

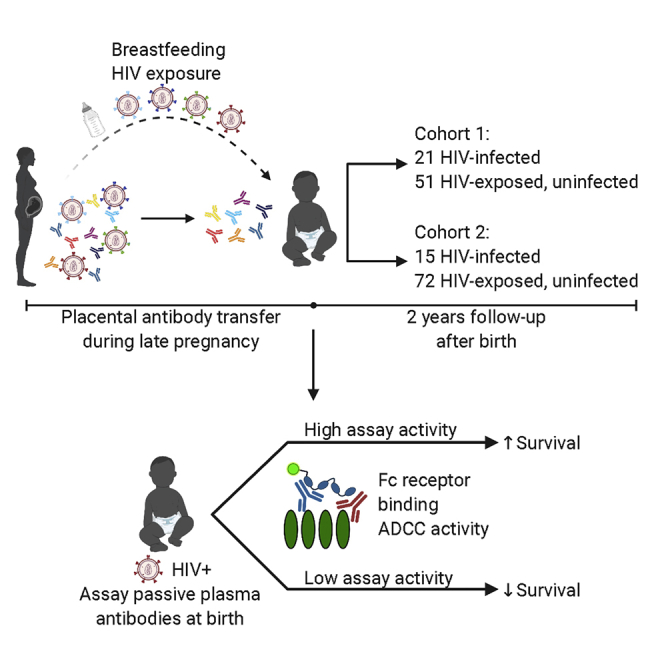
Summary
Defining immune responses that protect humans against diverse HIV strains has been elusive. Studying correlates of protection from mother-to-child transmission provides a benchmark for HIV vaccine protection because passively transferred HIV antibodies are present during infant exposure to HIV through breast milk. A previous study by our group illustrated that passively acquired antibody-dependent cellular cytotoxicity (ADCC) activity is associated with improved infant survival whereas neutralization is not. Here, we show, in another cohort and with two effector measures, that passively acquired ADCC antibodies correlate with infant survival. In combined analyses of data from both cohorts, there are highly statistically significant associations between higher infant survival and passively acquired ADCC levels (p = 0.029) as well as dimeric FcγRIIa (p = 0.002) or dimeric FcγRIIIa binding (p < 0.001). These results suggest that natural killer (NK) cell- and monocyte antibody-mediated effector functions may contribute to the observed survival benefit and support a role of pre-existing ADCC-mediating antibodies in clinical outcome.
Keywords: human immunodeficiency virus (HIV), antibody-dependent cellular cytotoxicity (ADCC), mother-to-child transmission (MTCT), passive antibody transfer, maternal antibodies, pediatric HIV, FcγR
Graphical abstract
Highlights
ADCC activity is associated with improved survival of HIV-infected infants
Binding to dimeric forms of Fcγ receptors is also associated with survival
These data support a role of ADCC in improved clinical outcome in infected infants
Mother-to-child transmission is a unique setting to define correlates of protection from HIV pathogenesis. Yaffe et al. show that passively acquired ADCC, measured by two assays, correlates with improved survival of HIV-infected infants in two breastfeeding cohorts. These results suggest that pre-existing, ADCC-mediating antibodies contribute to improved HIV-positive infant clinical outcome.
Introduction
Non-neutralizing antibodies (nnAbs) can mediate several cytotoxic effector functions, such as antibody-dependent cellular cytotoxicity (ADCC) and antibody-dependent phagocytosis (ADP), which play an important role in control of infectious diseases.1 nnAbs, particularly ADCC-mediating Abs, have been associated with protection from simian immunodeficiency virus (SIV) acquisition or progression to AIDS in several studies of macaques, a commonly used model for human HIV-1 (HIV) infection.2 Although early studies in humans also suggested that ADCC activity correlates with protection from HIV infection and/or slowed disease progression, human data regarding the role of HIV-specific ADCC-mediating Abs in infection and disease are still somewhat limited.3 Understanding the protective capacity of pre-existing nnAbs in HIV-exposed and infected individuals may help guide the design of an effective HIV vaccine.4
Our team has leveraged the unique setting of mother-to-child transmission (MTCT) to specifically investigate the role of ADCC-mediating Abs in outcomes in infants of mothers living with HIV.5 Maternal HIV-specific Abs are particularly relevant in this setting because they enter the infant circulation via transfer across the placenta and remain in circulation for months after birth.6, 7, 8, 9 We have shown previously that passively acquired infant plasma ADCC activity is correlated positively with infant survival but maternal plasma ADCC activity is not; nor are neutralization breadth and potency of infant or maternal plasma.9 In addition, maternal breast milk HIV-specific ADCC activity is correlated inversely with breastfeeding MTCT risk.10 These findings were observed using samples from the Nairobi Breastfeeding Trial (NBT), which was conducted prior to use of antiviral therapy for prevention of MTCT, and using HIV gp120 envelope protein in the rapid and fluorometric ADCC (RFADCC) assay, which may measure other non-neutralizing activities as well as ADCC.11, 12, 13, 14
Given the very limited data on immune correlates of protection in humans, we wanted to find out whether the findings from the NBT were generalizable to other cohorts and reproducible using a different ADCC assay. In a second cohort, the cytotoxic T lymphocyte (CTL) cohort,15 we again identified a significant association between passively acquired gp120-specific ADCC activity and infant survival. We also expanded our detection methods to include a dimeric, recombinant, soluble Fcγ receptor (FcγR) ELISA (dimeric FcγR) in addition to the RFADCC assay to measure the ADCC potential of passively acquired Abs.16,17 The dimeric FcγR ELISA results agreed with those of the RFADCC assay for both cohorts, and combined analyses using data from both cohorts identified a strong association between passively acquired ADCC activity and reduced infant mortality using both assays. These results provide further evidence for the therapeutic potential of pre-existing ADCC-mediating Abs in individuals that become HIV infected.
Results
Because the correlation of ADCC activity and infant outcome was detected previously in the NBT cohort using the RFADCC assay, we used the same assay in the current study with the goal of determining the generalizability of the association between RFADCC activity and infant outcomes. At the time of enrollment, clade A was dominant in Nairobi, Kenya, and clade D and recombinants were also common.18 We used gp120 derived from a clade A/D variant (BL035) isolated from an infant with HIV in the NBT cohort as the antigen in the studies of this cohort and showed that it gave results representative of gp120s from diverse strains in the RFADCC assay.9 To further validate this, we tested gp120 from a clade A strain, BG505, and observed a statistically significant association between passive ADCC antibodies and infant survival, as seen previously with BL035 and clade B SF162 gp120 antigens (Figure S1).9,19 In addition, we showed that prior results with SF162 gp120 antigen were reproducible in this study (Figure S1). These findings demonstrate a consistent association between infant ADCC activity and clinical outcome independent of the gp120 antigen tested.
Using the RFADCC assay and BL035 gp120 as the antigen, we measured the ADCC activity of 87 infants from the CTL cohort who were defined as HIV negative at birth, 15 of whom subsequently tested positive, and the other 72 remained HIV exposed, uninfected (HEU). As with the NBT cohort, we selected infants who tested HIV negative at birth so that we could assess the passively transferred levels of HIV-specific antibodies without interference from de novo responses. We included only infants with samples available from the first week of life, when HIV breastfeeding transmission risk and passively acquired ADCC activity are high.9,20 We examined the relationship between passively acquired ADCC levels in the first weeks of life with infection outcome and clinical outcome (in infants who subsequently acquired HIV [HIV+] in the CTL cohort). There was no statistically significant difference in ADCC activity between HEU and HIV+ infants (p = 0.73; Figure S2A), nor was passively acquired ADCC activity associated with HIV acquisition risk in a logistic regression analysis (odds ratio [OR], 1.00; p = 0.99; Table S1). In contrast, among HIV+ infants in the CTL cohort, those with ADCC activity above the median generally had longer survival times (Figure S2B), and there was a statistically significant association between pre-existing ADCC activity and improved survival after adjusting for maternal viral load (hazard ratio [HR], 0.97; p = 0.017; Cox proportional hazards model; Table 1).
Table 1.
Summary of Cox proportional hazards models of infant survival
| Assay | Cohort | HR (95% CI) | p Value |
|---|---|---|---|
| RFADCC | NBT | 0.94 (0.89, 0.99) | 0.024a |
| CTL | 0.97 (0.94, 0.99) | 0.017a | |
| combined | 0.97 (0.95, 0.99) | 0.029a | |
| Dimeric FcγRIIa ELISA | NBT | 0.81 (0.67, 0.97) | 0.020a |
| CTL | 0.74 (0.51, 1.07) | 0.11 | |
| combined | 0.83 (0.74, 0.93) | 0.0017a | |
| Dimeric FcγRIIIa ELISA | NBT | 0.80 (0.66, 0.97) | 0.024a |
| CTL | 0.77 (0.62, 0.95) | 0.017a | |
| combined | 0.82 (0.73, 0.92) | 0.00061a |
Shown are results of Cox proportional hazards modeling using assay activity + AUC maternal viral load as inputs and survival time and death as outcomes. HR, hazard ratio; CI, confidence interval. All statistics were rounded to the nearest two significant figures. The H131 and V158 variants were used for the dimeric FcγRIIa and FcγRIIIa ELISAs, respectively.
Statistically significant after Benjamini-Hochberg correction using a false discovery rate of 0.05. Data are from two technical and biological replicates. See also Figures S2 and S3 and Tables S1 and S2.
Because we used similar selection criteria for infants from both cohorts, we combined the RFADCC results obtained for HIV+ infants in the NBT and CTL cohorts (total n = 36) to determine whether there was an association between passively acquired ADCC activity and infant survival in the combined cohorts. Indeed, in the combined cohorts, we observed a statistically significant association between passively acquired ADCC activity and HIV+ infant survival when we compared infants with high (≥median) versus low (<median) ADCC activity (χ2 = 7.81, p = 0.0052; Figure 1) and when we used continuous data in Cox proportional hazards modeling adjusted for maternal viral load (HR, 0.97; p = 0.029; Table 1).
Figure 1.

Association between gp120-specific RFADCC activity and HIV+ infant outcome
Shown is a Kaplan-Meier survival curve for HIV+ infants with passively acquired ADCC activity at/above (blue) or below (red) the median from the NBT and CTL cohorts (n = 36). Clade A/D BL035 gp120 was used as the antigen. Black dots indicate time of censoring. Data are from two technical and biological replicates. See also Figure S1.
We next examined whether the effector potential of gp120-specific plasma antibodies measured by a different assay were correlated with infant outcome in the NBT cohort. For this, we employed the dimeric FcγR ELISA, which was developed to mimic the cross-linking of Fc receptors that occurs during initiation of effector functions.16 We used FcγRIIa, which is found on neutrophils, monocytes, and many other effector cell types, and FcγRIIIa, which is expressed by natural killer (NK) cells, CD16+ monocytes, and other leukocytes.21, 22, 23 There was no difference in median dimeric FcγR activity between HEU and HIV+ infants in the NBT cohort for FcγRIIa (p = 0.63) or FcγRIIIa (p = 0.90) via Mann-Whitney U test. Activity measured by this assay was also not associated with MTCT risk in logistic regression analysis for FcγRIIa (OR, 1.03; p = 0.58) or FcγRIIIa (OR, 1.03; p = 0.63; Table S1) after adjustment for maternal viral load. There was improved survival in HIV+ infants with higher activity measured in the dimeric FcyR ELISA that was statistically significant for FcγRIIa (HR, 0.81; p = 0.020) and FcγRIIIa (HR, 0.80; p = 0.024) in the NBT cohort (Table 1; Figure S3). All of these results are consistent with data from the RFADCC assay.9
We also examined ADCC activity using the dimeric FcγR ELISA assay in the CTL cohort, focusing only on HIV+ infants and survival as an outcome, given the lack of association between ADCC activity and infection risk as described previously described for the NBT cohort9 and reported here for the CTL cohort (Figure S2A; Table S1). In Cox proportional hazards modeling for the CTL cohort, there was a trend between FcγRIIa activity and infant survival (HR, 0.74; p = 0.11) and a statistically significant association of FcγRIIIa activity with infant survival (HR, 0.77; p = 0.017). Importantly, there was a statistically significant association between the activity of both receptors with survival when the two cohorts were combined: FcγRIIa (HR, 0.83; p = 0.0017) and FcγRIIIa (HR, 0.82; p = 0.00061). In combined survival analyses excluding the eight NBT infants infected after 6 months, when passive antibodies may be less relevant, there was still a statistically significant association between RFADCC (p = 0.021), dimeric FcγRIIa (p = 0.017), or dimeric FcγRIIIa (p = 0.0067) activity and infant survival. Kaplan-Meier curves are shown in Figure S3, and Cox proportional hazards survival analyses are summarized in Table 1. The results of models controlling for maternal CD4 count or CD4 count and viral load instead of viral load only were highly similar (Table S2).
As a final comparison of outcomes, we examined whether passively acquired RFADCC or dimeric FcγR ELISA activity differed between surviving and non-surviving HIV+ infants. Indeed, surviving HIV+ infants had significantly higher median RFADCC activity than infants who did not survive during the follow-up period (Figure 2A) as well as significantly higher ELISA activity for dimeric FcγRIIa (Figure 2B) and FcγRIIIa (Figure 2C). All comparisons between surviving and non-surviving infants were also significant in the separate cohorts (data not shown), with the exception of NBT infants for dimeric FcγRIIIa (p = 0.06).
Figure 2.

Comparison of gp120-specific RFADCC or dimeric FcγR activity in surviving and non-surviving HIV+ infants
(A) Normalized ADCC activity of surviving (blue, n = 20) and non-surviving (red, n = 15) HIV+ infants in the NBT and CTL cohorts.
(B) Area under the curve (AUC)-normalized dimeric FcγRIIa (H131 variant) ELISA activity for surviving (blue, n = 20) and non-surviving (red, n = 14) HIV+ infants in the NBT and CTL cohorts.
(C) AUC dimeric FcγRIIIa (variant V158) ELISA activity for surviving (blue, n = 20) and non-surviving (red, n = 14) HIV+ infants in the NBT and CTL cohorts.
Closed symbols represent infants from the NBT cohort, and open symbols represent infants from the CTL cohort. The sample for one HIV+ infant from the NBT cohort ran out before ELISAs were run. The median assay activity of groups in all panels was compared with Mann-Whitney U test. Clade A/D BL035 gp120 was used as the antigen. All p values are statistically significant after Benjamini-Hochberg correction using a false discovery rate of 0.05. Data are from two technical and biological replicates. See also Table S1.
Discussion
It has been challenging to define correlates of protection from HIV infection and disease progression in humans, where it will be necessary to protect against viruses that exhibit extensive diversity, which is difficult to capture in animal models.24 Despite this, growing evidence suggests that Fc-mediated effector functions may play an important role in protection and disease.3,23 For example, it has been shown in the RV144 trial that V1V2-specific nnAbs are correlated with protection from infection and that vaccinees exhibit engagement of FcγRIIa and FcγRIIIa by Abs recognizing a broad range of cross-clade antigens.17,25 With regard to disease outcome, long-term non-progressor status is also associated with development of de novo ADCC-mediating Abs.26, 27, 28 It has been difficult to study protection and disease progression simultaneously in the context of naturally circulating HIV. MTCT is a unique context in which the effect of pre-existing nnAbs on infection risk and disease outcome can be studied in the presence of endemic circulating HIV strains. Here we demonstrate that passively acquired ADCC antibody activity, measured by two different assays and in two different cohorts (individually or combined), is correlated with improved clinical outcome in infants who acquire HIV.
Passively acquired ADCC activity, measured with the RFADCC assay, was associated with improved HIV+ infant survival in the CTL cohort, similar to what has been observed previously in the NBT cohort in this assay.9 Importantly, this association was highly significant when the data using this assay from the two cohorts were combined. These findings suggest that the RFADCC assay measures a biologically relevant activity in this setting, although the specific nature of this activity in this assay remains poorly defined. This method uses gp120-coated target cells that may not completely resemble the epitopes presented by HIV-infected cells, and it measures NK cell- and monocyte-mediated effector functions.12, 13, 14 The findings presented here highlight the importance of better understanding the biological mechanisms underlying the activity that correlates with improved clinical outcomes.
The relationship between ADCC and infant outcome was also observed in both cohorts and in a combined analysis with a second assay that measures the ability of close-proximity antigen-antibody complexes to engage dimeric FcγRs, mimicking the clustering of FcγR required for activation of innate cells that mediate effector functions.16,29 The dimeric FcγR ELISA has been shown previously to correlate with multiple methods of measuring ADCC.17,30 Although ADCC activity is classically thought to be mediated by NK cells via FcγRIIIa, a growing amount of literature supports a potential role of FcγRIIa and monocytes in promoting ADCC and protective effects against HIV.1,13,17,31,32 Our finding that FcγRIIa binding was also associated with infant survival is consistent with the importance of multiple cellular triggers for Ab effector functions in the context of HIV infection. The highly concordant results among both cohorts and both assays may suggest that Fc-mediated effector functions, such as ADCC, play a role in limiting disease progression in infants with HIV.
The positive effect of passively acquired antibodies on infant infection is not unique to HIV. For example, ADCC-mediating maternal antibodies are associated with decreased herpes simplex virus (HSV) dissemination.33 Similarly, the degree of NK cell activation by RSV-specific maternal antibodies is inversely associated with infant RSV disease severity.34 ADCC has also been associated with improved outcomes in previous HIV MTCT studies; however, some of these studies were limited by imprecise determination of infant infectious status or study of maternal antibodies rather than the direct passive antibody levels in the infant.10,35, 36, 37 Here we provide strong support for a role of effector functions mediated by pre-existing antibodies in improving HIV clinical outcome. Given that there is cross-species variability in immunoglobulin-FcR interactions that could affect the relevance of study of HIV-specific ADCC antibodies in experimental systems, these human studies provide critical information for understanding which antibody activities are important for vaccine and therapy design.29,38
Limitations of study
This study has several limitations. Of the 35 infants who acquired HIV in our study, 7 had an estimated infection time of 6 months of age or greater, when passively acquired antibodies had largely waned.5 As described earlier, this did not appear to affect overall results because when the survival analyses were restricted to infants who acquired HIV before 6 months of age, the results were similar to those of the entire cohort(s). However, a larger study, focused on infants at risk and infected within the first 2 months of life, would more directly examine the effect of passive antibodies. A final caveat is use of the RFADCC assay, which relies on coated target cells, and an ELISA-based assay to measure ADCC activity mediated by passively transferred antibodies. Although it has been argued that these approaches are less biologically relevant than infection-based assays, both assays have been shown to be highly correlated with other measures of ADCC.16,17 Importantly, RFADCC activity was also associated with outcome measures in a number of human studies.9,10,39,40
STAR★Methods
Key resources table
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Antibodies | ||
| Polyclonal Anti-Human Immunodeficiency Virus Immune Globulin, Pooled Inactivated Human Sera (HIVIG) | NIH AIDS Reagent Program | Cat#3957 |
| Pierce High Sensitivity Streptavidin-HRP | ThermoFisher Scientific | Cat#21130 |
| Biological samples | ||
| Human plasma or cord blood samples from NBT cohort subjects | Nduati et al.20 | N/A |
| Human plasma or cord blood samples from CTL cohort subjects | John-Stewart et al.15 | N/A |
| Chemicals, eptides, and recombinant proteins | ||
| BG505.W6M.ENV.B1 (BG505) gp120 | Cambridge Biologics | Cat#01-01-1028 |
| BL035.W6M.ENV.C1 (BL035) gp120 | Immune Technology Corporation | Cat#IT-001-115p |
| SF162 gp120 | Immune Technology Corporation | Cat#IT-001-0028p |
| Critical commercial assays | ||
| Vybrant CFDA SE. Cell Tracer Kit | Invitrogen | Cat#V12883 |
| PKH26 Red Fluorescent Cell Linker Kit | Sigma Aldrich | Cat# MINI26-1KT |
| Experimental models: Cell lines | ||
| Human: CEM.NKR | NIH AIDS Reagent Program | Cat#458; RRID: CVCL_X622 |
| Software and algorithms | ||
| FlowJo v9.9 | TreeStar | RRID: SCR_008520 |
| Excel | Microsoft | RRID: SCR_016137 |
| Prism 8.0c | GraphPad | RRID: SCR_002798 |
| RStudio | RStudio | RRID: SCR_000432 |
| Other | ||
| 1-Step Ultra TMB-ELISA Substrate Solution | ThermoFisher Scientific | Cat# 34029 |
Resource availability
Lead contact
Further information and requests for resources and reagents should be directed to and will be fulfilled by the lead contact, Julie Overbaugh (joverbau@fredhutch.org).
Materials availability
This study did not generate new unique reagents.
Data and code availability
The custom RStudio code generated and used in this study is available upon request.
Experimental model and subject details
Human subjects
Plasma samples used in this study were from the Nairobi Breastfeeding Trial (NBT) and Cytotoxic T Lymphocyte (CTL) cohort. These studies enrolled HIV+ mothers late in pregnancy from 1992-1998 for NBT and 1999-2002 for CTL. Individuals were enrolled in late pregnancy, and mother-infant pairs were followed for two years of life in the NBT and for one year for CTL, with the exception that CTL infants who became infected in the first year were followed for an additional year.15,20 Both studies were carried out before the widespread availability of antiretroviral therapy (ART). Whereas all NBT participants did not receive ART, in CTL nearly all mothers received a short course of zidovudine prior to delivery for prevention of infant infection, which was consistent with contemporaneous standard of care. Approval to conduct this study was provided by Kenyatta National Hospital - University of Nairobi Ethics and Research Committee, the Fred Hutchinson Cancer Research Center Institutional Review Board, and/or the University of Washington Institutional Review Board. Study participants provided written informed consent prior to enrollment and for use of their data and samples for future studies.
Cell lines
CEM.NKR cells (RRID:CVCL_X622; originally derived from female human T-lymphoblastoid cells) were obtained from NIH AIDS Reagent Program (catalog #458) and grown at 37°C in RPMI 1640 media with added penicillin (100 U/mL), streptomycin (100 μg/mL), amphotericin B (250 ng/mL), L-glutamine (2mM), and fetal bovine serum (10%). These cells were not further authenticated in our hands.
Method details
Study Design
In both groups, a sub-cohort was selected for inclusion in this study based on infant criteria described in Milligan et al., which included HIV negative DNA/RNA testing at birth, breastfeeding history ≥ 3 months or until time of transmission, and availability of an infant sample from the first week of life.9 All eligible 72 cases from the NBT cohort were included in the study. In the CTL cohort, 161 mother-infant pairs initially met the same criteria, only 15 of which included infants who acquired HIV. Given that this large number of primarily non-transmitting pairs would make running all samples in parallel with the RFADCC assay difficult, the design was amended to a case-control design through selection of a random sample of 78 from the 161 mother-infant pairs. Six of 15 infected and 72 of 146 uninfected cases were randomly selected. The nine remaining infected cases were added back in after selection. In order to account for the non-random addition of infected cases, all samples were weighted according to Borgan II weighting.41 Infected cases were assigned a weight of 1, CTL HEUs were assigned a weight of 2.028, and all NBT samples were assigned a weight of 1. All infant samples from the CTL cohort were plasma, whereas 61 and 11 infant samples from NBT were cord blood and plasma, respectively. All samples were heat inactivated at 56°C for 1 hour. Insufficient volume remained for plasma of one infant in the NBT study to run the dimeric FcγR ELISAs on this sample.
Rapid and Fluorometric ADCC Assay
The rapid and fluorometric ADCC (RFADCC) assay was carried out as described previously.11,42 Briefly, CEM.NKR cells were stained with PKH26 cell linker dye (Sigma-Aldrich) and CFSE cytosolic dye (Vybrant CFDA-SE. Cell Tracer Kit, Invitrogen). Double-stained cells were then coated with Clade A/D BL035.W6M.ENV.C1 gp120 (Immune Tech; GenBank: DQ208480) at a ratio of 1.5ug gp120 per 100,000 cells for 1 hour at room temperature. Coated cells were washed and a total of 5,000 target cells were added to wells containing 100uL of plasma diluted at 1:5000 in RPMI media or 100-500ng/mL of control mAbs. After a 15-minute incubation, PBMCs from HIV negative donors were added at a ratio of 50:1 effector to target cells. RFADCC activity was allowed to occur for four hours at 37°C before cells were washed and fixed in 1% paraformaldehyde (Santa Cruz Biotechnology). Data were acquired via flow cytometry (BD Symphony). PKH and CFSE were detected in the PE and FITC channels, respectively. Data were analyzed using FlowJo (v.9.9, Treestar). ADCC was defined as the percentage of PKH+, CFSE- cells out of total PKH+ cells after subtracting the activity mediated against uncoated target cells (background), which was set to 3%–5%. ADCC activity was normalized to that mediated by Anti-HIV Immune Globulin (HIVIG, NIH ARP, Catalog #3957).
Dimeric FcγR ELISA
The ELISA using dimeric recombinant soluble FcγRIIIa and FcγRIIa ectodomains (dimeric FcγR) was adapted from Wines et al.16. Maxisorp 384-well plates were coated at 4°C overnight with BL035.W6M.ENV.C1 gp120 at 1ug/mL in PBS. Plates were washed five times between all steps in PBS-0.02% Tween-20-1mM EDTA (PBSE). Plates were blocked with 1% human serum albumin (Sigma Aldrich) in PBSE at 37°C for 1 hour. All samples/reagents used in subsequent steps were diluted in 1% bovine serum albumin (Sigma Aldrich) in PBSE. Plasma samples at 1:50, 1:100, or 1:200 dilutions and mAb controls were then incubated at 37°C for 1 hour. Biotinylated dimeric rsFcγRIIa variant H131 or rsFcγRIIIa variant V158 were prepared as previously described and added at 0.2 μg or 0.1 μg/mL, respectively, for 1 hour at 37°C.16 Next, plates were incubated with high sensitivity streptavidin-HRP (Pierce) at 1:10,000 dilution for 1 hour at 37°C. After a final wash, 1-Step TMB-Ultra (Thermo Fisher) was added to the wells. The reaction was stopped 5-10 minutes later using 1M H2SO4 (Sigma Aldrich). Absorbance was measured at 450nm optical density. The activity measured for wells coated with HIVIG was used to normalize measurements across experiments, after subtracting the background activity of wells in which no plasma/mAb was added. The area under the curve (AUC) for the activities at each dilution normalized to that of HIVIG was calculated and divided by 1000 for use in subsequent analyses.
Quantification and statistical analysis
Raw data was processed in Microsoft Excel. Statistical analyses were performed using RStudio (RStudio Team 2018) or GraphPad Prism v8. Graphs were generated using GraphPad Prism. In place of a single viral load, an area under the curve (AUC) for maternal viral loads was calculated to adjust for the potential impact of ART on maternal viral load and therefore HIV transmission risk. Linear regression on adjacent viral load time points was used to impute missing viral loads where necessary. For the CTL cohort, all AUC calculations used viral loads from pregnancy week 32 (P32), delivery, and one month postpartum. For the NBT cohort, AUC calculations used viral loads from P32 and delivery, week eight postpartum, or week 14 postpartum, where available. AUCs were divided by the total measurement time interval to account for the difference in measurement interval between samples. The AUC/week of maternal viral load was used in all subsequent transmission and survival analyses.
A binomial logistic regression of assay activity and viral load on infant infection status was performed to determine whether these factors were associated with HIV transmission. To determine whether assay activity was associated with survival of infants with HIV, assay activity and viral load were used as predictors in Cox-proportional hazards models of infant survival time. For models controlling for maternal CD4 count with or instead of viral load, the CD4 count closest to delivery was selected if data from multiple time points were available. Kaplan-Meier survival curves were also used to compare HIV+ infants with assay activity at/above or below the median via the log-rank test. Statistical significance was defined as a p value less than 0.05 and, where multiple comparisons were made, a Benjamini-Hochberg correction with a false discovery rate of 0.05 was applied. In all figures, N corresponds to the number of individuals. Except where noted, all data represent the mean of two biological replicates, with two technical replicates in each experiment. Additional details can be found in the figure legends.
Acknowledgments
We thank the participants and staff of the NBT and CTL cohorts. We thank Vrasha Chohan for initial help with the dimeric FcγR ELISA. This work was supported by NIH R01 HD103571 (to J.O.), training award T32 AI083203 (to Z.A.Y.), NIH F30AI136636 (to N.E.N.), and NHMRC project funding (to B.D.W. and P.M.H.). This research was also supported by the Flow Cytometry shared resource of the Fred Hutch/University of Washington Cancer Consortium (P30 CA015704). The graphical abstract was generated with BioRender.
Author contributions
Conceptualization, J.O. and G.J.-S.; methodology, Z.A.Y., N.E.N., J.S., B.D.W., P.M.H., and B.A.R.; investigation and data collection, Z.A.Y. and N.E.N.; clinical data and samples, G.J.-S., R.N., D.M.N., R.B., and C.F.; writing – original draft, Z.A.Y., N.E.N., and J.O.; writing – review & editing, Z.A.Y., N.E.N., J.O., G.J.-S., and B.A.R.; funding acquisition, J.O. and G.J.-S.; resources, J.O.; supervision, J.O.; comments on the manuscript, all authors.
Declaration of interests
The authors declare no competing interests.
Inclusion and diversity
One or more of the authors of this paper self-identifies as an underrepresented ethnic minority in science. One or more of the authors of this paper self-identifies as a member of the LGBTQ+ community. The author list of this paper includes contributors from the location where the research was conducted who participated in the data collection, design, analysis, and/or interpretation of the work.
Published: April 20, 2021
Footnotes
Supplemental information can be found online at https://doi.org/10.1016/j.xcrm.2021.100254.
Supplemental information
References
- 1.Lu L.L., Suscovich T.J., Fortune S.M., Alter G. Beyond binding: antibody effector functions in infectious diseases. Nat. Rev. Immunol. 2018;18:46–61. doi: 10.1038/nri.2017.106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Forthal D.N., Finzi A. Antibody-dependent cellular cytotoxicity in HIV infection. AIDS. 2018;32:2439–2451. doi: 10.1097/QAD.0000000000002011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lewis G.K. Role of Fc-mediated antibody function in protective immunity against HIV-1. Immunology. 2014;142:46–57. doi: 10.1111/imm.12232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fouda G.G., De Paris K., Levy O., Marchant A., Gray G., Permar S., Marovich M., Singh A. Immunological mechanisms of inducing HIV immunity in infants. Vaccine. 2020;38:411–415. doi: 10.1016/j.vaccine.2019.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Milligan C., Slyker J.A., Overbaugh J. The Role of Immune Responses in HIV Mother-to-Child Transmission. Adv. Virus Res. 2018;100:19–40. doi: 10.1016/bs.aivir.2017.10.001. [DOI] [PubMed] [Google Scholar]
- 6.Parekh B.S., Shaffer N., Coughlin R., Hung C.H., Krasinski K., Abrams E., Bamji M., Thomas P., Hutson D., Schochetman G., The NYC Perinatal HIV Transmission Study Group Dynamics of maternal IgG antibody decay and HIV-specific antibody synthesis in infants born to seropositive mothers. AIDS Res. Hum. Retroviruses. 1993;9:907–912. doi: 10.1089/aid.1993.9.907. [DOI] [PubMed] [Google Scholar]
- 7.Kourtis A.P., Bulterys M., Nesheim S.R., Lee F.K. Understanding the timing of HIV transmission from mother to infant. JAMA. 2001;285:709–712. doi: 10.1001/jama.285.6.709. [DOI] [PubMed] [Google Scholar]
- 8.Lynch J.B., Nduati R., Blish C.A., Richardson B.A., Mabuka J.M., Jalalian-Lechak Z., John-Stewart G., Overbaugh J. The breadth and potency of passively acquired human immunodeficiency virus type 1-specific neutralizing antibodies do not correlate with the risk of infant infection. J. Virol. 2011;85:5252–5261. doi: 10.1128/JVI.02216-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Milligan C., Richardson B.A., John-Stewart G., Nduati R., Overbaugh J. Passively acquired antibody-dependent cellular cytotoxicity (ADCC) activity in HIV-infected infants is associated with reduced mortality. Cell Host Microbe. 2015;17:500–506. doi: 10.1016/j.chom.2015.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mabuka J., Nduati R., Odem-Davis K., Peterson D., Overbaugh J. HIV-specific antibodies capable of ADCC are common in breastmilk and are associated with reduced risk of transmission in women with high viral loads. PLoS Pathog. 2012;8:e1002739. doi: 10.1371/journal.ppat.1002739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gómez-Román V.R., Florese R.H., Patterson L.J., Peng B., Venzon D., Aldrich K., Robert-Guroff M. A simplified method for the rapid fluorometric assessment of antibody-dependent cell-mediated cytotoxicity. J. Immunol. Methods. 2006;308:53–67. doi: 10.1016/j.jim.2005.09.018. [DOI] [PubMed] [Google Scholar]
- 12.Kramski M., Schorcht A., Johnston A.P., Lichtfuss G.F., Jegaskanda S., De Rose R., Stratov I., Kelleher A.D., French M.A., Center R.J. Role of monocytes in mediating HIV-specific antibody-dependent cellular cytotoxicity. J. Immunol. Methods. 2012;384:51–61. doi: 10.1016/j.jim.2012.07.006. [DOI] [PubMed] [Google Scholar]
- 13.Kramski M., Parsons M.S., Stratov I., Kent S.J. HIV-specific antibody immunity mediated through NK cells and monocytes. Curr. HIV Res. 2013;11:388–406. doi: 10.2174/1570162x113116660061. [DOI] [PubMed] [Google Scholar]
- 14.Alrubayyi A., Schuetz A., Lal K.G., Jongrakthaitae S., Paolino K.M., Ake J.A., Robb M.L., de Souza M.S., Michael N.L., Paquin-Proulx D., Eller M.A. A flow cytometry based assay that simultaneously measures cytotoxicity and monocyte mediated antibody dependent effector activity. J. Immunol. Methods. 2018;462:74–82. doi: 10.1016/j.jim.2018.08.012. [DOI] [PubMed] [Google Scholar]
- 15.John-Stewart G.C., Mbori-Ngacha D., Payne B.L., Farquhar C., Richardson B.A., Emery S., Otieno P., Obimbo E., Dong T., Slyker J. HV-1-specific cytotoxic T lymphocytes and breast milk HIV-1 transmission. J. Infect. Dis. 2009;199:889–898. doi: 10.1086/597120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wines B.D., Vanderven H.A., Esparon S.E., Kristensen A.B., Kent S.J., Hogarth P.M. Dimeric FcγR Ectodomains as Probes of the Fc Receptor Function of Anti-Influenza Virus IgG. J. Immunol. 2016;197:1507–1516. doi: 10.4049/jimmunol.1502551. [DOI] [PubMed] [Google Scholar]
- 17.McLean M.R., Madhavi V., Wines B.D., Hogarth P.M., Chung A.W., Kent S.J. Dimeric Fcγ Receptor Enzyme-Linked Immunosorbent Assay To Study HIV-Specific Antibodies: A New Look into Breadth of Fcγ Receptor Antibodies Induced by the RV144 Vaccine Trial. J. Immunol. 2017;199:816–826. doi: 10.4049/jimmunol.1602161. [DOI] [PubMed] [Google Scholar]
- 18.Neilson J.R., John G.C., Carr J.K., Lewis P., Kreiss J.K., Jackson S., Nduati R.W., Mbori-Ngacha D., Panteleeff D.D., Bodrug S. Subtypes of human immunodeficiency virus type 1 and disease stage among women in Nairobi, Kenya. J. Virol. 1999;73:4393–4403. doi: 10.1128/jvi.73.5.4393-4403.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Naiman N.E., Slyker J., Richardson B.A., John-Stewart G., Nduati R., Overbaugh J.M. Antibody-dependent cellular cytotoxicity targeting CD4-inducible epitopes predicts mortality in HIV-infected infants. EBioMedicine. 2019;47:257–268. doi: 10.1016/j.ebiom.2019.08.072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Nduati R., John G., Mbori-Ngacha D., Richardson B., Overbaugh J., Mwatha A., Ndinya-Achola J., Bwayo J., Onyango F.E., Hughes J., Kreiss J. Effect of breastfeeding and formula feeding on transmission of HIV-1: a randomized clinical trial. JAMA. 2000;283:1167–1174. doi: 10.1001/jama.283.9.1167. [DOI] [PubMed] [Google Scholar]
- 21.Nimmerjahn F., Gordan S., Lux A. FcγR dependent mechanisms of cytotoxic, agonistic, and neutralizing antibody activities. Trends Immunol. 2015;36:325–336. doi: 10.1016/j.it.2015.04.005. [DOI] [PubMed] [Google Scholar]
- 22.Golay J., Valgardsdottir R., Musaraj G., Giupponi D., Spinelli O., Introna M. Human neutrophils express low levels of FcγRIIIA, which plays a role in PMN activation. Blood. 2019;133:1395–1405. doi: 10.1182/blood-2018-07-864538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Su B., Dispinseri S., Iannone V., Zhang T., Wu H., Carapito R., Bahram S., Scarlatti G., Moog C. Update on Fc-Mediated Antibody Functions Against HIV-1 Beyond Neutralization. Front. Immunol. 2019;10:2968. doi: 10.3389/fimmu.2019.02968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Burton D.R., Poignard P., Stanfield R.L., Wilson I.A. Broadly neutralizing antibodies present new prospects to counter highly antigenically diverse viruses. Science. 2012;337:183–186. doi: 10.1126/science.1225416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Haynes B.F., Gilbert P.B., McElrath M.J., Zolla-Pazner S., Tomaras G.D., Alam S.M., Evans D.T., Montefiori D.C., Karnasuta C., Sutthent R. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N. Engl. J. Med. 2012;366:1275–1286. doi: 10.1056/NEJMoa1113425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lambotte O., Ferrari G., Moog C., Yates N.L., Liao H.X., Parks R.J., Hicks C.B., Owzar K., Tomaras G.D., Montefiori D.C. Heterogeneous neutralizing antibody and antibody-dependent cell cytotoxicity responses in HIV-1 elite controllers. AIDS. 2009;23:897–906. doi: 10.1097/QAD.0b013e328329f97d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wren L.H., Chung A.W., Isitman G., Kelleher A.D., Parsons M.S., Amin J., Cooper D.A., Stratov I., Navis M., Kent S.J., ADCC study collaboration investigators Specific antibody-dependent cellular cytotoxicity responses associated with slow progression of HIV infection. Immunology. 2013;138:116–123. doi: 10.1111/imm.12016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Madhavi V., Wines B.D., Amin J., Emery S., ENCORE1 Study Group, Lopez E., Kelleher A., Sydney LTNP Study Group, Center R.J., Hogarth P.M. HIV-1 Env- and Vpu-Specific Antibody-Dependent Cellular Cytotoxicity Responses Associated with Elite Control of HIV. J. Virol. 2017;91:e00700-17. doi: 10.1128/JVI.00700-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Bournazos S., Wang T.T., Dahan R., Maamary J., Ravetch J.V. Signaling by Antibodies: Recent Progress. Annu. Rev. Immunol. 2017;35:285–311. doi: 10.1146/annurev-immunol-051116-052433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Anand S.P., Prévost J., Baril S., Richard J., Medjahed H., Chapleau J.P., Tolbert W.D., Kirk S., Smith A.B., 3rd, Wines B.D. Two Families of Env Antibodies Efficiently Engage Fc-Gamma Receptors and Eliminate HIV-1-Infected Cells. J. Virol. 2018;93:e01823-18. doi: 10.1128/JVI.01823-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hogarth P.M., Pietersz G.A. Fc receptor-targeted therapies for the treatment of inflammation, cancer and beyond. Nat. Rev. Drug Discov. 2012;11:311–331. doi: 10.1038/nrd2909. [DOI] [PubMed] [Google Scholar]
- 32.Neidich S.D., Fong Y., Li S.S., Geraghty D.E., Williamson B.D., Young W.C., Goodman D., Seaton K.E., Shen X., Sawant S., HVTN 505 Team Antibody Fc effector functions and IgG3 associate with decreased HIV-1 risk. J. Clin. Invest. 2019;129:4838–4849. doi: 10.1172/JCI126391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kohl S. The neonatal human’s immune response to herpes simplex virus infection: a critical review. Pediatr. Infect. Dis. J. 1989;8:67–74. [PubMed] [Google Scholar]
- 34.van Erp E.A., Lakerveld A.J., de Graaf E., Larsen M.D., Schepp R.M., Hipgrave Ederveen A.L., Ahout I.M., de Haan C.A., Wuhrer M., Luytjes W. Natural killer cell activation by respiratory syncytial virus-specific antibodies is decreased in infants with severe respiratory infections and correlates with Fc-glycosylation. Clin. Transl. Immunol. 2020;9:e1112. doi: 10.1002/cti2.1112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ljunggren K., Moschese V., Broliden P.A., Giaquinto C., Quinti I., Fenyö E.M., Wahren B., Rossi P., Jondal M. Antibodies mediating cellular cytotoxicity and neutralization correlate with a better clinical stage in children born to human immunodeficiency virus-infected mothers. J. Infect. Dis. 1990;161:198–202. doi: 10.1093/infdis/161.2.198. [DOI] [PubMed] [Google Scholar]
- 36.Broliden K., Sievers E., Tovo P.A., Moschese V., Scarlatti G., Broliden P.A., Fundaro C., Rossi P. Antibody-dependent cellular cytotoxicity and neutralizing activity in sera of HIV-1-infected mothers and their children. Clin. Exp. Immunol. 1993;93:56–64. doi: 10.1111/j.1365-2249.1993.tb06497.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Tranchat C., Van de Perre P., Simonon-Sorel A., Karita E., Benchaïb M., Lepage P., Desgranges C., Boyer V., Trépo C. Maternal humoral factors associated with perinatal human immunodeficiency virus type-1 transmission in a cohort from Kigali, Rwanda, 1988-1994. J. Infect. 1999;39:213–220. doi: 10.1016/s0163-4453(99)90052-x. [DOI] [PubMed] [Google Scholar]
- 38.Crowley A.R., Ackerman M.E. Mind the Gap: How Interspecies Variability in IgG and Its Receptors May Complicate Comparisons of Human and Non-human Primate Effector Function. Front. Immunol. 2019;10:697. doi: 10.3389/fimmu.2019.00697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Madhavi V., Wren L.H., Center R.J., Gonelli C., Winnall W.R., Parsons M.S., Kramski M., Kent S.J., Stratov I. Breadth of HIV-1 Env-specific antibody-dependent cellular cytotoxicity: relevance to global HIV vaccine design. AIDS. 2014;28:1859–1870. doi: 10.1097/QAD.0000000000000310. [DOI] [PubMed] [Google Scholar]
- 40.Ruiz M.J., Salido J., Abusamra L., Ghiglione Y., Cevallos C., Damilano G., Rodriguez A.M., Trifone C., Laufer N., Giavedoni L.D. Evaluation of Different Parameters of Humoral and Cellular Immune Responses in HIV Serodiscordant Heterosexual Couples: Humoral Response Potentially Implicated in Modulating Transmission Rates. EBioMedicine. 2017;26:25–37. doi: 10.1016/j.ebiom.2017.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Borgan O., Langholz B., Samuelsen S.O., Goldstein L., Pogoda J. Exposure stratified case-cohort designs. Lifetime Data Anal. 2000;6:39–58. doi: 10.1023/a:1009661900674. [DOI] [PubMed] [Google Scholar]
- 42.Williams K.L., Stumpf M., Naiman N.E., Ding S., Garrett M., Gobillot T., Vézina D., Dusenbury K., Ramadoss N.S., Basom R. Identification of HIV gp41-specific antibodies that mediate killing of infected cells. PLoS Pathog. 2019;15:e1007572. doi: 10.1371/journal.ppat.1007572. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The custom RStudio code generated and used in this study is available upon request.