

Abstract
目的
基于网络药理学分析滋阴方治疗排卵障碍性不孕症的作用机制并设计临床对照试验观察滋阴方对该病的治疗疗效。
方法
利用TCMSP、PubChem、Genecards、STRING、Swiss Target Prediction、UniProt等数据库,获得滋阴方所有作用靶点和排卵障碍性不孕症相关的靶点。构建靶点蛋白质相互作用网络,进行GO生物学过程与KEGG通路富集分析,使用Cytoscape 3.6.0构建滋阴方治疗排卵障碍性不孕症的“中药-有效成分-靶点-通路”网络。同时设计临床对照试验,试验组及对照组均以克罗米芬联合人绝经期促性腺激素进行促排卵治疗,试验组在此基础上添加滋阴方,通过观察两组HMG用量、卵泡发育天数、扳机日单个优势卵泡雌二醇水平(E2)水平、周期排卵率、未破裂卵泡黄素化综合征发生率、周期妊娠率等指标,验证滋阴方的临床有效性。
结果
滋阴方共有22个活性成分,354个作用靶点,排卵障碍性不孕症靶点791个,取交集获得滋阴方治疗排卵障碍性不孕症作用靶点96个。根据Degree找出关键靶点为SRC、MAPK1、HSP90AA1、MAPK3、PTPN11、ESR1、AKT1、EGFR、NR3C1、KNG1。GO生物学过程富集分析显示,滋阴方治疗排卵障碍性不孕症主要集中在类固醇激素介导的信号通路、氧化、还原、凋亡等过程。KEGG通路富集分析表明富集最显著的信号通路PI3K-Akt信号通路、Rap1信号通路、HIF-1信号通路、雌激素信号通路、卵巢类固醇生成、类固醇激素生物合成。临床试验显示滋阴方能减少人绝经期促性腺激素用量,缩短卵泡发育天数,提高扳机日单个优势卵泡E2水平、周期排卵率及妊娠率。
结论
滋阴方中多种有效活性成分可以通过不同的途径发挥其促进卵泡发育的作用,从而提高排卵障碍不孕症患者排卵率及临床妊娠率,具有良好的临床应用前景。
Keywords: 网络药理, 排卵障碍, 不孕症, 滋阴方, 临床观察
Abstract
Objective
To explore the mechanisms of Ziyin recipe for treatment of ovulatory infertility based on network pharmacology analysis and evaluate the clinical efficacy of this recipe.
Methods
TCMSP, PubChem, Genecards, String, Swiss Target Prediction, and Uniprot databases were searched to identify all the action targets of Ziyin recipe and ovulatory infertility to construct the PPI network. Gene ontology (GO) and KEGG pathway enrichment analyses were performed and the "TCM-active ingredient-target-pathway" network was constructed using Cytoscape 3.6.0. We also designed a controlled clinical trial to verify the clinical effectiveness of Ziyin recipe. The patients were randomized into 2 groups to receive treatment with Western medicine including CC and HMG (control group) and additional treatment with Ziyin recipe, and the dosage of GN, follicular development days, E2 level of a single dominant follicle on trigger day, ovulation rate, the rate of LUFS, and clinical pregnancy rate were compared between the two groups.
Results
We identified 22 active ingredients and 354 targets of action in Ziyin recipe, 791 targets of ovulatory infertility, and 96 common targets of action shared by Ziyin recipe and ovulatory infertility. According to Degree, the key targets included SRC, MAPK1, HSP90aa1, MAPK3, PTPN11, ESR1, Akt1, EGFR, NR3C1 and KNG1. Enrichment analysis of GO biological process showed that Ziyin recipe mainly focused on steroid hormonemediated signaling pathway, oxidation, reduction, and apoptosis. The most significantly enriched signaling pathways included PI3K-Akt signaling pathway, RAP1 signaling pathway, HIF-1 signaling pathway, estrogen signaling pathway, ovarian steroid production, and steroid hormone biosynthesis. The results of the clinical trial showed that Ziyin recipe significantly reduced the dose of GN, accelerated follicle development, increased E2 level of single dominant follicle on the trigger day, and increased the cycle ovulation rate and pregnancy rate.
Conclusion
The therapeutic effects of Ziyin recipe are probably mediated through different pathways to promote follicle development, thus improving ovulation rate and clinical pregnancy rate of infertile patients with ovulatory disorders.
Keywords: network pharmacology, ovulation disorder, infertility, Ziyin recipe, clinical observation
女性无避孕性生活至少12月而未孕,称为不孕症,在我国发病率达7%~10%,其中排卵障碍占25%~ 35%[1]。临床上常通过促排卵方法促进卵泡成熟,常用药物有枸橼酸克罗米芬(CC)、来曲唑(LE)、促性腺激素(Gn)等,然而运用不当可能会引起卵巢过度刺激综合征(OHSS),多胎妊娠等。另外,即使运用促排卵药物,但仍有卵泡发育迟缓,或卵泡不发育等情况发生。对高龄女性促排卵药物的过度使用,可能增加乳腺癌和子宫内膜癌的潜在风险[2]。因此寻求有效、安全的促排卵方案显得尤为重要。滋阴方是导师谈勇教授经验方(专利号:ZL200410041406.0),是基于国医大师夏桂成教授“调周”理论而拟方,具体用药:当归、白芍、干地黄、山茱萸、菟丝子,经过多年临床实践,疗效确切。长期的临床观察证实其促进卵泡发育疗效较好[3-4]。相关实验研究滋阴方含药血清可明显增加颗粒细胞E2的分泌量, 降低P的分泌量,认为滋阴方可通过调节颗粒细胞的分泌功能进而改善类PCOS的卵巢功能[5]。然其治疗机制尚未完全阐明,仍有待继续明确。
网络药理学方法在中药复方研究领域中的应用较多, 为其现代化研究提供了新的思路与方法[6]。本研究拟通过网络药理学的分析,探讨其组方治疗该病可能机制,以期为后续动物实验提供方向,为临床提供理论依据。于此同时,设计临床对照试验,通过观察滋阴方对卵泡发育和周期结局的影响,进一步验证其临床有效性。
1. 资料和方法
1.1. 滋阴方活性成分的收集及筛选
利用TCMSP数据库(<a href="http://lsp.nwu.edu.cn/tcmsp.php" target="_blank">http://lsp.nwu.edu.cn/tcmsp.php</a>)收集滋阴方(当归、白芍、干地黄、山茱萸、菟丝子)的主要活性成分。筛选条件为:药物相似性(DL)>0.18,口服生物利用度(OB)≥30%作为筛选阈值。
利用PubChem(<a href="https://pubchem.nehi.nlm.nih.gov" target="_blank">https://pubchem.nehi.nlm.nih.gov</a>)对其分子结构进行确证,获得其Canonical SMILES字符串(SMILES)。
1.2. 滋阴方作用靶点的收集
我们利用化学相似性,通过Swiss Target Prediction(<a href="http://www.swisstargetprediction.ch/" target="_blank">http://www.swisstargetprediction.ch/</a>),将查询到的Canonical SMILES字符串输入进行靶点对接,获得上述中药的潜在靶点基因,即为活性成分作用靶点。用Excel整理药物活性成分-作用靶点。
1.3. 疾病靶点预测
我们运用GeneCards数据库(<a href="http://www.genecards.org/" target="_blank">http://www.genecards.org/</a>),以“ovulation failure infertility”为关键词进行搜索,收集与排卵障碍性不孕症相关的基因靶点信息,并导入UniProt数据库(<a href="https://www.uniprot.or-g/" target="_blank">https://www.uniprot.or-g/</a>),经名称-ID配对获得这些基因靶点的相应UniProt ID。将主要活性成分靶点和疾病靶点取交集,获得滋阴方治疗排卵障碍性不孕症的靶点。用excel整理药物活性成分-疾病靶点,导入Cytoscape3.6.0软件,进行网络风格化显示。
1.4. 构建靶基因PPI网络
我们使用STRING数据库(<a href="http://www.string-db.org/" target="_blank">http://www.string-db.org/</a>)进行蛋白-蛋白相互作用(PPI)分析,导入筛选出共同靶点,将蛋白种类设置为“homo sapiens”,最低相互作用阈值设为“highest confidence”(0.9),构建PPI网络图。导入Cytoscape软件,进行NetworkAnalyzer分析,然后根据Degree值筛选关键靶点。
1.5. GO、KEGG分析
我们通过DAVID(<a href="https://david.ncifcrf.gov/" target="_blank">https://david.ncifcrf.gov/</a>)网站,借助“Functional Annotation Tool”对共同靶基因进行GO富集分析,以<italic>P</italic> < 0.05和FDR < 0.05为标准筛选。并对潜在靶点进行KEGG分析,同样以<italic>P</italic> < 0.05和FDR < 0.05为筛选标准,通过R软件绘制气泡图展示结果。
1.6. 构建中药-有效成分-靶点-KEGG通路网络
我们利用cytoscape3.6.0软件分别构建中药-有效成分、有效成分-靶点、靶点-信号通路网络,利用“Merge”功能进行合并,构建中药-有效成分-靶点- KEGG通路网络。
1.7. 临床验证
1.7.1. 临床资料
选取2019年1~12月于江苏省中医院生殖医学科门诊就诊的排卵障碍性不孕症患者,共60例。排卵障碍性不孕症诊断标准:(1)无避孕性生活至少12个月而未孕,其中从未妊娠者称为原发性不孕;曾有过妊娠而后未避孕连续1年不孕者称为继发性不孕;(2)基础体温测定:单相或不典型双相型,温差小于0.3 ℃,缓慢上升持续时间小于10 d;(3)卵泡监测:周期第10天开始,B超连续监测卵泡,晚期或尿促黄体生成素(LH)峰值日优势卵泡平均直径 < 18 mm,或有成熟卵泡而无排卵征象;(4)血清孕酮低于黄体期水平。其中(1)必备,(2)~(4)项中符合其中2项者即可诊断。(参考第8版《妇产科学》《不孕不育专家推荐诊疗方案》、《人类辅助生殖技术规范》2003版)。
1.7.2. 纳入标准
年龄:21~35岁;符合上述诊断标准;至少3个自然周期B超监测无优势卵泡发育,即无排卵征象;基础促卵泡生成素(FSH):5~10 mU/mL;1年内子宫输卵管造影示至少有一侧输卵管通畅;受试者同意并签署知情同意书。
1.7.3. 排除标准
先天性生理缺陷或畸形、染色体异常、生殖器官器质性病变(子宫肌瘤、子宫腺肌症、卵巢囊肿、生殖器肿瘤及癌前病变等);高泌乳素血症、甲功异常;合并心血管、肝、肾、造血系统等严重原发疾病、精神病者;已知对所有药物过敏者;正在使用口服避孕药、糖皮质激素等激素药物者;正在参加其他药物临床试验者;配偶少弱畸精症者。
1.7.4. 剔除和脱落病例标准
患者未按规定用药,无法判定疗效或资料不全等影响疗效者;治疗过程中发生严重不良反应者。
1.7.5. 治疗方法
1.7.5.1. 分组方法
将60例病人按随机数字表法分为2组:试验组和对照组各30人。两组均采用西药促排治疗,试验组在此基础上于经后期加用中药滋阴方。本试验通过南京中医药大学附属医院伦理委员会审批(批件号:2018NL-120-03)。
1.7.5.2. 服药方法
(1)试验组促排卵方法:于经周第3~ 5天口服CC 1~2片/d,连服5 d,于经周第8天开始B超下监测卵泡,并根据卵泡生长情况,酌情添加促性腺激素(HMG)(乐宝得,规格75 U/支)。当优势卵泡直径≥18 mm时,肌注绒毛膜促性腺激素(HCG)5000~ 10 000 U,嘱患者24 h后同房,48 h后B超下确定卵泡是否排出,若卵泡排出后则开始口服地屈孕酮健黄体治疗,地屈孕酮服用方法为2次/d,2片/次,连服14 d。2周后测血清β-HCG,若β-HCG结果阳性提示妊娠,保胎治疗。若β-HCG阴性,则停服地屈孕酮,继续下1周期治疗。滋阴方服用方法:于月经干净后即服用经后滋阴方,当卵泡平均直径≥18 mm(肌注HCG日)时停用中药;(2)对照组:不服用中药,余治疗同试验组。两组均以3月经周期为观察期限,若妊娠,则终止试验。
1.7.6. 观察指标
HMG用量、卵泡发育天数、扳机日单个优势卵泡E2水平、周期排卵率、未破裂卵泡黄素化综合征(LUFS)发生率、周期妊娠率。
1.8. 统计学方法
应用SPSS23.0统计软件分析,计量资料用t检验,以均数±标准差表示,计数资料用卡方检验。P < 0.05为差异具有统计学意义。
2. 结果
2.1. 滋阴方活性成分
通过TCMSP数据库,结合“DL>0.18”与“OB >30%”筛选出滋阴方中可被口服吸收的48个活性成分,去除重复后得到40个活性成分。通过PubChem查找40个成分的SMILES字符串,得到有SMILES字符串的活性成分24个(表 1)。
1.
滋阴方活性成分列表
List of active ingredients of nourishing Yin prescription
| Mol ID | Molecule name | Molecular formula | OB (%) | DL | Traditional Chinese medicine in Ziyin recipe |
| OB: Oral bioavailability; DL: Drug likeness. | |||||
| MOL000358 | Beta-sitosterol | C29H50O | 36.91 | 0.75 | Chinese Angelica、White Paeony Root、Asiatic Cornelian Cherry Fruit、Cuscutae Semen |
| MOL000449 | Stigmasterol | C29H48O | 43.83 | 0.76 | Chinese Angelica、Rehmannia Glutinosa、Asiatic Cornelian Cherry Fruit |
| MOL001919 | 3S, 5R, 8R, 9R, 10S, 14S)-3, 17-dihydroxy-4, 4, 8, 10, 14-pentamethyl-2, 3, 5, 6, 7, 9-hexahydro-1H-cyclopenta[a]phenanthrene-15, 16-dione | C22H30O4 | 43.56 | 0.53 | White Paeony Root |
| MOL000211 | Mairin | C30H48O3 | 55.38 | 0.78 | White Paeony Root |
| MOL000359 | Sitosterol | C29H50O | 36.91 | 0.75 | White Paeony Root、Rehmannia Glutinosa、Asiatic Cornelian Cherry Fruit |
| MOL000422 | Kaempferol | C15H10O6 | 41.88 | 0.24 | White Paeony Root、Cuscutae Semen |
| MOL000492 | (+)-catechin | C15H14O6 | 54.83 | 0.24 | White Paeony Root |
| MOL001494 | Mandenol | C20H36O2 | 42 | 0.19 | Asiatic Cornelian Cherry Fruit |
| MOL001495 | Ethyl linolenate | C20H34O2 | 46.1 | 0.2 | Asiatic Cornelian Cherry Fruit |
| MOL001771 | poriferast-5-en-3beta-ol | C29H50O | 36.91 | 0.75 | Asiatic Cornelian Cherry Fruit |
| MOL002879 | Diop | C24H38O4 | 43.59 | 0.39 | Asiatic Cornelian Cherry Fruit |
| MOL002883 | Ethyl oleate (NF) | C20H38O2 | 32.4 | 0.19 | Asiatic Cornelian Cherry Fruit |
| MOL003137 | Leucanthoside | C22H22O11 | 32.12 | 0.78 | Asiatic Cornelian Cherry Fruit |
| MOL005481 | 2, 6, 10, 14, 18-pentamethylicosa-2, 6, 10, 14, 18-pentaene | C25H42 | 33.4 | 0.24 | Asiatic Cornelian Cherry Fruit |
| MOL005486 | 3, 4-Dehydrolycopen-16-al | C40H52O | 46.64 | 0.49 | Asiatic Cornelian Cherry Fruit |
| MOL005503 | Cornudentanone | C22H34O5 | 39.66 | 0.33 | Asiatic Cornelian Cherry Fruit |
| MOL005530 | Hydroxygenkwanin | C16H12O6 | 36.47 | 0.27 | Asiatic Cornelian Cherry Fruit |
| MOL001558 | Sesamin | C20H18O6 | 56.55 | 0.83 | Cuscutae Semen |
| MOL000184 | NSC63551 | C29H48O | 39.25 | 0.76 | Cuscutae Semen |
| MOL000354 | Isorhamnetin | C16H12O7 | 49.6 | 0.31 | Cuscutae Semen |
| MOL005043 | Campest-5-en-3beta-ol | C28H48O | 37.58 | 0.71 | Cuscutae Semen |
| MOL005440 | Isofucosterol | C29H48O | 43.78 | 0.76 | Cuscutae Semen |
| MOL000953 | CLR | C27H46O | 37.87 | 0.68 | Cuscutae Semen |
| MOL000098 | Quercetin | C15H10O7 | 46.43 | 0.28 | Cuscutae Semen |
2.2. 滋阴方活性成分靶点预测结果
借助Swiss Target Prediction网站,得到每个成分对应的基因靶点,导出数据,共1152个成分靶点,去重后剩余354个靶点。需要指出的是MOL000492没有预测到相关靶点,而MOL005486预测所得的靶点Probability均为0,故将二者予以剔除,剩余22个活性成分及354个靶点。
2.3. 排卵障碍性不孕症靶点预测结果
通过GeneCards数据库检索排卵障碍性不孕症,共得到789个与排卵障碍性不孕症相关的基因,作为疾病候选靶点基因。并提取各基因和排卵障碍性不孕症的相关度,将基因名导入UniProt数据库,经名称-ID配对后获得Uniprot ID 791个。将主要活性成分靶点和疾病靶点取交集,获得滋阴方治疗排卵障碍性不孕症的靶点96个(表 2)。应用R软件绘制疾病靶点与药物成分靶点韦恩图(图 1)。
2.
共同靶点基因列表
Common target genes list
| Uniprot ID | Gene name | Relevance score | Uniprot ID | Gene name | Relevance score |
| P11511 | CYP19A1 | 56.91 | P11473 | VDR | 8.8 |
| P10275 | AR | 49.95 | P28845 | HSD11B1 | 8.58 |
| P05093 | CYP17A1 | 36.45 | P02753 | RBP4 | 8.28 |
| P01375 | TNF | 32.92 | P80365 | HSD11B2 | 7.93 |
| P06401 | PGR | 31.18 | Q96PN6 | ADCY10 | 7.92 |
| P13569 | CFTR | 29.72 | Q07869 | PPARA | 7.84 |
| P04278 | SHBG | 29.12 | P10415 | BCL2 | 7.47 |
| P31749 | AKT1 | 28.75 | P08183 | ABCB1 | 7.45 |
| P06213 | INSR | 26.24 | P00749 | PLAU | 7.31 |
| P03372 | ESR1 | 26.05 | P30530 | AXL | 7.31 |
| P10721 | KIT | 20.16 | P28482 | MAPK1 | 7.21 |
| Q92731 | ESR2 | 19.57 | P32418 | SLC8A1 | 6.94 |
| P35228 | NOS2 | 17.58 | P10635 | CYP2D6 | 6.76 |
| Q06124 | PTPN11 | 15.73 | P55055 | NR1H2 | 6.63 |
| P49327 | FAS | 14.91 | P04626 | ERBB2 | 6.39 |
| P37231 | PPARG | 14.2 | P01042 | KNG1 | 6.36 |
| P08253 | MMP2 | 14.02 | P35869 | AHR | 6.27 |
| P14780 | MMP9 | 13.94 | P30559 | OXTR | 6.04 |
| P00533 | EGFR | 13.72 | P06493 | CDK1 | 5.67 |
| P08246 | ELANE | 12.4 | Q16678 | CYP1B1 | 5.3 |
| P18405 | SRD5A1 | 12.33 | P00747 | PLG | 5.23 |
| P00734 | F2 | 12.15 | P12931 | SRC | 5.2 |
| P14416 | DRD2 | 12 | P17948 | FLT1 | 5.1 |
| P35354 | PTGS2 | 11.88 | P04035 | HMGCR | 4.72 |
| P60568 | IL2 | 11.47 | P23219 | PTGS1 | 4.69 |
| P04150 | NR3C1 | 11.44 | O95718 | ESRRB | 4.6 |
| P08069 | IGF1R | 11.22 | P11413 | G6PD | 4.49 |
| P05164 | MPO | 11.13 | P11712 | CYP2C9 | 4.35 |
| P03956 | MMP1 | 9.53 | Q9NPH5 | NOX4 | 4.22 |
| P02766 | TTR | 9.5 | Q14432 | PDE3A | 4.11 |
| P35968 | KDR | 9.14 | P22303 | ACHE | 4.04 |
| Q00987 | MDM2 | 8.83 | P24666 | ACP1 | 3.94 |
| Q15303 | ERBB4 | 3.75 | O14684 | PTGES | 2.37 |
| P07900 | HSP90AA1 | 3.66 | Q02763 | TEK | 2.27 |
| P45452 | MMP13 | 3.37 | P11474 | ESRRA | 2.17 |
| P14060 | HSD3B1 | 3.3 | P08185 | SERPINA6 | 2.17 |
| Q8NER1 | TRPV1 | 3.23 | P14061 | HSD17B1 | 2.08 |
| Q99527 | GPER1 | 3.22 | O75469 | NR1I2 | 2.05 |
| P21397 | MAOA | 3.07 | P30307 | CDC25C | 2.01 |
| P27361 | MAPK3 | 3.04 | P10276 | RARA | 1.83 |
| P17252 | PRKACA | 3 | Q00534 | CDK6 | 1.73 |
| Q8WTV0 | SCARB1 | 2.87 | Q07820 | MCL1 | 1.63 |
| O00519 | FAAH | 2.82 | P34998 | CRHR1 | 1.63 |
| Q05469 | LIPE | 2.66 | P43116 | PTGER2 | 1.4 |
| P35372 | OPRM1 | 2.63 | Q03181 | PPARD | 1.34 |
| P34972 | CNR2 | 2.61 | P42330 | AKR1C3 | 0.62 |
| P33261 | CYP2C19 | 2.59 | P49286 | MTNR1B | 0.3 |
| P50281 | MMP14 | 2.47 | P48039 | MTNR1A | 0.3 |
1.
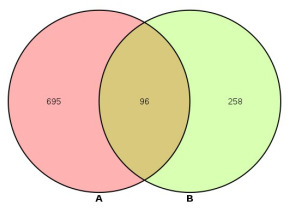
维恩图
Venn diagram.
2.4. 有效成分-疾病靶点网络
用Excel表格整理“成分-靶点”对,导入Cytoscape 3.6.0软件,进行网络风格化显示(图 2)。该网络共有节点118个,边405条,其中22个节点代表滋阴方活性成分,96个节点代表滋阴方治疗排卵障碍性不孕症的靶点。
2.
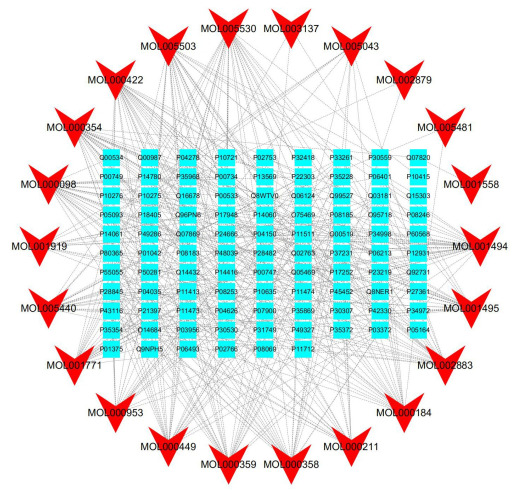
有效成分-疾病靶点网络
Network of active ingredient-disease target.
2.5. 构建PPI网络
我们将筛选出的96个共同靶点导入STRING数据库进行分析,将蛋白种类设置为“homo sapiens”,最低相互作用阈值设为“highest confidence”(0.9),获得PPI网络图(图 3A)。并导入Cytoscape软件,进行NetworkAnalyzer分析,然后根据Degree值筛选关键靶点,其中度值≥9的蛋白有10个,分别为SRC、MAPK1、HSP90AA1、MAPK3、PTPN11、ESR1、AKT1、EGFR、NR3C1、KNG1(图 3B)。
3.
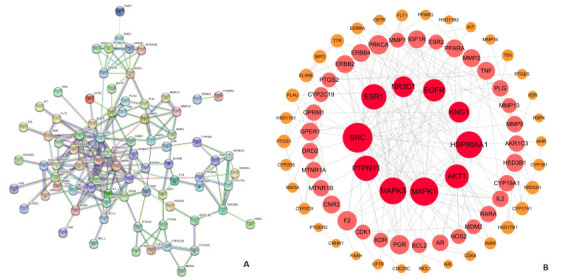
PPI网络图
PPI network diagram.A: STRING; B: Cytoscape.
2.6. GO分析
将96个共同靶基因导入DAVID网站,获得72条GO Term,其中关于生物学过程48条,细胞组成9条,分子功能15条。我们主要分析关键靶点生物学过程,发现主要集中在类固醇激素介导的信号通路、RNA聚合酶Ⅱ启动子的转录起始、ERK1和ERK2级联阳性调节、氧化还原过程、蛋白磷酸化的正调控、凋亡过程负调控、MAP激酶活性的正向调节、细胞内受体信号通路、跨膜受体蛋白酪氨酸激酶信号通路、类固醇代谢过程、对雌激素反应、RNA聚合酶Ⅱ启动子的转录的正调控、炎性反应负调控、对缺氧的反应、正向调控凋亡过程、正向调控细胞生长、糖皮质激素生物合成的过程、雄激素生物合成的过程、类固醇生物合成的过程等方面(表 3)。
3.
共同靶基因GO富集分析
GO enrichment analysis of common target genes
| Term | Count | P-Value | FDR |
| GO enrichment analysis of common target genes. | |||
| GO: 0043401~steroid hormone mediated signaling pathway | 12 | 1.38E-14 | 2.30E-11 |
| GO: 0006367~transcription initiation from RNA polymerase Ⅱ promoter | 14 | 2.62E-12 | 4.37E-09 |
| GO: 0070374~positive regulation of ERK1 and ERK2 cascade | 14 | 1.58E-11 | 2.64E-08 |
| GO: 0055114~oxidation-reduction process | 21 | 7.57E-11 | 1.26E-07 |
| GO: 0045429~positive regulation of nitric oxide biosynthetic process | 9 | 8.79E-11 | 1.47E-07 |
| GO: 0001934~positive regulation of protein phosphorylation | 12 | 1.21E-10 | 2.03E-07 |
| GO: 0043066~negative regulation of apoptotic process | 17 | 4.29E-09 | 7.17E-06 |
| GO: 0043406~positive regulation of MAP kinase activity | 8 | 3.70E-08 | 6.18E-05 |
| GO: 0030522~intracellular receptor signaling pathway | 7 | 6.26E-08 | 1.05E-04 |
| GO: 0007169~transmembrane receptor protein tyrosine kinase signaling pathway | 9 | 6.31E-08 | 1.06E-04 |
| GO: 0008202~steroid metabolic process | 7 | 1.35E-07 | 2.26E-04 |
| GO: 0043627~response to estrogen | 7 | 1.66E-06 | 0.002771787 |
| GO: 0045944~positive regulation of transcription from RNA polymerase Ⅱ promoter | 20 | 1.71E-06 | 0.002854344 |
| GO: 0050728~negative regulation of inflammatory response | 7 | 5.24E-06 | 0.00875908 |
| GO: 0001666~response to hypoxia | 9 | 5.42E-06 | 0.009060475 |
| GO: 0043065~positive regulation of apoptotic process | 11 | 6.86E-06 | 0.011465575 |
| GO: 0030307~positive regulation of cell growth | 7 | 7.49E-06 | 0.012524465 |
| GO: 0006704~glucocorticoid biosynthetic process | 4 | 1.39E-05 | 0.02328036 |
| GO: 0006702~androgen biosynthetic process | 4 | 2.71E-05 | 0.045354075 |
| GO: 0006694~steroid biosynthetic process | 5 | 2.94E-05 | 0.049082965 |
2.7. KEGG通路富集
将96个共同靶基因导入DAVID网站进行KEGG通路富集分析,以P < 0.05,FDR < 0.05为筛选标准,共得到富集通路11条(图 4、表 4),包括癌症通路、癌症中的蛋白聚糖、PI3K-Akt信号通路、Rap1信号通路、HIF-1信号通路、雌激素信号通路、卵巢类固醇生成、前列腺癌、膀胱癌、类固醇激素生物合成、神经胶质瘤。
4.
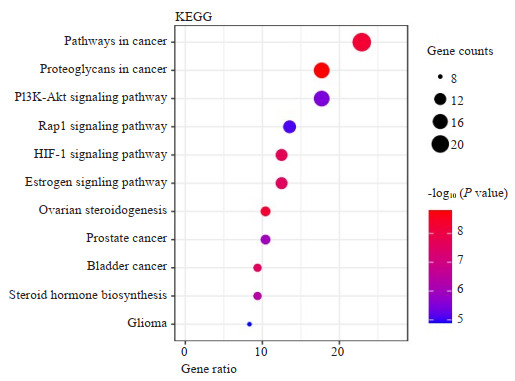
KEGG富集通路分析
KEGG enrichment pathway analysis.
4.
共同靶点KEGG通路分析
KEGG pathway analysis of common targets
| Term | Count | Genes | P | FDR |
| hsa05200: Pathways in cancer | 22 |
Q03181, P17252, P35354, P14780, P27361, P35228, P10721, P10275, P08069, P10276, P07900, Q00987, P04626, P08253, P28482, Q00534, P37231, P00533, P43116, P31749, P10415, P03956 |
5.38E-09 | 6.66E-06 |
| hsa05205: Proteoglycans in cancer | 17 |
P12931, P17252, P01375, P27361, P14780, Q15303, P08069, P03372, P35968, Q00987, P04626, Q06124, P08253, P28482, P00749, P00533, P31749 |
1.61E-09 | 1.99E-06 |
| hsa04151: PI3K-Akt signaling pathway | 17 |
P17252, P27361, P17948, Q07820, P10721, P08069, Q02763, P60568, P07900, P35968, Q00987, P28482, Q00534, P06213, P00533, P31749, P10415 |
3.18E-06 | 0.003940273 |
| hsa04015: Rap1 signaling pathway | 13 |
P10721, P08069, Q02763, P35968, P12931, P17252, P28482, P17948, P27361, P14416, P06213, P00533, P31749 |
7.80E-06 | 0.009655768 |
| hsa04066: HIF-1 signaling pathway | 12 |
P08069, Q02763, P04626, P17252, P28482, P27361, P17948, P06213, P35228, P00533, P31749, P10415 |
1.75E-08 | 2.17E-05 |
| hsa04915: Estrogen signaling pathway | 12 |
P07900, P03372, Q99527, P12931, Q92731, P08253, P28482, P27361, P14780, P35372, P00533, P31749 |
2.43E-08 | 3.01E-05 |
| hsa04913: Ovarian steroidogenesis | 10 |
P14060, P08069, P35354, P11511, P05093, Q16678, P06213, P42330, Q8WTV0, P14061 |
5.39E-09 | 6.67E-06 |
| hsa05215: Prostate cancer | 10 |
P10275, P08069, P07900, Q00987, P04626, P28482, P27361, P00533, P31749, P10415 |
1.02E-06 | 0.001260658 |
| hsa05219: Bladder cancer | 9 |
P12931, Q00987, P04626, P08253, P28482, P27361, P14780, P00533, P03956 |
2.43E-08 | 3.00E-05 |
| hsa00140: Steroid hormone biosynthesis | 9 |
P14060, P80365, P11511, P18405, P05093, P28845, Q16678, P42330, P14061 |
4.12E-07 | 5.09E-04 |
| hsa05214: Glioma | 8 |
P08069, Q00987, P17252, P28482, P27361, Q00534, P00533, P31749 |
1.24E-05 | 0.015289218 |
2.8. 中药-有效成分-靶点-通路网络及临床观察统计
为了阐明滋阴方作用于排卵障碍性不孕症的潜在机制,我们构建了由滋阴方中药-有效成分-靶点- KEGG通路组成的复杂网络(图 5)。该网络显示了多个通路中的相互作用,在该网络中,有134个节点(5个中药节点、22个有效成分节点、96个排卵障碍相关靶点节点和11条富集通路节点)和574条边。图 4和表 4显示了这11条信号通路的详细信息。通过该网络分析,表明了滋阴方中的5种中药协同作用,通过不同的信号通路途径治疗排卵障碍性不孕症,60例患者均完成临床观察,但在治疗过程中有2位对照组患者强烈要求服用中药,故最终试验组32人,对照组28人。其中试验组周期数84个,对照组72个。试验组HMG用量少于对照组(P < 0.05),卵泡发育天数短于对照组(P < 0.05),班级日单个优势卵泡E2水平高于对照组(P < 0.05),排卵率高于对照组(P < 0.05),妊娠率高于对照组(P < 0.05),而LUFS率未见明显差异(P>0.05)。临床观察统计结果见表 5。
5.
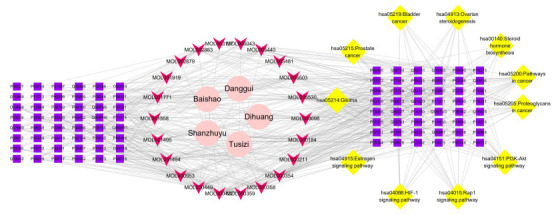
中药-有效成分-靶点-通路网络图
Network diagram of Chinese medicine-active ingredient-target-pathway.
5.
两组促排卵情况及周期结局比较
Comparation of ovulation induction and cycle outcome between experimental group and control group
| Observation index (unit) | Experimental group | Control group | P |
| Number of cycles (n) | 84 | 72 | |
| Dosage of HMG (U) | 295.54±108.34 | 392.19±116.72 | 0.000 |
| Follicular development days (days) | 15.14±2.17 | 16.47±2.03 | 0.000 |
| E2 level of single dominant follicle on trigger day (ng/L) | 315.86±47.11 | 297.63±32.54 | 0.006 |
| Cyclic ovulation rate | 82.1% (69/84) | 66.7% (48/72) | 0.026 |
| Rate of LUFS | 13.1% (11/84) | 25% (18/72) | 0.057 |
| Rate of pregnancy | 50% (16/32) | 25% (7/28) | 0.047 |
3. 讨论
西医治疗主要运用各种促排药物进行促排,中药在该病的治疗中亦发挥着重要的作用[7]。《素问·阴阳应象大论》“阴阳者,天地之道也”。国医大师夏桂成教授立足于中医“天人相应”的观点,重视阴阳消长转化在女性生殖节律的核心作用环节,经行过后即进入经后期,以阴长为前提,通过阴药的运用滋阴养血,奠定卵泡发育的基础[8]。导师经验方滋阴方组成主要为阴药,当归、白芍、干地黄为四物汤主药,重在补血养阴,山茱萸入肝肾经,重在补肝肾之阴,菟丝子为阳中之阴药,在于阳中补阴,正如张景岳“善补阴者, 必于阳中求阴, 则阴得阳升而泉源不竭”,共奏助养卵泡之效,以助妊娠。本方旨在养阴,重在补养动态之阴,若不加以助阳,纯以滋阴,则不易达到阴长趋向中高水平的要求,致后期卵泡发育停滞及经间期重阴转阳不利,致排卵障碍的发生。
本研究结果显示,滋阴方活性成分22个,具体如文中表 1。经查阅文献,发现Stigmasterol、beta-sitosterol与17β-雌二醇的结构有很高的相似性,因此可作为雌激素激动剂[9]。通过微量滴定板将beta-sitosterol生物转化为雄烯二酮(一种关键的药物类固醇中间体)[10]。kaempferol、quercetin、isorhamnetin都属于生物类黄酮,可抑制KGN细胞中雌激素的生物合成[11]。实验证明kaempferol可以通过PI3K/AKT通路促进原始卵泡活化和细胞增殖,降低体外培养的羊腔前卵泡的DNA碎片[12]。研究分析显示quercetin对猪和牛培养的颗粒细胞的影响,结果表明quercetin对可能具有物种特异性的基本卵巢细胞功能(增殖,凋亡和激素释放)具有直接作用[13]。有研究旨在测试膳食quercetin对热应激雌性兔用[13]。有研究旨在测试膳食quercetin对热应激雌性兔。
上述有效成分对应354个作用靶点,其中与排卵障碍性不孕症相关的共同靶点96个,其中包括SRC、MAPK1、HSP90AA1、MAPK3、PTPN11、ESR1、AKT1、EGFR、NR3C1、KNG1,为关键靶标。研究发现SRC蛋白分布在原始卵泡的卵质膜和颗粒细胞膜中,并且表达水平随原始卵泡的生长而增加,通过PI3K-PKC-ERK1/2途径在调节原始卵泡的活化和生长[15]。另有研究结果表明MAPK3/1蛋白在某些原始卵泡和所有生长的卵泡中表达[16]。用MAPK3/1信号抑制剂培养3 d可显著减少活化卵泡的数量,并伴有明显减少的颗粒细胞增殖和卵母细胞凋亡的增加。研究发现[17],母体持续暴露于环境化合物污染对胎儿卵巢发育有影响,导致原始卵泡储备减少,同时观察到卵巢蛋白的改变主要在胎儿卵巢蛋白和热休克家族的几个成员(HSPA4L,HSP90AA1和HSF1)。Idrees等[18]提及PTPN11在牛卵母细胞的成熟和胚胎发育中起着重要作用。雌激素是调节生育能力的必需激素。细胞对雌激素的反应是由雌激素受体α(ESR1)和雌激素受体β(ESR2)介导的。在小鼠和大鼠模型中,ESR1的破坏会导致雄性和雌性不育[19]。AKT1在卵巢支持细胞谱系中的分子作用仍然知之甚少。有学者为了深入了解其在此类细胞中的功能,通过耗尽了AKT1在鼠原代颗粒细胞中的表达,证实了AKT1参与这种卵巢细胞类型的代谢,细胞凋亡,细胞周期或细胞骨架动力学的调节[20]。有研究综述了EGF网络在卵母细胞发育和排卵级联调节中的作用,尤其着重于EGF网络对卵母细胞发育能力的影响,总结了EGF网络的急性上调是排卵级联反应的重要组成部分,因为它会将LH信号从卵泡周围传递到卵母细胞复合体[21]。EGF信号下调,抑制卵母细胞减数分裂成熟并同时提供减数分裂诱导信号。EGF网络还控制静止卵母细胞的转录翻译,这是卵母细胞能力不可或缺的过程。有研究观察到颗粒细胞中的NR3C1免疫染色从原始卵泡到窦卵泡逐渐增加,所有卵泡类别的卵母细胞和颗粒细胞中都存在NR3C1,除了在原始卵泡的颗粒细胞中。PCOS以女性不育和代谢异常为特征,是最常见的内分泌疾病之一[22]。有研究对PCOS患者中蛋白质变化的进行分析,共纳入43例PCOS患者和30例健康女性,证实了5种显著降低的蛋白质,其中包括KNG1[23]。
经过GO分析,发现其生物学过程主要集中在类固醇激素介导的信号通路、氧化、还原、凋亡等过程。根据KEGG分析的结果,与排卵障碍相关的通路包括PI3KAkt信号通路、Rap1信号通路、HIF-1信号通路、雌激素信号通路、卵巢类固醇生成、类固醇激素生物合成。缺氧诱导因子是细胞生长的主要调节因子。研究发现HIF是参与FSH对大鼠支持细胞增殖的调节[24]。研究证明HIF-1α在哺乳动物卵巢的卵泡生长发育过程中以细胞和阶段特异性方式表达[25]。通过免疫组织化学和蛋白质印迹分析、颗粒细胞培养等实验证明HIF-1α介导的信号通路可能是体内调节哺乳动物卵巢卵泡生长和发育的重要机制。一篇综述中提到从原始卵泡停滞到激活生长到卵母细胞成熟,哺乳动物雌性种系中基因组完整性的保留对于确保生殖成功至关重要[26]。由于卵母细胞是在出生前形成的,并且可能保持多年休眠状态,因此必须对防御机制进行监测并保持良好状态,PTEN/PI3K/Akt是控制原始卵泡募集和生长的主要信号途径,该途径还有助于细胞生长,存活和代谢,并有助于维持基因组完整性。雌激素信号通路对正常卵泡发育至关重要,研究表明在体外成熟培养基中添加E2后,可以看到卵丘扩展和随后卵母细胞发育的增加[27]。
我们通过临床试验发现,试验组的HMG用量少,卵泡发育天数短,扳机日单个优势卵泡E2水平高,周期排卵率及妊娠率亦高,较对照组均有统计学差异(P < 0.05)。因此对于排卵障碍性不孕症的患者,我们可以在西药促排治疗的同时加用中药滋阴方,帮助孕卵生长,提高妊娠率。
综上,滋阴方中多种有效活性可以通过不同的途径发挥其促进卵泡发育的作用。中医药治疗排卵障碍性不孕症有独特优势,多种中药配伍使用,协同增效,相对于西药促排卵其副作用明显减少,同时排卵率及妊娠率明显优于单纯西药,具有良好的临床应用前景。
Biography
罗倩倩,在读博士研究生,E-mail: 1530832700@qq.com
Funding Statement
国家自然科学基金(81873333);江苏省科教强卫工程“中医妇科临床医学中心”(YXZXB2016006)
Supported by National Natural Science Foundation of China (81873333)
Contributor Information
罗 倩倩 (Qianqian LUO), Email: 1530832700@qq.com.
谈 勇 (Yong TAN), Email: a1530832700@163.com.
夏 桂成 (Guicheng XIA), Email: x2000x@163.com.
References
- 1.王 泽华, 王 艳丽. 妇产科学. 北京: 人民卫生出版社; 2019. [王泽华, 王艳丽. 妇产科学[M]. 8版. 北京: 人民卫生出版社, 2019.] [Google Scholar]
- 2.胡 琳莉, 黄 国宁, 孙 海翔, et al. 促排卵药物使用规范(2016) 生殖医学杂志. 2017;26(4):302–7. doi: 10.3969/j.issn.1004-3845.2017.04.002. [胡琳莉, 黄国宁, 孙海翔, 等. 促排卵药物使用规范(2016)[J]. 生殖医学杂志, 2017, 26(4): 302-7.] [DOI] [Google Scholar]
- 3.唐 培培, 周 阁, 谈 勇, et al. 滋阴方联合标准长方案对输卵管因素不孕患者体外受精结局的影响. https://www.cnki.com.cn/Article/CJFDTOTAL-BXYY201904140.htm. 中华中医药杂志. 2019;34(4):1825–7. [唐培培, 周阁, 谈勇, 等. 滋阴方联合标准长方案对输卵管因素不孕患者体外受精结局的影响[J]. 中华中医药杂志, 2019, 34(4): 1825-7.] [Google Scholar]
- 4.唐 培培, 赵 娟, 谈 勇, et al. 滋阴方联合微刺激方案对卵巢低反应干预的临床研究. https://www.cnki.com.cn/Article/CJFDTOTAL-ANJS201805010.htm. 安徽科技学院学报. 2018;32(5):44–7. [唐培培, 赵娟, 谈勇, 等. 滋阴方联合微刺激方案对卵巢低反应干预的临床研究[J]. 安徽科技学院学报, 2018, 32(5): 44-7.] [Google Scholar]
- 5.童 星丽, 赵 娟, 谈 勇. 奠基方含药血清对类多囊卵巢综合征大鼠卵巢颗粒细胞分泌功能影响的研究. https://www.cnki.com.cn/Article/CJFDTOTAL-SHAN201302010.htm. 山西中医学院学报. 2013;14(2):21–3. [童星丽, 赵娟, 谈勇. 奠基方含药血清对类多囊卵巢综合征大鼠卵巢颗粒细胞分泌功能影响的研究[J]. 山西中医学院学报, 2013, 14(2): 21-3.] [Google Scholar]
- 6.李 秀云, 章 新友, 吴 地尧, et al. 中药复方网络药理学研究的文献分析. 中国药房. 2021;32(2):182–9. [李秀云, 章新友, 吴地尧, 等. 中药复方网络药理学研究的文献分析[J]. 中国药房, 2021, 32(2): 182-9.] [Google Scholar]
- 7.施 冬青. 排卵障碍性不孕症中医治疗研究进展. https://www.cnki.com.cn/Article/CJFDTOTAL-GYKX202012007.htm. 中国医药科学. 2020;10(12):42–5. [施冬青. 排卵障碍性不孕症中医治疗研究进展[J]. 中国医药科学, 2020, 10(12): 42-5.] [Google Scholar]
- 8.夏 桂成. 夏桂成实用中医妇科学. 北京: 中国中医药出版社; 2009. [夏桂成. 夏桂成实用中医妇科学[M]. 北京: 中国中医药出版社, 2009.] [Google Scholar]
- 9.Ju YH, Clausen LM, Allred KF, et al. Beta-sitosterol, beta-sitosterol glucoside, and a mixture of beta-sitosterol and beta-sitosterol glucoside modulate the growth of estrogen-responsive breast cancer cells in vitro and in ovariectomized athymic mice. http://www.ncbi.nlm.nih.gov/pubmed/15113961. J Nutr. 2004;134(5):1145–51. doi: 10.1093/jn/134.5.1145. [Ju YH, Clausen LM, Allred KF, et al. Beta-sitosterol, beta-sitosterol glucoside, and a mixture of beta-sitosterol and beta-sitosterol glucoside modulate the growth of estrogen-responsive breast cancer cells in vitro and in ovariectomized athymic mice[J]. J Nutr, 2004, 134(5): 1145-51.] [DOI] [PubMed] [Google Scholar]
- 10.Marques MPC, Fernandes P. Β-sitosterol bioconversion to androstenedione in microtiter plates. http://link.springer.com/protocol/10.1007/978-1-4939-7183-1_12. Methods Mol Biol. 2017;1645(4):167–76. doi: 10.1007/978-1-4939-7183-1_12. [Marques MPC, Fernandes P. Β-sitosterol bioconversion to androstenedione in microtiter plates[J]. Methods Mol Biol, 2017, 1645(4): 167-76.] [DOI] [PubMed] [Google Scholar]
- 11.Lu DF, Yang LJ, Wang F, et al. Inhibitory effect of luteolin on estrogen biosynthesis in human ovarian granulosa cells by suppression of aromatase(CYP19) http://pubs.acs.org/doi/10.1021/jf3022817. J Agric Food Chem. 2012;60(34):8411–8. doi: 10.1021/jf3022817. [Lu DF, Yang LJ, Wang F, et al. Inhibitory effect of luteolin on estrogen biosynthesis in human ovarian granulosa cells by suppression of aromatase(CYP19)[J]. J Agric Food Chem, 2012, 60(34): 8411-8.] [DOI] [PubMed] [Google Scholar]
- 12.Santos JMS, Lins TLBG, Barberino RS, et al. Kaempferol promotes primordial follicle activation through the phosphatidylinositol 3- kinase/protein kinase B signaling pathway and reduces DNA fragmentation of sheep preantral follicles cultured in vitro. http://onlinelibrary.wiley.com/doi/full/10.1002/mrd.23107. Mol Reprod Dev. 2019;86(3):319–29. doi: 10.1002/mrd.23107. [Santos JMS, Lins TLBG, Barberino RS, et al. Kaempferol promotes primordial follicle activation through the phosphatidylinositol 3- kinase/protein kinase B signaling pathway and reduces DNA fragmentation of sheep preantral follicles cultured in vitro[J]. Mol Reprod Dev, 2019, 86(3): 319-29.] [DOI] [PubMed] [Google Scholar]
- 13.Sirotkin AV, Hrabovszká S, Štochmaľová A, et al. Effect of quercetin on ovarian cells of pigs and cattle. http://d.wanfangdata.com.cn/periodical/ChlQZXJpb2RpY2FsRW5nTmV3UzIwMjEwMzAyEiAxZTdkOGEwMjhiYzM1ODdhYWIyYWU2OWZhYjFhZjA3YxoIdXI4b2xpMWk%3D. Anim Reprod Sci. 2019;205(3):44–51. doi: 10.1016/j.anireprosci.2019.04.002. [Sirotkin AV, Hrabovszká S, Štochmaľová A, et al. Effect of quercetin on ovarian cells of pigs and cattle[J]. Anim Reprod Sci, 2019, 205(3): 44-51.] [DOI] [PubMed] [Google Scholar]
- 14.Naseer Z, Ahmad E, Epikmen ET, et al. Quercetin supplemented diet improves follicular development, oocyte quality, and reduces ovarian apoptosis in rabbits during summer heat stress. http://www.ncbi.nlm.nih.gov/pubmed/28532829. Theriogenology. 2017;96(5):136–41. doi: 10.1016/j.theriogenology.2017.03.029. [Naseer Z, Ahmad E, Epikmen ET, et al. Quercetin supplemented diet improves follicular development, oocyte quality, and reduces ovarian apoptosis in rabbits during summer heat stress[J]. Theriogenology, 2017, 96(5): 136-41.] [DOI] [PubMed] [Google Scholar]
- 15.Du XY, Huang J, Xu LQ, et al. The proto-oncogene c-src is involved in primordial follicle activation through the PI3K, PKC and MAPK signaling pathways. http://rbej.biomedcentral.com/articles/10.1186/1477-7827-10-58. Reprod Biol Endocrinol. 2012;10(7):58. doi: 10.1186/1477-7827-10-58. [Du XY, Huang J, Xu LQ, et al. The proto-oncogene c-src is involved in primordial follicle activation through the PI3K, PKC and MAPK signaling pathways[J]. Reprod Biol Endocrinol, 2012, 10(7): 58.] [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 16.Zhao Y, Zhang Y, Li J, et al. MAPK3/1 participates in the activation of primordial follicles through mTORC1-KITL signaling. http://onlinelibrary.wiley.com/doi/full/10.1002/jcp.25868. J Cell Physiol. 2018;233(1):226–37. doi: 10.1002/jcp.25868. [Zhao Y, Zhang Y, Li J, et al. MAPK3/1 participates in the activation of primordial follicles through mTORC1-KITL signaling[J]. J Cell Physiol, 2018, 233(1): 226-37.] [DOI] [PubMed] [Google Scholar]
- 17.Bellingham M, Amezaga MR, Mandon-Pepin B, et al. Exposure to chemical cocktails before or after conception: -the effect of timing on ovarian development. http://europepmc.org/articles/PMC3731555. Mol Cell Endocrinol. 2013;376(1/2):156–72. doi: 10.1016/j.mce.2013.06.016. [Bellingham M, Amezaga MR, Mandon-Pepin B, et al. Exposure to chemical cocktails before or after conception: -the effect of timing on ovarian development[J]. Mol Cell Endocrinol, 2013, 376(1/2): 156-72.] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Idrees M, Xu LG, Song SH, et al. PTPN11(SHP2) is indispensable for growth factors and cytokine signal transduction during bovine oocyte maturation and blastocyst development. http://www.ncbi.nlm.nih.gov/pubmed/31635340. Cells. 2019;8(10):E1272. doi: 10.3390/cells8101272. [Idrees M, Xu LG, Song SH, et al. PTPN11(SHP2) is indispensable for growth factors and cytokine signal transduction during bovine oocyte maturation and blastocyst development[J]. Cells, 2019, 8(10): E1272.] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Rumi MAK, Singh P, Roby KF, et al. Defining the role of estrogen receptor β in the regulation of female fertility. http://europepmc.org/abstract/MED/28520870. Endocrinology. 2017;158(7):2330–43. doi: 10.1210/en.2016-1916. [Rumi MAK, Singh P, Roby KF, et al. Defining the role of estrogen receptor β in the regulation of female fertility[J]. Endocrinology, 2017, 158(7): 2330-43.] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Elzaiat M, Herman L, Legois B, et al. High-throughput exploration of the network dependent on AKT1 in mouse ovarian granulosa cells. http://www.sciencedirect.com/science/article/pii/S1535947620315413. Mol Cell Proteomics. 2019;18(7):1307–19. doi: 10.1074/mcp.RA119.001461. [Elzaiat M, Herman L, Legois B, et al. High-throughput exploration of the network dependent on AKT1 in mouse ovarian granulosa cells [J]. Mol Cell Proteomics, 2019, 18(7): 1307-19.] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Richani D, Gilchrist RB. The epidermal growth factor network: role in oocyte growth, maturation and developmental competence. http://europepmc.org/abstract/MED/29029246. Hum Reprod Update. 2018;24(1):1–14. doi: 10.1093/humupd/dmx029. [Richani D, Gilchrist RB. The epidermal growth factor network: role in oocyte growth, maturation and developmental competence[J]. Hum Reprod Update, 2018, 24(1): 1-14.] [DOI] [PubMed] [Google Scholar]
- 22.Pontes JT, Maside C, Lima LF, et al. Immunolocalization for glucocorticoid receptor and effect of cortisol on in vitro development of preantral follicles. http://www.sciencedirect.com/science/article/pii/S2451943X18302382. VetAnim Sci. 2019;7:100060. doi: 10.1016/j.vas.2019.100060. [Pontes JT, Maside C, Lima LF, et al. Immunolocalization for glucocorticoid receptor and effect of cortisol on in vitro development of preantral follicles[J]. VetAnim Sci, 2019, 7: 100060.] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zheng Q, Zhou FF, Cui XY, et al. Novel serum biomarkers detected by protein array in polycystic ovary syndrome with low progesterone level. http://europepmc.org/abstract/MED/29734187. Cell Physiol Biochem. 2018;46(6):2297–310. doi: 10.1159/000489619. [Zheng Q, Zhou FF, Cui XY, et al. Novel serum biomarkers detected by protein array in polycystic ovary syndrome with low progesterone level[J]. Cell Physiol Biochem, 2018, 46(6): 2297-310.] [DOI] [PubMed] [Google Scholar]
- 24.Gorga A, Rindone G, Regueira M, et al. HIF involvement in the regulation of rat Sertoli cell proliferation by FSH. http://www.sciencedirect.com/science/article/pii/S0006291X1831297X. Biochem Biophys Res Commun. 2018;502(4):508–14. doi: 10.1016/j.bbrc.2018.05.206. [Gorga A, Rindone G, Regueira M, et al. HIF involvement in the regulation of rat Sertoli cell proliferation by FSH[J]. Biochem Biophys Res Commun, 2018, 502(4): 508-14.] [DOI] [PubMed] [Google Scholar]
- 25.Zhang ZH, Chen LY, Wang F, et al. Expression of hypoxia-inducible factor-1α during ovarian follicular growth and development in Sprague-Dawley rats. Genet Mol Res. 2015;14(2):5896–909. doi: 10.4238/2015.June.1.7. [Zhang ZH, Chen LY, Wang F, et al. Expression of hypoxia-inducible factor-1α during ovarian follicular growth and development in Sprague-Dawley rats[J]. Genet Mol Res, 2015, 14(2): 5896-909.] [DOI] [PubMed] [Google Scholar]
- 26.Maidarti M, Anderson RA, Telfer EE. Crosstalk between PTEN/ PI3K/Akt signalling and DNA damage in the oocyte: implications for primordial follicle activation, oocyte quality and ageing. http://www.researchgate.net/publication/338600372_Crosstalk_between_PTENPI3KAkt_Signalling_and_DNA_Damage_in_the_Oocyte_Implications_for_Primordial_Follicle_Activation_Oocyte_Quality_and_Ageing/download. Cells. 2020;9(1):E200. doi: 10.3390/cells9010200. [Maidarti M, Anderson RA, Telfer EE. Crosstalk between PTEN/ PI3K/Akt signalling and DNA damage in the oocyte: implications for primordial follicle activation, oocyte quality and ageing[J]. Cells, 2020, 9(1): E200.] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pan Y, Wang M, Wang L, et al. Estrogen improves the development of yak(Bos grunniens) oocytes by targeting cumulus expansion and levels of oocyte-secreted factors during in vitro maturation. http://www.researchgate.net/publication/344305086_Estrogen_improves_the_development_of_yak_Bos_grunniens_oocytes_by_targeting_cumulus_expansion_and_levels_of_oocyte-secreted_factors_during_in_vitro_maturation. PLoS One. 2020;15(9):e0239151. doi: 10.1371/journal.pone.0239151. [Pan Y, Wang M, Wang L, et al. Estrogen improves the development of yak(Bos grunniens) oocytes by targeting cumulus expansion and levels of oocyte-secreted factors during in vitro maturation[J]. PLoS One, 2020, 15(9): e0239151.] [DOI] [PMC free article] [PubMed] [Google Scholar]