

Mucosal surfaces, which are frequently exposed to external antigens, are a major part of the immune system. The development of oral tolerance is a result of a complex interplay between a variety of innate and adaptive immune functions; deficiencies in these mechanisms may contribute to the development of allergic disorders. Although regulatory T cells have a clear role in enforcing oral tolerance, the role of the dominant immunoglobulin in the gut, immunoglobulin A (IgA), is less clear. This perspective will focus on IgA and its role in tolerance to oral antigens and development of food allergies.
IgA is the most abundantly produced antibody isotype and has two subsets, IgA1 and IgA2, of which IgA2 predominates in colonic secretions.1 It exists primarily in two forms: monomeric IgA in the serum and polymeric IgA in the mucosal surface. Secretory IgA is predominantly dimeric, with two monomers, of the same subsets, connected via a J chain and associated with secretory component. Gut-associated lymphoid tissue (GALT), composed of Peyer’s patches (PP) and other aggregates of lymphoid tissue, located throughout the intestine, are composed primarily of B lymphocytes, with a lesser amount of T lymphocytes, myeloid cells and innate lymphoid cells. Antigen is sampled via transcytosis through microfold cells (M cells), goblet cells, and macrophages and dendritic cells (DC); DCs in PPs and mesenteric lymph nodes (MLNs) present antigen and guide antigen-specific IgA development, which is promoted by transforming growth factor beta (TGF-ß), retinoic acid, and interleukin 10 (IL-10).1 Polymeric IgA, as well as IgM, is excreted into the gastrointestinal lumen and other mucosal surfaces via transcytosis through the polymeric immunoglobulin receptor.1
IgA is thought to have a variety of functions to maintain the epithelial barrier and promote intestinal homeostasis. One of the main forms of mucosal defense is immune exclusion, which limits bacterial colonization and penetration of antigen through the epithelial surface.1 Immune exclusion is accomplished primarily though secretory IgA, which can trap antigen. This, along with intestinal peristalsis, defensins, and mucin, results in removal of antigen from the epithelial surface and hinderance of potential immune responses. IgA also allows for intestinal homeostasis by shaping the composition of commensal micro-organisms.1 Given these demonstrated roles of IgA in the gut to microbes and toxins, it is presumed that IgA might also promote oral tolerance to food antigens.
The role of IgA in the development of atopic disorders has been investigated by examining patients with selective IgA deficiency. Results have been mixed but overall show that those with IgA deficiency or a delay in IgA production (due to immune system immaturity) are at an increased risk of atopic disorders, but there is limited data for food allergy. Systemic versus mucosal IgA production have different dynamics, with IgA production initiating in the first weeks of life in the gut after exposure to foreign antigens such as the commensals, but serum IgA responses mature slowly in the first 2 years and then more rapidly. Clinical studies have examined the association between mucosal IgA, measured as fecal or salivary IgA responses, and development of atopy, with conflicting results. Limited data suggest that in immune competent hosts (in whom IgA responses are intact), mucosal IgA was associated with tolerance to foods whereas elevated systemic IgA indicated increased level of sensitization. Clinical discoveries suggest that IgA may be involved in the development of tolerance overtime. IgA2 may counteract IgE responses in egg allergy in children who naturally “outgrew” their allergy compared to those who did not2, although more recent findings are conflicting. Upregulation of specific IgA (along with IgG) has been seen in those with clinical response to oral and sublingual food immunotherapy. It is not known whether mucosal IgA is an epiphenomenon (i.e. secondary effect) seen in development or loss of oral tolerance due to antigen exposure (resulting in specific IgA) or inflammation (resulting in non-specific IgA), or whether IgA plays a causal role in food allergy. While IgA can play a role in immune exclusion, other mechanisms may be involved, such as IgA control of gut microbiota and homeostasis or a modified response elicited by targeted antigen uptake as immune complexes1 (Figure 1). Mechanistic work on the role of mucosal IgA in food allergy and oral tolerance is needed, but studies in humans are challenging. In our work, we found little induction of food-specific IgA in the gut through oral exposure unless an adjuvant is co-administered with the food3, which is consistent with a number of published mouse models using different food antigens. This suggests that IgA is not a part of steady state oral tolerance in mice and highlights that a better mechanistic understanding of IgA-inducing factors in the gut is needed.
Figure 1. Mechanisms by which IgA may ameliorate the risk of food allergy.
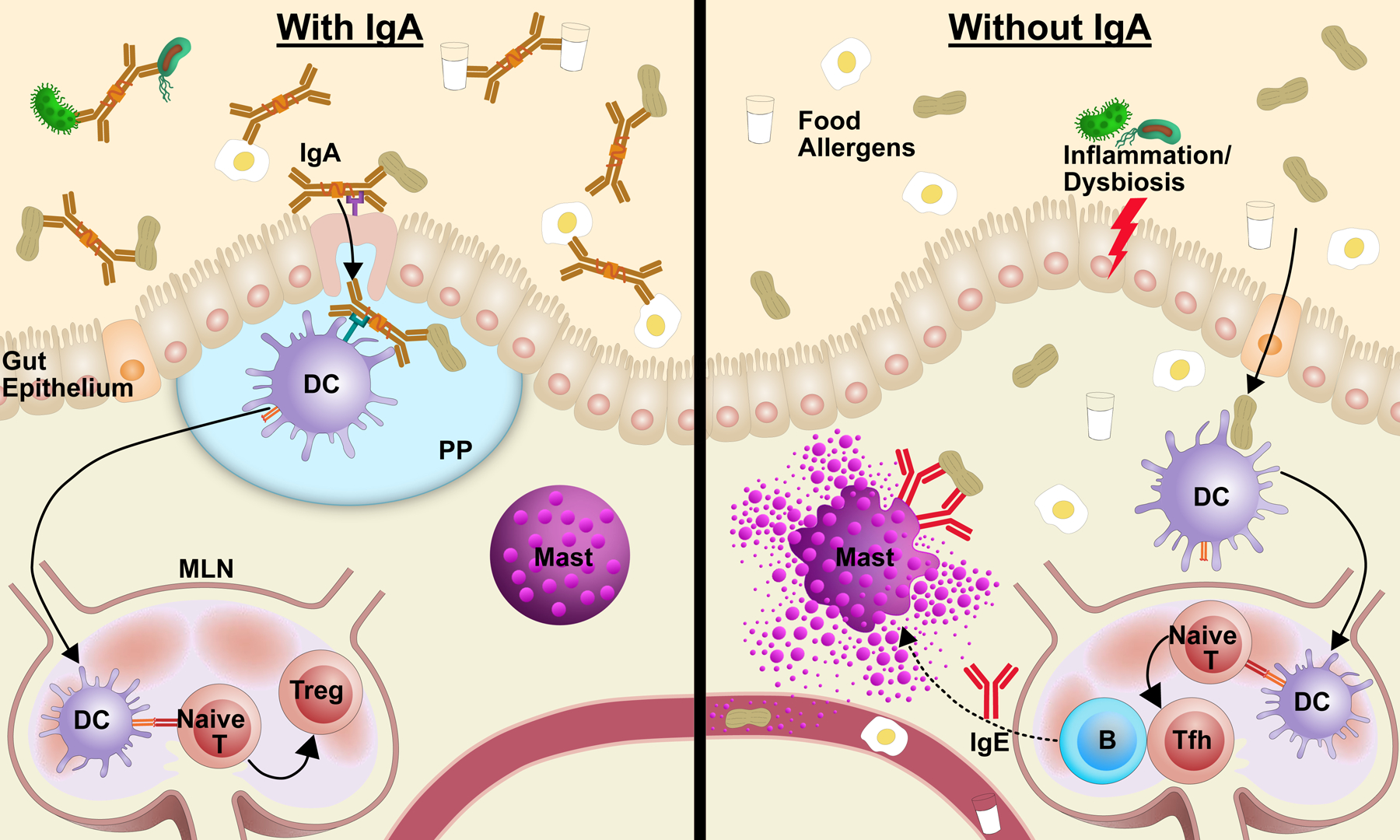
Left: Food-specific IgA may lead to immune exclusion, targeted uptake by PP, and shape microbiome, promoting tolerance. Right: Without food-specific IgA, food antigen may be taken up and presented under conditions that promote IgE sensitization.
Limited human data into the potential role for IgA as an active component promoting oral tolerance come from studies assessing the association of human milk IgA to food allergy. Antibodies in breast milk are the result of exposure to antigens in the maternal gut, as B cells in the mammary gland migrate from the intestine (via “enteromammary link”) and other mucosal surfaces. Those studies found that maternal cow’s milk (CM) avoidance resulted in lower levels of CM-specific IgA in human milk and lower CM-specific IgA was associated with the development of CM allergy4. In this report we showed that human milk IgA may decrease transcytosis of CM antigens in a gut epithelial cell model enhancing immune exclusion. In an animal model, we showed that breast milk IgA may contribute to targeted peanut antigen uptake to the Peyer’s patches.5
The maintenance of oral tolerance is an important and necessary function of the immune system and disruption is thought to play a role in the development of allergic hypersensitivities, such as food allergies. How exactly systemic and mucosal IgA responses contribute to development of oral tolerance versus sensitization to foods is unknown and an area of active investigation. A deeper understanding of these mechanisms would not only provide a better understanding of the gut immune system but may ultimately lead to advancements in allergy prevention and treatment.
Funding Source:
JP indicates no funding. EGL is supported by a T32 Institutional Training Grant, number 5T32AR007107. SCE is supported by the Ira & Dian Riklis Food Allergy Research award from FARE, the Sean N. Parker Center for Allergy and Asthma Research and a gift from the Colton Foundation. KMJ reports research funding for related work from the NIAID (U01 AI131344) and through Founders’ Distinguished Professorship in Pediatric Allergy.
Abbreviations:
- GI
gastrointestinal tract
- IgA
Immunoglobulin A
- GALT
Gut-associated lymphoid tissue
- M Cells
Microfold cells
- TGF-ß
transforming growth factor beta
- BLG
ß-lactoglobulin
- DC
Dendritic cell
- CM
Cow’s milk
- IL-10
Interleukin 10
- PP
Peyer’s patches
- Naïve T
naïve T cell
- Treg
regulatory T cell
- Tfh
T follicular helper cell
- Mast
mast cell
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Conflicts of interest: None
References:
- 1.Brandtzaeg P. Food allergy: separating the science from the mythology. Nature reviews.Gastroenterology & hepatology 2010; 7:380–400. [DOI] [PubMed] [Google Scholar]
- 2.Konstantinou GN, Nowak-Wegrzyn A, Bencharitiwong R, Bardina L, Sicherer SH, Sampson HA. Egg-white-specific IgA and IgA2 antibodies in egg-allergic children: is there a role in tolerance induction? Pediatr Allergy Immunol 2014; 25:64–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zhang B, Liu E, Gertie JA, Joseph J, Xu L, Pinker EY, et al. Divergent T follicular helper cell requirement for IgA and IgE production to peanut during allergic sensitization. Sci Immunol 2020; 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Jarvinen KM, Westfall JE, Seppo MS, James AK, Tsuang AJ, Feustel PJ, et al. Role of maternal elimination diets and human milk IgA in the development of cow’s milk allergy in the infants. Clin Exp Allergy 2014; 44:69–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Jarvinen KM, Westfall J, De Jesus M, Mantis NJ, Carroll JA, Metzger DW, et al. Role of Maternal Dietary Peanut Exposure in Development of Food Allergy and Oral Tolerance. PLoS One 2015; 10:e0143855. [DOI] [PMC free article] [PubMed] [Google Scholar]