression of its target cyclin G1, thereby inhibiting the binding of p53 to cyclin G1 and abrogating the p53-regulated suppression of HBV replication. Therefore, miR-122 can act as a host restriction factor in HBV infection, and its downregulation during HBV infection might contribute to viral persistence. Therefore, the induction and increase of miR-122 expression could provide an effective strategy for treatment of HBV infection and related liver diseases.192 In contrast, miR141 could downregulate HBV replication by targeting the peroxisome proliferator-stimulated receptor-α (PPARA). Hence, miR-141 could suppress HBV replication by reducing HBV promoter activity by downregulating PPARA. PPARA may be a promising host-oriented drug target for the development of novel HBV treatment, which could be mediated by miRNAs.193 miR-130a targets two metabolic modulators, peroxisome proliferator-stimulated receptor-γ and its respective co-activator PGC1-α, thereby preventing replication of HBV.194 miRNAs are important regulators in viral infections, which could provide an effective strategy for treatment of HBV infection and related diseases.
Aquaporins (AQPs) are membrane water transport channels that contribute to water secretion and adsorption in epithelial cells. A number of aquaporin subtypes can transfer other molecules, such as urea and glycerol. At present, there are 13 specific types of AQPs in mammals that are principally divided into three groups:195,196 (1) classical AQPs that are the original water-selective channels, including AQP0, AQP1, AQP2, AQP4, AQP5, AQP6, and AQP8; (2) aquaglyceroporins such as AQP3, AQP7, AQP9, and AQP10 that are capable of transporting glycerol and other small solutes; and (3) non-classical AQPs such as AQP11 and AQP12 that are located inside the cells. Nonetheless, researchers have not yet reached a complete understanding of their selectivity. Currently, the contribution of the AQPs to tumor progression is increasingly being investigated. Earlier investigations showed strong expression of the AQPs in tumor cells from various origins that were proposed to contribute to tumor-related edema, tumor cell migration, proliferation of tumors, as well as tumor angiogenesis.197,198 In addition, the expression of AQPs in various types of tumors is different due to the respective tissue-specific localization. Furthermore, the expression of AQP5 is upregulated in colon cancer tissues.
Zhang et al.199 addressed the role of miR-325-3p and AQP5 in the proliferation and apoptosis of HBV-infected hepatocellular carcinoma (HCC) cells. The results showed that miR-325-3p directly targeted AQP5 and decreased mRNA and the protein level of AQP5 that increased proliferation and inhibited apoptosis in HCC cells. Overexpression of miR-325-3p further inhibited proliferation and induced apoptosis. Introducing miR-325-3p into the cells inhibited growth and induced apoptosis in Huh7-1.3 and HepG2.2.15 cells via a direct decrease of AQP5 expression. Moreover, silencing the AQP5 expression duplicated the pro-apoptotic impact of miR-325-3p overexpression. HBV downregulates miR-325-3p and suppresses AQP5, resulting in inhibition of apoptosis and contributing to tumor progression.199 Therefore, AQPs may be a promising drug target for the development of a novel HBV therapy that uses miRNAs.
Avian leukosis viruses (ALVs)
ALVs are a type of avian retroviruses that can induce tumors in chickens.200 Chicken ALVs are categorized into six subgroups (A–E and J) with regard to the envelope glycoprotein that is responsible for the viral entry and is involved in host virus neutralization.201,202 During recent years, ALV-J has become epidemic in nature and affects commercial egg layers and local breeding flocks in China.203 Moreover, ALV-J causes myeloid leukosis and creates more severe problems in comparison to other virus subgroups.204 Studies have shown that ALV-J and other subgroups are among the leading causes of avian infections205, 206, 207 and can also cause immunosuppression by inhibiting dendritic cells (DCs).208 DCs are a type of professional antigen-presenting cells (APCs) that have multiple subtypes with different functions. DCs are capable of promoting an immune response in the presence of tissue damage signals or pathogens, and also inducing and maintaining immune tolerance.209 Moreover, DCs have been proposed to be the master regulators and initiators of the immune responses that link innate and adaptive immunity.210,211 In addition, their existence is crucial for maintaining immune homeostasis and preventing autoimmune responses to self-antigens.210 However, their function relies on suitable DC maturation that is often modulated by miRNAs.212, 213, 214, 215
The ALV provirus, as a retrovirus, is capable of random integration into the host genome that possibly leads to the deregulation of gene expression, in particular the expression of diverse regulatory factors such as miRNAs.216 Moreover, there is insufficient information about the mechanism of the ALV provirus integration, and ALV integration is frequently random. Nonetheless, a number of investigations have shown the integration of the provirus into specific sites upstream of some oncogenes (e.g., c-erb, c-myc, TERT, EGFR, MET, ZIC1).217,218 Therefore, ALV integration may modulate the expression of some miRNAs by unclear mechanisms. In addition, the process of virus infection, as well as replication, triggers the cells to alter their miRNA expression. Because some miRNAs, such as miR-146a and miR-146b, are responsible for DC apoptosis,219 the immunosuppressive effect of ALV-J on DCs may be attributed to the regulation of miRNAs involved in apoptosis pathways.
Liu et al.220 explored the miRNA-mediated apoptotic effects of ALV infection on cultured DCs in vitro. Based on their findings, ALV-J was shown to be capable of infecting the cells in the initial phases of differentiation, and infection with ALV-J caused apoptosis in the DCs. Moreover, miRNA sequencing of the non-infected and infected DCs showed 122 differentially expressed miRNAs, of which 115 were upregulated following ALV-J infection, and 7 miRNAs showed considerable downregulation. The miRNAs exhibiting the most upregulation were proposed to affect nutrient processing as well as metabolic functions. Therefore, ALV-J infection of the chicken DCs was capable of inducing apoptosis through altered miRNA expression, providing the motivation for additional study of the epigenetic effects of ALV-J-induced immunosuppression. Therefore, aberrant miRNA expression caused by ALV-J infection may interfere with DC differentiation and apoptosis, resulting in defective antigen presentation and suppression of the immune response.220
Chikungunya virus (CHIKV)
CHIKV is a re-emerging infectious disease caused by an alphavirus, resulting in large-scale outbreaks in the islands of the Indian Ocean, Southeast Asia, and currently in the Caribbean region.221, 222, 223 Moreover, CHIKV causes symptoms of debilitating arthritis that can endure from 1 week up to a year.224,225 CHIKV can infect both peripheral tissue as well as the central nervous system (CNS), which is most serious in newborns and children, and even in adults.225 For peripheral infections, CHIKV virus can affect the connective tissue of the joints, fibroblast cells of the dermis, as well as the muscle fascia. In addition, CHIKV induces delayed apoptosis in infected cells, and it uses the apoptotic blebs for spreading the infection to the neighboring cells.226, 227, 228 However, further studies are needed to fully investigate the CHIKV-mediated molecular pathways in fibroblasts.222,224
Sharma et al.229 carried out an investigation on the regulation of miRNAs in mouse fibroblast cells immediately following infection in order to identify the pathways targeted by CHIKV. In the next step, the researchers analyzed the expression of 760 separate miRNAs at 6 h after infection with CHIKV. Bioinformatic analysis was used to identify the signaling pathways that were targeted by the highly altered miRNAs. A singleplex miRNA assay was used to validate the miRNAs, and then western blot analysis was used to confirm the protein targets of the miRNAs. Among the 760 miRNAs studied, it was found that 75 miRNAs were significantly regulated following CHIKV infection, and of these 71 were upregulated in the infected cells. According to the functional pathway and network analysis, the miRNAs that were most altered contributed to apoptosis and signaling pathways, such as Toll-like receptors (TLRs), the PI3K/Akt pathway, mTOR, and JAK-STAT, that were proposed to be involved in the course of CHIKV infection. Finally, topoisomerase Iiβ was one target of the two downregulated miRNAs in CHIKV infection. Several miRNAs have been shown to be involved in processes that occur later after CHIKV infection, such as cellular proliferation, immune response, and apoptosis, which may be useful for treatment of CHIKV infections in the future.229
Influenza A virus (IAV)
IAV is one of the most common enveloped negative strand RNA viruses in the Orthomyxoviridae family. The IAV genome contains eight RNA sequences, with two of them encoding major surface glycoproteins known as neuraminidase (NA) and hemagglutinin (HA). By analysis of its antigenicity, this virus may be divided into 18 HA subtypes (H1–H18) and 11 NA subtypes (N1–N11). However, researchers have found a large number of permutations containing various HA and NA subtypes.230 Based on the virus strain, different IAVs have different pathogenicity for infection. For instance, less pathogenic avian influenza viruses (LPAIVs) result in only mild respiratory symptoms that can be occasionally followed by a gastrointestinal infection. Alternatively, the highly pathogenic avian influenza viruses (HPAIVs) such as H7N9 and H5N1 lead to multiorgan systemic infections, as well as fatal illnesses.231 Overall, IAV infections annually account for nearly 500,000 deaths throughout the world, and they also cause high rates of death in several animal species.232 The key clinical characteristic is the severity of pneumonia that manifests as cough, fever, and difficulties in breathing. Even though some investigations have shown that IAV can affect cell apoptosis in vitro and in vivo,158,233,234 there is not enough information about the respective molecular mechanism(s).
IAV infection is capable of altering miRNA expression in animal models as well as in cultured cells.235, 236, 237, 238 It is known that specific miRNAs are capable of positively or negatively influencing the immune response,239, 240, 241, 242 and some individual miRNAs can control apoptosis induced by virus infection by targeting cellular mRNAs and protein levels.158 Furthermore, changes in miRNA expression following viral infection profoundly influence the innate host response to viral infection. Nevertheless, the effects of miRNAs on cell apoptosis after IAV infection have been less well studied.
In one study, Fan and Wang243 addressed the molecular mechanism of miR-34a in IAV-induced apoptosis. They used a miRNA array to identify the miRNAs involved in IAV infection, and western blotting and a luciferase reporter assay to determine the target gene of miR-34a. After that, they transfected a miR-34a mimic into the IAV-infected A549 cells and examined the changes in target gene expression. According to the results, miR-34a was considerably downregulated in IAV-infected A549 cells, as confirmed by quantitative reverse transcriptase PCR (qRT-PCR) in vivo and in vitro. Afterward, transfection with a miR-34 mimic or an inhibitor in the IAV-infected A549 cells confirmed the effects on apoptosis. It was predicted that miR-34a was complementary to the 3′ UTR of the mRNA for Bax, and western blotting and a luciferase reporter assay confirmed the direct targeting of Bax by miR-34a. Western blotting showed that the upregulation of the Bax protein level was reversed by miR-34a overexpression. They proposed that miRNAs could provide therapeutic targets for influenza virus infections.243
Indirect regulation of the autophagy pathway by cellular miRNAs in viral infections
It has been found that autophagy contributes to the interactions between host cells and viruses. In the course of viral infection, autophagy is capable of performing either a proviral or an antiviral function. Autophagy can act as an anti-viral defense mechanism of the innate immune system, because it can lead to the preferential degradation of the sequestered viral structures within the cells to inhibit virus replication.244, 245, 246 It has been proposed that several viruses, such as poliovirus, coronaviruses, enteroviruses, DENV, and HCV, can all subvert the autophagy mechanism to promote their respective replication.116,247, 248, 249, 250 Autophagy has been found to have a role in the regulation of virus replication, and it also directly affects the generation of some inflammatory cytokines. Disrupting the normal autophagy pathways considerably increases the secretion of the proinflammatory cytokines IL-1β, IL-18, and IL-1α.251, 252, 253 The treatment of human and murine cells with the autophagy inhibitor 3-methyladenine (3-MA) largely suppressed the TLR-dependent secretion of IL-6 and TNF-α.254 Moreover, silencing of ATG7 decreased the generation of TLR-dependent IL-8 in intestinal epithelial cells.255 Autophagy also contributes to the regulation of type I IFN secretion. Recently it was shown that knockdown of either ATG7 or Beclin-1 increased the expression of IFN-α and IFN-β in HCV infected immortalized human hepatocytes (IHHs).256 The innate immune system is affected by autophagy, and how this affects the virus-host cell interaction requires further investigation. Herein, we summarize the existing knowledge about the miRNAs that regulate autophagy in virus-infected cells.
HBV and autophagy
Intracellular pathways, inflammatory cytokines, specific hormones, and other factors are known to be involved in the regulation HBV replication.257, 258, 259, 260 miRNAs have been found to be involved in the intracellular pathways modulating HBV transcription through regulation of transcription factors, nuclear receptors, and other gene products.261 miR-101 was found to repress the HBV replication via targeting the forkhead box O1 (FOXO1) transcription factor.262 miR-449a accelerated HBV replication via targeting CREB5, which in turn induced the expression of FXRα, a transcription factor known to facilitate HBV replication. CREB5 knockdown and overexpression confirmed that it was a negative regulator of HBV replication. Additionally, miR-449a overexpression inhibited proliferation, caused cell cycle arrest, and promoted HCC cell differentiation. The results indicated that epigenetically regulated miR-449a targeted CREB5 to increase FXRα expression, thereby promoting HBV replication and gene expression.190 Moreover, miR-224, miR-138, miR-224, miR-576, and miR-596 directly targeted the pre-genomic RNA (pgRNA) of HBV and inhibited virus replication. It has been proposed that in the early phase of HBV infection, HBx binding at or near to the p65/NF-κB sites within the miR-224 promoter leads to the repression of miR-224 expression and inhibits viral replication. However, inactivation of HBx leaves the miR-224 promoter free to respond to the TNF/LT-NF-κB signaling in HCCs, leading to the formation of highly dysplastic nodules.263 Nevertheless, this hypothesis remains to be confirmed in vivo.
The PI3K/Akt signaling pathway has been proposed to be a key cellular pathway in the regulation of HBV infection.264,265 In addition to the effects of apoptosis discussed in the previous section, autophagy is also involved in the regulation of HBV infection.266,267 IAV, HIV, HCV, and DENV can all take advantage of autophagy to enhance their replication.250,268, 269, 270, 271 Autophagy is thought to be modulated by the PI3K/Akt/mTOR pathway, which could facilitate HBV infection. For instance, autophagy was found to be increased by suppressing mTOR activity in eukaryotic cells.272,273
Zhang et al.274 investigated the role of miR-155 in HBV replication as well as the possible involvement of autophagy in this process. Moreover, downregulation of miR-155 increased HBV infection, while miR-155 transfection increased antigen expression, HBV replication, and progeny secretion in HepG2.2.15 cells. Additionally, miR-155 contributed to the suppressor of the cytokine signaling 1 (SOCS1)/Akt/mTOR axis and reinforced autophagy in HepG2.2.15 cells. Additionally, the autophagy inhibitor (3-MA) abolished the hepatitis B surface antigen (HBsAg) secretion triggered by miR-155. Hence, miR-155 helped HBV replication via reinforcing SOCS1-stimulated autophagy. It was proposed that chronic HBV could be treated or ameliorated by inhibiting miR-155 in the cells, and autophagy inhibitors (3-MA) could also be tested.274
Lin et al.275 examined whether the miR-99 family modulated HBV replication and investigated the underlying molecular mechanism. The expression of the miR-99 family was downregulated in hepatoma cells, in comparison to primary hepatocytes. Transfection of miR-99b, miR-100, and miR-99a enhanced HBV replication, antigen expression, and progeny secretion in hepatoma cells. Nonetheless, the miR-99 family did not influence HBV transcription or the activity of the HBV promoter, suggesting that these miRNAs modulated HBV replication at the post-transcriptional stage. In agreement with bioinformatic analysis, it was found that ectopic expression of the miR-99 family reduced the insulin growth factor (IGF)-1R/Akt/mTOR pathway signaling and suppressed insulin-stimulated activation in hepatoma cells. The miR-99 family increased autophagy via mTOR/ULK1 signaling and increased HBV replication.
Several miR-29 members, such as miR-29a, miR-29b, miR-29c, miR-29d, and miR-29e, have shown an anti-viral activity. Overexpression of miR-29c in HepG2.2.15 cells efficiently repressed the expression of TNF-α-induced protein 3 and inhibited HBV DNA replication. miR-29c overexpression suppressed proliferation and induced apoptosis.276 HCV infection downregulated miR-29, and the overexpression of miR-29 decreased the level of HCV RNA in hepatic stellate cells. HCV infection downregulates miR-29 in hepatocytes, and this may explain the increase in collagen synthesis by reducing the miR-29 levels in activated hematopoietic stem cells (HSCs). It is possible that treatment with a miR-29 mimic in vivo may suppress HCV and decrease fibrosis.277 Many studies have shown a correlation between the occurrence of fibrosis in different organs such as kidney,278 heart,279 and liver280 with members of the miR-29 family.
Duck enteritis virus (DEV) and autophagy
DEV is the causative pathogen of duck viral enteritis disease that causes significant economic losses in the duck farming industry, resulting in the death of the birds and in lower egg production.281,282 Additionally, DEV may lead to morbidity and mortality in swans, geese, and wild waterfowl, and it is considered a serious threat to waterfowl species.283, 284, 285
Wu et al.286 examined the possible role of miR-30a-5p in DEV infection. They used qRT-PCR to measure Beclin-1 mRNA expression and levels of miRNAs (miR-30a-5p). Researchers used a dual-luciferase reporter assay (DLRA) to determine miR-30a-5p-Beclin-1 target interaction and western blotting to analyze Beclin-1 expression and autophagy in duck embryonic fibroblasts (DEFs). Moreover, the median tissue culture infective dose (TCID50) was calculated to estimate DEV titers. The results showed considerable downregulation of miR-30a-5p and remarkable upregulation of Beclin-1 mRNA in the DEV-infected DEFs. In addition, a DLRA verified the direct targeting of the 3′ UTR of the Beclin-1 gene by miR-30a-5p. Overexpression of miR-30a-5p decreased the level of Beclin-1 protein expression and Beclin-1-mediated autophagy, and it eventually repressed DEV replication. Alternatively, transfection of a miR-30a-5p inhibitor increased Beclin-1-mediated autophagy and stimulated DEV replication over the entire course of DEV infection. It was concluded that miR-30a-5p was capable of inhibiting DEV replication via reduction of autophagy by targeting Beclin-1. This approach could be a new anti-viral treatment strategy against DEV infection.286
DENV and autophagy
Dengue is a common arthropod transmitted viral disease in tropical and subtropical areas of the world.287 In DENV infections, the patient can either suffer from dengue shock syndrome (DSS) and serious dengue hemorrhagic fever (DHF) or self-limiting dengue fever (DF).287 Researchers have considered cell autophagy to be necessary for efficient DENV replication.116,288, 289, 290 Autophagy is involved in DENV replication via alterations in cellular lipid metabolism that increase the generation of the ATP required for DENV replication.291 Most investigations have been carried out in liver cell lines, such as Huh7 and HepG2. However, other studies have given contradictory results, showing that the induction of autophagy decreased the DENV2 yield in the monocytic cell line U937.292 Consequently, the effects of autophagy on DENV replication appear to be mediated in a cell type-specific manner and require further work to understand the full implications.
miR-146a has already been shown to play a role in DENV replication inside monocytes via affecting the TRAF6 innate adaptor molecule.293 Pu et al.294 reported a new contribution of miRNA miR-146a to the negative regulation of cellular autophagy in DENV-infected A549 cells and THP-1 cells. Results showed that overexpression of miR-146a blocked DENV2-induced autophagy and, conversely, that locked nucleic acid (LNA)-mediated suppression of miR-146a abrogated these effects. In addition, overexpression of TRAF6, which is a target of miR-146a, reversed the suppressive effect of miR-146a on autophagy. Furthermore, treatment with recombinant IFN-β fully restored the autophagy activity in the TRAF6-silenced cells. In DENV2-infected A549 cells, autophagy enhanced the pro-inflammatory response with increased production of TNF-α and IL-6. It was concluded that miR-146a is a negative regulator of DENV-induced autophagy and that TRAF6 was a main target of this miRNA in the modulation of DENV autophagy.294
Bovine viral diarrhea virus (BVDV) and autophagy
BVDV is an enveloped, positive sense ssRNA virus belonging to the genus Pestivirus in the Flaviviridae family.295 BVDV causes multiple diseases in cattle, such as hemorrhagic syndrome, diarrhea, respiratory disorders, reproductive disorders, mucosal disease, and persistent infections, which cause considerable economic losses to the cattle industry.296 Moreover, infection with the BVDV strain NADL considerably induced autophagy in Madin-Darby bovine kidney (MDBK) cells and enhanced the level of expression of two autophagy-associated genes, ATG14 and BECN1. In addition, the initial phases of autophagy were involved with BVDV NADL replication in MDBK cells.297
Fu et al. examined whether the Bos taurus (cattle) bta-miR-29b was upregulated in MDBK cells after infection with the BVDV strain NADL.298 Six hours after infection, the miR-29b level in infected cells was found to be 2.3-fold higher compared to normal cells. The autophagy process in BVDV NADL-infected cells was significantly inhibited by lentivirus-induced upregulation of miR-29b via direct targeting of ATG14 and ATG9A, two major autophagy-related proteins. The results were confirmed by overexpression of ATG14 and ATG9A leading to upregulation of autophagy and increased BVDV NADL replication. BVDV NADL replication was induced in MDBK cells during the early stages of autophagy, and it was inhibited after suppression of autophagy. It was concluded that there was a new connection between viral replication and miR-29b.298
Indirect regulation of the anoikis pathway by cellular miRNAs after virus infection
As discussed above, once metastatic cancer cells are detached from the ECM, they become resistant to anoikis cell death, thereby penetrating into the lymph and blood circulation.299,300 Tropomyosin 1 (TPM1) has mostly been studied as a tumor suppressor gene, which inhibits the transformed phenotype and reduces anchorage-independent growth. In addition, TPM1 downregulation led to inhibition of anoikis in breast cancer cells.301, 302, 303 Interestingly, high molecular weight (HMW) TPM1 was downregulated by high levels of oncomir hsa-miR-2 in transformed cells.304,305 The HMW forms of TPM1 and TPM2 undergo translocation to the endothelial cell surface when activated by growth factors such as vascular endothelial cell growth factor (VEGF) or basic fibroblast growth factor (bFGF). A variety of ligands, including histidine-proline-rich glycoprotein (HPRG),306,307 cleaved HMW kininogen (HKa),308,309 and endostatin310 can then bind to TPMs on the cell surface, acting as their receptors. The anti-angiogenic effects of such binding can be abrogated by administration of engineered antibodies against the TPM1 and TPM2 cell surface receptors.311 Therefore, TPM1 can not only stabilize actin stress fibers in favor of anti-angiogenesis, but it can also promote anoikis301 and inhibit cancer development in primary breast tumor cells.302
Kaposi’s sarcoma (KS)-associated herpesvirus (KSHV) and anoikis
KSHV causes the vascular hyperplasic disease KS by infecting endothelial cells.52,312,313 KSHV may also infect B lymphocytes and contribute to the multicentric lymphoproliferative condition called Castleman’s disease,314 or to primary effusion lymphoma.315 It was found that endothelial cells that have been infected by KSHV undergo malignant transformation with a higher angiogenic activity.312,313 KSHV has been shown to be in the latent phase in most KS cells, which only express a limited number of viral proteins, but there are 18 mature KSHV miRNAs derived from 12 precursor (pre-)miRNAs.316 Some targets of KSHV miRNAs (miR-Ks) have been identified.317, 318, 319 In the course of KS, a major rearrangement of the host cytoskeleton is observed,320 and two gene expression microarray assays were used to show that cytoskeleton TPM1 was downregulated in KSHV infection of telomerase-immortalized microvascular endothelial (TIME) cells or lymphatic endothelial cells (LECs).321,322 Because anoikis is inhibited by TPM1 downregulation in breast cancer cells,301 and also by KSHV-induced expression of vFLIP,323 it is proposed that KSHV-infected cells show partial resistance to anoikis cell death.
Kieffer-Kwon et al.74 examined whether the HMW TPM1 isoforms, which they showed to be downregulated by two KSHV miRNAs (miR-K2 and miR-K5), were able to suppress anoikis. They detected a functional binding site for miR-K5 in the 3′ UTR of one TPM1 isoform. Although KSHV-infected cells showed relatively higher levels of TPM1 protein compared to normal cells, these infected cells expressed even higher levels of TPM1, once miR-K2 or miR-K5 was downregulated. Mechanistically, miR-K2/miR-K5-induced downregulation of TPM1 isoforms increases VEGFA expression and tube formation from non-anchored human umbilical vein endothelial cells (HUVECs), and it increases the survival rate of HUVECs via suppressing anoikis. Overall, it was concluded that miR-K2 and miR-K5 were probably involved in the development of KSHV pathogenesis.74
HBV and anoikis
HCC is a complex and heterogeneous disease caused by different risk factors, including viral infections. When HCC undergoes an invasive transformation, it leads to metastasis, post-surgical recurrence, and poor response rates to chemotherapy, with a corresponding poor prognosis and lower survival rate.324,325 HCC may be caused by HBV infection, because DNA repair, cell cycle progression, cellular adhesion, transcription modulation, signal transduction, and apoptotic pathways are all regulated by the small open reading frame of the HBV genome, HBV X protein (HBx).326, 327, 328, 329 HBV can activate cellular oncogenes, promote genome instability, regulate the host immune response, and facilitate other oncogenic processes, and therefore HBV is considered to be a major carcinogenic virus.330
A member of the serine protease inhibitor (serpin) superfamily, named mammary serine protease inhibitor (maspin) or serpin B5, is highly expressed in normal epithelial cells but is down-expressed in cancer cells such as HCC cells.331 The maspin level is negatively associated with angiogenesis, tumor growth, and tumor cell migration.332, 333, 334, 335 Maspin also upregulates integrin expression, thus promoting cell-fibronectin binding and reducing metastasis.335 Several studies have shown that maspin is downregulated in HCC cell lines;336, 337, 338, 339 however, the role of maspin in HCC development is not fully understood.
Chen et al.75 explored the pathway by which HBV-infected HCC cells could regulate maspin expression, as well as the secondary effects of this regulation. In accordance with previous studies, HCC cells exhibited a lower level of maspin expression. Mechanistically, miR-7, miR-21, and miR-107 were found to be upregulated by HBx, which, in turn, resulted in maspin downregulation. Moreover, analysis of patient samples showed a relationship between increased expression of miRNAs and maspin downregulation in HBV-infected HCC patients that was also linked to poor overall survival. The cancer cell resistance to anoikis, tumor metastasis, and chemoresistance were all enhanced by HBx-induced maspin downregulation. In addition, overexpression of maspin mediated by a lentivirus enhanced the sensitivity of the HBx-expressing HCC cells to anoikis, as well as increasing their viability. The data showed that downregulation of maspin via miRNAs is involved in the HBx-mediated malignancy and chemoresistance in HCC.75
Conclusions
Viral pathogens are a major threat to public health, either by directly causing viral infections or being responsible for several cancer types. The underlying mechanism explaining how viruses affect different cellular functions is of great importance. One of the most critical cell functions affected by viruses is programmed cell death, which has repeatedly been shown to be mediated by several miRNAs. miRNA-dependent regulation of cell death pathways has proved to be a complex yet crucial issue, with more and more miRNAs having been found to be involved in both positive and negative regulation of these pathways. In this review, we summarized the available data on the virus-regulated miRNAs that modulate cell death pathways, and that can be used for further development of diagnostic and therapeutic techniques.
As shown in Table 1, several miRNAs, such as miR-155, miR-34a, miR-193b, and miR-93, have been reported to be regulated by various viruses. For instance, miR-21 was upregulated by several different viruses, such as HBV, EBV, and GaHV-2. Nevertheless, its target, the tumor suppressor PDCD4, was the same regardless of the virus type. However, it was reported that miR-21 regulated by the same HBV had different targets in each study (e.g., PDCD4, PTEN, IL-12, maspin, Fas-L). It can also be concluded that the same miRNAs often participate in the regulatory pathways that affect different types of cell death. For instance, the upregulation of miR-21 by HBV led to the inhibition of both apoptosis and anoikis. HBV led to the downregulation of miR-192-3p and the upregulation of the closely related miR-192-5p; the former enhanced autophagy while the latter inhibited apoptosis. This may be due to the presence of connections between different cell death pathways, which has already been shown. Most intriguingly, the regulation of cell death pathways by specific miRNAs was found to be different depending on the type of virus. For instance, miR-34a downregulation by HPV 16 and HPV 18 led to inhibition of apoptosis, whereas miR-34a downregulation by H1N1 produced the opposite result. In other words, apoptosis could result from both upregulation and downregulation of miR-34a, depending on the type of host cell and viral pathogen.
Overall, the miRNAs that are involved in the modulation of cell death pathways after virus infection show a degree of complexity and a variety of functions. Further studies are required to fully understand the complex pathways and functional targets before any therapeutic applications can be tested.
Author contributions
H.M. was involved in the conception, design, statistical analysis, and drafting of the manuscript. N.R., L.S., S.A.A., M.M.-T., M.R.H., V.T., N.R., J.S.N., M.S.E., H.B.B., and H.R.M. contributed to data collection and manuscript drafting. M.R.H. critically edited the manuscript. All authors approved the final version of the manuscript for submission.
Declaration of interests
The authors declare no competing interests.
Contributor Information
Vida Tajiknia, Email: tajiknia@gmail.com.
Neda Rahimian, Email: rahimian.n@iums.ac.ir.
Hamed Mirzaei, Email: mirzaei-h@kaums.ac.ir.
References
- 1.Strauss J.H., Strauss E.G. Overview of viruses and virus infection. Second Edition. Viruses and Human Disease; 2007. pp. 1–33. Academic Press. [Google Scholar]
- 2.zur Hausen H. Viruses in human cancers. Science. 1991;254:1167–1173. doi: 10.1126/science.1659743. [DOI] [PubMed] [Google Scholar]
- 3.Heise M.T. Viral pathogenesis. Reference Module in Biomedical Sciences. 2014;2014 B978-0-12-801238-3.00079-9. [Google Scholar]
- 4.Overholtzer M., Mailleux A.A., Mouneimne G., Normand G., Schnitt S.J., King R.W., Cibas E.S., Brugge J.S. A nonapoptotic cell death process, entosis, that occurs by cell-in-cell invasion. Cell. 2007;131:966–979. doi: 10.1016/j.cell.2007.10.040. [DOI] [PubMed] [Google Scholar]
- 5.Zhang R., Chi X., Wang S., Qi B., Yu X., Chen J.L. The regulation of autophagy by influenza A virus. BioMed Res. Int. 2014;2014:498083. doi: 10.1155/2014/498083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lu J., Murakami M., Verma S.C., Cai Q., Haldar S., Kaul R., Wasik M.A., Middeldorp J., Robertson E.S. Epstein-Barr Virus nuclear antigen 1 (EBNA1) confers resistance to apoptosis in EBV-positive B-lymphoma cells through up-regulation of survivin. Virology. 2011;410:64–75. doi: 10.1016/j.virol.2010.10.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zhao Y., Wang Z., Zhang W., Zhang L. MicroRNAs play an essential role in autophagy regulation in various disease phenotypes. Biofactors. 2019;45:844–856. doi: 10.1002/biof.1555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ashrafizadeh M., Zarrabi A., Hashemipour M., Vosough M., Najafi M., Shahinozzaman M., Hushmandi K., Khan H., Mirzaei H. Sensing the scent of death: Modulation of microRNAs by curcumin in gastrointestinal cancers. Pharmacol. Res. 2020;160:105199. doi: 10.1016/j.phrs.2020.105199. [DOI] [PubMed] [Google Scholar]
- 9.Mirzaei H.R., Sahebkar A., Mohammadi M., Yari R., Salehi H., Jafari M.H., Namdar A., Khabazian E., Jaafari M.R., Mirzaei H. Circulating microRNAs in hepatocellular carcinoma: Potential diagnostic and prognostic biomarkers. Curr. Pharm. Des. 2016;22:5257–5269. doi: 10.2174/1381612822666160303110838. [DOI] [PubMed] [Google Scholar]
- 10.Amiri A., Pourhanifeh M.H., Mirzaei H.R., Nahand J.S., Moghoofei M., Sahebnasagh R., Mirzaei H., Hamblin M.R. Exosomes and lung cancer: Roles in pathophysiology, diagnosis and therapeutic applications. Curr. Med. Chem. 2021;28:308–328. doi: 10.2174/0929867327666200204141952. [DOI] [PubMed] [Google Scholar]
- 11.Li N., Long B., Han W., Yuan S., Wang K. microRNAs: Important regulators of stem cells. Stem Cell Res. Ther. 2017;8:110. doi: 10.1186/s13287-017-0551-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Vienberg S., Geiger J., Madsen S., Dalgaard L.T. MicroRNAs in metabolism. Acta Physiol. (Oxf.) 2017;219:346–361. doi: 10.1111/apha.12681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yuan X., Berg N., Lee J.W., Le T.T., Neudecker V., Jing N., Eltzschig H. MicroRNA miR-223 as regulator of innate immunity. J. Leukoc. Biol. 2018;104:515–524. doi: 10.1002/JLB.3MR0218-079R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhang B., Pan X., Cobb G.P., Anderson T.A. MicroRNAs as oncogenes and tumor suppressors. Dev. Biol. 2007;302:1–12. doi: 10.1016/j.ydbio.2006.08.028. [DOI] [PubMed] [Google Scholar]
- 15.Mariño G., Niso-Santano M., Baehrecke E.H., Kroemer G. Self-consumption: The interplay of autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2014;15:81–94. doi: 10.1038/nrm3735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Suvorova I.I., Pospelov V.A. AMPK/Ulk1-dependent autophagy as a key mTOR regulator in the context of cell pluripotency. Cell Death Dis. 2019;10:260. doi: 10.1038/s41419-019-1501-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cory S., Adams J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer. 2002;2:647–656. doi: 10.1038/nrc883. [DOI] [PubMed] [Google Scholar]
- 18.Riedl S.J., Shi Y. Molecular mechanisms of caspase regulation during apoptosis. Nat. Rev. Mol. Cell Biol. 2004;5:897–907. doi: 10.1038/nrm1496. [DOI] [PubMed] [Google Scholar]
- 19.Lockshin R.A., Zakeri Z. Caspase-independent cell death? Oncogene. 2004;23:2766–2773. doi: 10.1038/sj.onc.1207514. [DOI] [PubMed] [Google Scholar]
- 20.D’Arcy M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019;43:582–592. doi: 10.1002/cbin.11137. [DOI] [PubMed] [Google Scholar]
- 21.Coates J.M., Galante J.M., Bold R.J. Cancer therapy beyond apoptosis: autophagy and anoikis as mechanisms of cell death. J. Surg. Res. 2010;164:301–308. doi: 10.1016/j.jss.2009.07.011. [DOI] [PubMed] [Google Scholar]
- 22.Slattery M.L., Mullany L.E., Sakoda L.C., Wolff R.K., Samowitz W.S., Herrick J.S. Dysregulated genes and miRNAs in the apoptosis pathway in colorectal cancer patients. Apoptosis. 2018;23:237–250. doi: 10.1007/s10495-018-1451-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Elliott M.R., Ravichandran K.S. Clearance of apoptotic cells: Implications in health and disease. J. Cell Biol. 2010;189:1059–1070. doi: 10.1083/jcb.201004096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Elmore S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007;35:495–516. doi: 10.1080/01926230701320337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Martinvalet D., Zhu P., Lieberman J. Granzyme A induces caspase-independent mitochondrial damage, a required first step for apoptosis. Immunity. 2005;22:355–370. doi: 10.1016/j.immuni.2005.02.004. [DOI] [PubMed] [Google Scholar]
- 26.Jacobson M.D., Weil M., Raff M.C. Programmed cell death in animal development. Cell. 1997;88:347–354. doi: 10.1016/s0092-8674(00)81873-5. [DOI] [PubMed] [Google Scholar]
- 27.Nelson D.A., Tan T.T., Rabson A.B., Anderson D., Degenhardt K., White E. Hypoxia and defective apoptosis drive genomic instability and tumorigenesis. Genes Dev. 2004;18:2095–2107. doi: 10.1101/gad.1204904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Mathew R., Karantza-Wadsworth V., White E. Role of autophagy in cancer. Nat. Rev. Cancer. 2007;7:961–967. doi: 10.1038/nrc2254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Adams J.M., Cory S. Bcl-2-regulated apoptosis: mechanism and therapeutic potential. Curr. Opin. Immunol. 2007;19:488–496. doi: 10.1016/j.coi.2007.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Folkman J. Angiogenesis and apoptosis. Semin. Cancer Biol. 2003;13:159–167. doi: 10.1016/s1044-579x(02)00133-5. [DOI] [PubMed] [Google Scholar]
- 31.Liu F., Zhang S., Zhao Z., Mao X., Huang J., Wu Z., Zheng L., Wang Q. MicroRNA-27b up-regulated by human papillomavirus 16 E7 promotes proliferation and suppresses apoptosis by targeting polo-like kinase2 in cervical cancer. Oncotarget. 2016;7:19666–19679. doi: 10.18632/oncotarget.7531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Linnstaedt S.D., Gottwein E., Skalsky R.L., Luftig M.A., Cullen B.R. Virally induced cellular microRNA miR-155 plays a key role in B-cell immortalization by Epstein-Barr virus. J. Virol. 2010;84:11670–11678. doi: 10.1128/JVI.01248-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Pourhanifeh M.H., Vosough M., Mahjoubin-Tehran M., Hashemipour M., Nejati M., Abbasi-Kolli M., Sahebkar A., Mirzaei H. Autophagy-related microRNAs: Possible regulatory roles and therapeutic potential in and gastrointestinal cancers. Pharmacol. Res. 2020;161:105133. doi: 10.1016/j.phrs.2020.105133. [DOI] [PubMed] [Google Scholar]
- 34.Bagherian A., Mardani R., Roudi B., Taghizadeh M., Banfshe H.R., Ghaderi A., Davoodvandi A., Shamollaghamsari S., Hamblin M.R., Mirzaei H. Combination therapy with nanomicellar-curcumin and temozolomide for in vitro therapy of glioblastoma multiforme via Wnt signaling pathways. J. Mol. Neurosci. 2020;70:1471–1483. doi: 10.1007/s12031-020-01639-z. [DOI] [PubMed] [Google Scholar]
- 35.Klionsky D.J., Abdel-Aziz A.K., Abdelfatah S., Abdellatif M., Abdoli A., Abel S., Abeliovich H., Abildgaard M.H., Abudu Y.P., Acevedo-Arozena A. 4th edition. Autophagy; 2021. Guidelines for the use and interpretation of assays for monitoring autophagy. Published online February 8, 2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mizushima N., Levine B., Cuervo A.M., Klionsky D.J. Autophagy fights disease through cellular self-digestion. Nature. 2008;451:1069–1075. doi: 10.1038/nature06639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lee S.J., Kang W.Y., Yoon Y., Jin J.Y., Song H.J., Her J.H., Kang S.M., Hwang Y.K., Kang K.J., Joo K.M., Nam D.H. Natural killer (NK) cells inhibit systemic metastasis of glioblastoma cells and have therapeutic effects against glioblastomas in the brain. BMC Cancer. 2015;15:1011. doi: 10.1186/s12885-015-2034-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Laplante M., Sabatini D.M. mTOR signaling in growth control and disease. Cell. 2012;149:274–293. doi: 10.1016/j.cell.2012.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Dunlop E.A., Tee A.R. mTOR and autophagy: A dynamic relationship governed by nutrients and energy. Semin. Cell Dev. Biol. 2014;36:121–129. doi: 10.1016/j.semcdb.2014.08.006. [DOI] [PubMed] [Google Scholar]
- 40.Vanhaesebroeck B., Guillermet-Guibert J., Graupera M., Bilanges B. The emerging mechanisms of isoform-specific PI3K signalling. Nat. Rev. Mol. Cell Biol. 2010;11:329–341. doi: 10.1038/nrm2882. [DOI] [PubMed] [Google Scholar]
- 41.Yin Q., Feng W., Shen X., Ju S. Regulatory effects of lncRNAs and miRNAs on autophagy in malignant tumorigenesis. Biosci. Rep. 2018;38 doi: 10.1042/BSR20180516. BSR20180516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Degenhardt K., Mathew R., Beaudoin B., Bray K., Anderson D., Chen G., Mukherjee C., Shi Y., Gélinas C., Fan Y. Autophagy promotes tumor cell survival and restricts necrosis, inflammation, and tumorigenesis. Cancer Cell. 2006;10:51–64. doi: 10.1016/j.ccr.2006.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ouyang L., Shi Z., Zhao S., Wang F.T., Zhou T.T., Liu B., Bao J.K. Programmed cell death pathways in cancer: A review of apoptosis, autophagy and programmed necrosis. Cell Prolif. 2012;45:487–498. doi: 10.1111/j.1365-2184.2012.00845.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bhutia S.K., Mukhopadhyay S., Sinha N., Das D.N., Panda P.K., Patra S.K., Maiti T.K., Mandal M., Dent P., Wang X.Y. Autophagy: Cancer’s friend or foe? Adv. Cancer Res. 2013;118:61–95. doi: 10.1016/B978-0-12-407173-5.00003-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bhutia S.K., Kegelman T.P., Das S.K., Azab B., Su Z.Z., Lee S.G., Sarkar D., Fisher P.B. Astrocyte elevated gene-1 induces protective autophagy. Proc. Natl. Acad. Sci. USA. 2010;107:22243–22248. doi: 10.1073/pnas.1009479107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hebner C., Weaver V.M., Debnath J. Modeling morphogenesis and oncogenesis in three-dimensional breast epithelial cultures. Annu. Rev. Pathol. 2008;3:313–339. doi: 10.1146/annurev.pathmechdis.3.121806.151526. [DOI] [PubMed] [Google Scholar]
- 47.Yu L., Alva A., Su H., Dutt P., Freundt E., Welsh S., Baehrecke E.H., Lenardo M.J. Regulation of an ATG7-beclin 1 program of autophagic cell death by caspase-8. Science. 2004;304:1500–1502. doi: 10.1126/science.1096645. [DOI] [PubMed] [Google Scholar]
- 48.Frisch S.M., Francis H. Disruption of epithelial cell-matrix interactions induces apoptosis. J. Cell Biol. 1994;124:619–626. doi: 10.1083/jcb.124.4.619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Giancotti F.G. Complexity and specificity of integrin signalling. Nat. Cell Biol. 2000;2:E13–E14. doi: 10.1038/71397. [DOI] [PubMed] [Google Scholar]
- 50.Danial N.N., Korsmeyer S.J. Cell death: Critical control points. Cell. 2004;116:205–219. doi: 10.1016/s0092-8674(04)00046-7. [DOI] [PubMed] [Google Scholar]
- 51.Bissell M.J., Radisky D. Putting tumours in context. Nat. Rev. Cancer. 2001;1:46–54. doi: 10.1038/35094059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Buchheit C.L., Weigel K.J., Schafer Z.T. Cancer cell survival during detachment from the ECM: Multiple barriers to tumour progression. Nat. Rev. Cancer. 2014;14:632–641. doi: 10.1038/nrc3789. [DOI] [PubMed] [Google Scholar]
- 53.Frisch S.M., Screaton R.A. Anoikis mechanisms. Curr. Opin. Cell Biol. 2001;13:555–562. doi: 10.1016/s0955-0674(00)00251-9. [DOI] [PubMed] [Google Scholar]
- 54.Li A.E., Ito H., Rovira I.I., Kim K.S., Takeda K., Yu Z.Y., Ferrans V.J., Finkel T. A role for reactive oxygen species in endothelial cell anoikis. Circ. Res. 1999;85:304–310. doi: 10.1161/01.res.85.4.304. [DOI] [PubMed] [Google Scholar]
- 55.Guadamillas M.C., Cerezo A., Del Pozo M.A. Overcoming anoikis—Pathways to anchorage-independent growth in cancer. J. Cell Sci. 2011;124:3189–3197. doi: 10.1242/jcs.072165. [DOI] [PubMed] [Google Scholar]
- 56.Simpson C.D., Anyiwe K., Schimmer A.D. Anoikis resistance and tumor metastasis. Cancer Lett. 2008;272:177–185. doi: 10.1016/j.canlet.2008.05.029. [DOI] [PubMed] [Google Scholar]
- 57.Chiarugi P., Giannoni E. Anoikis: A necessary death program for anchorage-dependent cells. Biochem. Pharmacol. 2008;76:1352–1364. doi: 10.1016/j.bcp.2008.07.023. [DOI] [PubMed] [Google Scholar]
- 58.Kim Y.N., Koo K.H., Sung J.Y., Yun U.J., Kim H. Anoikis resistance: An essential prerequisite for tumor metastasis. Int. J. Cell Biol. 2012;2012:306879. doi: 10.1155/2012/306879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Desgrosellier J.S., Cheresh D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer. 2010;10:9–22. doi: 10.1038/nrc2748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lotti R., Marconi A., Truzzi F., Dallaglio K., Gemelli C., Borroni R.G., Palazzo E., Pincelli C. A previously unreported function of β1B integrin isoform in caspase-8-dependent integrin-mediated keratinocyte death. J. Invest. Dermatol. 2010;130:2569–2577. doi: 10.1038/jid.2010.195. [DOI] [PubMed] [Google Scholar]
- 61.Stupack D.G., Puente X.S., Boutsaboualoy S., Storgard C.M., Cheresh D.A. Apoptosis of adherent cells by recruitment of caspase-8 to unligated integrins. J. Cell Biol. 2001;155:459–470. doi: 10.1083/jcb.200106070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ratheesh A., Yap A.S. A bigger picture: classical cadherins and the dynamic actin cytoskeleton. Nat. Rev. Mol. Cell Biol. 2012;13:673–679. doi: 10.1038/nrm3431. [DOI] [PubMed] [Google Scholar]
- 63.Lu M., Marsters S., Ye X., Luis E., Gonzalez L., Ashkenazi A. E-cadherin couples death receptors to the cytoskeleton to regulate apoptosis. Mol. Cell. 2014;54:987–998. doi: 10.1016/j.molcel.2014.04.029. [DOI] [PubMed] [Google Scholar]
- 64.Grossmann J. Molecular mechanisms of “detachment-induced apoptosis—Anoikis. Apoptosis. 2002;7:247–260. doi: 10.1023/a:1015312119693. [DOI] [PubMed] [Google Scholar]
- 65.Kovacs E.M., Ali R.G., McCormack A.J., Yap A.S. E-cadherin homophilic ligation directly signals through Rac and phosphatidylinositol 3-kinase to regulate adhesive contacts. J. Biol. Chem. 2002;277:6708–6718. doi: 10.1074/jbc.M109640200. [DOI] [PubMed] [Google Scholar]
- 66.Luebke-Wheeler J.L., Nedredal G., Yee L., Amiot B.P., Nyberg S.L. E-cadherin protects primary hepatocyte spheroids from cell death by a caspase-independent mechanism. Cell Transplant. 2009;18:1281–1287. doi: 10.3727/096368909X474258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Onder T.T., Gupta P.B., Mani S.A., Yang J., Lander E.S., Weinberg R.A. Loss of E-cadherin promotes metastasis via multiple downstream transcriptional pathways. Cancer Res. 2008;68:3645–3654. doi: 10.1158/0008-5472.CAN-07-2938. [DOI] [PubMed] [Google Scholar]
- 68.Li G., Satyamoorthy K., Herlyn M. N-cadherin-mediated intercellular interactions promote survival and migration of melanoma cells. Cancer Res. 2001;61:3819–3825. [PubMed] [Google Scholar]
- 69.Freedman V.H., Shin S.I. Cellular tumorigenicity in nude mice: Correlation with cell growth in semi-solid medium. Cell. 1974;3:355–359. doi: 10.1016/0092-8674(74)90050-6. [DOI] [PubMed] [Google Scholar]
- 70.Mori S., Chang J.T., Andrechek E.R., Matsumura N., Baba T., Yao G., Kim J.W., Gatza M., Murphy S., Nevins J.R. Anchorage-independent cell growth signature identifies tumors with metastatic potential. Oncogene. 2009;28:2796–2805. doi: 10.1038/onc.2009.139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Eble J.A., Haier J. Integrins in cancer treatment. Curr. Cancer Drug Targets. 2006;6:89–105. doi: 10.2174/156800906776056518. [DOI] [PubMed] [Google Scholar]
- 72.Reddig P.J., Juliano R.L. Clinging to life: Cell to matrix adhesion and cell survival. Cancer Metastasis Rev. 2005;24:425–439. doi: 10.1007/s10555-005-5134-3. [DOI] [PubMed] [Google Scholar]
- 73.Valentijn A.J., Zouq N., Gilmore A.P. Anoikis. Biochem. Soc. Trans. 2004;32:421–425. doi: 10.1042/BST0320421. [DOI] [PubMed] [Google Scholar]
- 74.Kieffer-Kwon P., Happel C., Uldrick T.S., Ramalingam D., Ziegelbauer J.M. KSHV microRNAs repress tropomyosin 1 and increase anchorage-independent growth and endothelial tube formation. PLoS ONE. 2015;10:e0135560. doi: 10.1371/journal.pone.0135560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Chen W.S., Yen C.J., Chen Y.J., Chen J.Y., Wang L.Y., Chiu S.J., Shih W.L., Ho C.Y., Wei T.T., Pan H.L. miRNA-7/21/107 contribute to HBx-induced hepatocellular carcinoma progression through suppression of maspin. Oncotarget. 2015;6:25962–25974. doi: 10.18632/oncotarget.4504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Chen T.M., Pecoraro G., Defendi V. Genetic analysis of in vitro progression of human papillomavirus-transfected human cervical cells. Cancer Res. 1993;53:1167–1171. [PubMed] [Google Scholar]
- 77.Bartel D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi: 10.1016/s0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 78.Zhang H., Li Y., Huang Q., Ren X., Hu H., Sheng H., Lai M. miR-148a promotes apoptosis by targeting Bcl-2 in colorectal cancer. Cell Death Differ. 2011;18:1702–1710. doi: 10.1038/cdd.2011.28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Gong J., Zhang J.P., Li B., Zeng C., You K., Chen M.X., Yuan Y., Zhuang S.M. MicroRNA-125b promotes apoptosis by regulating the expression of Mcl-1, Bcl-w and IL-6R. Oncogene. 2013;32:3071–3079. doi: 10.1038/onc.2012.318. [DOI] [PubMed] [Google Scholar]
- 80.Li W.H., Wu H.J., Li Y.X., Pan H.G., Meng T., Wang X. MicroRNA-143 promotes apoptosis of osteosarcoma cells by caspase-3 activation via targeting Bcl-2. Biomed. Pharmacother. 2016;80:8–15. doi: 10.1016/j.biopha.2016.03.001. [DOI] [PubMed] [Google Scholar]
- 81.Funamizu N., Lacy C.R., Kamada M., Yanaga K., Manome Y. MicroRNA-203 induces apoptosis by upregulating Puma expression in colon and lung cancer cells. Int. J. Oncol. 2015;47:1981–1988. doi: 10.3892/ijo.2015.3178. [DOI] [PubMed] [Google Scholar]
- 82.Alanazi I., Hoffmann P., Adelson D.L. MicroRNAs are part of the regulatory network that controls EGF induced apoptosis, including elements of the JAK/STAT pathway, in A431 cells. PLoS ONE. 2015;10:e0120337. doi: 10.1371/journal.pone.0120337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kim Y., Lee J., Ryu H. Modulation of autophagy by miRNAs. BMB Rep. 2015;48:371–372. doi: 10.5483/BMBRep.2015.48.7.121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Singh S.V., Dakhole A.N., Deogharkar A., Kazi S., Kshirsagar R., Goel A., Moiyadi A., Jalali R., Sridhar E., Gupta T. Restoration of miR-30a expression inhibits growth, tumorigenicity of medulloblastoma cells accompanied by autophagy inhibition. Biochem. Biophys. Res. Commun. 2017;491:946–952. doi: 10.1016/j.bbrc.2017.07.140. [DOI] [PubMed] [Google Scholar]
- 85.Ye Z., Fang B., Pan J., Zhang N., Huang J., Xie C., Lou T., Cao Z. miR-138 suppresses the proliferation, metastasis and autophagy of non-small cell lung cancer by targeting Sirt1. Oncol. Rep. 2017;37:3244–3252. doi: 10.3892/or.2017.5619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Maiuri M.C., Zalckvar E., Kimchi A., Kroemer G. Self-eating and self-killing: crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007;8:741–752. doi: 10.1038/nrm2239. [DOI] [PubMed] [Google Scholar]
- 87.Kroemer G., Mariño G., Levine B. Autophagy and the integrated stress response. Mol. Cell. 2010;40:280–293. doi: 10.1016/j.molcel.2010.09.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Maiuri M.C., Le Toumelin G., Criollo A., Rain J.C., Gautier F., Juin P., Tasdemir E., Pierron G., Troulinaki K., Tavernarakis N. Functional and physical interaction between Bcl-XL and a BH3-like domain in Beclin-1. EMBO J. 2007;26:2527–2539. doi: 10.1038/sj.emboj.7601689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Germain M., Nguyen A.P., Le Grand J.N., Arbour N., Vanderluit J.L., Park D.S., Opferman J.T., Slack R.S. MCL-1 is a stress sensor that regulates autophagy in a developmentally regulated manner. EMBO J. 2011;30:395–407. doi: 10.1038/emboj.2010.327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Luo S., Rubinsztein D.C. Apoptosis blocks Beclin 1-dependent autophagosome synthesis: An effect rescued by Bcl-xL. Cell Death Differ. 2010;17:268–277. doi: 10.1038/cdd.2009.121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Cimmino A., Calin G.A., Fabbri M., Iorio M.V., Ferracin M., Shimizu M., Wojcik S.E., Aqeilan R.I., Zupo S., Dono M. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc. Natl. Acad. Sci. USA. 2005;102:13944–13949. doi: 10.1073/pnas.0506654102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Xu J., Liao X., Wong C. Downregulations of B-cell lymphoma 2 and myeloid cell leukemia sequence 1 by microRNA 153 induce apoptosis in a glioblastoma cell line DBTRG-05MG. Int. J. Cancer. 2010;126:1029–1035. doi: 10.1002/ijc.24823. [DOI] [PubMed] [Google Scholar]
- 93.Jin Z., Li Y., Pitti R., Lawrence D., Pham V.C., Lill J.R., Ashkenazi A. Cullin3-based polyubiquitination and p62-dependent aggregation of caspase-8 mediate extrinsic apoptosis signaling. Cell. 2009;137:721–735. doi: 10.1016/j.cell.2009.03.015. [DOI] [PubMed] [Google Scholar]
- 94.Gao Z., Gammoh N., Wong P.M., Erdjument-Bromage H., Tempst P., Jiang X. Processing of autophagic protein LC3 by the 20S proteasome. Autophagy. 2010;6:126–137. doi: 10.4161/auto.6.1.10928. [DOI] [PubMed] [Google Scholar]
- 95.Yousefi S., Perozzo R., Schmid I., Ziemiecki A., Schaffner T., Scapozza L., Brunner T., Simon H.U. Calpain-mediated cleavage of Atg5 switches autophagy to apoptosis. Nat. Cell Biol. 2006;8:1124–1132. doi: 10.1038/ncb1482. [DOI] [PubMed] [Google Scholar]
- 96.Cheng Y., Ren X., Zhang Y., Patel R., Sharma A., Wu H., Robertson G.P., Yan L., Rubin E., Yang J.M. eEF-2 kinase dictates cross-talk between autophagy and apoptosis induced by Akt inhibition, thereby modulating cytotoxicity of novel Akt inhibitor MK-2206. Cancer Res. 2011;71:2654–2663. doi: 10.1158/0008-5472.CAN-10-2889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Su H., Yang J.R., Xu T., Huang J., Xu L., Yuan Y., Zhuang S.M. MicroRNA-101, down-regulated in hepatocellular carcinoma, promotes apoptosis and suppresses tumorigenicity. Cancer Res. 2009;69:1135–1142. doi: 10.1158/0008-5472.CAN-08-2886. [DOI] [PubMed] [Google Scholar]
- 98.Frankel L.B., Wen J., Lees M., Høyer-Hansen M., Farkas T., Krogh A., Jäättelä M., Lund A.H. MicroRNA-101 is a potent inhibitor of autophagy. EMBO J. 2011;30:4628–4641. doi: 10.1038/emboj.2011.331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Chen L., Yan H.X., Yang W., Hu L., Yu L.X., Liu Q., Li L., Huang D.D., Ding J., Shen F. The role of microRNA expression pattern in human intrahepatic cholangiocarcinoma. J. Hepatol. 2009;50:358–369. doi: 10.1016/j.jhep.2008.09.015. [DOI] [PubMed] [Google Scholar]
- 100.Xu J., Wang Y., Tan X., Jing H. MicroRNAs in autophagy and their emerging roles in crosstalk with apoptosis. Autophagy. 2012;8:873–882. doi: 10.4161/auto.19629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.López J.A., Alvarez-Salas L.M. Differential effects of miR-34c-3p and miR-34c-5p on SiHa cells proliferation apoptosis, migration and invasion. Biochem. Biophys. Res. Commun. 2011;409:513–519. doi: 10.1016/j.bbrc.2011.05.036. [DOI] [PubMed] [Google Scholar]
- 102.Wilting S.M., Verlaat W., Jaspers A., Makazaji N.A., Agami R., Meijer C.J., Snijders P.J., Steenbergen R.D. Methylation-mediated transcriptional repression of microRNAs during cervical carcinogenesis. Epigenetics. 2013;8:220–228. doi: 10.4161/epi.23605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Wilting S.M., Miok V., Jaspers A., Boon D., Sørgård H., Lando M., Snoek B.C., van Wieringen W.N., Meijer C.J., Lyng H. Aberrant methylation-mediated silencing of microRNAs contributes to HPV-induced anchorage independence. Oncotarget. 2016;7:43805–43819. doi: 10.18632/oncotarget.9698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Malagobadan S., Nagoor N.H. Evaluation of microRNAs regulating anoikis pathways and its therapeutic potential. BioMed Res. Int. 2015;2015:716816. doi: 10.1155/2015/716816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Zhang L., Wang X., Chen P. miR-204 down regulates SIRT1 and reverts SIRT1-induced epithelial-mesenchymal transition, anoikis resistance and invasion in gastric cancer cells. BMC Cancer. 2013;13:290. doi: 10.1186/1471-2407-13-290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Alessi D.R., Sakamoto K., Bayascas J.R. LKB1-dependent signaling pathways. Annu. Rev. Biochem. 2006;75:137–163. doi: 10.1146/annurev.biochem.75.103004.142702. [DOI] [PubMed] [Google Scholar]
- 107.Cheng H., Liu P., Wang Z.C., Zou L., Santiago S., Garbitt V., Gjoerup O.V., Iglehart J.D., Miron A., Richardson A.L. SIK1 couples LKB1 to p53-dependent anoikis and suppresses metastasis. Sci. Signal. 2009;2:ra35. doi: 10.1126/scisignal.2000369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Wang Y., Li W., Zang X., Chen N., Liu T., Tsonis P.A., Huang Y. MicroRNA-204-5p regulates epithelial-to-mesenchymal transition during human posterior capsule opacification by targeting SMAD4. Invest. Ophthalmol. Vis. Sci. 2013;54:323–332. doi: 10.1167/iovs.12-10904. [DOI] [PubMed] [Google Scholar]
- 109.Cochrane D.R., Howe E.N., Spoelstra N.S., Richer J.K. Loss of miR-200c: A marker of aggressiveness and chemoresistance in female reproductive cancers. J. Oncol. 2010;2010:821717. doi: 10.1155/2010/821717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Howe E.N., Cochrane D.R., Richer J.K. Targets of miR-200c mediate suppression of cell motility and anoikis resistance. Breast Cancer Res. 2011;13:R45. doi: 10.1186/bcr2867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Tejero R., Navarro A., Campayo M., Viñolas N., Marrades R.M., Cordeiro A., Ruíz-Martínez M., Santasusagna S., Molins L., Ramirez J., Monzó M. miR-141 and miR-200c as markers of overall survival in early stage non-small cell lung cancer adenocarcinoma. PLoS ONE. 2014;9:e101899. doi: 10.1371/journal.pone.0101899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Jazdzewski K., Murray E.L., Franssila K., Jarzab B., Schoenberg D.R., de la Chapelle A. Common SNP in pre-miR-146a decreases mature miR expression and predisposes to papillary thyroid carcinoma. Proc. Natl. Acad. Sci. USA. 2008;105:7269–7274. doi: 10.1073/pnas.0802682105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Zhong H., Wang H.R., Yang S., Zhong J.H., Wang T., Wang C., Chen F.Y. Targeting Smad4 links microRNA-146a to the TGF-β pathway during retinoid acid induction in acute promyelocytic leukemia cell line. Int. J. Hematol. 2010;92:129–135. doi: 10.1007/s12185-010-0626-5. [DOI] [PubMed] [Google Scholar]
- 114.Xiao B., Zhu E.D., Li N., Lu D.S., Li W., Li B.S., Zhao Y.L., Mao X.H., Guo G., Yu P.W., Zou Q.M. Increased miR-146a in gastric cancer directly targets SMAD4 and is involved in modulating cell proliferation and apoptosis. Oncol. Rep. 2012;27:559–566. doi: 10.3892/or.2011.1514. [DOI] [PubMed] [Google Scholar]
- 115.Hou Z., Xie L., Yu L., Qian X., Liu B. MicroRNA-146a is down-regulated in gastric cancer and regulates cell proliferation and apoptosis. Med. Oncol. 2012;29:886–892. doi: 10.1007/s12032-011-9862-7. [DOI] [PubMed] [Google Scholar]
- 116.Fu Y., Xu W., Chen D., Feng C., Zhang L., Wang X., Lv X., Zheng N., Jin Y., Wu Z. Enterovirus 71 induces autophagy by regulating has-miR-30a expression to promote viral replication. Antiviral Res. 2015;124:43–53. doi: 10.1016/j.antiviral.2015.09.016. [DOI] [PubMed] [Google Scholar]
- 117.Kong Q., Wang W., Li P. Regulator role of HPV E7 protein on miR-21 expression in cervical carcinoma cells and its functional implication. Int. J. Clin. Exp. Pathol. 2015;8:15808–15813. [PMC free article] [PubMed] [Google Scholar]
- 118.Wang X., Wang H.-K., McCoy J.P., Banerjee N.S., Rader J.S., Broker T.R., Meyers C., Chow L.T., Zheng Z.M. Oncogenic HPV infection interrupts the expression of tumor-suppressive miR-34a through viral oncoprotein E6. RNA. 2009;15:637–647. doi: 10.1261/rna.1442309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Xu J., Wan X., Chen X., Fang Y., Cheng X., Xie X., Lu W. miR-2861 acts as a tumor suppressor via targeting EGFR/AKT2/CCND1 pathway in cervical cancer induced by human papillomavirus virus 16 E6. Sci. Rep. 2016;6:28968. doi: 10.1038/srep28968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Yeung C.L.A., Tsang T.Y., Yau P.L., Kwok T.T. Human papillomavirus type 16 E6 suppresses microRNA-23b expression in human cervical cancer cells through DNA methylation of the host gene C9orf3. Oncotarget. 2017;8:12158–12173. doi: 10.18632/oncotarget.14555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Liu C., Lin J., Li L., Zhang Y., Chen W., Cao Z., Zuo H., Chen C., Kee K. HPV16 early gene E5 specifically reduces miRNA-196a in cervical cancer cells. Sci. Rep. 2015;5:7653. doi: 10.1038/srep07653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Morgan E.L., Patterson M.R., Ryder E.L., Lee S.Y., Wasson C.W., Harper K.L., Li Y., Griffin S., Blair G.E., Whitehouse A., Macdonald A. MicroRNA-18a targeting of the STK4/MST1 tumour suppressor is necessary for transformation in HPV positive cervical cancer. PLoS Pathog. 2020;16:e1008624. doi: 10.1371/journal.ppat.1008624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Anastasiadou E., Stroopinsky D., Alimperti S., Jiao A.L., Pyzer A.R., Cippitelli C., Pepe G., Severa M., Rosenblatt J., Etna M.P. Epstein-Barr virus-encoded EBNA2 alters immune checkpoint PD-L1 expression by downregulating miR-34a in B-cell lymphomas. Leukemia. 2019;33:132–147. doi: 10.1038/s41375-018-0178-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Fujimura T., Kambayashi Y., Ohuchi K., Muto Y., Aiba S. Treatment of advanced melanoma: Past, present and future. Life (Basel) 2020;10:208. doi: 10.3390/life10090208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Yang G.-D., Huang T.-J., Peng L.-X., Yang C.-F., Liu R.-Y., Huang H.-B., Chu Q.Q., Yang H.J., Huang J.L., Zhu Z.Y. Epstein-Barr virus_encoded LMP1 upregulates microRNA-21 to promote the resistance of nasopharyngeal carcinoma cells to cisplatin-induced apoptosis by suppressing PDCD4 and Fas-L. PLoS ONE. 2013;8:e78355. doi: 10.1371/journal.pone.0078355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Yang F., Liu Q., Hu C.M. Epstein-Barr virus-encoded LMP1 increases miR-155 expression, which promotes radioresistance of nasopharyngeal carcinoma via suppressing UBQLN1. Eur. Rev. Med. Pharmacol. Sci. 2015;19:4507–4515. [PubMed] [Google Scholar]
- 127.Kim J.H., Kim W.S., Park C. Epstein-Barr virus latent membrane protein-1 protects B-cell lymphoma from rituximab-induced apoptosis through miR-155-mediated Akt activation and up-regulation of Mcl-1. Leuk. Lymphoma. 2012;53:1586–1591. doi: 10.3109/10428194.2012.659736. [DOI] [PubMed] [Google Scholar]
- 128.Kim S.-M., Hur D.Y., Hong S.-W., Kim J.H. EBV-encoded EBNA1 regulates cell viability by modulating miR34a-NOX2-ROS signaling in gastric cancer cells. Biochem. Biophys. Res. Commun. 2017;494:550–555. doi: 10.1016/j.bbrc.2017.10.095. [DOI] [PubMed] [Google Scholar]
- 129.Harris-Arnold A., Arnold C.P., Schaffert S., Hatton O., Krams S.M., Esquivel C.O., Martinez O.M. Epstein-Barr virus modulates host cell microRNA-194 to promote IL-10 production and B lymphoma cell survival. Am. J. Transplant. 2015;15:2814–2824. doi: 10.1111/ajt.13375. [DOI] [PubMed] [Google Scholar]
- 130.Yin D., Wang Y., Sai W., Zhang L., Miao Y., Cao L., Zhai X., Feng X., Yang L. HBx-induced miR-21 suppresses cell apoptosis in hepatocellular carcinoma by targeting interleukin-12. Oncol. Rep. 2016;36:2305–2312. doi: 10.3892/or.2016.5026. [DOI] [PubMed] [Google Scholar]
- 131.Damania P., Sen B., Dar S.B., Kumar S., Kumari A., Gupta E., Sarin S.K., Venugopal S.K. Hepatitis B virus induces cell proliferation via HBx-induced microRNA-21 in hepatocellular carcinoma by targeting programmed cell death protein4 (PDCD4) and phosphatase and tensin homologue (PTEN) PLoS ONE. 2014;9:e91745. doi: 10.1371/journal.pone.0091745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Tian Y., Xiao X., Gong X., Peng F., Xu Y., Jiang Y., Gong G. HBx promotes cell proliferation by disturbing the cross-talk between miR-181a and PTEN. Sci. Rep. 2017;7:40089. doi: 10.1038/srep40089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Nielsen K.O., Jacobsen K.S., Mirza A.H., Winther T.N., Størling J., Glebe D., Pociot F., Hogh B. Hepatitis B virus upregulates host microRNAs that target apoptosis-regulatory genes in an in vitro cell model. Exp. Cell Res. 2018;371:92–103. doi: 10.1016/j.yexcr.2018.07.044. [DOI] [PubMed] [Google Scholar]
- 134.Cao Y., Chen J., Wang D., Peng H., Tan X., Xiong D., Huang A., Tang H. Upregulated in hepatitis B virus-associated hepatocellular carcinoma cells, miR-331-3p promotes proliferation of hepatocellular carcinoma cells by targeting ING5. Oncotarget. 2015;6:38093–38106. doi: 10.18632/oncotarget.5642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Yue X., Lan F., Yang W., Yang Y., Han L., Zhang A., Liu J., Zeng H., Jiang T., Pu P., Kang C. Interruption of β-catenin suppresses the EGFR pathway by blocking multiple oncogenic targets in human glioma cells. Brain Res. 2010;1366:27–37. doi: 10.1016/j.brainres.2010.10.032. [DOI] [PubMed] [Google Scholar]
- 136.Liu N., Jiao T., Huang Y., Liu W., Li Z., Ye X. Hepatitis B virus regulates apoptosis and tumorigenesis through the microRNA-15a-Smad7-transforming growth factor beta pathway. J. Virol. 2015;89:2739–2749. doi: 10.1128/JVI.02784-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Qin X., Li C., Guo T., Chen J., Wang H.-T., Wang Y.-T., Xiao Y.S., Li J., Liu P., Liu Z.S., Liu Q.Y. Upregulation of DARS2 by HBV promotes hepatocarcinogenesis through the miR-30e-5p/MAPK/NFAT5 pathway. J. Exp. Clin. Cancer Res. 2017;36:148. doi: 10.1186/s13046-017-0618-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Liu N., Zhang J., Jiao T., Li Z., Peng J., Cui Z., Ye X. Hepatitis B virus inhibits apoptosis of hepatoma cells by sponging the microRNA 15a/16 cluster. J. Virol. 2013;87:13370–13378. doi: 10.1128/JVI.02130-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Zhang C., Liu P., Zhang C. Hepatitis B virus X protein up-regulates alpha-fetoprotein to promote hepatocellular carcinoma by targeting miR-1236 and miR-1270. J. Cell. Biochem. 2020;121:2489–2499. doi: 10.1002/jcb.29471. [DOI] [PubMed] [Google Scholar]
- 140.Sheng Y., Li J., Zou C., Wang S., Cao Y., Zhang J., Huang A., Tang H. Downregulation of miR-101-3p by hepatitis B virus promotes proliferation and migration of hepatocellular carcinoma cells by targeting Rab5a. Arch. Virol. 2014;159:2397–2410. doi: 10.1007/s00705-014-2084-5. [DOI] [PubMed] [Google Scholar]
- 141.Wu G., Yu F., Xiao Z., Xu K., Xu J., Tang W., Wang J., Song E. Hepatitis B virus X protein downregulates expression of the miR-16 family in malignant hepatocytes in vitro. Br. J. Cancer. 2011;105:146–153. doi: 10.1038/bjc.2011.190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Wu G., Huang P., Ju X., Li Z., Wang Y. Lin28B over-expression mediates the repression of let-7 by hepatitis B virus X protein in hepatoma cells. Int. J. Clin. Exp. Med. 2015;8:15108–15116. [PMC free article] [PubMed] [Google Scholar]
- 143.Yip W.-K., Cheng A.S.-L., Zhu R., Lung R.W.-M., Tsang D.P.-F., Lau S.S.-K., Chen Y., Sung J.G., Lai P.B., Ng E.K. Carboxyl-terminal truncated HBx regulates a distinct microRNA transcription program in hepatocellular carcinoma development. PLoS ONE. 2011;6:e22888. doi: 10.1371/journal.pone.0022888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Hu X.M., Yan X.H., Hu Y.W., Huang J.L., Cao S.W., Ren T.Y., Tang Y.T., Lin L., Zheng L., Wang Q. miRNA-548p suppresses hepatitis B virus X protein associated hepatocellular carcinoma by downregulating oncoprotein hepatitis B x-interacting protein. Hepatol. Res. 2016;46:804–815. doi: 10.1111/hepr.12618. [DOI] [PubMed] [Google Scholar]
- 145.Zhao J., Wang W., Huang Y., Wu J., Chen M., Cui P., Zhang W., Zhang Y. HBx elevates oncoprotein AEG-1 expression to promote cell migration by downregulating miR-375 and miR-136 in malignant hepatocytes. DNA Cell Biol. 2014;33:715–722. doi: 10.1089/dna.2014.2376. [DOI] [PubMed] [Google Scholar]
- 146.Xie Q., He X., Chang Y., Jiang X., Lin J. [HBx gene down-regulates miR-192 expression and inhibits apoptosis of human hepatoma cell line HepG2] Zhonghua Gan Zang Bing Za Zhi. 2011;19:857–860. doi: 10.3760/cma.j.issn.1007-3418.2011.11.015. [DOI] [PubMed] [Google Scholar]
- 147.Gao F., Sun X., Wang L., Tang S., Yan C. Downregulation of microRNA-145 caused by hepatitis B virus X protein promotes expression of CUL5 and contributes to pathogenesis of hepatitis B virus-associated hepatocellular carcinoma. Cell. Physiol. Biochem. 2015;37:1547–1559. doi: 10.1159/000438522. [DOI] [PubMed] [Google Scholar]
- 148.Fu X., Ouyang Y., Mo J., Li R., Fu L., Peng S. Upregulation of microRNA-328-3p by hepatitis B virus contributes to THLE-2 cell injury by downregulating FOXO4. J. Transl. Med. 2020;18:143. doi: 10.1186/s12967-020-02299-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Rezaei S., Mahjoubin-Tehran M., Aghaee-Bakhtiari S.H., Jalili A., Movahedpour A., Khan H., Moghoofei M., Shojaei Z., R Hamblin M., Mirzaei H. Autophagy-related microRNAs in chronic lung diseases and lung cancer. Crit. Rev. Oncol. Hematol. 2020;153:103063. doi: 10.1016/j.critrevonc.2020.103063. [DOI] [PubMed] [Google Scholar]
- 150.Zhang Y., Wei W., Cheng N., Wang K., Li B., Jiang X., Sun S. Hepatitis C virus-induced up-regulation of microRNA-155 promotes hepatocarcinogenesis by activating Wnt signaling. Hepatology. 2012;56:1631–1640. doi: 10.1002/hep.25849. [DOI] [PubMed] [Google Scholar]
- 151.Shiu T.-Y., Huang S.-M., Shih Y.-L., Chu H.-C., Chang W.-K., Hsieh T.-Y. Retraction. PLoS ONE. 2020;15:e0235375. [Google Scholar]
- 152.Liu D., Wu J., Liu M., Yin H., He J., Zhang B. Downregulation of miRNA-30c and miR-203a is associated with hepatitis C virus core protein-induced epithelial-mesenchymal transition in normal hepatocytes and hepatocellular carcinoma cells. Biochem. Biophys. Res. Commun. 2015;464:1215–1221. doi: 10.1016/j.bbrc.2015.07.107. [DOI] [PubMed] [Google Scholar]
- 153.Xie Z., Xiao Z., Wang F. Hepatitis C virus nonstructural 5A protein (HCV-NS5A) inhibits hepatocyte apoptosis through the NF-κb/miR-503/bcl-2 pathway. Mol. Cells. 2017;40:202–210. doi: 10.14348/molcells.2017.2299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Braconi C., Valeri N., Gasparini P., Huang N., Taccioli C., Nuovo G., Suzuki T., Croce C.M., Patel T. Hepatitis C virus proteins modulate microRNA expression and chemosensitivity in malignant hepatocytes. Clin. Cancer Res. 2010;16:957–966. doi: 10.1158/1078-0432.CCR-09-2123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Patra T., Meyer K., Ray R.B., Ray R. Hepatitis C virus mediated inhibition of miR-181c activates ATM signaling and promotes hepatocyte growth. Hepatology. 2020;71:780–793. doi: 10.1002/hep.30893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Tomita M. Important roles of cellular microRNA miR-155 in leukemogenesis by human T-cell leukemia virus type 1 infection. ISRN Microbiol. 2012;2012:978607. doi: 10.5402/2012/978607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Yeung M.L., Yasunaga J., Bennasser Y., Dusetti N., Harris D., Ahmad N., Matsuoka M., Jeang K.T. Roles for microRNAs, miR-93 and miR-130b, and tumor protein 53-induced nuclear protein 1 tumor suppressor in cell growth dysregulation by human T-cell lymphotrophic virus 1. Cancer Res. 2008;68:8976–8985. doi: 10.1158/0008-5472.CAN-08-0769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Guan Z., Shi N., Song Y., Zhang X., Zhang M., Duan M. Induction of the cellular microRNA-29c by influenza virus contributes to virus-mediated apoptosis through repression of antiapoptotic factors BCL2L2. Biochem. Biophys. Res. Commun. 2012;425:662–667. doi: 10.1016/j.bbrc.2012.07.114. [DOI] [PubMed] [Google Scholar]
- 159.Stik G., Dambrine G., Pfeffer S., Rasschaert D. The oncogenic microRNA oncomiR-21 overexpressed during Marek’s disease lymphomagenesis is transactivated by the viral oncoprotein Meq. J. Virol. 2013;87:80–93. doi: 10.1128/JVI.02449-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Chanda S., Nandi S., Chawla-Sarkar M. Rotavirus-induced miR-142-5p elicits proviral milieu by targeting non-canonical transforming growth factor beta signalling and apoptosis in cells. Cell. Microbiol. 2016;18:733–747. doi: 10.1111/cmi.12544. [DOI] [PubMed] [Google Scholar]
- 161.Ru J., Sun H., Fan H., Wang C., Li Y., Liu M., Tang H. miR-23a facilitates the replication of HSV-1 through the suppression of interferon regulatory factor 1. PLoS ONE. 2014;9:e114021. doi: 10.1371/journal.pone.0114021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Ramachandran S., Ilias Basha H., Sarma N.J., Lin Y., Crippin J.S., Chapman W.C., Mohanakumar T. Hepatitis C virus induced miR200c down modulates FAP-1, a negative regulator of Src signaling and promotes hepatic fibrosis. PLoS ONE. 2013;8:e70744. doi: 10.1371/journal.pone.0070744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Fan N., Wang J. MicroRNA 34a contributes to virus-mediated apoptosis through binding to its target gene Bax in influenza A virus infection. Biomed. Pharmacother. 2016;83:1464–1470. doi: 10.1016/j.biopha.2016.08.049. [DOI] [PubMed] [Google Scholar]
- 164.Du X., Wang H., Xu F., Huang Y., Liu Z., Liu T. Enterovirus 71 induces apoptosis of SH-SY5Y human neuroblastoma cells through stimulation of endogenous microRNA let-7b expression. Mol. Med. Rep. 2015;12:953–959. doi: 10.3892/mmr.2015.3482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Smith J.L., Grey F.E., Uhrlaub J.L., Nikolich-Zugich J., Hirsch A.J. Induction of the cellular microRNA, Hs_154, by West Nile virus contributes to virus-mediated apoptosis through repression of antiapoptotic factors. J. Virol. 2012;86:5278–5287. doi: 10.1128/JVI.06883-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Bakre A., Wu W., Hiscox J., Spann K., Teng M.N., Tripp R.A. Human respiratory syncytial virus non-structural protein NS1 modifies miR-24 expression via transforming growth factor-β. J. Gen. Virol. 2015;96:3179–3191. doi: 10.1099/jgv.0.000261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Wang J., Chen J., Liu Y., Zeng X., Wei M., Wu S., Xiong Q., Song F., Yuan X., Xiao Y. Hepatitis B virus induces autophagy to promote its replication by the axis of miR-192-3p-XIAP through NF kappa B signaling. Hepatology. 2019;69:974–992. doi: 10.1002/hep.30248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Chen L., Ming X., Li W., Bi M., Yan B., Wang X., Yang P., Yang B. The microRNA-155 mediates hepatitis B virus replication by reinforcing SOCS1 signalling-induced autophagy. Cell Biochem. Funct. 2020;38:436–442. doi: 10.1002/cbf.3488. [DOI] [PubMed] [Google Scholar]
- 169.Lan S.H., Wu S.Y., Zuchini R., Lin X.Z., Su I.J., Tsai T.F., Lin Y.J., Wu C.T., Liu H.S. Autophagy suppresses tumorigenesis of hepatitis B virus-associated hepatocellular carcinoma through degradation of microRNA-224. Hepatology. 2014;59:505–517. doi: 10.1002/hep.26659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Cui X., Wang X., Zhou X., Jia J., Chen H., Zhao W. miR-106a regulates cell proliferation and autophagy by targeting LKB1 in HPV-16-associated cervical cancer. Mol. Cancer Res. 2020;18:1129–1141. doi: 10.1158/1541-7786.MCR-19-1114. [DOI] [PubMed] [Google Scholar]
- 171.Moon S.L., Dodd B.J., Brackney D.E., Wilusz C.J., Ebel G.D., Wilusz J. Flavivirus sfRNA suppresses antiviral RNA interference in cultured cells and mosquitoes and directly interacts with the RNAi machinery. Virology. 2015;485:322–329. doi: 10.1016/j.virol.2015.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Pourhanifeh M.H., Mahjoubin-Tehran M., Karimzadeh M.R., Mirzaei H.R., Razavi Z.S., Sahebkar A., Hosseini N., Mirzaei H., Hamblin M.R. Autophagy in cancers including brain tumors: role of MicroRNAs. Cell Commun. Signal. 2020;18:88. doi: 10.1186/s12964-020-00587-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Dasgupta A., Wilson D.W. ATP depletion blocks herpes simplex virus DNA packaging and capsid maturation. J. Virol. 1999;73:2006–2015. doi: 10.1128/jvi.73.3.2006-2015.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Li X., Fu Z., Liang H., Wang Y., Qi X., Ding M., Sun X., Zhou Z., Huang Y., Gu H. H5N1 influenza virus-specific miRNA-like small RNA increases cytokine production and mouse mortality via targeting poly(rC)-binding protein 2. Cell Res. 2018;28:157–171. doi: 10.1038/cr.2018.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Triboulet R., Mari B., Lin Y.-L., Chable-Bessia C., Bennasser Y., Lebrigand K., Cardinaud B., Maurin T., Barbry P., Baillat V. Suppression of microRNA-silencing pathway by HIV-1 during virus replication. Science. 2007;315:1579–1582. doi: 10.1126/science.1136319. [DOI] [PubMed] [Google Scholar]
- 176.Brès V., Tagami H., Péloponèse J.M., Loret E., Jeang K.T., Nakatani Y., Emiliani S., Benkirane M., Kiernan R.E. Differential acetylation of Tat coordinates its interaction with the co-activators cyclin T1 and PCAF. EMBO J. 2002;21:6811–6819. doi: 10.1093/emboj/cdf669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Boden D., Pusch O., Lee F., Tucker L., Ramratnam B. Human immunodeficiency virus type 1 escape from RNA interference. J. Virol. 2003;77:11531–11535. doi: 10.1128/JVI.77.21.11531-11535.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Westerhout E.M., Ooms M., Vink M., Das A.T., Berkhout B. HIV-1 can escape from RNA interference by evolving an alternative structure in its RNA genome. Nucleic Acids Res. 2005;33:796–804. doi: 10.1093/nar/gki220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Cullen B.R. Is RNA interference involved in intrinsic antiviral immunity in mammals? Nat. Immunol. 2006;7:563–567. doi: 10.1038/ni1352. [DOI] [PubMed] [Google Scholar]
- 180.Hu G., Drescher K.M., Chen X.M. Exosomal miRNAs: Biological properties and therapeutic potential. Front. Genet. 2012;3:56. doi: 10.3389/fgene.2012.00056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Girardi E., López P., Pfeffer S. On the importance of host microRNAs during viral infection. Front. Genet. 2018;9:439. doi: 10.3389/fgene.2018.00439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Akhtar M.M., Micolucci L., Islam M.S., Olivieri F., Procopio A.D. Bioinformatic tools for microRNA dissection. Nucleic Acids Res. 2016;44:24–44. doi: 10.1093/nar/gkv1221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Hemida M.G., Ye X., Thair S., Yang D. Exploiting the therapeutic potential of microRNAs in viral diseases: Expectations and limitations. Mol. Diagn. Ther. 2010;14:271–282. doi: 10.1007/BF03256383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Nahand J.S., Mahjoubin-Tehran M., Moghoofei M., Pourhanifeh M.H., Mirzaei H.R., Asemi Z., Khatami A., Bokharaei-Salim F., Mirzaei H., Hamblin M.R. Exosomal miRNAs: Novel players in viral infection. Epigenomics. 2020;12:353–370. doi: 10.2217/epi-2019-0192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Pourhanifeh M.H., Mahjoubin-Tehran M., Shafiee A., Hajighadimi S., Moradizarmehri S., Mirzaei H., Asemi Z. MicroRNAs and exosomes: Small molecules with big actions in multiple myeloma pathogenesis. IUBMB Life. 2020;72:314–333. doi: 10.1002/iub.2211. [DOI] [PubMed] [Google Scholar]
- 186.Segal M., Slack F.J. Challenges identifying efficacious miRNA therapeutics for cancer. Expert Opin. Drug Discov. 2020;15:987–992. doi: 10.1080/17460441.2020.1765770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Deng W., Lu M. The role of microRNAs in hepatocyte metabolism and hepatitis B virus replication. Virol. Sin. 2016;31:472–479. doi: 10.1007/s12250-016-3924-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Lo Re V., 3rd, Newcomb C.W., Carbonari D.M., Roy J.A., Althoff K.N., Kitahata M.M., Reddy K.R., Lim J.K., Silverberg M.J., Mayor A.M. Determinants of liver complications among HIV/hepatitis B virus-coinfected patients. J. Acquir. Immune Defic. Syndr. 2019;82:71–80. doi: 10.1097/QAI.0000000000002094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Khan S., Madan M., Virmani S.K. Prevalence of hepatitis B virus, genotypes, and mutants in HBsAg-positive patients in Meerut, India. Iran. Biomed. J. 2019;23:354–361. doi: 10.29252/.23.5.354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Zhang X., Liu H., Xie Z., Deng W., Wu C., Qin B., Hou J., Lu M. Epigenetically regulated miR-449a enhances hepatitis B virus replication by targeting cAMP-responsive element binding protein 5 and modulating hepatocytes phenotype. Sci. Rep. 2016;6:25389. doi: 10.1038/srep25389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Zhang X., Zhang E., Ma Z., Pei R., Jiang M., Schlaak J.F., Roggendorf M., Lu M. Modulation of hepatitis B virus replication and hepatocyte differentiation by microRNA-1. Hepatology. 2011;53:1476–1485. doi: 10.1002/hep.24195. [DOI] [PubMed] [Google Scholar]
- 192.Wang S., Qiu L., Yan X., Jin W., Wang Y., Chen L., Wu E., Ye X., Gao G.F., Wang F. Loss of microRNA 122 expression in patients with hepatitis B enhances hepatitis B virus replication through cyclin G1-modulated P53 activity. Hepatology. 2012;55:730–741. doi: 10.1002/hep.24809. [DOI] [PubMed] [Google Scholar]
- 193.Hu W., Wang X., Ding X., Li Y., Zhang X., Xie P., Yang J., Wang S. MicroRNA-141 represses HBV replication by targeting PPARA. PLoS ONE. 2012;7:e34165. doi: 10.1371/journal.pone.0034165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Huang J.Y., Chou S.F., Lee J.W., Chen H.L., Chen C.M., Tao M.H., Shih C. MicroRNA-130a can inhibit hepatitis B virus replication via targeting PGC1α and PPARγ. RNA. 2015;21:385–400. doi: 10.1261/rna.048744.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Ishibashi K., Tanaka Y., Morishita Y. The role of mammalian superaquaporins inside the cell. Biochim. Biophys. Acta. 2014;1840:1507–1512. doi: 10.1016/j.bbagen.2013.10.039. [DOI] [PubMed] [Google Scholar]
- 196.Benga G. On the definition, nomenclature and classification of water channel proteins (aquaporins and relatives) Mol. Aspects Med. 2012;33:514–517. doi: 10.1016/j.mam.2012.04.003. [DOI] [PubMed] [Google Scholar]
- 197.Verkman A.S., Hara-Chikuma M., Papadopoulos M.C. Aquaporins—New players in cancer biology. J. Mol. Med. (Berl.) 2008;86:523–529. doi: 10.1007/s00109-008-0303-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Papadopoulos M.C., Saadoun S. Key roles of aquaporins in tumor biology. Biochim. Biophys. Acta. 2015;1848(10 Pt B):2576–2583. doi: 10.1016/j.bbamem.2014.09.001. [DOI] [PubMed] [Google Scholar]
- 199.Zhang Z., Han Y., Sun G., Liu X., Jia X., Yu X. MicroRNA-325-3p inhibits cell proliferation and induces apoptosis in hepatitis B virus-related hepatocellular carcinoma by down-regulation of aquaporin 5. Cell. Mol. Biol. Lett. 2019;24:13. doi: 10.1186/s11658-019-0137-1. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 200.Liu C., Zheng S., Wang Y., Jing L., Gao H., Gao Y., Qi X., Qin L., Pan W., Wang X. Detection and molecular characterization of recombinant avian leukosis viruses in commercial egg-type chickens in China. Avian Pathol. 2011;40:269–275. doi: 10.1080/03079457.2011.560932. [DOI] [PubMed] [Google Scholar]
- 201.Payne L.N., Howes K., Gillespie A.M., Smith L.M. Host range of Rous sarcoma virus pseudotype RSV(HPRS-103) in 12 avian species: Support for a new avian retrovirus envelope subgroup, designated J. J. Gen. Virol. 1992;73:2995–2997. doi: 10.1099/0022-1317-73-11-2995. [DOI] [PubMed] [Google Scholar]
- 202.Payne L.N., Nair V. The long view: 40 years of avian leukosis research. Avian Pathol. 2012;41:11–19. doi: 10.1080/03079457.2011.646237. [DOI] [PubMed] [Google Scholar]
- 203.Cui Z., Sun S., Wang J. Reduced serologic response to Newcastle disease virus in broiler chickens exposed to a Chinese field strain of subgroup J avian leukosis virus. Avian Dis. 2006;50:191–195. doi: 10.1637/7409-071305R1.1. [DOI] [PubMed] [Google Scholar]
- 204.Gao Y., Yun B., Qin L., Pan W., Qu Y., Liu Z., Wang Y., Qi X., Gao H., Wang X. Molecular epidemiology of avian leukosis virus subgroup J in layer flocks in China. J. Clin. Microbiol. 2012;50:953–960. doi: 10.1128/JCM.06179-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Abolnik C., Wandrag D.B. Avian gyrovirus 2 and avirulent Newcastle disease virus coinfection in a chicken flock with neurologic symptoms and high mortalities. Avian Dis. 2014;58:90–94. doi: 10.1637/10657-090313-Reg.1. [DOI] [PubMed] [Google Scholar]
- 206.Cui Z., Sun S., Zhang Z., Meng S. Simultaneous endemic infections with subgroup J avian leukosis virus and reticuloendotheliosis virus in commercial and local breeds of chickens. Avian Pathol. 2009;38:443–448. doi: 10.1080/03079450903349188. [DOI] [PubMed] [Google Scholar]
- 207.Isfort R.J., Qian Z., Jones D., Silva R.F., Witter R., Kung H.J. Integration of multiple chicken retroviruses into multiple chicken herpesviruses: Herpesviral gD as a common target of integration. Virology. 1994;203:125–133. doi: 10.1006/viro.1994.1462. [DOI] [PubMed] [Google Scholar]
- 208.Gao Y., Liu Y., Guan X., Li X., Yun B., Qi X., Wang Y., Gao H., Cui H., Liu C. Differential expression of immune-related cytokine genes in response to J group avian leukosis virus infection in vivo. Mol. Immunol. 2015;64:106–111. doi: 10.1016/j.molimm.2014.11.004. [DOI] [PubMed] [Google Scholar]
- 209.Mildner A., Jung S. Development and function of dendritic cell subsets. Immunity. 2014;40:642–656. doi: 10.1016/j.immuni.2014.04.016. [DOI] [PubMed] [Google Scholar]
- 210.Banchereau J., Steinman R.M. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. doi: 10.1038/32588. [DOI] [PubMed] [Google Scholar]
- 211.Watowich S.S., Liu Y.J. Mechanisms regulating dendritic cell specification and development. Immunol. Rev. 2010;238:76–92. doi: 10.1111/j.1600-065X.2010.00949.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Kuipers H., Schnorfeil F.M., Brocker T. Differentially expressed microRNAs regulate plasmacytoid vs. conventional dendritic cell development. Mol. Immunol. 2010;48:333–340. doi: 10.1016/j.molimm.2010.07.007. [DOI] [PubMed] [Google Scholar]
- 213.Jurkin J., Schichl Y.M., Koeffel R., Bauer T., Richter S., Konradi S., Gesslbauer B., Strobl H. miR-146a is differentially expressed by myeloid dendritic cell subsets and desensitizes cells to TLR2-dependent activation. J. Immunol. 2010;184:4955–4965. doi: 10.4049/jimmunol.0903021. [DOI] [PubMed] [Google Scholar]
- 214.Lu C., Huang X., Zhang X., Roensch K., Cao Q., Nakayama K.I., Blazar B.R., Zeng Y., Zhou X. miR-221 and miR-155 regulate human dendritic cell development, apoptosis, and IL-12 production through targeting of p27kip1, KPC1, and SOCS-1. Blood. 2011;117:4293–4303. doi: 10.1182/blood-2010-12-322503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Hashimi S.T., Fulcher J.A., Chang M.H., Gov L., Wang S., Lee B. MicroRNA profiling identifies miR-34a and miR-21 and their target genes JAG1 and WNT1 in the coordinate regulation of dendritic cell differentiation. Blood. 2009;114:404–414. doi: 10.1182/blood-2008-09-179150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Pajer P., Pecenka V., Králová J., Karafiát V., Průková D., Zemanová Z., Kodet R., Dvorák M. Identification of potential human oncogenes by mapping the common viral integration sites in avian nephroblastoma. Cancer Res. 2006;66:78–86. doi: 10.1158/0008-5472.CAN-05-1728. [DOI] [PubMed] [Google Scholar]
- 217.Justice J., 4th, Malhotra S., Ruano M., Li Y., Zavala G., Lee N., Morgan R., Beemon K. The MET gene is a common integration target in avian leukosis virus subgroup J-induced chicken hemangiomas. J. Virol. 2015;89:4712–4719. doi: 10.1128/JVI.03225-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Li Y., Liu X., Yang Z., Xu C., Liu D., Qin J., Dai M., Hao J., Feng M., Huang X. The MYC, TERT, and ZIC1 genes are common targets of viral integration and transcriptional deregulation in avian leukosis virus subgroup J-induced myeloid leukosis. J. Virol. 2014;88:3182–3191. doi: 10.1128/JVI.02995-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Park H., Huang X., Lu C., Cairo M.S., Zhou X. MicroRNA-146a and microRNA-146b regulate human dendritic cell apoptosis and cytokine production by targeting TRAF6 and IRAK1 proteins. J. Biol. Chem. 2015;290:2831–2841. doi: 10.1074/jbc.M114.591420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Liu D., Dai M., Zhang X., Cao W., Liao M. Subgroup J avian leukosis virus infection of chicken dendritic cells induces apoptosis via the aberrant expression of microRNAs. Sci. Rep. 2016;6:20188. doi: 10.1038/srep20188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Chhabra M., Mittal V., Bhattacharya D., Rana U., Lal S. Chikungunya fever: A re-emerging viral infection. Indian J. Med. Microbiol. 2008;26:5–12. doi: 10.4103/0255-0857.38850. [DOI] [PubMed] [Google Scholar]
- 222.Assunção-Miranda I., Cruz-Oliveira C., Da Poian A.T. Molecular mechanisms involved in the pathogenesis of alphavirus-induced arthritis. BioMed Res. Int. 2013;2013:973516. doi: 10.1155/2013/973516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Fischer M., Staples J.E., Arboviral Diseases Branch, National Center for Emerging and Zoonotic Infectious Diseases, CDC Notes from the field: Chikungunya virus spreads in the Americas—Caribbean and South America, 2013–2014. MMWR Morb. Mortal. Wkly. Rep. 2014;63:500–501. [PMC free article] [PubMed] [Google Scholar]
- 224.Jaffar-Bandjee M.C., Ramful D., Gauzere B.A., Hoarau J.J., Krejbich-Trotot P., Robin S., Ribera A., Selambarom J., Gasque P. Emergence and clinical insights into the pathology of Chikungunya virus infection. Expert Rev. Anti Infect. Ther. 2010;8:987–996. doi: 10.1586/eri.10.92. [DOI] [PubMed] [Google Scholar]
- 225.Das T., Jaffar-Bandjee M.C., Hoarau J.J., Krejbich Trotot P., Denizot M., Lee-Pat-Yuen G., Sahoo R., Guiraud P., Ramful D., Robin S. Chikungunya fever: CNS infection and pathologies of a re-emerging arbovirus. Prog. Neurobiol. 2010;91:121–129. doi: 10.1016/j.pneurobio.2009.12.006. [DOI] [PubMed] [Google Scholar]
- 226.Joubert P.E., Werneke S., de la Calle C., Guivel-Benhassine F., Giodini A., Peduto L., Levine B., Schwartz O., Lenschow D., Albert M.L. Chikungunya-induced cell death is limited by ER and oxidative stress-induced autophagy. Autophagy. 2012;8:1261–1263. doi: 10.4161/auto.20751. [DOI] [PubMed] [Google Scholar]
- 227.Joubert P.E., Werneke S.W., de la Calle C., Guivel-Benhassine F., Giodini A., Peduto L., Levine B., Schwartz O., Lenschow D.J., Albert M.L. Chikungunya virus-induced autophagy delays caspase-dependent cell death. J. Exp. Med. 2012;209:1029–1047. doi: 10.1084/jem.20110996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Krejbich-Trotot P., Denizot M., Hoarau J.J., Jaffar-Bandjee M.C., Das T., Gasque P. Chikungunya virus mobilizes the apoptotic machinery to invade host cell defenses. FASEB J. 2011;25:314–325. doi: 10.1096/fj.10-164178. [DOI] [PubMed] [Google Scholar]
- 229.Sharma A., Balakathiresan N.S., Maheshwari R.K. Chikungunya virus infection alters expression of microRNAs involved in cellular proliferation, immune response and apoptosis. Intervirology. 2015;58:332–341. doi: 10.1159/000441309. [DOI] [PubMed] [Google Scholar]
- 230.Wu Y., Wu Y., Tefsen B., Shi Y., Gao G.F. Bat-derived influenza-like viruses H17N10 and H18N11. Trends Microbiol. 2014;22:183–191. doi: 10.1016/j.tim.2014.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Mostafa A., Abdelwhab E.M., Mettenleiter T.C., Pleschka S. Zoonotic potential of influenza A viruses: A comprehensive overview. Viruses. 2018;10:497. doi: 10.3390/v10090497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Smith D.J., Lapedes A.S., de Jong J.C., Bestebroer T.M., Rimmelzwaan G.F., Osterhaus A.D., Fouchier R.A. Mapping the antigenic and genetic evolution of influenza virus. Science. 2004;305:371–376. doi: 10.1126/science.1097211. [DOI] [PubMed] [Google Scholar]
- 233.Han X., Li Z., Chen H., Wang H., Mei L., Wu S., Zhang T., Liu B., Lin X. Influenza virus A/Beijing/501/2009(H1N1) NS1 interacts with β-tubulin and induces disruption of the microtubule network and apoptosis on A549 cells. PLoS ONE. 2012;7:e48340. doi: 10.1371/journal.pone.0048340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Mori I., Komatsu T., Takeuchi K., Nakakuki K., Sudo M., Kimura Y. In vivo induction of apoptosis by influenza virus. J. Gen. Virol. 1995;76:2869–2873. doi: 10.1099/0022-1317-76-11-2869. [DOI] [PubMed] [Google Scholar]
- 235.Wang Y., Brahmakshatriya V., Zhu H., Lupiani B., Reddy S.M., Yoon B.J., Gunaratne P.H., Kim J.H., Chen R., Wang J., Zhou H. Identification of differentially expressed miRNAs in chicken lung and trachea with avian influenza virus infection by a deep sequencing approach. BMC Genomics. 2009;10:512. doi: 10.1186/1471-2164-10-512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Li Y., Chan E.Y., Li J., Ni C., Peng X., Rosenzweig E., Tumpey T.M., Katze M.G. MicroRNA expression and virulence in pandemic influenza virus-infected mice. J. Virol. 2010;84:3023–3032. doi: 10.1128/JVI.02203-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Brogaard L., Heegaard P.M., Larsen L.E., Mortensen S., Schlegel M., Dürrwald R., Skovgaard K. Late regulation of immune genes and microRNAs in circulating leukocytes in a pig model of influenza A (H1N2) infection. Sci. Rep. 2016;6:21812. doi: 10.1038/srep21812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Skovgaard K., Cirera S., Vasby D., Podolska A., Breum S.Ø., Dürrwald R., Schlegel M., Heegaard P.M. Expression of innate immune genes, proteins and microRNAs in lung tissue of pigs infected experimentally with influenza virus (H1N2) Innate Immun. 2013;19:531–544. doi: 10.1177/1753425912473668. [DOI] [PubMed] [Google Scholar]
- 239.Zhao L., Zhu J., Zhou H., Zhao Z., Zou Z., Liu X., Lin X., Zhang X., Deng X., Wang R. Identification of cellular microRNA-136 as a dual regulator of RIG-I-mediated innate immunity that antagonizes H5N1 IAV replication in A549 cells. Sci. Rep. 2015;5:14991. doi: 10.1038/srep14991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Gui S., Chen X., Zhang M., Zhao F., Wan Y., Wang L., Xu G., Zhou L., Yue X., Zhu Y., Liu S. Mir-302c mediates influenza A virus-induced IFNβ expression by targeting NF-κB inducing kinase. FEBS Lett. 2015;589(24 Pt B):4112–4118. doi: 10.1016/j.febslet.2015.11.011. [DOI] [PubMed] [Google Scholar]
- 241.Li Z., Zhang J., Su J., Liu Y., Guo J., Zhang Y., Lu C., Xing S., Guan Y., Li Y. MicroRNAs in the immune organs of chickens and ducks indicate divergence of immunity against H5N1 avian influenza. FEBS Lett. 2015;589:419–425. doi: 10.1016/j.febslet.2014.12.019. [DOI] [PubMed] [Google Scholar]
- 242.Zhang X., Dong C., Sun X., Li Z., Zhang M., Guan Z., Duan M. Induction of the cellular miR-29c by influenza virus inhibits the innate immune response through protection of A20 mRNA. Biochem. Biophys. Res. Commun. 2014;450:755–761. doi: 10.1016/j.bbrc.2014.06.059. [DOI] [PubMed] [Google Scholar]
- 243.Fan N., Wang J. MicroRNA 34a contributes to virus-mediated apoptosis through binding to its target gene Bax in influenza A virus infection. Biomed. Pharmacother. 2016;83:1464–1470. doi: 10.1016/j.biopha.2016.08.049. [DOI] [PubMed] [Google Scholar]
- 244.Deretic V. Multiple regulatory and effector roles of autophagy in immunity. Curr. Opin. Immunol. 2009;21:53–62. doi: 10.1016/j.coi.2009.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Lee H.K., Lund J.M., Ramanathan B., Mizushima N., Iwasaki A. Autophagy-dependent viral recognition by plasmacytoid dendritic cells. Science. 2007;315:1398–1401. doi: 10.1126/science.1136880. [DOI] [PubMed] [Google Scholar]
- 246.De Leo A., Colavita F., Ciccosanti F., Fimia G.M., Lieberman P.M., Mattia E. Inhibition of autophagy in EBV-positive Burkitt’s lymphoma cells enhances EBV lytic genes expression and replication. Cell Death Dis. 2015;6:e1876. doi: 10.1038/cddis.2015.156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Lee Y.R., Lei H.Y., Liu M.T., Wang J.R., Chen S.H., Jiang-Shieh Y.F., Lin Y.S., Yeh T.M., Liu C.C., Liu H.S. Autophagic machinery activated by dengue virus enhances virus replication. Virology. 2008;374:240–248. doi: 10.1016/j.virol.2008.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Taylor M.P., Kirkegaard K. Modification of cellular autophagy protein LC3 by poliovirus. J. Virol. 2007;81:12543–12553. doi: 10.1128/JVI.00755-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Reggiori F., Monastyrska I., Verheije M.H., Calì T., Ulasli M., Bianchi S., Bernasconi R., de Haan C.A., Molinari M. Coronaviruses Hijack the LC3-I-positive EDEMosomes, ER-derived vesicles exporting short-lived ERAD regulators, for replication. Cell Host Microbe. 2010;7:500–508. doi: 10.1016/j.chom.2010.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Dreux M., Gastaminza P., Wieland S.F., Chisari F.V. The autophagy machinery is required to initiate hepatitis C virus replication. Proc. Natl. Acad. Sci. USA. 2009;106:14046–14051. doi: 10.1073/pnas.0907344106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Harris J., Hartman M., Roche C., Zeng S.G., O’Shea A., Sharp F.A., Lambe E.M., Creagh E.M., Golenbock D.T., Tschopp J. Autophagy controls IL-1β secretion by targeting pro-IL-1β for degradation. J. Biol. Chem. 2011;286:9587–9597. doi: 10.1074/jbc.M110.202911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Nakahira K., Haspel J.A., Rathinam V.A., Lee S.J., Dolinay T., Lam H.C., Englert J.A., Rabinovitch M., Cernadas M., Kim H.P. Autophagy proteins regulate innate immune responses by inhibiting the release of mitochondrial DNA mediated by the NALP3 inflammasome. Nat. Immunol. 2011;12:222–230. doi: 10.1038/ni.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Saitoh T., Fujita N., Jang M.H., Uematsu S., Yang B.G., Satoh T., Omori H., Noda T., Yamamoto N., Komatsu M. Loss of the autophagy protein Atg16L1 enhances endotoxin-induced IL-1β production. Nature. 2008;456:264–268. doi: 10.1038/nature07383. [DOI] [PubMed] [Google Scholar]
- 254.Crişan T.O., Plantinga T.S., van de Veerdonk F.L., Farcaş M.F., Stoffels M., Kullberg B.J., van der Meer J.W., Joosten L.A., Netea M.G. Inflammasome-independent modulation of cytokine response by autophagy in human cells. PLoS ONE. 2011;6:e18666. doi: 10.1371/journal.pone.0018666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Li C., Zhou H.-M. The role of manganese superoxide dismutase in inflammation defense. Enzyme Res. 2011;2011:387176. doi: 10.4061/2011/387176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Shrivastava S., Raychoudhuri A., Steele R., Ray R., Ray R.B. Knockdown of autophagy enhances the innate immune response in hepatitis C virus-infected hepatocytes. Hepatology. 2011;53:406–414. doi: 10.1002/hep.24073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Yang Q., Zhang Q., Zhang X., You L., Wang W., Liu W., Han Y., Ma C., Xu W., Chen J. HoxA10 facilitates SHP-1-catalyzed dephosphorylation of p38 MAPK/STAT3 to repress hepatitis B virus replication by a feedback regulatory mechanism. J. Virol. 2019;93:e01607-18. doi: 10.1128/JVI.01607-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Saeed U., Kim J., Piracha Z.Z., Kwon H., Jung J., Chwae Y.J., Park S., Shin H.J., Kim K. Parvulin 14 and parvulin 17 bind to HBx and cccDNA and upregulate hepatitis B virus replication from cccDNA to virion in an HBx-dependent manner. J. Virol. 2019;93:e01840-18. doi: 10.1128/JVI.01840-18. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 259.Yu H.B., Jiang H., Cheng S.T., Hu Z.W., Ren J.H., Chen J. AGK2, a SIRT2 inhibitor, inhibits hepatitis B virus replication in vitro and in vivo. Int. J. Med. Sci. 2018;15:1356–1364. doi: 10.7150/ijms.26125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Li X., Xie T., Gao L., Ma C., Yang X., Liang X. Prostaglandin E2 facilitates hepatitis B virus replication by impairing CTL function. Mol. Immunol. 2018;103:243–250. doi: 10.1016/j.molimm.2018.08.009. [DOI] [PubMed] [Google Scholar]
- 261.Zamani P., Oskuee R.K., Atkin S.L., Navashenaq J.G., Sahebkar A. MicroRNAs as important regulators of the NLRP3 inflammasome. Prog. Biophys. Mol. Biol. 2020;150:50–61. doi: 10.1016/j.pbiomolbio.2019.05.004. [DOI] [PubMed] [Google Scholar]
- 262.Yin B., Liu W., Yu P., Liu C., Chen Y., Duan X., Liao Z., Chen Y., Wang X., Pan X., Tao Z. Association between human papillomavirus and prostate cancer: A meta-analysis. Oncol. Lett. 2017;14:1855–1865. doi: 10.3892/ol.2017.6367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Guerrieri F., Belloni L., D’Andrea D., Pediconi N., Le Pera L., Testoni B., Scisciani C., Floriot O., Zoulim F., Tramontano A., Levrero M. Genome-wide identification of direct HBx genomic targets. BMC Genomics. 2017;18:184. doi: 10.1186/s12864-017-3561-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Guo H., Zhou T., Jiang D., Cuconati A., Xiao G.H., Block T.M., Guo J.T. Regulation of hepatitis B virus replication by the phosphatidylinositol 3-kinase-Akt signal transduction pathway. J. Virol. 2007;81:10072–10080. doi: 10.1128/JVI.00541-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Rawat S., Bouchard M.J. The hepatitis B virus (HBV) HBx protein activates AKT to simultaneously regulate HBV replication and hepatocyte survival. J. Virol. 2015;89:999–1012. doi: 10.1128/JVI.02440-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266.Fu L., Fu X., Mo J., Li X., Li R., Peng S. miR-146a-5p enhances hepatitis B virus replication through autophagy to promote aggravation of chronic hepatitis B. IUBMB Life. 2019;71:1336–1346. doi: 10.1002/iub.2044. [DOI] [PubMed] [Google Scholar]
- 267.Chen X., Hu Y., Zhang W., Chen K., Hu J., Li X., Liang L., Cai X., Hu J., Wang K. Cisplatin induces autophagy to enhance hepatitis B virus replication via activation of ROS/JNK and inhibition of the Akt/mTOR pathway. Free Radic. Biol. Med. 2019;131:225–236. doi: 10.1016/j.freeradbiomed.2018.12.008. [DOI] [PubMed] [Google Scholar]
- 268.Chen M., Hong M.J., Sun H., Wang L., Shi X., Gilbert B.E., Corry D.B., Kheradmand F., Wang J. Essential role for autophagy in the maintenance of immunological memory against influenza infection. Nat. Med. 2014;20:503–510. doi: 10.1038/nm.3521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Fischl W., Bartenschlager R. Exploitation of cellular pathways by Dengue virus. Curr. Opin. Microbiol. 2011;14:470–475. doi: 10.1016/j.mib.2011.07.012. [DOI] [PubMed] [Google Scholar]
- 270.Kyei G.B., Dinkins C., Davis A.S., Roberts E., Singh S.B., Dong C., Wu L., Kominami E., Ueno T., Yamamoto A. Autophagy pathway intersects with HIV-1 biosynthesis and regulates viral yields in macrophages. J. Cell Biol. 2009;186:255–268. doi: 10.1083/jcb.200903070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Sir D., Tian Y., Chen W.L., Ann D.K., Yen T.S., Ou J.H. The early autophagic pathway is activated by hepatitis B virus and required for viral DNA replication. Proc. Natl. Acad. Sci. USA. 2010;107:4383–4388. doi: 10.1073/pnas.0911373107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272.Alers S., Löffler A.S., Wesselborg S., Stork B. Role of AMPK-mTOR-Ulk1/2 in the regulation of autophagy: cross talk, shortcuts, and feedbacks. Mol. Cell. Biol. 2012;32:2–11. doi: 10.1128/MCB.06159-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 273.Neufeld T.P. TOR-dependent control of autophagy: biting the hand that feeds. Curr. Opin. Cell Biol. 2010;22:157–168. doi: 10.1016/j.ceb.2009.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274.Zhang R., Zhu Y., Li Y., Liu W., Yin L., Yin S., Ji C., Hu Y., Wang Q., Zhou X. Human umbilical cord mesenchymal stem cell exosomes alleviate sepsis-associated acute kidney injury via regulating microRNA-146b expression. Biotechnol. Lett. 2020;42:669–679. doi: 10.1007/s10529-020-02831-2. [DOI] [PubMed] [Google Scholar]
- 275.Lin Y., Deng W., Pang J., Kemper T., Hu J., Yin J., Zhang J., Lu M. The microRNA-99 family modulates hepatitis B virus replication by promoting IGF-1R/PI3K/Akt/mTOR/ULK1 signaling-induced autophagy. Cell Microbiol. 2017;19 doi: 10.1111/cmi.12709. [DOI] [PubMed] [Google Scholar]
- 276.Wang C.M., Wang Y., Fan C.G., Xu F.F., Sun W.S., Liu Y.G., Jia J.H. miR-29c targets TNFAIP3, inhibits cell proliferation and induces apoptosis in hepatitis B virus-related hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2011;411:586–592. doi: 10.1016/j.bbrc.2011.06.191. [DOI] [PubMed] [Google Scholar]
- 277.Bandyopadhyay S., Friedman R.C., Marquez R.T., Keck K., Kong B., Icardi M.S., Brown K.E., Burge C.B., Schmidt W.N., Wang Y., McCaffrey A.P. Hepatitis C virus infection and hepatic stellate cell activation downregulate miR-29: miR-29 overexpression reduces hepatitis C viral abundance in culture. J. Infect. Dis. 2011;203:1753–1762. doi: 10.1093/infdis/jir186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 278.Qin W., Chung A.C., Huang X.R., Meng X.M., Hui D.S., Yu C.M., Sung J.J., Lan H.Y. TGF-β/Smad3 signaling promotes renal fibrosis by inhibiting miR-29. J. Am. Soc. Nephrol. 2011;22:1462–1474. doi: 10.1681/ASN.2010121308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 279.van Rooij E., Sutherland L.B., Thatcher J.E., DiMaio J.M., Naseem R.H., Marshall W.S., Hill J.A., Olson E.N. Dysregulation of microRNAs after myocardial infarction reveals a role of miR-29 in cardiac fibrosis. Proc. Natl. Acad. Sci. USA. 2008;105:13027–13032. doi: 10.1073/pnas.0805038105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280.Roderburg C., Urban G.W., Bettermann K., Vucur M., Zimmermann H., Schmidt S., Janssen J., Koppe C., Knolle P., Castoldi M. Micro-RNA profiling reveals a role for miR-29 in human and murine liver fibrosis. Hepatology. 2011;53:209–218. doi: 10.1002/hep.23922. [DOI] [PubMed] [Google Scholar]
- 281.Wang G., Qu Y., Wang F., Hu D., Liu L., Li N., Yue R., Li C., Liu S. The comprehensive diagnosis and prevention of duck plague in northwest Shandong province of China. Poult. Sci. 2013;92:2892–2898. doi: 10.3382/ps.2013-03356. [DOI] [PubMed] [Google Scholar]
- 282.Dhama K., Kumar N., Saminathan M., Tiwari R., Karthik K., Kumar M.A., Palanivelu M., Shabbir M.Z., Malik Y.S., Singh R.K. Duck virus enteritis (duck plague)—A comprehensive update. Vet. Q. 2017;37:57–80. doi: 10.1080/01652176.2017.1298885. [DOI] [PubMed] [Google Scholar]
- 283.Kaleta E.F., Kuczka A., Kühnhold A., Bunzenthal C., Bönner B.M., Hanka K., Redmann T., Yilmaz A. Outbreak of duck plague (duck herpesvirus enteritis) in numerous species of captive ducks and geese in temporal conjunction with enforced biosecurity (in-house keeping) due to the threat of avian influenza A virus of the subtype Asia H5N1. Dtsch. Tierarztl. Wochenschr. 2007;114:3–11. [PubMed] [Google Scholar]
- 284.Keymer I.F., Gough R.E. Duck virus enteritis (anatid herpesvirus infection) in mute swans (Cygnus olor) Avian Pathol. 1986;15:161–170. doi: 10.1080/03079458608436275. [DOI] [PubMed] [Google Scholar]
- 285.Metwally S.A. Duck virus enteritis (duck plague) In: Swayne D.E., Glisson J.R., McDougald L.R., Nolan L.K., Suarez D.L., Nair V., editors. Diseases of Poultry. 13th Edition. Wiley-Blackwell; 2013. pp. 431–440. [Google Scholar]
- 286.Wu X., Jia R., Wang M., Chen S., Liu M., Zhu D., Zhao X., Yang Q., Wu Y., Yin Z. Downregulation of microRNA-30a-5p contributes to the replication of duck enteritis virus by regulating Beclin-1-mediated autophagy. Virol. J. 2019;16:144. doi: 10.1186/s12985-019-1250-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 287.Halstead S.B. Dengue. Lancet. 2007;370:1644–1652. doi: 10.1016/S0140-6736(07)61687-0. [DOI] [PubMed] [Google Scholar]
- 288.Khakpoor A., Panyasrivanit M., Wikan N., Smith D.R. A role for autophagolysosomes in dengue virus 3 production in HepG2 cells. J. Gen. Virol. 2009;90:1093–1103. doi: 10.1099/vir.0.007914-0. [DOI] [PubMed] [Google Scholar]
- 289.Panyasrivanit M., Khakpoor A., Wikan N., Smith D.R. Co-localization of constituents of the dengue virus translation and replication machinery with amphisomes. J. Gen. Virol. 2009;90:448–456. doi: 10.1099/vir.0.005355-0. [DOI] [PubMed] [Google Scholar]
- 290.Heaton N.S., Perera R., Berger K.L., Khadka S., Lacount D.J., Kuhn R.J., Randall G. Dengue virus nonstructural protein 3 redistributes fatty acid synthase to sites of viral replication and increases cellular fatty acid synthesis. Proc. Natl. Acad. Sci. USA. 2010;107:17345–17350. doi: 10.1073/pnas.1010811107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Heaton N.S., Randall G. Dengue virus-induced autophagy regulates lipid metabolism. Cell Host Microbe. 2010;8:422–432. doi: 10.1016/j.chom.2010.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Panyasrivanit M., Greenwood M.P., Murphy D., Isidoro C., Auewarakul P., Smith D.R. Induced autophagy reduces virus output in dengue infected monocytic cells. Virology. 2011;418:74–84. doi: 10.1016/j.virol.2011.07.010. [DOI] [PubMed] [Google Scholar]
- 293.Wu S., He L., Li Y., Wang T., Feng L., Jiang L., Zhang P., Huang X. miR-146a facilitates replication of dengue virus by dampening interferon induction by targeting TRAF6. J. Infect. 2013;67:329–341. doi: 10.1016/j.jinf.2013.05.003. [DOI] [PubMed] [Google Scholar]
- 294.Pu J., Wu S., Xie H., Li Y., Yang Z., Wu X., Huang X. miR-146a Inhibits dengue-virus-induced autophagy by targeting TRAF6. Arch. Virol. 2017;162:3645–3659. doi: 10.1007/s00705-017-3516-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Lackner T., Müller A., Pankraz A., Becher P., Thiel H.J., Gorbalenya A.E., Tautz N. Temporal modulation of an autoprotease is crucial for replication and pathogenicity of an RNA virus. J. Virol. 2004;78:10765–10775. doi: 10.1128/JVI.78.19.10765-10775.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296.Perdrizet J.A., Rebhun W.C., Dubovi E.J., Donis R.O. Bovine virus diarrhea—Clinical syndromes in dairy herds. Cornell Vet. 1987;77:46–74. [PubMed] [Google Scholar]
- 297.Fu Q., Shi H., Zhang H., Ren Y., Guo F., Qiao J., Jia B., Wang P., Chen C. Autophagy during early stages contributes to bovine viral diarrhea virus replication in MDBK cells. J. Basic Microbiol. 2014;54:1044–1052. doi: 10.1002/jobm.201300750. [DOI] [PubMed] [Google Scholar]
- 298.Fu Q., Shi H., Ni W., Shi M., Meng L., Zhang H., Ren Y., Guo F., Wang P., Qiao J. Lentivirus-mediated Bos taurus bta-miR-29b overexpression interferes with bovine viral diarrhoea virus replication and viral infection-related autophagy by directly targeting ATG14 and ATG9A in Madin-Darby bovine kidney cells. J. Gen. Virol. 2015;96:85–94. doi: 10.1099/vir.0.067140-0. [DOI] [PubMed] [Google Scholar]
- 299.Ma Z., Liu Z., Myers D.P., Terada L.S. Mechanotransduction and anoikis: Death and the homeless cell. Cell Cycle. 2008;7:2462–2465. doi: 10.4161/cc.7.16.6463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 300.Meredith J.E., Jr., Fazeli B., Schwartz M.A. The extracellular matrix as a cell survival factor. Mol. Biol. Cell. 1993;4:953–961. doi: 10.1091/mbc.4.9.953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Raval G.N., Bharadwaj S., Levine E.A., Willingham M.C., Geary R.L., Kute T., Prasad G.L. Loss of expression of tropomyosin-1, a novel class II tumor suppressor that induces anoikis, in primary breast tumors. Oncogene. 2003;22:6194–6203. doi: 10.1038/sj.onc.1206719. [DOI] [PubMed] [Google Scholar]
- 302.Mahadev K., Raval G., Bharadwaj S., Willingham M.C., Lange E.M., Vonderhaar B., Salomon D., Prasad G.L. Suppression of the transformed phenotype of breast cancer by tropomyosin-1. Exp. Cell Res. 2002;279:40–51. doi: 10.1006/excr.2002.5583. [DOI] [PubMed] [Google Scholar]
- 303.Bharadwaj S., Thanawala R., Bon G., Falcioni R., Prasad G.L. Resensitization of breast cancer cells to anoikis by tropomyosin-1: Role of Rho kinase-dependent cytoskeleton and adhesion. Oncogene. 2005;24:8291–8303. doi: 10.1038/sj.onc.1208993. [DOI] [PubMed] [Google Scholar]
- 304.Zhu S., Si M.L., Wu H., Mo Y.Y. MicroRNA-21 targets the tumor suppressor gene tropomyosin 1 (TPM1) J. Biol. Chem. 2007;282:14328–14336. doi: 10.1074/jbc.M611393200. [DOI] [PubMed] [Google Scholar]
- 305.Wang M., Li W., Chang G.Q., Ye C.S., Ou J.S., Li X.X., Liu Y., Cheang T.Y., Huang X.L., Wang S.M. MicroRNA-21 regulates vascular smooth muscle cell function via targeting tropomyosin 1 in arteriosclerosis obliterans of lower extremities. Arterioscler. Thromb. Vasc. Biol. 2011;31:2044–2053. doi: 10.1161/ATVBAHA.111.229559. [DOI] [PubMed] [Google Scholar]
- 306.Doñate F., Juarez J.C., Guan X., Shipulina N.V., Plunkett M.L., Tel-Tsur Z., Shaw D.E., Morgan W.T., Mazar A.P. Peptides derived from the histidine-proline domain of the histidine-proline-rich glycoprotein bind to tropomyosin and have antiangiogenic and antitumor activities. Cancer Res. 2004;64:5812–5817. doi: 10.1158/0008-5472.CAN-04-0440. [DOI] [PubMed] [Google Scholar]
- 307.Mallery S.R., Morse M.A., Wilson R.F., Pei P., Ness G.M., Bradburn J.E., Renner R.J., Schuller D.E., Robertson F.M. AIDS-related Kaposi’s sarcoma cells rapidly internalize endostatin, which co-localizes to tropomysin microfilaments and inhibits cytokine-mediated migration and invasion. J. Cell. Biochem. 2003;89:133–143. doi: 10.1002/jcb.10489. [DOI] [PubMed] [Google Scholar]
- 308.Zhang J.C., Claffey K., Sakthivel R., Darzynkiewicz Z., Shaw D.E., Leal J., Wang Y.C., Lu F.M., McCrae K.R. Two-chain high molecular weight kininogen induces endothelial cell apoptosis and inhibits angiogenesis: partial activity within domain 5. FASEB J. 2000;14:2589–2600. doi: 10.1096/fj.99-1025com. [DOI] [PubMed] [Google Scholar]
- 309.Zhang J.C., Donate F., Qi X., Ziats N.P., Juarez J.C., Mazar A.P., Pang Y.P., McCrae K.R. The antiangiogenic activity of cleaved high molecular weight kininogen is mediated through binding to endothelial cell tropomyosin. Proc. Natl. Acad. Sci. USA. 2002;99:12224–12229. doi: 10.1073/pnas.192668299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.MacDonald N.J., Shivers W.Y., Narum D.L., Plum S.M., Wingard J.N., Fuhrmann S.R., Liang H., Holland-Linn J., Chen D.H., Sim B.K. Endostatin binds tropomyosin. A potential modulator of the antitumor activity of endostatin. J. Biol. Chem. 2001;276:25190–25196. doi: 10.1074/jbc.M100743200. [DOI] [PubMed] [Google Scholar]
- 311.Doñate F., McCrae K., Shaw D.E., Mazar A.P. Extracellular tropomyosin: A novel common pathway target for anti-angiogenic therapy. Curr. Cancer Drug Targets. 2004;4:543–553. doi: 10.2174/1568009043332781. [DOI] [PubMed] [Google Scholar]
- 312.Guilluy C., Zhang Z., Bhende P.M., Sharek L., Wang L., Burridge K., Damania B. Latent KSHV infection increases the vascular permeability of human endothelial cells. Blood. 2011;118:5344–5354. doi: 10.1182/blood-2011-03-341552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Wang L., Damania B. Kaposi’s sarcoma-associated herpesvirus confers a survival advantage to endothelial cells. Cancer Res. 2008;68:4640–4648. doi: 10.1158/0008-5472.CAN-07-5988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 314.Soulier J., Grollet L., Oksenhendler E., Cacoub P., Cazals-Hatem D., Babinet P., d’Agay M.F., Clauvel J.P., Raphael M., Degos L. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in multicentric Castleman’s disease. Blood. 1995;86:1276–1280. [PubMed] [Google Scholar]
- 315.Cesarman E., Chang Y., Moore P.S., Said J.W., Knowles D.M. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in AIDS-related body-cavity-based lymphomas. N. Engl. J. Med. 1995;332:1186–1191. doi: 10.1056/NEJM199505043321802. [DOI] [PubMed] [Google Scholar]
- 316.Umbach J.L., Cullen B.R. In-depth analysis of Kaposi’s sarcoma-associated herpesvirus microRNA expression provides insights into the mammalian microRNA-processing machinery. J. Virol. 2010;84:695–703. doi: 10.1128/JVI.02013-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 317.Ziegelbauer J.M. Functions of Kaposi’s sarcoma-associated herpesvirus microRNAs. Biochim. Biophys. Acta. 2011;1809:623–630. doi: 10.1016/j.bbagrm.2011.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 318.Liang D., Lin X., Lan K. Looking at Kaposi’s sarcoma-associated herpesvirus-host interactions from a microRNA viewpoint. Front. Microbiol. 2012;2:271. doi: 10.3389/fmicb.2011.00271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 319.Ramalingam D., Kieffer-Kwon P., Ziegelbauer J.M. Emerging themes from EBV and KSHV microRNA targets. Viruses. 2012;4:1687–1710. doi: 10.3390/v4091687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 320.Sharma-Walia N., Naranatt P.P., Krishnan H.H., Zeng L., Chandran B. Kaposi’s sarcoma-associated herpesvirus/human herpesvirus 8 envelope glycoprotein gB induces the integrin-dependent focal adhesion kinase-Src-phosphatidylinositol 3-kinase-rho GTPase signal pathways and cytoskeletal rearrangements. J. Virol. 2004;78:4207–4223. doi: 10.1128/JVI.78.8.4207-4223.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 321.Carroll P.A., Brazeau E., Lagunoff M. Kaposi’s sarcoma-associated herpesvirus infection of blood endothelial cells induces lymphatic differentiation. Virology. 2004;328:7–18. doi: 10.1016/j.virol.2004.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 322.Cheng F., Pekkonen P., Laurinavicius S., Sugiyama N., Henderson S., Günther T., Rantanen V., Kaivanto E., Aavikko M., Sarek G. KSHV-initiated notch activation leads to membrane-type-1 matrix metalloproteinase-dependent lymphatic endothelial-to-mesenchymal transition. Cell Host Microbe. 2011;10:577–590. doi: 10.1016/j.chom.2011.10.011. [DOI] [PubMed] [Google Scholar]
- 323.Efklidou S., Bailey R., Field N., Noursadeghi M., Collins M.K. vFLIP from KSHV inhibits anoikis of primary endothelial cells. J. Cell Sci. 2008;121:450–457. doi: 10.1242/jcs.022343. [DOI] [PubMed] [Google Scholar]
- 324.Lara-Pezzi E., Gómez-Gaviro M.V., Gálvez B.G., Mira E., Iñiguez M.A., Fresno M., Martínez-A C., Arroyo A.G., López-Cabrera M. The hepatitis B virus X protein promotes tumor cell invasion by inducing membrane-type matrix metalloproteinase-1 and cyclooxygenase-2 expression. J. Clin. Invest. 2002;110:1831–1838. doi: 10.1172/JCI200215887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 325.Lee J.O., Lee K.W., Oh D.Y., Kim J.H., Im S.A., Kim T.Y., Bang Y.J. Combination chemotherapy with capecitabine and cisplatin for patients with metastatic hepatocellular carcinoma. Ann. Oncol. 2009;20:1402–1407. doi: 10.1093/annonc/mdp010. [DOI] [PubMed] [Google Scholar]
- 326.Diao J., Garces R., Richardson C.D. X protein of hepatitis B virus modulates cytokine and growth factor related signal transduction pathways during the course of viral infections and hepatocarcinogenesis. Cytokine Growth Factor Rev. 2001;12:189–205. doi: 10.1016/s1359-6101(00)00034-4. [DOI] [PubMed] [Google Scholar]
- 327.Hodgson A.J., Keasler V.V., Slagle B.L. Premature cell cycle entry induced by hepatitis B virus regulatory HBx protein during compensatory liver regeneration. Cancer Res. 2008;68:10341–10348. doi: 10.1158/0008-5472.CAN-08-2695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328.Wang W.H., Hullinger R.L., Andrisani O.M. Hepatitis B virus X protein via the p38MAPK pathway induces E2F1 release and ATR kinase activation mediating p53 apoptosis. J. Biol. Chem. 2008;283:25455–25467. doi: 10.1074/jbc.M801934200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 329.Cheng P., Li Y., Yang L., Wen Y., Shi W., Mao Y., Chen P., Lv H., Tang Q., Wei Y. Hepatitis B virus X protein (HBx) induces G2/M arrest and apoptosis through sustained activation of cyclin B1-CDK1 kinase. Oncol. Rep. 2009;22:1101–1107. doi: 10.3892/or_00000542. [DOI] [PubMed] [Google Scholar]
- 330.Bréchot C. Pathogenesis of hepatitis B virus-related hepatocellular carcinoma: old and new paradigms. Gastroenterology. 2004;127(5, Suppl 1):S56–S61. doi: 10.1053/j.gastro.2004.09.016. [DOI] [PubMed] [Google Scholar]
- 331.Yu L.R., Zeng R., Shao X.X., Wang N., Xu Y.H., Xia Q.C. Identification of differentially expressed proteins between human hepatoma and normal liver cell lines by two-dimensional electrophoresis and liquid chromatography-ion trap mass spectrometry. Electrophoresis. 2000;21:3058–3068. doi: 10.1002/1522-2683(20000801)21:14<3058::AID-ELPS3058>3.0.CO;2-U. [DOI] [PubMed] [Google Scholar]
- 332.Bailey C.M., Khalkhali-Ellis Z., Seftor E.A., Hendrix M.J. Biological functions of maspin. J. Cell. Physiol. 2006;209:617–624. doi: 10.1002/jcp.20782. [DOI] [PubMed] [Google Scholar]
- 333.Blandamura S., Giacomelli L., Leo G., Segato P., Ninfo V. Nuclear maspin detection in renal cell tumours: Possible diagnostic role and correlation with p53 status. Histopathology. 2006;49:274–282. doi: 10.1111/j.1365-2559.2006.02504.x. [DOI] [PubMed] [Google Scholar]
- 334.Blacque O.E., Worrall D.M. Evidence for a direct interaction between the tumor suppressor serpin, maspin, and types I and III collagen. J. Biol. Chem. 2002;277:10783–10788. doi: 10.1074/jbc.M110992200. [DOI] [PubMed] [Google Scholar]
- 335.Seftor R.E., Seftor E.A., Sheng S., Pemberton P.A., Sager R., Hendrix M.J. maspin suppresses the invasive phenotype of human breast carcinoma. Cancer Res. 1998;58:5681–5685. [PubMed] [Google Scholar]
- 336.Futscher B.W., Oshiro M.M., Wozniak R.J., Holtan N., Hanigan C.L., Duan H., Domann F.E. Role for DNA methylation in the control of cell type specific maspin expression. Nat. Genet. 2002;31:175–179. doi: 10.1038/ng886. [DOI] [PubMed] [Google Scholar]
- 337.Luo J.L., Tan W., Ricono J.M., Korchynskyi O., Zhang M., Gonias S.L., Cheresh D.A., Karin M. Nuclear cytokine-activated IKKα controls prostate cancer metastasis by repressing Maspin. Nature. 2007;446:690–694. doi: 10.1038/nature05656. [DOI] [PubMed] [Google Scholar]
- 338.Huang W.C., Chen W.S., Chen Y.J., Wang L.Y., Hsu S.C., Chen C.C., Hung M.C. Hepatitis B virus X protein induces IKKα nuclear translocation via Akt-dependent phosphorylation to promote the motility of hepatocarcinoma cells. J. Cell. Physiol. 2012;227:1446–1454. doi: 10.1002/jcp.22860. [DOI] [PubMed] [Google Scholar]
- 339.Sharma G., Mirza S., Parshad R., Srivastava A., Gupta S.D., Pandya P., Ralhan R. Clinical significance of Maspin promoter methylation and loss of its protein expression in invasive ductal breast carcinoma: Correlation with VEGF-A and MTA1 expression. Tumour Biol. 2011;32:23–32. doi: 10.1007/s13277-010-0087-8. [DOI] [PubMed] [Google Scholar]