

Abstract
Cyclopeptides or cyclic peptides are polypeptides formed by ring closing of terminal amino acids. A large number of natural cyclopeptides have been reported to be highly effective against different cancer cells, some of which are renowned for their clinical uses. Compared to linear peptides, cyclopeptides have absolute advantages of structural rigidity, biochemical stability, binding affinity as well as membrane permeability, which contribute greatly to their anticancer potency. Therefore, the discovery and development of natural cyclopeptides as anticancer agents remains attractive to academic researchers and pharmaceutical companies. Herein, we provide an overview of anticancer cyclopeptides that were discovered in the past 20 years. The present review mainly focuses on the anticancer efficacies, mechanisms of action and chemical structures of cyclopeptides with natural origins. Additionally, studies of the structure–activity relationship, total synthetic strategies as well as bioactivities of natural cyclopeptides are also included in this article. In conclusion, due to their characteristic structural features, natural cyclopeptides have great potential to be developed as anticancer agents. Indeed, they can also serve as excellent scaffolds for the synthesis of novel derivatives for combating cancerous pathologies.
Keywords: cyclopeptides, anticancer, natural products
1. Introduction
Cancer has become an enormous burden in society and a leading cause of death in the world. Apart from aging, the risk factors of modern daily life such as smoking, unhealthy diet and mental stress also play significant roles in the growing incidence of cancer [1]. According to the World Health Organization (WHO), nearly 10 million of people were killed by cancer in 2020. New cancer cases reached to 18 million in 2018, and are expected to grow to up to 27.5 million in 2040 [2,3]. Conventional therapeutic approaches for the treatment of cancer include chemotherapy, radiotherapy and surgical resection. In recent years, advancement in cancer treatment has been applied with some novel therapies and choices for patients to improve their survival chances, such as immunotherapy, targeted therapy, hormone therapy, stem cell transplant and precision medicine [4]. However, low specificity to cancer cells, high toxicities to normal tissues as well as drug resistance are often the most problematic concerns [5,6,7]. Therefore, studies on the development of new drugs and therapies for cancer with higher efficacy are urgent and crucial.
From ancient times, taking Chinese Medicine as an example, people have been familiar with medicinal properties of natural resources and the benefits of using them to treat diseases [8]. Hence, due to their extraordinary chemical diversity with different origins, natural products are of great potential to be developed as new drugs for cancer prevention and treatment [9]. Natural cyclopeptides, due to their special chemical properties and therapeutic potency have attracted great attention from both academic researchers and pharmaceutical companies in recent years.
Cyclopeptides, also known as cyclic peptides, are polypeptides formed from amino acids arranged in a cyclic ring structure [10]. Most natural cyclopeptides are found to be assembled with 4–10 amino acids. However, some natural cyclopeptides can contain dozens of amino acids [11,12,13,14]. In the past 20 years, hundreds of ribosomally synthesized cyclopeptides have been isolated from a variety of living creatures with various biological activities. In fact, there are two biosynthetic mechanisms of cyclopeptides—ribosomal and non-ribosomal pathways. The ribosomal pathway merely requires the 21 basic proteinogenic amino acids for the synthesis of cyclopeptides, whereas the non-ribosomal pathway depends on non-ribosomal peptide synthetases (NRPSs) in incorporating functional groups, such as hydroxyl and N-methyl groups, to the existing peptides [15,16]. A large number of cyclopeptides have been isolated and demonstrated to deliver a wide spectrum of bioactivities. Very often, they can be served as antibiotics (e.g., microcin 25 from Escherichia coli) [17], protease inhibitors (e.g., SFTI-1 from Helianthus annuus) [18], boosting agents for the innate immune system (e.g., θ-defensins from Macaca mulatta) [19], anticancer agents (e.g., chlamydocin from Diheterospora chlamydosporia) [20] and so on. According to previous reports, cyclization is an important process for peptide stabilization [21]. Compared with linear peptides, cyclic peptides may have a better biological potency due to the rigidity of their conformation [22]. Moreover, the rigidity of cyclopeptides may also increase the binding affinity to their target molecules, allowing enhanced receptor selectivity as rigid structures often decrease the interaction entropy to a negative Gibbs free energy. Furthermore, cyclopeptides are resistant to exopeptidases because they contain no amino/carbonyl termini in their skeletons, so that they exert more stable binding capability than linear peptides do. Owing to the absence of the amino/carbonyl termini, the less flexible backbone of cyclopeptides even enhances their resistance to endopeptidases, and therefore, cyclopeptides are considered as useful biochemical tools with high specificity [23]. In particular, when serving as an anticancer agent, the cyclic structure of the peptide eliminates the effect of charged termini and thus increases membrane permeability [24]. As a result, cyclopeptides can penetrate to the tumors without much hurdle while exhibiting their anti-proliferative effect [25].
Overall, the structural rigidity, biochemical stability, binding affinity and membrane permeability are the predominant advantages of cyclopeptides in comparison of linear peptides and other chemicals for therapeutic purposes [10]. Owing to these exceptional characteristics, natural cyclopeptides are considered as promising lead compounds for the development of novel anticancer drugs. With the efforts from researchers around the world, a great number of cyclopeptides with anticancer effects have been identified from various living entities. In this review, we summarize the recent development of cyclopeptides with anticancer effects according to their origins.
2. Marine-Derived Anticancer Cyclopeptides
Marine organisms are the vast source of many chemical products because of the diversity of species and their abundance [26]. In fact, the marine environment though somehow mysterious, promotes the fruitful development of a great number of natural products, which may possess novel structures, chemical properties and biological effects [27]. The marine-derived cyclopeptides have received great attention due to their high potency in anti-microbial, anti-inflammatory and anticancer activities [28,29,30,31]. This section mainly discusses the discovery of marine-derived cyclopeptides with anticancer properties in the last 20 years.
2.1. Anticancer Cyclopeptides Derived from Sponge
As sessile marine filter feeders, sponges possess decent abilities to defense against foreign invaders such as viruses, bacteria and fungi [32]. Marine sponges belong to the phylum Porifera, which is the source of many bioactive marine natural products [33]. It is estimated that about 5000 different new compounds have been discovered from sponges each year [32]. Among them, cyclopeptides are an important portion of the sponge-derived compounds that are accompanied with broad biological effects including anticancer activities [34]. The structures of these sponge-derived natural cyclopeptides compounds are shown in Figure 1 and their in vitro activities against different cancer cells are listed in Table 1.
Figure 1.



Structures of cyclopeptides 1–52 derived from sponges.
Table 1.
Marine-derived anticancer cyclopeptides.
| Name | Biological Source | Anticancer Activity | Reference |
|---|---|---|---|
| Anticancer Cyclopeptides Derived from Sponge | |||
| Azumamide A (1) | Mycale izuensis | HDAC inhibitory activity against K562 cells (IC50 = 0.045 μM); Cytostatic effects on WiDr (IC50 = 5.8 μM) and K562 (IC50 = 4.5 μM) cells | [35,36,37,38,39] |
| Azumamide E (2) | Mycale izuensis | HDAC inhibitory activity against K562 cells (IC50 = 0.045 μM) | [35,36,37,38,39] |
| Microsclerodermin A (3) | Microscleroderma herdmani | Induction of apoptosis in AsPC-1 (IC50 = 2.3 µM), BxPC-3 (IC50 = 0.8 µM) and PANC-1 (IC50 = 4.0 µM) cells. | [40,41,42] |
| Leucamide A (4) | Leucetta microraphis | Inhibitory activities to HM02 (GI50 = 8.5 µM), HepG2 (GI50 = 9.7 µM) and Huh7 (GI50 = 8.3 µM) cells. | [43,44] |
| Carteritin A (5) | Sponge, Stylissa carteri | Cytotoxicity against HCT116 (IC50 = 1.3 µM), RAW264 (IC50 = 1.5 µM) and HeLa (IC50 = 0.7 µM) cells | [45] |
| Stylissatin B (6) | Stylissa massa | Inhibitory effects on HCT116, HepG2, BGC823, NCI-H1650, A2780 and MCF7 cells. (IC50 = 2.3 to 10.6 µM) | [46] |
| Cupolamide A (7) | Theonella cupola | Cytotoxicity against P388 (IC50 = 7.5 µM) cell. | [47] |
| Geodiamolide H (8) | Geodia corticostylifera | Anti-proliferative activitiy against T47D (EC50 = 38.36 nM) and MCF7 (EC50 = 89.96 nM) cells | [48,49,50] |
| Theopapuamide (9) | Theonella swinhoei | Cytotoxicity against CEM-TART (EC50 = 0.5 µM) and HCT116 (EC50 = 0.9 µM) cells. | [51] |
| Homophymines A-E (10–14) & A1-E1 (15–19) | Homophymia sp. | Anti-proliferative against several cancer cell lines (IC50 = 2 to 100 nM), among which PC3 and OV3 are the most sensitive. | [52,53] |
| Pipestelide A (20) | Pipestela candelabra | Cytotoxicity against KB (IC50 = 0.1 µM) cell. | [54] |
| Neamphamides B-D (21–23) | Neamphius huxleyi | Cytotoxicity against A549, LNCaP and PC3 cells. (IC50 = 91 to 230 nM) | [55,56] |
| Callipeltins N (24) and O (25) | Asteropus sp. | Cytotoxicity against A2058, HT29, MCF7 and MRC-5 cells. (IC50 = 0.16 to 0.21 µM and 0.48 to 2.08 µM, respectively) | [57] |
| Pipecolidepsins A (26) and B (27) | Homophymia lamellosa | Cytotoxicity against A549 (GI50 = 0.6 and 0.04 µM, respectively), HT29 (GI50 = 1.12 and 0.01 µM, respectively) and MDA-MB-231 (GI50 = 0.7 and 0.02 µM, respectively) cells. | [58,59,60,61] |
| Halipeptin D (28) | Leiosella cf. arenifibrosa | Cytotoxicity against HCT116 cell line (IC50 = 7 nM) and BMS ODCP with an average IC50 value of 420 nM. | [62,63] |
| Reniochalistatin E (29) | Reniochalina stalagmitis | Cytotoxicity against RPMI-8226 (IC50 = 4.9 μM) and MGC-803 (IC50 = 9.7 μM) cells. | [64,65] |
| Callyaerin G (30) | Callyspngia aerizusa | Cytotoxicity against L5178Y (ED50 = 4.1 mM) and HeLa (ED50 = 41.8 mM) cells. | [66] |
| Callyaerins E (31) and H (32) | Callyspngia aerizusa | Cytotoxicity against L5178Y (ED50 = 0.39 and 0.48 μM, respectively). | [67] |
| Scleritodermin A (33) | Scleritoderma nodosum | Cytotoxicity against HCT116 (IC50 = 1.9 μM), HCT116/VM46 (IC50 = 5.6 μM), A2780 (IC50 = 0.94 μM) and SKBR3 (IC50 = 0.67 μM) cells. | [68,69,70] |
| Calyxamides A (34) and B (35) | Discodermia calyx | Cytotoxicity against P388 (IC50 = 3.9 and 0.9 μM, respectively) cell. | [71] |
| Keramamide E (36) | Theonella sp. | Cytotoxicity against L1210 (IC50 = 1.42 µM) and KB (IC50 = 1.38 µM) cells. | [72] |
| Keramamides K (37) and L (38) | Theonella sp. | Cytotoxicity against L1210 (IC50 = 0.77 and 0.5 µM, respectively) and KB (IC50 = 0.45 and 0.97 µM, respectively) cells. | [73] |
| Keramamides M (39) and N (40) | Theonella sp. | Cytotoxicity against L1210 (IC50 = 2.02 and 2.33 µM, respectively) and KB (IC50 = 5.05 and 6.23 µM, respectively) cells. | [74,75] |
| Axinellins A (41) and B (42) | Axinella carteri | Cytotoxicity against NSLC-N6 (IC50 = 16.7 and 29.8 µM, respectively) cell. | [76,77] |
| Stylopeptide 2 (43) | Stylotella sp. | Inhibition of 23% BT-549 cell growth and 44% Hs578T cell growth at one dose (10−5 M). | [78] |
| Stylissamide X (44) | Stylissa sp. | Anti-migration effects on HeLa cell. | [79,80] |
| Aciculitins A-C (45–47) | Aciculites orientalis | Cytotoxicity against HCT116 (IC50 = 0.37 µM) cell. | [81] |
| Nazumazoles A-C (48–50) | Theonella swinhoei | Cytotoxicity against P388 (IC50 = 0.83 µM) cell. | [82] |
| Theonellamide G (51) | Theonella swinhoei | Cytotoxicity against HCT116 (IC50 = 6.0 µM) cell. | [83] |
| Koshikamide B (52) | Theonella sp. | Cytotoxicity against HCT116 (IC50 = 3.62 µM) and P388 (IC50 = 0.22 µM) cells. | [84] |
| Anticancer Cyclopeptides Derived from Ascidians/Tunicates | |||
| Mollamide (53) | Ascidian, Didemnum molle | Cytotoxicity against P388 (IC50 = 1.24 µM), A549 (IC50 = 3.1 µM), HT29 (IC50 = 3.1 µM) and CV1 (IC50 = 3.1 µM) cells. | [87,88] |
| Mollamide B (54) | Tunicate, Didemnum molle | Growth inhibition of H460, MCF7 and SF-268 cells. | [89] |
| Trunkamide A (55) | Ascidian, Lissoclinum sp. | Cytotoxicity against A549, P388, HT29 and MEL28 cells. (IC50 = 0.6 to 1.19 µM) | [90,91] |
| Prepatellamide A (56) | Ascidian, Lissoclinum patella | Cytotoxicity against P388 cells. (IC50~6.57 µM) | [92] |
| Vitilevuamide (57) | Ascidian, Didemnum cuculiferum | Cytotoxicity against HCT116 (IC50 = 6.24 nM), A549 (IC50 = 0.12 µM), SKMEL-5 (IC50 = 0.31 µM) and A498 (IC50 = 3.12 µM) cells. | [93,94] |
| Tamandarin A (58) | Ascidian, Didemnidae sp. | Cytotoxicity against BX-PC3 (IC50 = 1.69 nM), DU145 (IC50 = 1.29 nM), UM-SCC-10B (IC50 = 0.94 nM) cells. | [95,96,97] |
| Comoramides A-B (59–60) & Mayotamides A-B (61–62) | Ascidian, Didemnum molle | Cytotoxicity against A549, HT29 and MEL28 cells. (IC50 = 7.22 to 14.97 µM) | [98] |
| Patellin 6 (63) | Ascidian, Lissoclinum patella | Cytotoxicity against P388, A549, HT29 and CV1 cells. (IC50~2.08 µM) | [99] |
| Cycloforskamide (64) | Sea slug, Pleurobranchus forskalii | Cytotoxicity against P388 cell (IC50 = 5.8 µM). | [100] |
| Anticancer Cyclopeptides Derived from Mollusks | |||
| Keenamide A (65) | Mollusk, Pleurobranchus forskali | Anti-proliferative activity against A549, P388, MEL20 and HT29 cells. (IC50 = 4.03 to 8.05 µM) | [103] |
| Aurilide (66) | Sea hare, Dolabella auricularia | Cytotoxicity against HeLa S3 (IC50 = 0.013 µM) cell. Inhibitory activity against ovarian, renal and prostate cancer cells in NCI 60 cell lines. | [104,105] |
| Dolastatin 16 (67) | Sea hare, Dolabella auricularia | Cytotoxicity against H460 (GI50 = 1.09 nM), KM20L2 (GI50 = 1.37 nM), SF-295 (GI50 = 5.92 nM) and SKMEL-5 (GI50 = 3.75 nM) cells. Anti-proliferative effects against 5 human leukemia cell lines | [106] |
| Doliculide (68) | Sea hare, Dolabella auricularia | Cytotoxicity against HeLa S3 (IC50 = 1.62 nM) cell. | [107,108,109,110] |
| Kulokekahilide-1 (69) | Mollusk, Philinopsis speciosa | Cytotoxicity against P388 (IC50 = 2.2 µM) cell. | [111] |
| Kulokekahilide-2 (70) | Mollusk, Philinopsis speciosa | Cytotoxicity against P388 (IC50 = 4.2 nM), SK-OV-3 (IC50 = 7.5 nM), MDA-MB-435 (IC50 = 14.6 nM) and A-10 (IC50 = 59.1 nM) cells. | [112,113,114] |
| Kahalalide R1 (71) | Sea slug, Elysia grandifolia | Cytotoxicity against MCF7 (IC50 = 0.14 µM), L1578Y (IC50 = 4.26 nM) cells. | [117] |
| Anticancer Cyclopeptides Derived from Marine Algae | |||
| Galaxamide (72) | Algae, Galaxaura filamentosa | Inhibitory activity against GRC-1 (IC50 = 7.18 μM) and HepG2 (IC50 = 7.81 μM) cells. | [119,120] |
Azumamides A (1) and E (2) were two cyclotetrapeptides isolated from the marine sponge Mycale izuensis. These two compounds have been demonstrated to exhibit potent inhibitory activities against histone deacetylase (HDAC) enzymes [35]. It is widely acknowledged that overexpression of HDACs may lead to the occurrence of heterochromatin and thus different types of cancers [36]. In consequence, HDAC inhibitors are suggested with a role in cancer growth inhibition, and their anticancer mechanisms are mainly associated with their induction of apoptotic events in cancer cells [37]. According to a previous study, azumamide A showed anti-proliferative effect on K562 (human bone marrow chronic myelogenous leukemia) cells with IC50 values of 0.045 μM, and cytostatic effect on WiDr (human colon cancer) and K562 cells with IC50 values of 5.8 and 4.5 μM, respectively [35]. Furthermore, azumamides A and E obtained from total synthesis were demonstrated to exert significant zinc-binding affinity to HDACs, which may contribute to the rational design for synthesizing potent anticancer agents [38,39].
Microsclerodermin A (3), a cyclic hexapeptide, first isolated from the lithistid sponge Microscleroderma herdmani, was found to be a potent antifungal agent [40,41]. This compound was also reported to have anti-proliferative activities against lung, leukemia and pancreatic cancer cells. In human pancreatic adenocarcinoma cells (AsPC-1), the level of phosphorylated NFκB was significantly reduced by the incubation with microsclerodermin A (IC50 = 2.3 µM) whereas apoptosis was strongly induced. In the human pancreatic adenocarcinoma cells BxPC-3 (IC50 = 0.8 µM) and human pancreatic epithelioid carcinoma PANC-1 cells (IC50 = 4.0 µM), microsclerodermin A exhibited remarkable inhibitions at low IC50 values. In human umbilical vein endothelial cells (HUVECs), this cyclopeptide was shown to have no effect of angiogenesis [42]. The above biological investigations demonstrated that microsclerodermin A is a good anticancer lead compound for further development.
Leucamide A (4) is a cytotoxic cyclic heptapeptide with potent cytotoxicity, which was first purified from the Australian marine sponge Leucetta microraphis. In the study of Kehraus and co-workers, this peptide was found to exhibit inhibitory activities against human mucin-producing gastric cells (HM02), and human liver cancer cells (HepG2 and Huh7) with GI50 values of 8.5, 9.7 and 8.3 µM, respectively [43]. In the consecutive year, the total synthesis of leucamide A was accomplished, and provided a viable approach to the sufficient supply of this compound for further comprehensive investigations [44].
Afifi et al. isolated a new cyclic heptapeptide, named carteritin A (5), from the marine sponge Stylissa carteri. Carteritin A exhibited cytotoxicities against human colorectal (HCT116), mouse macrophage (RAW264) and human cervical (HeLa) cancer cells with IC50 values of 1.3, 1.5 and 0.7 µM, respectively [45].
Stylissatin B (6), a novel cytotoxic cycloheptapeptide, was obtained from the sponge Stylissa massa. This compound displayed inhibitory effects on HCT116, HepG2, gastric (BGC823), lung (NCI-H1650), ovarian (A2780) and breast (MCF7) human cancer cells with IC50 values ranging 2.3 to 10.6 µM [46]. These results implicated that significant structural modification is needed to improve the moderate activity of Stylissatin B.
Derived from the sponge Theonella cupola, cupolamide A (7) was found to be a biologically active cyclic heptapeptide. This compound was cytotoxic against mouse lymphoma cells (P388) with an IC50 value of 7.5 µM but no cytotoxic against thrombin. Moreover, the resemblance of cupolamide A to γ-aminobutyric acid (GABA) unveils its potential effect on the central nervous system, which requires further investigation [47].
Geodiamolide H (8), a cyclic depsipeptide isolated from the Brazilian sponge Geodia corticostylifera, was firstly identified as a neurotoxic and hemolytic agent [48]. This compound showed a potent anti-proliferative activity against the human ductal carcinoma T47D (EC50 = 38.36 nM) and MCF7 (EC50 = 89.96 nM) cells via altering the cytoskeleton of cancer cells by actin depolymerization. However, no such deleterious effect was observed in normal cell lines even when higher concentrations was used (e.g., 136 nM) [49]. Subsequently, the results of another study further demonstrated that geodiamolide H exhibited significant inhibitory effects on the migration and invasion of breast cancer cells (Hs578T) that were poorly differentiated and highly aggressive [50]. These studies revealed geodiamolide H as a potential lead compound for further development of therapeutic use.
Theopapuamide (9), isolated from the lithistid sponge Theonella swinhoei, was also a new cytotoxic depsipeptide with a cyclic ring structure. This cyclopeptide exhibited significant cytotoxicity against human leukemia (CEM-TART) (EC50 = 0.5 µM) and HCT116 (EC50 = 0.9 µM) cells. When compared with other depsipeptide with HIV-inhibitory activities, the β-methoxytyrosine residue of this compound appears to be of great significance for its anti-HIV biological effect [51].
A group of structurally similar cyclodepsipeptides named homophymines A-E (10–14) and A1-E1 (15–19) isolated from the sponge Homophymia sp., were found to be potent anti-proliferative agents against a wide panel of cancer cell lines with IC50 values in the range of 2–100 nM. Among these cell lines, human prostate (PC3) and ovarian (OV3) human cancer cell lines were the most sensitive to this group of cyclopeptides. Furthermore, the overexpression of P-glycoprotein (P-gp) proteins did not affect the intracellular concentrations of homophymines [52]. Apart from cytotoxic activities, homophymine A also exhibited cytoprotective activity against HIV-1 infection (IC50 = 75 nM) [53]. In the family of homophymins, their general chemical structures are complicated but structural differences among them are subtle. Because of their significant anticancer potential, further structure activity relationship (SAR) assessment, bioactivity evaluation and mechanism of action studies should be performed.
Jaspamide is an early-discovered cyclic depsipeptide from the sponge Jaspis splendens, which delivered pronounced inhibitory activities against breast and prostate cancer cells [31]. Pipestelide A (20), a cyclodepsipeptide derived from the marine sponge Pipestela candelabra, was biosynthetically related to jaspamide. When subjected to cytotoxicity assay, this compound showed cytotoxicity against the human nasopharyngeal epidermoid carcinoma (KB) cells with an IC50 value of 0.1 µM, which was less potent than jaspamide [54].
Three cytotoxic cyclodepsipeptides, neamphamides B-D (21–23), were purified from the Australian sponge Neamphius huxleyi. These three compounds were cytotoxic to human lung (A549), cervical (HeLa) and prostate (LNCaP and PC3) cancer cells with IC50 values varying from 91 to 230 nM. Nevertheless, at sub-nanomolar concentrations (100 nM for 48 h and 72 h), neaphamide D was observed to induce proliferation in human lung cancer A549 cells. Therefore, these neamphamindes should be discreetly utilized [55]. Apart from the cytotoxic effects, neaphamide D also showed a great antimicrobial potency against the growth of Mycobacterium smegmatis and M. bovis BCG [56].
Collected in Solomon Islands, the sponge Asteropus sp. yielded two new derivatives of callipeltin A, which were named cyclic depsipeptides callipeltins N (24) and O (25). Their cytotoxicities against human melanoma (A2058), colorectal (HT29), and breast (MCF7) cancer cells as well as non-malignant MRC-5 fibroblast cells were accompanied with IC50 values ranging from 0.16 to 0.21 µM for callipeltins N and ranging from 0.48 to 2.08 µM for callipeltins O. The slight difference of cytotoxicities between malignant cancer cells and non-malignant cells indicated that these two compounds can provide minimal specificity.Structure-wise, the biological potencies of the two compounds was highly related to the positions of the methylGln group [57].
Pipecolidepsins A (26) and B (27), two head-to-side-chain cyclodepsipeptides with significant anticancer potential, were isolated from the sponge Homophymia lamellosa. Both compounds were tested for their cytotoxic effects against A549, HT29 and human breast cancer (MDA-MB-231) cells. The results demonstrated that pipecolidepsin B (GI50 = 0.04, 0.01 and 0.02 µM, respectively) was more cytotoxic than pipecolidepsin A (GI50 = 0.6, 1.12 and 0.7 µM, respectively), though pipecolidepsin B contains only one more hydroxy group. The SAR assessment further revealed that the replacement of residue at C-3 was an important step for gaining higher hydrophilic properties in pipecolidepsin B [58], whereas the γ-amino acid in pipecolidepsin A was the major functional group for its cytotoxicity [59], While exerting cytotoxic activity, pipecolidepsin A was shown to induce intense membrane damage and necrotic cell death [60]. Due to the early discovery of pipecolidepsin A, a total solid-phase synthesis of this compound had been previously accomplished [61]. Considering the great anticancer potential of pipecolidepsin B, the further understanding of its biological property, total synthetic strategy as well as other pharmacological actions is of high research interest.
Halipeptin D (28) was a cyclic depsipeptide isolated from the sponge Leiosella cf. arenifibrosa by Faulkner and Manam [62]. This natural product was reported to exhibit significant cytotoxicity against HCT116 cells (IC50 = 7 nM) and BMS ODCP (oncology diverse cell panel) with an average IC50 value of 420 nM. However, the synthetic halipeptin D and other halipeptins (e.g., halipeptins A-C) did not show potent cytotoxicities. Based on the distinct results, it is reasonable to assume that the previous naturally derived compound might have been contaminated with other cytotoxic agents during harvest [63]. The discrepancy of cytotoxicity between the synthetic and naturally derived compounds is not surprising as such happens somehow. As a result, biological properties of halipeptin D remained uncertain and required further investigations.
The cyclic peptide reniochalistatin E (29) was isolated from the marine sponge Reniochalina stalagmitis by Zhan and co-workers. Differed from other congeners (reniochalistatins A-D) of the same origin, this compound showed in vitro cytotoxicity against RPMI-8226 (human myeloma) and MGC-803 (human gastric carcinoma) cells with IC50 values of 4.9 and 9.7 μM, respectively [64]. The subsequent total synthesis of reniochalistatin E provided some inspiring methods for the synthetic strategies of cyclopeptides with similar structures [65].
From the freeze-dried Indonesian sponge Callyspngia aerizusa, a cyclic peptide called callyaerin G (30) was purified. Biological studies showed that this compound was cytotoxic to mouse lymphoma cells (L5178Y) and HeLa cells with ED50 values of 0.41 and 4.17 μM, respectively [66]. Callyaerins E (31) and H (32), were purified using another bioassay-guided fractionation of the same sponge Callyspngia aerizusa. Callyaerins E and H exhibited strong anti-proliferative activities against L5178Y cells with ED50 values of 0.39 and 0.48 µM, respectively. Similar to the three compounds above, other proline-rich callyaerins yielded from the same sponge also showed high antimicrobial activities [67]. Accordingly, the biological significance of proline residue in cyclic peptides becomes a trendy research topic. The mechanism of callyaerins in biological actions definitely deserves further investigation.
Obtained from the lithistid sponge Scleritoderma nodosum, scleritodermin A (33) was a cyclic peptide with anticancer potential. By means of cell viability assay, this compound was demonstrated to exert cytotoxicity in various cancer cell lines including HCT116, HCT116/VM46 (human vinblastin-resistant colon cancer cell), A2780 and SKBR3 (human breast cancer cells) with IC50 values of 1.9, 5.6, 0.94 and 0.67 µM, respectively. Regarding its molecular mechanism, scleritodermin A induced G2/M phase arrest and inhibited tubulin polymerization [68]. By using different moieties and fragments, the total synthesis of scleritodermin A had been accomplished [69,70], and consequently provided a sufficient supply of this compound for further comprehensive studies of its other biological properties and the synthesis of analogs or derivatives.
Calyxamides A (34) and B (35), two novel cyclic peptides with bioactivities, were first isolated from the marine sponge Discodermia calyx in Japan. When incubated with leukemia P388 cells, both compounds yielded moderate cytotoxicities with IC50 values of 3.9 and 0.9 µM, respectively. Subsequently, the analysis of the 16S rDNA sequence indicated that the biosynthesis of calyxamides A and B were possibly related to Candidatus Entotheonella sp. inhabiting in the Theonella genus [71]. Hence, further biological studies of these compounds would be relied on the bulky isolation from their natural source.
A group of compounds named keramamides were yielded from the Okinawa marine sponge Theonella sp., which possess a variety of bioactivities. Keramamide E (36) was reported to display inhibitory activities against mouse lymphocytic leukemia (L1210) and KB cells with IC50 values of 1.42 and 1.38 µM, respectively [72]. Subsequent explorations on another Theonella sponge led to the purification of keramamides K (37) and L (38). Keramamide K contains a (1-Me)Trp residue, which is rarely seen in any naturally derived compound whilst keramamide L is a natural cyclopeptide possessing a MeCtrp residue. These two special cyclopeptides exhibited strong cytotoxicity against L1210 (IC50 = 0.77 and 0.5 µM, respectively) and KB (IC50 = 0.45 and 0.97 µM, respectively) cells [73]. Keramamides M (39) and N (40) were two other congeners from the Theonella sponge that were discovered by the same research team. Both cyclic peptides contained a sulfate ester, which is also a rare residue to be found in compounds derived from marine sponge. Keramamides M and N showed moderate cytotoxicities against L1210 (IC50 = 2.02 and 2.33 µM, respectively) and KB (IC50 = 5.05 and 6.23 µM, respectively) cells [74]. The total synthesis of different keramamides had been completed by the same research group [75]. From the results of structural elucidation, some intriguing constituents of these cyclic peptides exhibited anticancer potentials against some other cancer cell lines. In order to highlight the aspect of their anticancer effect, more types of cancer cell lines and mechanism of their biological actions should be exploited.
The marine sponge Axinella carteri collected near Vanuatu islands yielded two proline-containing cyclopeptides, which were called axinellins A (41) and B (42). Both compounds were found to display moderate cytotoxicity against non-small lung cancer cells (NSLC-N6) with IC50 values of 16.7 and 29.8 µM, respectively [76]. The first synthesis of axinellin A was achieved shortly after its discovery. However, the synthetic compound did not show cytotoxicity as the naturally occurring peptide did [77]. Therefore, further investigation should be performed for clarifying such discrepancy on cytotoxicities between the synthetic and the natural axinellin A.
The proline-rich cyclopeptide stylopeptide 2 (43) was isolated from the marine sponge Stylotella sp. in Papua New Guinea. According to the study of Brennan et al., this compound inhibited 23% growth of BT-549 and 44% growth of Hs578T human breast cancer cells in the National Cancer Institute one-dose (10−5 M) 60-cell-line assay [78]. However, further dose-dependent cell viability assays should be carried out for a more accurate evaluation of its cytotoxicity and anticancer potential.
Stylissamide X (44) was a cyclic proline-rich octapeptide discovered in the Indonesian marine sponge of Stylissa species. Although only little inhibitory activity was observed in the viability assay of HeLa cells, stylissamide X induced significant anti-migration effects on the same cell line in the wound-healing assay [79]. As a new member of the stylissamides family, the total synthetic approach of it became extremely important for securing a good supply of materials for further comprehensive chemical and biological studies [80]. More systemic cell viability assays and detailed evaluation of its anti-migration properties would be of similar importance for the mode of action studies of this type of compounds.
Aciculitins A-C (45–47) were three bicyclic peptides extracted from the lithistid sponge Aciculites orientalis with an unusual histidino-tyrosin bridge. They were also the first glycopeptidolipids obtained from a marine origin. In regards to their biological activities, aciculitins A-C were cytotoxic against HCT116 (IC50 = 0.37 µM) and inhibitory to the growth of bacterium Candida albicans. Comparing with other compounds of the same origin, aciculitins A-C possess a histidine residue which is responsible for their predominant bioactivities [81].
Nazumazoles A-C (48–50) were three novel bicyclic pentapeptides isolated from a mixture derived from the marine sponge Theonella swinhoei. The mixture of nazumazoles A-C showed cytotoxic activities against P388 cells with an IC50 value of 0.83 µM. The reduction of either the ketone or thiol group in the nazumazoles led to a significant decrease of the cytotoxicity of the mixture [82]. Based on the results above, separation of individual nazumazoles A-C is undoubtedly needed to validate the anticancer potential of this type of compounds.
Theonellamide G (51) was a novel bicyclic glycopeptide isolated from the Red Sea Sponge Theonella swinhoei. When compared the positive anticancer control etoposide (2.0 µM), theonellamide G merely yielded a moderate cytotoxicity against HCT116 with an IC50 value of 6.0 µM. However, this compound showed a potent antifungal activity against the wild and amphotericin B-resistant strains of Candida albicans [83]. These results added some fresh insights into the diverse biological activities of this natural compound, which may inspire further studies on the pharmacological actions and SAR for this class of compounds.
Koshikamide B (52) was a 17-residue cyclic peptide lactone purified from a marine sponge of the genus Theonella. This compound was the first natural product containing the constitution of Nδ-carbamoylasparagine. Importantly, the 2-(3-amino-2-hydroxy-5-oxopyrrolidin-2-yl) propionic acid (AHPP) residue of koshikamide B was a unique moiety in peptide lactones. Koshikamide B exhibited cytotoxicity in HCT116 and P388 cancer cells with IC50 values of 3.62 and 0.22 µM, respectively [84].
2.2. Anticancer Cyclopeptides Derived from Ascidians/Tunicates
Ascidians are also known as tunicates. They are found in high-current fields, and firmly fixed to rocks. About 3000 species of ascidians have been reported so far [85]. As more and more compounds were isolated from the ascidians, the US National Cancer Institute (NCI) estimated that about 1% of the marine natural products (MNPs) showed anticancer activities [86]. This section focuses on the ascidians derived cyclopeptides with promising anticancer potential. The structures of these compounds are present in Figure 2 and their activities are listed in Table 1.
Figure 2.

Structures of cyclopeptides (53–64) derived from ascidians/tunicates.
Mollamide (53) was a cyclopeptide isolated from the ascidian Didemnum molle. This compound displayed a moderate cytotoxicity against several cancer cell lines, with IC50 values of 1.24 μM against P388 cells, and 3.1 μM against A549, HT29 and monkey kidney CV1 cells. The anti-proliferative property of mollamide was plausibly associated with its ability to inhibit RNA synthesis in the cancer cells [87,88]. Another research study on the same Indonesian tunicate Didemnum molle led to the isolation of a novel congener, which was a cyclic hexapeptide called mollamide B (54). Mollamide B (100 μM) displayed significant growth inhibition against human lung carcinoma (H460), MCF7 and glioblastoma (SF-268) cells by 29%, 44% and 42%, respectively. Other than its anticancer activities, mollamide B also exhibited notable anti-HIV (HIV-1, EC50 = 48.7 μM) and anti-malarial (Plasmodium falciparum, IC50 = 2.87 μM) properties [89]. For the future perspective of its anticancer potential, the specificity of mollamide B acting against different cancer cell lines should be investigated.
Trunkamide A (55) is an analog of mollamide with a similar cyclopeptide structure that is naturally produced by ascidians of Lissoclinum sp. Compared to mollamide, trunkamide A was reported to have a more promising antitumor activity [90]. Due to its significant in vitro cytotoxicity against several human-derived cancer cell lines including A549, P388, HT29 and MEL28 (human melanoma) with IC50 values ranging from 0.6 to 1.19 μM, trunkamide A had been evaluated for its antitumor efficacy in several preclinical trials [91].
Fu et al. purified a novel cyclic peptide called prepatellamide A (56), from Lissoclinum patella. Although the structure of this peptide had been determined, its biological properties were incompletely understood. The IC50 value of prepatallamide A against P388 cells was 6.57 µM [92]. Other than this piece of result, no further information on prepatallamide A could be obtained. Therefore, the bioactivity studies of this compound are desperately needed.
Vitilevuamide (57), another ascidian derivative with a bicyclic structure, was found in two species of ascidians, Didemnum cuculiferum and Polysyncranton lithostrotum. It displayed potent cytotoxicity against HCT116 cells, with an IC50 value of 6.24 nM. In A549, SKMEL-5 (human melanoma) and A498 (human kidney carcinoma) cells, vitilevuamide exhibited a less potent anti-proliferative effect as its IC50 values were determined to be 0.12, 0.31 and 3.12 μM, respectively. In addition, the compound showed a percentage increase in the mean lifespan (%ILS) of 70 at the concentration of 30 μg/mL in a P388-xenograft in vivo experiment in mice, which indicated its significant anticancer potential [93]. Regarding its anticancer mechanisms, vitilevuamide inhibited tubulin polymerization and induced cell cycle arrest at the G2/M phase [94]. As the supply of the naturally derived vitilevuamide is limited, further evaluations on this compound are largely hindered. Due to its complex structure, identification of its interactive site(s) with tubulin and methods of chemical synthesis are in slow progress.
Tamandarin A (58) was first isolated from a Brazilian marine ascidian of the family Didemnidae. This compound was cytotoxic to three cancer cell lines, BX-PC3 (pancreatic carcinoma), DU145 (prostate carcinoma) and UM-SCC-10B (laryngeal squamous cell carcinoma) with IC50 values of 1.69, 1.29 and 0.94 nM, respectively. Didemnin B, another renowned tunicate-derived cyclodepsipeptide, had been promoted into clinical trials several years ago owing to its significant anticancer potential [95]. With slightly lower IC50 values than didemnin B, tamandarin A can also be a good anticancer candidate [96,97]. Although its mechanism of action remains unclear, the structural similarity between tamandarin A and didemnin B may represent their similar mechanisms of action in cancer cells, which warrants it for further biological investigations.
Rudi’s research group found two groups of cyclic hexapeptides, comoramides A and B (59–60) and mayotamides A and B (61–62), which were isolated from the ascidian Didemnum molle collected at different spots. Cytotoxicity screening revealed that these two groups of compounds showed cytotoxic activities against A549, HT29 and MEL28 cell lines with IC50 values ranging from 7.22 to 14.97 µM [98].
Patellin 6 (63) was a cytotoxic cyclic hexapeptide isolated from the colonial ascidian Lissoclinum patella. Although patellins 1-5 exhibited no cytotoxicity, patellin 6 was highly cytotoxic to P388, A549, HT29 and CV1 cells with the IC50 values around 2.08 µM. In addition, patellin 6 inhibited the activity of topoisomerase II with an IC50 value of 2.6 µM [99].
Cycloforskamide (64) was a macrocyclic dodecapeptide isolated from the sea slug Pleurobranchus forskalii collected in Japan. This compound showed cytotoxic effect against P388 cells with an IC50 value of 5.8 µM. In addition, from an ecological perspective, cycloforskamide is presumed to chelate toxic metals and plays a detoxification role [100].
2.3. Anticancer Cyclopeptides Derived from Mollusks
As the largest marine phylum and the secondary largest phylum of invertebral animals, mollusks possess highly diverse anatomical structures while having various behavior and habitats [101]. Surprisingly, a great amount of enzymes, polysaccharides, lipids and peptides with broad therapeutic uses have been discovered from mollusks. No wonder mollusks are considered a great resource for the discovery of bioactive compounds [102]. Many previous studies reported that mollusk-derived compounds showed significant anticancer properties, and many of them are indeed cyclopeptides. This section lists a wide range of mollusk-derived cyclopeptides with decent anticancer activities. The structures of these compounds are shown in Figure 3 and their activities are listed in Table 1.
Figure 3.

Structures of cyclopeptides (65–71) derived from ascidians/tunicates.
Keenamide A (65) is a cyclic hexapeptide isolated from Pleurobranchus forskali. This compound elicited anti-proliferative activities against tumor cell lines A549, P388, MEL20 (uveal melanoma) and HT29 with IC50 values ranging from 4.03 to 8.05 μM; however, its molecular mechanisms remained elusive [103].
Aurilide (66) is a 26-membered cyclic peptide extracted from the internal organs of sea hare Dolabella auricularia. It exhibited significant cytotoxicity against HeLa S3 tumor cells with an IC50 value of 0.013 µM. In the NCI-60 assay, this compound showed a strong inhibitory activity against the ovarian, renal as well as prostate cancer cells. However, no significant antitumor effect was observed in the subsequent in vivo experiments in xenograft mice. In regards to its antitumor mechanism, aurilide promoted microtubule stabilization rather than inducing a direct tubulin interaction [104]. Sato et al. found that aurilide could selectively bind to prohibitin-1 (PHB1) in mitochondria and activate proteolytic processing of OPA1, resulting in mitochondrial apoptosis [105]. However, further studies are needed to clarify the obvious diverging results obtained from the in vitro and in vivo tests of aurilide. In addition, detailed studies of mechanism and SAR are also needed to develop this type of cyclopeptides as druggable anticancer agents.
Dolastatin 16 (67) is a cyclodepsipeptide originated from the sea hare Dolabella auricularia in Papua New Guinea. When tested against NCI cancer cell lines, this compound exhibited strong inhibitory activities against H460 (GI50 = 1.09 nM), colon adenocarcinoma KM20L2 (GI50 = 1.37 nM), glioblastoma SF-295 (GI50 = 5.92 nM) and SKMEL-5 (GI50 = 3.75 nM) cells. Moreover, dolastatin 16 also showed comparable anti-proliferative effects against five human leukemia cell lines [106]. Although studies on the molecular mechanisms or pharmacological actions of dolastatin 16 are rather limited, this highly potent bioactive compound is worthy for further development into an anticancer agent.
Doliculide (68), a potent cytotoxic cyclodepsipeptide, was previously isolated from the Japanese sea hare Dolabella auricularia. This compound showed very potent cytotoxicity against HeLaS3 cells with an IC50 value of 1.62 nM [107]. It is worth noting that the first total synthesis of doliculide was accomplished soon after its isolation [108]. Actin is an abundant protein in many eukaryotic cells, and it is also an important target in many cancer studies. In order to target actin polymerization, Matcha et al. developed an efficient strategy for synthesizing (-)-doliculide, which was proven with great actin binding ability [109]. When applied to breast cancer cell lines MCF7 and MDA-MB-231, the synthetic doliculide showed a decent anti-proliferative activity with IC50 values of 93 and 77 nM, respectively. Induction of apoptotic events and migratory impairment were also observed in both cell lines when incubated with doliculide [110]. Collectively, the above experimental results unveiled the high anticancer potential of doliculide.
Kulokekahilide-1 (69) was a cyclic depsipeptide purified from the cephalaspidean mollusk Philinopsis speciosa. This compound showed cytotoxic activity against P388 cells with an IC50 value of 2.2 µM [111]. Kulokekahilide-2 (70), a much more potent cytotoxic cyclic depsipeptide than 69 was isolated from the same origin two years later. The IC50 values of kulokekahilide-2 in P388, human ovarian cancer (SK-OV-3), human breast cancer (MDA-MB-435) and rat myoblast (A-10) cells were determined to be 4.2, 7.5, 14.6 and 59.1 nM, respectively [112]. Due to the potent anticancer potential, kulokekahilide-2 and its derivatives were totally synthesized in subsequent studies. According to the SAR assessments and cytotoxicity tests performed by Umehara et al., the IC50 values of the synthesized kulokekahilide-2 against A549, K562 and MCF7 cells were found to be 0.0021 nM, 0.0031 nM and 0.22 nM. These low IC50 values indicated that this compound was of great anticancer potential. Interestingly, the cyclic structure of this peptide accompanied by the chirality at the 21 position largely contributes to its super potent cytotoxicity [113]. According to the NCI COMPARE analysis the mechanisms of action of kulokekahilide-2 appeared rather different from those of the conventional anticancer agents such as aurilide (66), palauamide (182) and lagunamide A (152) [114]. Nevertheless, kulokekahilide-2 and its derivatives are considered good anticancer candidates for testing in vivo efficacies in future human clinical trials. In the long run, further studies on specific mechanism of action, exploration of new derivatives as well as structural modifications would be of high significance for the development of kulokekahilide-2 as therapeutic agent for cancer treatment.
Kahalalide F is an intensively studied cyclic peptide that was first isolated from Elysia rufescens in 1993 [115]. Owing to its remarkable anticancer potential, several phase II clinical trials for kahalalide F are ongoing whilst a few had been completed [116]. As an analog of kahalalide F, kahalalide R1 (71) is also a cyclic depsipeptide, which was reported to be isolated from a species of sea slug E. grandifolia in 2006. Both kahalalide R1 and kahalalide F were tested for their cytotoxicities against MCF7 cells, and the results showed that they had comparable IC50 values of 0.14 and 0.22 µM, respectively. Furthermore, kahalalide R1 also exhibited anti-proliferative activity in mouse lymphoma L1578Y cell line with an IC50 value of 4.26 nM. The anti-proliferative potency of kahalalide R1 against cancer cells was almost the same potent as kahalalide F [117]. Nevertheless, further in vivo and preclinical studies on kahalalide R1 are urgently needed for a thorough validation of its anticancer potential.
2.4. Anticancer Cyclopeptides Derived from Marine Algae
In the plant kingdom, marine algae are the most ancient members responsible for maintaining the stability of the marine ecosystem though they are merely simple organisms possessing chlorophyll. Many bioactive compounds have also been derived from marine algae, therefore these microorganisms have attracted a lot of attention in the recent years for the exploration of algal products [118]. The structures of renowned bioactive algal compounds are provided in Figure 4 and the activities of these compounds are listed in Table 1.
Figure 4.
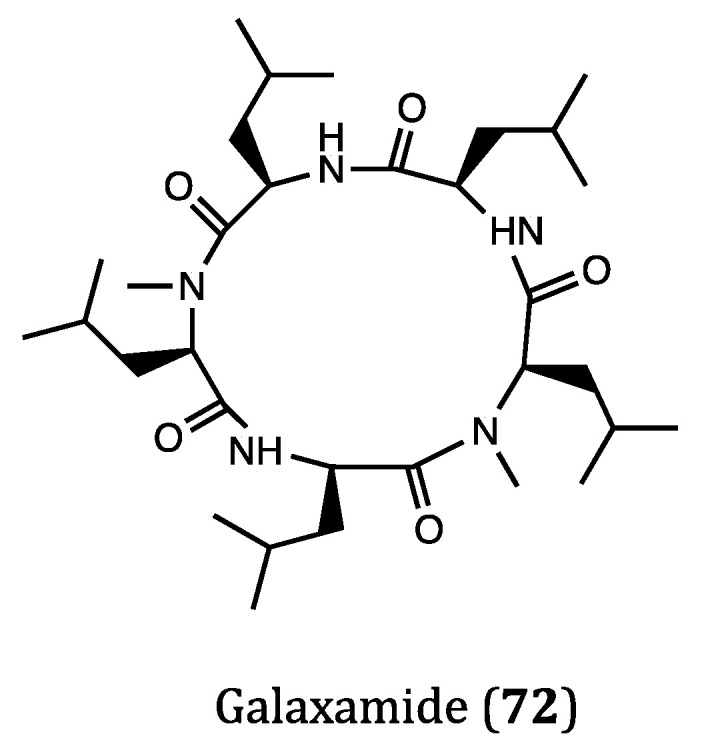
Structure of cyclopeptide (72) derived from marine algae.
Galaxamide (72) was a representative algae-derived cyclic pentapeptide discovered in marine algae Galaxaura filamentosa, which showed moderate inhibitory activities against GRC-1 (human renal cell carcinoma, IC50 = 7.18 μM) and HepG2 (IC50 = 7.81 μM) cell lines [119]. In the study of Lunagariya et al. [120], they investigated the mechanism of action of galaxamide on MCF7 cells in detail. Their results demonstrated that galaxamide induced apoptosis via the oxidative stress-mediated signaling pathway, by which the mitochondrial membrane potential was disrupted in response to the overwhelmed production of reactive oxygen species (ROS). Further, cell cycle arrest at G1 phase was also observed after the treatment with galaxamide due to ROS production, which contributed much to the apoptosis of MCF7 cells. Several synthetic analogs of galaxamide were found to be more potent than the natural scaffold, thus this type of compounds is worthy for further studies as novel candidates for the treatment of breast cancer.
3. Terrestrial Plant-Derived Anticancer Cyclopeptides
Plants are undeniably an important source for natural products. Nowadays, traditional plant-based medicines are still prevalently used for healthcare and medical treatment around the world. Over 28,000 species of plants have been recorded with differenct therapeutic purposes [121]. Therefore, plants constituents have long been regarded as the mainstream materials for drug discovery. In particular, over one third of current anticancer drugs are derived from plant natural products [122]. In this section, the recently isolated plant-derived cyclopeptides and their anticancer properties are discussed. The structures of these compounds are shown in Figure 5 and their activities are listed in Table 2.
Figure 5.

Structures of cyclopeptides (73–117) derived from terrestrial plant.
Table 2.
Terrestrial plant-derived anticancer cyclopeptides.
| Name | Biological Source | Anticancer Activity | Reference |
|---|---|---|---|
| Cherimolacyclopeptides C-F (73–76) | Seeds of Annona cherimola | Cytotoxicity against KB cell. (IC50 = 0.017 to 0.97 µM) | [123,124,125] |
| Integerrimide C (77) | Latex of Jatropha integerrima | Cytotoxicity against KB (IC50 = 1.7 µM) cell. | [126] |
| LOB3 (78) | Flaxseed oil | Cytotoxicty against A375, SKBR3 and MCF7 cells. | [128,129] |
| Cambodine A (79) | Root bark of Ziziphus cambodiana | Cytotoxicity against BC-1 (IC50 = 11.1 µM) cell. | [130] |
| Longicalycinin A (80) | Dianthus superbus | Growth inhibitory activity against HepG2 (IC50 = 22.13 μM) cell. | [131,132,133] |
| Dianthin E (81) | Dianthus superbus | Cytotoxicity against HepG2 (IC50 = 3.51 μM) cell. | [134] |
| [1-8-NαC]-Zanriorb A1 (82) | Leaves of Zanthoxylum riedelianum | Cytotoxicity against Jurkat leukemia T cell (IC50 = 218 nM). | [136] |
| Justicianenes B-D (83–85) | Justicia procumbens L. | Justicianene D displayed cytotoxicity against MCF-7 cells (IC50 = 90 μM). | [137] |
| GCP-1 (86) | Ginseng | Cytotoxicity against SGC-7901 cells (IC50 = 37.8 μM). | [138] |
| Rubipodanin A (87) | Roots and rhizomes of Rubia podantha | Cytotoxicity against HeLa, A549 and SGC-7901 cells. (IC50 = 3.80 to 7.22 μM) | [143] |
| Rubipodanin B (88) | Rubia podantha | Cytotoxicity against MDA-MB-231 (IC50 = 1.47 μM), SW620 (IC50 = 0.69 μM) and HepG2 (IC50 = 3.37 μM) cells. | [144] |
| RA-XII (89) | Rubia yunnanensis | Inhibitory effects on 4T1 ((IC50 = 606 nM and 96 nM for 24 and 48 h, respectively), SW260 and HT29 cells. | [145,146,147] |
| RA-XXV (90) | Roots of Rubia cordifolia L. | Cytotoxicity against HL-60 ((IC50 = 62 nM) and HCT116 cells ((IC50 = 28 nM). | [149] |
| RA-XXVI (91) | Roots of Rubia cordifolia L. | Cytotoxicity against HL-60 ((IC50 = 66 nM) and HCT116 cells ((IC50 = 51 nM). | [149] |
| HB7 (92) | Hedyotis biflora | Cytotoxicity against BxPC3 (IC50 = 0.68 μM), Capan2 (IC50 = 0.45 μM), MOH-1 (IC50 = 0.33 μM) and PANC1 (IC50 = 0.36 μM) cells. | [150] |
| Cliotides T1-T4 (93–96), cliotide T7 (97), cliotide T10 (98), cliotide T12 (99) | Clitoria ternatea | Cliotides T1-T4: cytotoxicity against HeLa (IC50 = 0.6 to 8.0 μM) cell. Cliotides T2, T4, T7, T10 and T12: cytotoxicity against A549 (IC50 = 0.21 to 7.59 μM) and A549/paclitaxel (IC50 = 0.45 to 7.92 μM) cells. |
[151,152] |
| psyle E (100) | Psychotria leptothyrsa var. longicarpa | Cytotoxicity against U937-GTB IC50 = 0.76 μM) cell. | [153] |
| Vigno 5 (101) | Viola ignobilis | Pro-apoptotic activity on HeLa cell. | [154,155] |
| Cycloviolacin O2 (CyO2) (102) | Viola odorata L. | Cytotoxicity against U937 GTB (IC50 = 0.75 μM), CCRF-CEM, NCI-H69 and HT29 cells. | [156,157,158,159,160] |
| Viphi A-G (103–109) | Viola philippica | Cytotoxicity against MM96L, HeLa, BGC-823, HFF-1 cells, but viphi D and E showed no activity against BGC-823 cell. (IC50 = 1.03 to 7.92 μM) | [161] |
| Vaby A (110) and D (111) | Viola abyssinica | Cytotoxicity against U-937 (IC50 = 2.6 and 7.6 μM, respectively) cell. | [162] |
| Vibi G (112) and H (113) | Viola biflora | Cytotoxicity against U-937 GTB (IC50 = 0.96 and 1.6 μM, respectively) cell. | [163] |
| Vitri A (114) | Viola tricolor | Cytotoxicity against U-937 GTB (IC50 = 0.6 µM) and RPMI-8226/s (IC50 = 1 µM) cells. Cytotoxicity against U251, MDA-MB-231, A549, DU145 and BEL7402 cells (IC50 = 3.07 to 6.03 μM). |
[164,165] |
| Varv A (115) and E (116) | Viola tricolor | Cytotoxicity against U-937 GTB (IC50 = 6 and 4 µM, respectively) and RPMI-8226/s (IC50 = 3 and 4 µM, respectively) cells. | [164] |
| Vitri F (117) | Viola tricolor | Cytotoxicity against U251, MDA-MB-231, A549, DU145 and BEL7402 cel ls (IC50 = 2.74 to 6.31 μM). | [165] |
Cherimolacyclopeptides C-F (73–76) were cyclic heptapeptides obtained from the seeds of Annona cherimola in 2004 and 2005. All four compounds exhibited significant in vitro cytotoxicity against KB cells with IC50 values ranging from 0.017 to 0.97 μM [123,124,125]. Although these compounds showed anticancer potential, no further screening on other cancer cell lines or pharmacological investigations of these congeners have been reported thereafter.
Integerrimide C (77), a novel cycloheptapeptide, has been recently isolated from the latex of Jatropha integerrima. The cytotoxicity bioassay indicated its inhibitory effect against KB cells was associated with an IC50 value of 1.7 μM [126]. The discovery of integerrimide C was actually the continuation of studies on the latex of J. integerrima. Previously reported cyclopeptides integerrimides A and B were also the isolates of this particular species and exhibited moderate inhibitory activities against human melanoma (IPC-298) cell proliferation (up to 40% at 50 μM) and human pancreatic carcinoma (Capan II) cell migration (30% and 20% at 50 μM, respectively) [127].
Linoorbitides (LOBs) are a group of cyclopeptides found in flaxseed oil. Actually, flaxseed and its oil have been considered and utilized as anticancer products for many years [128]. In order to investigate whether flaxseed-derived LOBs are cytotoxic to cancer cells, Denis et al. [129]. tested the cytotoxicity of four cyclopeptides derived from flaxseed against A375 (melanoma), SKBR3 and MCF7 cells. Their study showed that LOB3 (78) possessed the highest in vitro potency among the test compounds, yet the minimum concentration of LOBs in serum had to be reached about 400–500 μg/mL for the delivery of its effectiveness. Such concentration is unlikely to be used as a drug through oral administration in humans. Nevertheless, as topical medication, LOB3 could be a potential agent for the treatment of melanoma and other skin cancers. Although flaxseed is well known for its health benefits against cancer, searching for specific compounds derived from flaxseed with high potency is still under way.
Cambodine A (79) was a novel 14-membered ring cyclopeptide extracted from the root barks of Ziziphus cambodiana. The in vitro bioassay showed that it was moderately cytotoxic against the BC-1 (lymphoma) cells with an IC50 value of 11.1 µM, whilst no toxicity was shown against the non-cancerous Vero cells [130].
The species Dianthus superbus is an important anti-inflammatory and diuretic herb in traditional Chinese medicine. Longicalycinin A (80), a cytotoxic cyclopeptide, was first isolated from the extract of D. superbus several years ago. The natural compound showed a growth inhibitory activity against HepG2 cancer cells with an IC50 value of 22.13 µM [131]. This compound had been successfully synthesized by the solid-phase methodology. When acting against in vitro Dalton’s lymphoma ascites (DLA) and Enrlich’s ascites carcinoma (EAC) cells, CTC50 values were determined to be 2.62 and 6.17 µM, respectively [132]. In the study of Tehrani et al., they demonstrated that the synthesized linear and cyclic disulfide heptapeptides of longicalycinin A showed similar inhibitory effect against HepG2 (IC50 = 16.97 and 16.91 µM, respectively) and HT29 (IC50 = 20.38 and 27.68 µM, respectively) cells. However, considering their vulnerability and proapoptotic actions against normal cells (skin fibroblast cells), the linear disulfide heptapeptides were more tempting for promotion as novel anticancer agents [133]. Although longicalycinin A and its analogs can be totally synthesized, the mechanism of actions as well as the in vivo actions of these compounds are yet to be thoroughly studied.
Similar to longicalycinin A, dianthin E (81) is another cyclic peptide isolated from Dianthus superbus. With the IC50 values of >33.3 µM against Hep3B (human hepatic carcinoma), MCF7, A549 and MDA-MB-231 cells, this compound exhibited a selective in vitro cytotoxic activity against HepG2 cells (IC50 = 3.51 µM) [134].
Orbitides are a group of plant cyclic peptides possessing a signature short chain of 5 to 11 residues, they are biosynthesized by ribosomes and characterized by their N-to-C amide bonds, but not disulfide bonds [135]. [1-8-NαC]-Zanriorb A1 (82) was a novel orbitide isolated from the leaves of Zanthoxylum riedelianum with proapoptotic effects. This compound was found to be cytotoxic against Jurkat leukemia T cells with an IC50 value of 218 nM. Regarding its molecular mechanism, it induced remarkable cell death, which would be partially inhibited by the caspase inhibitor Z-VAD-FMK. Moreover, it also induced apoptosis via reducing the levels of mitochondrial membrane potential (Ψmit) and deactivating caspase-3 [136].
Three novel cyclopeptide alkaloids, justicianenes B-D (83–85) were isolated from the Justicia procumbens L. These three compounds were evaluated their cytotoxicity against human breast cancer MCF-7, cervix carcinoma HeLa, and lung cancer A549 and H460. However, only justicianene D displayed weak cyctotoxicity against MCF-7 cells with IC50 of 90 μM [137].
Seven novel ginseng cyclopeptides (GCPs) were isolated and showed anti-proliferative activities in gastric cancer SGC-7901 cells, in which GCP-1 (86) showed the most potency with IC50 value of 37.8 μM. Regarding its molecular mechanism, it induces apoptosis by activating the caspases and regulating thioredoxin (Trx)-dependent pathways, including signal-regulating kinase 1 (ASK1), mitogen-activated protein kinases (MAPKs)-p38 and JNK pathways [138].
Cyclotides are the macrocyclic cysteine-rich peptides derived from plants, and featured with three disulfide bonds, 28 to 37 amino acids and a head-to-tail cyclized backbone in a knotted arrangement [139]. The typical structure of cyclotides makes them broadly bioactive and remarkably stable against enzymatic and thermal degradation [140].
In traditional Chinese medicine, the roots and rhizomes of Rubia plants have been used for thousands of years to treat menoxenia, contusion, rheumatism and tuberculosis [141]. Rubiaceae-type cyclopeptides (RAs) are referred to as natural cyclopeptides derived from Rubia. Up to now, 56 RAs have been isolated from Rubia plants. Due to their characteristic bicyclic structures and significant anticancer activities, RAs have gained much attention in recent years [142]. Rubipodanin A (87), a novel cyclic hexapeptide obtained from the roots and rhizomes of R. podantha, is the first naturally identified N-desmonomethyl RA. This compound was tested to be cytotoxic against three cancer cell lines including HeLa, A549 and SGC-7901 (human gastric cancer cell), with IC50 values ranging from 3.80 to 7.22 μM. However, when compared to the previously known cyclopeptide RA-V, in which two N-methyl groups are present in its backbone skeleton, the cytotoxicity of rubipodanin A was considered much weaker. In regards to its mechanisms, rubipodanin A exhibited a down-regulating effect on the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) pathway, which largely contributed to its anticancer activity [143]. Derived from the same plant species, another new RA with potent cytotoxicity was designated rubipodanin B (88). This compound showed IC50 values of 1.47 μM, 0.69 μM and 3.37 μM in MDA-MB-231, SW620 (human colorectal adenocarcinoma) and HepG2 cells respectively. Nevertheless, the anticancer activity of rubipodanin B was not associated with any down-regulation of the NF-κB signaling pathway [144]. Another cyclopeptide compound from RA family is RA-XII (89), which derived from R. yunnanensis, showed inhibitory effects on tumor growth and metastasis IC50 value of 606 nM and 96 nM on human breast cancer 4T1 cells for 24 and 48 h, respectively [145,146,147]. RA-XII also exerts antitumor activity by supressing autophagy via activating Akt-mTOR and NF-κB pathways in colocrectal cancer SW620 and HT29 cells [148]. RA-XXV (90) and RA-XXVI (91) are two new bicyclic hexapeptides, which isolated from the roots of R. cordifolia L. RA-XXV showed cytotoxic activity against human promyelocytic leukemia HL-60 and human colorectal carcinoma HCT 116 cells with IC50 of 62 nM and 28 nM, respectively. RA-XXVI showed cytotoxic activity agains HL-60 and HCT 116 cells with with IC50 of 66 nM and 51 nM, respectively [149]. To further determine RA compounds underlying mechanism, a wide screening of its molecular targets is desperately needed.
The intensive studies of famous traditional Chinese herb Hedyotis biflora had led to the isolation of HB7 (92), which is considered a potential anticancer cyclotide. By means of MTT assay, HB7 showed in vitro cytotoxicity against four pancreatic cancer cell lines BxPC3, Capan2, MOH-1 and PANC1 with IC50 values of 0.68, 0.45, 0.33 and 0.36 µM, respectively. At a non-toxic concentration of 0.05 µM, HB7 even inhibited cellular migration and invasion of Capan2 cells. When extrapolated to the in vivo xenograft mouse models, this cyclotide was observed to significantly suppress tumor growth without exhibiting any obvious organ injury or toxicity. Based on the results above, the cytotoxicity of HB7 was plausibly related to the different net charges of the cyclotide [150]. Because of its decent anticancer potential, the specific underlying mechanism of action of HB7 deserves further investigation.
Cliotides T1-T4 (93–96) were four novel cyclotides isolated from the tropical plant Clitoria ternatea, belonging to the Fabaceae family. With the prevalent distribution in almost every tissue of C. ternate, the extraction of these four cyclotides is rather efficient. These cyclotides have been confirmed to be heat-stable cysteine-rich peptides. When incubated with HeLa cells, all these four compounds yielded significant cytotoxicity with IC50 values ranging from 0.6 to 8.0 µM. Among the four, T1 and T4 were the most potent ones (IC50 = 0.6 µM) [151]. Cliotides T2 (94), T4 (96), T7 (97), T10 (98) and T12 (99) also exhibited significant cytotoxicity towards A549 cells (IC50 values ranging from 0.21 to 7.59 µM) as well as paclitaxel-resistant A549 cells (IC50 values ranging from 0.45 to 7.92 µM). Intriguingly, the IC50 values of these five test compounds were decreased by two to four folds when incubated with paclitaxel [152]. Nevertheless, the chemosensitizing ability of the cliotides became evidenced in drug-resistant cell lines, which deserves further studies on the aspects of compound charge status and in vivo efficacy. Apart from anticancer potential, cliotides, T1 to T4 in particular, also exhibited notable antimicrobial properties, particularly E. coli [152].
Among the cyclotides isolated from Psychotria leptothyrsa var. longicarpa, psyle E (100) was the most cytotoxic one. The IC50 value of psyle E against human lymphoma cell line U937-GTB was determined to be 0.76 µM. According to the reported SAR assessment, the linear structure of psyle E was required to maintain its potent cytotoxicity [153].
Vigno 5 (101) was a cyclotide discovered in Viola ignobilis [154]. This compound showed its in vitro cytotoxicity against HeLa cells with an IC50 value around 2.5 µM. Due to the complexity of its chemical structure and technical limitation, only the primary structure of vigno 5 had been elucidated so far. In the study of Esmaeili el al., vigno 5 induced apoptosis in a dose-dependent manner accompanied by nuclear shrinkage, DNA fragmentation, caspase activation and the cleavage of PARP. The vigno 5 induced-apoptosis was found to be caspase-dependent, explicitly caspase-3. To the induction of apoptotic events and mitochondrial dysfunction, a decreased level of anti-apoptotic Bcl-2 and an increased level of pro-apoptotic Bax were observed in the vigno 5-treated HeLa cells [155].
Cycloviolacin O2 (CyO2, 102), a potent cytotoxic cyclotide, was first isolated from the plant Viola odorata L. (Violaceae). The presence of glutamic acid in cycloviolacin O2 contributed much to its cytotoxic activity against U937 GTB (human lymphoma) cells with an IC50 value of 0.75 μM [156]. Besides, CyO2 also exhibited concentration-dependent cytotoxicities towards CCRF-CEM (leukemia cell line), NCI-H69 (small cell lung cancer) and HT29 cells with IC50 values of 0.3, 1.2 and 5.3 μM, respectively [157]. Disruption of the U937 cell membranes by CyO2 further indicated its membrane-disrupting activity while exerting cytotoxicity [158]. Gerlach et al. even demonstrated that CyO2 induced pore formation specifically in highly proliferating tumor cells [159]. In addition, CyO2 was also reported to exhibit bioactivities against gram-negative bacteria and HIV-1 virus via multipleregulatory mechanisms [153,160]. However, another study on the evaluation of toxicity and antitumor activity of CyO2 in mice showed that its antitumor effects were little or even absent at sublethal doses. It is worth noting that quick lethality of CyO2 was observed at 2 mg/kg whilst no abnormal signs were seen at the dose of 1.5 mg/kg in the experimental mice [157]. Thus, there is a great disparity between the results of the in vitro and in vivo experiments. For further investigation of this cyclotide, problems such as low in vivo efficacy need to be addressed.
Characterized by the macrocyclic backbones, cyclotides viphis A-G (103–109) were isolated from Viola philippica. These eight compounds were cytotoxic to human melanoma (MM96L), HeLa and human gastric adenocarcinoma (BGC-823) cells and human normal fibroblast cells (HFF-1). When applied at 3.24 and 3.17 µM, viphis D-E showed no activity against BGC-823 cell; however all these eight cyclotides displayed cytotoxicities against many cancer cell lines with IC50 values ranging from 1.03 to 15.5 µM. From the results of SAR assessments, hydrophobicity and glutamic acid residue of loop 1 are suggested of great importance for their cytotoxic activities. Further biochemical assays revealed that these viphis could be influenced by minimal sequential changes for alterations of their bioactivities [161].
Vabys A (110) and D (111) were two cyclotides derived from Viola abyssinica, a plant growing at an altitude over 3400 m. Both compounds contain charged residues; however different net charges lead to different bioactivities. Vabys A and D exhibited cytotoxicities against U-937 lymphoma cells in a dose-dependent manner, with IC50 values of 2.6 and 7.6 µM, respectively [162].
Isolated from the alpine violet Viola biflora, vibis G (112) and H (113) were two bracelet cyclotides. These two compounds showed similar cytotoxic potency (IC50 = 0.96 and 1.6 µM, respectively) against lymphoma U-937 GTB cells [163].
A bioactivity-guided fractionation of Viola tricolor led to the isolation of a cluster of highly similar cyclotides, among which vitri A (114), varv A (115) and varv E (116) were found to be cytotoxic agents. Two human cancer cell lines, U-937 GTB (IC50 = 0.6, 6 and 4 µM, respectively) and myeloma RPMI-8226/s (IC50 = 1, 3 and 4 µM, respectively) were used in the cytotoxicity assays. Sequence determination and SAR analysis demonstrated that differences in the net charges and cationic amino acid residues in these cyclotides are extremely crucial for their cytotoxicities [164]. Assessment of the cytotoxic activities against human malignant glioblastoma (U251), MDA-MB-231, A549, DU145 and human hepatoma (BEL7402) cell lines showed the inhibitory activities of vitri A (114) with IC50 values ranging from 0.97 to 1.91 µM and vitri F (117) with IC50 values ranging from 0.85 to 1.97 µM. Importantly, the distribution of highly hydrophobic residues on the surface was highlighted for the cytotoxic activities of these cyclotides [165]. Undeniably, plant-derived cyclotides are the intriguing candidates for drug development as anticancer therapeutics. Owing to the abundant source of cyclotides, V. tricolor warrants further studies for the identification of new bioactive compounds and their anticancer mechanism of actions.
4. Bacteria-Derived Anticancer Cyclopeptides
Bacteria are single celled micro-organisms that thrive in diverse environments. In fact, a significant number of bacteria are from oceans. It is hard to imagine that there are actually 40 million bacterial cells living in 1 g of soil and 1 million bacterial cells in 1 mL of freshwater [166]. Diverse habitats and complex relationships with other organisms, such as antagonistic actions towards some worms and molecules, cause bacteria to produce many different types of metabolites, and some of them are highly bioactive for pharmaceutical uses in humans [167]. In recent years, numerous bioactive cyclopeptides have been isolated from bacteria possessing anticancer properties. In this section, these cyclopeptides and their cytotoxicities against cancer cells are discussed. The chemical structures of these cyclopeptides are provided in Figure 6 and their activities were listed in Table 3.
Figure 6.

Structure of cyclopeptides (118–119) derived from non-marine bacteria.
Table 3.
Bacteria-derived anticancer cyclopeptides.
| Name | Biological Source | Anticancer Activity | Reference |
|---|---|---|---|
| Anticancer Cyclopeptides Derived from Non-Marine Bacteria | |||
| YM753 (spiruchostatin A/OBP-801) (118) | Pseudomonas sp. | In vitro growth inhibitory ceffects on many cancer cells including WiDr, melanoma (GI50 = 5.0 nM), Calu-3 (GI50 = 1.6 nM), HOS (GI50 = 3.8 nM) cells and etc.; In vivo antitumor activity on WiDr xenograft model. | [168,169,170,171] |
| Telomestatin (119) | Streptomyces anulatus 3533-SV4 | Cytotoxicity against MiaPaCa (IC50 = 0.5 µM) cell and four neuroblastoma cells (IC50 = 0.8 to 4.0 µM). Specific telomerase inhibitor in cancer cells inhibition (inhibit cellular senescence). |
[172,173,174,175,176,177,178] |
| Anticancer Cyclopeptides Derived from Marine Bacteria | |||
| Thiocoraline (120) | Gram-positive bacteria Micromonospora sp. ACM2-092 and Micromonospora sp. ML1 | Cytotoxicity against P388 (IC50 = 1.72 nM), A549 (IC50 = 1.72 nM), MEL28 (IC50 = 1.72 nM) and HT29 (IC50 = 8.64 nM) and MTC-TT (IC50 = 7.6 nM) cells. | [181,182,183,184,185] |
| Ohmyungsamycins A (121) and B (122) | Streptomyces sp. | Cytotoxicity against HCT116, A549, SNU-638, MDA-MB-231 and SK-HEP-1 cells. (IC50 = 359 to 816 nM and 12.4 to 16.8 µM, respectively) | [186,187] |
| Arenamides A (123) and B (124) | Salinispora arenicola | Cytotoxicity against HCT116 (IC50 = 1.97 and 2.99 μM, respectively) cell. | [188,189] |
| Piperazimycins A-C (125–127) | Streptomyces sp. | Cytotoxicity against HCT116 cell. (Average GI50 = 0.1 µM) Piperazimycin A showed a mean GI50 of 100 nM in NCI 60 cancer cell line panel screening. |
[190,191] |
| Kailuins A-D (128–131) | Gram-negative bacterium BH-107 | Inhibitory activities against A549, MCF7 and HT29 cells. (IC50 = 2.66 to 5.52 µM) | [192] |
| Mixirins A-C (132–134) | Bacillus genus | Anti-proliferative activity of HCT116 (IC50 = 0.65, 1.6 and 1.26 μM, respectively) cell. | [193] |
| Bacilohydrin A (135) | Bacillus sp. SY27F | Cytotoxicity against HepG2, MCF7 and DU145 cells. (IC50 = 50.3 to 175.1 nM) | [195] |
| Dolyemycins A (136) and B (137) | Streptomyces griseus subsp. griseus HYS31 | Anti-proliferative activities against A549 cells. (IC50 = 1 and 1.2 µM, respectively) | [196] |
| Sungsanpin (138) | Streptomyces sp. | Anti-invasion effects on A549 cell. | [197] |
| Marthiapeptide A (139) | Actinomycete Marinactinospora thermotolerans | Cytotoxicity against SF-268, MCF7, NCI-H460 and HepG2 cell. (IC50 = 0.38 to 0.52 µM) | [198,199] |
| Mechercharstatin A (140) | Thermoactinomyces sp. YM3-251 | Cytotoxicity against Jurkat cell (IC50 = 4.6 × 10−8 M), A549 (IC50 = 0.03 µM), HT29 (IC50 = 0.04 µM) and MDA-MB-231 (IC50 = 0.09 µM) cells. | [198,200,201] |
| Urukthapelstatin A (141) | Mechercharimyces asporophorigenens YM11-542 | Cytotoxicity against several cancer cell lines. (Mean GI50 = 1.55 μM) | [202,203,204,205] |
| Anticancer Cyclopeptides Derived from Cyanobacteria | |||
| Microcystin-LR (142) | Mainly from Microcystis aeruginosa | Cytotoxicity against BxPC-3 (IC50 = 83.50 nM) and MIA PaCa2 (IC50 = 2.14 µM) and transfected HeLa (IC50 = 1 nM) cells. | [211,212,213,214,215,216] |
| Apratoxin A (143) | Lyngbya sp. | Cytotoxicity against KB (IC50 = 0.52 nM), LoVo (IC50 = 0.36 nM) and HCT116 (IC50 = 1.21 nM) cells. | [217,218,219,220,221] |
| Apratoxins B (144) and C (145) | Lyngbya sp. | Cytotoxicity against KB (IC50 = 21.3 and 1.0 nM, respectively) and LoVo (IC50= 10.8 and 0.73 nM, respectively) cells. | [222] |
| Apratoxin D (146) | Lyngbya majuscula and Lyngbya sordida | Cytotoxicity against H460 (IC50 = 2.6 nM) cell. | [223] |
| Apratoxin E (147) | Lyngbya bouillonii strain PS372 | Cytotoxicity against HT29 (IC50 = 21 nM), HeLa (IC50 = 72 nM) and U2OS (IC50 = 59 nM) cells. | [224] |
| Apratoxins F (148) and G (149) | Lyngbya bouillonii | Cytotoxicity against H460 (IC50 = 2 and 14 nM, respectively) and HCT116 cells. | [225] |
| Aurilide B (150) | Lyngbya majuscula | In vitro cytotoxicity against H460 (LC50 = 0.04 µM) and neuro-2a (LC50 = 0.01 µM) cells. In vivo net tumor cell killing activity |
[226,227] |
| Aurilide C (151) | Lyngbya majuscula | Cytotoxicity against H460 (LC50 = 0.13 µM) and neuro-2a (LC50 = 0.05 µM) cells. | [226] |
| Lagunamide A (152) | Lyngbya majuscula | Cytotoxicity against P388, A549, PC3, HCT8 and SK-OV3 cells. (IC50 = 1.64 to 6.4 nM) | [228,229] |
| Lagunamide B (153) | Lyngbya majuscula | Cytotoxicity against P388 (IC50 = 20.5 nM) and HCT8 (IC50 = 5.2 nM) cells. | [228,229] |
| Lagunamide C (154) | Lyngbya majuscula | Cytotoxicity against P388, A549, PC3, HCT8 and SK-OV3 cells (IC50 = 2.1 to 24.2 nM). | [228,230] |
| Homodolastatin 16 (155) | Lyngbya majuscula | Cytotoxicity against WHCO1 (IC50 = 4.19 µM), WHCO6 (IC50 = 9.85 µM) and ME180 (IC50 = 8.09 µM) cells. | [231] |
| Lyngbyabellin A (156) | Lyngbya majuscula | Cytotoxicity against KB (IC50 = 0.04 µM) and LoVo (IC50 = 0.72 µM) cells. Specific cytoskeletal-disrupting effects to microfilaments in A-10 cells. |
[232,233] |
| Lyngbyabellins D (157), F (158) and H (159) | Lyngbya majuscula | Cytotoxicity against H460 and KB cells. (IC50 or LC50 = 0.1 to 0.4 µM) | [234] |
| Lyngbyabellin N (160) | Moorea bouillonii | Cytotoxicity against HCT116 (IC50 = 40.9 nM) cell. | [235] |
| Palmyramide A (161) | Lyngbya majuscula | Modest cytotoxicity against H460 cell (IC50 = 39.7 µM). Blockage of the voltage-gated sodium channel in neuro-2a cell. |
[236] |
| Hoiamide A (162) | Lyngbya majuscula and phormidium gracile | Cytotoxicity against H460 (IC50 = 11.2 µM) cell. | [237,238,239] |
| Hoiamide B (163) | A sample of marine cyanobacteria collected in Gallows reef, unclear species | Cytotoxicity against H460 (IC50 = 8.3 µM) cell. | [237,238] |
| Wewakazole (164) | Lyngbya majuscula | Cytotoxicity against H460 (IC50 = 10 µM) cell. | [240,241,242,243] |
| Wewakazole B (165) | Moorea peoducens | Cytotoxicity against H460 (IC50 = 1.0 µM) and MCF7 (IC50 = 0.58 µM) cells. | [241,242,243] |
| Hectochlorin (166) | Lyngbya majuscula | Average GI50 value of 5.1 µM in the tests against a panel of NCI 60 tumor cell lines. Cytotoxicity against KB (ED50 = 0.86 µM) and NCI-H187 (ED50 = 1.20 µM) cells. |
[244,245,246,247] |
| Desmethoxymajusculamide C (DMMC) (167) | Lyngbya majuscula | Selectivity and cytotoxicity against HCT116 (IC50 = 0.016 µM), H460 (IC50 = 0.094 µM) and MDA-MB-435 (IC50 = 0.23 µM) cells. | [248] |
| Hantupeptin A (168) | Lyngbya majuscula | Cytotoxicity against MOLT-4 (IC50 = 32 nM) and MCF7 (IC50 = 4.0 µM) cells. | [249] |
| Hantupeptins B (169) and C (170) | Lyngbya majuscula | Cytotoxicity against MOLT-4 (IC50 = 0.2 and 3.0 µM, respectively) and MCF7 (IC50 = 0.5 and 1.0 µM, respectively) cells. | [249,250] |
| Laxaphycin A (171) | Lyngbya majuscula | Obvious biological synergism when acting against DLD1 and MDR cells in the presence of laxaphycins A and B. | [251,252] |
| Laxaphycin B (172) | Lyngbya majuscula | Cytotoxicity against CCRF-CEM (IC50 = 1.11 µM), CEM/VLB100 (IC50 = 1.02 µM) and CEM/VM-1 (IC50 = 1.37 µM) cells. Anti-proliferative effects on A549, MCF7, PA1 and PC3 cells. (IC50 = 0.19 to 1.0 µM) Obvious biological synergism when acting against DLD1 and MDR cells in the presence of laxaphycins A and B. |
[251,252] |
| Laxaphycins B2 (173) and B3 (174) | Lyngbya majuscula | Moderate cytotoxicities against CCRF-CEM, CEM/VLB100 and CEM/VM-1 cells. | [252] |
| Laxaphycin B4 (175) | Hormothamnion enteromorphoides | Growth-inhibitory activity and synergistic effect with laxaphycin A when acting against HCT116 (IC50 = 1.7 µM) cell. | [253] |
| Obyanamide (176) | Lyngbya confervoides | Cytotoxicity against KB (IC50 = 0.97 µM) and LoVo (IC50 = 5.24 µM) cells. | [254,255] |
| Grassypeptolide A (177) | Lyngbya confervoides | Cytotoxicity against U2OS (IC50 = 2.2 µM), HeLa (IC50 = 1.0 µM), HT29 (IC50 = 1.5 µM) and IMR-32 (IC50 = 4.2µM) cells. | [256,257,258] |
| Grassypeptolides B (178) and C (179) | Lyngbya confervoides | Anti-proliferative effects on HT29 (IC50 = 2.97 µM and 76.7 nM, respectively) and HeLa (IC50 = 2.93 µM and 44.6 nM, respectively) cells. | [257] |
| Grassypeptolides D (180) and E (181) | Leptolyngbya sp. | Cytotoxicity against HeLa (IC50 = 335 and 192 nM, respectively) and neuro-2a (IC50 = 599 and 407 nM, respectively) cells. | [259] |
| Palauamide (182) | Lyngbya sp. | Cytotoxicity against KB (IC50 = 13 nM) cell. | [260,261,262] |
| Coibamide A (183) | Leptolyngbya sp. | Cytotoxicity against NCI-H460 and neuro-2a cells. (LC50 < 17.9 nM) | [263,264,265] |
| Ulongamides A-E (184–188) | Lyngbya sp. | Cytotoxicity against KB (IC50~1 µM) and LoVo (IC50~5 µM) cells. | [266,267,268] |
| Ulongapeptin (189) | Lyngbya sp. | Cytotoxicity against KB (IC50 = 0.63 µM) cell. | [269] |
| Odoamide (190) | Okeania sp. | Cytotoxicity against HeLa S3 (IC50 = 26.3 nM) cell. | [270,271] |
| Cryptophycin 1 (191) | Nostoc sp. | Broad-spectrum cytotoxicity against cell lines such as L1210 (IC50 = 4 pM), SKOV3 (IC50 = 7 pM), MCF7 (IC50 = 16 pM) and etc. | [272,273,274,275,276,277,278,279] |
| Largazole (192) | Symploca sp. | Cytotoxicity against MDA-MB-231 (GI50 = 122 nM, LC50 = 272 nM), NmuMG (GI50 = 7.7 nM, LC50 = 117 nM), U2OS (GI50 = 55 nM, LC50 = 94 nM), NIH3T3 (GI50 = 480 nM, LC50 > 8 µM), HCT116 (GI50 = 0.08 µM) and A549 (GI50 = 0.32 µM) cells. | [280,281,282,283,284,285] |
| Symplocamide A (193) | Symploca sp. | Cytotoxicity against H460 (IC50 = 40 nM) and neuro-2a (IC50 = 29 nM) cells. | [286,287] |
| Tasipeptins A (194) and B (195) | Symploca sp. | Cytotoxicity against KB (IC50 = 0.93 and 0.82 µM, respectively) cell. | [288,289] |
| Veraguamides A-G (196–202) | Symploca cf. hydnoides | Cytotoxicity against HT29 (IC50 = 0.84 to 49 µM) and HeLa (IC50 = 0.54 to 49 µM) cells. | [290] |
| Floridamide (203) | Moorea producens | Cytotoxicity against H460 and neuro-2a cells. (EC50 = 1.89 × 10−5 µM) | [291] |
| Viequeamide A (204) | Rivularia sp. | Cytotoxicity against H460 (IC50 = 60 nM) cell. | [292,293] |
4.1. Anticancer Cyclopeptides Derived from Non-Marine Bacteria
YM753 (spiruchostatin A/OBP-801) (118), a disulfide bond-containing bicyclic depsipeptide, was isolated from Pseudomonas sp. and identified as a gene-enhancing substance at first. Subsequent studies also demonstrated it also exerted TGF-β-like activity and HDAC inhibitory properties [168,169]. As a HDAC inhibitor, YM753 induced histone acetylation through cell cycle arrest at the G1 and G2/M phases and enhancing p21WAF1/Cip1 expression and apoptosis in WiDr cells [169]. When applied with celecoxib as a combo treatment, a synergetic anti-proliferative effect was observed in the high-grade bladder cancer cells. Such anti-proliferation of cancer cells was found to be caspase dependent in both in vitro and in vivo experiments [170]. The combinatory treatment with OBP-801 and fibroblast growth factor receptor (FGFR) inhibitor BGJ298, a significant attenuation on cell growth was obtained in the muscle-invasive bladder cancer cells, and a regulation of the caspase and Bim pathways was demonstrated [171]. Accordingly, these evidences have demonstrated the great anticancer potential of combo remedies YM753/OBP-801/spiruchostatin A.
Telomestatin (119) was a macrocyclic peptide first isolated from Streptomyces anulatus 3533-SV4 and had been shown to act as a telomerase inhibitor in cancer cells [172]. Telomerase is indeed a significant enzyme responsible for cellular senescence and tumorigenic conversion. Telomestatin can specifically bind to the telomeric G-quadruplex structure at the 3′ telomere end and inhibit the activity of telomerase [173]. When acting against MiaPaCa (human pancreatic carcinoma) cells, the IC50 value of telomestatin was determined to be 0.5 μM [174]. This compound also exhibited significant cytotoxicity against four neuroblastoma cell lines with IC50 values ranging from 0.8 to 4.0 μM. Telomere shortening, cell cycle arrest and apoptosis were observed after long-term incubation with telomestain at non-cytotoxic concentrations [175]. Due to its potent cytotoxicity against cancer cells, telomestatin is considered as a potential therapeutic agent in treating cancers [176]. The pharmacological characteristics of telomestatin promotes it to be served as a reference standard substance in the in vitro telomere shortening activity assays [177]. Although a great number of telomerase inhibitors have been isolated from the nature, telomestatin remains the strongest cytotoxic agent of its kind. However, the limited production has impeded the wide uses of telomestatin. Fortunately, the total synthesis of telomestatin was achieved by the coupling reactions of cysteine-containing trisoxazole amine and serine-containing trisoxazole carboxylic acid in 2006 [178]. In a research reported in 2017, the biosynthesis of telometstatin was further proposed and carried out using a cluster of genes by assembling the amino acids cysteine, serine and threonine, which may be provide a promising approach for mass production of this compound for future preclinical and clinical trials [176].
4.2. Anticancer Cyclopeptides Derived from Marine Bacteria
As the smallest autonomous organisms in the ocean, marine bacteria can exist independently or symbiotically with other marine microorganisms [177,179]. Because marine bacteria are always the excellent origins of natural therapeutic agents, they are described as the chemical gold [180]. Numerous compounds, including cyclopeptides, with anticancer activities have been extracted from marine bacteria. The chemical structures of these cyclopeptides are provided in Figure 7 and their activities were listed in Table 3.
Figure 7.

Structure of cyclopeptides (120–141) derived from marine bacteria.
Thiocoraline (120) was a potential antitumor cyclic depsipeptide produced by the the Gram-positive bacteria Micromonospora sp. ACM2-092 and Micromonospora sp. ML1 [181]. This compound exerted cytotoxicities against P388, A549, MEL28 and HT29 cells with IC50 values of 1.72, 1.72, 1.72 and 8.64 nM, respectively [182]. By using the human colorectal adenocarcinoma LoVo and SW620 cells as the cellular models, thiocoraline was found to induce cell cycle arrest at the G1 phase. Its anticancer effects may also be related to its inhibition of DNA polymerase α activities [183]. To the human pancreatic carcinoid BON and pulmonary carcinoid H727 cells, the thiocoraline treatment led to increased expression levels of poly ADP ribose polymerase (PARP) and X-linked inhibitor of apoptosis protein (XIAP), indicating that the anti-proliferative activity of this compound was highly associated with the induction of apoptosis [184]. Furthermore, in the aggressive neuroendocrine tumor medullary thyroid cancer MTC-TT cells, the IC50 value of the compound was determined to be 7.6 nM. The studies on its mechanism of action demonstrated that thiocoraline induced induce cell cycle arrest at the G1 phase and activated the NOTCH pathway [185]. Overall, the findings above illustrated that thilcoraline is worthy for further investigation to reveal its anticancer potential as a palliative therapeutic agent.
Ohmyungsamycins A (121) and B (122) were two new cyclodepsipeptides purified from the marine bacteria of Streptomyces sp. of a volcanic island area in Korea. Both cyclodepsipeptides showed selective cytotoxicity towards cancer cells over normal cells as no notable cytotoxicity (IC50 > 40 µM) against normal lung epithelial cells (MRC-5) was observed. When applied to the HCT116, A549, SNU-638 (human gastric adenocarcinoma), MDA-MB-231 and SK-HEP-1 (human liver adenocarcinoma) cells, ohmyungsamycin A (IC50 values ranging from 359 to 816 nM) exhibited inhibitory activities much higher than ohmyungmycin B (IC50 values ranging from 12.4 to 16.8 µM). The only structural difference between these two compounds is that the N-methylvaline moiety on the side chain of ohmyungmycin A is replaced by the N,N-dimethylvaline moiety in ohmyungmycin B. In the CRC HCT116 cells, ohmyungmycin A was found 78 times more potent than the positive reference drug etoposide. On the other hand, ohmyungmycin A also displayed much better antibacterial activities than ohmyungmycin B. Collectively, the N-methyl group at the terminus of ohmyungmycin B is a hinder to its bioactivities [186]. As total syntheses of ohmyungmycins A and B had been established, investigations of these two compounds could be comprehensively performed. A few reports even suggested that these two cyclopeptides deliver antituberculosis properties [187]. Therefore, studies on synthesizing their analogs with higher selectivity and bioactivities and exploring their mechanisms of actions are inspired.
Arenamides A (123) and B (124) were two novel cyclohexadepsipeptides isolated from the marine bacterial strain Salinispora arenicola. These two compounds merely exhibited moderate in vitro cytotoxicity against HCT116 cells with IC50 values of 19.7 and 29.8 µM, respectively. However, arenamides A and B were able to block tumor necrosis factor (TNF)-induced activities at rather low concentrations (IC50 = 5.51 and 2.64 µM, respectively). When the lipopolysaccharides (LPS)-challenged RAW 264.7 macrophages were treated with arenamides A or B, the generation of nitric oxide (NO) and prostaglandin E2 (PGE2), so as the major oxidative mediators of the NFκB pathway, were significantly inhibited whereas cell viability was not influenced. The above data suggested these two compounds carry chemopreventive potential [188]. The successful total synthesis of arenamide A has been reported [189]. which provided an alternative approach to obtain this compound as well as to synthesize its anologues.
Piperazimycins A-C (125–127) were three cytotoxic hexadepsipeptides isolated from the genus Streptomyces living in the marine sediment near the island of Guam. They showed in vitro cytotoxicities against HCT116 accompanied with an average GI50 value of 0.1 µM. Such low value unveiled the significant anticancer potency of this type of compounds. When screened against the NCI 60 cancer cell line panel, piperazimycin A exhibited a mean GI50 value of only 100 nM. Although piperazimycin A was shown with a selective growth inhibition against solid tumor cell lines over leukemia cell lines, the general mode of cytotoxicity was observed in the NCI cell line panel assay [190]. Further comprehensive studies on piperazimycin A can be proceeded as the total synthesis of piperazimycin A had been accomplished [191].
Obtained from a piece of driftwood collected at Kailua beach in Oahu, a Gram-negative bacterium BH-107 yielded four cyclic acyldepsipeptides kailuins A-D (128–131). These compounds represent a new type of lipopeptides with five amino acids and a β-acyloxy fatty acid moiety arranged in a 19-membered ring. All these four compounds displayed inhibitory activities against A549, MCF7 and HT29 cells with GI50 values ranging from 2.66–5.52 µM [192].
Mixirins A-C (132–134) were three cyclic acylpeptides obtained from marine bacteria of the Bacillus genus. The anti-proliferative activities of these three compounds were evaluated in HCT116 cells whereas their IC50 values were determined to be 0.65, 1.6 and 1.26 µM, respectively [193].
Four cyclopeptides were isolated from the ethyl acetate extract of Bacillus velezensis sp. RA5401. All four compounds showed anti-proliferative effects against human breast cancer MDA-MB-231 cells. Their molecular-docking studies showed that some G-protein coupled receptors (GPCRs) could be the potential binding targets. However, the IC50 of these four compounds and the mechanism of actions against cancer cells remain unknown and are worthy for further studies [194].
Bacillus sp. SY27F collected from the Indian Ocean hydrothermal vent yielded bacilohydrin A (135), a novel cyclic lipopeptide with significant anticancer potential. Among the six tested cancer cell lines, bacilohydrin A exhibited rather high cytotoxicity against the HepG2, MCF7 and DU145 cells, with IC50 values varying from 50.3 to 175.1 nM. In addition to its cytotoxic effect, cell shrinkage, condensation and DNA fragmentation were also observed as the result of apoptosis [195]. More studies are therefore needed to explore its detailed mechanism of action and therapeutic potential for cancer treatment.
Dolyemycins A (136) and B (137) are cyclopeptides isolated from Streptomyces griseus subsp, griseus HYS31 by bio-guided isolation. Both compounds showed anti-proliferative activities against human lung cancer A549 cells with IC50 values of 1.0 and 1.2 µM, respectively [196].
Sungsanpin (138) is a lasso peptide that contains a cyclic peptide and a tail. This compound was first isolated from the bacteria of Streptomyces sp. In the deep-sea sediment. In the cell invasion assay of A549 cells, the invading activity of the cells was significantly inhibited by sungsanpin. The anti-invasive mechanism of this compound was related to its ability to increase the expression levels of tissue inhibitor of metallopeptidase-1 (TIMP-1) and TIMP-2. Consequently, the degradation of matrix metalloproteinases (MMPs) was inhibited, and thus resulting in a decreased metastasis of the A549 cells [197].
Marthiapeptide A (139) was a novel cyclic peptide isolated from a marine-derived actinomycete Marinactinospora thermotolerans. This compound displayed significant cytotoxicity towards SF-268, MCF7, NCI-H460 (human lung tumor cells) and HepG2 cancer cells with IC50 values ranging from 0.38 to 0.52 μM. Its cytotoxic potency was actually 5 to 10 folds higher than the positive control compound—cisplatin. In addition to its potent cytotoxicity, marthiapeptide A was demonstrated to exert anti-microbial activity against a panel of Gram-positive bacteria [198]. A rational synthetic route of marthiapeptide A had been described to produce a sufficient amount of this compound for further bioactivity evaluation [199]. The synthesis of additional analogs may be inspired by the preparation of marthiapeptide A and its macrocyclic backbone.
Another cytotoxic cyclic peptide named mechercharstatin A (former name, mechercharmycin A) (140) was isolated from the marine-derived Thermoactinomyces sp. YM3-251. This compound exhibited strong cytotoxicity against A549 and Jurkat cells with IC50 values of 0.040 and 0.046 μM, respectively [200]. The GI50 values of mechercharstatin A against A549, HT29 and MDA-MB-231 cells were determined to be 0.03, 0.04 and 0.09 μM, respectively. The anti-proliferative action of mechercharstatin A was associated with cell cycle arrest at the G2 phase and apoptotic events [201]. Although the chemical structure of mechercharstatin A is highly similar to telomestatin (119), a telomerase inhibitor described previously in this section, no significant inhibition on telomerase, DNA polymerase or reverse transcriptase was observed after the treatment with mechercharstatin A [198]. Therefore, more specific and concrete pharmacological studies are needed in order to elucidate its special mechanism of action.
Urukthapelstatin A (141) was another a cytotoxic cyclic peptide purified from the cultured mycelia of the marine-derived Thermoactinomycetaceae bacterium Mechercharimyces asporophorigenens YM11-542. This compound was assayed to be cytotoxic against a spectrum of cancer cell lines with the mean GI50 value of 1.55 μM, among which lung and ovarian cancer cell lines were the most sensitive. This peptide was structurally similar to telomestatin (119) as well; however, urukthapelstatin A showed no significant effect on telomerase inhibition. Thus, the cytotoxic activity of this compound is derived a mechanism very different from telomestatin, and its actions are more or less like the ones of mechercharstatin A (140) [202]. Regarding the synthetic aspect, both total synthesis and biomimmetic synthesis of urukthapelstatin A have been achieved correspondingly, which allow an efficient production of new analogs and further extensive SAR studies [203,204,205].
4.3. Anticancer Cyclopeptides Derived from Cyanobacteria
Cyanobacteria are prokaryotes with ability to produce oxygen through photosynthesis [206]. There are around 150 genera and 2000 species in the phylum cyanobacteria. A great number of peptides found in cyanobacteria have been confirmed to be cytotoxic against human cancer cells, and some of them were used as the structural scaffolds for the synthesis of novel anticancer agents [207]. The chemical structures of these cyclopeptides are provided in Figure 8 and their activities were listed in Table 3.
Figure 8.



Structure of cyclopeptides (142–204) derived from cyanobacteria.
Microcystins are a kind of cyanopeptides being the most extensively studied due to their severe toxicological influence in the ecosystem [208]. Accumulating evidence showed that such toxins can cause human poisoning because they widely exist in drinking water even after filtration [209]. Microcystins are a category of toxins with a cyclic heptapeptide structure produced by some cyanobacterial blooms. Over 80 microcystins have been isolated so far [210]. Among them, microcystin-LR (142) is the most common encountered variant owing to its potent toxicity. Microcystin-LR is a cyclic heptapeptide mainly produced by Microcystis aeruginosa [211]. It has been broadly reported that micrcystin-LR is a significant pathological agent causing carcinogenesis and organ injury [212]. When human fetal L-02 hepatocytes were chronically exposed to low-dose microcystin-LR, increased tumor formation was observed as a result of gankyrin activation [213]. Moreover, microcystin-LR has been suggested with great potential for immunotoxicity and may lead to various diseases and even cancers [211]. However, microcystin-LR has also been reported with notable cytotoxicity against a wide spectrum of cancer cells. Particularly, microcystin-LR was demonstrated to directly interact with organic-anion-transporting polypeptides (OATP1B1 and 1B3), and consequently resulted in remarkable anti-proliferative and pro-apoptotic effects in pancreatic cancer cells. The IC50 values of microcystin-LR against both human pancreatic cancer BxPC-3 and MIA PaCa2 cells were determined to be 83.50 nM and 2.14 µM, respectively [214]. Niedermeyer et al. found that OATP1B1 and 1B3 are the essential membrane transporters for microcystin to be uptaken into human cells. When compared to the normal HeLa cells, microcystin-LR was more potent in inhibiting the proliferation of HeLa cells overexpressing OATP1B1, with an IC50 value of around 1 nM. Thus, microcystin-LR is of great potential to be an anticancer agent against OATP1B1 transporter-overexpressing tumor cells [215]. Furthermore, OATP1B3 transporters-overexpressing cancer cells were found particularly sensitive to a series of microcystin analogs [216]. Taken together, microcystin-LR, as well as its analogs are worthy for further studies on their anticancer mechanisms.
Apratoxins are a novel family of cytotoxic cyclodepsipeptides, and they were isolated from cyanobacterium Lyngbya sp. with potent anti-proliferative effects. Apratoxin A (143) was a natural peptide possessing a signature cyclic structure purified from the secondary metabolites of marine cyanobacterium Lyngbya majuscula. Apratoxin A showed significant in vitro cytotoxicity against KB and LoVo cancer cells with IC50 values of 0.52 and 0.36 nM, respectively. Nevertheless, this antitumor cyclopeptide was toxic and poorly tolerated in xenograft mouse models [217]. Mechanistic studies have become the core researching area of this compound. The in vitro anti-proliferative activity of Apratoxin A was associated with the G1 cell cycle arrest and apoptosis via an inhibition of signal transducer and activator of transcription 3 (STAT3) [218]. Liu et al. [219] discovered that apratoxin A could induce degradation of cancer-related receptors, tyrosine kinases, by inhibiting their N-glycosylation and cotranslational translocation in the secretory pathway. A subsequent biochemical study demonstrated that apratoxin A prevented the protein cotranslational translocation by a direct blockade of the protein translocation channel Sec61α [220]. Further, Huang et al. [221] found that the in vivo toxicity of apratoxin A was the result of severe pancreatic atrophy upon the high-level exposure of the drug. They also identified Sec61 complex was a molecular target of apratoxin A in the secretory pathway. Taken together, the cytotoxicity mechanisms of apratoxin A are complicated, and its in vivo application remains controversial. However, apratoxin A is an inspiring tool to the study of the secretory pathway and cotranslational translocation inhibition for the development of anticancer drugs. Two other members of the family, apratoxins B (144) and C (145) are also derived from the secondary metabolites of Lyngbya sp. These two cyclopeptides also delivered cytotoxicities against human cancer cell lines, KB (IC50 = 21.3 and 1.0 nM, respectively) and LoVo cells (IC50 = 10.8 and 0.73 nM, respectively). The results indicated that apratoxin C was less potent than apratoxins A and B, though apratoxin C is different from apratoxin B by only the position of a methyl group [222]. Likewise, isolated from the marine cyanobacteria L. majuscula and L. sordida, apratoxin D (146) has a similar structure to apratoxin A. It exerted potent cytotoxicity to cancer cells as well. Its IC50 value against H460 cells was determined to be 2.6 nM. From the SAR assessments of apratoxins A-D, we noticed that the additional lipopeptide tail was responsible for the cytotoxic activities of this type of compounds; however, Suyama et al. made a contrary claim [223]. This SAR information is extremely useful to the design of additional analogs. Apratoxin E (147) was another congener isolated from L. bouillonii strain PS372. However, comparing to the IC50 values of apratoxin A (IC50 = 1.4, 10 and 10 nM, respectively), apratoxin E (IC50 = 21, 72 and 59 nM, respectively) showed less potency when testing against the HT29, HeLa and U2OS (human bone osteosarcoma epithelial) cells. The cell viability test results indicated that the loss of a hydroxy group and the formation of a conjugated double bond in apratoxin E attributed to the significant reduction of bioactivities [224]. Apratoxins F (148) and G (149) were two novel cytotoxins obtained from L. bouillonii cyanobacterium collected in the Central Pacific. When incubated with H460 cancer cells, the inhibitory IC50 values of apratoxins F and G were found to be 2 and 14 nM, respectively; however, they are less cytotoxic to HCT116 cells [apratoxins F (IC50 = 36.7 nM) and G (insufficient to test the IC50 value)]. According to the SAR assessment results, the N-methyl alanine residue was not the key functional group for the cytotoxicity of apratoxins [225]. Overall, disparate moieties and regions are usually crucial for biological activities, which can be of great use for further synthesis and drug development.
Unlike aurilide (66) isolated from sea hare, aurilides B (150) and C (151) were obtained from cyanobacterium L. majuscula. Both aurilides B and C were cyclic depsipeptides with potent in vitro cytotoxic activities. In H460 cells and mouse neuroblastoma (neuro-2a) cells, the LC50 values of aurilide B were found to be 0.04 and 0.01 µM, respectively whereas the LC50 values of aurilide C were found to be 0.13 and 0.05 µM, respectively. Moreover, aurilide B was demonstrated to induce microfilament network disruption against A-10 smooth muscle cells with an IC50 value of 2.9 µM. In an in vivo experiment, aurilide B was shown to deliver net tumor cell killing activity in the NCI’s hollow fiber assay [226]. A quantitative shRNA screening experiment illustrated that cytotoxicity of aurilide B was derived from mitochondrial apoptosis, which was tightly regulated by the gene sodium/potassium-transporting ATPase subunit alpha-1 (ATP1A1) [227]. Collectively, the promising bioassay data suggested that aurilide B has potential for further preclinical studies as a novel anticancer agent.
Lagunamides A-C (152–154) were cyclic depsipeptides isolated from L. majuscula with anti-proliferative effects against several cancer cell lines [228]. Tripathi et al. assessed the inhibitory activities of lagunamides A against the cancer cell lines P388, A549, PC3, HCT8 (human colorectal adenocarcinoma) as well as the SK-OV3 cells. The IC50 values of lagunamides A were found ranging from 1.64 to 6.4 nM. Lagunamide B was only tested for its cytotoxicities against P388 and HCT8 cell lines; its IC50 values were determined to be 20.5 and 5.2 nM, respectively. Subsequent studies revealed that the cytotoxic effects of lagunamides A and B was highly associated with the induction of mitochondrial apoptosis [229]. As for lagunamide C, its IC50 values varied from 2.1 nM to 24.2 nM against a panel of cancer cell lines such as P388, A549, PC3, HCT8 and SK-OV3 [230].
Another cyclic depsipeptide identified from L. majuscula was homodolastatin 16 (155), which was a homologue of the potent anticancer agent dolastatin 16. Compared to dolastatin 16, homodolastatin 16 only exhibited moderate activities against esophageal (WHCO1 and WHCO6) and cervical (ME180) cancer cell lines with IC50 values of 4.19, 9.85 and 8.09 µM, respectively [231].
Lyngbyabellin A (156) was another cytotoxic cyclic depsipeptide derived from Guamanian strain of L. majuscula. Other than cytotoxic activities against KB cells (IC50 = 0.04 µM) and LoVo cells (IC50 = 0.72 µM), lyngbyabellin A also exhibited specific cytoskeletal-disrupting effects to microfilaments in A-10 cells at concentrations ranging from 0.01 to 7.24 µM [232]. To further investigate the properties of this natural cyanobacterium metabolite, the total synthesis of lyngbyabellin A was subsequently achieved [233]. Similar to lyngbyabellin A, lyngbyabellins D (157), F (158) and H (159) were also isolated from L. majuscula. They exhibited cytotoxicity against H460 and KB cancer cells with IC50 or LC50 values ranging from 0.1 to 0.4 µM [234]. In the subsequent studies, Choi et al. identified lyngbyabellin N (160) from Moorea bouillonii, a rather new cytotoxic member of the lyngbyabellin class. This compound showed a potent anticancer potential against the CRC HCT116 cells with IC50 value of 40.9 nM [235]. Although a series of lyngbyabellins had been proved with anticancer properties, limited studies on these congeners largely restricted the ultimate exploration of these compounds as potential therapeutic agents.
Palmyramide A (161), another cyclic depsipeptide, was first isolated from L. majuscula. In the H460 cell line, palmyramide A displayed merely a weak cytotoxicity (IC50 = 39.7 µM). However, in neuro-2a cells, palmyramide A led to the blockage of the voltage-gated sodium channel with an IC50 value of 17.2 µM [236]. As a result, significant bioactivity improvements and further mechanistic studies of palmyramide A are needed.
Hoiamides A (162) and B (163) were two cyclic depsipeptides isolated from two distinct collections of marine cyanobacteria obtained from Papua New Guinea. Hoiamide A was a potent bioactive cyclic depsipeptide found in the assemblage of Lyngbya majuscula and Phormidium gracile [237], while hoiamide B was extracted from a sample of marine cyanobacteria collected in Gallows reef, the species of which was still unclear [238]. In a previous report, hoiamide A was shown to strongly inhibit voltage-gated sodium channels so that to activate the Na+ influx in mouse neocortical neurons [237]. In addition, Cao et al. found that in neocortical neurons, hoiamide A induced neuronal death through activating the c-Jun N-terminal kinase (JNK) and caspase signaling pathways [239]. In the aspect of their anticancer effects, both hoiamides A and B showed cytotoxicity against H460 cells with IC50 values of 11.2 and 8.3 µM, respectively [238]. Collectively, hoiamides A and B possess only moderate anticancer properties, which are still insufficient for further drug promotion.
Wewakazole (164) was a cyclic dodecapeptide derived from L. majuscula in Papua New Guinea [240]. A mass spectrometry-guided isolation afforded the isolation of wewakazole B (165), another novel cyclic peptide congener of wewakazole, from cyanobacterium Moorea peoducens. As cyanobactins, the most prominent bioactivity of the two compounds is cytotoxicity. When testing against H460 cells, wewakazole B (IC50 = 1.0 µM) was more cytotoxic than wewakazole (IC50 = 10 µM). Besides, wewakazole B also exhibited cytotoxicity against MCF7 cells with an IC50 value of 0.58 µM. Different from wewakazole, wewakazole B was inactive in siderophore assay [241]. The diverging bioactivities implicated that the iron-binding ability in their structures is crucial and deserves a detailed investigation. In recent years, both wewakazole and wewakazole B have been totally synthesized. With the availability of such synthetic strategy, steady and sufficient supply of these cyanobactins can be warranted for further biological evaluations [242,243].
Hectochlorin (166) was isolated from L. majuscula. It is a cyclic lipopeptide with moderate cytotoxicity against several cancer cell lines. Hectochlorin showed an average GI50 value of 5.1 µM in the NCI-60 screening assay; the colon, melanoma, ovarian and renal cell lines were the most sensitive to this compound [244]. In another bioassay, hectochlorin displayed cytotoxicity against KB and NCI-H187 (human small cell lung cancer) cells with ED50 values of 0.86 and 1.20 µM, respectively [245]. Further, this cyclic lipopeptide was able to induce actin polymerization and affect cell cycle regulation in those highly dividing cells. Apart from its cytotoxicity to cancer cells, hectochlorin was also found to be biologically active to yeasts and fungi [244]. Total synthesis and biosynthesis at the genetic level have been extensively studied in order to secure an abundant supply of this compound for further chemical and biological investigations [246,247].
Desmethoxymajusculamide C (DMMC) (167), a cyclic depsipeptide found in the process of a cytotoxicity-guided fractionation, was isolated from L. majuscula. The cyclic DMMC exhibited potent selectivity and cytotoxicity against the cancer cell lines HCT116, H460 and MDA-MB-435 with IC50 values of 0.02, 0.063 and 0.22 µM, respectively. Furthermore, a ring-opened form of DMMC was produced by means of base hydrolysis. Remarkably, the linear form of DMMC shared the same potency of cytotoxicity against the same panel of cancer cell lines (IC50 = 0.016, 0.094 and 0.23 µM, respectively). When acting against A10 cells, both cyclic and linear DMMCs disrupted the cellular microfilament networks and caused dramatic alterations in cell morphology, which were shown as the result of cellular apoptosis. Overall, these biological data indicated the potential anticancer efficacy of both cyclic and linear forms of DMMC [248]. The significant antitumor activities of DMMC are in need of further investigations, such as SAR and in vivo studies.
Hantupeptin A (168), another cyclodepsipeptide isolated from L. majuscula, was first discovered in Singapore. The compound displayed in vitro cytotoxicities against human lymphoblastic leukemia (MOLT-4) and MCF7 cells with IC50 values of 32 nM and 4.0 µM, respectively [249]. A subsequent bioassay-guided investigation of the same cyanobacterium led to purification of two other novel cyclic peptides, hantupeptins B (169) and C (170). Both compounds exhibited cytotoxicities when tested against MOLT-4 (IC50 = 0.2 and 3.0 µM, respectively) and MCF7 (IC50 = 0.5 and 1.0 µM, respectively) cells [250]. Furthermore, hantupeptins A-C also showed 100% brine shrimp toxicity when tested at 10 and 100 ppm [249,250].
Two cyclic peptides isolated from L. majuscula were designated as laxaphycins A (171) and B (172). Laxaphycin B showed significant cytotoxicities against the CCRF-CEM drug-sensitive human leukemic lymphoblasts (IC50 = 1.11 µM), as well as the vinblastine-resistant subline overexpressing Pgp CEM/VLB100 (IC50 = 1.02 µM) and CEM/VM-1 subline with DNA topoisomerase II alteration (IC50 = 1.37 µM). However, laxaphycin A was found to be inactive towards these cell lines at 20 µM [251]. Subsequently, Bonnard et al. demonstrated the anti-proliferative effect of laxaphycin B on several solid tumor cancer cell lines, including A549, MCF7, PA1 (human ovarian teratocarcinoma) and PC3, with IC50 values ranging from 0.19 to 1.0 µM. Furthermore, obvious biological synergism was observed on DLD1 (human colorectal adenocarcinoma) and MDR leukemic cells in the presence of laxaphycins A and B. Similar to the compounds mentioned previously, laxaphycins B2 (173) and B3 (174) were also isolated from the same assemblage of the cyanobacteria. These two novel cyclic peptides, displayed merely moderate cytotoxicities against the three leukemic cell lines above [252]. Laxaphycin B4 (175) was a new member of B-type laxaphycins, and it was discovered from the marine cyanobacterium Hormothamnion enteromorphoides. This compound yielded growth-inhibitory activity against HCT116 (IC50 = 1.7 µM); synergistic effect was obtained when it was used in combination with laxaphycin A [253].
Obyanamide (176) was a cytotoxic cyclic depsipeptide isolated in L. confervoides. From the MTT assay results, we noticed that this compound exhibited cytotoxicity against KB and LoVo cells with IC50 values of 0.97 and 5.24 µM, respectively [254]. To further promote biochemical utilization of obyanamide, several analogs were synthesized for biological evaluation, among which the cyclic analog 9b possessed the most potent activity. According to the SAR assessments, β-amino acid was an essential moiety for the bioactivities of obyanamide and its analogs [255]. Compared to the nature-derived obyanamide, its analogs are of greater significance for further biological studies.
Grassypeptolide A (177) was a macrocyclic depsipeptide with anticancer potential isolated from L. confervoides. Anti-proliferative activities of grasssypeptolide A were investigated in the U2OS (IC50 = 2.2 µM), HeLa (IC50 = 1.0 µM), HT29 (IC50 = 1.5 µM) and IMR-32 (IC50 = 4.2µM) cancer cell lines [256]. Subsequently, another study on the extract of the same cyanobacterium led to the purification of grassypeptolides A-C (177–179). The in vitro anti-proliferative effects of these compounds were evaluated in HT29 (IC50 = 1.22 µM, 2.97 µM and 76.7 nM, respectively) and HeLa (IC50 = 1.01 µM, 2.93 µM and 44.6 nM, respectively) cells. According to the cell viability assay results, grassypeptolide C was 16-23 times more potent than grassypeptolide A. Moreover, both grasspeptolides A and C could induce cell cycle arrest at the G1 phase at relatively low concentrations (10 µM and 316 nM, respectively); they could also induce the G2/M phase arrest at higher concentrations (31.6 µM and 1 µM, respectively). These two compounds have the capability to bind to metal ions such as Cu2+ and Zn2+, which may denote a partial mechanism of their cytotoxicities [257]. The study Liu et al. revealed that grassypeptolide A increased the cleavage of PARP and decreased the levels of bcl-2 and bcl-xL. As for the induction of cell cycle arrest, it downregulated cyclin D, but upregulated P27 and P21 [258]. Grassypeptolides D (180) and E (181), two novel members of the family, were isolated from the marine cyanobacterium Leptolyngbya sp. These two compounds were cytotoxic to HeLa cells (IC50 = 335 and 192 nM, respectively) and mouse neuro-2a blastoma cells (IC50 = 599 and 407 nM, respectively) [259]. To promote these grassypeptolides for further drug development, total synthesis and mechanistic studies are desperately needed.
Palauamide (182), a cyclic depsipeptide extracted from a bioassay-guided fractionation, was confirmed as a potent cytotoxin isolated from Lyngbya sp. in Palau. When cultured with KB cells, palauamide yielded significant cytotoxicity with an IC50 value of 13 nM [260]. Palauamide was totally synthesized, along with its diastereomers and other analogs, which also showed potent cytotoxicities [261,262]. However, more analogs are in need to be synthesized for further elucidation of their biological potential and SAR studies.
Coibamide A (183) was a cyclic depsipeptide that was first isolated from the marine filamentous cyanobacterium Leptolyngbya sp. in Coiba National Park, Panama. The preliminary biological activity screening showed that this compound displayed potent cytotoxicity against NCI-H460 and mouse neuro-2a cells with LC50 < 17.9 nM. The evaluation against the NCI’s in vitro 60 cancer cell line panel highlighted its remarkable anti-proliferative response. From the flow cytometric analysis, coibamide A was shown to induce cell cycle arrest at the G1 phase [263]. Subsequently, Hau et al. [264] demonstrated that coibamide A induced mammalian target of rapamycin (mTOR)-independent autophagy and cell death in human glioblastoma cells. In the glioblastoma xenograft models, the expression levels of vascular epithelial growth factor A/vascular epithelial growth factor receptor 2 (VEGFA/VEGFR2) were markedly suppressed upon the treatment with coibamide A, so that the size of tumor xenografts was largely reduced from 2000 mm3 to 200-300 mm3. However, different degrees of weight loss were observed in the animal experiment. Such result implicated that targeted delivery and toxicity profile should be improved for drug development [265].
Ulongamides A-E (184–188) were five β-amino acid-containing cyclopeptides derived from the marine cyanobacterium Lyngbya sp. in Palau. These ulongamides exhibited moderate in vitro cytotoxicity against KB and LoVo cells with IC50 values around 1 and 5 µM [266]. The total synthesis of ulongamide A as well as its two congeners (ulongmides B and C) had been previously accomplished [267,268]. In the long run, structural optimization is needed to improve the bioactivity potency of this type of compounds.
Ulongapeptin (189), another cytotoxic cyclopeptide, was isolated from Lyngbya sp. in Palau. This compound yielded cytotoxicity against KB cells with an IC50 value of 0.63 µM [269]. No further synthetic or biological approaches of this compound have been reported thereafter.
The marine cyanobacterium Okeania sp. is the origin of the cytotoxic cyclodepsipeptide odoamide (190). As another new member of aurilide class, odoamide has been reported with a significant cytotoxicity against HeLa S3 cells (IC50 = 26.3 nM) and a moderate toxicity to brine shrimp [270]. From the SAR assessment of synthetic odoamide isomers, the in vitro cytotoxicity was mostly affected by the serum protein binding of odoamide derivatives. Nevertheless, the macrocyclic structure of odoamide and derivatives is not crucial to the alteration of membrane permeability [271].
Cryptophycin 1 (191), a cyclic depsipeptide isolated from cyanobacterium Nostoc sp., was first discovered as an antifungal agent two decades ago. Subsequent studies indicated that cryptophycin 1 was also an anti-proliferative agent with significant antitumor activities [272,273]. A great number of experiments showed that cryptophycin 1 yielded inhibitory activities against various cancer cell lines such as L1210 (IC50 = 4 pM), SKOV3 (IC50 = 7 pM) and MCF7 (IC50 = 16 pM). Some recent reports demonstrated that cryptophycin 1 provided an anticancer potency ~100-fold higher than the mainstay anticancer drug paclitaxel; therefore, a great interest was immediately prompted to investigate the mechanism of action of this compound [274,275]. According to the study of Kerksiek et al., cryptophycin 1 was an effective inhibitor of tubulin polymerization, by which microtubules were depolymerized into linear polymers [276]. When exposed to stoichiometric amounts of proteins, cryptophycin 1 elicited extensive aggregates of tubulin [277]. Even at picomolar concentrations, cryptophycin 1 was found to cause cell cycle arrest at the G2/M phase via its actions on the microtubules [275]. Though inhibitory effect on macromolecular synthesis of cryptophycin 1 had been observed in cancer cells, the slight downregulation of DNA, RNA and proteins was not well correlated to its anticancer potency [278]. Up to now, several analogs of cryptophycin 1, such as cryptophycin 52, have been synthesized in order to achieve higher anticancer potency [279].
Largazole (192) is a cyclic depsipeptide isolated from the cyanobacterium of Symploca sp. This compound was found to have good selectivity against different types of cancer cells. For instance, largazole was less cytotoxicy to the nontransformed murine mammary epithelial MDA-MB-231 cells (GI50 = 122 nM, LC50 = 272 nM), but it was more susceptible to the invasive transformed human mammary epithelial (NmuMG) cells (GI50 = 7.7 nM, LC50 = 117 nM). Similarly, this cyclopeptide displayed higher inhibitory effect against U2OS transformed fibroblastic osteosarcoma (GI50 = 55 nM, LC50 = 94 nM) than NIH3T3 nontransformed fibroblasts (GI50 = 480 nM, LC50 > 8 µM) [280]. Based on the pharmacological studies carried out by Bowers et al., largazole was considered a potent inhibitor of class I HDACs [281]. In the subsequent SAR assessments of largazole and its derivatives, largazole was shown to exhibit good selectivity towards cancer cell lines HCT116 and A549 with GI50 values of 0.08 µM and 0.32 µM, respectively [282]. The in vitro cytotoxicity of largazole in the HCT116 cells was plausibly derived from its induction of cell cycle arrest and apoptosis. When tested in other CRC cells (e.g., HT29 and HCT15), this compound was found to induce histone H3 hyperacetylation. The further in vivo studies indicated that the anticancer effect of largazole was highly associated with an attenuation of the protein kinase B (AKT) pathway and a modulation of cell cycle regulators [283]. It is widely accepted that ubiquitination of protein is important for the vast majority of eukaryotic cellular functions; regular protein ubiquitination is halted in many cancers. Liu and his co-workers reported that largazole was able to inhibit the ubiquitin activating enzyme E1 to disturb ubiquitin conjugation with p27Kip1 and telomeric repeat-binding factor 1 (TRF1) in vitro [284]. In addition, largazole was demonstrated to induce the expression of osteoblast differentiation markers, such as ALP. Taken together, largazole is suggested to activate osteogenesis [285]. While showing several intriguing and multidisciplinary functions, largazole is undoubtedly a potential therapeutic agent for further development.
Studies on a cyanobacterium of Symploca sp. From Papua New Guinea led to the isolation of a novel cyclopeptide—symplocamide A (193). This compound showed potent cytotoxicity against H460 and neuro-2a cells with IC50 values of 40 nM and 29 nM, respectively. In fact, symplocamide A shares a structure very similar to protease inhibitors; thus, it can be served as a potent chymotrypsin inhibitor (IC50 = 0.38 µM). The role of protease inhibition in cancer therapy is worth to be studied. On the other hand, symplocamide A was reported to show moderate anti-parasite activities against malaria, Chagas disease and leishmaniasis [286]. Solid phase total synthesis of symplocamide A had been accomplished by Kaiser and his co-workers, which has laid a groundwork for further studies of this type of compounds [287].
Two other cyclic depsipeptides had been isolated from Symploca sp. in Palau, and they were named tasipeptins A (194) and B (195). These two compounds exhibited cytotoxicities against KB cells with IC50 values of 0.93 and 0.82 µM, respectively [288]. Due to the presence of Ahp, a symbolic structural moiety of cyanobacterial metabolites, tasipeptins A and B were considered as potential protease inhibitors. The total synthesis of these two compounds had been completed by Kaiser and his co-workers [289]. It is believed that ever-growing investigations on Ahp-containing cyclic depsipeptides can make good contributions to the further rational design of novel anticancer lead compounds.
Cytotoxicity-directed fractionation of cyanobacterium Symploca cf. hydnoides from Guam led to the isolation of a series of cyclic depsipeptides, veraguamides A-G (196–202). The results of several cell viability assays showed that veraguamides A-G displayed weak to moderate cytotoxicities against HT29 and HeLa cells with IC50 values varying from 0.84 to 49 µM and 0.54 to 49 µM, respectively. From the SAR assessments, the seven semi-synthetic analogs of veraguamide G explicitly denoted the sensitive positions of veraguamides that were responsible for their cytotoxic activities [290].
Floridamide (203), a novel cyclic depsipeptide containing four amino acid units, was discovered in a bioassay-guided investigation of Moorea producens, a species of filamentous cyanobacteria collected from Florida, USA. This compound displayed cytotoxic activities against H460 and mouse neuro-2a neuroblastoma cells with the EC50 values of around 1.89 × 10−5 µM [291].
Viequeamide A (204) was a highly toxic cyclic depsipeptide found in the cyanobacterium Rivularia sp. near the island of Vieques. It was found to be strongly cytotoxic against H460 cells with an IC50 value of 60 nM. As a member of the “kulolide superfamily”, the biological properties of viequeamied A were reported to be highly divergent [292]. The total synthesis of this compound had been already achieved, so that the steady and sufficient supply should facilitate further intriguing studies on SAR assessment and mechanism of actions of this type of compounds [293].
5. Fungi-Derived Anticancer Cyclopeptides
Fungi are eukaryotic organisms with worldwide distribution and different habitats. It is estimated that biodiversity of fungi is between 2.2 to 3.8 million species around the world [294]. Moreover, fungi belong to the most prolific producers of secondary metabolites [295]. Due to their roles acting as food, medicines and pathogens, as well as their biodiversity, fungi can be abundant sources of natural therapeutic agents. Many recently isolated cyclopeptides from fungi exhibited potent anticancer activities and they will be discussed in this section. The structures of these compounds were drawn in Figure 9 and their activities were presented in Table 4.
Figure 9.

Structure of cyclopeptides (205–219) derived from non-marine fungi.
Table 4.
Fungi-derived anticancer cyclopeptides.
| Name | Biological Source | Anticancer Activity | Reference |
|---|---|---|---|
| Anticancer Cyclopeptides Derived from Non-Marine Fungi | |||
| 1-alaninechlamydocin (205) | Tolypocladium sp. | Cytotoxicity against MIA PaCa-2 (GI50 = 5.3 nM, LC50 = 22 nM) cell. | [296] |
| Pseudoxylallemycin C (206) | Pseudoxylaria sp. X802 | Anti-proliferative effect to K562 (GI50 = 6.8 µM) cell. Cytotoxicity against HeLa (CC50 = 16.68 µM) cell. | [297] |
| Cycloaspeptides F (207) and G (208) | Isaria farinosa | Inhibitory activities toward MCF7 (GI50 = 18.7 and 15.2 µM, respectively) cell. | [298] |
| Malformin A1 (MA1) (209) | Aspergillus niger | Cytotoxicity against PC3 (IC50 = 130 nM), LNCaP (IC50 = 90 nM), HeLa (IC50 = 9.48 nM) and P388 (IC50 = 132.9 nM) cells. | [299,300,301] |
| Malformin E (210) | Aspergillus tamarii | Cytotoxicity against MCF7 (IC50 = 0.65 µM) and A549 (IC50 = 2.42 µM) cells. | [302] |
| Neo-N-methylsansalvamide (211) | Fusarium solani KCCM90040 | Cytotoxicity against A549, SK-OV-3, SK-MEL-2 and MES-SA cells. (EC50 = 10.0 to 14.74 µM) Synergistic effect of neo-N-methylsansalvamide and paclitaxel when acting against MES-SA, HCT15, MES-SA/DX5 and HCT15/CL05 cells. |
[303,304] |
| Cylindrocyclin A (212) | Cyclindrocarpon sp. strain A101-96 | Cytotoxicity against COLO-320, HL-60, L1210 and human T lymphocyte Jurkat cells. (IC50~11 µM) | [305] |
| Gymnopeptides A (213) and B (214) | Mushroom, Gymnopus fusipes | Anti-proliferative activities against HeLa, A431, T47D, MCF7 and MDA-MB-231 cells. (IC50 = 18.0 to 88.4 nM and 14.0 to 44.3 nM, respectively) | [306,307] |
| Phomopsin F (215) | Diaporthe toxica | Impact on viability and vitability of HepG2 (IC50 = 6.1 µM) cell. | [308] |
| Pteratides I-IV (216–219) | Pterula sp. | Cytotoxicity against P388 (IC50 = 41 nM, 40 nM, 7.4 µM and 2.9 µM, respectively) cell. | [309] |
| Anticancer Cyclopeptides Derived from Marine Fungi | |||
| Asperterrestide A (220) | Aspergillus terreus SCSGAF0162 | Cytotoxicity against U937 (IC50 = 6.4 μM) and MOL-4 (IC50 = 6.2 μM) cells. | [310,311] |
| Cordyheptapeptide A (221) | Cordyceps sp. BCC 1788 | Anti-proliferative activities to KB (IC50 = 0.78 μM), BC (IC50 = 0.2 μM), NCI-H187 (IC50 = 0.18 μM) and Vero (IC50 = 14 μM) cells. | [312,313] |
| Cordyheptapeptide B (222) | Cordyceps sp. BCC 16173 | Anti-proliferative activities to KB (IC50 = 2.0 μM), BC (IC50 = 0.66 μM), NCI-H187 (IC50 = 3.1 μM) and Vero (IC50 = 1.6 μM) cells. | [312,313] |
| Cordyheptapeptide C (223) | Acremonium persicinum | Cytotoxicity against SF-268 (IC50 = 3.7 μM) and MCF7 (IC50 = 3.0 μM) cells. | [314] |
| Cordyheptapeptide E (224) | Acremonium persicinum | Cytotoxicity against SF-268 (IC50 = 3.2 μM), MCF7 (IC50 = 2.7 μM) and NCI-H460 (IC50 = 4.5 μM) cells. | [314] |
| Scytalidamides A (225) and B (226) | Scytalidium sp. | Cytotoxicity against HCT116 (IC50 = 2.7 and 11.0 µM, respectively) cell. Moderate cytotoxicitty in NCI 60 cell line panel. (Mean GI50 = 7.9 and 4.1 µM, respectively) |
[315] |
| Zygosporamide (227) | Zygosporium masonii | Cytotoxicity against SF-268 (IC50 = 6.5 nM) and RXF393 (IC50 ≤ 5.0 nM) cells. | [316,317] |
| Sansalvamide A (228) | Fusarium sp. | Cytotoxicity against HCT116, COLO 205 and SK-MEL-2 (IC50 = 9.7, 3.46 and 5.84 μM, respectively) cells. | [318,319,320,321,322,323] |
| Clavatustide A (229) | Aspergillus clavatus | Anti-proliferative activity against Hep G2, SMMC-7721 and Bel-7402 cells. (IC50~32 μM) | [324] |
| Clavatustide B (230) | Aspergillus clavatus | Anti-proliferative activity against Hep G2, SMMC-7721 and Bel-7402 cells. (IC50~32 μM) Cell growth inhibitory activity against PANC-1 and PC3 cells. |
[324,325] |
| Sclerotiotide M (231) | Aspergillus ochraceopetaliformis DSW-2 | Cytotoxicity against HPAC and BXPC3 cells. (IC50 > 20 μM) | [326] |
| Scopularides A (232) and B (233) | Scopulariopsis brevicaulis | Viability inhibition of several cancer cell lines including Colo357, Panc89 and HT29 cells at a concentration of 14.87 μM. | [327] |
| IB-01212 (234) | Clonostachys sp. ESNA-A009 | Inhibitory activity against several cancer cell lines including LNCaP, SKBR3, HT29 and HeLa with the GI50 values on the order of 10−8 M. | [328,329,330,331] |
| Microsporins A (235) and B (236) | Microsporum cf. gypseum | Cytotoxicity especially against HCT116 cell. (IC50 = 1.12 and 15.82 μM, respectively) | [332,333,334] |
5.1. Anticancer Cyclopeptides Derived from Non-Marine Fungi
1-Alaninechlamydocin (205) was a cyclic tetrapeptide isolated from the fungi of Tolypocladium sp. obtained from the Great Lakes. The compound displayed predominant anti-proliferative and cytotoxic activities toward MIA PaCa-2 cells with GI50 and LC50 values of 5.3 and 22 nM, respectively. Moreover, as a potent HDAC inhibitor, 1-alaninechlamydocin induced apoptosis and cell cycle arrest at phase G2/M as well. HDAC inhibitory activity also primarily contributed to in vitro bioactivities of the compound [296].
Pseudoxylaria sp. X802, a termite-associated fungus, yielded six new cyclic tetrapeptides, among which pseudoxylallemycin C (206) showed moderate cytotoxicity. The compound exhibited anti-proliferative effect to K562 cells with the GI50 value of 6.8 µM and cytotoxicity to HeLa cells with CC50 value of 16.68 µM. However, human umbilical vein endothelial cells (HUVEC) was also sensitive to pseudoxylallemycin C (GI50 = 6.96 µM), which demonstrated low specificity of the compound [297].
Cycloaspeptides F (207) and G (208) were two new members of cycloaspeptides—the ABA-containing cyclic pentapeptides discovered in different fungi. These two compounds were derived from the fungus Isaria farinosa, an inhabitant of Cordyceps sinensis obtained from Linzhi, Tibet. They showed inhibitory activities toward MCF7 cells with respective GI50 values of 18.7 and 15.2 µM respectively, which were comparable to the positive control 5FU (GI50 = 15 µM) [298].
Malformin A1 (MA1) (209), a cyclopentapeptide with versatile biological activities, was originally isolated from the terrestrial plant fungus Aspergillus niger. Liu et al. investigated its anticancer mechanism of action in human prostate cancer PC3 (IC50 = 130 nM) aand LNCaP (IC50 = 90 nM) cell lines. Coexistence of apoptosis, necrosis and autophagy were all induced by mitochondrial damage, which illustrated the mechanism of MA1-mediated cell death in prostate cancer cells. In addition, stimulation of the AMP-activated serine/threonine protein kinase/mammalian target-of-rapamycin (AMPK/mTOR) pathway by adding oxidative stress or decreasing ATP could lead to MA1-mediated autophagy [299]. In human colorectal cancer cells (SW480 and DKO1), invasive and oncogenic phenotypes were modified through p38 signaling pathway [300]. Recently, the marine-derived fungus A. tubingenesis also yielded MA1 with significant cytotoxicity against HeLa (IC50 = 9.48 nM) and P388 (IC50 = 132.9 nM) cell lines [301]. Malformin A1 is an early-discovered cyclic peptide with diverse biological properties and sufficient studies have demonstrated its bioactivities as well as mechanism of action. However, for further promotion to clinical stage, more supporting data in animal models shall be carried out. Another new member of the malformin class, designated as malformin E (210), was isolated from the endophytic fungus A. tamarii collected from the roots of Ficus carica. The compound also showed in vitro cytotoxic effects on MCF7 and A549 cells, with IC50 values of 0.65 and 2.42 µM, respectively. Moreover, malformin E also exhibited antimicrobial activities against several strains of bacteria and fungi [302].
Another new analog of sansalvamide A (228), neo-N-methylsansalvamide (211), was isolated from the fungus Fusarium solani KCCM90040 derived from the potato in Korea. The new cyclic pentadepsipeptide was able to inhibit in vitro cell growth of A549, SK-OV-3, SK-MEL-2 (human melanoma) and MES-SA (human uterine sarcoma) cells with EC50 values ranging from 10.0 to 14.74 µM [303]. Moreover, synergistic effects of neo-N-methylsansalvamide and paclitaxel were also observed when acting against MES-SA, HCT15 (colorectal adenocarcinoma) and two multidrug resistance sublines (MES-SA/DX5 (human uterine corpus sarcoma) and HCT15/CL05). The EC50 values of paclitaxel added with 3 µM neo-N-methylsansalvamide were significantly decreased by several orders of magnitude [304]. As a result, although the cytotoxic effect of the cyclic peptide alone was not so dominant, significant potency in reversing in vitro multidrug resistance can be promoted for further development as a promising agent with clinical function.
Cylindrocyclin A (212), a cyclic nonapeptide, was isolated from Cyclindrocarpon sp. strain A101-96 during the period of a screening of new metabolites with cytotoxic effects from fungal extracts. Six cancer cell lines were assayed for its anticancer potential, while the compound only showed moderate cytotoxicity to colorectal adenocarcinoma (COLO-320), human leukemia (HL-60), L1210 and human T lymphocyte Jurkat cell lines with IC50 value around 11 µM. Neither antimicrobial or nematicidal effect was observed for cylindrocyclin A [305].
Studies on the mushroom Gymnopus fusipes led to the isolation of two novel octadecapeptides, gymnopeptides A (213) and B (214). Both compounds are octadecacyclopeptides with natural cyclic β hairpins. Gymnopeptides A and B displayed significant anti-proliferative activities against HeLa, A431 (human epidermoid carcinoma), T47D, MCF7 and MDA-MB-231 cells with IC50 values ranging from 18.0 to 88.4 nM and 14.0 to 44.3 nM, respectively [306]. Due to their unprecedented structures and dominant growth inhibitory activity on cancer cells, to make sufficient supply for further research, total synthesis of the two compounds were successfully accomplished subsequently [307]. As a result, it is fully justified to further explore their bioactive roles and mechanism of actions.
Phomopsin F (215), a cyclic hexapeptide derived from the fungus Diaporthe toxica, was also an N-methylated derivative of the mycotoxin phomopsin A. The initial in vitro screening of cytotoxicity revealed its impact on viability and vitability of HepG2 cells (IC50 = 6.1 µM) [308].
Four cytotoxic cyclodepsipeptides, pteratides I-IV (216–219), were purified from a basidiomycete identified as Pterula species. Pterides I and II showed remarkable cytotoxicity against P388 cell line with IC50 values of 41 and 40 nM, respectively. Nevertheless, compared to the first two compounds, pteratides III and IV were much less active (IC50 = 7.4 and 2.9 µM, respectively) due to their much different structural discrepancy to pteratides I and II [309]. Practically, these compounds can be good candidates as anticancer agents, but their mechanism of actions as well as the underlying SAR remains to be further investigated.
5.2. Anticancer Cyclopeptides Derived from Marine Fungi
A cyclic tetrapeptide called asperterrestide A (220), with entire composition of non-protein amino acids, was first isolated from the marine-derived fungus Aspergillus terreus SCSGAF0162 (Figure 10). The compound exhibited cytotoxicity against U937 and MOL-4 (acute lymphoblastic leukemia) cell lines with IC50 values of 6.4 and 6.2 μM, respectively. Moreover, the growth of the influenza virus strains H1N1 and H3N2 could also be inhibited by asperterrestide A [310]. Just like other cyclic tetrapeptide natural products, synthesis of the compound remains challenging [311].
Figure 10.

Structure of cyclopeptides (220–236) derived from marine fungi.
Cordyheptapeptides A (221) and B (222) were two bioactive cycloheptapeptides previously isolated from two different insect pathogenic fungi Cordyceps sp. BCC 1788 and 16173, respectively. The two cyclic peptides displayed anti-proliferative activities against KB, BC (human lymphoma), NCI-H187 and Vero (normal kidney) cells with IC50 values of cordyheptapeptide A measured as 0.78, 0.2, 0.18 and 14 µM, respectively, the IC50 values of cordyheptapeptide B as 2.0, 0.66, 3.1 and 1.6 µM, respectively. In addition, cordyheptapeptide A also exhibited moderate antimalarial activities [312]. Total synthesis of the two compounds, according to subsequent reports, had been accomplished together with other pharmacological investigations. Intriguingly, both synthetic compounds were of cytotoxic potential acting against Dalton’s lymphoma ascites (DLA) (CTC50 = 10.6 and 7.4 µM, respectively) and Ehrlich’s ascites carcinoma (EAC) (CTC50 = 14.6 and 13.56 µM, respectively) cell lines [312,313]. Accordingly, further studies could add more insights into their mechanisms of action. Chen et al. discovered two other novel congeners, Cordyheptapeptides C (223) and E (224), from a marine-derived fungus Acremonium persicinum in 2012. Cordyheptapeptide C displayed cytotoxicity against SF-268 and MCF7 with IC50 values of 3.7 and 3.0 μM, respectively. And the other one showed similar cytotoxicity against SF-268, MCF7 and NCI-H460 cell lines (IC50 = 3.2, 2.7 and 4.5 μM, respectively) [314]. However, no subsequent pharmacological study and in vivo experiments associated with the two compounds have been reported.
Scytalidamides A (225) and B (226) were two cyclic heptapeptides extracted from the culture broth of the marine fungus Scytalidium sp. from the Bahamas. When incubated with HCT116 cell line, the two compounds displayed in vitro cytotoxicity with IC50 values of 2.7 and 11.0 µM, respectively. Tested in the NCI 60 cell line panel, scytalidamides A and B exhibited moderate cytotoxicity with mean GI50 values of 7.9 and 4.1 µM, respectively. The screening test also demonstrated the most sensitive cell lines were MOLT-4 (IC50 = 3.0 µM) for scytalidamide A and UACC-257 (IC50 = 3.0 µM) for scytalidamide B [315].
Zygosporamide (227), a cyclic pentadepsipeptide produced by the marine-derived fungus Zygosporium masonii, was shown to possess potent anticancer properties. Evaluated in the NCI cell line assays (median GI50 = 9.1 µM), the compound yielded significant in vitro cytotoxicity against central nervous system (CNS) cancer cell line SF-268 (IC50 = 6.5 nM) and renal cancer cell line RXF393 (IC50 ≤ 5.0 nM) with high selectivity [316]. The total synthesis of zygosporamide was accomplished. However, there was a notable (about 700-fold) discrepancy of the IC50 values between the natural and synthetic cyclic peptides when acting against SF-268 [317].
Sansalvamide A (228), a natural cyclic depsipeptide, was isolated first from a marine fungus of Fusarium sp. living on the seagrass Halodule wrightii. The compound showed cytotoxicity against several cancer cells lines, including HCT116, COLO 205 and SK-MEL-2 cells with IC50 values of 9.7, 3.46 and 5.84 μM, respectively [318]. Sansalvamide A caused in vitro pancreatic cancer cell death by upregulation of p21 and downregulation of cyclinsD1, E, A and CDK4, which was consistent with G0/G1 cell cycle arrest [319]. Additionally, it was also identified as an inhibitor of a topoisomerase in the pathogenic poxvirus molluscum contagiosum virus (MCV) [320]. However, whether such inhibitory effect was partly related to its anticancer effect is still unclear [319]. The total synthesis of sansalvamide A had also been achieved, which laid substantial groundwork for subsequent SAR study of its derivatives [321]. Several derivatives of sansalvamide A were reported to possess much higher efficacy and selectivity against different cancer cells than the natural product [321,322,323]. Accordingly, sansalvamide A is considered as a lead compound to synthesise its congeners to improve its anticancer potential.
Clavatustides A (229) and B (230) were two novel cyclodepsipeptides derived from the fungus Aspergillus clavatus. Xenograpsus testudinatus, a kind of hydrothermal vent crab, is the habitat where they found the fungus. Jiang et al. determined that both compounds suppressed the proliferation of HCC (hepatocellular carcinoma) cell lines (Hep G2, SMMC-7721 and Bel-7402) in a dose-dependent as well as time-dependent manner, with IC50 values around 32 μM. Compared with human normal hepatocyte, HCC cell lines, especially HepG2 cells, were more sensitive in clavatustides-induced cell proliferation suppression. Their proliferation inhibitory property was due to suppression of G1/S phase transition [324]. Clavatustide B also exhibited cell growth inhibitory activity against PANC-1 and PC3, the chemo- and radio-therapy resistant cell lines. Cyclin E2 was found to be involved in clavatustide B-induced cell cycle arrest. With special origin and unique structure, these two compounds warrant further investigation to explore their underlying mechanisms and other potential bioactivities. Synthesis of clavatustides A and B analogs is also a meaningful approach to propel their future clinical application [325].
A novel cyclic tripeptide, sclerotiotide M (231) was isolated from the fungus Aspergillus ochraceopetaliformis DSW-2 and the compound only showed weak cytotoxic activities against human pancreatic cancer HPAC and BXPC3 cell lines with IC50 values over 20 μM [326].
Two cyclodepsipeptides, scopularides A (232) and B (233), were isolated from the fungus Scopulariopsis brevicaulis, which was found in the marine sponge Tethya aurantium. Both compounds significantly inhibited viability of several cancer cell lines including Colo357 (pancreatic adenosquamous carcinoma), Panc89 (pancreatic tumor) and HT29 cells at a concentration of 14.87 μM [327]. However, regorous and standard cytotoxic experimental procedures are needed to determine their anticancer potential.
The cytotoxic cyclodepsipeptide IB-01212 (234) was first isolated from the marine fungus Clonostachys sp. ESNA-A009. It was highly active against several cancer cell lines including LNCaP, SKBR3, HT29 and HeLa with the GI50 values on the order of 10−8 M [328]. In addition, IB-01212 also showed significant leishmanicidal activity through an apoptotic-like process, which added fresh insights to develop this compound as potential novel chemotherapeutic agent against Leishmania [329]. The synthetic and SAR studies of IB-01212 demonstrated great cytotoxic significance of its symmetric structure. The N-methylation on the amino acids of IB-01212 were found to enhance its cytotoxic effect, which were corroborating evidence for further discovery and synthesis of new analogs for anticancer drug development [330,331].
Microsporins A (235) and B (236) were cyclopeptides isolated from the marine fungus Microsporum cf. gypseum in bryozoan Bugula sp. from Virgin Islands in the US. Both were shown to be potent HDAC inhibitors and demonstrated cytotoxicity especially against HCT116 cell line, with IC50 values of 1.12 and 15.82 μM, respectively [332]. To extensively explore their potential bioactive properties, efforts towards the synthesis of microsporins and their isomers have been reported [333,334].
6. Conclusions and Future Perspectives
This review included the recently discovered natural anticancer cyclopeptides, which have been summarized and categorized according to their origins. An abundant number of cyclopeptides with diverging chemical and biological characteristics can be obtained from the aqueous and terrestrial portions of our nature. Cyclopeptides derived from natural sources have drawn great attention due to their high therapeutic potential for drug development. Today, many studies have shown that natural cyclopeptides are effective in the treatment and preventive measures of cancerous pathologies. Other biological activities such as antiviral, antibacterial and antimalarial effects have also been extensively observed whilst some are considered promising. Hence, detailed mechanism studies and structural modification of cyclopeptides are desperately needed for the development of cyclopeptide as therapeutic drug candidates. Nonetheless, the current research studies on the anticancer effects of natural cyclopeptides are just the tip of the iceberg. The natural cyclopeptides are undoubtedly a rich source as bioactive compound leads gifted from nature to benefit human welfare.
Author Contributions
All authors contributed equally to this work. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Research Grants Council of the Hong Kong Special Administrative Region, China (Project No. HKBU 12142016).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Torre L.A., Bray F., Siegel R.L., Ferlay J., Lortet-tieulent J., Jemal A. Global Cancer Statistics, 2012, CA a Cancer. J. Clin. 2015;65:87–108. doi: 10.3322/caac.21262. [DOI] [PubMed] [Google Scholar]
- 2.Bray F., Ferlay J., Soerjomataram I., Siegel R.L., Torre L.A., Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018;68:394–424. doi: 10.3322/caac.21492. [DOI] [PubMed] [Google Scholar]
- 3.American Cancer Society Global Cancer Facts & Figures 4th Edition. Atlanta: American Cancer Society. [(accessed on 12 April 2021)];2018 Available online: https://www.cancer.org/content/dam/cancer-org/research/cancer-facts-and-statistics/global-cancer-facts-and-figures/global-cancer-facts-and-figures-4th-edition.pdf.
- 4.Appold K. Cancer Treatment—Top 4 Developments to Watch. [(accessed on 12 April 2021)];2017 Available online: https://www.managedhealthcareexecutive.com/view/cancer-treatment-developments.
- 5.Kang T.H., Mao C.P., He L., Tsai Y.C., Liu K., La V., Wu T.C., Hung C.F. Tumor-targeted delivery of IL-2 by NKG2D leads to accumulation of antigen-specific CD8+ T cells in the tumor loci and enhanced anti-tumor effects. PLoS ONE. 2012;7:e35141. doi: 10.1371/journal.pone.0035141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Amit D., Tamir S., Hochberg A. Development of targeted therapy for a broad spectrum of solid tumors mediated by a double promoter plasmid expressing diphtheria toxin under the control of IGF2-P4 and IGF2-P3 regulatory sequences. Int. J. Clin. Exp. Med. 2013;6:110–118. doi: 10.1186/1479-5876-8-134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Peer D., Karp J.M., Hong S., Farokhzad O.C., Margalit R., Langer R. Nanocarriers as an emerging platform for cancer therapy. Nat. NanoTechnol. 2007;2:751–760. doi: 10.1038/nnano.2007.387. [DOI] [PubMed] [Google Scholar]
- 8.Pawar S.V., Ho J.C.H., Yadav G.D., Yadav V.G. The impending renaissance in discovery & development of natural products. Curr. Top. Med. Chem. 2017:251–267. doi: 10.2174/15680266166661605301. [DOI] [PubMed] [Google Scholar]
- 9.Butler M.S. The role of natural product chemistry in drug discovery. J. Nat. Prod. 2004;67:2141–2153. doi: 10.1021/np040106y. [DOI] [PubMed] [Google Scholar]
- 10.Joo S.H. Cyclic peptides as therapeutic agents and biochemical tools. Biomol. Ther. 2012;20:19–26. doi: 10.4062/biomolther.2012.20.1.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wipf P. Synthetic studies of biologically active marine cyclopeptides. Chem. Rev. 1995;95:2115–2134. doi: 10.1021/cr00038a013. [DOI] [Google Scholar]
- 12.Pomilio A., Battista M., Vitale A. Naturally-occurring cyclopeptides: Structures and bioactivity. Curr. Org. Chem. 2006;10:2075–2121. doi: 10.2174/138527206778742669. [DOI] [Google Scholar]
- 13.Tran D., Selsted M.E., Dorrestein P.C., Pavel A. Cycloquest: Identification of cyclopeptides via database search of their mass spectra against genome databases. NIH Public Access. 2012;10:4505–4512. doi: 10.1021/pr200323a.Cycloquest. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Tan N.H., Zhou J. Plant cyclopeptides. Chem. Rev. 2006;106:840–895. doi: 10.1021/cr040699h. [DOI] [PubMed] [Google Scholar]
- 15.Marahiel M.A., Stachelhaus T., Mootz H.D. Modular peptide synthetases involved in nonribosomal peptide synthesis. Chem. Rev. 1997;97:2651–2674. doi: 10.1021/cr960029e. [DOI] [PubMed] [Google Scholar]
- 16.Hur G.H., Vickery C.R., Burkart M.D. Explorations of catalytic domains in non-ribosomal peptide synthetase enzymology. Nat. Prod. Rep. 2012;29:1074–1098. doi: 10.1039/c2np20025b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Salomon R.A., Farias R.N. Microcin 25, a novel antimicrobial peptide produced by Escherichia coli. J. Bacteriol. 1992 doi: 10.1128/JB.174.22.7428-7435.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Luckett S., Garcia R.S., Barker J.J., Konarev A.V., Shewry P.R., Clarke A.R., Brady R.L. High-resolution structure of a potent, cyclic proteinase inhibitor from sunflower seeds. J. Mol. Biol. 1999;290:525–533. doi: 10.1006/jmbi.1999.2891. [DOI] [PubMed] [Google Scholar]
- 19.Tang Y.Q., Yuan J., Ösapay G., Ösapay K., Tran D., Miller C.J., Ouellette A.J., Selsted M.E. A cyclic antimicrobial peptide produced in primate leukocytes by the ligation of two truncated α-defensins. Science. 1999;286:498–502. doi: 10.1126/science.286.5439.498. [DOI] [PubMed] [Google Scholar]
- 20.Closse A., Huguenin R. Isolierung und Strukturaufklärung von Chlamydocin. Helv. Chim. Acta. 1974;57:533–545. doi: 10.1002/hlca.19740570306. [DOI] [PubMed] [Google Scholar]
- 21.Mollica A., Costante R., Stefanucci A., Novellino E. Cyclotides: A natural combinatorial peptide library or a bioactive sequence player? J. Enzym. Inhib. Med. Chem. 2015;30:575–580. doi: 10.3109/14756366.2014.954108. [DOI] [PubMed] [Google Scholar]
- 22.Edman P. Chemistry of amino acids and peptides. Annu. Rev. Biochem. 1959;28:69–96. doi: 10.1146/annurev.bi.28.070159.000441. [DOI] [PubMed] [Google Scholar]
- 23.Trabi M., Craik D.J. Circular proteins-No end in sight. Trends Biochem. Sci. 2002;27:132–138. doi: 10.1016/S0968-0004(02)02057-1. [DOI] [PubMed] [Google Scholar]
- 24.Rezai T., Yu B., Millhauser G.L., Jacobson M.P., Lokey R.S. Testing the conformational hypothesis of passive membrane permeability using synthetic cyclic peptide diastereomers. J. Am. Chem. Soc. 2006;128:2510–2511. doi: 10.1021/ja0563455. [DOI] [PubMed] [Google Scholar]
- 25.Áron R. Ph.D. Thesis. University of Toronto; Toronto, ON, Canada: 2014. Towards Targeted Photodynamic Therapy: Synthesis and Characterization of Aziridine Aldehyde-Cyclized Cancer-Targeting Peptides and Bacteriochlorin Photosensitizers. [Google Scholar]
- 26.Malaker A., Ahmad S.A.I. Therapeutic potency of anticancer peptides derived from marine organism. Int. J. Eng. Appl. Sci. 2013;2:53–65. [Google Scholar]
- 27.Liu J., Gu B., Yang L., Yang F., Lin H., Yang F. New anti-inflammatory cyclopeptides from a sponge-derived fungus Aspergillus violaceofuscus. Front. Chem. 2018;6:1–8. doi: 10.3389/fchem.2018.00226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Desriac F., Jé C., Balnois E., Brillet B., le Chevalier P. Antimicrobial peptides from marine proteobacteria. Mar. Drugs. 2013;11:3632–3660. doi: 10.3390/md11103632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wu W., Zhen Z., Niu T., Zhu X., Gao Y., Yan J., Chen Y., Yan X., Chen H. κ-Carrageenan enhances lipopolysaccharide-induced interleukin-8 secretion by stimulating the Bcl10-NF-κB pathway in HT-29 cells and aggravates C. freundii-Induced inflammation in mice. Mediat. Inflamm. 2017;2017:8634865. doi: 10.1155/2017/8634865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Randazzo A., Bifulco G., Giannini C., Bucci M., Cirino G., Gomez-paloma L., Ponte V., Salerno F., Ii F., Montesano V.D., et al. Halipeptins A and B: Two novel potent anti-inflammatory cyclic depsipeptides from the Vanuatu marine sponge Haliclona species. J. Am. Chem. Soc. 2001;123:10870–10876. doi: 10.1021/ja010015c. [DOI] [PubMed] [Google Scholar]
- 31.Xing H., Tong M., Jiang N., Zhang X., Hu H., Pan H., Li D. Antitumour bioactive peptides isolated from marine organisms. Clin. Exp. Pharmacol. Physiol. 2017;44:1077–1082. doi: 10.1111/1440-1681.12808. [DOI] [PubMed] [Google Scholar]
- 32.Laport M., Santos O., Muricy G. Marine sponges: Potential sources of new antimicrobial drugs. Curr. Pharm. BioTechnol. 2009;10:86–105. doi: 10.2174/138920109787048625. [DOI] [PubMed] [Google Scholar]
- 33.Mehbub M.F., Lei J., Franco C., Zhang W. Marine Sponge derived natural products between 2001 and 2010: Trends and opportunities for discovery of bioactives. Mar. Drugs. 2014;12:4539–4577. doi: 10.3390/md12084539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kang H.K., Choi M.C., Seo C.H., Park Y. Therapeutic properties and biological benefits of marine-derived anticancer peptides. Int. J. Mol. Sci. 2018;19:919. doi: 10.3390/ijms19030919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Nakao Y., Yoshida S., Matsunaga S., Shindoh N., Terada Y., Nagai K., Yamashita J.K., Ganesan A., van Soest R.W.M., Fusetani N. Azumamides A-E: Histone deacetylase inhibitory cyclic tetrapeptides from the marine sponge Mycale izuensis. Angew. Chem. Int. Ed. 2006;45:7553–7557. doi: 10.1002/anie.200602047. [DOI] [PubMed] [Google Scholar]
- 36.Sriraksa R., Limpaiboon T. Histone deacetylases and their inhibitors as potential therapeutic drugs for cholangiocarcinoma-cell line findings. Asian Pac. J. Cancer Prev. 2013;14:2503–2508. doi: 10.7314/APJCP.2013.14.4.2503. [DOI] [PubMed] [Google Scholar]
- 37.Abdalla M.A. Medicinal significance of naturally occurring cyclotetrapeptides. J. Nat. Med. 2016;70:708–720. doi: 10.1007/s11418-016-1001-5. [DOI] [PubMed] [Google Scholar]
- 38.Maulucci N., Chini M.G., di Micco S., Izzo I., Cafaro E., Russo A., Gallinari P., Paolini C., Nardi M.C., Casapullo A., et al. Molecular insights into azumamide E histone deacetylases inhibitory activity. J. Am. Chem. Soc. 2007;129:3007–3012. doi: 10.1021/ja0686256. [DOI] [PubMed] [Google Scholar]
- 39.Wen S., Carey K.L., Nakao Y., Fusetani N., Packham G., Ganesan A. Total synthesis of azumamide A and azumamide E, evaluation as histone deacetylase inhibitors, and design of a more potent analogue. Org. Lett. 2007;9:1105–1108. doi: 10.1021/ol070046y. [DOI] [PubMed] [Google Scholar]
- 40.Zhang X., Jacob M.R., Rao R.R., Wang Y.H., Agarwal A.K., Newman D.J., Khan I.A., Clark A.M., Li X.C. Antifungal cyclic peptides from the marine sponge Microscleroderma herdmani [Corrigendum] Res. Rep. Med. Chem. 2013;3:9–10. doi: 10.2147/rrmc.s44029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bewley C.A., Detritus C., Faulkner D.J. Microsclerodermins A and B. Antifungal cyclic peptides from the lithistid sponge Microscleroderma sp. J. Am. Chem. Soc. 1994;116:7631–7636. doi: 10.1021/ja00096a020. [DOI] [Google Scholar]
- 42.Guzmán E.A., Maers K., Roberts J., Kemami-Wangun H.V., Harmody D., Wright A.E. The marine natural product microsclerodermin A is a novel inhibitor of the nuclear factor kappa B and induces apoptosis in pancreatic cancer cells. Investig. New Drugs. 2015;33:86–94. doi: 10.1007/s10637-014-0185-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kehraus S., Ko G.M., Wright A.D., Bonn D., Woerheide G., Reef B. Leucamide A: A new cytotoxic heptapeptide from the Australian sponge Leucetta microraphis leucamide. J. Org. Chem. 2002;67:4989–4992. doi: 10.1021/jo020058r. [DOI] [PubMed] [Google Scholar]
- 44.Wang W., Nan F. First total synthesis of leucamide A. J. Org. Chem. 2003;68:1636–1639. doi: 10.1021/jo026799+. [DOI] [PubMed] [Google Scholar]
- 45.Afifi A.H., El-Desoky A.H., Kato H., Mangindaan R.E.P., de Voogd N.J., Ammar N.M., Hifnawy M.S., Tsukamoto S. Carteritins A and B, cyclic heptapeptides from the marine sponge Stylissa carteri. Tetrahedron Lett. 2016;57:1285–1288. doi: 10.1016/j.tetlet.2016.02.031. [DOI] [Google Scholar]
- 46.Sun J., Cheng W., de Voogd N.J., Proksch P., Lin W. Stylissatins B–D, cycloheptapeptides from the marine sponge Stylissa massa. Tetrahedron Lett. 2016;57:4288–4292. doi: 10.1016/j.tetlet.2016.08.024. [DOI] [Google Scholar]
- 47.Bonnington L.S., Tanaka J., Higa T., Kimura J., Yoshimura Y., Nakao Y., Yoshida W.Y., Scheuer P.J. Cupolamide A: A cytotoxic cyclic heptapeptide from two samples of the sponge Theonella cupola. J. Org. Chem. 2002;62:7765–7767. doi: 10.1021/jo9710285. [DOI] [Google Scholar]
- 48.Rangel M., Konno K., Brunaldi K., Procopio J., de Freitas J.C. Neurotoxic activity induced by a haemolytic substance in the extract of the marine sponge Geodia corticostylifera. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005;141:207–215. doi: 10.1016/j.cca.2005.06.004. [DOI] [PubMed] [Google Scholar]
- 49.Rangel M., Prado M.P., Konno K., Naoki H., Freitas J.C., Machado-Santelli G.M. Cytoskeleton alterations induced by Geodia corticostylifera depsipeptides in breast cancer cells. Peptides. 2006;27:2047–2057. doi: 10.1016/j.peptides.2006.04.021. [DOI] [PubMed] [Google Scholar]
- 50.Freitas V.M., Rangel M., Bisson L.F., Jaeger R.G., Machado-Santelli G.M. The geodiamolide H, derived from Brazilian sponge Geodia corticostylifera, regulates actin cytoskeleton, migration and invasion of breast cancer cells cultured in three-dimensional environment. J. Cell. Physiol. 2008;216:583–594. doi: 10.1002/jcp.21432. [DOI] [PubMed] [Google Scholar]
- 51.Ratnayake A.S., Bugni T.S., Feng X., Harper M.K., Skalicky J.J., Mohammed K.A., Andjelic C.D., Barrows L.R., Ireland C.M. Theopapuamide, a cyclic depsipeptide from a Papua New Guinea lithistid sponge Theonella swinhoei. J. Nat. Prod. 2006;69:1582–1586. doi: 10.1021/np060229d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zampella A., Sepe V., Bellotta F., Luciano P., D’Auria M.V., Cresteil T., Debitus C., Petek S., Poupat C., Ahond A. Homophymines B-E and A1-E1, a family of bioactive cyclodepsipeptides from the sponge Homophymia sp. Org. Biomol. Chem. 2009;7:4037–4044. doi: 10.1039/b910015f. [DOI] [PubMed] [Google Scholar]
- 53.Zampella A., Sepe V., Luciano P., Bellotta F., Monti M.C., D’Auria M.V., Jepsen T., Petek S., Adeline M.T., Laprévôte O., et al. Homophymine A, an anti-HIV cyclodepsipeptide from the sponge Homophymia sp. J. Org. Chem. 2008;73:5319–5327. doi: 10.1021/jo800583b. [DOI] [PubMed] [Google Scholar]
- 54.Sorres J., Martin M.T., Petek S., Levaique H., Cresteil T., Ramos S., Thoison O., Debitus C., Al-Mourabit A. Pipestelides A–C: Cyclodepsipeptides from the pacific marine sponge Pipestela candelabra. J. Nat. Prod. 2012;75:759–763. doi: 10.1021/np200714m. [DOI] [PubMed] [Google Scholar]
- 55.Tran T.D., Pham N.B., Fechner G., Zencak D., Vu H.T., Hooper J.N.A., Quinn R.J. Cytotoxic cyclic depsipeptides from the Australian marine sponge Neamphius huxleyi. J. Nat. Prod. 2012;75:2200–2208. doi: 10.1021/np3006474. [DOI] [PubMed] [Google Scholar]
- 56.Yamano Y., Arai M., Kobayashi M. Neamphamide B, new cyclic depsipeptide, as an anti-dormant mycobacterial substance from a Japanese marine sponge of Neamphius sp. Bioorganic Med. Chem. Lett. 2012;22:4877–4881. doi: 10.1016/j.bmcl.2012.05.071. [DOI] [PubMed] [Google Scholar]
- 57.Stierhof M., Hansen K.Ø., Sharma M., Feussner K., Subko K., Díaz-Rullo F.F., Isaksson J., Pérez-Victoria I., Clarke D., Hansen E., et al. New cytotoxic callipeltins from the Solomon Island marine sponge Asteropus sp. Tetrahedron. 2016;72:6929–6934. doi: 10.1016/j.tet.2016.09.016. [DOI] [Google Scholar]
- 58.Coello L., Reyes F., Martín M.J., Cuevas C., Fernández R. Isolation and structures of pipecolidepsins A and B, cytotoxic cyclic depsipeptides from the madagascan sponge Homophymia lamellosa. J. Nat. Prod. 2014;77:298–303. doi: 10.1021/np400888e. [DOI] [PubMed] [Google Scholar]
- 59.Gimeno M.P. Ph.D. Thesis. University of Barcelona; Barcelona, Spain: 2013. Synthesis, Structural Elucidation and Biological Evaluation of Pipecolidepsin A and Synthesis, Structural Elucidation and Biological Evaluation of Pipecolidepsin A and Phakellistatin 19. [Google Scholar]
- 60.Molina-Guijarro J.M., Moneo V., Martinez-Leal J.F., Cuevas C., Garcia-Fernandez L.F., Galmarini C.M. Pipecolidepsin A, stellatolide A and irvalec: New cyclodepsipeptides of marine origin with antitumor activity. Eur. J. Cancer. 2014;50:24. doi: 10.1016/S0959-8049(14)70184-4. [DOI] [Google Scholar]
- 61.Pelay-Gimeno M., García-Ramos Y., Martin M.J., Spengler J., Molina-Guijarro J.M., Munt S., Francesch A.M., Cuevas C., Tulla-Puche J., Albericio F. The first total synthesis of the cyclodepsipeptide pipecolidepsin A. Nat. Commun. 2013;4:1–10. doi: 10.1038/ncomms3352. [DOI] [PubMed] [Google Scholar]
- 62.Nicolaou K.C., Schlawe D., Kim D.W., Longbottom D.A., de Noronha R.G., Lizos D.E., Manam R.R., Faulkner D.J. Total synthesis of halipeptins: Isolation of halipeptin D and synthesis of oxazoline halipeptin analogues. Chem. A Eur. J. 2005;11:6198–6211. doi: 10.1002/chem.200500624. [DOI] [PubMed] [Google Scholar]
- 63.Nicolaou K.C., Lizos D.E., Kim D.W., Schlawe D., de Noronha R.G., Longbottom D.A., Rodriquez M., Bucci M., Cirino G. Total synthesis and biological evaluation of halipeptins A and D and analogues. J. Am. Chem. Soc. 2006;128:4460–4470. doi: 10.1021/ja060064v. [DOI] [PubMed] [Google Scholar]
- 64.Zhan K., Jiao W., Yang F., Li J., Wang S., Li Y., Han B., Lin H. Reniochalistatins A−E, cyclic peptides from the marine sponge Reniochalina stalagmitis. J. Nat. Prod. 2014;77:2678–2684. doi: 10.1021/np5006778. [DOI] [PubMed] [Google Scholar]
- 65.Fatino A., Baca G., Weeramange C., Rafferty R.J. Total synthesis of reniochalistatin E. J. Nat. Prod. 2017;80:3234–3240. doi: 10.1021/acs.jnatprod.7b00656. [DOI] [PubMed] [Google Scholar]
- 66.Ibrahim S.R.M., Edrada-Ebel R.A., Mohamed G.A., Youssef D.T.A., Wray V., Proksch P. Callyaerin G, a new cytotoxic cyclic peptide from the marine sponge Callyspongia aerizusa. Arkivoc. 2008;12:164–171. doi: 10.3998/ark.5550190.0009.c18. [DOI] [Google Scholar]
- 67.Ibrahim S.R.M., Min C.C., Teuscher F., Ebel R., Kakoschke C., Lin W., Wray V., Edrada-Ebel R., Proksch P. Callyaerins A-F and H, new cytotoxic cyclic peptides from the Indonesian marine sponge Callyspongia aerizusa. Bioorganic Med. Chem. 2010;18:4947–4956. doi: 10.1016/j.bmc.2010.06.012. [DOI] [PubMed] [Google Scholar]
- 68.Schmidt E.W., Raventos-Suarez C., Bifano M., Menendez A.T., Fairchild C.R., Faulkner D.J. Scleritodermin A, a cytotoxic cyclic peptide from the Lithistid sponge Scleritoderma nodosum. J. Nat. Prod. 2004;67:475–478. doi: 10.1021/np034035z. [DOI] [PubMed] [Google Scholar]
- 69.Sellanes D., Manta E., Serra G. Toward the total synthesis of Scleritodermin A: Preparation of the C1-N15 fragment. Tetrahedron Lett. 2007;48:1827–1830. doi: 10.1016/j.tetlet.2007.01.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Liu S., Cui Y.M., Nan F.J. Total synthesis of the originally proposed and revised structures of scleritodermin A. Org. Lett. 2008;10:3765–3768. doi: 10.1021/ol801419m. [DOI] [PubMed] [Google Scholar]
- 71.Kimura M., Wakimoto T., Egami Y., Tan K.C., Ise Y., Abe I. Calyxamides A and B, cytotoxic cyclic peptides from the marine sponge Discodermia calyx. J. Nat. Prod. 2012;75:290–294. doi: 10.1021/np2009187. [DOI] [PubMed] [Google Scholar]
- 72.Kobayashi J., Itagaki F., Shigemori I., Takao T., Shimonishi Y. Keramamides E, G, H, and J, new cyclic peptides containing an oxazole or a thiazole ring from a Theonella sponge. Tetrahedron. 1995;51:2525–2532. doi: 10.1016/0040-4020(95)00017-3. [DOI] [Google Scholar]
- 73.Uemoto H., Yahiro Y., Shigemori H., Tsuda M., Takao T., Shimonishi Y., Kobayashi J. Keramamides K and L, new cyclic peptides containing unusual tryptophan residue from Theonella sponge. Tetrahedron. 1998;54:6719–6724. doi: 10.1016/S0040-4020(98)00358-5. [DOI] [Google Scholar]
- 74.Tsuda M., Ishiyama H., Masuko K., Takao T., Shimonishi Y., Kobayashi J. Keramamides M and N, two new cyclic peptides with a sulfate ester from Theonella sponge. Tetrahedron. 1999;55:12543–12548. doi: 10.1016/S0040-4020(99)00744-9. [DOI] [Google Scholar]
- 75.Junk L., Kazmaier U. Total Synthesis of Keramamides A and L from a common precursor by late-stage indole synthesis and configurational revision. Angew. Chem. Int. Ed. 2018;57:11432–11435. doi: 10.1002/anie.201806657. [DOI] [PubMed] [Google Scholar]
- 76.Randazzo A., Dal F., Orru S., Gomez-paloma L. Axinellins A and B: New proline-containing antiproliferative cyclopeptides from the Vanuatu sponge Axinella carteri. Eur. J. Org. Chem. 1998;3:2659–2665. doi: 10.1002/(SICI)1099-0690(199811)1998:11<2659::AID-EJOC2659>3.0.CO;2-H. [DOI] [Google Scholar]
- 77.Fairweather K.A., Sayyadi N., Roussakis C., Jolliffe K.A. Synthesis of the cyclic heptapeptide axinellin A. Tetrahedron. 2010;66:935–939. doi: 10.1016/j.tet.2009.11.090. [DOI] [Google Scholar]
- 78.Brennan M.R., Costello C.E., Maleknia S.D., Pettit G.R., Erickson K.L. Stylopeptide 2, a proline-rich cyclodecapeptide from the sponge Stylotella sp. J. Nat. Prod. 2008;71:453–456. doi: 10.1021/np0704856. [DOI] [PubMed] [Google Scholar]
- 79.Arai M., Yamano Y., Fujita M., Setiawan A., Kobayashi M. Stylissamide X, a new proline-rich cyclic octapeptide as an inhibitor of cell migration, from an Indonesian marine sponge of Stylissa sp. Bioorganic Med. Chem. Lett. 2012;22:1818–1821. doi: 10.1016/j.bmcl.2011.10.023. [DOI] [PubMed] [Google Scholar]
- 80.Huang T., Zou Y., Wu M.C., Zhao Q.J., Hu H.G. Total synthesis of proline-rich cyclic octapeptide stylissamide X. Chem. Nat. Compd. 2015;51:523–526. doi: 10.1007/s10600-015-1329-1. [DOI] [Google Scholar]
- 81.Bewley C.A., He H., Williams D.H., Faulkner D.J. Aciculitins A–C: Cytotoxic and antifungal cyclic peptides from the lithistid sponge Aciculites orientalis. J. Am. Chem. Soc. 1996;118:4314–4321. doi: 10.1021/ja953628w. [DOI] [Google Scholar]
- 82.Fukuhara K., Takada K., Okada S., Matsunaga S. Nazumazoles A-C, cyclic pentapeptides dimerized through a disulfide bond from the marine sponge Theonella swinhoei. Org. Lett. 2015;17:2646–2648. doi: 10.1021/acs.orglett.5b01020. [DOI] [PubMed] [Google Scholar]
- 83.Youssef D.T.A., Shaala L.A., Mohamed G.A., Badr J.M., Bamanie F.H., Ibrahim S.R.M. Theonellamide G, a potent antifungal and cytotoxic bicyclic glycopeptide from the red sea marine sponge Theonella swinhoei. Mar. Drugs. 2014;12:1911–1923. doi: 10.3390/md12041911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Araki T., Matsunaga S., Nakao Y., Furihata K., West L., Faulkner D.J., Fusetani N. Koshikamide B, a cytotoxic peptide lactone from a marine sponge Theonella sp. J. Org. Chem. 2008;73:7889–7894. doi: 10.1021/jo801032n. [DOI] [PubMed] [Google Scholar]
- 85.Shenkar N., Swalla B.J. Global diversity of Ascidiacea. PLoS ONE. 2011;6:e20657. doi: 10.1371/journal.pone.0020657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Marino A., Rajendran N.M. Natural products diversity of marine ascidians (Tunicates; Ascidiacea) and successful drugs in clinical development. Nat. Prod. Bioprospect. 2017;7:1–111. doi: 10.1007/s13659-016-0115-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Hockless D.C.R., Skelton B.W., White A.H. Studies of australian ascidians. IV. mollamide, a cytotoxic cyclic heptapeptide from the compound ascidian. Aust. J. Chem. 1994;47:61–69. doi: 10.1071/CH9940061. [DOI] [Google Scholar]
- 88.McKeever B., Pattenden G. Total synthesis of the cytotoxic cyclopeptide mollamide, isolated from the sea squirt Didemnum molle. Tetrahedron. 2003;59:2701–2712. doi: 10.1016/S0040-4020(03)00293-X. [DOI] [Google Scholar]
- 89.Donia M.S., Wang B., Dunbar D.C., Desai P.V., Patny A., Avery M., Hamann M.T. Mollamides B and C, cyclic hexapeptides from the indonesian tunicate Didemnum molle. J. Nat. Prod. 2008;71:941–945. doi: 10.1021/np700718p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.McKeever B., Pattenden G. Total synthesis of trunkamide A, a novel thiazoline-based prenylated cyclopeptide metabolite from Lissoclinum sp. Tetrahedron. 2003;59:2713–2727. doi: 10.1016/S0040-4020(03)00294-1. [DOI] [Google Scholar]
- 91.Bowden B.F., Garcia G.D. A Cyclic Hepta-Peptide Derivative from Colonial Ascidians, Lissoclinum sp. EP0894092B1. European Patent. 2002 Apr 3;
- 92.Fu X., Su J., Zeng L. Prepatellamide A, a new cyclic peptide from the ascidian Lissoclinum patella. Sci. China Ser. B Chem. 2008;43:643–648. doi: 10.1007/BF02969512. [DOI] [Google Scholar]
- 93.Fernandez A.M. Ph.D. Thesis. The University of Utah; Salt Lake City, UT, USA: 1996. Isolation and Characterization of Vitilevuamide from the Ascidians Didemnum cuculliferum and Polysyncraton lithostrotum. [Google Scholar]
- 94.Edler M.C., Fernandez A.M., Lassota P., Ireland C.M., Barrows L.R. Inhibition of tubulin polymerization by vitilevuamide, a bicyclic marine peptide, at a site distinct from colchicine, the vinca alkaloids, and dolastatin 10. Biochem. Pharmacol. 2002;63:707–715. doi: 10.1016/S0006-2952(01)00898-X. [DOI] [PubMed] [Google Scholar]
- 95.Pangestuti R., Kim S.K. Bioactive peptide of marine origin for the prevention and treatment of non-communicable diseases. Mar. Drugs. 2017;15:67. doi: 10.3390/md15030067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Vervoort H., Fenical W., Epifanio R.D.A. Tamandarins A and B: New cytotoxic depsipeptides from a Brazilian ascidian of the family Didemnidae. J. Org. Chem. 2000;65:782–792. doi: 10.1021/jo991425a. [DOI] [PubMed] [Google Scholar]
- 97.Liang B., Richard D.J., Portonovo P.S., Joullie M.M. Total Syntheses and biological investigations of tamandarins A and B and tamandarin A analogs. J. Am. Chem. Soc. 2001;123:4469–4474. doi: 10.1021/ja010222c. [DOI] [PubMed] [Google Scholar]
- 98.Rudi A., Aknin M., Gaydou E.M., Kashman Y. Four new cytotoxic cyclic hexa- and heptapeptides from the marine ascidian Didemnum molle. Tetrahedron. 1998;54:13203–13210. doi: 10.1016/S0040-4020(98)00801-1. [DOI] [Google Scholar]
- 99.Carroll A., Coll J., Bourne D., Macleod J., Zabriskie T., Ireland C., Bowden B. Patellins 1–6 and trunkamide A: Novel cyclic hexa-, hepta- and octa-peptides from colonial ascidians, Lissoclinum sp. Aust. J. Chem. 1996;49:659–667. doi: 10.1071/CH9960659. [DOI] [Google Scholar]
- 100.Tan K.C., Wakimoto T., Takada K., Ohtsuki T., Uchiyama N., Goda Y., Abe I. Cycloforskamide, a cytotoxic macrocyclic peptide from the sea slug Pleurobranchus forskalii. J. Nat. Prod. 2013;76:1388–1391. doi: 10.1021/np400404r. [DOI] [PubMed] [Google Scholar]
- 101.Rosenberg G. A New critical estimate of named species-level diversity of the recent Mollusca. Am. Malacol. Bull. 2014;32:308–322. doi: 10.4003/006.032.0204. [DOI] [Google Scholar]
- 102.Chakraborty S., Ghosh U. Oceans: A store house of drugs-a review. J. Pharm. Res. 2010;3:1293–1296. [Google Scholar]
- 103.Wesson K.J., Hamann M.T. Keenamide A, a bioactive cyclic peptide from the marine mollusk Pleurobranchus forskalii. J. Nat. Prod. 1996;59:629–631. doi: 10.1021/np960153t. [DOI] [PubMed] [Google Scholar]
- 104.Suenaga K., Mutou T., Shibata T., Itoh T., Fujita T., Takada N., Hayamizu K., Takagi M., Irifune T. Aurilide, a cytotoxic depsipeptide from the sea hare Dolabella auricularia: Isolation, structure determination, synthesis, and biological activity. Tetrahedron. 2004;60:8509–8527. doi: 10.1016/j.tet.2004.06.125. [DOI] [Google Scholar]
- 105.Sato S.I., Murata A., Orihara T., Shirakawa T., Suenaga K., Kigoshi H., Uesugi M. Marine natural product aurilide activates the opa1-mediated apoptosis by binding to prohibitin. Chem. Biol. 2011;18:131–139. doi: 10.1016/j.chembiol.2010.10.017. [DOI] [PubMed] [Google Scholar]
- 106.Pettit G.R., Xu J.P., Hogan F., Williams M.D., Doubek D.L., Schmidt J.M., Cerny R.L., Boyd M.R. Isolation and structure of the human cancer cell growth inhibitory cyclodepsipeptide dolastatin 16. J. Nat. Prod. 1997;60:752–754. doi: 10.1021/np9700230. [DOI] [PubMed] [Google Scholar]
- 107.Ishiwata H., Nemoto T., Ojika M., Yamada K. Isolation and stereostructure of doliculide, a cytotoxic cyclodepsipeptide from the Japanese sea hare Dolabella auricularia. J. Org. Chem. 1994;59:4710–4711. doi: 10.1021/jo00096a002. [DOI] [Google Scholar]
- 108.Ishiwata H., Sone H., Kigoshi H., Yamada K. Total synthesis of doliculide, a potent cytotoxic cyclodepsipeptide from the Japanese sea hare Dolabella auricularia. J. Org. Chem. 1994;59:4712–4713. doi: 10.1021/jo00096a003. [DOI] [Google Scholar]
- 109.Matcha K., Madduri A.V.R., Roy S., Ziegler S., Waldmann H., Hirsch A.K.H., Minnaard A.J. Total synthesis of (-)-doliculide, structure-activity relationship studies and its binding to F-actin. ChemBioChem. 2012;13:2537–2548. doi: 10.1002/cbic.201200512. [DOI] [PubMed] [Google Scholar]
- 110.Foerster F., Braig S., Chen T., Altmann K.H., Vollmar A.M. Pharmacological characterization of actin-binding (-)-doliculide. Bioorganic Med. Chem. 2015;22:5117–5122. doi: 10.1016/j.bmc.2014.03.003. [DOI] [PubMed] [Google Scholar]
- 111.Kimura J., Takada Y., Inayoshi T., Nakao Y., Goetz G., Yoshida W.Y., Scheuer P.J. Kulokekahilide-1, a cytotoxic depsipeptide from the cephalaspidean mollusk Philinopsis speciosa. J. Org. Chem. 2002;67:1760–1767. doi: 10.1021/jo010176z. [DOI] [PubMed] [Google Scholar]
- 112.Nakao Y., Yoshida W.Y., Takada Y., Kimura J., Yang L., Mooberry S.L., Scheuer P.J. Kulokekahilide-2, a cytotoxic depsipeptide from a cephalaspidean mollusk Philinopsis speciosa. J. Nat. Prod. 2004;67:1332–1340. doi: 10.1021/np049949f. [DOI] [PubMed] [Google Scholar]
- 113.Umehara M., Negishi T., Tashiro T., Nakao Y., Kimura J. Structure-related cytotoxic activity of derivatives from kulokekahilide-2, a cyclodepsipeptide in Hawaiian marine mollusk. Bioorganic Med. Chem. Lett. 2012;22:7422–7425. doi: 10.1016/j.bmcl.2012.10.058. [DOI] [PubMed] [Google Scholar]
- 114.Umehara M., Negishi T., Maehara Y., Nakao Y., Kimura J. Stereochemical analysis and cytotoxicity of kulokekahilide-2 and its analogues. Tetrahedron. 2013;69:3045–3053. doi: 10.1016/j.tet.2013.01.089. [DOI] [Google Scholar]
- 115.Von Schwarzenberg K., Vollmar A.M. Targeting apoptosis pathways by natural compounds in cancer: Marine compounds as lead structures and chemical tools for cancer therapy. Cancer Lett. 2013;332:295–303. doi: 10.1016/j.canlet.2010.07.004. [DOI] [PubMed] [Google Scholar]
- 116.Martín-Algarra S., Espinosa E., Rubió J., López J.J.L., Manzano J.L., Carrión L.A., Plazaola A., Tanovic A., Paz-Ares L. Phase II study of weekly kahalalide F in patients with advanced malignant melanoma. Eur. J. Cancer. 2009;45:732–735. doi: 10.1016/j.ejca.2008.12.005. [DOI] [PubMed] [Google Scholar]
- 117.Gao J., Hamann M.T. Chemistry and biology of lipids. Chem. Rev. 2009;53:35–43. doi: 10.1021/cr100187n. [DOI] [Google Scholar]
- 118.El Gamal A.A. Biological importance of marine algae. Saudi Pharm. J. 2010;18:1–25. doi: 10.1016/j.jsps.2009.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Xu W.J., Liao X.J., Xu S.H., Diao J.Z., Du B., Zhou X.L., Pan S.S. Isolation, structure determination, and synthesis of galaxamide, a rare cytotoxic cyclic pentapeptide from a marine algae Galaxaura filamentosa. Org. Lett. 2008;10:4569–4572. doi: 10.1021/ol801799d. [DOI] [PubMed] [Google Scholar]
- 120.Lunagariya J., Liao X., Long W., Zhong S., Bhadja P., Li H., Zhao B., Xu S. Cytotoxicity study of cyclopentapeptide analogues of marine natural product galaxamide towards human breast cancer cells. Oxid. Med. Cell. Longev. 2017;2017 doi: 10.1155/2017/8392035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Allkin B. State of the World’s Plants 201. Royal Botanic Gardens, Kew; London, UK: 2017. Useful Plants–Medicines: At Least 28,187 Plant Species are Currently Recorded as Being of Medicinal Use. [PubMed] [Google Scholar]
- 122.Wu D., Gao Y., Qi Y., Chen L., Ma Y., Li Y. Peptide-based cancer therapy: Opportunity and challenge. Cancer Lett. 2014;351:13–22. doi: 10.1016/j.canlet.2014.05.002. [DOI] [PubMed] [Google Scholar]
- 123.Wélé A., Zhang Y., Ndoye I., Brouard J.P., Pousset J.L., Bodo B. A cytotoxic cyclic heptapeptide from the seeds of Annona cherimola. J. Nat. Prod. 2004;67:1577–1579. doi: 10.1021/np040068i. [DOI] [PubMed] [Google Scholar]
- 124.Wélé A., Ndoye I., Zhang Y., Brouard J.P., Bodo B. Cherimolacyclopeptide D, a novel cycloheptapeptide from the seeds of Annona cherimola. Phytochemistry. 2005;66:693–696. doi: 10.1016/j.phytochem.2004.12.011. [DOI] [PubMed] [Google Scholar]
- 125.Wélé A., Zhang Y., Brouard J.P., Pousset J.L., Bodo B. Two cyclopeptides from the seeds of Annona cherimola. Phytochemistry. 2005;66:2376–2380. doi: 10.1016/j.phytochem.2005.06.011. [DOI] [PubMed] [Google Scholar]
- 126.Idrissa N., Adama D., Mamadou B., Rokhaya S.G., Yoro T. Novel cytotoxic cycloheptapeptide from the latex of Jatropha integerrima. J. Chem. Pharm. Res. 2016;8:135–139. [Google Scholar]
- 127.Mongkolvisut W., Sutthivaiyakit S., Leutbecher H., Mika S., Klaiber I., Möller W., Rösner H., Beifuss U., Conrad J. Integerrimides A and B, cyclic heptapeptides from the latex of Jatropha integerrima. J. Nat. Prod. 2006;69:1435–1441. doi: 10.1021/np0602012. [DOI] [PubMed] [Google Scholar]
- 128.Goyal A., Sharma V., Upadhyay N., Gill S., Sihag M. Flax and flaxseed oil: An ancient medicine & modern functional food. J. Food Sci. Technol. 2014;51:1633–1653. doi: 10.1007/s13197-013-1247-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Okinyo-Owiti D.P., Dong Q., Ling B., Jadhav P.D., Bauer R., Maley J.M., Reaney M.J.T., Yang J., Sammynaiken R. Evaluating the cytotoxicity of flaxseed orbitides for potential cancer treatment. Toxicol. Rep. 2015;2:1014–1018. doi: 10.1016/j.toxrep.2015.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Lomchoey N., Panseeta P., Boonsri P., Apiratikul N., Prabpai S., Kongsaeree P., Suksamrarn S. New bioactive cyclopeptide alkaloids with rare terminal unit from the root bark of: Ziziphus cambodiana. RSC Adv. 2018;8:18204–18215. doi: 10.1039/C7RA13050C. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Hsieh P.-W., Chang F.-R., Wu C.-C., Li C.-M., Wu K.-Y., Chen S.-L., Yen H.-F., Wu Y.-C. Longicalycinin A, a New cytotoxic cyclic peptide from Dianthus superbus var. longicalycinus (MAXIM.) WILL. Chem. Pharm. Bull. 2005;53:336–338. doi: 10.1248/cpb.53.336. [DOI] [PubMed] [Google Scholar]
- 132.Ahmad W. Solid-phase total synthesis of cyclic pentapeptide longicalycinin A, by using 2-chlorotrityl chloride resin. J. Cancer Res. Exp. Oncol. 2014;5:8–19. doi: 10.5897/JCREO2013.0102. [DOI] [Google Scholar]
- 133.HoushdarTehrani M.H., Bamoniri A., Mirjalili B.B.F., Gholibeikian M. Synthesis of linear and cyclic disulfide heptapeptides of longicalycinin a and evaluation of toxicity on cancerous cells HepG2 and HT-29. Iran. J. Pharm. Res. 2018;17:956–963. [PMC free article] [PubMed] [Google Scholar]
- 134.Hsieh P.W., Chang F.R., Wu C.C., Wu K.Y., Li C.M., Chen S.L., Wu Y.C. New cytotoxic cyclic peptides and dianthramide from Dianthus superbus. J. Nat. Prod. 2004;67:1522–1527. doi: 10.1021/np040036v. [DOI] [PubMed] [Google Scholar]
- 135.Shim Y.Y., Young L.W., Arnison P.G., Gilding E., Reaney M.J.T. Proposed systematic nomenclature for orbitides. J. Nat. Prod. 2015;78:645–652. doi: 10.1021/np500802p. [DOI] [PubMed] [Google Scholar]
- 136.Beirigo P.J.S.D., Torquato H.F.V., Santos C.H.C.D., de Carvalho M.G., Castro R.N., Paredes-Gamero E.J., de Sousa P.T., Jacinto M.J., da Silva V.C. [1-8-NαlC]-Zanriorb A1, a proapoptotic orbitide from leaves of Zanthoxylum riedelianum. J. Nat. Prod. 2016;79:1454–1458. doi: 10.1021/acs.jnatprod.5b00177. [DOI] [PubMed] [Google Scholar]
- 137.Lv J.P., Yang S., Dong J.X., Jin H. New cyclopeptide alkaloids from the whole plant of Justicia procumbens L. Nat. Prod. Res. 2020:1–9. doi: 10.1080/14786419.2020.1758090. [DOI] [PubMed] [Google Scholar]
- 138.Liu Z., Fu J., Xiao S., Wang D. Structural characterization of ginseng cyclopeptides and detection of capability to induce apoptosis in gastrointestinal cancer cells. RSC Adv. 2019;9:29847–29855. doi: 10.1039/C9RA03965A. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 139.Craik D.J., Daly N.L., Bond T., Waine C. Plant cyclotides: A unique family of cyclic and knotted proteins that defines the cyclic cystine knot structural motif. J. Mol. Biol. 1999;294:1327–1336. doi: 10.1006/jmbi.1999.3383. [DOI] [PubMed] [Google Scholar]
- 140.Craik D.J., Cemazar M., Wang C.K.L., Daly N.L. The cyclotide family of circular miniproteins: Nature’s combinatorial peptide teplate. Biopolymer. 2006;84:250–266. doi: 10.1002/bip.20451. [DOI] [PubMed] [Google Scholar]
- 141.Zhao S.M., Kuang B., Fan J.T., Yan H., Xu W.Y., Tan N.H. Antitumor cyclic hexapeptides from Rubia plants: History, chemistry, and mechanism (2005–2011) Chim. Int. J. Chem. 2011;65:952–956. doi: 10.2533/chimia.2011.952. [DOI] [PubMed] [Google Scholar]
- 142.Figueiredo P.O., Matos M.D.F.C., Perdomo R.T., Kato W.H., Barros M.V.G.O., Garcez F.R., Garcez W.S. Rubiaceae-type cyclopeptides from Galianthe thalictroides. J. Nat. Prod. 2016;79:1165–1169. doi: 10.1021/acs.jnatprod.5b00849. [DOI] [PubMed] [Google Scholar]
- 143.Wang Z., Zhao S.M., Zhao L.M., Chen X.Q., Zeng G.Z., Tan N.H. Rubipodanin A, the first natural ndesmonomethyl rubiaceae-type cyclopeptide from rubia podantha, indicating an important role of the n9- methyl group in the conformation and bioactivity. PLoS ONE. 2015;10:e0144950. doi: 10.1371/journal.pone.0144950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Hu Y.Y., Feng L., Wang J., Zhang X.J., Wang Z., Tan N.H. Rubipodanin B, a new cytotoxic cyclopeptide from Rubia podantha. Chem. Biodivers. 2019;16:16–21. doi: 10.1002/cbdv.201800438. [DOI] [PubMed] [Google Scholar]
- 145.Wang Y., Guo D., He J., Song L., Chen H., Zhang Z., Tan N. Inhibition of fatty acid synthesis arrests colorectal neoplasm growth and metastasis: Anti-cancer therapeutical effects of natural cyclopeptide RA-XII. Biochem. Biophys. Res. Commun. 2019;512:819–824. doi: 10.1016/j.bbrc.2019.03.088. [DOI] [PubMed] [Google Scholar]
- 146.Leung H.W., Zhao S.M., Yue G.G.L., Lee J.K.M., Fung K.P., Leung P.C., Tan N.H., Lau C.B.S. RA-XII inhibits tumour growth and metastasis in breast tumour-bearing mice via reducing cell adhesion and invasion and promoting matrix degradation. Sci. Rep. 2015;5:1–17. doi: 10.1038/srep16985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Song L., Wang Z., Wang Y., Guo D., Yang J., Chen L., Tan N. Natural cyclopeptide RA-XII, a new autophagy inhibitor, suppresses protective autophagy for enhancing apoptosis through AMPK/mTOR/P70S6K pathways in HepG2 Cells. Molecules. 2017;22:1934. doi: 10.3390/molecules22111934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Wang J., Wang J., Li L., Feng L., Wang Y.R., Wang Z., Tan N.H. RA-XII, a bicyclic hexapeptidic glucoside isolated from Rubia yunnanensis Diels, exerts antitumor activity by inhibiting protective autophagy and activating Akt-mTOR pathway in colorectal cancer cells. J. Ethnopharmacol. 2021;266:113438. doi: 10.1016/j.jep.2020.113438. [DOI] [PubMed] [Google Scholar]
- 149.Hitotsuyanagi Y., Hirai M., Odagiri M., Komine M., Hasuda T., Fukaya H., Takeya K. RA-XXV and RA-XXVI, Bicyclic hexapeptides from Rubia cordifolia L.: Structure, synthesis, and conformation. Chem. Asian J. 2019;14:205–215. doi: 10.1002/asia.201801466. [DOI] [PubMed] [Google Scholar]
- 150.Ding X., Bai D., Qian J. Novel cyclotides from Hedyotis biflora inhibit proliferation and migration of pancreatic cancer cell in vitro and in vivo. Med. Chem. Res. 2014;23:1406–1413. doi: 10.1007/s00044-013-0746-6. [DOI] [Google Scholar]
- 151.Nguyen G.K.T., Zhang S., Nguyen N.T.K., Nguyen P.Q.T., Chiu M.S., Hardjojo A., Tam J.P. Discovery and characterization of novel cyclotides originated from chimeric precursors consisting of albumin-1 chain a and cyclotide domains in the Fabaceae family. J. Biol. Chem. 2011;286:24275–24287. doi: 10.1074/jbc.M111.229922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Zhang S., Xiao K.Z., Jin J., Zhang Y., Zhou W. Chemosensitizing activities of cyclotides from clitoria ternatea in paclitaxel-resistant lung cancer cells. Oncol. Lett. 2013;5:641–644. doi: 10.3892/ol.2012.1042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Gerlach U., Burman S.L., Debasis Monda R., Goransson D. Characterization and bioactivity of cyclotides from Psychotria leptothyrsa (Rubiaceae) J. Nat. Prod. 2010;73:1207–1213. doi: 10.1021/np9007365. [DOI] [PubMed] [Google Scholar]
- 154.Hashempour H., Koehbach J., Daly N.L., Ghassempour A., Gruber C.W. Characterizing circular peptides in mixtures: Sequence fragment assembly of cyclotides from a violet plant by MALDI-TOF/TOF mass spectrometry. Amino Acids. 2013;44:581–595. doi: 10.1007/s00726-012-1376-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Esmaeili M.A., Abagheri-Mahabadi N., Hashempour H., Farhadpour M., Gruber C.W., Ghassempour A. Viola plant cyclotide vigno 5 induces mitochondria-mediated apoptosis via cytochrome C release and caspases activation in cervical cancer cells. Fitoterapia. 2016;109:162–168. doi: 10.1016/j.fitote.2015.12.021. [DOI] [PubMed] [Google Scholar]
- 156.Herrmann A., Svangård E., Claeson P., Gullbo J., Bohlin L., Göransson U. Key role of glutamic acid for the cytotoxic activity of the cyclotide cycloviolacin O2. Cell. Mol. Life Sci. 2006;63:235–245. doi: 10.1007/s00018-005-5486-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Burman R., Svedlund E., Felth J., Hassan S., Herrmann A., Clark R.J., Craik D.J., Bohlin L., Claeson P., Göransson U., et al. Evaluation of toxicity and antitumor activity of cycloviolacin O2 in mice. Biopolymers. 2010;94:626–634. doi: 10.1002/bip.21408. [DOI] [PubMed] [Google Scholar]
- 158.Svangård E., Burman R., Gunasekera S., Lövborg H., Gullbo J., Göransson U. Mechanism of action of cytotoxic cyclotides: Cycloviolacin O2 disrupts lipid membranes. J. Nat. Prod. 2007;70:643–647. doi: 10.1021/np070007v. [DOI] [PubMed] [Google Scholar]
- 159.Gerlach S.L., Rathinakumar R., Chakravarty G., Göransson U., Wimley W.C., Darwin S.P., Mondal D. Anticancer and chemosensitizing abilities of cycloviolacin 02 from Viola odorata and psyle cyclotides from Psychotria leptothyrsa. Biopolymers. 2010;94:617–625. doi: 10.1002/bip.21435. [DOI] [PubMed] [Google Scholar]
- 160.Gerlach S.L., Yeshak M., Göransson U., Roy U., Izadpanah R., Mondal D. Cycloviolacin O2 (CyO2) suppresses productive infection and augments the antiviral efficacy of nelfinavir in HIV-1 infected monocytic cells. Biopolymers. 2013;100:471–479. doi: 10.1002/bip.22325. [DOI] [PubMed] [Google Scholar]
- 161.He W., Yue L., Zeng G., Daly N.L., Craik D.J., Tan N. Peptides Isolation and characterization of cytotoxic cyclotides from Viola philippica. Peptides. 2011;32:1719–1723. doi: 10.1016/j.peptides.2011.06.016. [DOI] [PubMed] [Google Scholar]
- 162.Yeshak M.Y., Burman R., Asres K., Göransson U. Cyclotides from an extreme habitat: Characterization of cyclic peptides from Viola abyssinica of the Ethiopian highlands. J. Nat. Prod. 2011;74:727–731. doi: 10.1021/np100790f. [DOI] [PubMed] [Google Scholar]
- 163.Herrmann A., Burman R., Mylne J.S., Karlsson G., Gullbo J., Craik D.J., Clark R.J., Göransson U. The alpine violet, Viola biflora, is a rich source of cyclotides with potent cytotoxicity. Phytochemistry. 2008;69:939–952. doi: 10.1016/j.phytochem.2007.10.023. [DOI] [PubMed] [Google Scholar]
- 164.Svangård E., Göransson U., Hocaoglu Z., Gullbo J., Larsson R., Claeson P., Bohlin L. Cytotoxic cyclotides from Viola tricolor. J. Nat. Prod. 2004;67:144–147. doi: 10.1021/np030101l. [DOI] [PubMed] [Google Scholar]
- 165.Jun T., Wang C.K., Pan X., Yan H., Zeng G., Xu W., He W., Chan L.Y., Zeng G., Daly N.L., et al. Isolation and characterization of cytotoxic cyclotides from Viola tricolor. Peptides. 2010;32:1719–1723. doi: 10.1016/j.peptides.2011.06.016. [DOI] [PubMed] [Google Scholar]
- 166.Karoum F., Commissiong J.W., Neff N.H., Wyatt R.J. Biochemical evidence for uncrossed and crossed locus coeruleus projections to the spinal cord. Brain Res. 1980;196:237–241. doi: 10.1016/0006-8993(80)90730-1. [DOI] [PubMed] [Google Scholar]
- 167.Cryan J.F., Dinan T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. NeuroSci. 2012;13:701–712. doi: 10.1038/nrn3346. [DOI] [PubMed] [Google Scholar]
- 168.Masuoka Y., Nagai A., Shin-ya K., Furihata K., Nagai K., Suzuki K., Hayakawa Y., Seto H. Spiruchostatins A and B, novel gene expression-enhancing substances produced by Pseudomonas sp. Tetrahedron Lett. 2001;42:41–44. doi: 10.1016/S0040-4039(00)01874-8. [DOI] [Google Scholar]
- 169.Shindoh N., Mori M., Terada Y., Oda K., Amino N., Kita A., Taniguchi M., Sohda K.Y., Nagai K., Sowa Y., et al. YM753, a novel histone deacetylase inhibitor, exhibits antitumor activity with selective, sustained accumulation of acetylated histones in tumors in the WiDr xenograft model. Int. J. Oncol. 2008;32:545–555. doi: 10.3892/ijo.32.3.545. [DOI] [PubMed] [Google Scholar]
- 170.Toriyama S., Horinaka M., Yasuda S., Taniguchi T., Aono Y., Takamura T., Morioka Y., Miki T., Ukimura O., Sakai T. A histone deacetylase inhibitor, OBP-801, and celecoxib synergistically inhibit the cell growth with apoptosis via a DR5-dependent pathway in bladder cancer cells. Mol. Cancer Ther. 2016;15:2066–2075. doi: 10.1158/1535-7163.MCT-16-0010. [DOI] [PubMed] [Google Scholar]
- 171.Takamura T., Horinaka M., Yasuda S., Toriyama S., Aono Y., Sowa Y., Miki T., Ukimura O., Sakai T. FGFR inhibitor BGJ398 and HDAC inhibitor OBP-801 synergistically inhibit cell growth and induce apoptosis in bladder cancer cells. Oncol. Rep. 2018;39:627–632. doi: 10.3892/or.2017.6127. [DOI] [PubMed] [Google Scholar]
- 172.Shin-ya K., Wierzba K., Matsuo K. Telomestatin, a Novel Telomerase inhibitor from Streptomyces anulatus. J. Am. Chem. Soc. 2001;123:1262–1263. doi: 10.1021/ja005780q. [DOI] [PubMed] [Google Scholar]
- 173.Kim M.Y., Vankayalapati H., Shin-Ya K., Wierzba K., Hurley L.H. Telomestatin, a potent telomerase inhibitor that interacts quite specifically with the human telomeric intramolecular G-quadruplex. J. Am. Chem. Soc. 2002;124:2098–2099. doi: 10.1021/ja017308q. [DOI] [PubMed] [Google Scholar]
- 174.Liu W., Sun D., Hurley L.H. Binding of G-quadruplex-interactive agents to distinct G-quadruplexes induces different biological effects in MiaPaCa cells. Nucleosides Nucleotides Nucleic Acids. 2005;24:1801–1815. doi: 10.1080/15257770500267238. [DOI] [PubMed] [Google Scholar]
- 175.Binz N., Shalaby T., Rivera P., Shin-Ya K., Grotzer M.A. Telomerase inhibition, telomere shortening, cell growth suppression and induction of apoptosis by telomestatin in childhood neuroblastoma cells. Eur. J. Cancer. 2005;41:2873–2881. doi: 10.1016/j.ejca.2005.08.025. [DOI] [PubMed] [Google Scholar]
- 176.Amagai K., Ikeda H., Hashimoto J., Kozone I., Izumikawa M., Kudo F., Eguchi T., Nakamura T., Osada H., Takahashi S., et al. Identification of a gene cluster for telomestatin biosynthesis and heterologous expression using a specific promoter in a clean host. Sci. Rep. 2017;7:3382. doi: 10.1038/s41598-017-03308-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Arndt G.M., MacKenzie K.L. New prospects for targeting telomerase beyond the telomere. Nat. Rev. Cancer. 2016;16:508–524. doi: 10.1038/nrc.2016.55. [DOI] [PubMed] [Google Scholar]
- 178.Doi T., Yoshida M., Shin-ya K., Takahashi T. Total synthesis of (R)-telomestatin. Org. Lett. 2006;8:4165–4167. doi: 10.1021/ol061793i. [DOI] [PubMed] [Google Scholar]
- 179.Ducklow H.W. Bacterioplankton. Encycl. Ocean. Sci. 2001:217–224. doi: 10.1006/rwos.2001.0493. [DOI] [Google Scholar]
- 180.Joseph A., Guedes A.C., Malcata F.X. Biotechnological and pharmacological applications of biotoxins and other bioactive molecules from dinoflagellates. Mar. Drug. 2017;15:393. doi: 10.3390/md15120393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Lombó F., Velasco A., Castro A., de la Calle F., Braña A.F., Sánchez-Puelles J.M., Méndez C., Salas J.A. Deciphering the biosynthesis pathway of the antitumor thiocoraline from a marine actinomycete and its expression in two Streptomyces species. ChemBioChem. 2006;7:366–376. doi: 10.1002/cbic.200500325. [DOI] [PubMed] [Google Scholar]
- 182.Romero F., Espliego F., Baz J.P., de Quesada T.G., Grávalos D., la Calle F.D., Fernández-Puentes J.L. Thiocoraline, a New depsipeptide with antitumor activity produced by a marine Micromonospora. I. taxonomy, fermentation, isolation, and biological activities. J. Antibiot. 1997;50:734–737. doi: 10.7164/antibiotics.50.734. [DOI] [PubMed] [Google Scholar]
- 183.Erba E., Bergamaschi D., Ronzoni S., Faretta M., Taverna S., Bonfanti M., Catapano C.V., Faircloth G., Jimeno J., D’Incalci M. Mode of action at thiocoraline, a natural marine compound with anti-tumour activity. Br. J. Cancer. 1999;80:971–980. doi: 10.1038/sj.bjc.6690451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Sohn J.A., Zarebczan B., Wyche T.P., Bugni T.S., Kunnimalaiyaan M., Jaskula-Sztul R., Chen H. Thiocoraline regulates neuroendocrine phenotype and inhibits proliferation in carcinoid tumor cells. J. Surg. Res. 2012;172:237. doi: 10.1016/j.jss.2011.11.366. [DOI] [Google Scholar]
- 185.Tesfazghi S., Eide J., Dammalapati A., Korlesky C., Wyche T.P., Bugni T.S., Chen H., Jaskula-Sztul R. Thiocoraline alters neuroendocrine phenotype and activates the Notch pathway in MTC-TT cell line. Cancer Med. 2013;2:734–743. doi: 10.1002/cam4.118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Um S., Choi T.J., Kim H., Kim B.Y., Kim S.H., Lee S.K., Oh K.B., Shin J., Oh D.C. Ohmyungsamycins A and B: Cytotoxic and antimicrobial cyclic peptides produced by Streptomyces sp. from a volcanic island. J. Org. Chem. 2013;78:12321–12329. doi: 10.1021/jo401974g. [DOI] [PubMed] [Google Scholar]
- 187.Hur J., Jang J., Sim J., Son W.S., Ahn H.C., Kim T.S., Shin Y.H., Lim C., Lee S., An H., et al. Conformation-enabled total syntheses of ohmyungsamycins A and B and structural revision of ohmyungsamycin B. Angew. Chem. Int. Ed. 2018;57:3069–3073. doi: 10.1002/anie.201711286. [DOI] [PubMed] [Google Scholar]
- 188.Asolkar R.N., Freel K.C., Jensen P.R., Fenical W., Kondratyuk E.-J., Park T.P., Pezzuto J.M. Arenamides A-C, cytotoxic NF-κB inhibitors from the marine Actinomycete salinispora. J. Nat. Prod. 2009;72:396–402. doi: 10.1021/np800617a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Chandrasekhar S., Pavankumarreddy G., Sathish K. Total synthesis of arenamide A and its diastereomer. Tetrahedron Lett. 2009;50:6851–6854. doi: 10.1016/j.tetlet.2009.09.119. [DOI] [Google Scholar]
- 190.Miller E.D., Kauffman C.A., Jensen P.R., Fenical W. Piperazimycins: Cytotoxic hexadepsipeptides from a marine-derived bacterium of the genus Streptomyces. J. Org. Chem. 2007;72:323–330. doi: 10.1021/jo061064g. [DOI] [PubMed] [Google Scholar]
- 191.Li W., Gan J., Ma D. Total synthesis of piperazimycin a: A cytotoxic cyclic hexadepsipeptide. Angew. Chem. Int. Ed. 2009;48:8891–8895. doi: 10.1002/anie.200904603. [DOI] [PubMed] [Google Scholar]
- 192.Harrigan G.G., Harrigan B.L., Davidson B.S. Kailuins A-D, new cyclic acyldepsipeptides from cultures of a marine-derived bacterium. Tetrahedron. 1997;53:1577–1582. doi: 10.1016/S0040-4020(96)01136-2. [DOI] [Google Scholar]
- 193.Zhang H.L., Hua H.M., Pei Y.H., Yao X.S. Three new cytotoxic cyclic acylpeptides from marine Bacillus sp. Chem. Pharm. Bull. 2004;52:1029–1030. doi: 10.1248/cpb.52.1029. [DOI] [PubMed] [Google Scholar]
- 194.Rehman N.U., Abed R.M.M., Hussain H., Khan H.Y., Khan A., Khan A.L., Ali M., Al-Nasri A., Al-Harrasi K., Al-Rawahi A.N., et al. Anti-proliferative potential of cyclotetrapeptides from Bacillus velezensis RA5401 and their molecular docking on G-protein-coupled receptors. Microb. Pathog. 2018;123:419–425. doi: 10.1016/j.micpath.2018.07.043. [DOI] [PubMed] [Google Scholar]
- 195.Zhou H., He Y., Tian Y., Cong B., Yang H. Bacilohydrin A, a new cytotoxic cyclic lipopeptide of surfactins class produced by bacillus sp. sy27f from the indian ocean hydrothermal vent. Nat. Prod. Commun. 2019;14:141–146. doi: 10.1177/1934578X1901400137. [DOI] [Google Scholar]
- 196.Liu X.d., Gu K.b., Xia S.S., Zhang D.J., Li Y.G. Dolyemycins A and B, two novel cyclopeptides isolated from Streptomyces griseus subsp. griseus HYS31. J. Antibiot. 2018;71:838–845. doi: 10.1038/s41429-018-0071-x. [DOI] [PubMed] [Google Scholar]
- 197.Um S., Kim Y.J., Kwon H., Wen H., Kim S.H., Kwon H.C., Park S., Shin J., Oh D.C. Sungsanpin, a lasso peptide from a deep-sea Streptomycete. J. Nat. Prod. 2013;76:873–879. doi: 10.1021/np300902g. [DOI] [PubMed] [Google Scholar]
- 198.Zhou X., Huang H., Chen Y., Tan J., Song Y., Zou J., Tian X., Hua Y., Ju J. Marthiapeptide A, an anti-infective and cytotoxic polythiazole cyclopeptide from a 60 L scale fermentation of the deep sea-derived Marinactinospora thermotolerans SCSIO 00652. J. Nat. Prod. 2012;75:2251–2255. doi: 10.1021/np300554f. [DOI] [PubMed] [Google Scholar]
- 199.Zhang Y., Islam M.A., McAlpine S.R. Synthesis of the natural product marthiapeptide A. Org. Lett. 2015;17:5149–5151. doi: 10.1021/acs.orglett.5b02574. [DOI] [PubMed] [Google Scholar]
- 200.Kanoh K., Matsuo Y., Adachi K., Imagawa H., Nishizawa M., Shizuri Y. Mechercharmycins A and B, cytotoxic substances from marine-derived Thermoactinomyces sp. YM3-251. J. Antibiot. 2005;58:289–292. doi: 10.1038/ja.2005.36. [DOI] [PubMed] [Google Scholar]
- 201.Hernández D., Altuna M., Cuevas C., Aligué R., Albericio F., Álvarez M. Synthesis and activity of cytotoxic mechercharmycin A analogs. J. Med. Chem. 2008;51:5722–5730. doi: 10.1021/jm800513w. [DOI] [PubMed] [Google Scholar]
- 202.Matsuo Y., Kanoh K., Imagawa H., Adachi K., Nishizawa M., Shizuri Y. Urukthapelstatin A, a novel cytotoxic substance from marine-derived Mechercharimyces asporophorigenens YM11-542: II. Physico-chemical properties and structural elucidation. J. Antibiot. 2007;60:256–260. doi: 10.1038/ja.2007.31. [DOI] [PubMed] [Google Scholar]
- 203.Schwenk S., Ronco C., Oberheide A., Arndt H.-D. Biomimetic synthesis of urukthapelstatin A by Aza-Wittig ring contraction. J. Subst. Abuse Treat. 2016;13:287–288. doi: 10.1002/ejoc.201600994. [DOI] [Google Scholar]
- 204.Pan C.M., Lin C.C., Kim S.J., Sellers R.P., McAlpine S.R. Progress toward the synthesis of urukthapelstatin A and two analogues. Tetrahedron Lett. 2012;53:4065–4069. doi: 10.1016/j.tetlet.2012.05.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Lin C., Tantisantisom W., Mcalpine S.R. Total synthesis and biological activity of natural product urukthapelstatin A. Org. Lett. 2013;15:3574–3577. doi: 10.1021/ol401412v. [DOI] [PubMed] [Google Scholar]
- 206.Hamilton T.L., Bryant D.A., Macalady J.L. The role of biology in planetary evolution: Cyanobacterial primary production in low-oxygen Proterozoic oceans. Environ. Microbiol. 2016;18:325–340. doi: 10.1111/1462-2920.13118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Sainis I., Fokas D., Vareli K., Tzakos A.G., Kounnis V., Briasoulis E. Cyanobacterial cyclopeptides as lead compounds to novel targeted cancer drugs. Mar. Drugs. 2010;8:629–657. doi: 10.3390/md8030629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Janssen E.M. Cyanobacterial peptides beyond microcystins e A review on co-occurrence, toxicity, and challenges for risk assessment. Water Res. 2019;151:488–499. doi: 10.1016/j.watres.2018.12.048. [DOI] [PubMed] [Google Scholar]
- 209.Falconer I.R. Health Risk Assessment of Cyanobacterial (Blue-green Algal) Toxins in health risk assessment of cyanobacterial (blue-green algal) toxins in drinking water. Int. J. Environ. Res. Public Health. 2005;2:43–50. doi: 10.3390/ijerph2005010043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Dawson R.M. The toxicology of microcystins. Toxicon. 1998;36:953–962. doi: 10.1016/S0041-0101(97)00102-5. [DOI] [PubMed] [Google Scholar]
- 211.Lone Y., Bhide M., Koiri R.K. Microcystin-LR induced immunotoxicity in mammals. J. Toxicol. 2016;2016 doi: 10.1155/2016/8048125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Piyathilaka M.A.P.C., Pathmalal M.M., Tennekoon K.H., de Silva B.G.D.N.K., Samarakoon S.R., Chanthirika S. Microcystin-LR-induced cytotoxicity and apoptosis in human embryonic kidney and human kidney adenocarcinoma cell lines. Microbiology. 2015;161:819–828. doi: 10.1099/mic.0.000046. [DOI] [PubMed] [Google Scholar]
- 213.Physiology C. Chronic microcystin-LR exposure induces hepatocarcinogenesis via increased gankyrin in vitro and in vivo. Cell. Physiol. Biochem. 2018;49:1420–1430. doi: 10.1159/000493446. [DOI] [PubMed] [Google Scholar]
- 214.Kounnis V., Chondrogiannis G., Mantzaris M.D., Tzakos A.G., Fokas D., Papanikolaou N.A., Galani V., Sainis I., Briasoulis E. Microcystin lr shows cytotoxic activity against pancreatic cancer cells expressing the membrane OATP1B1 and OATP1B3 transporters. Anticancer Res. 2015;35:5857–5865. [PubMed] [Google Scholar]
- 215.Niedermeyer T.H.J., Daily A., Swiatecka-hagenbruch M., Moscow J.A. Selectivity and potency of microcystin congeners against OATP1B1 and OATP1B3 expressing cancer cells. PLoS ONE. 2014;9:e91476. doi: 10.1371/journal.pone.0091476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Monks N.R., Moscow J.A. Microcystins as Agents for Treatment of Cancer. US9006173B2. U.S. Patent. 2015 Apr 14;
- 217.Luesch H., Yoshida W.Y., Moore R.E., Paul V.J., Corbett T.H. Total structure determination of apratoxin A, a potent novel cytotoxin from the marine cyanobacterium Lyngbya majuscula. J. Am. Chem. Soc. 2001;123:5418–5423. doi: 10.1021/ja010453j. [DOI] [PubMed] [Google Scholar]
- 218.Luesch H., Chanda S.K., Raya R.M., DeJesus P.D., Orth A.P., Walker J.R., Belmonte J.C.I., Schultz P.G. A functional genomics approach to the mode of action of apratoxin A. Nat. Chem. Biol. 2006;2:158–167. doi: 10.1038/nchembio769. [DOI] [PubMed] [Google Scholar]
- 219.Liu Y., Law B.K., Luesch H. Apratoxin A aeversibly inhibits the secretory pathway by preventing cotranslational translocation. Mol. Pharmacol. 2009;76:91–104. doi: 10.1124/mol.109.056085. [DOI] [PubMed] [Google Scholar]
- 220.Paatero A.O., Kellosalo J., Dunyak B.M., Almaliti J., Gestwicki J.E., Gerwick W.H., Taunton J., Paavilainen V.O. Apratoxin Kills Cells by Direct Blockade of the Sec61 protein translocation channel. Cell Chem. Biol. 2016;23:561–566. doi: 10.1016/j.chembiol.2016.04.008. [DOI] [PubMed] [Google Scholar]
- 221.Huang K.-C., Chen Z., Jiang Y., Akare S., Kolber-Simonds D., Condon K., Agoulnik S., Tendyke K., Shen Y., Wu K.-M., et al. Apratoxin A shows novel pancreas-targeting activity through the binding of Sec 61. Mol. Cancer Ther. 2016;15:1208–1216. doi: 10.1158/1535-7163.MCT-15-0648. [DOI] [PubMed] [Google Scholar]
- 222.Luesch H., Yoshida W.Y., Moore E., Paul V.J. New apratoxins of marine cyanobacterial origin from Guam and Palau. Bioorg. Med. Chem. 2002;10:1973–1978. doi: 10.1016/S0968-0896(02)00014-7. [DOI] [PubMed] [Google Scholar]
- 223.Suyama T.L., Engene N., Wingerd J.S., Matainaho T. Apratoxin D, a potent cytotoxic cyclodepsipeptide from Papua New Guinea collections of the marine cyanobacteria Lyngbya majuscula and Lyngbya sordida. J. Nat. Prod. 2008;71:1099–1103. doi: 10.1021/np800121a. [DOI] [PubMed] [Google Scholar]
- 224.Matthew S., Schupp P.J., Luesch H. Apratoxin E, a Cytotoxic peptolide from a Guamanian collection of the marine cyanobacterium Lyngbya bouillonii. J. Nat. Prod. 2008;71:1113–1116. doi: 10.1021/np700717s. [DOI] [PubMed] [Google Scholar]
- 225.Tidgewell K., Engene N., Byrum T., Media J., Doi T., Valeriote F.A., Gerwick W.H. Evolved diversification of a modular natural product pathway: Apratoxins F and G, two cytotoxic cyclic depsipeptides from a Palmyra collection of Lyngbya bouillonii. ChemBioChem. 2012;11:1458–1466. doi: 10.1002/cbic.201000070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Han B., Gross H., Goeger D.E., Mooberry S.L., Gerwick W.H. Aurilides B and C, cancer cell toxins from a Papua New Guinea collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2006;69:572–575. doi: 10.1021/np0503911. [DOI] [PubMed] [Google Scholar]
- 227.Takase S., Kurokawa R., Arai D., Kanto K.K., Okino T., Nakao Y., Kushiro T., Yoshida M., Matsumoto K. A quantitative shRNA screen identifies ATP1A1 as a gene that regulates cytotoxicity by aurilide B. Sci. Rep. 2017;7:2002. doi: 10.1038/s41598-017-02016-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Huang X., Huang W., Li L., Sun X., Song S., Xu Q., Zhang L., Wei B., Deng X. Structure determinants of lagunamide A for anticancer activity and its molecular mechanism of mitochondrial apoptosis. Mol. Pharm. 2016;13:3756–3763. doi: 10.1021/acs.molpharmaceut.6b00564. [DOI] [PubMed] [Google Scholar]
- 229.Tripathi A., Fang W., Leong D.T., Tan L.T. Biochemical studies of the lagunamides, potent cytotoxic cyclic depsipeptides from the marine cyanobacterium Lyngbya majuscula. Mar. Drugs. 2012;10:1126–1137. doi: 10.3390/md10051126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Tripathi A., Puddick J., Prinsep M.R., Rottmann M., Ping K., Chen D.Y., Tong L. Phytochemistry lagunamide C, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Lyngbya majuscula. Phytochemistry. 2011;72:2369–2375. doi: 10.1016/j.phytochem.2011.08.019. [DOI] [PubMed] [Google Scholar]
- 231.Davies-Coleman M.T., Dzeha T.M., Gray C.A., Hess S., Pannell L.K., Hendricks D.T., Arendse C.E. Isolation of homodolastatin 16, a new cyclic depsipeptide from a Kenyan collection of Lyngbya majuscula. J. Nat. Prod. 2003;66:712–715. doi: 10.1021/np030014t. [DOI] [PubMed] [Google Scholar]
- 232.Moore R.E., Mooberry S.L., Yoshida W.Y., Luesch H., Paul V.J. Isolation, structure determination, and biological activity of lyngbyabellin A from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2002;63:611–615. doi: 10.1021/np990543q. [DOI] [PubMed] [Google Scholar]
- 233.Yokokawa F., Sameshima H., Shioiri T. Total synthesis of lyngbyabellin A, a potent cytotoxic metabolite from the marine cyanobacterium Lyngbya majuscula. Tetrahedron Lett. 2001;63:1440–1443. doi: 10.1016/S0040-4039(01)00678-5. [DOI] [Google Scholar]
- 234.Gogineni V., Hamann M.T. Marine natural product peptides with therapeutic potential: Chemistry, biosynthesis, and pharmacology. Biochim. Biophys. Acta Gen. Subj. 2018;1862:81–196. doi: 10.1016/j.bbagen.2017.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Choi H., Mevers E., Byrum T., Valeriote F.A., Gerwick W.H. Lyngbyabellins K-N from two Palmyra atoll collections of the marine cyanobacterium Moorea bouillonii. Eur. J. Org. Chem. 2012;2012:5141–5150. doi: 10.1002/ejoc.201200691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Taniguchi M., Nunnery J.K., Engene N., Esquenazi E., Byrum T., Dorrestein P.C., Gerwick W.H. Palmyramide A, a Cyclic depsipeptide from a Palmyra atoll collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010;73:393–398. doi: 10.1021/np900428h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Pereira A., Cao Z., Murray T.F., Gerwick W.H. Hoiamide A, a sodium channel activator of unusual architecture from a consortium of two Papua New Guinea cyanobacteria. Chem. Biol. 2009;16:893–906. doi: 10.1016/j.chembiol.2009.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Mangoni A., Shuman C.F., Engene N., Pereira A.R., Gerwick W.H., Cao Z., Murray T.F., Byrum T., Choi H., Matainaho T. The Hoiamides, structurally intriguing neurotoxic lipopeptides from Papua New Guinea marine cyanobacteria. J. Nat. Prod. 2010;73:1411–1421. doi: 10.1021/np100468n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Cao Z., Li X., Zou X., Greenwood M., Gerwick W.H., Murray T.F. Involvement of JNK and caspase activation in hoiamide A-induced neurotoxicity in neocortical neurons. Mar. Drugs. 2015;13:903–919. doi: 10.3390/md13020903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Nogle L.M., Marquez B.L., Gerwick W.H. Wewakazole, a novel cyclic dodecapeptide from a papua new guinea Lyngbya majuscula. Org. Lett. 2003;5:3–6. doi: 10.1021/ol026811k. [DOI] [PubMed] [Google Scholar]
- 241.Lopez J.A.V., Al-Lihaibi S.S., Alarif W.M., Abdel-Lateff A., Nogata Y., Washio K., Morikawa M., Okino T. Wewakazole B, a cytotoxic cyanobactin from the cyanobacterium Moorea producens collected in the Red Sea. J. Nat. Prod. 2016;79:1213–1218. doi: 10.1021/acs.jnatprod.6b00051. [DOI] [PubMed] [Google Scholar]
- 242.Nayani K., Hussaini S.D.A. Synthesis of cytotoxic cyanobactin, Wewakazole B. Tetrahedron Lett. 2017;58:1166–1169. doi: 10.1016/j.tetlet.2017.02.012. [DOI] [Google Scholar]
- 243.Inman M., Dexter H.L., Moody C.J. Total synthesis of the cyclic dodecapeptides wewakazole and wewakazole B. Org. Lett. 2017;19:3454–3457. doi: 10.1021/acs.orglett.7b01393. [DOI] [PubMed] [Google Scholar]
- 244.Marquez B.L., Watts K.S., Yokochi A., Roberts M.A., Verdier-Pinard P., Jimenez J.I., Hamel E., Scheuer P.J., Gerwick W.H. Structure and absolute stereochemistry of hectochlorin, a potent stimulator of actin assembly. J. Nat. Prod. 2002;65:866–871. doi: 10.1021/np0106283. [DOI] [PubMed] [Google Scholar]
- 245.Suntornchashwej S., Chaichit N., Isobe M., Suwanborirux K. Hectochlorin and morpholine derivatives from the Thai sea hare, Bursatella leachii. J. Nat. Prod. 2005;68:951–955. doi: 10.1021/np0500124. [DOI] [PubMed] [Google Scholar]
- 246.Ramaswamy A.V., Sorrels C.M., Gerwick W.H. Cloning and biochemical characterization of the hectochlorin biosynthetic gene cluster from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2007;70:1977–1986. doi: 10.1021/np0704250. [DOI] [PubMed] [Google Scholar]
- 247.Cetusic J.R.P., Green F.R., Graupner P.R., Oliver M.P. Total synthesis of hectochlorin. Org. Lett. 2002;4:1307–1310. doi: 10.1021/ol025604h. [DOI] [PubMed] [Google Scholar]
- 248.Simmons T.L., Nogle L.M., Media J., Valeriote F.A., Mooberry S.L., Gerwick W.H. Desmethoxymajusculamide C, a cyanobacterial depsipeptide with potent cytotoxicity in both cyclic and ring-opened forms. J. Nat. Prod. 2009;72:1011–1016. doi: 10.1021/np9001674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Tripathi A., Puddick J., Prinsep M.R., Lee P.P.F., Lik T.T. Hantupeptin A, a cytotoxic cyclic depsipeptide from a Singapore collection of Lyngbya majuscula. J. Nat. Prod. 2009;72:29–32. doi: 10.1021/np800448t. [DOI] [PubMed] [Google Scholar]
- 250.Tripathi A., Puddick J., Prinsep M.R., Lee P.P.F., Tan L.T. Hantupeptins B and C, cytotoxic cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. Phytochemistry. 2010;71:307–311. doi: 10.1016/j.phytochem.2009.10.006. [DOI] [PubMed] [Google Scholar]
- 251.Bonnard I., Rolland M., Francisco C., Banaigs B. Total structure and biological properties of laxaphycins A and B, cyclic lipopeptides from the marine cyanobacterium Lyngbya majuscula. Int. J. Pept. Res. Ther. 1997;4:289–292. doi: 10.1007/BF02442891. [DOI] [Google Scholar]
- 252.Bonnard I., Rolland M., Salmon J.M., Debiton E., Barthomeuf C., Banaigs B. Total structure and inhibition of tumor cell proliferation of laxaphycins. J. Med. Chem. 2007;50:1266–1279. doi: 10.1021/jm061307x. [DOI] [PubMed] [Google Scholar]
- 253.Cai W., Matthew S., Chen Q.Y., Paul V.J., Luesch H. Discovery of new A- and B-type laxaphycins with synergistic anticancer activity. Bioorganic Med. Chem. 2018;26:2310–2319. doi: 10.1016/j.bmc.2018.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Williams P.G., Yoshida W.Y., Moore R.E., Paul V.J. Isolation and structure determination of obyanamide, a novel cytotoxic cyclic depsipeptide from the marine cyanobacterium Lyngbya confervoides. J. Nat. Prod. 2002;65:29–31. doi: 10.1021/np0102253. [DOI] [PubMed] [Google Scholar]
- 255.Zhang W., Ding N., Li Y. Synthesis and biological evaluation of analogues of the marine cyclic depsipeptide obyanamide. J. Pept. Sci. 2011;17:533–539. doi: 10.1002/psc.1361. [DOI] [PubMed] [Google Scholar]
- 256.Kwan J.C., Rocca J.R., Abboud K.A., Paul V.J., Luesch H. Total structure determination of grassypetolide, a new marine cyanobacterical cytotoxin. Org. Lett. 2008;10:789–792. doi: 10.1021/ol702946d. [DOI] [PubMed] [Google Scholar]
- 257.Kwan J.C., Ratnayake R., Abboud K.A., Paul V.J., Luesch H. Grassypeptolides A-C, cytotoxic bis-thiazoline containing marine cyclodepsipeptides. J. Org. Chem. 2010;75:8012–8023. doi: 10.1021/jo1013564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Liu H., Liu Y., Wang Z., Xing X., Maguire A.R., Luesch H., Zhang H., Xu Z., Ye T. Total synthesis and biological evaluation of grassypeptolide A. Chem. A Eur. J. 2013;19:6774–6784. doi: 10.1002/chem.201203667. [DOI] [PubMed] [Google Scholar]
- 259.Thornburg C.C., Thimmaiah M., Shaala L.A., Hau A.M., Malmo J.M., Ishmael J.E., Youssef D.T.A., McPhail K.L. Cyclic Depsipeptides, grassypeptolides D and E and Ibu-epidemethoxylyngbyastatin 3, from a Red Sea Leptolyngbya cyanobacterium. J. Nat. Prod. 2011;74:1677–1685. doi: 10.1021/np200270d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Williams P.G., Yoshida W.Y., Quon M.K., Moore R.E., Paul V.J. The Structure of Palau’ amide, a potent cytotoxin from a species of the marine cyanobacterium Lyngbya. J. Nat. Prod. 2003;66:1545–1549. doi: 10.1021/np034001r. [DOI] [PubMed] [Google Scholar]
- 261.Sugiyama H., Watanabe A., Teruya T., Suenaga K. Synthesis of Palau’amide and its diastereomers: Confirmation of its stereostructure. Tetrahedron Lett. 2009;50:7343–7345. doi: 10.1016/j.tetlet.2009.10.059. [DOI] [Google Scholar]
- 262.Zou B., Long K., Ma D. Total synthesis and cytotoxicity studies of a cyclic depsipeptide with proposed structure of Palau’ amide. Org. Lett. 2005;7:2003–2006. doi: 10.1021/ol051685g. [DOI] [PubMed] [Google Scholar]
- 263.Medina R.A., Goeger D.E., Hills P., Mooberry S.L., Huang N., Romero L.I., Ortega-Barría E., Gerwick W.H., McPhail K.L. Coibamide A, a potent antiproliferative cyclic depsipeptide from the panamanian marine cyanobacterium Leptolyngbya sp. J. Am. Chem. Soc. 2008;130:6324–6325. doi: 10.1021/ja801383f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Hau A.M., Greenwood J.A., Löhr C.V., Serrill J.D., Proteau P.J., Ganley I.G., McPhail K.L., Ishmael J.E. Coibamide A induces mTOR-independent autophagy and cell death in human glioblastoma cells. PLoS ONE. 2013;8:e65250. doi: 10.1371/journal.pone.0065250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Serrill J.D., Wan X., Hau A.M., Jang H.S., Coleman D.J., Indra A.K., Alani A.W.G., McPhail K.L., Ishmael J.E. Coibamide A, a natural lariat depsipeptide, inhibits VEGFA/VEGFR2 expression and suppresses tumor growth in glioblastoma xenografts. Investig. New Drugs. 2016;34:24–40. doi: 10.1007/s10637-015-0303-x. [DOI] [PubMed] [Google Scholar]
- 266.Luesch H., Williams P.G., Yoshida W.Y., Moore R.E., Paul V.J. Ulongamides A-F, new β-amino acid-containing cyclodepsipeptides from Palauan collections of the marine cyanobacterium Lyngbya sp. J. Nat. Prod. 2002;65:996–1000. doi: 10.1021/np0200461. [DOI] [PubMed] [Google Scholar]
- 267.Alvarado C., Hernández G., Díaz E., Soano J.D., Vilchis-Reyes M.A., Martínez-Urbina M.A., Guzmán A. Synthesis of O-Me ulongamide B and O-Me ulongamide C, natural modified cyclodepsipeptides. Synth. Commun. 2013;43 doi: 10.1080/00397911.2011.618284. [DOI] [Google Scholar]
- 268.Alvarado C., Díaz E., Guzmán Á. Total synthesis of ulongamide A, a cyclic depsipeptide isolated from marine cyanobacteria Lyngbya sp. Tetrahedron Lett. 2007;48:603–607. doi: 10.1016/j.tetlet.2006.11.117. [DOI] [Google Scholar]
- 269.Williams P.G., Yoshida W.Y., Quon M.K., Moore R.E., Paul V.J. Ulongapeptin, a Cytotoxic cyclic depsipeptide from a Palauan marine cyanobacterium Lyngbya sp. J. Nat. Prod. 2003;4:651–654. doi: 10.1021/np030050s. [DOI] [PubMed] [Google Scholar]
- 270.Sueyoshi K., Kaneda M., Sumimoto S., Oishi S., Fujii N., Suenaga K., Teruya T. Odoamide, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Okeania sp. Tetrahedron. 2016;72:5472–5478. doi: 10.1016/j.tet.2016.07.031. [DOI] [Google Scholar]
- 271.Kaneda M., Kawaguchi S., Fujii N., Ohno H., Oishi S. Structure–activity relationship study on odoamide: Insights into the bioactivities of aurilide-family hybrid peptide–polyketides. ACS Med. Chem. Lett. 2018;9:365–369. doi: 10.1021/acsmedchemlett.8b00028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 272.Shih C., Teicher B.A. Cryptophycins: A novel class of potent antimitotic antitumor depsipeptides. Curr. Pharm. Des. 2001;7:1259–1276. doi: 10.2174/1381612013397474. [DOI] [PubMed] [Google Scholar]
- 273.Georg G.I. Halohydrin analogues of cryptophycin 1: Synthesis and biological activity. Bioorg. Med. Chem. Lett. 1998;8:1959–1962. doi: 10.1016/S0960-894X(98)00356-4. [DOI] [PubMed] [Google Scholar]
- 274.Cells D., Smith C.D., Zhang X., Mooberry S.L., Patterson G.M.L., Moore R.E. Cryptophycin: A new antimicrotubule agent active against drug-resistant cells. Cancer Res. 1994;54:3779–3784. [PubMed] [Google Scholar]
- 275.Panda D., Himes R.H., Moore R.E., Wilson L., Jordan M.A. Mechanism of action of the unusually potent microtubule inhibitor. Biochemistry. 1997;21:12948–12953. doi: 10.1021/bi971302p. [DOI] [PubMed] [Google Scholar]
- 276.Kerksiek K., Mejillano M.R., Schwartz R.E., Georg G.I., Himes R.H. Interaction of cryptophycin 1 with tubulin and microtubules. FEBS Lett. 1995;377:59–61. doi: 10.1016/0014-5793(95)01271-0. [DOI] [PubMed] [Google Scholar]
- 277.Bai R., Schwartz R.E., Kepler J.A., Pettit G.R., Hamel E. Characterization of the interaction of cryptophycin 1 with tubulin: Binding in the vinca domain, competitive inhibition of dolastatin 10 binding, and an unusual aggregation reaction. Cancer Res. 1996;56:4398–4406. [PubMed] [Google Scholar]
- 278.Brenda J., Richard A., Frederick A. Cryptophycin 1 cellular levels and effects in vitro using L1210 cells. Investig. New. Drugs. 1999;16:199–204. doi: 10.1023/a:1006148127527. [DOI] [PubMed] [Google Scholar]
- 279.Ilson L.E.W. Antiproliferative mechanism of action of cryptophycin-52: Kinetic stabilization of microtubule dynamics by high-affinity binding to microtubule ends. Proc. Natl. Acad. Sci. USA. 1998;95:9313–9318. doi: 10.1073/pnas.95.16.9313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280.Taori K., Paul V.J., Luesch H. Structure and activity of largazole, a potent antiproliferative agent from the Floridian marine cyanobacterium Symploca sp. J. Am. Chem. Soc. 2008;130:1806–1807. doi: 10.1021/ja7110064. [DOI] [PubMed] [Google Scholar]
- 281.Bradner J.E., Schreiber S.L., West N., Williams R.M., Taunton J., Bowers A. Total synthesis and biological mode of action of largazole: A potent class I histone deacetylase inhibitor. J. Am. Chem. Soc. 2008;130:11219–11222. doi: 10.1021/ja8033763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Zeng X., Yin B., Hu Z., Liao C., Liu J. Total synthesis and biological evaluation of largazole and derivatives. Org. Lett. 2010;12:1368–1371. doi: 10.1021/ol100308a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283.Byeon S., Law B.K., Liu Y., Hong J., Salvador L.A., Ying Y., Kwan J.C., Luesch H. Anticolon cancer activity of largazole, a marine-derived tunable histone deacetylase inhibitor. J. Pharmacol. Exp. Ther. 2010;335:351–361. doi: 10.1124/jpet.110.172387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 284.Wang W., Parker S.J., Zhang G., Kuchta R.D., Ungermannova D., Nasveschuk C.G., Quade B., Liu X., Phillips A.J. Largazole and its derivatives selectively inhibit ubiquitin activating enzyme (E1) PLoS ONE. 2012;7:e29208. doi: 10.1371/journal.pone.0029208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 285.Hong J., Luesch H. Largazole: From discovery to broad-spectrum therapy. Nat. Prod. Rep. 2012;29:449–456. doi: 10.1039/c2np00066k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286.Linington R.G., Edwards D.J., Shuman C.F., McPhail K.L., Matainaho T., Gerwick W.H. Symplocamide A, a potent cytotoxin and chymotrypsin inhibitor from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2008;71:22–27. doi: 10.1021/np070280x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 287.Stolze S.C., Meltzer M., Ehrmann M., Kaiser M. Solid phase total synthesis of the 3-amino-6-hydroxy-2-piperidone (Ahp) cyclodepsipeptide and protease inhibitor symplocamide A. Chem. Commun. 2010;46:8857–8859. doi: 10.1039/c0cc02889d. [DOI] [PubMed] [Google Scholar]
- 288.Williams P.G., Yoshida W.Y., Moore R.E., Paul V.J. Tasipeptins A and B: New cytotoxic depsipeptides from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2003;66:620–624. doi: 10.1021/np020582t. [DOI] [PubMed] [Google Scholar]
- 289.Köcher S., Rey J., Bongard J., Tiaden A.N., Meltzer M., Richards P.J., Ehrmann M., Kaiser M. Tailored Ahp-cyclodepsipeptides as potent non-covalent serine protease inhibitors. Angew. Chem. Int. Ed. 2017;56:8555–8558. doi: 10.1002/anie.201701771. [DOI] [PubMed] [Google Scholar]
- 290.Salvador L.A., Biggs J.S., Paul V.J., Luesch H. Veraguamides A-G, cyclic hexadepsipeptides from a dolastatin 16-producing cyanobacterium Symploca cf. hydnoides from Guam. J. Nat. Prod. 2011;74:917–927. doi: 10.1021/np200076t. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Sabry O.M., Goeger D.E., Gerwick W.H. Biologically active new metabolites from a Florida collection of Moorea producens. Nat. Prod. Res. 2017;25:555–561. doi: 10.1080/14786419.2016.1207074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Boudreau P.D., Byrum T., Liu W.T., Dorrestein P.C., Gerwick W.H. Viequeamide a, a cytotoxic member of the kulolide superfamily of cyclic depsipeptides from a marine button cyanobacterium. J. Nat. Prod. 2012;75:1560–1570. doi: 10.1021/np300321b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293.Wang D., Song S., Tian Y., Xu Y., Miao Z., Zhang A. Total synthesis of the Marine cyclic depsipeptide viequeamide A. J. Nat. Prod. 2013 doi: 10.1021/np4001027. [DOI] [PubMed] [Google Scholar]
- 294.Lücking D.L.H.R. Fungal diversity revisited: 2.2 to 3.8 million species. Neurol. India. 2015;63:1004–1005. doi: 10.1128/microbiolspec.FUNK-0052-2016.Correspondence. [DOI] [Google Scholar]
- 295.Keller N.P., Turner G., Bennett J.W. Fungal secondary metabolism-From biochemistry to genomics. Nat. Rev. Microbiol. 2005 doi: 10.1038/nrmicro1286. [DOI] [PubMed] [Google Scholar]
- 296.Du L., Risinger A.L., King J.B., Powell D.R., Cichewicz R.H. A potent HDAC inhibitor, 1-alaninechlamydocin, from a Tolypocladium sp. induces G2/M cell cycle arrest and apoptosis in MIA PaCa-2 cells. J. Nat. Prod. 2014;77:1753–1757. doi: 10.1021/np500387h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297.Guo H., Kreuzenbeck N.B., Otani S., Garcia-Altares M., Dahse H.M., Weigel C., Aanen D.K., Hertweck C., Poulsen M., Beemelmanns C. Pseudoxylallemycins A-F, cyclic tetrapeptides with rare allenyl modifications isolated from Pseudoxylaria sp. X802: A competitor of fungus-growing termite cultivars. Org. Lett. 2016;18:3338–3341. doi: 10.1021/acs.orglett.6b01437. [DOI] [PubMed] [Google Scholar]
- 298.Zhang Y., Liu S., Liu H., Liu X., Che Y. Cycloaspeptides F and G, cyclic pentapeptides from a Cordyceps-colonizing isolate of Isaria farinosa. J. Nat. Prod. 2009;72:1364–1367. doi: 10.1021/np900205m. [DOI] [PubMed] [Google Scholar]
- 299.Liu Y., Wang M., Wang D., Li X., Wang W., Lou H., Yuan H. Malformin A1 promotes cell death through induction of apoptosis, necrosis and autophagy in prostate cancer cells. Cancer Chemother. Pharmacol. 2016;77:63–75. doi: 10.1007/s00280-015-2915-4. [DOI] [PubMed] [Google Scholar]
- 300.Park S.Y., Oh H.H., Park Y.L., Yu H.M., Myung D.S., Cho S.B., Lee W.S., Park D., Joo Y.E. Malformin A1 treatment alters invasive and oncogenic phenotypes of human colorectal cancer cells through stimulation of the p38 signaling pathway. Int. J. Oncol. 2017;51:959–966. doi: 10.3892/ijo.2017.4070. [DOI] [PubMed] [Google Scholar]
- 301.Notarte K., Nakao Y., Yaguchi T., Bungihan M., Suganuma K., dela Cruz T.E. Trypanocidal activity, cytotoxicity and histone modifications induced by malformin A1 isolated from the marine-derived fungus Aspergillus tubingensis IFM 63452. Mycosphere. 2017;8:111–120. doi: 10.5943/mycosphere/8/1/10. [DOI] [Google Scholar]
- 302.Ma Y.M., Liang X.A., Zhang H.C., Liu R. Cytotoxic and antibiotic cyclic pentapeptide from an endophytic Aspergillus tamarii of Ficus carica. J. Agric. Food Chem. 2016;64:3789–3793. doi: 10.1021/acs.jafc.6b01051. [DOI] [PubMed] [Google Scholar]
- 303.Song H.H., Lee H.S., Lee C. A new cytotoxic cyclic pentadepsipeptide, neo-N-methylsansalvamide produced by Fusarium solani KCCM90040, isolated from potato. Food Chem. 2011;126:472–478. doi: 10.1016/j.foodchem.2010.11.023. [DOI] [Google Scholar]
- 304.Lee H.S., Phat C., Choi S.U., Lee C. Synergistic effect of a novel cyclic pentadepsipeptide, neo N-methylsansalvamide, and paclitaxel on human multidrug resistance cancer cell lines. Anticancer Drugs. 2013;24:455–460. doi: 10.1097/CAD.0b013e32835f060d. [DOI] [PubMed] [Google Scholar]
- 305.Weber D., Erosa G., Sterner O., Anke T. Cylindrocyclin A, a new cytotoxic cyclopeptide from Cylindrocarpon sp. J. Antibiot. 2006;59:495–499. doi: 10.1038/ja.2006.69. [DOI] [PubMed] [Google Scholar]
- 306.Ványolós A., Dékány M., Kovács B., Krámos B., Bérdi P., Zupkó I., Hohmann J., Béni Z. Gymnopeptides A and B, Cyclic octadecapeptides from the mushroom Gymnopus fusipes. Org. Lett. 2016;18:2688–2691. doi: 10.1021/acs.orglett.6b01158. [DOI] [PubMed] [Google Scholar]
- 307.Pan Z., Wu C., Wang W., Cheng Z., Yao G., Liu K., Li H., Fang L., Su W. Total synthesis and stereochemical assignment of gymnopeptides A and B. Org. Lett. 2017;19:4420–4423. doi: 10.1021/acs.orglett.7b01742. [DOI] [PubMed] [Google Scholar]
- 308.Schloß S., Hackl T., Herz C., Lamy E., Koch M., Rohn S., Maul R. Detection of a toxic methylated derivative of phomopsin A produced by the legume-infesting fungus Diaporthe toxica. J. Nat. Prod. 2017;80:1930–1934. doi: 10.1021/acs.jnatprod.6b00662. [DOI] [PubMed] [Google Scholar]
- 309.Chen C.H., Lang G., Mitova M.I., Murphy A.C., Cole A.L.J., Din L.B., Blunt J.W., Munro M.H.G. Pteratides I-IV, new cytotoxic cyclodepsipeptides from the Malaysian basidiomycete Pterula sp. J. Org. Chem. 2006;71:7947–7951. doi: 10.1021/jo060843u. [DOI] [PubMed] [Google Scholar]
- 310.He F., Bao J., Zhang X.Y., Tu Z.C., Shi Y.M., Qi S.H. Asperterrestide A, a cytotoxic cyclic tetrapeptide from the marine-derived fungus Aspergillus terreus SCSGAF0162. J. Nat. Prod. 2013;76:1182–1186. doi: 10.1021/np300897v. [DOI] [PubMed] [Google Scholar]
- 311.Ohsawa K., Sugai M., Zhang L., Masuda Y., Yoshida M., Doi T. Total synthesis and structural revision of cyclotetrapeptide asperterrestide A. J. Org. Chem. 2019 doi: 10.1021/acs.joc.9b00526. [DOI] [PubMed] [Google Scholar]
- 312.Isaka M., Srisanoh U., Lartpornmatulee N., Boonruangprapa T. ES-242 derivatives and cycloheptapeptides from Cordyceps sp. strains BCC 16173 and BCC 16176. J. Nat. Prod. 2007;70:1601–1604. doi: 10.1021/np070357h. [DOI] [PubMed] [Google Scholar]
- 313.Rukachaisirikul V., Chantaruk S., Tansakul C., Saithong S., Chaicharernwimonkoon L., Pakawatchai C., Isaka M., Intereya K. A cyclopeptide from the insect pathogenic fungus Cordyceps sp. BCC 1788. J. Nat. Prod. 2006;69:305–307. doi: 10.1021/np050433l. [DOI] [PubMed] [Google Scholar]
- 314.Chen Z., Song Y., Chen Y., Huang H., Zhang W., Ju J. Cyclic heptapeptides, cordyheptapeptides C-E, from the marine-derived fungus Acremonium persicinum SCSIO 115 and their cytotoxic activities. J. Nat. Prod. 2012;75:1215–1219. doi: 10.1021/np300152d. [DOI] [PubMed] [Google Scholar]
- 315.Dahiya R., Gautam H. Solution phase synthesis and bioevaluation of cordyheptapeptide B. Bull. Pharm. Res. 2011;1:1–10. [Google Scholar]
- 316.Oh D.C., Jensen P.R., Fenical W. Zygosporamide, a cytotoxic cyclic depsipeptide from the marine-derived fungus Zygosporium masonii. Tetrahedron Lett. 2006;47:8625–8628. doi: 10.1016/j.tetlet.2006.08.113. [DOI] [Google Scholar]
- 317.Wang Y., Zhang F., Zhang Y., Liu J.O., Ma D. Synthesis and antitumor activity of cyclodepsipeptide zygosporamide and its analogues. Bioorganic Med. Chem. Lett. 2008;18:4385–4387. doi: 10.1016/j.bmcl.2008.06.066. [DOI] [PubMed] [Google Scholar]
- 318.Belofsky G.N., Jensen P.R., Fenical W. Sansalvamide: A new cytotoxic cyclic depsipeptide produced by a marine fungus of the genus Fusarium. Tetrahedron Lett. 1999;40:2913–2916. doi: 10.1016/S0040-4039(99)00393-7. [DOI] [Google Scholar]
- 319.Heiferman M.J., Salabat M.R., Ujiki M.B., Strouch M.J., Cheon E.C., Silverman R.B., Bentrem D.J. Sansalvamide induces pancreatic cancer growth arrest through changes in the cell cycle. Anticancer Res. 2010;30:73–78. [PubMed] [Google Scholar]
- 320.Hwang Y., Rowley D., Rhodes D., Gertsch J., Fenical W., Bushman F. Mechanism of inhibition of a poxvirus topoisomerase by the marine natural product sansalvamide A. Mol. Pharmacol. 1999;55:1049–1053. doi: 10.1124/mol.55.6.1049. [DOI] [PubMed] [Google Scholar]
- 321.Chen L., Zheng M., Zhou Y., Liu H., Jiang H. Ionic-liquid-supported total synthesis of sansalvamide A peptide. Synth. Commun. 2008;38:239–248. doi: 10.1080/00397910701749633. [DOI] [Google Scholar]
- 322.Carroll C.L., Johnston J.V.C., Kekec A., Brown J.D., Parry E., Cajica J., Medina I., Cook K.M., Corral R., Pan P.S., et al. Synthesis and cytotoxicity of novel sansalvamide A derivatives. Org. Lett. 2005;7:3481–3484. doi: 10.1021/ol051161g. [DOI] [PubMed] [Google Scholar]
- 323.Pan P.S., Vasko R.C., Lapera S.A., Johnson V.A., Sellers R.P., Lin C.C., Pan C.M., Davis M.R., Ardi V.C., McAlpine S.R. A comprehensive study of sansalvamide A derivatives: The structure-activity relationships of 78 derivatives in two pancreatic cancer cell lines. Bioorganic Med. Chem. 2009;17:5806–5825. doi: 10.1016/j.bmc.2009.07.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 324.Jiang W., Ye P., Chen C.T.A., Wang K., Liu P., He S., Wu X., Gan L., Ye Y., Wu B. Two novel hepatocellular carcinoma cycle inhibitory cyclodepsipeptides from a hydrothermal vent crab-associated fungus Aspergillus clavatus C2WU. Mar. Drugs. 2013;11:4761–4772. doi: 10.3390/md11124761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 325.Chettu S.K., Konidena L.N.S., Korupolu R.B., Rao N.S.K., Doddipalla R., Gandham H.B., Guduru R. Ring opening of benzoxazinones: An improved and efficient synthesis of clavatustides A & B. Tetrahedron Lett. 2017;58:3418–3420. doi: 10.1016/j.tetlet.2017.07.059. [DOI] [Google Scholar]
- 326.Liao Y., Hong X., Yang J., Qin J.J., Zhang B., Lin J., Shao Z., Wang W. Structure elucidation of a novel cyclic tripeptide from the marine-derived fungus Aspergillus ochraceopetaliformis DSW-2. Nat. Prod. Res. 2020:1–7. doi: 10.1080/14786419.2020.1869969. [DOI] [PubMed] [Google Scholar]
- 327.Yu Z., Lang G., Kajahn I., Schmaljohann R., Imhoff J.F. Scopularides A and B, cyclodepsipeptides from a marine sponge-derived fungus, Scopulariopsis brevicaulis. J. Nat. Prod. 2008;71:1052–1054. doi: 10.1021/np070580e. [DOI] [PubMed] [Google Scholar]
- 328.Cruz L.J., Insua M.M., Baz J.P., Trujillo M., Rodriguez-Mias R.A., Oliveira E., Giralt E., Albericio F., Cañedo L.M. IB-01212, a new cytotoxic cyclodepsipeptide isolated from the marine fungus Clonostachys sp. ESNA-A009. J. Org. Chem. 2006;71:3335–3338. doi: 10.1021/jo051600p. [DOI] [PubMed] [Google Scholar]
- 329.Luque-Ortega J.R., Cruz L.J., Albericio F., Rivas L. The antitumoral depsipeptide IB-01212 kills Leishmania through an apoptosis-like process involving intracellular targets. Mol. Pharm. 2010;7:1608–1617. doi: 10.1021/mp100035f. [DOI] [PubMed] [Google Scholar]
- 330.Nabika R., Oishi S., Misu R., Ohno H., Fujii N. Synthesis of IB-01212 by multiple N-methylations of peptide bonds. Bioorganic Med. Chem. 2014;22:6156–6162. doi: 10.1016/j.bmc.2014.08.036. [DOI] [PubMed] [Google Scholar]
- 331.Cruz L.J., Francesch A., Cuevas C., Albericio F. Synthesis and structure-activity relationship of cytotoxic marine cyclodepsipeptide IB-01212 analogues. ChemMedChem. 2007;2:1076–1084. doi: 10.1002/cmdc.200700025. [DOI] [PubMed] [Google Scholar]
- 332.Gu W., Cueto M., Jensen P.R., Fenical W., Silverman R.B. Microsporins A and B: New histone deacetylase inhibitors from the marine-derived fungus Microsporum cf. gypseum and the solid-phase synthesis of microsporin A. Tetrahedron. 2007;63:6535–6541. doi: 10.1016/j.tet.2007.04.025. [DOI] [Google Scholar]
- 333.Mukherjee J.P., Sil S., Chattopadhyay S.K. A modular approach to cyclic tetrapeptides related to histone deacetylase inhibition: Synthesis of epi-microsporin A. Tetrahedron Lett. 2016;57:739–742. doi: 10.1016/j.tetlet.2016.01.005. [DOI] [Google Scholar]
- 334.Swaroop P.S., Tripathy S., Jachak G., Reddy D.S. Efforts towards the synthesis of microsporin B: Ready access to both the enantiomers of the key amino acid fragment. Tetrahedron Lett. 2014;55:4777–4779. doi: 10.1016/j.tetlet.2014.06.093. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.