

Abstract
Human milk represents a cornerstone for growth and development of infants, with extensive array of benefits. In addition to exceptionally nutritive and bioactive components, human milk encompasses a complex community of signature bacteria that helps establish infant gut microbiota, contributes to maturation of infant immune system, and competitively interferes with pathogens. Among bioactive constituents of milk, human milk oligosaccharides (HMOs) are particularly significant. These are non-digestible carbohydrates forming the third largest solid component in human milk. Valuable effects of HMOs include shaping intestinal microbiota, imparting antimicrobial effects, developing intestinal barrier, and modulating immune response. Moreover, recent investigations suggest correlations between HMOs and milk microbiota, with complex links possibly existing with environmental factors, genetics, geographical location, and other factors. In this review, and from a physiological and health implications perspective, milk benefits for newborns and mothers are highlighted. From a microbiological perspective, a focused insight into milk microbiota, including origins, diversity, benefits, and effect of maternal diet is presented. From a metabolic perspective, biochemical, physiological, and genetic significance of HMOs, and their probable relations to milk microbiota, are addressed. Ongoing research into mechanistic processes through which the rich biological assets of milk promote development, shaping of microbiota, and immunity is tackled.
Keywords: human milk, breastfeeding, human milk benefits, milk microbiota, human milk oligosaccharides, maternal diet
1. Introduction to Human Milk and Importance of Breastfeeding
Over the past decades, human breast milk has been widely agreed upon as being the normal and optimal dietary start for infants, with unparalleled biological effects driven by the combined action of its nutritional and bioactive components [1]. The positive effects of breastfeeding on infants from the nutritional, physiological, and developmental viewpoints have been confirmed [2]. Worldwide, health authorities have recognized the value and benefits of breastfeeding [3]. The World Health Organization (WHO) and the United Nations Children’s Fund (UNICEF) recommend the initiation of breastfeeding within the first hour of birth and advise on exclusive breastfeeding for the first six months of life, without any other food or liquids, including water. Although the introduction of complementary food is safe starting at six months, the WHO and UNICEF recommend to pursue breastfeeding for up to two years [4]. In the breastfeeding week 2020 message, under the theme “support breastfeeding for a healthier planet”, the WHO called for protection, promotion, and support of women’s access to skilled breastfeeding, in line with the multifaceted benefits of this sustainable, biological food system [5]. Likewise, the American Academy of Pediatrics (AAP) continuously reinforces breastfeeding and mother’s milk as the normal standards for infant feeding and nutrition. It recommends exclusive breastfeeding for six months, followed by continuation of breastfeeding as complementary foods are introduced, with possible extension of breastfeeding for one year of age or beyond, as desired [6].
Human milk holds a plethora of health benefits, both on the short- and the long-term. The relationship between breastfeeding and infant’s health is based on its nutritional and non-nutritional components that have diverse roles [7]. Human milk contains the precise mix of nutrients, including carbohydrates, proteins, fats, vitamins, minerals as well as water, that together secure a dynamic composition, well developed to fulfil needs of the infant growth [8]. The main nutrient constituents of milk are shown in Table 1 [8,9,10,11]. Further to nutrients, human milk contains many bioactive constituents like immunoglobulins, growth factors, microRNAs, and human milk oligosaccharides (HMOs). These form a complex system linking the mother’s lifestyle, such as diet and gut microbiota, with outcomes on infant growth, gut microbiota, immunity, and other developmental features [12]. In the infantile gut, a human milk-directed microbiota develops, forming initial doses for seeding the mature gut microbiome. Additionally, the high concentration and structural diversity of HMOs that reach the infant colon, initiate a series of health effects [13]. To this end, this review article presents up-to-date information about human milk benefits, human milk microbiota, and HMOs, and revises possible associations between microbial milk profiles and both maternal diet and HMOs.
Table 1.
| Macronutrients | |
| Protein | 0.9 g/dL |
| Fat | 3.5 g/dL |
| Carbohydrates (mainly glucose) | 6.7 g/dL |
| Minerals | |
| Calcium | 200–250 mg/L |
| Magnesium | 30–35 mg/L |
| Phosphorus | 120–140 mg/L |
| Sodium | 120–250 mg/L |
| Potassium | 400–550 mg/L |
| Iron | 0.3–0.9 mg/L |
| Fluoride | 4–15 µg/L |
| Fat-Soluble Vitamins | |
| Vitamin A | 0.3–0.6 mg/L |
| Vitamin D | 15 IU/day (in exclusively breastfed infants) or 0.33 µg/L |
| Vitamin E | 3–8 mg/L |
| Vitamin K | 2–3 µg/L |
| Water-Soluble Vitamins | |
| Ascorbic acid | 100 mg/L |
| Thiamine (vitamin B1) | 200 µg/L |
| Riboflavin (vitamin B2) | 0.35–0.39 mg/L |
| Niacin | 1.8–6 mg/L |
| Vitamin B6 | 0.09–0.31 mg/L |
| Folate | 80–140 µg/L |
| Vitamin B12 | 0.5–1 µg/L |
2. Human Milk Benefits
A priceless natural regimen presented by a mother to her newborn, human milk offers an extensive array of health benefits [14]. Milk from mothers whose diet is sufficient and well-balanced supplies all the necessary nutrients suitable for ideal growth (except vitamin D, whose intake should be 400 IU/day for all breastfed infants) [15]. Any amount of breastfeeding is better than none, and superior benefits accumulate with increased duration [3]. In addition to its supreme nutritive value, current scientific evidence about breastfeeding supports many beneficial effects, not only for infants who are breastfed on both short- and long-term, but also for mothers who breastfeed [16].
2.1. Short-Term Benefits for Infants
Breastfeeding possesses obvious short-term benefits for infant health, mostly in decreasing mortality and morbidity resulting from infectious diseases, such as gastrointestinal and respiratory tract infections, as well as otitis media, with a level of evidence described as convincing [15].
The protection conferred by breastfeeding against diarrhea is certainly one of the most consistent findings in the epidemiological literature [3,15,17,18,19]. A systematic review published by the WHO in 2013 suggests that breastfeeding substantially protects against morbidity/mortality from diarrhea, with a reduction of 50% in morbidity and 80–90% in mortality and hospital admissions, compared to infants with less or no breastfeeding. Such protection was higher among infants who were exclusively breastfed in the first six months of life and was provided by more intense breastfeeding. These robust results were observed across high and low-income settings [17]. An integrative review by Santos and Colleagues [20] in 2015 also replicated similar results, confirming the importance of breast feeding in the prevention of diarrhea in children under six months, especially among those who were exclusively breastfed. Such studies reinforce the recommendations for exclusive breastfeeding during the first six months of life as a key intervention for infant survival. The protective effect of breastfeeding against gastrointestinal infections has been attributed to the positive role of the microbial and immunological milk constituents and to the lack of contamination from baby bottles [19].
In addition to prevention of diarrhea, it is well documented that breastfeeding is associated with a significantly lower risk for lower respiratory tract infections and otitis media, compared with non-breastfed children [21]. Breast milk is associated with around 30% reduction in morbidity, 50% in hospital admissions, and 60% in mortality from these infections, suggesting that breastfeeding affects not only the incidence of these infections, but also their severity [17]. A systematic literature review and meta-analysis showed that suboptimal breastfeeding increased the risk of morbidity and mortality from pneumonia across different infant age groups. In particular, pneumonia mortality was higher among non-breastfed compared to exclusively breastfed infants 0–5 months of age, and among non-breastfed compared to breastfed infants and young children 6–23 months of age [22]. In a study from UK, higher risk of respiratory infection was found among infants exclusively breastfed for 4–6 months, but who stopped breastfeeding by six months, adding to the evidence of continued milk benefits after six months [23]. Furthermore, evidence shows that breastfeeding protects against acute otitis media until two years of age, but protection is greater for exclusive breastfeeding and breastfeeding of longer duration [24]. In a recent cohort analysis from Denmark in 2020, involving 815 mother-infant pairs, a strong association between breastfeeding and hospitalizations due to lower respiratory tract infection was found particularly in the first year of life [25]. In an interventional study to promote successful breastfeeding, Zivich and Colleagues showed that breastfeeding reduced incidence of mild as well as severe episodes of both respiratory infection and diarrhea in the first six months of life [26].
Recently, with the upsurge of Coronavirus Disease 2019 (COVID-19), caused by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), recommendations about breastfeeding in infected mothers were endorsed [27]. Breastfeeding in postpartum women with SARS-CoV-2 is recommended by the WHO [28] and by the US Centers for Disease Control and Prevention (CDC) [29] for the newborn as long as health of the mother and newborn allow it. This belongs to the same context of the protective effect of mother’s milk against infections and is built on the general evidence that benefits of breastfeeding substantially outweigh the potential risks for SARS-CoV-2 transmission. Appropriate respiratory hygiene measures for infected mothers always have to be considered with direct breastfeeding [30].
Apart from infection, the American Academy of Family Physicians reports reduced incidence of atopic dermatitis (eczema) in breastfed infants [3]. Although there is evidence that exclusive breastfeeding for 3 to 4 months reduces incidence of atopic dermatitis in the first two years of life, long-term advantages for exclusive breastfeeding for prevention of atopic skin disease are not clear. Furthermore, there are no definite conclusions regarding the role of breastfeeding in preventing nor postponing the onset of specific types of food allergies in newborns [31]. In a nationwide cohort studied in Japan in 2020, breastfeeding had prophylactic effects on food allergy only among high-risk children (defined as having positive family history and eczema presentation prior to one year) with infantile eczema, whereas prolonged breastfeeding may increase the risk of food allergy [32].
Another possible benefit of human milk is being a protective factor against sudden infant death syndrome, which is the leading death cause in the United States in infants of ages 1 month to 1 year [33]. Several literature reviews have documented that sudden infant death syndrome risk decreases with breastfeeding [34,35,36], and the effect is stronger when breastfeeding is prolonged [37]. Such positive effect of breastfeeding has been deliberated in physiologic sleep studies, which have revealed that breastfed infants have lower arousal thresholds than their formula-fed counterparts, probably providing a survival mechanism for protection against this condition [38,39].
2.2. Long-Term Benefits for Infants
In addition to growth and infection prevention on the short-term, research into human milk also indicates that it may influence noncommunicable diseases or conditions later in life, perhaps offering a lifetime of benefits [40]. For instance, the relationship between breastfeeding and obesity prevention has long been sought by researchers. According to meta-analysis, breastfeeding appears to a significantly protect against obesity in childhood, although evidence on this role is still controversial, and the underlying mechanisms are still unclear [41,42]. Recently, in a prospective six-year study from Spain that followed a cohort of infants from birth up to six years of age, full breastfeeding was associated with a significant decrease in obesity. The delay in introduction of bottle feeding showed a probable protective effect against obesity at 6 years of age [43]. In trying to explain the role of breastfeeding in prevention of obesity, epidemiological data illustrate that breastfed children have healthier dietary patterns compared to formula-fed children, probably reducing their risk of obesity. These healthier patterns are, in part, caused by early opportunities for flavor learning afforded by breastfeeding. In particular, the flavors of the mother’s diet are transmitted to the infant via breastmilk, providing an additional reinforcement and flavor learning upon repeated exposure to a wide variety of flavors. The flavors experienced by the infant modulate later food preferences, acceptance of solid food, and culture onto which the infant is weaned [44]. Plus, breastfeeding favors the establishment of diverse microbial communities in infant’s gut that play a crucial role in preventing obesity via their various mechanisms of harvesting energy from food, affecting patterns of fatty acid oxidation, modulating bile acid metabolism, and influencing satiety and lipogenesis. It is still debatable to define which microbial category is responsible for obesity, and studies in this regard show discrepancies [45].
Studies linking human milk to protection against gastrointestinal tract diseases are also numerous. For the past two decades, human milk have been associated with reduced incidence in necrotizing enterocolitis in preterm infants [46,47]. Such infants’ susceptibility to enterocolitis stems from their gastrointestinal and immune system immaturity. It is thought that an exclusive human milk diet, consisting of mother’s own milk and/or donor human milk alone or fortified with a human milk-based fortifier, compensates for these immature systems by many pathways like stimulating intestinal motility, composed from lowering gastric pH, decreasing epithelial penetrability, and altering the composition of the gut microbiota [48]. In a multicenter study in a cohort of over 1000 patients with Crohn’s disease, a history of breastfeeding was inversely associated with complicated pediatric symptoms [49]. The evidence of the link between Crohn’s disease prevention and breastfeeding, however, is described as insufficient, and more reliable data are needed [15]. Similarly, limited evidence exists for the role of breastfeeding in reducing incidence of celiac disease, and further investigation is needed to indicate whether such protection only delays the onset of the disease or if it can provide permanent protection [50].
Metabolic disorders and tumors also had their share in research trying to investigate late benefits of breastfeeding. In a meta-analysis, Horta and Colleagues [40] found that breastfeeding decreased the odds of type 2 diabetes mellitus and based on high-quality studies, but no associations were found for total cholesterol or blood pressure. Breast milk has long-chain polyunsaturated fatty acids (LCPUFAs), whose supplementation is associated with a reduction in blood pressure. In addition, LCPUFAs would induce early changes in skeletal muscle which can be protective against insulin resistance and type 2 diabetes [51]. As far as cancer is concerned, and with leukemia, which accounts for 30% of childhood cancers, a meta-analysis showed that breastfeeding for six months or longer may help reduce the incidence of childhood leukemia by up to 19% [52]. Furthermore, a pediatric oncology team assessed relation between breastfeeding and cancer in a group of 300 children with cancer (leukemia, lymphoma, and solid organ cancers including brain, bone, liver, kidney, and others), and an approximately equal number of age- and sex-matched controls. Breastfeeding duration of the control group was found to be significantly longer than the patient group [53].
Perhaps the possible influence of breastfeeding on cognitive development is a subject that has not ceased to provoke considerable scientific debate. Despite substantial disparity of reported results, a positive correlation between breastfeeding and the levels of intelligence and cognitive development in children exists, although with a number of confounders like maternal age, education, socioeconomic status, and IQ [15,21]. In general, children who are breastfed for longer than six months are known to show better cognitive outcomes and a lower risk to develop attention-deficit/hyperactivity disorder [54]. Researchers from McGill University, and based on the largest randomized, interventional trial ever conducted in the field of human milk benefits, provide robust evidence that prolonged, exclusive breastfeeding enhances children’s cognitive development up to age of 6.5 years [55]. However, in a study from UK analyzing children till age of nine years breastfeeding was not associated with IQ advantage nor difference in neurological soft signs. The higher IQ associated with breastfeeding was probably accounted for by some confounding maternal and socio-economic factors [56]. The actual mechanisms linking improved cognitive development with breastfeeding remain unknown; however, it is suggested that human milk LCPUFAs are incorporated in relatively large amounts during early growth of the brain and the retina [57], and more specifically, the white matter of the brain [21]. Some of these acids, such docosahexaenoic acid (DHA) and arachidonic acid (ARA), present in human milk, may play a role in growth and development of eye, brain and nerve; these acids also exhibit potent biological activity that modulates various cellular and tissue processes [58,59]. Interestingly, and also in the field of neurologic development, a recent meta-analysis in 2020 concluded that breastfeeding for 12–24 months was associated with a significant decrease in autism spectrum disorder risk [60]. Furthermore, studies show that breastfed infants, as compared to bottle-fed infants, show higher vigor, including social approach, activity and the intensity of reaction [61]. The duration of breastfeeding was shown to correlate negatively with parent-reported antisocial conduct and aggressive behavior in children aged 4 to 11 years [62]. These and other findings continue to verify that human milk is not simply a meal at the breast, but rather a biological fluid with significant and far-reaching effects into child’s health, psychology, and social behavior.
It is worth noting that the public health significance of breastfeeding originates from a big pool of studies that are mostly observational. Large interventional or randomized controlled trials on breastfeeding are usually hindered by ethical issues. Therefore, the evidence for short-term outcomes mentioned above, such as reduced rates of infection, can be classified as strong. However, the longer term effects are probably less firm, due to difficulty in completely accounting for confounding factors [16].
2.3. Maternal Benefits
While the nutritional and physical health benefits conveyed by human milk are well established for infants, accumulating research reveals that breastfeeding mothers benefit as well. As an evidence of such profits, it was reported that mothers with premature weaning have health risks of breast and ovarian cancers, hypertension, hyperlipidemia, diabetes mellitus, myocardial infarction, and obesity, with these risks being highest in those who do not breastfeed at all [63].
Breastfeeding reduces overall risk of breast cancer [64]. In a meta-analysis, exclusive breastfeeding among parous women was found to reduce the risk of breast cancer compared with parous women who do not exclusively breastfeed [65]. Furthermore, mothers positive for the BRCA1 mutation and breastfeeding for at least one year have 37% lower breast cancer risk compared to carriers of the mutation who do not breastfeed [66]. The reduced risk of breast cancer is most evident in postmenopausal women and is directly proportional to the duration of lactation. The protective effect of breastfeeding is attributed to differentiation of breast cells, decreased number of ovulatory cycles, and estrogen excretion through milk, reducing overall exposure to this hormone [67]. In addition to breast cancer, evidence suggests that breastfeeding reduces risk of ovarian cancer [63]. Modugno and Colleagues [68] concluded that breastfeeding for as few as three months was associated with reduced risk of ovarian cancer, and the association was long-lived. The probable mechanism of breastfeeding in ovarian cancer protection may be due to absence of ovulation. Monthly ovulation might increase the odds of genetic mutations, and ovarian overstimulation by elevated gonadotropins may trigger hyperproliferation, with possible malignant transformation [69].
Prolonged breastfeeding has beneficial effects in reducing the long-term risk of coronary artery disease. Parous women who breastfeed for five or more months in at least one pregnancy seem to be at decreased risk of coronary artery disease, whereas parous women who either did not breastfeed at all or discontinued breastfeeding early seem to be at elevated risk [70]. The European Society of Cardiology concluded that breastfeeding women had a lower risk of coronary artery disease later in life compared to those who do not breastfeed [71]. Data from studies in both animals and humans suggest that lactation may alter maternal sugar and lipid homeostasis and may exert effects on blood pressure regulation. This might have significant effects on lipid homeostasis, with lower triglyceride and higher HDL cholesterol levels, possibly reducing risks of heart diseases [72]. Breastfeeding also affects maternal risks of hypertension and diabetes mellitus [63,64,73]. Park and Choi [74] studied a population of over 3000 nonsmoking postmenopausal women aged 50 years or above, and showed that more children breastfed and longer breastfeeding duration were associated with lower risk of hypertension. A systematic review and meta-analysis determined that variable breastfeeding durations have different protective effects against maternal hypertension, especially if continued beyond 12 months [75]. Longer duration of breastfeeding, whether full or partial, is associated with lower maternal risk of hypertension and heart disease irrespective of pre-pregnancy body mass index and abdominal adiposity seven years after delivery [76]. Further, mothers who breastfeed have less visceral obesity and smaller waist circumference, probably accounting for a lower maternal risk of type 2 diabetes mellitus [77], and this was recently proved in a prospective cohort of women followed for 14.2 years [78]. Few common mechanisms of relationships between breastfeeding and maternal risks of hypertension and diabetes were suggested. For example, maternal metabolism (e.g., fat accumulation and insulin resistance) may be reversed by breastfeeding, which decreases both diseases risk. During pregnancy, visceral fat accumulates, insulin resistance rises, and lipid and triglyceride levels increase. These changes appear to retune more rapidly, and more entirely, with lactation—the so called “reset hypothesis” [79]. Another theory is that oxytocin, needed in glucose homeostasis and the release of which is stimulated by breastfeeding, may be associated with decreased risk of type 2 diabetes mellitus [80]. It is worth mentioning that milk production, which consumes about 500 kcal per day for an infant who is exclusively breastfed, reduces maternal obesity later in life [81].
Apart from cardiovascular health, breastfeeding affects psychological well-being. Breastfeeding mothers report less anxiety, less negative mood, and less stress, as well as increased sleep duration and reduced sleep disturbances when compared to formula-feeding mothers [82,83]. On another note, studies on post-partum depression demonstrate that breastfeeding may protect mothers from this disorder, and researchers have strived to explain the biological processes that explain this protection. For example, lactation attenuates neuro-endocrine responses to stress, and this may be related to fewer post-partum depressive symptoms [84,85]. Moreover, early breastfeeding cessation was linked to higher risk of post-partum depression [86]. It is the psychological pressure to exclusively breastfeed that contributes to postpartum depression symptoms in mothers unable to achieve their breastfeeding intentions [87,88]. In a prospective follow-up for eight weeks postpartum, mothers with breastfeeding problems (including mastitis, nipple pain, need for frequent expressing of milk, or over-supply or under-supply of milk) showed poor mental health [89].
With all the aforementioned benefits of human milk on infant and maternal wellness, breastfeeding continues to form a cornerstone for prevention of many short- and long-term health risks. It is a real investment in health, rather than a mere lifestyle decision. The promotion of human milk as a highly active biological milieu is substantially imperative, and further research into its numerous potential benefits is always appealing.
Figure 1 summarizes data from Section 2.1, Section 2.2 and Section 2.3 of this review pertaining to benefits of human milk and breastfeeding.
Figure 1.

A summarized description of the benefits of breastfeeding and effects of human milk on the early and long-term infant health, as well as benefits for the mother.
3. A Focused Insight into Human Milk Microbiota
In addition to its diverse nutritional and health benefits, human milk is characterized by a rich microbiota which constitutes a source for the infantile gut microbiota [90]. Except for initial observations of occurrence of microbes in milk in the 1950s, while studying the possibility of transmission of infections to infants via breastfeeding [91], human milk was thought of as a sterile fluid [92]. Around the beginning of the twenty-first century, a study in Spain by Martin and Colleagues [93] demonstrated, for the first time, the presence of the lactic acid bacterium, Lactobacillus gasseri, from milk and stools of mother–infant pairs, indicating that breast milk formed a potential source for infant microbiota. The non-sterility of breast milk has, ever since this initial observation, started to become extensively researched, and regarded as a protective factor arising from the wide range of microbes, in addition to the universally acknowledged nutritional role [94].
3.1. The Origin of Human Milk Microbiota
The origin of microbial populations in human milk is not completely understood and remains debatable. However, milk microbial populations have been proposed to originate endogenously, from the maternal digestive system through a complex pathway involving immune cells, or from the mother’s skin or infant’s mouth [95], or from the breast tissue itself [92]. It is plausible that more than one pathway contribute to the bacterial contents of milk [96].
3.1.1. The Entero-Mammary Pathway
The discovery of anaerobic species, such as bifidobacteria, in human milk suggested the transfer of microbiota from the mother’s gut to milk, especially since these anaerobes are not commonly cultivated from breast skin swabs [90]. A physiological translocation mechanism is thought to carry maternal intestinal microbiota to the lactating mammary gland. This pathway is supported by the presence of bacterial communities in colostrum, collected even before first infant suckling occurs [97]. The unique process of translocation is aided by physiological and hormonal variations during late pregnancy and increased permeability of the intestinal epithelial lining [98], and is also immunologically mediated. Such active migration of microbes involves mononuclear cells, dendritic cells, and CD18 cells, delivering nonpathogenic bacteria from the intestine to the lactating mammary gland [99]. Dendritic cells are capable of penetrating the gut epithelium by loosening the tight junctions between intestinal epithelial cells and taking up bacteria from the gut lumen. Bacteria are then transported by macrophages to mesenteric lymph nodes and finally to the mammary gland [98,100].
3.1.2. Retrograde Origin
Ultrasound imaging studies of nursing mothers have proved retrograde back flow of milk from the infant oral cavity due to infant suckling, suggesting a backward movement that transfers bacteria from infant’s mouth into the mother’s mammary gland [101]. This theory of milk microbiota origin is supported by similarities between the infant oral microbiota and human milk microbiota. For instance, Streptococcus, an abundant bacterial genus of breast milk, also dominates the salivary bacteria. Nevertheless, whether both bacterial communities share the same species and strains of Streptococcus remains to be determined [92,102].
3.1.3. Transfer from Maternal Skin
This is a traditionally acceptable model of human milk microbiota origin, supporting that milk microbiota can result from contamination from maternal skin during the process of infant suckling, or a seeding process of milk from maternal skin. This theory is based on the similarities among the adult skin microbiota and milk microbiota, especially among the genera Staphylococcus, Corynebacterium, and Propionibacterium [103,104].
3.1.4. Mammary Tissue Origin
In 2014, Urbaniak and Colleagues [105] showed, using culture and 16S rRNA gene sequencing, the presence of various genera of bacteria in breast tissue of Canadian and Irish women aged 18–90, with or without history of lactation, and without signs or symptoms of infection. Other investigations also showed that the breast tissue itself has an established microbiota, which is quite different from that of breast skin tissue and breast skin swabs [106]. Investigations on nipple aspirate fluid collected aseptically by application of negative pressure on the nipple, showed presence of bacterial DNA, probably from breast ducts [107]. It is probable that the existence of six to eight ductal openings on the surface of the human nipple permits microbes environmental microbes to access the ductal system of the breast [108]. It is also likely that such microbiota may influence the one present in human milk [92].
In summary, regardless of the relative contribution of the above four sources to microbes in milk, it has become generally acceptable that milk microbiota is not a secondary contamination, but rather a discrete microbiota, which is distinct from others in both the mother and the infant.
3.2. Types of Microbes in Human Milk
Both the early, culture-based methods, as well as the new, genomic-based and sophisticated techniques, such as next-generation sequencing, have been applied to analyze human milk microbiota and embrace its diversity [109]. Breast milk is a niche to several hundreds of bacterial species, and harbors about 1000 colony-forming units of bacteria/mL [110]. Following initial exposure to microbes upon birth, breast milk is the next immediate important source of various microbes to seed the infant’s gut [103]. As an evidence of this vertical transfer from milk microbiota to infant intestine, it was found that infants who primarily breastfeed during the first month of life, share 28% of their stool microbes with microbes of their mother’s milk. The number of shared microbes rises with the amount of daily breast milk intake in a dose-related manner, and microbes in the infant gut most resemble those from their own mother. The types and proportions of different microbes in human milk exhibit interindividual variation [103,111]. The major types are presented below.
3.2.1. Bacteria in Human Milk
A very recent systematic review in 2019 determined the bacterial repertoire of human milk, showing the presence of about 820 species, mainly belonging to Gram-positive Firmicutes and Gram-negative Proteobacteria [112]. Firmicutes include members of the genera Staphylococcus, Streptococcus, Lactobacillus, Bifidobacterium, Enterococcus, Veillonella, Gemella, Clostridium, and others. On the other hand, Proteobacteria include Escherichia, Pseudomonas, Enterobacter, Serratia, Ralstonia, Sphingomonas, Bradyrhizobium, and others. Additionally, milk contains Actinobacteria, like Actinomyces, Corynebacterium, and Propionibacterium, Fusobacteria such as Leptotrichia, as well as Bacteroidetes, such as Prevotella [113]. Since 2011, scientific consensus has been molded towards the existence of a “core” bacteriome in human milk, consisting of the following nine genera: Staphylococcus, Streptococcus, Serratia, Pseudomonas, Corynebacterium, Ralstonia, Propionibacterium, Sphingomonas, and Bradyrhizobium. These represent about half of the microbial milk community, although their abundance might diverge among milk [87,102]. In terms of relative proportions, Streptococcus and Staphylococcus species are the most common, and this is universally true, irrespective of variability in geographic location or analytical methods of milk analysis, whether culture or molecular based. They were followed by Bifidobacterium, Lactobacillus, Propionibacterium, Enterococcus, and members of the Enterobacteriaceae family [114]. During breastfeeding course, the overall bacterial diversity decreases compared with colostrum and transition milk. Moreover, the composition of the bacteria shifts from skin and enteric organisms such as Staphylococcus and Streptococcus, to infant mouth and skin organisms, such as Veillonella, Leptotrichia, and Prevotella, which increase as lactation progresses and mature milk is secreted [115].
3.2.2. Fungi in Human Milk
Most studies of human milk microbiome have overlooked fungi, and they have not been thoroughly assessed. In a cohort of Australian women, Candida was detected in breast milk in both culture and PCR-based methods [116]. Furthermore, a metagenomic analysis of milk samples showed not only fungal-related reads of DNA, but reads related to Archea and protozoa as well [117]. In 2020, a cohort of 271 samples of human milk revealed, through DNA sequencing, the presence of fungi in over 20% of samples, dominated by the genera Candida, Alternaria, and Rhodotorula [118]. In a study comparing transient and mature human milk samples, the most abundant fungal species in transient milk were Saccharomyces cerevisiae and Aspergillus glaucus. While A. glaucus was the second most common species in mature milk, S. cerevisiae disappeared, and Penicillium rubens appeared as the most abundant species, suggesting variable composition of milk mycobiome with maturity [119]. Further research is needed to establish the prevalence and role of fungi in human milk.
3.2.3. Human Milk Virome
Recent evidence is emerging to show that the neonatal virome is modulated by breastfeeding [120], and human milk contains viruses transmitted from the mother to the infant to colonize the infant gastrointestinal tract [121]. These viruses include eukaryotic viruses, bacteriophages, and other viral particles [122]. Specifically, bacteriophages form a majority, and they have the ability to kill bacteria or provide them with potentially beneficial genes [109].
3.3. Milk Microbiota Diversity and Associated Factors
A myriad of environmental, genetic, and immune factors personalizes a mother’s milk microbiome. Among different women, and in the same woman experiencing diverse physiological, hormonal, and pathological conditions, an inconsistency is expected among communities of milk microbiota [92].
Primarily, maternal factors contribute to differences in milk microbiota composition. Khodayar-Pardo and Colleagues [123] showed that Cesarean section births were associated with total bacterial concentrations in milk higher than vaginal delivery, and significantly higher levels of Streptococcus, but significantly lower levels of Bifidobacterium. This was associated in some studies with risk of immune-mediated and inflammatory disorders in infants born by Cesarean delivery [124]. Possibly, physiological stress or hormonal signals of delivery could influence the microbial transmission process to milk [115]. The difference in composition of milk microbiota among vaginal deliveries and Cesarean section, however, was not significant in other investigations [125], warranting further analysis. A fail-safe mechanism has been proposed, whereby the mother passes along her bacterial imprint irrespective of how the baby is born, eliminating major differences in milk microbiome composition with mode of delivery [125]. The differences in milk microbiota among cesarean and vaginal deliveries are, however, thought to be only short lived when reflected on infant gut microbiome. By one month, vaginal and cesarean section infants cannot be separated on the basis of composition of their microbiota [126]. Based on available evidence, it is difficult to draw clear-cut conclusions regarding milk microbiota composition and both mode of delivery and milk maturity.
Maternal body weight also influences milk microbiota. Milk from obese mothers was found to include a bacterial community of lower diversity compared with milk from normal-weight mothers, according to findings of a randomized controlled trial, but was associated with higher numbers of Staphylococcus and lower numbers of Bifidobacterium [115]. The link between gut microbiota and obesity may possibly be extended to milk, substantiating the conception that obesity influences the milk microbiota via its influence on the gut microbiota [115,127]. Indeed, maternal diet influences milk microbiota, and this is elaborated in Section 3.4.
In 2015, and also relating to maternal factors on milk microbiota composition, Olivares and Colleagues [128] showed that levels of bifidobacteria were reduced in milk of mothers with celiac disease compared to healthy mothers, and this could, theoretically, diminish the protective effects of breastfeeding on the child’s risk of developing celiac disease. Further, breast milk favored Clostridium leptum, Bifidobacterium longum, and Bifidobacterium breve gut colonization, and these were associated with the HLA type of low genetic risk for celiac disease [129]. In an analysis of intrapartum antibiotic administration on human milk microbiota, it was found that the species Bifidobacterium was uniquely found in breast milk samples of mothers who did not receive antibiotics. Such observation may be significant, since reduced fecal bifidobacteria in early infancy may be associated with higher risk of non-communicable diseases in childhood, like atopy and overweight [130].
On another note, maternal postnatal psychosocial distress may alter maternal gut microbiota, which, in turn, may affect bacteria present in milk. Browne and Colleagues [131] found no significant differences in the relative proportions of major bacterial genera between women with high and low levels of psychosocial distress. However, progressive and distinct decrease in Firmicutes, Proteobacteria, and Bacteroidetes at the phylum level, and increase in Acinetobacter, Flavobacterium, and Lactobacillus at the genus level, were evident in milk samples of women with low psychosocial distress. High maternal psychosocial distress was also related to significantly lower bacterial diversity in milk three months after delivery. These findings suggest a likely relationship between maternal psychosocial distress and milk microbiota, indicating that post-partum psychological symptoms may impact infantile development and health, through their influence on milk.
Milk bacterial profiles do not appear to significantly differ in relation to maternal age, infant gender, or race/ethnicity in a given geographical region [109], but do differ across geographic locations of Europe, Africa, and Asia. Across these continents, Kumar and Colleagues [132] analyzed milk samples of 80 women from 4 different countries (China, Finland, South Africa, and Spain). Spanish women had highest abundance of Bacteroidetes, whereas Chinese women had highest abundance of Actinobacteria. Women who had a cesarean section had higher amount of Proteobacteria as observed in the milk of the Spanish and South African women. Interestingly, and in the emerging field of human milk mycobiome, a core of four genera was shared across milk samples from the above four countries, consisting of Malassezia, Davidiella, Sistotrema, and Penicillium, which are also present in the infant gut, supporting potential role of breast milk in the initial seeding of fungal species in the infant gut [133]. In a very recent investigation from Dubai [134], the genus Hydrogenophaga (of the beta-Proteobacteria), previously reported in breast cancer tissue [106], was significantly higher in the breast milk of local women compared to expatriates living in Dubai. This finding may shed a light on possible influence of race and lifestyle on human milk microbiota, but the importance of such data needs to be further explored.
Other determining factors of human milk microbiota have been investigated. In one survey, of milk samples collected at different stages of lactation, Bifidobacterium spp. concentration was significantly higher in milk samples from term gestations compared to preterm ones, indicating that gestational age plays a role in structuring milk microbiota [123]. This was not reproducible in another study on Canadian women, whereby comparison of bacterial profiles between preterm and term births showed no statistically significant differences [125]. The number of lactobacilli- or bifidobacteria-positive samples was significantly lower in women treated with antibiotics during pregnancy or lactation [135]. This emphasizes that consideration needs to be given to the impact of drugs administered to the breastfeeding mother, not only on potential consequences for the infant health, but also in diverting the normal milk microbiota [109].
To summarize the above, a review about the multitude of factors influencing human milk composition was very nicely and recently elaborated by Zimmermann and Curtis [136] in 2020. This review of 44 studies recognized some evidence that gestational age, infant sex, delivery mode, parity, lactation stage, diet, body mass index, composition of breast milk, geographic location, HIV infection, and method of collection affect composition of the breast milk microbiota. However, many studies were small and findings possibly conflicting, indicating need for further research setting some benchmarks on the internal and external players affecting the nature of human milk microbiota.
3.4. The Effect of Maternal Diet on Human Milk Microbiota and on Intestinal Microbiota of Infants
In addition to the numerous physiological, medical and environmental factors described above, it was shown that the assembly of microbes in human milk is also affected by maternal diet [137]. The associations between numerous macro- and micronutrients in maternal diet and the milk microbiota have been investigated.
In fact, maternal diet affects the concentration of certain substances, like fatty acids, in milk. In other words, maternal nutrient intake may indirectly help shape the bacterial community membership in human milk simply because of its impact on milk nutrient composition. For example, Kumar and Colleagues [132] documented multiple associations between human milk fatty acid profiles and variations in milk microbiota. Monounsaturated fatty acids of milk were negatively associated with Proteobacteria, but positively associated with Lactobacillus genus.
Additionally, maternal nutrient intake affects gastrointestinal bacterial communities, which in turn, may become part of the milk microbiota via the entero-mammary pathway (Section 3.1.1). For example, Carrothers and Colleagues [138] provided initial evidence for associations between maternal nutrition and maternal fecal microbial community structure during lactation. Increased intake of pantothenic acid, riboflavin, vitamin B6, and vitamin B12 were related to increased relative abundance of Prevotella and decreased relative abundance of Bacteroides in the maternal gut. Intake of copper, magnesium, manganese, and molybdenum were positively associated with Firmicutes and negatively associated with Bacteroidetes. Overall, findings steadily suggest that high consumption of a more nutrient-rich and calorie-loaded diet was positively associated with relative abundance of Firmicutes. It was suggested that such maternal dietary factors that directly influence the maternal gastrointestinal bacterial community, might also indirectly affect the milk microbiota [137].
Correlations between maternal dietary intake and milk microbiota composition have been validated by several studies. In one investigation from Brazil, postpartum women went through a validated food frequency questionnaire that covered the whole pregnancy period, and correlations were obtained between their diet and their milk microbiota. A global, significant association with milk microbiota diversity was observed for the intake of ascorbic acid during pregnancy [139]. In light of ascorbic acid importance, higher maternal consumption of citrus fruits, as well as of vegetables and β-carotene during pregnancy were protective against eczema in the offspring, in line with the positive effects of ascorbic acid on the human immune system [140]. Furthermore, positive correlations were found between Bifidobacterium in the milk and intake of polyunsaturated and linoleic fatty acids during lactation [139]. This is consistent with the conversion of polyunsaturated fatty acids to conjugated linoleic acid and conjugated linolenic acid, known to favor Bifidobacterium growth [141].
In a study of 21 healthy breastfeeding women at Washington State University and University of Idaho, diet records, collected over 9 dietary assessments over six months postpartum, were correlated with milk microbiota. The results showed that relative abundances of several bacterial groups were associated with changes in maternal dietary intake. For instance, intake of saturated fatty acids and monounsaturated fatty acids were inversely associated with the relative abundance of Corynebacterium; total carbohydrates, disaccharides, and lactose were negatively associated with Firmicutes; and protein consumption was positively correlated with the increase in Gemella, a genus belonging to the Streptococcaceae family [137].
In an interesting animal model of diet followed for 31 months, it was demonstrated that alone, diet may modulate mammary gland microbiota. Mediterranean diet resulted in approximately 10-fold increase in mammary gland Lactobacillus abundance compared with mammary tissue from Western diet-fed animals [142]. Lactobacillus is often thought of as commensal organism and is commonly included in probiotic formulations, where it directly contributes to beneficial nutritional, physiological, microbiological, and immunological effects in the host [143]. Again, with Lactobacillus transferred from maternal gut to human milk, and then from milk to intestinal tract of infants, a dynamic pathway exists in which maternal diet will play a major role in determining the profile of infant gut microbiota [144].
Another recent study reinforcing the hypothesis that maternal diet affects milk microbiota found, at one-month postpartum, significant negative correlation between Streptococcus in maternal milk and the intake of unsaturated fatty acids and the abundance of oleic acid. This was explained by the fact that streptococci may be sensitive to the antibacterial effects of fatty acids, causing a direct inhibition [145]. The same group reported that Bifidobacterium, pivotal in development of the infant gut microbiome, was detected, at extremely low abundances, in milk samples [145]. This observation is in favor of the current view that maternal milk provides a microbial source for colonization of the infant intestine. However, the main effect of milk on the baby’s gut composition does not necessarily come from the milk bacteria only, but rather from other milk components, as well, that help to enrich specific bacterial groups. This could explain why bacterial breast milk composition and fecal microbiota composition are not exactly the same, but correlations do exist [146]. In light of benefits of Bifidobacterium, a trial investigated maternal supplementation of this bacterium as well as Lactobacillus to maternal diet four weeks prior to the due date and for three months after delivery. The intervention did not significantly affect overall composition of breast milk microbiota transferred to the infant during breastfeeding, analyzed by 16S rRNA sequencing, thus questioning the substantial effect of probiotic supplementation to maternal diet on microbiota composition of breast milk [147].
The currently available reports on maternal diet and microbiota of human milk are still few and reports on the role of nutrients in the metabolism of bacteria in human milk are much needed. Furthermore, the somehow incomplete knowledge regarding the availability of macro- and micronutrients in milk, which are directly useful for bacteria, further complicate data interpretation. The relationship between maternal diet and milk microbiota has much yet to be investigated, and in-depth studies of various nutritive components and their concentrations on microbiota of milk are warranted.
3.5. Beneficial Effects of Milk Microbiota on Infant Health
The commensal and beneficial microbes in human milk play a pioneer role in shaping infant health, and perhaps represent an example of how intimate contact with the microbial world is necessary for normal development in early life [148]. Human milk microbiota affects the establishment of the largest human microbial reservoir in the gastrointestinal tract; also, the reciprocal relation between milk microbiota and gut microbiota is reflected on the development of immunity and protection against pathogens.
After birth, milk microbiota forms the most important determinant of infant gut colonization, which occurs in a stepwise fashion. With initial exposure to microbes transferred to the infant gut from mother’s milk, colonization of the intestine of infants starts, and may be essential for the maturation of the gut-associated lymphoid tissue, homeostasis of the intestinal epithelium, and development of intestinal physiology [149]. In the first few days after birth, the infant intestine is characterized by a heterogeneous population of microbes characterized by facultative anaerobes that belong to Enterobacteriaceae, Streptococcus, Enterococcus, and Staphylococcus that thrive on oxygen availability in the newborn gut. Escherichia coli, Enterococcus faecalis, and Enterococcus faecium are the most characterized species among first colonizers. Gradual oxygen consumption by such facultative anaerobes creates a reduced oxygen environment that allows expansion of obligate anaerobes such as Bifidobacterium, Bacteroides, and Clostridium. Intestinal colonization undergoes further changes upon introduction of solid food. The genus Bifidobacterium is detected in the first few months, predominates by 12 months, then declines towards the second year, at the end of which the infant microbiota becomes more diverse [150]. It is essential here to note the distinction between gut microbiota of breastfed and formula-fed infants: It is well known today that Bifidobacterium is the predominant intestinal genus in both feeding modes [151], with species variations. The species Bifidobacterium longum, Bifidobacterium infantis, Bifidobacterium breve, and Bifidobacterium bifidum are commonly detected in breastfed babies, while Bifidobacterium adolescentis and Bifidobacterium pseudocatenulatum, commonly seen among the intestinal adult microbiota, predominate in formula-fed babies [152]. Collectively, formula-fed infants in general have relatively stable and diverse microbial intestinal communities with higher levels of facultative anaerobes and strict anaerobes when compared to breast-fed infants. Fecal samples from breast-fed infants are less complex, with higher numbers of aerobic organisms, and with more critical changes in microbial composition up to the first year. Once weaning, or the introduction of solid foods into the feeding pattern, begins, differences in microbial communities between breastfed and formula-fed infants diminish, and the microbial profile shifts towards the adult intestinal microbiome [153].
The establishment and development of bifidobacterial and lactic acid bacteria in infant’s gut from viable inocula in human milk was demonstrated by several studies. For example, a study of Spanish full-term breastfed infants evaluated similarity between fecal bacteria and those from corresponding milk samples. It was shown that in one-day-old newborns, Enterococcus and Streptococcus were the microorganisms most frequently isolated, while from 10 days until 3 months of age, bifidobacteria became predominant. In corresponding breast-milk, Streptococcus genus was most frequently isolated, and Lactobacillus and Bifidobacterium were also obtained [154]. With an approximate viable bacterial density of 2–4 log colony-forming units/mL of human milk, resulting in a projected daily ingestion of 5–7 log cells with regular breastfeeding, it is expected that neonatal gut microbiota echoes the bacterial composition of breast milk [155]. A trend consistent with breastfeeding is that breast milk selects for a highly adapted intestinal microbiota, dominated by bifidobacteria and termed a “milk-oriented microbiota” [150]. When a disbalance occurs in such orientation health changes may result, for example, preterm infants with altered microbiota are susceptible to necrotizing enterocolitis due to the immaturity of their gastrointestinal and immune systems [48]. In 2020, a meta-analysis of necrotizing enterocolitis in premature infants showed that a mixture of Bifidobacterium and Lactobacillus could reduce morbidity, illustrating role of human milk microbiota in health and disease [156].
The human milk microbiota has a putative role in prevention of infections in newborns, and this occurs through its contribution to the competitive exclusion of pathogens and its involvement in the maturation of the immune system [155]. One possible example of the competitive exclusion of pathogens is the genus Staphylococcus. Several Staphylococcus species, especially S. epidermidis, colonize human milk and the intestine of breastfed infants. In an analysis of milk and stool samples from mother–infant pairs, Jiménez and Colleagues [157] found that S. epidermidis was the predominant species in the milk and stool of breast-fed infants, while it was less prevalent in those of formula fed-infants. The presence of adhesion-related genes in S. epidermidis was very high, while the biofilm-related operon icaD and the gene mecA were only detected only rarely the S. epidermidis strains. Hence, the bacterial attachment factors were present but less commonly the virulence or antibiotic resistance determinants. The commensal staphylococcal strains provided by breast milk to the infant gut may successfully compete with potentially harmful strains. In another, more recent investigation on intestinal Enterococcus from breastfed infants, the commensal enterococci harbored antibiotic resistance and virulence genes. However, the patterns of these genes were not consistent with those described for antibiotic-resistant hospital-associated enterococci, and none were resistant to vancomycin. The frequency of virulence determinants like hemolysin and gelatinase was also low, while some genes linked to colonization were abundant. Taken together, these findings suggest a possible benefit of enterococci in preparing the infant gut for effective opposition against pathogens [158]. It is possible that the pre-colonization of infants with maternal commensal strains will help later in preventing acquisition of infection by more virulent ones [155]. Interestingly, and in the field of HIV infection, lactobacilli cultured from human milk were able to inhibit HIV-1 infection in vitro by blockade of CCR5 co-receptor, and to a lesser extent CXCR4 or both coreceptors, suggesting a probable role for lactobacilli in mucosal protection against HIV-1 [159].
In summary, an association between human milk microbiota, and consequently gut microbiota, and infant health and disease is an important contributor to infant hemostasis. In addition to the relationships described above, ongoing studies are continuing to investigate influence of microbiota on infant irritable bowel syndrome, inflammatory bowel disorder, and type 1 diabetes mellitus [160]. The functions of human milk microbiota and HMOs are summarized in Figure 2 below.
Figure 2.

Functions of human milk microbiota and human milk oligosaccharides (HMOs).
3.6. Probiotics of Human Milk
The term “Probiotic” means “for life” and it is currently used to name bacteria associated with beneficial effects for humans and animals. In 2001, the Food and Agriculture Organization of the United Nations (FAO) and the WHO defined probiotics as “live microorganism which, when administered in adequate amounts confer a health benefit on the host” [161]. This definition was revised in 2014 by the International Scientific Association for Probiotics and Prebiotics, including for probiotics “microorganism for which there are scientific evidence of safety and efficacy [162]. Probiotics that have been principally studied in humans include species from Lactobacillus and Bifidobacterium genera, both of which have been used safely for quite a long time [90]. Probiotic administration early in life may be effective in prevention and treatment of some disorders, leading to a correct microbial colonization while the gut microbiota is still being established [163]. Increasingly administered to infants, probiotics are intended to decrease the risk of certain ailments like necrotizing enterocolitis and late-onset sepsis in preterm infants, colic and antibiotic-associated diarrhea in term infants, in addition to some chronic diseases of childhood such as asthma and atopic disease [164]. The mechanisms responsible for probiotic action still require full elucidation; however, they include modification of the gut microbiota and normalizing its perturbation, competitive adherence to the mucosa and epithelium for pathogen exclusion, strengthening of the gut epithelial barrier, improving the digestion process by complementing the functions of absent digestive enzymes, and modulating the immune system to offer an advantage to the host [162,165]. There are agreed-upon criteria that a probiotic must harbor to be considered efficacious, such as capacity to survive in the GI tract, resistance to stomach acidity, lack of mobile antibiotic resistance genes, and demonstration of health benefits through efficacy testing followed by clinical trials [90,166].
Human milk-derived bacterial strains can be considered as potential probiotics; isolation of strains from milk for subsequent use in infant health and nutrition markets is used [90]. For example, human milk strains of Lactobacillus reuteri, a well-studied probiotic found in the gut and breast milk, can reduce the production of pro-inflammatory cytokines, promote T cell development, and decrease the microbial translocation across the intestinal epithelium from the gut lumen to tissues, thereby reducing inflammation. Notably, the decrease in L. reuteri in humans in the past decades was correlated with an increase in prevalence of inflammatory disorders. Direct supplementation of L. reuteri may be an attractive preventive and/or therapeutic model against inflammatory diseases [167]. In a murine model of asthma developed in 2020, Li and Colleagues [168] showed that oral administration of L. reuteri was more effective in asthma prevention than five other Lactobacillus species, where it reduced the risk of asthma by modulating specific gut microbiota to improve the immune environment of the lungs. L. reuteri supplementation, may, therefore, be a candidate against asthma and other allergic diseases. The same organism from human milk attenuated weight gain, fat accumulation, hypertriglyceridemia, and hypercholesterolemia in mice, thus opening a new horizon for the development of relevant foods to prevent metabolic disorders [169]. In a randomized controlled trial, L. reuteri administration was preventive in reducing pediatric consultations, parental discomfort, and the use of pain relievers for infant colic [170].
Apart from L. reuteri, Lactobacillus fermentum strains from human milk have been studied as probiotic candidates, and proved to be safe, well tolerated and useful for the prophylaxis against community-acquired infections [171]. They also showed cholesterol-lowering effects in simulated models of liver and gastrointestinal tract [172]. Both L. fermentum, and another milk-derived species, Lactobacillus salivarius, enhanced both natural and acquired immune responses, via activation of natural killer and T cell subsets and induction of a broad array of cytokines in peripheral blood mononuclear cells in vitro [173]. An interesting investigation of milk-derived Lactobacillus casei and Lactobacillus paracasei strains, demonstrated anticancer and antioxidant effect in HeLa cell lines of cervical cancer [174]. Likewise, anticancer potential was demonstrated by Rajoka and Colleagues [175] for Lactobacillus rhamnosus isolated from human milk, through induction of apoptosis and down-regulation of the bcl-2 protooncogene, suggesting potential anticancer capability. A review of available studies on the use of probiotics in infantile acute gastroenteritis and antibiotic-associated diarrhea found that L. rhamnosus had the highest compelling evidence of efficacy in reducing duration of gastroenteritis by one day, and was the most effective probiotic in prevention of diarrhea [176].
As for bifidobacteria of human milk, studies are also available to support their use as probiotics. Maldonado and Colleagues [177] conducted a randomized controlled trial on the addition of a strain of Bifidobacterium breve, originally isolated from human milk, to infant formula. They concluded that this probiotic reduced crying rates, is safe, and induces beneficial effects on health. Bifidobacterium longum, isolated from human breast milk, had anti-oncogenic and tumor suppressor potential in murine colorectal cancer [178]. Park and Colleagues [179] showed that a probiotics formula containing both B. longum and Lactobacillus acidophilus alleviated fever, vomiting, and diarrhea with no adverse events in hospitalized infants with rotavirus infection.
Although Bifidobacterium and Lactobacillus strains from human milk persist as the most commonly sought bacteria for probiotic use, the health endorsing effects of other bacteria from milk is also under investigation. Enterococcus faecium, a probable probiotic isolated from breast milk was evaluated both in vitro and in vivo for its safety. The strain was non-hemolytic, sensitive to majority of antibiotics, and showed no alteration of normal growth and development in male rats. No adverse effects on general status nor behavior; additionally, no significant changes were noted in the hematological results, blood biochemistry, organ weights, and histopathology, and none of the vital organs of treated animals displayed signs of bacteremia nor infectivity. These findings indicated the candidature of E. faecium as a potential safe probiotic [180]. In another analysis of E. faecium and E. faecalis strains isolated from breast milk, these bacteria inhibited the growth of Escherichia coli, Listeria monocytogenes, Salmonella typhi, Staphylococcus aureus, Shigella dysenteriae, and Streptococcus agalactiae. However, phenotypic and genotypic virulence analysis indicated hyaluronidase enzyme production and vancomycin resistance in E. faecalis, calling for careful monitoring of probiotic strains for safety parameters [181]. Moreover, a study of cells and cytoplasmic fractions of E. faecalis and Staphylococcus hominis isolated from breast milk against MCF-7 breast cancer cell line revealed significant decrease in cellular proliferation in concentration- and time-dependent style. Morphological signals of apoptosis such as cell shrinkage, cell death, and membrane blebbing were observed in over one-third of the tumor cells [182].
In short, the available data on possible probiotics isolated from human milk is abundant. Further, focused evaluation of safety and efficacy of this rich milieu of microbes is needed to determine which members are good alternative nutraceuticals with health-promoting profiles or promising therapeutic indices.
4. A Review of Human Milk Oligosaccharides (HMOs)
In addition to the ideal balance of nutrients and the abundance of microbes present in human milk, rendering it a perfect food and an optimum medium for growth and immunity, multiple bioactive components also exist in this biofluid, such as immunoglobulins, cytokines, microRNAs, hormones, lactoferrin, and others [183]. Among these, perhaps HMOs constitute a major fraction and a supreme biochemical component that has prompted chemical, microbiological, and medical interest since the beginning of the twentieth century [184]. The major properties of HMOs and their benefits are discussed below.
4.1. Overview and Chemical Composition of HMOs
HMOs form a category of unconjugated, multifunctional, nondigestible, structurally diverse glycans that are unique to humans [185]. Quantitatively, HMOs represent an approximate 20% of total carbohydrate content of breast milk, and are the third largest solid component, after lactose and fats, amounting to 20–25 g/L in colostrum, and gradually decreasing to 5–15 g/L in mature milk [90,186]. The milk of mothers giving birth to premature newborns generally has higher HMO concentrations than that of mothers of term babies [187], although the concentrations were not significantly different in at least one report [188].
Nowadays, over 200 HMOs have been identified, with variable structure and composition in milk between mothers and across the lactation period [183]. The chemical composition of HMOs is shown in Figure 3. Structurally, five monosaccharides form the building blocks of HMOs; these are D-glucose (Glc), D-galactose (Gal), N-acetylglucosamine (GlcNAc), L-fucose (Fuc), and sialic acid, which is exclusively found in humans as N-acetylneuraminic acid (Neu5Ac) [150,184,189]. All these monosaccharides are added to the precursor molecule lactose, which forms the core molecule, and are organized into various structures. The lactose core may be elongated enzymatically by β1-3 linkage to lacto-N-biose, or by β1-6 linkage to N-acetyllactosamine. The HMO structure can be elongated further via addition of: (1) lacto-N-biose and N-acetyllactosamine units (by β1-3 and β1-6 linkages); (2) Fuc connected with α1-2, α1-3, or α1-4 linkages; (3) and/or sialic acid residues attached by α2-3 or α2-6 linkages at terminal positions [190].
Figure 3.

Basic structures of Human Milk Oligosaccharides (HMOs). The core HMO structures are shown in yellow and blue in the center of the figure. The upper panel shows fucosylated HMOs, and the lower panel shows sialylated ones. The building blocks are shown in the lowermost part of the figure [191].
HMOs can be linear or branched. Elongation with lacto-N-biose appears to terminate the HMO chain, whereas N-acetyllactosamine can be further elongated by the addition of any of the two disaccharides. A β1-6 linkage between two disaccharide units introduces chain branching to HMOs. Branched structures are called iso-HMOs, while linear structures without branches are called para-HMOs. Lactose or the elongated oligosaccharide chain can be fucosylated with α1-2, α1-3, or α1-4 linkages, and/or sialylated with α2-3 or α2-6 linkages. Elongation or branching could lead to HMOs up to 15 monosaccharides long [192]. Nearly all HMOs contain lactose at the reducing end, which can be elongated by the addition of GlcNAc and Gal, forming either type 1 (Galβ1-3GlcNAc) or type 2 (Galβ1-4GlcNAc) chains in β1-3 or β1-6 linkages. Furthermore, Fuc and Neu5Ac can be attached to the HMO core or directly to the lactose reducing end [193]. HMOs can be subdivided into three fractions:
-
(a)
A fraction of 35–50% of the total HMO content; these are neutral and contain fucose at the terminal position. These are called neutral (fucosylated) HMOs.
-
(b)
A fraction of 42–55% of the total HMO content; these are neutral N-containing HMOs and contain N-acetylglucosamine at the terminal position. These are called neutral (nonfucosylated) HMOs.
-
(c)
A fraction of 12–14% of total HMO content; these are acidic and contain sialic acid at the terminal position (e.g., 2′-sialyllactose). These are called acidic (sialylated) HMOs [192,194].
Fucosylated HMOs account for about 70% of total HMOs by weight, while sialylated HMOs account for 10–20% [195]. Examples of neutral and acidic HMOs and their basic structures are shown in Table 2 and Figure 3.
Table 2.
Examples of the three different fractions of HMOs, showing neutral (fucosylated and nonfucosylated) and acidic compounds.
| Neutral Fucosylated HMOs | Neutral Nonfucosylated HMOs | Acidic HMOs |
|---|---|---|
| 2′-Fucosyllactose (2′-FL) | Lacto-N-biose (LNB) | Disialyllacto-N-tetraose (DSLNT) |
| 3-Fucosyllactose (3-FL) | Lacto-N-tetraose (LNT) | Sialyllacto-N-neo-tetraose b (LST b) |
| Lacto-N-fucopentaose I (LNFP I) | Lacto-N-neotetraose(LNnT) | Sialyllacto-N-neo-tetraose c (LST c) |
| Lacto-N-fucopentaose II (LNFP II) | Lacto-N-hexaose (LNH) | Sialyl lacto-N-tetraose a (LST a) |
| Lacto-N-fucopentaose III (LNFP III) | Lacto-N-neohexaose (LNnH) | 3′-Sialyllactose (3′-SL) |
| Difucosyllactose (DFL) | 6′-Sialyllactose (6′-SL) |
4.2. Milk Groups Related to Lewis Blood Group-Dependent HMOs: Definition and Relevance to HMOs Research
The HMOs composition and proportion are unique in the milk of each mother, being synthesized in the mammary gland by activity of specific glycosyltransferases, which sequentially add GlcNAc, Gal, Fuc, and Neu5Ac to the basic acceptor molecule, lactose [186]. The profile of HMOs from each mother reflects some blood group characteristics and appears to be genetically determined [192].
First, the link between HMO types and blood groups has been very well elucidated in literature, specifically pertaining to the Lewis blood group antigen [185]. By definition, blood group antigens are specific carbohydrate structures located on red blood cell surface, and constitute secondary gene products, where the primary gene products are the various glycosyltransferase enzymes that attach sugar molecules to the oligosaccharide chain [196]. Among various blood group antigens, the Lewis blood group system is encoded by the Le gene (also called the FUT3 gene). In the mammary gland, the Le gene expresses a fucosyltransferase that adds Fuc in α1-4/3 linkage to synthesize fucosylated HMOs [197].
Second, another gene related to HMOs secretion is the secretor gene, designated by Se. Other blood group antigens, namely the A, B and H, are α1-2-linked fucose containing glycans present on red blood cells in individuals, representing respectively the blood groups A, B, and H. The enzyme fucosyltransferase 1 encoded by the FUT1 gene is responsible for the synthesis of ABH antigens on red blood cells. However, the ABH antigens are also expressed in mucus, milk, and other secretions, where their expression is dependent on another enzyme, fucosyltransferase 2 (secretor type α1-2-fucosyltransferase) encoded by the FUT2 gene [198]. Therefore, in the mammary gland, the Se and Le genes encode respectively the enzymes FUT2 and FUT3, which are involved in the biosynthesis of fucosylated HMOs [199].
Genetic mutations in Se and Le gene affect the synthesis of FUT2 and FUT3 enzymes, thereby affecting the chemical structure of HMOs. While secretor mothers with normal Se and Le genes secrete all types of HMOs, mutations in Se result in inactive FUT2; as such, milk from non-secretor (Se−) women contain no or very limited quantities of α1-2 fucosylated HMOs. Mutations in Le result in inactive FUT3; as such milk from Lewis-negative (Le−) women contains no or very limited quantities α1-4 fucosylated HMOs [200]. Depending on the activity of FUT2 and FUT 3 enzymes in the breast tissue, HMOs composition can be classified into four phenotypes, also referred to as four milk groups, as shown in Table 3 [193,201]. It is worth noting that the α1-3 fucosylated HMOs, such as 3′-fucosyllactose (3′-FL) and difucosyl-para-lacto-N-neohexaose (DFpLNnH), occur in all four SeLe groups, since their synthesis apparently is not affected by the Se and Le genes [199].
Table 3.
The four different milk groups based on genotypes of Le and Se genes, with prevalence, type of linkages in HMOs, and HMO examples. Prevalence is dependent on data from Thurl et al. and refers to the European population [201].
| Milk Group | Prevalence | Fucosylated HMO Linkages | Examples of Fucosylated HMOs |
|---|---|---|---|
| Se+Le+ | 70% | α1-2 α1-4 α1-3 |
2′-Fucosyllactose (2′-FL) Lacto-N-difuco-hexaose I (LNDFH I) Lacto-N-fucopentaose II (LNFP II) 3′-Fucosyllactose (3′-FL) Difucosyllactose (DFL) |
| Se-Le+ | 20% | α1-4 α1-3 |
Lacto-N-difuco-hexaose II (LNDFH II) LNFP II 3′-FL |
| Se+Le- | 10% | α1-2 α1-3 |
Lacto-N-fucopentaose I (LNFP I) 2′-FL 3′-FL |
| Se-Le- | 1% | α1-3 only | Difucosyl-para-lacto-N-neohexaose (DFpLNnH) 3′-FL |
In a study comparing HMOs concentration across milk groups, the total amounts were highest in Se+Le+ milk at day 4 (23.4 g/L) and lowest in Se-Le- (11.3 g/L), with noticeable decline in concentrations in both groups during the first month of lactation (15% and 22%, respectively). Although decreased over time, the levels in Se+Le+ group continued to be highest within the first month, relative to all three groups, indicating that the concentration of total HMOs is linked to the maternal blood type characteristics [187]. While the prevalence of milk groups stated in Table above was detected by earlier studies, and mostly confirmed for European populations [201,202], a higher frequency of expression of recessive Lewis negative and non-secretors was detected in West African populations [194]. Also, a recent investigation from Dubai found the Se+Le+ type to be the most abundant (60%), followed by the Se+Le- type (23%). On the other hand, the non-secretor group was almost equally subdivided between Se–Le+ and Se–Le- groups [134].
In another recent study on breastfeeding Brazilian women [199], HMOs concentrations were highly variable even in women of the same SeLe phenotype, and unprecedented data on HMOs associations with maternal weight, infant’s weight, allergic disease, time postpartum, and sex were revealed. Allergic diseases in the mother, socioeconomic status, infant sex, and home pets were not associated with HMOs concentrations neither in Se+ nor in Se− women. In Se+ women, some positive correlations were observed between 2′-FL and maternal BMI. In the Se− group, significant correlations were mostly negative, involving 6′-SL and infants’ clinical variables such as gestational age, weight, and height.
Another hypothesis regarding normal HMO concentrations and profiles is that they might vary geographically. In a large observational study on health breastfeeding women from 11 nationalities, McGuire and Colleagues [195] reported that 3-fucosyllactose concentration was at least 4 times higher in milk collected in Sweden than in that collected in rural Gambia, while disialyllacto-N-tetraose (DSLNT) was about 4 times lower in Sweden compared to the latter. Maternal age, weight, and body mass index were correlated with several HMOs, and multiple differences in HMOs were shown between ethnically and genetically similar populations living in different locations, suggesting that the environment may play a role in regulating synthesis of HMOs. Targeted genomic analysis, as well as diligent examination of sociocultural, behavioral, and environmental factors is warranted to figure out the roles of these parameters in determining HMO profiles.
As such, both genetic and non-genetic factors influence HMOs composition and concentrations, and few studies have targeted this area. The HMO relations to Le and Se genotypes as well as other factors like gestational age, weight, parity, geographical location, gender, and lactation time remain challenging, especially with their effects on infant health, as described below.
4.3. Physiological Importance and Benefits of HMOs for Infant Health
With their unique and composite carbohydrate structure, HMOs resist gastrointestinal hydrolysis and digestion by gastric acidity and pancreatic and brush-border enzymes, hence are not absorbable in significant amounts. This allows HMOs to reach infant gut and produce a variety of beneficial effects [189].
4.3.1. HMOs and Gut Microbiota Development
Essentially, HMOs serve as metabolic substrates for the growth of beneficial microorganisms in the infant intestine [90]. In infants who are breastfed, bifidobacteria predominate, and such dominance is the result of the presence of a bifidogenic agent in human milk. HMOs constitute such an agent, whereby they are fermented, in the large intestine, by bifidobacterial species [183]. The main products of this fermentation are acetic and lactic acids, which reduce the pH in the intestine, inhibiting the growth of pathogens. In addition, other short chain fatty acids are formed by fermentation, like butyric and propionic acid, where butyric acid is an important energy source for colonocytes. Hence, HMOs are the source of essential molecules needed for maintenance of intestinal health [203].
The ability of bifidobacteria to utilize HMOs depends on both intracellular and extracellular pathways [185]. B. infantis and B. bifidum deploy HMO utilization strategy, which involves translocation of intact oligosaccharides to the intracellular medium using transporters. Once internalized into the bacteria, HMOs are degraded via several intracellular glycosyl hydrolases, and the released monosaccharides are fermented producing ATP, and secreting acetate and lactate as end products. In contrast, the HMO consumption by B. bifidum depends upon initial extracellular processing, primarily via secreted glycosyl hydrolases that decompose HMO into mono- and disaccharides. The resultant products are then imported for intracellular catabolism [204]. When B. bifidum was incubated in HMOs-containing fecal cultures, the extracellular degradation mechanism produced other sugars that stimulated growth of different bifidobacteria. B. bifidum was able to mediate cross-feeding or sharing of HMOs degradants within bifidobacterial communities, increasing their diversity [205]. Recently, an analysis of different HMO degrading enzymes (glycosidase, phosphorylase, fucosidase, sialidase, galactosidase and others), transporters, and gene sets for HMO degradation was performed for at least six different species of bifidobacteria. The abundance of the HMO assimilation mechanisms and HMO-related genes was linked with Bifidobacterium-rich microbiota. Although the knowledge about HMO transporters is still incomplete, the accumulated data stress on the importance of varied HMO assimilation pathways for the formation of infant gut microbiota [206].
In addition to the influence of HMOs on bifidobacteria, other bacterial groups have also been studied. Thongaram and Colleagues [207] showed, using a high-throughput, low volume growth assay that L. acidophilus utilized extracellular β-galactosidase, encoded by lacL gene, to cleave the terminal galactose of LNnT for growth, leaving lacto-N-triose II in the medium. Schwab and Colleagues [208] investigated, in a human study, the interaction between bifidobacteria and Eubacterium hallii, one of the first butyrate producers in infantile gut. They demonstrated that E. hallii consumes acetate, lactate and 1,2-propanediol, which are fermentation products of HMOs by bifidobacteria, and eventually produces butyrate and propionate. These findings suggested E. hallii as a metabolically versatile species in infants, and which utilizes intermediates of bifidobacterial HMO fermentation. Taken together, these data provide insights into how HMOs shape the infant intestinal microbiota.
4.3.2. HMOs and Infection Prevention
In addition to the nutritional role of HMOs on the intestinal microbiota of infants, there is increasing evidence that their interaction with pathogens is important in terms of reducing infection [186]. In doing so, and by being similar to human cell surface glycoconjugates utilized by microbes for attachment to intestinal cells, HMOs act as decoy receptors that bind pathogens [185,192]. For example, 2′-FL inhibits Campylobacter jejuni binding to intestinal mucosa both ex vivo and in vivo, and quenches the inflammation and sequalae induced by this bacterium, causing diarrhea to which young infants worldwide are most susceptible [209]. Furthermore, in a study about bioengineered 2′-FL and 3-FL, 2′-FL inhibited adhesion of C. jejuni, enteropathogenic E. coli, Salmonella enterica, and Pseudomonas aeruginosa to an intestinal human cell line. Moreover, adherence of P. aeruginosa to a human respiratory epithelial cell line was significantly prevented by 2′-FL and 3-FL [210].
Not only bacteria, but protozoa may be affected as well by the anti-adherence effect of HMOs in the infant intestine. HMOs were able to detach Entamoeba histolytica and rescue human intestinal epithelial cells from its destruction in a dose-dependent manner. The cytoprotective effects of HMOs were structure-specific, where Lacto-N-tetraose rescued up to 80 % of the cells, while HMOs with fucose α1-2-linked to the terminal galactose did not show an effect. Galacto-oligosaccharides (GOS), which are added to infant formula to mimic some of the HMOs beneficial effects, abolished E. histolytica attachment and cytotoxicity. These results explain why breast-fed infants have lower risk of E. histolytica infections, and suggest HMOs and GOS as well-tolerated, safe, stable, and valuable alternatives to anti-amoebic agents [211]. HMOs also inhibit infectivity of reoviruses, by blocking their attachment to host cells, and this blockade is serotype-specific [212].
More recently, it has been documented that some HMOs exhibit bacteriostatic properties against group B Streptococcus (GBS). GBS is known to be a leading cause of invasive bacterial infections in newborns, and is vertically acquired during childbirth, secondary to colonization in the vagina. Specific non-sialylated HMOs, in synergy with specific conventional antibiotics agents, may modify particular GBS components in a mode that impairs growth kinetics [213]. In growth and biofilm assays, HMOs showed antibacterial and antibiofilm activity, not only against GBS, but also against Acinetobacter baumannii and Staphylococcus aureus [214]. Additionally, in a quantitative assay of fungal invasiveness, HMOs were effective in prevention of the invasion and damage of the premature infant intestine by Candida albicans hyphae, perhaps providing a model of reducing fungal pathogenesis [215].
4.3.3. HMOs and Immunomodulatory Effect
The immune system of the infant is functionally immature and naïve, and HMOs may play a role in neonatal gastrointestinal and systemic immune development and physiology [216]. The importance of HMOs may be explained, not only by inhibiting the adhesion of microorganisms to the intestinal mucosa, but also by the expression of genes that are involved in inflammation [192]. HMOs can bind to cell surface receptors expressed on epithelial and immune system cells, and thereby modulate neonatal immunity in the infant gut [217]. For example, in a study of HMO effect on colonic epithelial cells, HMOs influenced the expression of multiple cytokines and chemokines, adhesion molecules and receptors. These results suggested that HMOs have a valuable contribution to the development and maturation of the intestinal immune response [218].
To explore the effect of HMOs on the components of immune signaling, human milk was tested on enterocyte cell lines treated with pro-inflammatory molecules, namely, tumor necrosis factor-α (TNF-α) or interleukin-1β (IL-1β), or infected with Listeria or Salmonella. HMOs reduced TNF-α- and IL-1β-induced inflammatory responses to 25–26% and pathogen-induced IL-8 to 36–39%. HMOs reduced the inflammatory response to Salmonella infection by the intestinal tissue, suggesting them as strong physiologic anti-inflammatory agents in human milk, and as contributors to innate immune modulation [219].
In terms of effect on immune cells, HMOs isolated from pooled human milk, induced semi-maturation of human monocytes-derived dendritic cells, and elevated levels of IL-10, IL-27 and IL-6 but not IL-12p70 and TNF-α, thereby playing a modulatory role in development of the neonatal immune system [220]. Recently, 2′-FL, abundantly available in human milk, enhanced Th1-type IFNγ and regulatory IL-10 secretion of peripheral blood mononuclear cells. Dendritic cells exposed to 2′-FL instructed the secretion of IFNγ and IL-10 from CD4+ T-cells, suggesting the growth of a regulatory Th1 response [221].
The effect of HMOs on the immune system may be, as well, associated with the effect of particular receptors. Galectins are glycan-binding proteins involved in intracellular signaling, cell–cell communication processes, cellular proliferation and survival. As endogenous lectins, they are expressed and released by different cell types, including immune, tumor, and endothelial cells [222]. Some HMOs contain β1-3- or β1-4-linked galactose at the non-reducing end, which can be the potential target for galectin-mediated interactions. The binding affinities of 31 free HMOs with the galectins Gal-1, Gal3, and Gal-7 were studied by catch-and-release electrospray ionization mass spectrometry (CaR-ESI-MS). In such analysis, HMOs showed high binding affinity to galectins. Whether HMOs are capable of modulating galectin-mediated immune responses still needs further study but galectins may prove as potential HMO receptors for immune system mediation [223].
4.3.4. HMOs and Intestinal Barrier Function
One important function of HMOs is to promote development of gut barrier function, where the gastrointestinal tract is capable of digestion and absorption of nutrients, while simultaneously acting as a barrier against intruders and toxic agents [185]. HMOs support the intestinal barrier function, initially immature at birth, indirectly by influencing composition of the microbiota, and directly through modulation of intestinal cells. HMOs, through metabolism into short chain fatty acids and cross feeding with butyrate-producing bacteria, induce butyrate production in the colon. Butyrate is an essential metabolite, as it forms the preferable energy source for colon epithelial cells, contributes to the preservation of the gut barrier, and displays immunomodulatory effects [224]. Therefore, HMOs support barrier function through promotion of the formation of fermentation products, especially by Bifidobacterium [225].
Another possible pathway of intestinal barrier evolution by HMOs is stimulating the development of the epithelial glycocalyx on gut epithelial cells, which can contribute to gut homeostasis and form a physical barrier on top of the gut epithelium. The glycocalyx present on the neonatal gut epithelium offers binding sites for commensals and may prevent adhesion of pathogens as well as serve as a barrier for toxins and enzymes. If the glycocalyx components are not properly developed, this may elevate the risk of gastrointestinal disorders [226]. In 2019, Wu and Colleagues [227] demonstrated that HMOs increase mucin levels and decrease bacterial attachment in a murine model of necrotizing enterocolitis. HMOs directly induced the expression of chaperone proteins including protein disulfide isomerase, whose suppression removed the protective effects of HMOs on barrier function as well as on necrotizing enterocolitis protection in vivo. Such results provided insights to the possible mechanism by which HMOs protect the neonatal intestinal barrier through upregulation of mucins. The aforementioned evidence suggests that HMO supplementation may be a valuable approach to build up the gut barrier and perhaps restore or strengthen its function in conditions associated with occurrence of a ”leaky gut” in humans. Such condition includes ageing, irritable bowel syndrome, or inflammatory bowel disease. However, the exact mechanism of these effects remains to be largely explored [224].
4.4. HMOs and Prebiotics
Introduced in 1995, the concept of a prebiotic was described as “a non-digestible food ingredient that beneficially affects the host by selectively stimulating the growth and/or activity of one or a limited number of bacteria in the colon, and thus improves host health” [228]. This definition, armed with major scientific and clinical developments, has significantly evolved over the past two decades [229], until, it was expanded in 2017 by The International Scientific Association for Probiotics and Prebiotics (ISAPP), which currently uses an updated definition for a prebiotic as “a substrate that is selectively utilized by host microorganisms conferring a health benefit” [230].
Prebiotics must be resistant to gastric acidity, to hydrolysis by host enzymes and to gastrointestinal absorption. HMOs have the interesting feature of meeting all three criteria [183]. In fact, research has shown that HMOs, specifically 2′-FL, are even more powerful than standard commercial prebiotics, such as fructo-oligosaccharides and have multiple functions, including immune, gut, and cognition benefits [231]. Despite their beneficial effects, among HMOs, only 2′-FL, LNnT, and 3′-SL are currently added to infant formula, mainly because the complexity of HMOs makes their commercial synthesis still challenging and expensive (Table 4). 2′-FL was a logical starting point for HMO production due to its abundance in milk and its simple structure. LNnT is less abundant but is easier to synthesize. Nondigestible fibers such as GOS and inulin-type fructans are nowadays added to commercially available infant formulas to provide an alternative to HMOs and their benefits [185,232]. However, advances in HMO synthesis are desirable, and various disciplines have been utilized to prepare single HMO compounds by chemical or enzymatic syntheses or by whole-cell biotransformation using recombinant bacterial cells. These attempts utilize lactose as an acceptor molecule, and fucosyltransferases to produce 2′-fucosyllactose, 3-fucosyllactose, or more complex fucosylated core structures. Sialylated HMO can be produced by enzymatically using sialyltransferases and trans-sialidases. Core structures as lacto-N-tetraose can be obtained by glycosyltransferases from chemical donor compounds or by multi-enzyme cascades [191]. Whole cell biotransformation [233] and recombinant techniques [234] are also additional promising options to synthesize HMOs.
Table 4.
The three HMOs approved for use in infant formula, with dates of approval by the European Food Safety Authority and the US Food and Drug Administration.
| HMO | Structure | Approval in EU (European Food Safety Authority) | Approval in USA (US Food and Drug Administration) |
|---|---|---|---|
| 2′-Fucosyllactose (2′-FL) |
|
2015 | 2016 |
| Lacto-N-neotetraose (LNnT) |
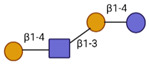
|
2015 | 2016 |
| 3′-Sialyllactose (3′-SL) |
|
- | 2019 |
 .
.
Although attempts to synthesize HMOs represent an initial phase in tapering the compositional gap between human milk and infant formula, it is unclear whether one or two HMO molecules will be capable of inducing the complex actions exerted by the rich blend of HMOs that breastfed infants ingest in human milk. Interestingly, 2′-FL, LNnT, and combinations of both used in a simulated ecosystem of adult human intestine were shown to increase bifidobacterial levels and to decrease levels of IL-6 and cellular permeability [224]. Still, a low diversity of HMOs in breast milk was associated with necrotizing enterocolitis in infants fed exclusively with breast milk [235]. Thus, as more HMOs become commercially available from chemical, microbial, or biotechnological synthesis sources, it is anticipated that more oligosaccharides will be added to infant formula, whether as single compounds or in synergy with additional prebiotics [216]. Plus, the use of human milk-based fortifiers having a full spectrum of HMOs, similar to that in breast milk, is quite tempting to nourish infants and support their gut development and maturity and warrants further investigation. Moreover, the capacity of HMOs to modulate the immune function and to reinforce the gut barrier might support their ability to afford health paybacks in adults, rather than in infants only [224]. Despite that the focus on HMO supplementation to date has been on infants, the transfer of HMOs to adults as well seems on the rise, and commercialization of HMOs is slowly, but surely, coming to make a case for HMO adult supplementation.
5. Milk Microbiota and HMOs: Are There Any Correlations?
As stated above, the composition of human milk is dependent on genetics, diet, lifestyle, lactation period, and other factors. Certain milk components, such as HMOs and bacteria seem also to vary depending on diet, delivery mode, geographical area, and other factors. However, the complex microbiota–HMOs interactions, and their consequences for infant’s health on both short- and long-term, stay uncertain [236]. To put it in a simple manner, milk metabolites, especially HMOs, may influence milk microbial communities, and these, in turn, influence intestinal microbiota of infants, probably favoring specific bacterial genera [237]. However, the interplay among these components is yet to be understood. In recent years, investigations aiming at characterization of relationships among HMOs and bacteria in milk has been ongoing.
In their study of the milk microbiota and HMO relationships in women from several areas of the northwest regions of the US, Williams and Colleagues [237], found that total HMOs and 2′-FL were positively correlated with relative proportion of Staphylococcus, while 3′-SL was inversely correlated with proportions of Ralstonia and Novosphingobium. However, with only 16 milk samples analyzed, the authors suggested that a more thorough interpretation of the repercussions of various HMOs on milk microbiota is needed. In a recent pilot observational study on Spanish breastfeeding women, the predominant HMOs in milk of secretor mothers were 2′-FL and lacto-N-fucopentaose I, whereas the milk of non-secretor mothers was characterized by lacto-N-fucopentaose II and lacto-N-difucohexaose II. Differences in microbiota types and concentrations were related to secretor/non-secretor status. Lactobacilli, enterococci, and streptococci were lower in non-secretor compared to secretor samples, while Bifidobacterium was less prevalent in non-secretor samples. Despite little differences on diversity and richness, non-secretor samples had less Actinobacteria and more Enterobacteriaceae, Lactobacillaceae, and Staphylococcaceae. This study supported the limited data on the complex interaction between microbiota and HMO during lactation, specifically with secretor status, although the potential biological influence on the newborn remains indiscernible [238]. In human colostrum of women from Finland, HMO content analyzed by high-pressure liquid chromatography and microbiota types analyzed by quantitative PCR, showed higher total HMO concentration associated with higher counts of Bifidobacterium. Furthermore, positive correlations were observed among sialylated HMOs and B. breve, and among non-fucosylated/non-sialylated HMOs and B. longum. Furthermore, positive correlations were observed among fucosylated HMOs and Akkermansia muciniphila, and among fucosylated/sialylated HMOs and S. aureus. Both oligosaccharides and microbes provided a concise inoculum that will provide for the compositional development of the infant gut microbiota [239]. In another study, lactobacilli and bifidobacteria, obtained from commercial probiotic supplements or from culture collections, were capable of fermentation of HMOs. B. longum ssp. infantis and B. infantis were able to ferment 3′-sialyllactose, 6′-sialyllactose, 2′-fucosyllactose, and 3′-fucosyllactose. B. infantis degraded almost 90% of 2′-fucosyllactose but left most of the fucose, as revealed by HPLC. Among bifidobacteria, only the B. infantis strains and B. breve were able to ferment lacto-N-neotetraose (LNnT). On the other hand, among lactobacilli, L. acidophilus was the most efficient at utilizing LNnT. The extracellular β-galactosidase of L. acidophilus, encoded by lacL, cleaved the terminal galactose of LNnT for growth, leaving lacto-N-triose II in the media. Moreover, the inactivation of lacL gene abolished growth of L. acidophilus on LNnT [207]. Not only do these results underwrite the increased scientific awareness of HMO-microbiota interactions, but they also emphasize the promise for prospective combinations of probiotics and HMOs, in a likely analogy to human milk constituents.
To ascertain the interaction between metabolomic profile of milk, including HMOs, and milk microbiota, an investigation on milk from different populations analyzed milk content using nuclear magnetic resonance spectroscopy (NMR). Specific interrelations between HMOs and milk microbiota were identified. Gammaproteobacteria positively correlated with lacto-N-fucopentaose I, and 2-fucosyllactose, although these HMOs negatively associated with Alpha and Betaproteobacteria and Bacilli. Further, the Actinobacteria in the human breast milk were negatively associated with lacto-N-fucopentaose I and 2-fucosyllactose acetate. These findings are in agreement with the existing, although limited data on complex relations between the milk microbiome and various types of HMOs during lactation [236]. In milk of mothers from Dubai, negative correlations were detected for the genus Atopobium versus sialyl-lacto-N-tetraose, for Leptotrichia and Veillonella versus 6′-sialyl-lactose, and for Porphyromonas versus lacto-N-hexaose. It could be hypothesized that, because they were able to use that particular HMO, they proliferated in the mammary gland; this, nevertheless, remains to be investigated [134]. In another investigation, Streptococcus and Staphylococcus showed an increase in relative abundance in relation to high consumption of the fucosylated HMOs, namely, DFL, LNFPII, and LNFPIII. The presence of HMOs may stimulate the growth of breast milk-associated Staphylococcus by activating growth-promoting signals without being actively metabolized by this strain. Streptococcus and Staphylococcus that cross-feed on HMO metabolites may play a role as well, although at the present time, no studies show direct evidence of that [240].
Scientific research on associations between milk microbiota profiles and HMOs has sometimes elucidated mechanisms of HMO utilization, rather than only the correlations. Perhaps the best example of this is the utilization of HMOs by Bifidobacterium in breast milk, which has been relatively well described. The dominant Bifidobacterium species can utilize various HMO components as their solitary carbohydrate source [241]. These species basically include strains of B. bifidum, B. longum subsp. infantis, and B. breve. B. bifidum extracellularly hydrolyzes complex HMO structures, including LNT and LNnT, using glycosyl hydrolases, followed by internalization and intracellular degradation and metabolism of the resulting mono- and disaccharides, such as LNB. Furthermore, B. longum subsp. infantis internalizes intact LNT, LNnT, and LNB, then uses a series of sequential hydrolytic/phosphorolytic reactions starting from the nonreducing end of the carbohydrate structures for degradation and metabolic processing. However, B. infantis is also known to take up and utilize fucosyl- and sialyl-lactose [242]. To this end, a multicomponent transcriptional regulation system that controls the recently identified pathways of HMO metabolism was described in B. breve. These include three transcriptional regulators, with the corresponding operator and associated promoter sequences, the latter governing the transcription of the genetic elements involved in LNnT/LNB metabolism. This result, not only provides some vision of the regulatory mechanisms present in bifidobacteria, but also offers an example of a system of consecutive steps regulating microbial carbohydrate metabolism [243].
The above results show high variability in HMOs and milk microbiota profiles, with possible existing correlations. It is anticipated that larger trials, more diverse populations, and targeted genomic analyses will elucidate such correlations in a more definitive manner. These trials should also be able to establish whether such correlations are dependent on genomic, sociocultural, or environmental features.
6. Future Implications in Milk Microbiota and HMOs Research
It is of no doubt that human milk harbors extraordinary features which make it a treasured fluid, whose amazing assets and roles are yet to be fully clarified. Research on human milk will continue to elucidate additional benefits and correlations among components, and such area of scientific investigation is very promising. Perhaps the driving force for such research lies in the incessant attempts to produce an industrial infant formula that mimics the nutritional breast milk composition as much as possible, to be able to substitute breastfeeding when this process is not possible or is solely inadequate [244]. Randomized controlled trials on milk benefits continue to be realized, and some effects under investigation are the analgesic effect of human milk [245] or milk odor [246,247], and the lessened need for antibiotics up to the first year of life [248].
In the field of milk microbiota and HMOs, characterization of the valuable organisms of the human microbiota and their utilization of defined HMOs on a strain-specific basis is essential. Furthermore, the number of randomized trials that have assessed the impact of HMOs on infant health is very limited. Furthermore, the number of subjects studied for determination of the safety and tolerance of HMO-supplemented formulas is not enough to generate actual evidence of the potential prophylactic role of HMOs against infection [192]. As previously mentioned, limited supply of HMOs currently impedes extensive trials on these compounds to develop targeted HMO-based prebiotic therapy [249,250]. Continued development of large-scale HMOs synthesis via different enzymatic engineering pathways should facilitate human trials, especially interventional ones, in the coming years [251]. New modalities of synthesis such as loop engineering [252] and the use of specific fucosidases [253,254] to synthesize specific HMO structures are being explored. Such new means, coupled with increased understanding of the dynamic interrelationship between HMO structure and the developing intestinal microbiota, shall enable the design and testing of novel prebiotic agents based on HMOs in clinical settings in the near future.
7. Conclusions
In brief, human milk encompasses a wealth of biologically active components that contribute to multiple advantages, far beyond infant nutrition. The past decade has witnessed an explosion of research and interest in the human milk microbiota. This fluid represents a unique resource for translational medicine, from research on the bench into clinical applications such as pro- and pre-biotics [255]. With its rich assortment of biologically active molecules and demonstrated benefits, both in newborn life and later into adulthood, ongoing characterization of the mechanisms through which milk components enhance development, shaping of microbiota, and immunity is on the rise. Additional investigations regarding links between the human milk microbiota and remarkably bioactive molecules, such as HMOs, are highly warranted.
Funding
This research was funded in part by an unrestricted grant from Prolacta Bioscience. The APC was funded by Zayed University.
Conflicts of Interest
The author declares no conflict of interest. The funders had no role in the design and the writing of the review.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Mosca F., Giannì M.L. Human Milk: Composition and Health Benefits. Pediatr. Med. Chir. 2017;39:155. doi: 10.4081/pmc.2017.155. [DOI] [PubMed] [Google Scholar]
- 2.Johnson A., Kirk R., Rosenblum K.L., Muzik M. Enhancing Breastfeeding Rates among African American Women: A Systematic Review of Current Psychosocial Interventions. Breastfeed Med. 2015;10:45–62. doi: 10.1089/bfm.2014.0023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Westerfield K.L., Koenig K., Oh R. Breastfeeding: Common Questions and Answers. Am. Fam. Phys. 2018;98:368–373. [PubMed] [Google Scholar]
- 4.Bagci Bosi A.T., Eriksen K.G., Sobko T., Wijnhoven T.M.A., Breda J. Breastfeeding Practices and Policies in WHO European Region Member States. Public Health Nutr. 2016;19:753–764. doi: 10.1017/S1368980015001767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.World Breastfeeding Week 2020 Message. [(accessed on 19 September 2020)]; Available online: https://www.who.int/news-room/detail/31-07-2020-world-breastfeeding-week-2020-message.
- 6.Section on Breastfeeding Breastfeeding and the Use of Human Milk. Pediatrics. 2012;129:e827–e841. doi: 10.1542/peds.2011-3552. [DOI] [PubMed] [Google Scholar]
- 7.Ojo-Okunola A., Cacciatore S., Nicol M.P., du Toit E. The Determinants of the Human Milk Metabolome and Its Role in Infant Health. Metabolites. 2020;10:77. doi: 10.3390/metabo10020077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ballard O., Morrow A.L. Human Milk Composition: Nutrients and Bioactive Factors. Pediatr. Clin. 2013;60:49–74. doi: 10.1016/j.pcl.2012.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gidrewicz D.A., Fenton T.R. A Systematic Review and Meta-Analysis of the Nutrient Content of Preterm and Term Breast Milk. BMC Pediatr. 2014;14:216. doi: 10.1186/1471-2431-14-216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Dror D.K., Allen L.H. Overview of Nutrients in Human Milk. Adv. Nutr. 2018;9:278S–294S. doi: 10.1093/advances/nmy022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Picciano M.F. Representative Values for Constituents of Human Milk. Pediatr. Clin. N. Am. 2001;48:263–264. doi: 10.1016/S0031-3955(05)70299-6. [DOI] [PubMed] [Google Scholar]
- 12.Gay M.C.L., Koleva P.T., Slupsky C.M., Toit E.D., Eggesbo M., Johnson C.C., Wegienka G., Shimojo N., Campbell D.E., Prescott S.L., et al. Worldwide Variation in Human Milk Metabolome: Indicators of Breast Physiology and Maternal Lifestyle? Nutrients. 2018;10:1151. doi: 10.3390/nu10091151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Musilova S., Rada V., Vlkova E., Bunesova V. Beneficial Effects of Human Milk Oligosaccharides on Gut Microbiota. Benef. Microbes. 2014;5:273–283. doi: 10.3920/BM2013.0080. [DOI] [PubMed] [Google Scholar]
- 14.Bhandari N., Prajapati R. Prevalence of Exclusive Breast Feeding and Its Associated Factors among Mothers. Kathmandu Univ. Med. J. KUMJ. 2018;16:166–170. [PubMed] [Google Scholar]
- 15.Gertosio C., Meazza C., Pagani S., Bozzola M. Breastfeeding and Its Gamut of Benefits. Minerva. Pediatr. 2016;68:201–212. [PubMed] [Google Scholar]
- 16.Binns C., Lee M., Low W.Y. The Long-Term Public Health Benefits of Breastfeeding. Asia Pac. J. Public Health. 2016;28:7–14. doi: 10.1177/1010539515624964. [DOI] [PubMed] [Google Scholar]
- 17.Horta B.L., Victora C.G. Short-Term Effects of Breastfeeding. A Systematic Review on the Benefits of Breastfeeding on Diarrhoea and Pneumonia Mortality. World Health Organization; Geneva, Switzerland: 2013. [Google Scholar]
- 18.Sankar M.J., Sinha B., Chowdhury R., Bhandari N., Taneja S., Martines J., Bahl R. Optimal Breastfeeding Practices and Infant and Child Mortality: A Systematic Review and Meta-Analysis. Acta Paediatr. 2015;104:3–13. doi: 10.1111/apa.13147. [DOI] [PubMed] [Google Scholar]
- 19.Lamberti L.M., Fischer Walker C.L., Noiman A., Victora C., Black R.E. Breastfeeding and the Risk for Diarrhea Morbidity and Mortality. BMC Public Health. 2011;11(Suppl. 3):S15. doi: 10.1186/1471-2458-11-S3-S15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Santos F.S., Santos F.C.S., dos Santos L.H., Leite A.M., de Mello D.F. Breastfeeding and Protection against Diarrhea: An Integrative Review of Literature. Einstein Sao Paulo. 2015;13:435–440. doi: 10.1590/S1679-45082015RW3107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rouw E., von Gartzen A., Weißenborn A. The importance of breastfeeding for the infant. Bundesgesundheitsblatt Gesundh. Gesundh. 2018;61:945–951. doi: 10.1007/s00103-018-2773-4. [DOI] [PubMed] [Google Scholar]
- 22.Lamberti L.M., Zakarija-Grković I., Fischer Walker C.L., Theodoratou E., Nair H., Campbell H., Black R.E. Breastfeeding for Reducing the Risk of Pneumonia Morbidity and Mortality in Children under Two: A Systematic Literature Review and Meta-Analysis. BMC Public Health. 2013;13(Suppl. 3):S18. doi: 10.1186/1471-2458-13-S3-S18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Quigley M.A., Carson C., Sacker A., Kelly Y. Exclusive Breastfeeding Duration and Infant Infection. Eur. J. Clin. Nutr. 2016;70:1420–1427. doi: 10.1038/ejcn.2016.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bowatte G., Tham R., Allen K.J., Tan D.J., Lau M., Dai X., Lodge C.J. Breastfeeding and Childhood Acute Otitis Media: A Systematic Review and Meta-Analysis. Acta Paediatr. 2015;104:85–95. doi: 10.1111/apa.13151. [DOI] [PubMed] [Google Scholar]
- 25.Christensen N., Bruun S., Søndergaard J., Christesen H.T., Fisker N., Zachariassen G., Sangild P.T., Husby S. Breastfeeding and Infections in Early Childhood: A Cohort Study. Pediatrics. 2020;146 doi: 10.1542/peds.2019-1892. [DOI] [PubMed] [Google Scholar]
- 26.Zivich P., Lapika B., Behets F., Yotebieng M. Implementation of Steps 1–9 to Successful Breastfeeding Reduces the Frequency of Mild and Severe Episodes of Diarrhea and Respiratory Tract Infection Among 0–6 Month Infants in Democratic Republic of Congo. Matern. Child Health J. 2018;22:762–771. doi: 10.1007/s10995-018-2446-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Davanzo R., Moro G., Sandri F., Agosti M., Moretti C., Mosca F. Breastfeeding and Coronavirus Disease-2019: Ad Interim Indications of the Italian Society of Neonatology Endorsed by the Union of European Neonatal & Perinatal Societies. Matern. Child Nutr. 2020;16:e13010. doi: 10.1111/mcn.13010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.World Health Organization Breastfeeding and COVID-19. [(accessed on 30 January 2021)]; Available online: https://www.who.int/news-room/commentaries/detail/breastfeeding-and-covid-19.
- 29.CDC Coronavirus Disease (COVID-19) and Breastfeeding. [(accessed on 30 January 2021)]; Available online: https://www.cdc.gov/breastfeeding/breastfeeding-special-circumstances/maternal-or-infant-illnesses/covid-19-and-breastfeeding.html.
- 30.Fernández-Carrasco F.J., Vázquez-Lara J.M., González-Mey U., Gómez-Salgado J., Parrón-Carreño T., Rodríguez-Díaz L. Coronavirus Covid-19 Infection and Breastfeeding: An Exploratory Review. [(accessed on 20 September 2020)]; Available online: https://pubmed.ncbi.nlm.nih.gov/32458823/ [PubMed]
- 31.Greer F.R., Sicherer S.H., Burks A.W., Committee on Nutrition. Section on Allergy and Immunology The Effects of Early Nutritional Interventions on the Development of Atopic Disease in Infants and Children: The Role of Maternal Dietary Restriction, Breastfeeding, Hydrolyzed Formulas, and Timing of Introduction of Allergenic Complementary Foods. Pediatrics. 2019;143 doi: 10.1542/peds.2019-0281. [DOI] [PubMed] [Google Scholar]
- 32.Matsumoto N., Yorifuji T., Nakamura K., Ikeda M., Tsukahara H., Doi H. Breastfeeding and Risk of Food Allergy: A Nationwide Birth Cohort in Japan. Allergol. Int. 2020;69:91–97. doi: 10.1016/j.alit.2019.08.007. [DOI] [PubMed] [Google Scholar]
- 33.Carlin R.F., Moon R.Y. Risk Factors, Protective Factors, and Current Recommendations to Reduce Sudden Infant Death Syndrome: A Review. JAMA Pediatr. 2017;171:175–180. doi: 10.1001/jamapediatrics.2016.3345. [DOI] [PubMed] [Google Scholar]
- 34.Alm B., Wennergren G., Möllborg P., Lagercrantz H. Breastfeeding and Dummy Use Have a Protective Effect on Sudden Infant Death Syndrome. Acta Paediatr. 2016;105:31–38. doi: 10.1111/apa.13124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Horne R.S.C. Sudden Infant Death Syndrome: Current Perspectives. Intern. Med. J. 2019;49:433–438. doi: 10.1111/imj.14248. [DOI] [PubMed] [Google Scholar]
- 36.Adams S.M., Ward C.E., Garcia K.L. Sudden Infant Death Syndrome. Am. Fam. Phys. 2015;91:778–783. [PubMed] [Google Scholar]
- 37.Hauck F.R., Thompson J.M.D., Tanabe K.O., Moon R.Y., Vennemann M.M. Breastfeeding and Reduced Risk of Sudden Infant Death Syndrome: A Meta-Analysis. Pediatrics. 2011;128:103–110. doi: 10.1542/peds.2010-3000. [DOI] [PubMed] [Google Scholar]
- 38.Franco P., Scaillet S., Wermenbol V., Valente F., Groswasser J., Kahn A. The Influence of a Pacifier on Infants’ Arousals from Sleep. J. Pediatr. 2000;136:775–779. [PubMed] [Google Scholar]
- 39.Horne R.S.C., Parslow P.M., Ferens D., Watts A.-M., Adamson T.M. Comparison of Evoked Arousability in Breast and Formula Fed Infants. Arch. Dis. Child. 2004;89:22–25. [PMC free article] [PubMed] [Google Scholar]
- 40.Horta B.L., Loret de Mola C., Victora C.G. Long-Term Consequences of Breastfeeding on Cholesterol, Obesity, Systolic Blood Pressure and Type 2 Diabetes: A Systematic Review and Meta-Analysis. Acta Paediatr. 2015;104:30–37. doi: 10.1111/apa.13133. [DOI] [PubMed] [Google Scholar]
- 41.Marseglia L., Manti S., D’Angelo G., Cuppari C., Salpietro V., Filippelli M., Trovato A., Gitto E., Salpietro C., Arrigo T. Obesity and Breastfeeding: The Strength of Association. Women Birth. 2015;28:81–86. doi: 10.1016/j.wombi.2014.12.007. [DOI] [PubMed] [Google Scholar]
- 42.Yan J., Liu L., Zhu Y., Huang G., Wang P.P. The Association between Breastfeeding and Childhood Obesity: A Meta-Analysis. BMC Public Health. 2014;14:1267. doi: 10.1186/1471-2458-14-1267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ortega-García J.A., Kloosterman N., Alvarez L., Tobarra-Sánchez E., Cárceles-Álvarez A., Pastor-Valero R., López-Hernández F.A., Sánchez-Solis M., Claudio L. Full Breastfeeding and Obesity in Children: A Prospective Study from Birth to 6 Years. Child Obes. 2018;14:327–337. doi: 10.1089/chi.2017.0335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ak V. Does Breastfeeding Shape Food Preferences? Links to Obesity. [(accessed on 20 September 2020)]; Available online: https://pubmed.ncbi.nlm.nih.gov/28903109/
- 45.Dahiya D.K., Puniya M., Shandilya U.K., Dhewa T., Kumar N., Kumar S., Puniya A.K., Shukla P. Gut Microbiota Modulation and Its Relationship with Obesity Using Prebiotic Fibers and Probiotics: A Review. Front. Microbiol. 2017;8:563. doi: 10.3389/fmicb.2017.00563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Patel A.L., Kim J.H. Human Milk and Necrotizing Enterocolitis. Semin. Pediatr. Surg. 2018;27:34–38. doi: 10.1053/j.sempedsurg.2017.11.007. [DOI] [PubMed] [Google Scholar]
- 47.Cotten C.M. Modifiable Risk Factors in Necrotizing Enterocolitis. Clin. Perinatol. 2019;46:129–143. doi: 10.1016/j.clp.2018.10.007. [DOI] [PubMed] [Google Scholar]
- 48.Maffei D., Schanler R.J. Human Milk Is the Feeding Strategy to Prevent Necrotizing Enterocolitis! Semin. Perinatol. 2017;41:36–40. doi: 10.1053/j.semperi.2016.09.016. [DOI] [PubMed] [Google Scholar]
- 49.Lindoso L., Mondal K., Venkateswaran S., Somineni H.K., Ballengee C., Walters T.D., Griffiths A., Noe J.D., Crandall W., Snapper S., et al. The Effect of Early-Life Environmental Exposures on Disease Phenotype and Clinical Course of Crohn’s Disease in Children. Am. J. Gastroenterol. 2018;113:1524–1529. doi: 10.1038/s41395-018-0239-9. [DOI] [PubMed] [Google Scholar]
- 50.Silano M., Agostoni C., Sanz Y., Guandalini S. Infant Feeding and Risk of Developing Celiac Disease: A Systematic Review. BMJ Open. 2016;6:e009163. doi: 10.1136/bmjopen-2015-009163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Murff H.J., Edwards T.L. Endogenous Production of Long-Chain Polyunsaturated Fatty Acids and Metabolic Disease Risk. Curr. Cardiovasc. Risk Rep. 2014;8 doi: 10.1007/s12170-014-0418-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Amitay E.L., Keinan-Boker L. Breastfeeding and Childhood Leukemia Incidence: A Meta-Analysis and Systematic Review. JAMA Pediatr. 2015;169:e151025. doi: 10.1001/jamapediatrics.2015.1025. [DOI] [PubMed] [Google Scholar]
- 53.Küçükçongar A., Oğuz A., Pınarlı F.G., Karadeniz C., Okur A., Kaya Z., Çelik B. Breastfeeding and Childhood Cancer: Is Breastfeeding Preventative to Childhood Cancer? Pediatric Hematol. Oncol. 2015;32:374–381. [PubMed] [Google Scholar]
- 54.Bar S., Milanaik R., Adesman A. Long-Term Neurodevelopmental Benefits of Breastfeeding. Curr. Opin. Pediatr. 2016;28:559–566. doi: 10.1097/MOP.0000000000000389. [DOI] [PubMed] [Google Scholar]
- 55.Kramer M.S., Aboud F., Mironova E., Vanilovich I., Platt R.W., Matush L., Igumnov S., Fombonne E., Bogdanovich N., Ducruet T., et al. Breastfeeding and Child Cognitive Development: New Evidence from a Large Randomized Trial. Arch. Gen. Psychiatry. 2008;65:578–584. doi: 10.1001/archpsyc.65.5.578. [DOI] [PubMed] [Google Scholar]
- 56.Holme A., MacArthur C., Lancashire R. The Effects of Breastfeeding on Cognitive and Neurological Development of Children at 9 Years. Child Care Health Dev. 2010;36:583–590. doi: 10.1111/j.1365-2214.2009.01068.x. [DOI] [PubMed] [Google Scholar]
- 57.Koletzko B., Rodriguez-Palmero M. Polyunsaturated Fatty Acids in Human Milk and Their Role in Early Infant Development. J. Mammary Gland Biol. Neoplasia. 1999;4:269–284. doi: 10.1023/A:1018749913421. [DOI] [PubMed] [Google Scholar]
- 58.Bernard J.Y., Armand M., Peyre H., Garcia C., Forhan A., De Agostini M., Charles M.-A., Heude B., EDEN Mother-Child Cohort Study Group (Etude des Déterminants pré- et postnatals précoces du développement et de la santé de l’Enfant) Breastfeeding, Polyunsaturated Fatty Acid Levels in Colostrum and Child Intelligence Quotient at Age 5–6 Years. J. Pediatr. 2017;183:43–50.e3. doi: 10.1016/j.jpeds.2016.12.039. [DOI] [PubMed] [Google Scholar]
- 59.Hadley K.B., Ryan A.S., Forsyth S., Gautier S., Salem N. The Essentiality of Arachidonic Acid in Infant Development. Nutrients. 2016;8:216. doi: 10.3390/nu8040216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ghozy S., Tran L., Naveed S., Quynh T.T.H., Helmy Zayan A., Waqas A., Sayed A.K.H., Karimzadeh S., Hirayama K., Huy N.T. Association of Breastfeeding Status with Risk of Autism Spectrum Disorder: A Systematic Review, Dose-Response Analysis and Meta-Analysis. Asian J. Psychiatr. 2020;48:101916. doi: 10.1016/j.ajp.2019.101916. [DOI] [PubMed] [Google Scholar]
- 61.Kielbratowska B., Kazmierczak M., Michalek J., Preis K. Temperament and the Mother-Infant Dyad: Associations with Breastfeeding and Formula Feeding with a Bottle. Infant Ment. Health J. 2015;36:243–250. doi: 10.1002/imhj.21508. [DOI] [PubMed] [Google Scholar]
- 62.Shelton K.H., Collishaw S., Rice F.J., Harold G.T., Thapar A. Using a Genetically Informative Design to Examine the Relationship between Breastfeeding and Childhood Conduct Problems. Eur. Child Adolesc. Psychiatry. 2011;20:571–579. doi: 10.1007/s00787-011-0224-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Schwarz E.B., Nothnagle M. The Maternal Health Benefits of Breastfeeding. Am. Fam. Phys. 2015;91:603–604. [PubMed] [Google Scholar]
- 64.Victora C.G., Bahl R., Barros A.J.D., França G.V.A., Horton S., Krasevec J., Murch S., Sankar M.J., Walker N., Rollins N.C., et al. Breastfeeding in the 21st Century: Epidemiology, Mechanisms, and Lifelong Effect. Lancet. 2016;387:475–490. doi: 10.1016/S0140-6736(15)01024-7. [DOI] [PubMed] [Google Scholar]
- 65.Unar-Munguía M., Torres-Mejía G., Colchero M.A., González de Cosío T. Breastfeeding Mode and Risk of Breast Cancer: A Dose-Response Meta-Analysis. J. Hum. Lact. 2017;33:422–434. doi: 10.1177/0890334416683676. [DOI] [PubMed] [Google Scholar]
- 66.Pan H., He Z., Ling L., Ding Q., Chen L., Zha X., Zhou W., Liu X., Wang S. Reproductive Factors and Breast Cancer Risk among BRCA1 or BRCA2 Mutation Carriers: Results from Ten Studies. Cancer Epidemiol. 2014;38:1–8. doi: 10.1016/j.canep.2013.11.004. [DOI] [PubMed] [Google Scholar]
- 67.Manrique Tejedor J., Figuerol Calderó M.I., Cuéllar De Frutos A. Breastfeeding as a method of breast cancer prevention. Rev. Enferm. 2015;38:32–38. [PubMed] [Google Scholar]
- 68.Modugno F., Goughnour S.L., Wallack D., Edwards R.P., Odunsi K., Kelley J.L., Moysich K., Ness R.B., Brooks M.M. Breastfeeding Factors and Risk of Epithelial Ovarian Cancer. Gynecol. Oncol. 2019;153:116–122. doi: 10.1016/j.ygyno.2019.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sung H.K., Ma S.H., Choi J.-Y., Hwang Y., Ahn C., Kim B.-G., Kim Y.-M., Kim J.W., Kang S., Kim J., et al. The Effect of Breastfeeding Duration and Parity on the Risk of Epithelial Ovarian Cancer: A Systematic Review and Meta-Analysis. J. Prev. Med. Public Health. 2016;49:349–366. doi: 10.3961/jpmph.16.066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Rajaei S., Rigdon J., Crowe S., Tremmel J., Tsai S., Assimes T.L. Breastfeeding Duration and the Risk of Coronary Artery Disease. J. Womens Health Larchmt. 2019;28:30–36. doi: 10.1089/jwh.2018.6970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Peters S.A., van der Schouw Y.T., Wood A.M., Sweeting M.J., Moons K.G., Weiderpass E., Arriola L., Benetou V., Boeing H., Bonnet F., et al. Parity, Breastfeeding and Risk of Coronary Heart Disease: A Pan-European Case-Cohort Study. Eur. J. Prev. Cardiol. 2016;23:1755–1765. doi: 10.1177/2047487316658571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Stuebe A.M., Michels K.B., Willett W.C., Manson J.E., Rexrode K., Rich-Edwards J.W. Duration of Lactation and Incidence of Myocardial Infarction in Middle-to-Late Adulthood. Am. J. Obstet. Gynecol. 2009;200:138.e1–138.e8. doi: 10.1016/j.ajog.2008.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Poudel R.R., Shrestha D. Breastfeeding for Diabetes Prevention. J. Pak. Med. Assoc. 2016;66:S88–S90. [PubMed] [Google Scholar]
- 74.Park S., Choi N.-K. Breastfeeding and Maternal Hypertension. Am. J. Hypertens. 2018;31:615–621. doi: 10.1093/ajh/hpx219. [DOI] [PubMed] [Google Scholar]
- 75.Qu G., Wang L., Tang X., Wu W., Sun Y. Association Between Duration of Breastfeeding and Maternal Hypertension: A Systematic Review and Meta-Analysis. Breastfeed Med. 2018;13:318–326. doi: 10.1089/bfm.2017.0180. [DOI] [PubMed] [Google Scholar]
- 76.Kirkegaard H., Bliddal M., Støvring H., Rasmussen K.M., Gunderson E.P., Køber L., Sørensen T.I.A., Nohr E.A. Breastfeeding and Later Maternal Risk of Hypertension and Cardiovascular Disease—The Role of Overall and Abdominal Obesity. Prev. Med. 2018;114:140–148. doi: 10.1016/j.ypmed.2018.06.014. [DOI] [PubMed] [Google Scholar]
- 77.Jäger S., Jacobs S., Kröger J., Fritsche A., Schienkiewitz A., Rubin D., Boeing H., Schulze M.B. Breast-Feeding and Maternal Risk of Type 2 Diabetes: A Prospective Study and Meta-Analysis. Diabetologia. 2014;57:1355–1365. doi: 10.1007/s00125-014-3247-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Luo J., Hendryx M., LeBlanc E.S., Shadyab A.H., Qi L., Sealy-Jefferson S., Manson J.E. Associations Between Parity, Breastfeeding, and Risk of Maternal Type 2 Diabetes Among Postmenopausal Women. Obstet. Gynecol. 2019;134:591–599. doi: 10.1097/AOG.0000000000003407. [DOI] [PubMed] [Google Scholar]
- 79.Stuebe A.M., Rich-Edwards J.W. The Reset Hypothesis: Lactation and Maternal Metabolism. Am. J. Perinatol. 2009;26:81–88. doi: 10.1055/s-0028-1103034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wang T., Xu Y., Xu M., Ning G., Lu J., Dai M., Xu B., Sun J., Sun W., Lai S., et al. Circulating Prolactin and Risk of Type 2 Diabetes: A Prospective Study. Am. J. Epidemiol. 2016;184:295–301. doi: 10.1093/aje/kwv326. [DOI] [PubMed] [Google Scholar]
- 81.Bobrow K.L., Quigley M.A., Green J., Reeves G.K., Beral V., Million Women Study Collaborators Persistent Effects of Women’s Parity and Breastfeeding Patterns on Their Body Mass Index: Results from the Million Women Study. Int. J. Obes. 2013;37:712–717. doi: 10.1038/ijo.2012.76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Groër M.W. Differences between Exclusive Breastfeeders, Formula-Feeders, and Controls: A Study of Stress, Mood, and Endocrine Variables. Biol. Res. Nurs. 2005;7:106–117. doi: 10.1177/1099800405280936. [DOI] [PubMed] [Google Scholar]
- 83.Doan T., Gardiner A., Gay C.L., Lee K.A. Breast-Feeding Increases Sleep Duration of New Parents. J. Perinat. Neonatal. Nurs. 2007;21:200–206. doi: 10.1097/01.JPN.0000285809.36398.1b. [DOI] [PubMed] [Google Scholar]
- 84.Figueiredo B., Dias C.C., Brandão S., Canário C., Nunes-Costa R. Breastfeeding and Postpartum Depression: State of the Art Review. J. Pediatr. Rio J. 2013;89:332–338. doi: 10.1016/j.jped.2012.12.002. [DOI] [PubMed] [Google Scholar]
- 85.Da Silva Tanganhito D., Bick D., Chang Y.-S. Breastfeeding Experiences and Perspectives among Women with Postnatal Depression: A Qualitative Evidence Synthesis. Women Birth. 2020;33:231–239. doi: 10.1016/j.wombi.2019.05.012. [DOI] [PubMed] [Google Scholar]
- 86.Lara-Cinisomo S., McKenney K., Di Florio A., Meltzer-Brody S. Associations Between Postpartum Depression, Breastfeeding, and Oxytocin Levels in Latina Mothers. Breastfeed Med. 2017;12:436–442. doi: 10.1089/bfm.2016.0213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Diez-Sampedro A., Flowers M., Olenick M., Maltseva T., Valdes G. Women’s Choice Regarding Breastfeeding and Its Effect on Well-Being. Nurs. Womens Health. 2019;23:383–389. doi: 10.1016/j.nwh.2019.08.002. [DOI] [PubMed] [Google Scholar]
- 88.Silva C.S., Lima M.C., Sequeira-de-Andrade L.A.S., Oliveira J.S., Monteiro J.S., Lima N.M.S., Santos R.M.A.B., Lira P.I.C. Association between Postpartum Depression and the Practice of Exclusive Breastfeeding in the First Three Months of Life. J. Pediatr. Rio J. 2017;93:356–364. doi: 10.1016/j.jped.2016.08.005. [DOI] [PubMed] [Google Scholar]
- 89.Cooklin A.R., Amir L.H., Nguyen C.D., Buck M.L., Cullinane M., Fisher J.R.W., Donath S.M., CASTLE Study Team Physical Health, Breastfeeding Problems and Maternal Mood in the Early Postpartum: A Prospective Cohort Study. Arch. Womens Ment. Health. 2018;21:365–374. doi: 10.1007/s00737-017-0805-y. [DOI] [PubMed] [Google Scholar]
- 90.Lyons K.E., Ryan C.A., Dempsey E.M., Ross R.P., Stanton C. Breast Milk, a Source of Beneficial Microbes and Associated Benefits for Infant Health. Nutrients. 2020;12:39. doi: 10.3390/nu12041039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Rantasalo I., Kauppinen M.A. The Occurrence of Staphylococcus Aureus in Mother’s Milk. Ann. Chir. Gynaecol. Fenn. 1959;48:246–258. [PubMed] [Google Scholar]
- 92.Ojo-Okunola A., Nicol M., du Toit E. Human Breast Milk Bacteriome in Health and Disease. Nutrients. 2018;10:1643. doi: 10.3390/nu10111643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Martín R., Langa S., Reviriego C., Jimínez E., Marín M.L., Xaus J., Fernández L., Rodríguez J.M. Human Milk Is a Source of Lactic Acid Bacteria for the Infant Gut. J. Pediatr. 2003;143:754–758. doi: 10.1016/j.jpeds.2003.09.028. [DOI] [PubMed] [Google Scholar]
- 94.Civardi E., Garofoli F., Tzialla C., Paolillo P., Bollani L., Stronati M. Microorganisms in Human Milk: Lights and Shadows. J. Matern. Fetal. Neonatal. Med. 2013;26(Suppl. 2):30–34. doi: 10.3109/14767058.2013.829693. [DOI] [PubMed] [Google Scholar]
- 95.Ruiz L., García-Carral C., Rodriguez J.M. Unfolding the Human Milk Microbiome Landscape in the Omics Era. Front. Microbiol. 2019;10:1378. doi: 10.3389/fmicb.2019.01378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Moossavi S., Sepehri S., Robertson B., Bode L., Goruk S., Field C.J., Lix L.M., de Souza R.J., Becker A.B., Mandhane P.J., et al. Composition and Variation of the Human Milk Microbiota Are Influenced by Maternal and Early-Life Factors. Cell Host Microbe. 2019;25:324–335.e4. doi: 10.1016/j.chom.2019.01.011. [DOI] [PubMed] [Google Scholar]
- 97.Damaceno Q.S., Souza J.P., Nicoli J.R., Paula R.L., Assis G.B., Figueiredo H.C., Azevedo V., Martins F.S. Evaluation of Potential Probiotics Isolated from Human Milk and Colostrum. Probiotics Antimicrob. Proteins. 2017;9:371–379. doi: 10.1007/s12602-017-9270-1. [DOI] [PubMed] [Google Scholar]
- 98.Rodríguez J.M. The Origin of Human Milk Bacteria: Is There a Bacterial Entero-Mammary Pathway during Late Pregnancy and Lactation? Adv. Nutr. 2014;5:779–784. doi: 10.3945/an.114.007229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Perez P.F., Doré J., Leclerc M., Levenez F., Benyacoub J., Serrant P., Segura-Roggero I., Schiffrin E.J., Donnet-Hughes A. Bacterial Imprinting of the Neonatal Immune System: Lessons from Maternal Cells? Pediatrics. 2007;119:e724–e732. doi: 10.1542/peds.2006-1649. [DOI] [PubMed] [Google Scholar]
- 100.Rescigno M., Urbano M., Valzasina B., Francolini M., Rotta G., Bonasio R., Granucci F., Kraehenbuhl J.P., Ricciardi-Castagnoli P. Dendritic Cells Express Tight Junction Proteins and Penetrate Gut Epithelial Monolayers to Sample Bacteria. Nat. Immunol. 2001;2:361–367. doi: 10.1038/86373. [DOI] [PubMed] [Google Scholar]
- 101.Geddes D.T. The Use of Ultrasound to Identify Milk Ejection in Women—Tips and Pitfalls. Int. Breastfeed J. 2009;4:5. doi: 10.1186/1746-4358-4-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Hunt K.M., Foster J.A., Forney L.J., Schütte U.M.E., Beck D.L., Abdo Z., Fox L.K., Williams J.E., McGuire M.K., McGuire M.A. Characterization of the Diversity and Temporal Stability of Bacterial Communities in Human Milk. PLoS ONE. 2011;6:e21313. doi: 10.1371/journal.pone.0021313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Pannaraj P.S., Li F., Cerini C., Bender J.M., Yang S., Rollie A., Adisetiyo H., Zabih S., Lincez P.J., Bittinger K., et al. Association Between Breast Milk Bacterial Communities and Establishment and Development of the Infant Gut Microbiome. JAMA Pediatr. 2017;171:647–654. doi: 10.1001/jamapediatrics.2017.0378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Demmelmair H., Jiménez E., Collado M.C., Salminen S., McGuire M.K. Maternal and Perinatal Factors Associated with the Human Milk Microbiome. Curr. Dev. Nutr. 2020;4:nzaa027. doi: 10.1093/cdn/nzaa027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Urbaniak C., Cummins J., Brackstone M., Macklaim J.M., Gloor G.B., Baban C.K., Scott L., O’Hanlon D.M., Burton J.P., Francis K.P., et al. Microbiota of Human Breast Tissue. Appl. Environ. Microbiol. 2014;80:3007–3014. doi: 10.1128/AEM.00242-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Hieken T.J., Chen J., Hoskin T.L., Walther-Antonio M., Johnson S., Ramaker S., Xiao J., Radisky D.C., Knutson K.L., Kalari K.R., et al. The Microbiome of Aseptically Collected Human Breast Tissue in Benign and Malignant Disease. Sci. Rep. 2016;6:30751. doi: 10.1038/srep30751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Chan A.A., Bashir M., Rivas M.N., Duvall K., Sieling P.A., Pieber T.R., Vaishampayan P.A., Love S.M., Lee D.J. Characterization of the Microbiome of Nipple Aspirate Fluid of Breast Cancer Survivors. Sci. Rep. 2016;6:28061. doi: 10.1038/srep28061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Going J.J., Moffat D.F. Escaping from Flatland: Clinical and Biological Aspects of Human Mammary Duct Anatomy in Three Dimensions. J. Pathol. 2004;203:538–544. doi: 10.1002/path.1556. [DOI] [PubMed] [Google Scholar]
- 109.Le Doare K., Holder B., Bassett A., Pannaraj P.S. Mother’s Milk: A Purposeful Contribution to the Development of the Infant Microbiota and Immunity. Front. Immunol. 2018;9:361. doi: 10.3389/fimmu.2018.00361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Jeurink P.V., van Bergenhenegouwen J., Jiménez E., Knippels L.M.J., Fernández L., Garssen J., Knol J., Rodríguez J.M., Martín R. Human Milk: A Source of More Life than We Imagine. Benef. Microbes. 2013;4:17–30. doi: 10.3920/BM2012.0040. [DOI] [PubMed] [Google Scholar]
- 111.Jost T., Lacroix C., Braegger C.P., Rochat F., Chassard C. Vertical Mother-Neonate Transfer of Maternal Gut Bacteria via Breastfeeding. Environ. Microbiol. 2014;16:2891–2904. doi: 10.1111/1462-2920.12238. [DOI] [PubMed] [Google Scholar]
- 112.Togo A., Dufour J.-C., Lagier J.-C., Dubourg G., Raoult D., Million M. Repertoire of Human Breast and Milk Microbiota: A Systematic Review. Future Microbiol. 2019;14:623–641. doi: 10.2217/fmb-2018-0317. [DOI] [PubMed] [Google Scholar]
- 113.Latuga M.S., Stuebe A., Seed P.C. A Review of the Source and Function of Microbiota in Breast Milk. Semin. Reprod. Med. 2014;32:68–73. doi: 10.1055/s-0033-1361824. [DOI] [PubMed] [Google Scholar]
- 114.Fitzstevens J.L., Smith K.C., Hagadorn J.I., Caimano M.J., Matson A.P., Brownell E.A. Systematic Review of the Human Milk Microbiota. Nutr. Clin. Pract. 2017;32:354–364. doi: 10.1177/0884533616670150. [DOI] [PubMed] [Google Scholar]
- 115.Cabrera-Rubio R., Collado M.C., Laitinen K., Salminen S., Isolauri E., Mira A. The Human Milk Microbiome Changes over Lactation and Is Shaped by Maternal Weight and Mode of Delivery. Am. J. Clin. Nutr. 2012;96:544–551. doi: 10.3945/ajcn.112.037382. [DOI] [PubMed] [Google Scholar]
- 116.Amir L.H., Donath S.M., Garland S.M., Tabrizi S.N., Bennett C.M., Cullinane M., Payne M.S. Does Candida and/or Staphylococcus Play a Role in Nipple and Breast Pain in Lactation? A Cohort Study in Melbourne, Australia. BMJ Open. 2013;3 doi: 10.1136/bmjopen-2012-002351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Jiménez E., de Andrés J., Manrique M., Pareja-Tobes P., Tobes R., Martínez-Blanch J.F., Codoñer F.M., Ramón D., Fernández L., Rodríguez J.M. Metagenomic Analysis of Milk of Healthy and Mastitis-Suffering Women. J. Hum. Lact. 2015;31:406–415. doi: 10.1177/0890334415585078. [DOI] [PubMed] [Google Scholar]
- 118.Moossavi S., Fehr K., Derakhshani H., Sbihi H., Robertson B., Bode L., Brook J., Turvey S.E., Moraes T.J., Becker A.B., et al. Human Milk Fungi: Environmental Determinants and Inter-Kingdom Associations with Milk Bacteria in the CHILD Cohort Study. BMC Microbiol. 2020;20:146. doi: 10.1186/s12866-020-01829-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Dinleyici M., Pérez-Brocal V., Arslanoglu S., Aydemir O., Ozumut S.S., Tekin N., Vandenplas Y., Moya A., Dinleyici E.C. Human Milk Mycobiota Composition: Relationship with Gestational Age, Delivery Mode, and Birth Weight. Benef. Microbes. 2020;11:151–162. doi: 10.3920/BM2019.0158. [DOI] [PubMed] [Google Scholar]
- 120.Liang G., Zhao C., Zhang H., Mattei L., Sherrill-Mix S., Bittinger K., Kessler L.R., Wu G.D., Baldassano R.N., DeRusso P., et al. The Stepwise Assembly of the Neonatal Virome Is Modulated by Breastfeeding. Nature. 2020;581:470–474. doi: 10.1038/s41586-020-2192-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Pannaraj P.S., Ly M., Cerini C., Saavedra M., Aldrovandi G.M., Saboory A.A., Johnson K.M., Pride D.T. Shared and Distinct Features of Human Milk and Infant Stool Viromes. Front. Microbiol. 2018;9:1162. doi: 10.3389/fmicb.2018.01162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Mohandas S., Pannaraj P.S. Beyond the Bacterial Microbiome: Virome of Human Milk and Effects on the Developing Infant. Milk Mucosal Immun. Microbiome Impact Neonate. 2020;94:86–93. doi: 10.1159/000504997. [DOI] [PubMed] [Google Scholar]
- 123.Khodayar-Pardo P., Mira-Pascual L., Collado M.C., Martínez-Costa C. Impact of Lactation Stage, Gestational Age and Mode of Delivery on Breast Milk Microbiota. J. Perinatol. 2014;34:599–605. doi: 10.1038/jp.2014.47. [DOI] [PubMed] [Google Scholar]
- 124.Decker E., Engelmann G., Findeisen A., Gerner P., Laass M., Ney D., Posovszky C., Hoy L., Hornef M.W. Cesarean Delivery Is Associated with Celiac Disease but Not Inflammatory Bowel Disease in Children. Pediatrics. 2010;125:e1433–e1440. doi: 10.1542/peds.2009-2260. [DOI] [PubMed] [Google Scholar]
- 125.Urbaniak C., Angelini M., Gloor G.B., Reid G. Human Milk Microbiota Profiles in Relation to Birthing Method, Gestation and Infant Gender. Microbiome. 2016;4:1. doi: 10.1186/s40168-015-0145-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Chu D.M., Ma J., Prince A.L., Antony K.M., Seferovic M.D., Aagaard K.M. Maturation of the Infant Microbiome Community Structure and Function across Multiple Body Sites and in Relation to Mode of Delivery. Nat. Med. 2017;23:314–326. doi: 10.1038/nm.4272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Gérard P. Gut Microbiota and Obesity. Cell. Mol. Life Sci. 2016;73:147–162. doi: 10.1007/s00018-015-2061-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Olivares M., Albrecht S., De Palma G., Ferrer M.D., Castillejo G., Schols H.A., Sanz Y. Human Milk Composition Differs in Healthy Mothers and Mothers with Celiac Disease. Eur. J. Nutr. 2015;54:119–128. doi: 10.1007/s00394-014-0692-1. [DOI] [PubMed] [Google Scholar]
- 129.Valitutti F., Cucchiara S., Fasano A. Celiac Disease and the Microbiome. Nutrients. 2019;11:2403. doi: 10.3390/nu11102403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Hermansson H., Kumar H., Collado M.C., Salminen S., Isolauri E., Rautava S. Breast Milk Microbiota Is Shaped by Mode of Delivery and Intrapartum Antibiotic Exposure. Front. Nutr. 2019;6:4. doi: 10.3389/fnut.2019.00004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Browne P.D., Aparicio M., Alba C., Hechler C., Beijers R., Rodríguez J.M., Fernández L., de Weerth C. Human Milk Microbiome and Maternal Postnatal Psychosocial Distress. Front. Microbiol. 2019;10:2333. doi: 10.3389/fmicb.2019.02333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Kumar H., du Toit E., Kulkarni A., Aakko J., Linderborg K.M., Zhang Y., Nicol M.P., Isolauri E., Yang B., Collado M.C., et al. Distinct Patterns in Human Milk Microbiota and Fatty Acid Profiles Across Specific Geographic Locations. Front. Microbiol. 2016;7:1619. doi: 10.3389/fmicb.2016.01619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Boix-Amorós A., Puente-Sánchez F., du Toit E., Linderborg K.M., Zhang Y., Yang B., Salminen S., Isolauri E., Tamames J., Mira A., et al. Mycobiome Profiles in Breast Milk from Healthy Women Depend on Mode of Delivery, Geographic Location, and Interaction with Bacteria. Appl. Environ. Microbiol. 2019;85 doi: 10.1128/AEM.02994-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Ayoub Moubareck C., Lootah M., Tahlak M., Venema K. Profiles of Human Milk Oligosaccharides and Their Relations to the Milk Microbiota of Breastfeeding Mothers in Dubai. Nutrients. 2020;12:1727. doi: 10.3390/nu12061727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Soto A., Martín V., Jiménez E., Mader I., Rodríguez J.M., Fernández L. Lactobacilli and Bifidobacteria in Human Breast Milk: Influence of Antibiotherapy and Other Host and Clinical Factors. J. Pediatric Gastroenterol. Nutr. 2014;59:78–88. doi: 10.1097/MPG.0000000000000347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Zimmermann P., Curtis N. Breast Milk Microbiota: A Review of the Factors That Influence Composition. J. Infect. 2020;81:17–47. doi: 10.1016/j.jinf.2020.01.023. [DOI] [PubMed] [Google Scholar]
- 137.Williams J.E., Carrothers J.M., Lackey K.A., Beatty N.F., York M.A., Brooker S.L., Shafii B., Price W.J., Settles M.L., McGuire M.A., et al. Human Milk Microbial Community Structure Is Relatively Stable and Related to Variations in Macronutrient and Micronutrient Intakes in Healthy Lactating Women. J. Nutr. 2017;147:1739–1748. doi: 10.3945/jn.117.248864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Carrothers J.M., York M.A., Brooker S.L., Lackey K.A., Williams J.E., Shafii B., Price W.J., Settles M.L., McGuire M.A., McGuire M.K. Fecal Microbial Community Structure Is Stable over Time and Related to Variation in Macronutrient and Micronutrient Intakes in Lactating Women123. J. Nutr. 2015;145:2379–2388. doi: 10.3945/jn.115.211110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Padilha M., Danneskiold-Samsøe N.B., Brejnrod A., Hoffmann C., Cabral V.P., Iaucci J.d.M., Sales C.H., Fisberg R.M., Cortez R.V., Brix S., et al. The Human Milk Microbiota Is Modulated by Maternal Diet. Microorganisms. 2019;7:502. doi: 10.3390/microorganisms7110502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Miyake Y., Sasaki S., Tanaka K., Hirota Y. Consumption of Vegetables, Fruit, and Antioxidants during Pregnancy and Wheeze and Eczema in Infants. Allergy. 2010;65:758–765. doi: 10.1111/j.1398-9995.2009.02267.x. [DOI] [PubMed] [Google Scholar]
- 141.Coakley M., Ross R.P., Nordgren M., Fitzgerald G., Devery R., Stanton C. Conjugated Linoleic Acid Biosynthesis by Human-Derived Bifidobacterium Species. J. Appl. Microbiol. 2003;94:138–145. doi: 10.1046/j.1365-2672.2003.01814.x. [DOI] [PubMed] [Google Scholar]
- 142.Shively C.A., Register T.C., Appt S.E., Clarkson T.B., Uberseder B., Clear K.Y.J., Wilson A.S., Chiba A., Tooze J.A., Cook K.L. Consumption of Mediterranean versus Western Diet Leads to Distinct Mammary Gland Microbiome Populations. Cell Rep. 2018;25:47–56.e3. doi: 10.1016/j.celrep.2018.08.078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Zhang Z., Lv J., Pan L., Zhang Y. Roles and Applications of Probiotic Lactobacillus Strains. Appl. Microbiol. Biotechnol. 2018;102:8135–8143. doi: 10.1007/s00253-018-9217-9. [DOI] [PubMed] [Google Scholar]
- 144.Bäckhed F., Roswall J., Peng Y., Feng Q., Jia H., Kovatcheva-Datchary P., Li Y., Xia Y., Xie H., Zhong H., et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe. 2015;17:690–703. doi: 10.1016/j.chom.2015.04.004. [DOI] [PubMed] [Google Scholar]
- 145.Babakobi M.D., Reshef L., Gihaz S., Belgorodsky B., Fishman A., Bujanover Y., Gophna U. Effect of Maternal Diet and Milk Lipid Composition on the Infant Gut and Maternal Milk Microbiomes. Nutrients. 2020;12:2539. doi: 10.3390/nu12092539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Turroni F., Milani C., Duranti S., Mancabelli L., Mangifesta M., Viappiani A., Lugli G.A., Ferrario C., Gioiosa L., Ferrarini A., et al. Deciphering Bifidobacterial-Mediated Metabolic Interactions and Their Impact on Gut Microbiota by a Multi-Omics Approach. ISME J. 2016;10:1656–1668. doi: 10.1038/ismej.2015.236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Simpson M.R., Avershina E., Storrø O., Johnsen R., Rudi K., Øien T. Breastfeeding-Associated Microbiota in Human Milk Following Supplementation with Lactobacillus Rhamnosus GG, Lactobacillus Acidophilus La-5, and Bifidobacterium Animalis Ssp. Lactis Bb-12. J. Dairy Sci. 2018;101:889–899. doi: 10.3168/jds.2017-13411. [DOI] [PubMed] [Google Scholar]
- 148.Rautava S. Early Microbial Contact, the Breast Milk Microbiome and Child Health. J. Dev. Orig. Health Dis. 2016;7:5–14. doi: 10.1017/S2040174415001233. [DOI] [PubMed] [Google Scholar]
- 149.Gueimonde M., Laitinen K., Salminen S., Isolauri E. Breast Milk: A Source of Bifidobacteria for Infant Gut Development and Maturation? Neonatology. 2007;92:64–66. doi: 10.1159/000100088. [DOI] [PubMed] [Google Scholar]
- 150.Pacheco A.R., Barile D., Underwood M.A., Mills D.A. The Impact of the Milk Glycobiome on the Neonate Gut Microbiota. Annu. Rev. Anim. Biosci. 2015;3:419–445. doi: 10.1146/annurev-animal-022114-111112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Turroni F., Peano C., Pass D.A., Foroni E., Severgnini M., Claesson M.J., Kerr C., Hourihane J., Murray D., Fuligni F., et al. Diversity of Bifidobacteria within the Infant Gut Microbiota. PLoS ONE. 2012;7:e36957. doi: 10.1371/journal.pone.0036957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Turroni F., Foroni E., Pizzetti P., Giubellini V., Ribbera A., Merusi P., Cagnasso P., Bizzarri B., de’Angelis G.L., Shanahan F., et al. Exploring the Diversity of the Bifidobacterial Population in the Human Intestinal Tract. Appl. Environ. Microbiol. 2009;75:1534–1545. doi: 10.1128/AEM.02216-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Chong C.Y.L., Bloomfield F.H., O’Sullivan J.M. Factors Affecting Gastrointestinal Microbiome Development in Neonates. Nutrients. 2018;10:274. doi: 10.3390/nu10030274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Solís G., de Los Reyes-Gavilan C.G., Fernández N., Margolles A., Gueimonde M. Establishment and Development of Lactic Acid Bacteria and Bifidobacteria Microbiota in Breast-Milk and the Infant Gut. Anaerobe. 2010;16:307–310. doi: 10.1016/j.anaerobe.2010.02.004. [DOI] [PubMed] [Google Scholar]
- 155.Bergmann H., Rodríguez J.M., Salminen S., Szajewska H. Probiotics in Human Milk and Probiotic Supplementation in Infant Nutrition: A Workshop Report. Br. J. Nutr. 2014;112:1119–1128. doi: 10.1017/S0007114514001949. [DOI] [PubMed] [Google Scholar]
- 156.Jiao X., Fu M.-D., Wang Y.-Y., Xue J., Zhang Y. Bifidobacterium and Lactobacillus for Preventing Necrotizing Enterocolitis in Very-Low-Birth-Weight Preterm Infants: A Systematic Review and Meta-Analysis. World J. Pediatr. 2020;16:135–142. doi: 10.1007/s12519-019-00297-5. [DOI] [PubMed] [Google Scholar]
- 157.Jiménez E., Delgado S., Maldonado A., Arroyo R., Albújar M., García N., Jariod M., Fernández L., Gómez A., Rodríguez J.M. Staphylococcus Epidermidis: A Differential Trait of the Fecal Microbiota of Breast-Fed Infants. BMC Microbiol. 2008;8:143. doi: 10.1186/1471-2180-8-143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Landete J.M., Peirotén Á., Medina M., Arqués J.L., Rodríguez-Mínguez E. Virulence and Antibiotic Resistance of Enterococci Isolated from Healthy Breastfed Infants. Microb. Drug Resist. 2018;24:63–69. doi: 10.1089/mdr.2016.0320. [DOI] [PubMed] [Google Scholar]
- 159.Martín V., Maldonado A., Fernández L., Rodríguez J.M., Connor R.I. Inhibition of Human Immunodeficiency Virus Type 1 by Lactic Acid Bacteria from Human Breastmilk. Breastfeed Med. 2010;5:153–158. doi: 10.1089/bfm.2010.0001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Milani C., Duranti S., Bottacini F., Casey E., Turroni F., Mahony J., Belzer C., Delgado Palacio S., Arboleya Montes S., Mancabelli L., et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017;81 doi: 10.1128/MMBR.00036-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria; Proceedings of the Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria; Cordoba, Argentina. 1–4 October 2001. [Google Scholar]
- 162.Hill C., Guarner F., Reid G., Gibson G.R., Merenstein D.J., Pot B., Morelli L., Canani R.B., Flint H.J., Salminen S., et al. Expert Consensus Document. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014;11:506–514. doi: 10.1038/nrgastro.2014.66. [DOI] [PubMed] [Google Scholar]
- 163.Bozzi Cionci N., Baffoni L., Gaggìa F., Di Gioia D. Therapeutic Microbiology: The Role of Bifidobacterium Breve as Food Supplement for the Prevention/Treatment of Paediatric Diseases. Nutrients. 2018;10:1723. doi: 10.3390/nu10111723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Halloran K., Underwood M.A. Probiotic Mechanisms of Action. Early Hum. Dev. 2019;135:58–65. doi: 10.1016/j.earlhumdev.2019.05.010. [DOI] [PubMed] [Google Scholar]
- 165.Bermudez-Brito M., Plaza-Díaz J., Muñoz-Quezada S., Gómez-Llorente C., Gil A. Probiotic Mechanisms of Action. Ann. Nutr. Metab. 2012;61:160–174. doi: 10.1159/000342079. [DOI] [PubMed] [Google Scholar]
- 166.Montalban-Arques A., De Schryver P., Bossier P., Gorkiewicz G., Mulero V., Gatlin D.M., Galindo-Villegas J. Selective Manipulation of the Gut Microbiota Improves Immune Status in Vertebrates. Front. Immunol. 2015;6:512. doi: 10.3389/fimmu.2015.00512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Mu Q., Tavella V.J., Luo X.M. Role of Lactobacillus Reuteri in Human Health and Diseases. Front. Microbiol. 2018;9:757. doi: 10.3389/fmicb.2018.00757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Li L., Fang Z., Liu X., Hu W., Lu W., Lee Y.-K., Zhao J., Zhang H., Chen W. Lactobacillus Reuteri Attenuated Allergic Inflammation Induced by HDM in the Mouse and Modulated Gut Microbes. PLoS ONE. 2020;15:e0231865. doi: 10.1371/journal.pone.0231865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Li S., Qi C., Zhu H., Yu R., Xie C., Peng Y., Yin S.-W., Fan J., Zhao S., Sun J. Lactobacillus Reuteri Improves Gut Barrier Function and Affects Diurnal Variation of the Gut Microbiota in Mice Fed a High-Fat Diet. Food Funct. 2019;10:4705–4715. doi: 10.1039/C9FO00417C. [DOI] [PubMed] [Google Scholar]
- 170.Savino F., Ceratto S., Poggi E., Cartosio M.E., Cordero di Montezemolo L., Giannattasio A. Preventive Effects of Oral Probiotic on Infantile Colic: A Prospective, Randomised, Blinded, Controlled Trial Using Lactobacillus Reuteri DSM 17938. Benef. Microbes. 2015;6:245–251. doi: 10.3920/BM2014.0090. [DOI] [PubMed] [Google Scholar]
- 171.López-Huertas E. Safety and Efficacy of Human Breast Milk Lactobacillus Fermentum CECT 5716. A Mini-Review of Studies with Infant Formulae. Benef. Microbes. 2015;6:219–224. doi: 10.3920/BM2014.0091. [DOI] [PubMed] [Google Scholar]
- 172.Asan-Ozusaglam M., Gunyakti A. Lactobacillus Fermentum Strains from Human Breast Milk with Probiotic Properties and Cholesterol-Lowering Effects. Food Sci. Biotechnol. 2019;28:501–509. doi: 10.1007/s10068-018-0494-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Pérez-Cano F.J., Dong H., Yaqoob P. In Vitro Immunomodulatory Activity of Lactobacillus Fermentum CECT5716 and Lactobacillus Salivarius CECT5713: Two Probiotic Strains Isolated from Human Breast Milk. Immunobiology. 2010;215:996–1004. doi: 10.1016/j.imbio.2010.01.004. [DOI] [PubMed] [Google Scholar]
- 174.Riaz Rajoka M.S., Zhao H., Lu Y., Lian Z., Li N., Hussain N., Shao D., Jin M., Li Q., Shi J. Anticancer Potential against Cervix Cancer (HeLa) Cell Line of Probiotic Lactobacillus Casei and Lactobacillus Paracasei Strains Isolated from Human Breast Milk. Food Funct. 2018;9:2705–2715. doi: 10.1039/C8FO00547H. [DOI] [PubMed] [Google Scholar]
- 175.Riaz Rajoka M.S., Zhao H., Mehwish H.M., Li N., Lu Y., Lian Z., Shao D., Jin M., Li Q., Zhao L., et al. Anti-Tumor Potential of Cell Free Culture Supernatant of Lactobacillus Rhamnosus Strains Isolated from Human Breast Milk. Food Res. Int. 2019;123:286–297. doi: 10.1016/j.foodres.2019.05.002. [DOI] [PubMed] [Google Scholar]
- 176.Guarino A., Guandalini S., Lo Vecchio A. Probiotics for Prevention and Treatment of Diarrhea. J. Clin. Gastroenterol. 2015;49(Suppl. 1):S37–S45. doi: 10.1097/MCG.0000000000000349. [DOI] [PubMed] [Google Scholar]
- 177.Maldonado J., Gil-Campos M., Maldonado-Lobón J.A., Benavides M.R., Flores-Rojas K., Jaldo R., Jiménez Del Barco I., Bolívar V., Valero A.D., Prados E., et al. Evaluation of the Safety, Tolerance and Efficacy of 1-Year Consumption of Infant Formula Supplemented with Lactobacillus Fermentum CECT5716 Lc40 or Bifidobacterium Breve CECT7263: A Randomized Controlled Trial. BMC Pediatr. 2019;19:361. doi: 10.1186/s12887-019-1753-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Fahmy C.A., Gamal-Eldeen A.M., El-Hussieny E.A., Raafat B.M., Mehanna N.S., Talaat R.M., Shaaban M.T. Bifidobacterium Longum Suppresses Murine Colorectal Cancer through the Modulation of OncomiRs and Tumor Suppressor MiRNAs. Nutr. Cancer. 2019;71:688–700. doi: 10.1080/01635581.2019.1577984. [DOI] [PubMed] [Google Scholar]
- 179.Park M.S., Kwon B., Ku S., Ji G.E. The Efficacy of Bifidobacterium Longum BORI and Lactobacillus Acidophilus AD031 Probiotic Treatment in Infants with Rotavirus Infection. Nutrients. 2017;9:887. doi: 10.3390/nu9080887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Khalkhali S., Mojgani N. In Vitro and in Vivo Safety Analysis of Enterococcus Faecium 2C Isolated from Human Breast Milk. Microb. Pathog. 2018;116:73–77. doi: 10.1016/j.micpath.2018.01.012. [DOI] [PubMed] [Google Scholar]
- 181.Khalkhali S., Mojgani N. Bacteriocinogenic Potential and Virulence Traits of Enterococcus Faecium and E. Faecalis Isolated from Human Milk. Iran. J. Microbiol. 2017;9:224–233. [PMC free article] [PubMed] [Google Scholar]
- 182.Hassan Z., Mustafa S., Rahim R.A., Isa N.M. Anti-Breast Cancer Effects of Live, Heat-Killed and Cytoplasmic Fractions of Enterococcus Faecalis and Staphylococcus Hominis Isolated from Human Breast Milk. Cell. Dev. Biol. Anim. 2016;52:337–348. doi: 10.1007/s11626-015-9978-8. [DOI] [PubMed] [Google Scholar]
- 183.Wiciński M., Sawicka E., Gębalski J., Kubiak K., Malinowski B. Human Milk Oligosaccharides: Health Benefits, Potential Applications in Infant Formulas, and Pharmacology. Nutrients. 2020;12:266. doi: 10.3390/nu12010266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Bode L. Human Milk Oligosaccharides: Every Baby Needs a Sugar Mama. Glycobiology. 2012;22:1147–1162. doi: 10.1093/glycob/cws074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Cheng L., Akkerman R., Kong C., Walvoort M.T.C., de Vos P. More than Sugar in the Milk: Human Milk Oligosaccharides as Essential Bioactive Molecules in Breast Milk and Current Insight in Beneficial Effects. Crit. Rev. Food Sci. Nutr. 2020:1–17. doi: 10.1080/10408398.2020.1754756. [DOI] [PubMed] [Google Scholar]
- 186.Thurl S., Munzert M., Boehm G., Matthews C., Stahl B. Systematic Review of the Concentrations of Oligosaccharides in Human Milk. Nutr. Rev. 2017;75:920–933. doi: 10.1093/nutrit/nux044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Gabrielli O., Zampini L., Galeazzi T., Padella L., Santoro L., Peila C., Giuliani F., Bertino E., Fabris C., Coppa G.V. Preterm Milk Oligosaccharides during the First Month of Lactation. Pediatrics. 2011;128:e1520–e1531. doi: 10.1542/peds.2011-1206. [DOI] [PubMed] [Google Scholar]
- 188.Nakhla T., Fu D., Zopf D., Brodsky N.L., Hurt H. Neutral Oligosaccharide Content of Preterm Human Milk. Br. J. Nutr. 1999;82:361–367. doi: 10.1017/S0007114599001609. [DOI] [PubMed] [Google Scholar]
- 189.Bering S.B. Human Milk Oligosaccharides to Prevent Gut Dysfunction and Necrotizing Enterocolitis in Preterm Neonates. Nutrients. 2018;10:1461. doi: 10.3390/nu10101461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Smilowitz J.T., Lebrilla C.B., Mills D.A., German J.B., Freeman S.L. Breast Milk Oligosaccharides: Structure-Function Relationships in the Neonate. Annu. Rev. Nutr. 2014;34:143–169. doi: 10.1146/annurev-nutr-071813-105721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Sprenger G.A., Baumgärtner F., Albermann C. Production of Human Milk Oligosaccharides by Enzymatic and Whole-Cell Microbial Biotransformations. J. Biotechnol. 2017;258:79–91. doi: 10.1016/j.jbiotec.2017.07.030. [DOI] [PubMed] [Google Scholar]
- 192.Plaza-Díaz J., Fontana L., Gil A. Human Milk Oligosaccharides and Immune System Development. Nutrients. 2018;10:1038. doi: 10.3390/nu10081038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Bode L., Jantscher-Krenn E. Structure-Function Relationships of Human Milk Oligosaccharides. Adv. Nutr. 2012;3:383S–391S. doi: 10.3945/an.111.001404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Totten S.M., Zivkovic A.M., Wu S., Ngyuen U., Freeman S.L., Ruhaak L.R., Darboe M.K., German J.B., Prentice A.M., Lebrilla C.B. Comprehensive Profiles of Human Milk Oligosaccharides Yield Highly Sensitive and Specific Markers for Determining Secretor Status in Lactating Mothers. J. Proteome Res. 2012;11:6124–6133. doi: 10.1021/pr300769g. [DOI] [PubMed] [Google Scholar]
- 195.McGuire M.K., Meehan C.L., McGuire M.A., Williams J.E., Foster J., Sellen D.W., Kamau-Mbuthia E.W., Kamundia E.W., Mbugua S., Moore S.E., et al. What’s Normal? Oligosaccharide Concentrations and Profiles in Milk Produced by Healthy Women Vary Geographically. Am. J. Clin. Nutr. 2017;105:1086–1100. doi: 10.3945/ajcn.116.139980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Denomme G.A. Molecular Basis of Blood Group Expression. Transfus. Apher. Sci. 2011;44:53–63. doi: 10.1016/j.transci.2010.12.010. [DOI] [PubMed] [Google Scholar]
- 197.Ewald D.R., Sumner S.C.J. Blood Type Biochemistry and Human Disease. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016;8:517–535. doi: 10.1002/wsbm.1355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Wacklin P., Mäkivuokko H., Alakulppi N., Nikkilä J., Tenkanen H., Räbinä J., Partanen J., Aranko K., Mättö J. Secretor Genotype (FUT2 Gene) Is Strongly Associated with the Composition of Bifidobacteria in the Human Intestine. PLoS ONE. 2011;6:e20113. doi: 10.1371/journal.pone.0020113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Tonon K.M., de Morais M.B., Abrão A.C.F.V., Miranda A., Morais T.B. Maternal and Infant Factors Associated with Human Milk Oligosaccharides Concentrations According to Secretor and Lewis Phenotypes. Nutrients. 2019;11:1358. doi: 10.3390/nu11061358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Kobata A. Structures and Application of Oligosaccharides in Human Milk. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010;86:731–747. doi: 10.2183/pjab.86.731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Thurl S., Henker J., Siegel M., Tovar K., Sawatzki G. Detection of Four Human Milk Groups with Respect to Lewis Blood Group Dependent Oligosaccharides. Glycoconj. J. 1997;14:795–799. doi: 10.1023/A:1018529703106. [DOI] [PubMed] [Google Scholar]
- 202.Stahl B., Thurl S., Henker J., Siegel M., Finke B., Sawatzki G. Detection of Four Human Milk Groups with Respect to Lewis-Blood-Group-Dependent Oligosaccharides by Serologic and Chromatographic Analysis. Adv. Exp. Med. Biol. 2001;501:299–306. doi: 10.1007/978-1-4615-1371-1_37. [DOI] [PubMed] [Google Scholar]
- 203.Kulinich A., Liu L. Human Milk Oligosaccharides: The Role in the Fine-Tuning of Innate Immune Responses. Carbohydr. Res. 2016;432:62–70. doi: 10.1016/j.carres.2016.07.009. [DOI] [PubMed] [Google Scholar]
- 204.Garrido D., Ruiz-Moyano S., Lemay D.G., Sela D.A., German J.B., Mills D.A. Comparative Transcriptomics Reveals Key Differences in the Response to Milk Oligosaccharides of Infant Gut-Associated Bifidobacteria. Sci. Rep. 2015;5:13517. doi: 10.1038/srep13517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Gotoh A., Katoh T., Sakanaka M., Ling Y., Yamada C., Asakuma S., Urashima T., Tomabechi Y., Katayama-Ikegami A., Kurihara S., et al. Sharing of Human Milk Oligosaccharides Degradants within Bifidobacterial Communities in Faecal Cultures Supplemented with Bifidobacterium Bifidum. Sci. Rep. 2018;8:13958. doi: 10.1038/s41598-018-32080-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Sakanaka M., Gotoh A., Yoshida K., Odamaki T., Koguchi H., Xiao J.-Z., Kitaoka M., Katayama T. Varied Pathways of Infant Gut-Associated Bifidobacterium to Assimilate Human Milk Oligosaccharides: Prevalence of the Gene Set and Its Correlation with Bifidobacteria-Rich Microbiota Formation. Nutrients. 2019;12:71. doi: 10.3390/nu12010071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Thongaram T., Hoeflinger J.L., Chow J., Miller M.J. Human Milk Oligosaccharide Consumption by Probiotic and Human-Associated Bifidobacteria and Lactobacilli. J. Dairy Sci. 2017;100:7825–7833. doi: 10.3168/jds.2017-12753. [DOI] [PubMed] [Google Scholar]
- 208.Schwab C., Ruscheweyh H.-J., Bunesova V., Pham V.T., Beerenwinkel N., Lacroix C. Trophic Interactions of Infant Bifidobacteria and Eubacterium Hallii during L-Fucose and Fucosyllactose Degradation. Front. Microbiol. 2017;8:95. doi: 10.3389/fmicb.2017.00095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Yu Z.-T., Nanthakumar N.N., Newburg D.S. The Human Milk Oligosaccharide 2′-Fucosyllactose Quenches Campylobacter Jejuni-Induced Inflammation in Human Epithelial Cells HEp-2 and HT-29 and in Mouse Intestinal Mucosa. J. Nutr. 2016;146:1980–1990. doi: 10.3945/jn.116.230706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Weichert S., Jennewein S., Hüfner E., Weiss C., Borkowski J., Putze J., Schroten H. Bioengineered 2′-Fucosyllactose and 3-Fucosyllactose Inhibit the Adhesion of Pseudomonas Aeruginosa and Enteric Pathogens to Human Intestinal and Respiratory Cell Lines. Nutr. Res. 2013;33:831–838. doi: 10.1016/j.nutres.2013.07.009. [DOI] [PubMed] [Google Scholar]
- 211.Jantscher-Krenn E., Lauwaet T., Bliss L.A., Reed S.L., Gillin F.D., Bode L. Human Milk Oligosaccharides Reduce Entamoeba Histolytica Attachment and Cytotoxicity in Vitro. Br. J. Nutr. 2012;108:1839–1846. doi: 10.1017/S0007114511007392. [DOI] [PubMed] [Google Scholar]
- 212.Iskarpatyoti J.A., Morse E.A., McClung R.P., Ikizler M., Wetzel J.D., Contractor N., Dermody T.S. Serotype-Specific Differences in Inhibition of Reovirus Infectivity by Human-Milk Glycans Are Determined by Viral Attachment Protein Σ1. Virology. 2012;433:489–497. doi: 10.1016/j.virol.2012.08.036. [DOI] [PubMed] [Google Scholar]
- 213.Lin A.E., Autran C.A., Szyszka A., Escajadillo T., Huang M., Godula K., Prudden A.R., Boons G.-J., Lewis A.L., Doran K.S., et al. Human Milk Oligosaccharides Inhibit Growth of Group B Streptococcus. J. Biol. Chem. 2017;292:11243–11249. doi: 10.1074/jbc.M117.789974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Ackerman D.L., Craft K.M., Doster R.S., Weitkamp J.-H., Aronoff D.M., Gaddy J.A., Townsend S.D. Antimicrobial and Antibiofilm Activity of Human Milk Oligosaccharides against Streptococcus Agalactiae, Staphylococcus Aureus, and Acinetobacter Baumannii. ACS Infect. Dis. 2018;4:315–324. doi: 10.1021/acsinfecdis.7b00183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Gonia S., Tuepker M., Heisel T., Autran C., Bode L., Gale C.A. Human Milk Oligosaccharides Inhibit Candida Albicans Invasion of Human Premature Intestinal Epithelial Cells. J. Nutr. 2015;145:1992–1998. doi: 10.3945/jn.115.214940. [DOI] [PubMed] [Google Scholar]
- 216.Donovan S.M., Comstock S.S. Human Milk Oligosaccharides Influence Neonatal Mucosal and Systemic Immunity. Ann. Nutr. Metab. 2016;69(Suppl. 2):42–51. doi: 10.1159/000452818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Triantis V., Bode L., van Neerven R.J.J. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018;6:190. doi: 10.3389/fped.2018.00190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Lane J.A., O’Callaghan J., Carrington S.D., Hickey R.M. Transcriptional Response of HT-29 Intestinal Epithelial Cells to Human and Bovine Milk Oligosaccharides. Br. J. Nutr. 2013;110:2127–2137. doi: 10.1017/S0007114513001591. [DOI] [PubMed] [Google Scholar]
- 219.Newburg D.S., Ko J.S., Leone S., Nanthakumar N.N. Human Milk Oligosaccharides and Synthetic Galactosyloligosaccharides Contain 3′-, 4-, and 6′-Galactosyllactose and Attenuate Inflammation in Human T84, NCM-460, and H4 Cells and Intestinal Tissue Ex Vivo. J. Nutr. 2016;146:358–367. doi: 10.3945/jn.115.220749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Xiao L., van De Worp W.R., Stassen R., van Maastrigt C., Kettelarij N., Stahl B., Blijenberg B., Overbeek S.A., Folkerts G., Garssen J., et al. Human Milk Oligosaccharides Promote Immune Tolerance via Direct Interactions with Human Dendritic Cells. Eur. J. Immunol. 2019;49:1001–1014. doi: 10.1002/eji.201847971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Ayechu-Muruzabal V., Overbeek S.A., Kostadinova A.I., Stahl B., Garssen J., Van’t Land B., Willemsen L.E.M. Exposure of Intestinal Epithelial Cells to 2′-Fucosyllactose and CpG Enhances Galectin Release and Instructs Dendritic Cells to Drive Th1 and Regulatory-Type Immune Development. Biomolecules. 2020;10:784. doi: 10.3390/biom10050784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Johannes L., Jacob R., Leffler H. Galectins at a Glance. J. Cell. Sci. 2018;131 doi: 10.1242/jcs.208884. [DOI] [PubMed] [Google Scholar]
- 223.El-Hawiet A., Chen Y., Shams-Ud-Doha K., Kitova E.N., St-Pierre Y., Klassen J.S. High-Throughput Label- and Immobilization-Free Screening of Human Milk Oligosaccharides Against Lectins. Anal. Chem. 2017;89:8713–8722. doi: 10.1021/acs.analchem.7b00542. [DOI] [PubMed] [Google Scholar]
- 224.Šuligoj T., Vigsnæs L.K., Abbeele P.V.d., Apostolou A., Karalis K., Savva G.M., McConnell B., Juge N. Effects of Human Milk Oligosaccharides on the Adult Gut Microbiota and Barrier Function. Nutrients. 2020;12:2808. doi: 10.3390/nu12092808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Lewis E.D., Richard C., Larsen B.M., Field C.J. The Importance of Human Milk for Immunity in Preterm Infants. Clin. Perinatol. 2017;44:23–47. doi: 10.1016/j.clp.2016.11.008. [DOI] [PubMed] [Google Scholar]
- 226.Kong C., Elderman M., Cheng L., de Haan B.J., Nauta A., de Vos P. Modulation of Intestinal Epithelial Glycocalyx Development by Human Milk Oligosaccharides and Non-Digestible Carbohydrates. Mol. Nutr. Food Res. 2019;63:e1900303. doi: 10.1002/mnfr.201900303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Wu R.Y., Li B., Koike Y., Määttänen P., Miyake H., Cadete M., Johnson-Henry K.C., Botts S.R., Lee C., Abrahamsson T.R., et al. Human Milk Oligosaccharides Increase Mucin Expression in Experimental Necrotizing Enterocolitis. Mol. Nutr. Food Res. 2019;63:e1800658. doi: 10.1002/mnfr.201800658. [DOI] [PubMed] [Google Scholar]
- 228.Gibson G.R., Roberfroid M.B. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995;125:1401–1412. doi: 10.1093/jn/125.6.1401. [DOI] [PubMed] [Google Scholar]
- 229.Miqdady M., Al Mistarihi J., Azaz A., Rawat D. Prebiotics in the Infant Microbiome: The Past, Present, and Future. Pediatric Gastroenterol. Hepatol. Nutr. 2020;23:1–14. doi: 10.5223/pghn.2020.23.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Gibson G.R., Hutkins R., Sanders M.E., Prescott S.L., Reimer R.A., Salminen S.J., Scott K., Stanton C., Swanson K.S., Cani P.D., et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017;14:491–502. doi: 10.1038/nrgastro.2017.75. [DOI] [PubMed] [Google Scholar]
- 231.Reverri E.J., Devitt A.A., Kajzer J.A., Baggs G.E., Borschel M.W. Review of the Clinical Experiences of Feeding Infants Formula Containing the Human Milk Oligosaccharide 2′-Fucosyllactose. Nutrients. 2018;10:1346. doi: 10.3390/nu10101346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Akkerman R., Faas M.M., de Vos P. Non-Digestible Carbohydrates in Infant Formula as Substitution for Human Milk Oligosaccharide Functions: Effects on Microbiota and Gut Maturation. Crit. Rev. Food Sci. Nutr. 2019;59:1486–1497. doi: 10.1080/10408398.2017.1414030. [DOI] [PubMed] [Google Scholar]
- 233.Baumgärtner F., Jurzitza L., Conrad J., Beifuss U., Sprenger G.A., Albermann C. Synthesis of Fucosylated Lacto-N-Tetraose Using Whole-Cell Biotransformation. Bioorg. Med. Chem. 2015;23:6799–6806. doi: 10.1016/j.bmc.2015.10.005. [DOI] [PubMed] [Google Scholar]
- 234.Petschacher B., Nidetzky B. Biotechnological Production of Fucosylated Human Milk Oligosaccharides: Prokaryotic Fucosyltransferases and Their Use in Biocatalytic Cascades or Whole Cell Conversion Systems. J. Biotechnol. 2016;235:61–83. doi: 10.1016/j.jbiotec.2016.03.052. [DOI] [PubMed] [Google Scholar]
- 235.Wejryd E., Martí M., Marchini G., Werme A., Jonsson B., Landberg E., Abrahamsson T.R. Low Diversity of Human Milk Oligosaccharides Is Associated with Necrotising Enterocolitis in Extremely Low Birth Weight Infants. Nutrients. 2018;10:1556. doi: 10.3390/nu10101556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Gómez-Gallego C., Morales J.M., Monleón D., du Toit E., Kumar H., Linderborg K.M., Zhang Y., Yang B., Isolauri E., Salminen S., et al. Human Breast Milk NMR Metabolomic Profile across Specific Geographical Locations and Its Association with the Milk Microbiota. Nutrients. 2018;10:1355. doi: 10.3390/nu10101355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Williams J.E., Price W.J., Shafii B., Yahvah K.M., Bode L., McGuire M.A., McGuire M.K. Relationships Among Microbial Communities, Maternal Cells, Oligosaccharides, and Macronutrients in Human Milk. J. Hum. Lact. 2017;33:540–551. doi: 10.1177/0890334417709433. [DOI] [PubMed] [Google Scholar]
- 238.Cabrera-Rubio R., Kunz C., Rudloff S., García-Mantrana I., Crehuá-Gaudiza E., Martínez-Costa C., Collado M.C. Association of Maternal Secretor Status and Human Milk Oligosaccharides With Milk Microbiota: An Observational Pilot Study. J. Pediatric Gastroenterol. Nutr. 2019;68:256–263. doi: 10.1097/MPG.0000000000002216. [DOI] [PubMed] [Google Scholar]
- 239.Aakko J., Kumar H., Rautava S., Wise A., Autran C., Bode L., Isolauri E., Salminen S. Human Milk Oligosaccharide Categories Define the Microbiota Composition in Human Colostrum. Benef. Microbes. 2017;8:563–567. doi: 10.3920/BM2016.0185. [DOI] [PubMed] [Google Scholar]
- 240.Borewicz K., Gu F., Saccenti E., Arts I.C.W., Penders J., Thijs C., van Leeuwen S.S., Lindner C., Nauta A., van Leusen E., et al. Correlating Infant Faecal Microbiota Composition and Human Milk Oligosaccharide Consumption by Microbiota of One-Month Old Breastfed Infants. Mol. Nutr. Food Res. 2019:e1801214. doi: 10.1002/mnfr.201801214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Sela D.A., Mills D.A. Nursing Our Microbiota: Molecular Linkages between Bifidobacteria and Milk Oligosaccharides. Trends Microbiol. 2010;18:298–307. doi: 10.1016/j.tim.2010.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.James K., Motherway M.O., Bottacini F., van Sinderen D. Bifidobacterium Breve UCC2003 Metabolises the Human Milk Oligosaccharides Lacto-N-Tetraose and Lacto-N-Neo-Tetraose through Overlapping, yet Distinct Pathways. Sci. Rep. 2016;6:38560. doi: 10.1038/srep38560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.James K., O’Connell Motherway M., Penno C., O’Brien R.L., van Sinderen D. Bifidobacterium Breve UCC2003 Employs Multiple Transcriptional Regulators To Control Metabolism of Particular Human Milk Oligosaccharides. Appl. Environ. Microbiol. 2018;84 doi: 10.1128/AEM.02774-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Martin C.R., Ling P.-R., Blackburn G.L. Review of Infant Feeding: Key Features of Breast Milk and Infant Formula. Nutrients. 2016;8:279. doi: 10.3390/nu8050279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Collados-Gómez L., Ferrera-Camacho P., Fernandez-Serrano E., Camacho-Vicente V., Flores-Herrero C., García-Pozo A.M., Jiménez-García R. Randomised Crossover Trial Showed That Using Breast Milk or Sucrose Provided the Same Analgesic Effect in Preterm Infants of at Least 28 Weeks. Acta Paediatr. 2018;107:436–441. doi: 10.1111/apa.14151. [DOI] [PubMed] [Google Scholar]
- 246.Baudesson de Chanville A., Brevaut-Malaty V., Garbi A., Tosello B., Baumstarck K., Gire C. Analgesic Effect of Maternal Human Milk Odor on Premature Neonates: A Randomized Controlled Trial. J. Hum. Lact. 2017;33:300–308. doi: 10.1177/0890334417693225. [DOI] [PubMed] [Google Scholar]
- 247.Zhang S., Su F., Li J., Chen W. The Analgesic Effects of Maternal Milk Odor on Newborns: A Meta-Analysis. Breastfeed Med. 2018;13:327–334. doi: 10.1089/bfm.2017.0226. [DOI] [PubMed] [Google Scholar]
- 248.Berger B., Porta N., Foata F., Grathwohl D., Delley M., Moine D., Charpagne A., Siegwald L., Descombes P., Alliet P., et al. Linking Human Milk Oligosaccharides, Infant Fecal Community Types, and Later Risk To Require Antibiotics. mBio. 2020;11 doi: 10.1128/mBio.03196-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Yu Z.-T., Chen C., Newburg D.S. Utilization of Major Fucosylated and Sialylated Human Milk Oligosaccharides by Isolated Human Gut Microbes. Glycobiology. 2013;23:1281–1292. doi: 10.1093/glycob/cwt065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Zeuner B., Meyer A.S. Enzymatic Transfucosylation for Synthesis of Human Milk Oligosaccharides. Carbohydr. Res. 2020;493:108029. doi: 10.1016/j.carres.2020.108029. [DOI] [PubMed] [Google Scholar]
- 251.Zeuner B., Teze D., Muschiol J., Meyer A.S. Synthesis of Human Milk Oligosaccharides: Protein Engineering Strategies for Improved Enzymatic Transglycosylation. Molecules. 2019;24:33. doi: 10.3390/molecules24112033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 252.Zeuner B., Vuillemin M., Holck J., Muschiol J., Meyer A.S. Loop Engineering of an α-1,3/4-l-Fucosidase for Improved Synthesis of Human Milk Oligosaccharides. Enzyme Microb. Technol. 2018;115:37–44. doi: 10.1016/j.enzmictec.2018.04.008. [DOI] [PubMed] [Google Scholar]
- 253.Zeuner B., Muschiol J., Holck J., Lezyk M., Gedde M.R., Jers C., Mikkelsen J.D., Meyer A.S. Substrate Specificity and Transfucosylation Activity of GH29 α-l-Fucosidases for Enzymatic Production of Human Milk Oligosaccharides. New Biotechnol. 2018;41:34–45. doi: 10.1016/j.nbt.2017.12.002. [DOI] [PubMed] [Google Scholar]
- 254.Guzmán-Rodríguez F., Alatorre-Santamaría S., Gómez-Ruiz L., Rodríguez-Serrano G., García-Garibay M., Cruz-Guerrero A. Employment of Fucosidases for the Synthesis of Fucosylated Oligosaccharides with Biological Potential. Biotechnol. Appl. Biochem. 2019;66:172–191. doi: 10.1002/bab.1714. [DOI] [PubMed] [Google Scholar]
- 255.Hill D.R., Newburg D.S. Clinical Applications of Bioactive Milk Components. Nutr. Rev. 2015;73:463–476. doi: 10.1093/nutrit/nuv009. [DOI] [PMC free article] [PubMed] [Google Scholar]