

Abstract
Background
Acute type A aortic dissection (ATAAD) is one of the most lethal cardiovascular diseases, and its molecular mechanism remains unclear.
Methods
Differentially expressed genes (DEGs) between ATAAD and control were detected by limma R package in GSE52093, GSE153434, GSE98770, and GSE84827, respectively. The coexpression network of DEGs was identified by the WGCNA package. Enrichment analysis was performed for module genes that were positively correlated with ATAAD using clusterProfiler R package. In addition, differentially methylated markers between aortic dissection and control were identified by ChAMP package. After comparing with ATAAD-related genes, a protein-protein interaction (PPI) network was established based on the STRING database. The genes with the highest connectivity were identified as hub genes. Finally, differential immune cell infiltration between ATAAD and control was identified by ssGSEA.
Results
From GSE52093 and GSE153434, 268 module genes were obtained with consistent direction of differential expression and high correlation with ATAAD. They were significantly enriched in T cell activation, HIF-1 signaling pathway, and cell cycle. In addition, 2060 differentially methylated markers were obtained from GSE84827. Among them, 77 methylation markers were ATAAD-related DEGs. Using the PPI network, we identified MYC, ITGA2, RND3, BCL2, and PHLPP2 as hub genes. Finally, we identified significantly differentially infiltrated immune cells in ATAAD.
Conclusion
The hub genes we identified may be regulated by methylation and participate in the development of ATAAD through immune inflammation and oxidative stress response. The findings may provide new insights into the molecular mechanisms and therapeutic targets for ATAAD.
1. Introduction
Aortic dissection (AD) is a serious invasive vascular disease with high mortality and limited treatment options [1]. The incidence of aortic dissection ranges from between 3.5 and 6/100,000 person-years in the general population to as high as 10/100,000 person-years in the elderly [2]. Usually, aortic dissection is caused by intimal tear, which further causes blood to flow into the media layer of the aorta, resulting in the separation of the layers within the aortic wall [3]. When the ascending aorta is involved, this dissection is known as Stanford type A aortic dissection (STAAD) [4]. Surgical mortality for acute Stanford type A aortic dissection (ATAAD) is relatively high, despite advances in medical and surgical treatment over the past 30 years [5]. Theoretically, once acute STAAD is diagnosed, patients should undergo emergency surgical treatment immediately [6]. However, limited by geographical, economic, and technical conditions, not all patients can receive timely treatment.
Early clinical symptoms of ATAAD may mimic those of other diseases, such as acute coronary syndrome, pulmonary embolism, or pneumothorax, often leading to delayed diagnosis [7–9]. When ATAAD is detected early and treated promptly, the chance of survival is greatly improved [10–12]. However, even in experienced cardiac centers, the early mortality rate for surgical treatment of acute aortic dissection is around 10%, and many patients still die before surgery [13]. Therefore, we believe that identifying early prognostic biomarkers can leverage patient characteristics and symptoms to optimize treatment strategies [14, 15].
At present, the molecular mechanism of ATAAD remains unclear. Chronic inflammation of the aortic lining has been reported to cause aneurysm growth, leading to aortic dissection [16–18]. In animal models, adventitial inflammation characterized by neutrophil aggregation can promote tissue damage, leading to aortic dilation and rupture [19]. In addition, the medial integrity maintained by collagen and elastin cross-linking is one of the keys in preventing aortic dissection [20]. Bone marrow mesenchymal stem cells are also potential contributors to aortic repair [21].
It is increasingly believed that human disease states are not caused by a single change but by the multifactorial regulation of biological systems [22]. In many cardiovascular diseases, important epigenetic modifications, including methylation, have been shown to affect the development or progression of the disease [23]. Methylation modification of the gene may serve as a diagnostic and prognostic marker in patients with aortic dissection [24].
Weighted gene coexpression network analysis (WGCNA) is a widely used method to build coexpression pairwise correlation matrices [25]. Exclusively based on coexpression analysis, it will better represent genes with a small effect size acting together [26]. WGCNA provides a systems-level insight into the signaling networks that may be associated with a phenotype of interest [27].
The network-based approach provided a powerful option for systematic analysis to identify candidate target genes. The aim of this study was to identify DEGs and related methylation modifications in ATAAD compared with healthy controls. At the same time, the molecular mechanisms involved in gene expression changes were discussed. This study is helpful in identifying new DNA methylation markers and improving both our understanding and the treatment level of ATAAD.
2. Materials and Methods
2.1. Data Sources
Aortic dissection data were collected from the Gene Expression Omnibus (GEO) database. We screened datasets with a sample size greater than 5. GSE52093 included gene expression data of dissected ascending aorta from patients with acute Stanford type A aortic dissection (n = 7) and normal controls (n = 5). GSE98770 included gene expression data of dissected ascending aorta from patients with acute type A aortic dissection (ATAAD) (n = 6) and gene expression data of nondissected ascending aorta obtained from transplant donors (n = 5). GSE153434 included gene expression data of dissected ascending aorta from patients with Stanford type A aortic dissection (n = 10) and normal control samples (n = 10).
2.2. Differential Gene Expression Analysis
The differential expression analysis between aortic dissection and healthy controls was performed by using the R software package limma [28]. The genes with P ≤ 0.05 (up-/downregulated) were extracted as differentially expressed genes (DEGs) [29, 30].
2.3. WGCNA
A gene coexpression network was constructed through the WGCNA package [31] using differentially expressed genes. The genes with similar expression behavior were divided into different modules. After determining the soft thresholding, the network was developed. Module-trait relationships were calculated using a Pearson correlation between modules and clinical trait. P value < 0.05 was regarded as significant.
2.4. Enrichment Analysis
Module genes were analyzed using the clusterProfiler R package [32] for Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG). The Gene Ontology (GO) enrichment results included the biological process (BP), cellular component (CC), and molecular function (MF) [33]. A term with P < 0.05 was considered a functionally enriched term.
2.5. Methylation Data Analysis
GSE84274 included methylation profiling of ascending aorta from 6 normal and 12 aortic dissection patients. The difference of methylation sites between aortic dissection and healthy controls was analyzed by the ChAMP software package [34]. Adjust (adj) P value < 0.05 was regarded as significant.
2.6. PPI Network Construction
We utilized the Search Tool for the Retrieval of Interacting Genes (STRING) database (http://string-db.org) to construct a protein-protein interaction (PPI) network for module genes, with a combined score > 0.4. The PPI network was visualized through Cytoscape software (Version 3.7.0) [35–38]. The genes with the top 5 degrees for connecting other genes in the network were considered as hub genes.
2.7. Single-Sample Gene Set Enrichment Analysis (ssGSEA)
To investigate the immune infiltration landscape of acute type A aortic dissection, ssGSEA was performed to evaluate the level of immune infiltration in a sample according to immune cell-specific marker genes [39]. Infiltration levels for immune cells were quantified using the ssGSEA implementation in gsva R package. P value < 0.05 was considered significant.
3. Results
3.1. Coexpression Network of Differentially Expressed Genes
To obtain genes related to acute type A aortic dissection, we compared them with healthy controls. A total of 4913 differentially expressed genes were obtained in GSE52093 (Figure 1(a)). We selected β = 18 as the soft thresholding to ensure that the network can obey the scale-free criteria (Figure 1(b)). The created network included three modules (Figure 1(c)). Then, 4682 differentially expressed genes were obtained in GSE153434 (Figure 1(d)). Setting β = 10 as the soft thresholding, we got 9 modules (Figures 1(e) and 1(f)).
Figure 1.
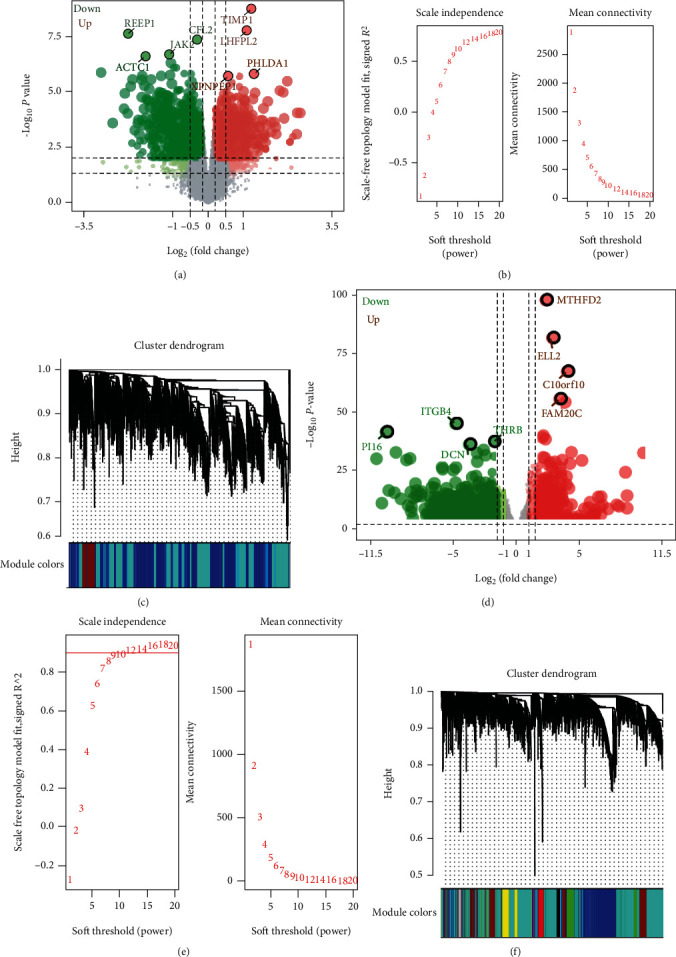
WGCNA network for differentially expressed genes. (a) Volcano map of differentially expressed genes between acute type A aortic dissection and controls in GSE52093. Red nodes are significantly upregulated genes, and green nodes are significantly downregulated genes. (b) The soft-threshold power versus scale-free topology model fit index and mean connectivity for GSE52093. The left image shows the scale-free fit index (y-axis) as a function of the soft-thresholding power (x-axis). The right image shows the average connectivity (degree, y-axis) as a function of the soft-thresholding power (x-axis). (c) Module clustering tree of differentially expressed genes in GSE52093. (d) Volcano map of differentially expressed genes between acute type A aortic dissection and normal controls in GSE153434. (e) The soft-threshold power versus scale-free topology model fit index and mean connectivity for GSE153434. (f) Module clustering tree of differentially expressed genes in GSE153434.
3.2. Biological Functions of Module Genes
The correlation analysis found that MEturquoise (module 2) of GSE52093 had the strongest correlation with ATAAD (Figure 2(a)). MEbrown (module 1), MEyellow (module 5), MEgreen (module 4), and MEblack (module 7) in GSE153434 were positively correlated with ATAAD (Figure 2(b)). Then, we obtained 268 common genes that expressed in the same direction (upregulated or downregulated expression) in these modules (Figure 2(c)). They may have a stronger association with ATAAD. Enrichment analysis revealed that common genes were mainly enriched in response to oxygen levels, T cell activation, leukocyte migration, and NIK/NF-kappaB signaling biological functions (Figure 2(d)). In addition, the p53 signaling pathway, the HIF-1 signaling pathway, the FoxO signaling pathway, and the cell cycle of the KEGG pathways were also significantly enriched (Figure 2(e)).
Figure 2.
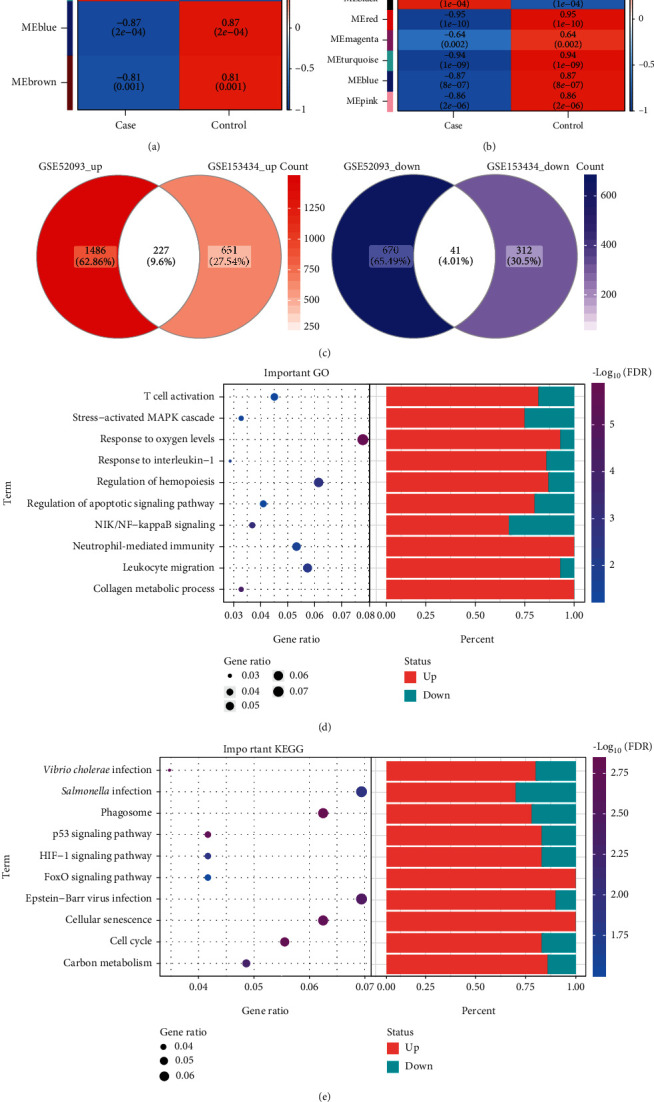
The enrichment analysis of related genes for acute type A aortic dissection. The correlation between module and clinical trait in GSE52093 (a) and in GSE153434 (b). Red represents positive correlation, and blue represents negative correlation. (c) Genes up- or downregulated simultaneously from two datasets in modules which positively correlated with acute type A aortic dissection. (d) The main biological processes of common gene enrichment. (e) The main KEGG pathway of common gene enrichment. Red bars represent upregulated terms, and green bars represent downregulated terms.
3.3. Methylated ATAAD-Related Genes
By comparing the differences between aortic dissection patients and controls, we obtained 46,845 differentially methylated positions (DMPs) (Figure 3(a)). Most DMPs were concentrated at the chr1 position (Figure 3(b)). We identified 2060 genes with opposite methylation and transcription levels as methylation markers (Figure 3(c)). Interestingly, among these methylation markers, we found that 77 genes were common genes (Figure 3(d)). Using the PPI network, we identified the top five genes with the highest connectivity as hub genes (Figure 4(a)). Compared with the control, MYC, ITGA2, and RND3 were upregulated in ATAAD, and BCL2 and PHLPP2 were downregulated (Figure 4(b)). The AUC values of hub genes were greater than 0.8 in both datasets, which may have a diagnostic role for ATAAD (Figure 4(c)).
Figure 3.
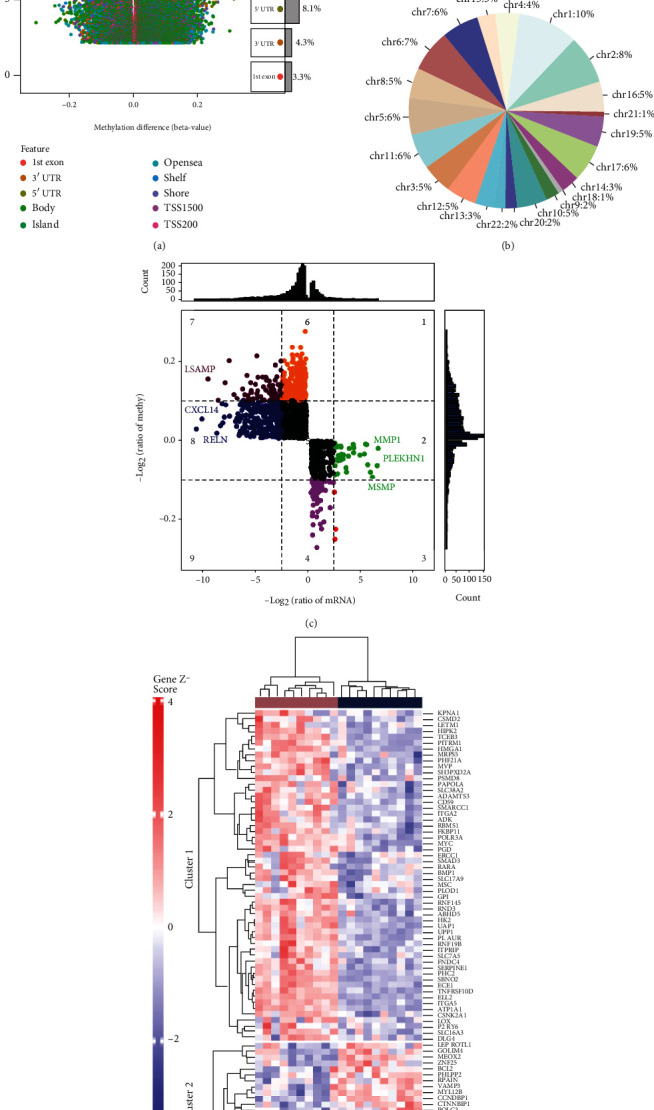
Acute type A aortic dissection-related methylation markers. (a) Differential methylation positions between aortic dissection patients and controls. (b) Proportion of differentially methylated positions in different chromosomes. (c) Genes whose transcription levels are opposite to the methylation level. (d) Heatmap of the expression of methylation markers in GSE153434. Red nodes are significantly upregulated genes, and blue nodes are significantly downregulated genes.
Figure 4.
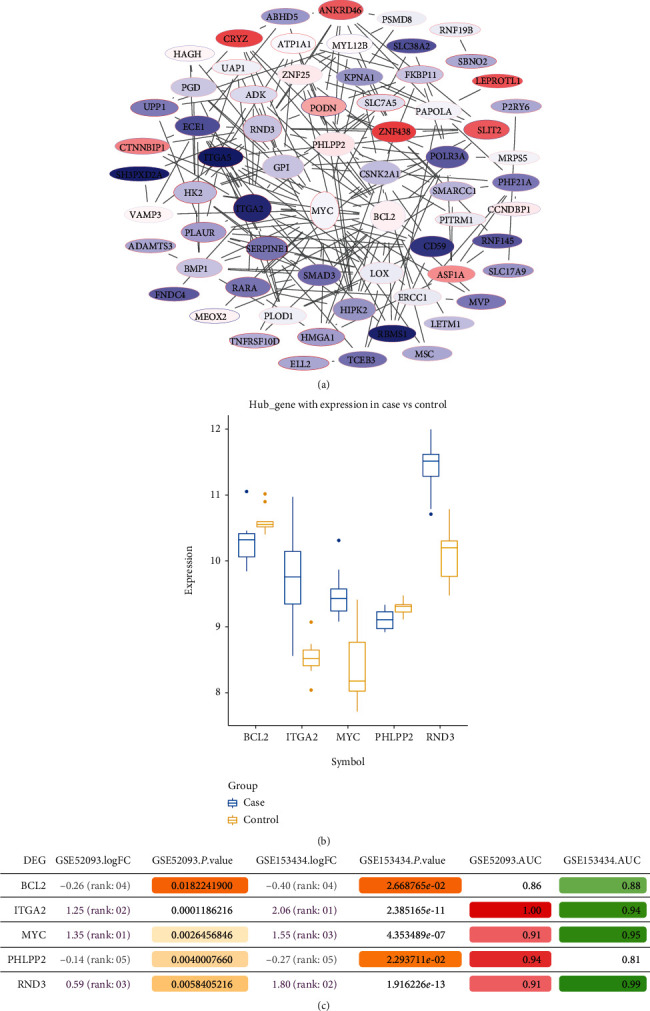
Identification of hub genes for acute type A aortic dissection. (a) The PPI network of 77 methylation markers based on STRING database. (b) The expression of hub genes in GSE153434. (c) The AUC values of hub genes in two datasets. The darker orange color represents a smallerPvalue. The darker red color represents a greater AUC value compared to what the gene has in GSE52093. The darker green color represents a greater AUC value compared to what the gene has in GSE153434.
3.4. Immune Cell Infiltration Difference in ATAAD
Differences in immune cell infiltration were found in ATAAD patients compared with controls (Figure 5(a)). Th1 cells, B cells, T helper cells, T cells, DC, iDC, Tgd, eosinophils, and NK cells were significantly downregulated. The different directions of Th1 cells, Tgd, T cells, T helper cells, iDC, DC, and B cells were consistent in GSE52093, GSE98770, and GSE153434 (Figure 5(b)). These immune cells were clustered into four categories, and there was a positive or negative correlation between the cells (Figure 5(c)). In ATAAD, iDC and macrophages had the strongest positive correlation, while in the control group, iDC and neutrophils had the strongest positive correlation (Figure 5(d)). The correlation analysis results between immune cells and hub genes showed that Th2 cells had the strongest correlation with ITGA2, while NK cells and Th17 cells had the strongest correlation with BCL2 (Figure 5(e)).
Figure 5.
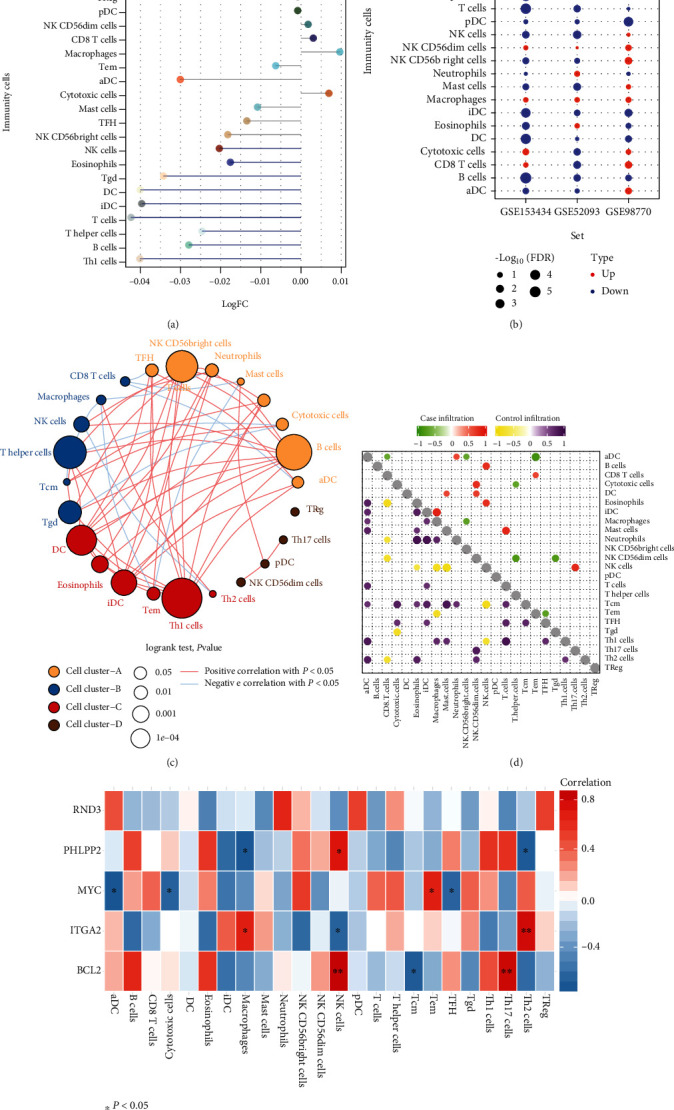
Differences in immune cell infiltration between acute type A aortic dissection and controls. (a) Immune cell infiltration differences between ATAAD and control. Blue columns represent significant differences. Blue lines represent significantly downregulated infiltration of immune cells. (b) Immune cell infiltration differences were validated in three datasets. Red node represents upregulation, and blue node represents downregulation. (c) Cluster diagram of differentially infiltrated immune cells. (d) Correlation of differentially infiltrated immune cells in ATAAD or control. (e) Correlation between differentially infiltrated immune cells and hub genes. Red represents positive correlation between immune cells, and blue line represents negative correlation. ∗P < 0.05; ∗∗P < 0.01.
4. Discussion
Repair of acute type A aortic dissection remains a challenge with high operative mortality [40]. As ATAAD is one of the most elusive and life-threatening vascular diseases, a better understanding of the molecular mechanisms of ATAAD is essential to improve clinical efficacy. In this study, genes with higher correlation with ATAAD were identified by comparing gene expression differences between ATAAD and controls. These genes were mainly associated with immune inflammation. The genes modified by methylation were screened as important genes to construct a PPI network, and five hub genes were identified. In addition, by comparing the difference of immune cell infiltration between ATAAD and control, we also similarly found that immune cells played an important regulatory role in the disease process [41].
Among the ATAAD-related biological functions we identified, T cell activation had been confirmed by other studies [42–44]. Different T cell subsets may play different roles in the development of ATAAD. Elevated white blood cell count is associated with poor prognosis in ATAAD [45, 46]. Inflammatory cells and cytokines, white blood cell count, and neutrophil count have been reported to be responsible for preoperative hypoxemia in ATAAD [47]. Increased inflammatory response is a key factor in promoting the occurrence and development of ATAAD [48]. High inflammatory biomarkers were observed in patients at onset, indicating that the inflammatory response started early in ATAAD [49]. More severe inflammation and oxidative stress reactions occur in obese ATAAD patients [50]. Inflammation and hypoxia are often interdependent [51]. Our findings also suggested that the HIF-1 signaling pathway was activated during ATAAD, thereby aggravating aortic dissection [1]. Therefore, we believe that inflammation and oxidative stress may play an important role in the process of ATAAD.
Notably, upregulated MYC, ITGA2, and RND3 and downregulated BCL2 and PHLPP2 were identified as hub genes of the PPI network. Studies had shown that MYC was indeed upregulated in ATAAD [52]. MYC signaling is involved in vascular smooth muscle cell (VSMC) dysfunction, vasoconstriction, and vascular remodeling in aortic dissection [53]. ITGA2 interacts with collagen in tumors, promotes cell migration, and promotes apoptosis-free resistance [54, 55]. Although there was no direct evidence that ITGA2 was associated with ATAAD, aortic disease was associated with collagen content or structure [56]. It suggested that ITGA2 may act on the development of ATAAD through collagen. In addition, ITGA3 and ITGA5 were identified as new biomarkers for the onset of acute aortic dissection [57]. RND3 played an important role in blocking cell cycle distribution, inhibiting cell growth, and inducing apoptosis and differentiation [58, 59]. Abnormal expression of RND3 may be the main cause of some cardiovascular diseases [60]. The BCL2 protein family influenced the apoptosis of vascular smooth muscle cells in human aortic dissection [61]. PHLPP2 had been reported as a therapeutic target for cancer and cardiovascular diseases [62, 63]. Although no relationship between PHLPP2 and ATAAD had been reported, our findings suggested that its downregulation may be a risk factor for ATAAD.
Most studies used microarray technology to compare diseased and normal aortic tissues and found some signs of ATAAD differentially expressed genes [16, 22]. However, the regulatory mechanisms of differentially expressed genes remain to be determined. The hub genes identified in this study were all modified by methylation. Alterations in gene methylation may mediate the involvement of vascular smooth muscle cells and inflammatory cells in the development of aortic dissection [24]. At present, there are relatively few studies on the regulation of methylation in ATAAD, and we believe that hub genes may be regulated by methylation and thus participate in the progress of ATAAD.
Like other studies, our study also had some limitations. First, these results were obtained only by bioinformatics analysis and were not confirmed by molecular experiments. In addition, although hub genes, potential methylation regulators, and related biological functions of ATAAD had been identified, there was still a long way to go to translate these findings into clinical applications.
It was worth emphasizing that comprehensive network analysis provided a new perspective to understand the molecular basis of ATAAD and promised to elucidate the complex relationship between DEGs in complex diseases. Hub genes were regulated by methylation and participated in the development of ATAAD through immune inflammation and oxidative stress responses. This study will help to identify new DNA methylation markers and improve the understanding and treatment level of ATAAD.
5. Conclusion
There were a large number of differentially expressed genes in ATAAD patients, which mainly regulated immune inflammation and oxidative stress functions. In particular, MYC, ITGA2, RND3, BCL2, and PHLPP2 were regulated by methylation in ATAAD. Differential expression of these genes may be associated with the progression of ATAAD, which may be a diagnostic biomarker and a new therapeutic target for ATAAD.
Acknowledgments
This work was funded by the National Key R&D Program of China (2019YFF0216303).
Data Availability
The data used in our study could be found in GSE52093 and GSE98770.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- 1.Lian G., Li X., Zhang L., et al. Macrophage metabolic reprogramming aggravates aortic dissection through the HIF1α-ADAM17 pathway. eBioMedicine. 2019;49:291–304. doi: 10.1016/j.ebiom.2019.09.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chiu P., Miller D. C. Evolution of surgical therapy for Stanford acute type A aortic dissection. Annals of cardiothoracic surgery. 2016;5(4):275–295. doi: 10.21037/acs.2016.05.05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Parve S., Ziganshin B. A., Elefteriades J. A. Overview of the current knowledge on etiology, natural history and treatment of aortic dissection. The Journal of Cardiovascular Surgery. 2017;58(2):238–251. doi: 10.23736/S0021-9509.17.09883-4. [DOI] [PubMed] [Google Scholar]
- 4.Jiang W. J., Ren W. H., Liu X. J., et al. Disruption of mechanical stress in extracellular matrix is related to Stanford type A aortic dissection through down-regulation of Yes-associated protein. Aging (Albany NY) 2016;8(9):1923–1939. doi: 10.18632/aging.101033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Moeller E., Nores M., Stamou S. C. Repair of acute type-A aortic dissection in the present era: outcomes and controversies. AORTA Journal. 2019;7(6):155–162. doi: 10.1055/s-0039-3401810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Coady M. A., Rizzo J. A., Goldstein L. J., Elefteriades J. A. Natural history, pathogenesis, and etiology of thoracic aortic aneurysms and dissections. Cardiology Clinics. 1999;17(4):615–635. doi: 10.1016/S0733-8651(05)70105-3. vii. [DOI] [PubMed] [Google Scholar]
- 7.Fukui T. Management of acute aortic dissection and thoracic aortic rupture. Journal of Intensive Care. 2018;6(1):p. 15. doi: 10.1186/s40560-018-0287-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Spittell P. C., Spittell J. A., Jr., Joyce J. W., et al. Clinical features and differential diagnosis of aortic dissection: experience with 236 cases (1980 through 1990) Mayo Clinic Proceedings. 1993;68(7):642–651. doi: 10.1016/S0025-6196(12)60599-0. [DOI] [PubMed] [Google Scholar]
- 9.Clough R. E., Nienaber C. A. Management of acute aortic syndrome. Nature Reviews. Cardiology. 2015;12(2):103–114. doi: 10.1038/nrcardio.2014.203. [DOI] [PubMed] [Google Scholar]
- 10.Nienaber C. A., Clough R. E., Sakalihasan N., et al. Aortic dissection. Nature Reviews. Disease Primers. 2016;2(1):p. 16053. doi: 10.1038/nrdp.2016.53. [DOI] [PubMed] [Google Scholar]
- 11.Hagan P. G., Nienaber C. A., Isselbacher E. M., et al. The International Registry of Acute Aortic Dissection (IRAD): new insights into an old disease. JAMA. 2000;283(7):897–903. doi: 10.1001/jama.283.7.897. [DOI] [PubMed] [Google Scholar]
- 12.Luo F., Zhou X. L., Li J. J., Hui R. T. Inflammatory response is associated with aortic dissection. Ageing Research Reviews. 2009;8(1):31–35. doi: 10.1016/j.arr.2008.08.001. [DOI] [PubMed] [Google Scholar]
- 13.Okita Y. Current surgical results of acute type A aortic dissection in Japan. Annals of cardiothoracic surgery. 2016;5(4):368–376. doi: 10.21037/acs.2016.06.02. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gu C., Shi X., Dai C., et al. RNA m6A modification in cancers: molecular mechanisms and potential clinical applications. The Innovation. 2020;1(3):p. 100066. doi: 10.1016/j.xinn.2020.100066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gu C., Chen C. Methylation in lung cancer: a brief review. Methods in Molecular Biology. 2020;2204:91–97. doi: 10.1007/978-1-0716-0904-0_8. [DOI] [PubMed] [Google Scholar]
- 16.Kimura N., Futamura K., Arakawa M., et al. Gene expression profiling of acute type A aortic dissection combined with in vitro assessment. European Journal of Cardio-Thoracic Surgery. 2017;52(4):810–817. doi: 10.1093/ejcts/ezx095. [DOI] [PubMed] [Google Scholar]
- 17.Kermani T. A., Warrington K. J., Crowson C. S., et al. Predictors of dissection in aortic aneurysms from giant cell arteritis. Journal of Clinical Rheumatology. 2016;22(4):184–187. doi: 10.1097/RHU.0000000000000381. [DOI] [PubMed] [Google Scholar]
- 18.Tavora F., Jeudy J., Gocke C., Burke A. Takayasu aortitis with acute dissection and hemopericardium. Cardiovascular Pathology. 2005;14(6):320–323. doi: 10.1016/j.carpath.2005.08.006. [DOI] [PubMed] [Google Scholar]
- 19.Anzai A., Shimoda M., Endo J., et al. Adventitial CXCL1/G-CSF expression in response to acute aortic dissection triggers local neutrophil recruitment and activation leading to aortic rupture. Circulation Research. 2015;116(4):612–623. doi: 10.1161/CIRCRESAHA.116.304918. [DOI] [PubMed] [Google Scholar]
- 20.Hiratzka L. F., Bakris G. L., Beckman J. A., et al. 2010 ACCF/AHA/AATS/ACR/ASA/SCA/SCAI/SIR/STS/SVM guidelines for the diagnosis and management of patients with thoracic aortic disease: a report of the American College of Cardiology Foundation/American Heart Association Task Force on Practice Guidelines, American Association for Thoracic Surgery, American College of Radiology,American Stroke Association, Society of Cardiovascular Anesthesiologists, Society for Cardiovascular Angiography and Interventions, Society of Interventional Radiology, Society of Thoracic Surgeons,and Society for Vascular Medicine. Journal of the American College of Cardiology. 2010;55(14):e27–e129. doi: 10.1016/j.jacc.2010.02.015. [DOI] [PubMed] [Google Scholar]
- 21.Yang J., Zou S., Liao M., Qu L. Transcriptome sequencing revealed candidate genes relevant to mesenchymal stem cells’ role in aortic dissection patients. Molecular Medicine Reports. 2018;17(1):273–283. doi: 10.3892/mmr.2017.7851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Pan S., Wu D., Teschendorff A. E., et al. JAK2-centered interactome hotspot identified by an integrative network algorithm in acute Stanford type A aortic dissection. PLoS One. 2014;9(2, article e89406) doi: 10.1371/journal.pone.0089406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Boileau A., Lindsay M. E., Michel J. B., Devaux Y. Epigenetics in ascending thoracic aortic aneurysm and dissection. AORTA Journal. 2018;6(1):1–12. doi: 10.1055/s-0038-1639610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li N., Lin H., Zhou H., et al. Efficient detection of differentially methylated regions in the genome of patients with thoracic aortic dissection and association with MMP2 hypermethylation. Experimental and Therapeutic Medicine. 2020;20(2):1073–1081. doi: 10.3892/etm.2020.8753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Feltrin A. S., Tahira A. C., Simões S. N., Brentani H., Martins D. C., Jr. Assessment of complementarity of WGCNA and NERI results for identification of modules associated to schizophrenia spectrum disorders. PLoS One. 2019;14(1, article e0210431) doi: 10.1371/journal.pone.0210431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chaste P., Klei L., Sanders S. J., et al. A genome-wide association study of autism using the Simons Simplex Collection: does reducing phenotypic heterogeneity in autism increase genetic homogeneity? Biological Psychiatry. 2015;77(9):775–784. doi: 10.1016/j.biopsych.2014.09.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Liang W., Sun F., Zhao Y., Shan L., Lou H. Identification of susceptibility modules and genes for cardiovascular disease in diabetic patients using WGCNA analysis. Journal Diabetes Research. 2020;2020, article 4178639:1–11. doi: 10.1155/2020/4178639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ritchie M. E., Phipson B., Wu D. I., et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic acids research. 2015;43(7, article e47) doi: 10.1093/nar/gkv007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gu C., Shi X., Huang Z., et al. A comprehensive study of construction and analysis of competitive endogenous RNA networks in lung adenocarcinoma. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics. 2020;1868(8, article 140444) doi: 10.1016/j.bbapap.2020.140444. [DOI] [PubMed] [Google Scholar]
- 30.Gu C., Huang Z., Chen X., et al. TEAD4 promotes tumor development in patients with lung adenocarcinoma via ERK signaling pathway. Biochimica et Biophysica Acta - Molecular Basis of Disease. 2020;1866(12, article 165921) doi: 10.1016/j.bbadis.2020.165921. [DOI] [PubMed] [Google Scholar]
- 31.Langfelder P., Horvath S. WGCNA: an R package for weighted correlation network analysis. BMC Bioinformatics. 2008;9(1):p. 559. doi: 10.1186/1471-2105-9-559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yu G., Wang L. G., Han Y., He Q. Y. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS. 2012;16(5):284–287. doi: 10.1089/omi.2011.0118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gu C., Shi X., Dang X., et al. Identification of common genes and pathways in eight fibrosis diseases. Frontiers in Genetics. 2020;11, article 627396 doi: 10.3389/fgene.2020.627396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Tian Y., Morris T. J., Webster A. P., et al. ChAMP: updated methylation analysis pipeline for Illumina BeadChips. Bioinformatics. 2017;33(24):3982–3984. doi: 10.1093/bioinformatics/btx513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Liang S., Shi X., Yu C., et al. Identification of novel candidate genes in heterotaxy syndrome patients with congenital heart diseases by whole exome sequencing. Biochimica et Biophysica Acta - Molecular Basis of Disease. 2020;1866(12, article 165906) doi: 10.1016/j.bbadis.2020.165906. [DOI] [PubMed] [Google Scholar]
- 36.Shi X., Huang T., Wang J., et al. Next-generation sequencing identifies novel genes with rare variants in total anomalous pulmonary venous connection. eBioMedicine. 2018;38:217–227. doi: 10.1016/j.ebiom.2018.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Shi X., Cheng L., Jiao X. T., et al. Rare copy number variants identify novel genes in sporadic total anomalous pulmonary vein connection. Frontiers in Genetics. 2018;9:p. 559. doi: 10.3389/fgene.2018.00559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zhang L., Shi X., Gu C., et al. Identification of cell-to-cell interactions by ligand-receptor pairs in human fetal heart. Biochimica et Biophysica Acta - Molecular Basis of Disease. 2020;1866(12):p. 165917. doi: 10.1016/j.bbadis.2020.165917. [DOI] [PubMed] [Google Scholar]
- 39.Bindea G., Mlecnik B., Tosolini M., et al. Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. Immunity. 2013;39(4):782–795. doi: 10.1016/j.immuni.2013.10.003. [DOI] [PubMed] [Google Scholar]
- 40.Yang B., Norton E. L., Shih T., et al. Late outcomes of strategic arch resection in acute type A aortic dissection. The Journal of Thoracic and Cardiovascular Surgery. 2019;157(4):1313–1321.e2. doi: 10.1016/j.jtcvs.2018.10.139. e2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shen Y. H., LeMaire S. A. Molecular pathogenesis of genetic and sporadic aortic aneurysms and dissections. Current Problems in Surgery. 2017;54(3):95–155. doi: 10.1067/j.cpsurg.2017.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ye J., Wang Y., Wang Z., et al. Circulating Th1, Th2, Th9, Th17, Th22, and Treg levels in aortic dissection patients. Mediators of Inflammation. 2018;2018:10. doi: 10.1155/2018/5697149.5697149 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wu D., Choi J. C., Sameri A., et al. Inflammatory cell infiltrates in acute and chronic thoracic aortic dissection. Aorta. 2013;1(6):259–267. doi: 10.12945/j.aorta.2013.13-044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.He R., Guo D. C., Estrera A. L., et al. Characterization of the inflammatory and apoptotic cells in the aortas of patients with ascending thoracic aortic aneurysms and dissections. The Journal of Thoracic and Cardiovascular Surgery. 2006;131(3):671–678.e2. doi: 10.1016/j.jtcvs.2005.09.018. [DOI] [PubMed] [Google Scholar]
- 45.Suzuki K., Kimura N., Mieno M., et al. Factors related to white blood cell elevation in acute type A aortic dissection. PLoS One. 2020;15(2, article e0228954) doi: 10.1371/journal.pone.0228954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ma M., Shi J., Feng X., Wang J., Liu L., Wei X. The elevated admission white blood cell count relates to adverse surgical outcome of acute Stanford type A aortic dissection. Journal of Cardiothoracic Surgery. 2020;15(1):p. 48. doi: 10.1186/s13019-020-1078-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Duan X. Z., Xu Z. Y., Lu F. L., et al. Inflammation is related to preoperative hypoxemia in patients with acute Stanford type A aortic dissection. Journal of Thoracic Disease. 2018;10(3):1628–1634. doi: 10.21037/jtd.2018.03.48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Han L., Dai L., Zhao Y. F., et al. CD40L promotes development of acute aortic dissection via induction of inflammation and impairment of endothelial cell function. Aging (Albany NY) 2018;10(3):371–385. doi: 10.18632/aging.101394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Sbarouni E., Georgiadou P., Analitis A., Voudris V. High neutrophil to lymphocyte ratio in type A acute aortic dissection facilitates diagnosis and predicts worse outcome. Expert Review of Molecular Diagnostics. 2015;15(7):965–970. doi: 10.1586/14737159.2015.1042367. [DOI] [PubMed] [Google Scholar]
- 50.Wu Z., Wang Z., Wu H., et al. Obesity is a risk factor for preoperative hypoxemia in Stanford A acute aortic dissection. Medicine (Baltimore) 2020;99(11, article e19186) doi: 10.1097/MD.0000000000019186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Colgan S. P., Taylor C. T. Hypoxia: an alarm signal during intestinal inflammation. Nature Reviews. Gastroenterology & Hepatology. 2010;7(5):281–287. doi: 10.1038/nrgastro.2010.39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Weis-Müller B. T., Modlich O., Drobinskaya I., et al. Gene expression in acute Stanford type A dissection: a comparative microarray study. Journal of Translational Medicine. 2006;4(1):p. 29. doi: 10.1186/1479-5876-4-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Feng J., Ge S., Zhang L., Che H., Liang C. Aortic dissection is associated with reduced polycystin-1 expression, an abnormality that leads to increased ERK phosphorylation in vascular smooth muscle cells. European Journal of Histochemistry. 2016;60(4):p. 2711. doi: 10.4081/ejh.2016.2711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Huang Y. L., Liang C. Y., Ritz D., et al. Collagen-rich omentum is a premetastatic niche for integrin α2-mediated peritoneal metastasis. eLife. 2020;9 doi: 10.7554/eLife.59442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Yu J., Zhang C., Yu Q., Yu H., Zhang B. ADAR1 p110 enhances adhesion of tumor cells to extracellular matrix in hepatocellular carcinoma via up-regulating ITGA2 expression. Medical Science Monitor. 2019;25:1469–1479. doi: 10.12659/MSM.911944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Tsamis A., Krawiec J. T., Vorp D. A. Elastin and collagen fibre microstructure of the human aorta in ageing and disease: a review. J R Soc Interface. 2013;10(83):p. 20121004. doi: 10.1098/rsif.2012.1004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Xing L., Xue Y., Yang Y., et al. TMT-based quantitative proteomic analysis identification of integrin alpha 3 and integrin alpha 5 as novel biomarkers in pathogenesis of acute aortic dissection. BioMed Research International. 2020;2020:12. doi: 10.1155/2020/1068402.1068402 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Yue X., Yang X., Lin X., et al. Rnd3 haploinsufficient mice are predisposed to hemodynamic stress and develop apoptotic cardiomyopathy with heart failure. Cell Death & Disease. 2014;5(6, article e1284) doi: 10.1038/cddis.2014.235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Chiarugi P., Giannoni E. Anoikis: a necessary death program for anchorage-dependent cells. Biochemical Pharmacology. 2008;76(11):1352–1364. doi: 10.1016/j.bcp.2008.07.023. [DOI] [PubMed] [Google Scholar]
- 60.Jie W., Andrade K. C., Lin X., Yang X., Yue X., Chang J. Pathophysiological functions of Rnd3/RhoE. Comprehensive Physiology. 2015;6(1):169–186. doi: 10.1002/cphy.c150018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Durdu S., Deniz G. C., Balci D., et al. Apoptotic vascular smooth muscle cell depletion via BCL2 family of proteins in human ascending aortic aneurysm and dissection. Cardiovascular Therapeutics. 2012;30(6):316. doi: 10.1111/1755-5922.12007. [DOI] [PubMed] [Google Scholar]
- 62.Wang H., Gu R., Tian F., et al. PHLPP2 as a novel metastatic and prognostic biomarker in non-small cell lung cancer patients. Thoracic cancer. 2019;10(11):2124–2132. doi: 10.1111/1759-7714.13196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Yeh S. T., Zambrano C. M., Koch W. J., Purcell N. H. PH domain leucine-rich repeat protein phosphatase 2 (PHLPP2) regulates G-protein-coupled receptor kinase 5 (GRK5)-induced cardiac hypertrophy in vitro. The Journal of Biological Chemistry. 2018;293(21):8056–8064. doi: 10.1074/jbc.M117.809913. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data used in our study could be found in GSE52093 and GSE98770.