

Abstract
Purpose
Glaucoma remains a poorly understood disease, and identifying biomarkers for early diagnosis is critical to reducing the risk of glaucoma-related visual impairment and blindness. The aim of this review is to provide current metabolic profiles for glaucoma through a summary and analysis of reported metabolites associated with glaucoma.
Methods
We searched PubMed and Web of Science for metabolomics studies of humans on glaucoma published before November 11, 2020. Studies were included if they assessed the biomarkers of any types of glaucoma and performed mass spectrometry-based or nuclear magnetic resonance–based metabolomics approach. Pathway enrichment analysis and topology analysis were performed to generate a global view of metabolic signatures related to glaucoma using the MetaboAnalyst 3.0.
Results
In total, 18 articles were included in this review, among which 13 studies were focused on open-angle glaucoma (OAG). Seventeen metabolites related to OAG were repeatedly identified, including seven amino acids (arginine, glycine, alanine, lysine, methionine, phenylalanine, tyrosine), two phosphatidylcholine (PC aa C34:2, PC aa C36:4), three complements (acetylcarnitine, propionylcarnitine, butyrylcarnitine), carnitine, glutamine, hypoxanthine, spermine, and spermidine. The pathway analysis implied a major role of amino metabolism in OAG pathophysiology and revealed the metabolic characteristics between different biological samples.
Conclusions
In this review, we summarize existing metabolomic studies related to glaucoma biomarker identification and point out a series of metabolic disorders in OAG patients, providing information on the molecular mechanism changes in glaucoma. Additional studies are needed to validate existing findings, and future research will need to explore the potential overlap between different biological fluids.
Keywords: glaucoma, metabolomics, potential biomarkers, metabolic pathways
Glaucoma is a main cause of irreversible blindness, contributing to about 12% of all global blindness.1 It is estimated that the global prevalence of glaucoma in adults aged 40 to 80 years is 3.54%.2 The number of people with glaucoma worldwide is projected to increase to 111.8 million in 2040, disproportionally affecting Asians and Africans.2 Disease risk factors, such as increasing age, elevated intraocular pressure, inflammation, high myopia, and family history have been reported to be associated with glaucoma.3,4 However, glaucoma is generally asymptomatic at early stages and always leads to severe vision impairment at advanced stages. Thus identifying biomarkers for early diagnosis of glaucoma or new therapeutic targets is critical to reducing the risk of visual impairment and blindness from the disease.5,6
Metabolomics is the study of metabolic products that all cellular processes leave behind in a biological sample7 and could be used as a complement to the currently available diagnostic, classification, and progression detection tools.8 In the past few years, metabolomics has been proved to be useful in the area of biomarker discovery for diseases including cancers, cardiovascular diseases, and endocrine diseases.9–12 In ophthalmology, metabolomics has been applied to investigate metabolic signatures and potential biomarkers for various eye diseases as well,13–15 and exploratory studies have suggested metabolic alterations in vitamin metabolism, fatty acid oxidation, and glutaminolysis in patients with glaucoma.16–18 However, the current application of glaucoma metabonomics research is limited by the sample size of the study, the lack of validation of results, the evaluation of long-term markers, and time-efficient and cost-effective analysis techniques. We conducted this systematic review to summarize the current metabolic profiles of glaucoma in humans and to find potential glaucoma biomarkers. Although the values of many differential metabolites and metabolic pathways are not completely confirmed, the results may be helpful for understanding the disease pathophysiology, as well as developing target-based drugs and susceptibility biomarkers for diagnostic applications for this vision-threatening eye disease.
Methods
Search Strategy
We performed a systematic search of electronic databases such as PubMed and Web of Science to identify metabolomics studies on glaucoma published before 11st November 2020 using the keywords: “metabolomics” or “metabonomics” or “metabolic profiling” or “metabolic profiles” or “metabolic biomarkers” or “metabolome” and “glaucoma.” Additional sources were obtained from the reference lists of the included literature. All articles, including those obtained from electronic databases and through cross-referencing, were independently searched and scanned by two authors (Y.W. and X.W.H.) to determine their suitability for inclusion in the review. Disagreements were addressed by discussion with a senior author (C.W.P.).
Inclusion and Exclusion Criteria
Studies were eligible for inclusion if they assessed the biomarker of any types of glaucoma and performed mass spectrometry (MS)–based or nuclear magnetic resonance (NMR)–based metabolomics approach on human beings. We excluded animal studies, vitro model studies, review articles, abstracts without full texts, and the reports of drug therapy response and other experiments. Moreover, only literature published in English was considered.
Quality Assessment
Based on QUADOMIC described by Lumbreras et al.,19 the quality of the studies was assessed with a 16-item scale. The criteria include sample characteristics, differential conditions in preanalytical, clinical, and physiological characteristics of research subjects, and overfitting during research and analysis. Studies achieved with 70% or more scores were included in this review.
Data Extraction and Pathway Analysis
A review of the findings of studies included in the final list of the sources was performed, including study subjects and their essential information (i.e., age, gender ratio, and study area), the types of glaucoma, biological samples, detection and analysis platforms, sample size, and creating a summary of major metabolites they concluded. The pathway analysis was derived from integrating of metabolites from different biological sample sources using the MetaboAnalyst software (version 3.0) on the basis of Kyoto Encyclopedia of Genes and Genomes and Human Metabolome Database. Studies with negative results were excluded from pathway analysis.
Results
Characteristics of the Included Studies
Eighteen studies were identified on the basis of the inclusion and exclusion criteria.17,18,20–35 The flow chart for selection of the studies included in this review is shown in Figure 1. All the studies were designed as case-control studies with the sample sizes ranging from 19 to 506. Among the 18 studies, 13 focused on open-angle glaucoma (OAG),17,20–31 two treated any glaucoma as cases,18,35 and the other three investigated primary angle-closure glaucoma (PACG), primary congenital glaucoma (PCG), and pseudoexfoliation glaucoma, respectively.32–34 Among the 13 studies on OAG, four were performed with aqueous humor (AH) samples,20–23 three with serum samples,24–26 three with plasma samples,17,27,28 and the other three with tear samples,29 occipital brain region,30 and optic nerve,31 respectively. Characteristics of the studies are summarized in Table 1.
Figure 1.

Flow chart of the study screening.
Table 1.
Basic Characteristics of Literature Included
| First Authors (Year) | Country | Case (n) | Co ntrol (n) | Sample | Mean Age, Year Case/Control | F/M Case/Control |
|---|---|---|---|---|---|---|
| Cabrerizo (2017)20 | Spain | OAG (10) | Myopia (10) | AH | 68.8:55.9 | 6/4:6/4 |
| Buisset (2019)21 | France | POAG (26) | Cataract (26) | AH | 74.69:74.92 | 12/14:12/14 |
| Myer (2020)22 | USA | POAG (23) | Cataract (35) | AH | 73.74:70.71 | 3/20:0/35 |
| Pan (2020)23 | China | POAG (16) | Cataract (24) | AH | 72.5:74.2 | 9/16:15/9 |
| Burgess (2015)24 | USA | POAG (72) | Healthy (72) | Plasma | 67.8:68.5 | 45/27:41/31 |
| Leruez (2018)25 | France | POAG (36) | Cataract (27) | Plasma | 72.00:73.04 | 21/15:12/15 |
| Kouassi Nzoughet (2019)26 | France | POAG (34) | Cataract (30) | Plasma | — | — |
| Javadiyan (2012)27 | Australia | POAG (211) | Healthy (295) | Serum | 78:76 | 100/111:160/135 |
| Umeno (2019)17 | Japan | POAG (198) | Control (119) | Serum | 70.4:70.6 | 106/92:83/36 |
| Gong (2020)28 | China | POAG (30) | Healthy (30) | Serum | 54.77:53.80 | 16/14:16/14 |
| Rossi (2019)29 | Italy | POAG (16) | Healthy (17) | Tear | 64.63:61.53 | 9/7:10/7 |
| Boucard (2007)30 | Netherlands | POAG (7) | Control (12) | Occipital brain region | 73:62 | 1/6:4/8 |
| Sidek (2016)31 | USA | Mild POAG (15) Severe POAG (15) | Healthy (15) | ON | 64.9&69.7:53.9 | 9/6&6/9:3/12 |
| Rong (2017)32 | China | PACG (38) | Healthy (48) | Serum | 60.45:60.25 | 19/19:24/24 |
| Chen (2019)33 | China | PCG (30) | Cataract (20) | AH | — | — |
| Myer (2020)34 | USA | PEXG (31) | Cataract (25) | AH | — | — |
| Barbosa Breda (2020)35 | Belgium | Glaucoma (54) | Cataract (29) | AH | 71:75 | 38/16:16/13 |
| Doganay (2012)18 | USA | Glaucoma (29) | Healthy (13) | LGB & Vitreous body | 65.8:62.8 | 13/16:7/6 |
PACG, primary angle-closure glaucoma; PEXG, pseudoexfoliation glaucoma; PCG, primary congenital glaucoma; ON, optic nerve; LGB, lateral geniculate body.
Quality of the Included Studies
The results of the quality assessment on studies that have identified significant differential metabolites using the QUADOMICS tool are shown in Supplementary Table S1. All these diagnostic studies assessed using QUADOMICS in this systematic review were phase I. Thirteen studies presented clear inclusion and exclusion criteria for study subjects.17,20–25,27,28,32–35 Five studies failed to provide clinical and physiological factors in detail.22,23,25,33,34 Ten studies described their approach in dealing with overfitting.17,20–23,25–27,34,35
Metabolic Biomarkers Identified for Glaucoma
All studies included in this review clearly described the analytical platforms on which the studies were performed and the changes of significant differential metabolites identified. Thirteen studies on OAG applied MS analytical platforms, consisting of eight studies using liquid chromatogram–mass spectrometry (LC-MS),17,20,21,24–27,29 two studies using gas chromatography mass spectrometry (GC-MS),23,28 and one study using LC-MS combined with NMR platform.22 In addition, two studies performed magnetic resonance spectroscopy examination by placing region of interest into occipital brain region30 and optic nerve,31 respectively. Rong et al.32 and Chen et al.33 adopted GC-MS to study the metabolomics on primary angle-closure glaucoma and primary congenital glaucoma, respectively, whereas Barbosa Breda et al.35 and Doganay et al.18 chose NMR and magnetic resonance spectroscopy as the identification method for any glaucoma, respectively. In addition, Myer et al.34 concerned the metabolic portrait of pseudoexfoliation by a combination of LC-MS and NMR. Reported alterations in the levels of some metabolites in glaucoma that could be potential candidate biomarkers for clinical practice are summarized in Supplementary Table S2, and the available fold-changes for differential metabolites of glaucoma to controls are listed in Supplementary Table S3. In total, 144 differential metabolites were extracted from 18 included studies.
Seventeen high-frequency (reported twice or more) differential metabolites extracted from 11 studies on OAG17,20–29 are displayed in Table 2. Arginine is the most significant metabolite that has been repeatedly reported up to four times,22,25,26,29 followed by glycine, which is reported with three hits.21,22,28 However, their changing orientation had some disagreements in different biological samples. The increasing of butyrylcarnitine (C4), lysine, propionylcarnitine (C3), PC aa C34:2 and PC aa C36:4, and the decreasing of phenylalanine, spermine, and spermidine were consistently observed in different studies. Also, alanine, acetylcarnitine (C2), carnitine, glutamine, hypoxanthine, methionine, and tyrosine, which were hit in two studies, are important differential metabolites. Rong et al.32 compared the predictive performance using palmitoleic acid, linoleic acid, gamma-linolenic acid, arachidonic acid and four combined metabolites, which generated area under receiver operating characteristic values of 0.724, 0.742, 0.797, 0.673, and 0.867, respectively. Leruez et al.25 used metabolites with the most important coefficients obtained with the least absolute shrinkage and selection operator method to distinguish cases and control, which exhibited an area under receiver operating characteristic of 95.71% in the training set and 91.78% in the validation set.
Table 2.
High-Frequency Differential Metabolites Related to OAG
| Metabolite Name | HMDB ID | Hits | Biological Samples to Be Analyzed |
|---|---|---|---|
| Arginine | HMDB0000517 | 4 | (↑): AH22, Plasma25,26; (↓): Tear29 |
| Glycine | HMDB0000123 | 3 | (↑): AH21,22; (↓): Serum28 |
| Alanine | HMDB0000161 | 2 | (↑): AH21; (↓): Tear29 |
| Acetylcarnitine | HMDB0000201 | 2 | (↑): AH21; (↓): Tear29 |
| Butyrylcarnitine | HMDB0002013 | 2 | (↑): AH21, Plasma25 |
| Carnitine | HMDB0000062 | 2 | (↑): AH21; (↓): Tear29 |
| Glutamine | HMDB0000641 | 2 | (↑): AH21; (↓): AH22 |
| Hypoxanthine | HMDB0000157 | 2 | (↑): Plasma26; (↓): Serum28 |
| Lysine | HMDB0000182 | 2 | (↑): AH22, Serum28 |
| Methionine | HMDB0000696 | 2 | (↑): Plasma25; (↓): Tear29 |
| Propionylcarnitine | HMDB0000824 | 2 | (↑): AH21, Plasma25 |
| PC aa C34:2 | HMDB0007880 | 2 | (↑): AH21, Plasma25 |
| PC aa C36:4 | HMDB0007982 | 2 | (↑): AH21, Plasma25 |
| Phenylalanine | HMDB0000159 | 2 | (↓): AH22, Tear29 |
| Spermine | HMDB0001256 | 2 | (↓): AH21, Plasma25 |
| Spermidine | HMDB0001257 | 2 | (↓): Plasma25, AH23 |
| Tyrosine | HMDB0000158 | 2 | (↑): Plasma25; (↓): Tear29 |
Pathway Analysis
We integrated the metabolomic information of 11 studies on OAG17,20–29 and detailed information of the pathways enriched at the significance level of 0.05 is presented in Supplementary Tables S4, S5, S6 and S7. In this analysis, 52, 24, nine, and nine differential metabolites, which were reported in AH, plasma, serum and tear, respectively, were imported to MetaboAnalyst for pathway enrichment analysis and pathway topology analysis to generate a global view of metabolic signatures of OAG. As a result, 11 metabolic pathways related to AH were significantly enriched, including taurine and hypotaurine metabolism; glycine, serine, and threonine metabolism; and alanine, aspartate, and glutamate metabolism (Fig. 2). In terms of studies using plasma samples, four metabolic pathways, including sphingolipid metabolism, arginine and proline metabolism, and beta-alanine metabolism were significantly enriched (Fig. 3). Meanwhile, glutathione metabolism, glyoxylate and dicarboxylate metabolism, aminoacyl-tRNA biosynthesis, and biotin metabolism were significantly enriched in studies using serum (Fig. 4). Regarding the analysis performed in metabolites identified in tear samples, five metabolic pathways were significantly enriched, among which phenylalanine, tyrosine, and tryptophan biosynthesis showed the greatest impacts (Fig. 5).
Figure 2.
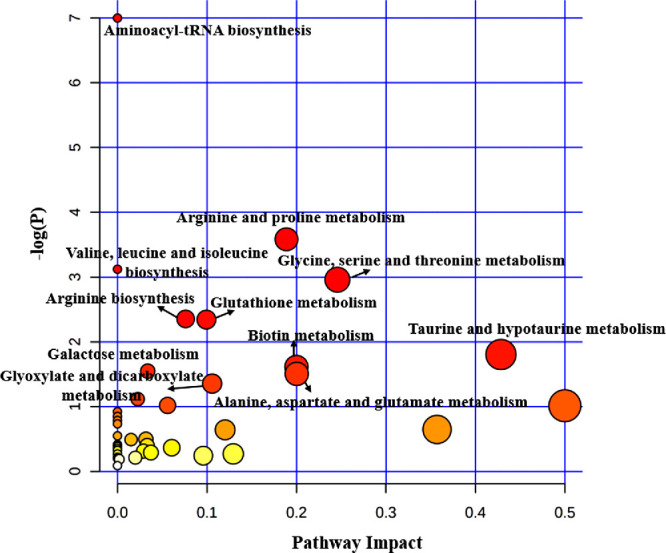
Pathway analysis for significant metabolites of OAG using AH.
Figure 3.
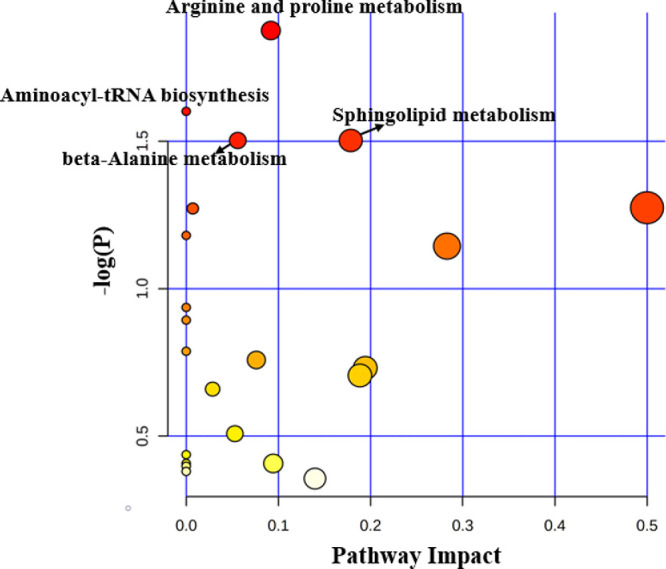
Pathway analysis for significant metabolites of OAG using plasma.
Figure 4.
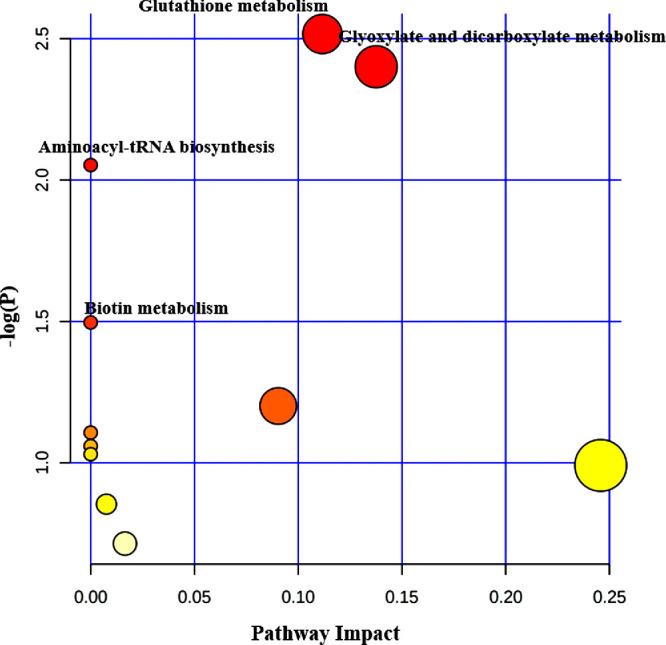
Pathway analysis for significant metabolites of OAG using serum.
Figure 5.
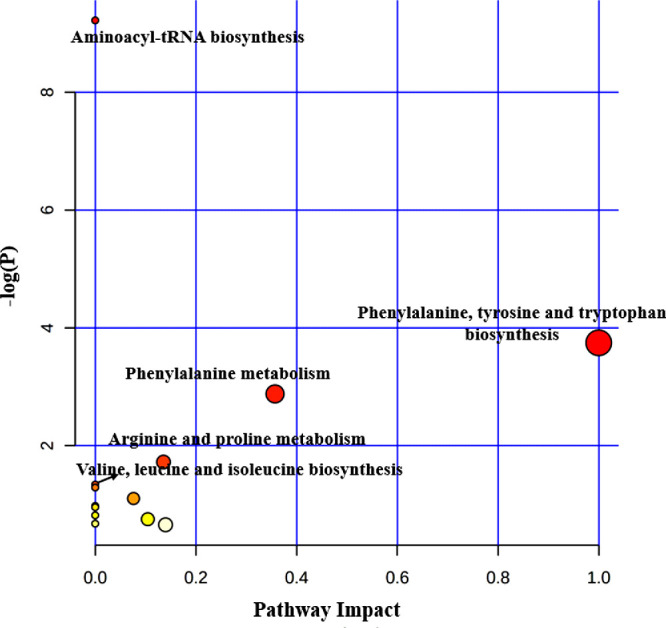
Pathway analysis for significant metabolites of OAG using tear.
Notably, aminoacyl-tRNA biosynthesis was significantly enriched in all four biological samples. Also, we observed arginine and proline metabolism consistently enriched in AH, plasma, and tear samples. In addition, we found three pathways (glutathione metabolism, glyoxylate and dicarboxylate metabolism, and biotin metabolism) significantly enriched in both AH and serum samples, and valine, leucine, and isoleucine biosynthesis pathway were significantly enriched in both AH and tear samples.
Discussion
In this review, we summarized 15 metabolomics studies on glaucoma and took a look at the profiling of valuable metabolic pathways of OAG. As a result, 17 metabolites identified in at least two studies were listed to be high-frequency differential metabolites of OAG, and arginine is the most promising metabolic biomarker with the highest rate of repetition. In the pathway analysis, we found some metabolism pathways that were significantly enriched in different samples, suggesting a series of metabolic dysregulations in OAG patients. Considering that glaucoma remains a disorder with limited therapeutic options, the findings of this review contribute to the further understanding of the molecular characteristics of disease, which will contribute to the understanding of disease mechanisms.
Metabolic Dysregulations Implicated in OAG Pathogenic Mechanisms
According to the 17 high-frequency differential metabolites and the 24 significantly enriched metabolism pathways, amino acid dysregulation was the most prominent feature of OAG. Arginine is an important integral in the pathway of arginine and proline metabolism, which has been reported to be involved in the nitric oxide production pathway. Nitric oxide is an essential antioxidant, regulating AH dynamics, ocular blood flow, neuronal visual processing, and the apoptosis of retinal ganglion cell death.36,37 In addition, the disorder of spermidine and spermine may be the reason for the disorder of methionine and arginine, as they are both precursors of the biosynthesis of spermidine and spermine. Previous reports have already identified the role of spermidine and spermine as a metabolic regulator of mitochondria with regard to apoptosis and the mitochondrial membrane potential and one of the central elements of the POAG signature.25,38 Likewise, glycine was also identified as a high-frequency biomarker. A few studies have revealed that the level of glycine is associated with neurotransmission on the retina, and the dual effect of glycine in both inhibitory and excitatory neurotransmission is based on the precise regulation of its extracellular concentration.39,40 Moreover, it was also indicated that glycine could participate in cell oxidative stress by inducing lipid peroxidation and decreasing glutathione concentrations.41
Biotin metabolism was also shown to be crucial for the development of OAG. As a cofactor for five carboxylases, biotin is thought to be involved not only in fatty acid biosynthesis, gluconeogenesis, and catabolism of amino acids and fatty acids42 but also in the maintenance of the normal structure and function of mitochondria.43 Chronic complement activation has been suggested to be a significant feature seen in glaucoma.44 This is consistent with our results that C2, C3, and C4, which are the activating unit of complements, were repeatedly identified in different studies. In addition, although aminoacyl-tRNA biosynthesis was significantly enriched in the four biofluids, its biological interpretation is still obscure. The mechanism of this metabolism pathway in the development of glaucoma might be related to the regulatory role of aminoacyl-tRNA synthetases in both cytosolic and mitochondrial translation.45
Metabolic Changes Among Different Biological Samples
The levels of differential metabolites showed differences in the direction of change in different samples. According to our summary, the high-frequency metabolites of AH have always tended to change in the opposite direction to tears. According to the analysis of Csosz et al.,46 tear was different from AH in terms of the proinflammatory mechanism point of view, so that tear could not substitute for aqueous humor for diagnostic purposes. Interestingly, even in blood samples, the differential metabolites in plasma were not identical to that in serum. Several metabolites, including amino acids, glycerophospholipids, lipoproteins, and energy pathway metabolites, have been reported to be distinguished in the serum and plasma samples.47–49 It might be explained by the influence of distinct collecting methods, different biochemical processing, and different constituents such as platelet-derived metabolites.49 However, it is well known that some circulating metabolites and eye metabolites could be connected either through the blood retina barrier or through sharing common mechanisms, such as by the same enzymes. AH is a transparent fluid present in the anterior chamber of the eye that provides nutrition and eliminates the metabolic waste moving from the eye to the blood by circulating through the vascular and lymphatic systems. This could partly explain the great synchronous variation trend of high-frequency differential metabolites from AH and plasma.
Biofluid testing is a less-damaging diagnostic method, compared to histological testing. Among the commonly used biofluids in ophthalmology, AH is perhaps the most closely linked to the pathological tissue and thus could provide information on normal and pathological conditions. However, AH is usually collected during an invasive eye surgery, which has a risk of complications (e.g., persistent diplopia and corneal edema),50 preventing its application for the purpose of screening or follow-up. Blood is also a valuable biological sample for metabolomics because it is easy to collect, and plasma seems to be highly reproducible.51 In addition, tears provide a more available and noninvasive choice for collection, although the sampling procedure still needs to improve. In all, because these different biological samples possess both shared and unique components, these differences may be an important consideration for biofluid selection and metabolite identification in metabolic phenotyping studies.
Clinical Implications of Glaucoma Using Metabolomics
Glaucoma is an age-related disease that seriously threatens the vision of the elderly. In light of the growing number of senior citizens in many countries, the disease burden of glaucoma is increasing, and a large proportion of this population has undetected or untreated glaucoma.52,53 Tang et al.54 performed decision-analytic Markov models to evaluate costs and benefits of glaucoma screening and found that community-based screening for glaucoma is likely to be cost-effective in both urban and rural settings.54 Metabolomics provides a promising method of analyzing accessible biological samples for the investigation of glaucoma biomarkers in predictive, preventive, and personalized medicine. The advancements of NMR and MS have enabled us to detect more and more metabolites and thus helped us obtain a comprehensive view on the metabolic changes in glaucoma. Meanwhile, the increasing understanding of metabolism pathways could promote an understanding of the mechanisms of glaucoma disease. Although lowering the intraocular pressure is regarded as the most efficacious and acceptable form of treatment,55 there are other mechanisms that might lead to glaucoma. Besides, the interaction between many natural substances and specific metabolic pathways could arouse the emerging of drug discovery and adjuvant therapy for treatment of glaucoma.56
Limitations and Future Direction
The current reports of metabolic alterations serve significant clues to the early diagnostic capabilities. However, several limitations of the existing metabolomics studies on glaucoma should be addressed. First, different researches chose differential metabolites using different methods, including metabolomics technologies and statistical analysis, resulting in difficulty in replication and bias in the identification of potential biomarkers. Second, most studies did not provide quantitative information on metabolites, so we couldn't integrate the changes in metabolite concentration associated with glaucoma risk here. Third, all the studies included were cross-sectional, thus limiting the power of verifying causal relationships between differential metabolites and glaucoma. Fourth, most studies focused on OAG, and there is a lack of data on other types of glaucoma.
Application of metabolomics on glaucoma is a field that can benefit enormously from personalized medicine.1 However, the development of glaucoma metabolomics has sadly lagged behind compared to other fields such as oncology. More studies, especially longitudinal cohort metabolomics and clinical intervention metabolomics studies, should be carried out to explore novel biomarkers of glaucoma and determine target metabolic pathways for prevention and treatment. Besides, we should pay more attention to the potential overlap or interaction of metabolism in different biological samples to enhance our understanding of the complex mechanisms underpinning glaucoma pathology. Furthermore, integrating metabolomics with other omics platforms, including genomics, transcriptomics, and proteomics, will provide services for precision medicine.
Conclusions
To sum up, several metabolites and metabolic pathways have been indicated to be associated with OAG in this review. These high-frequency differential metabolites are promising to as biomarkers of OAG for early diagnosis, and the prominent metabolic pathway may provide new idea for the explanation of the pathogenesis and the discovery of new therapeutic targets. However, applying these potential biomarkers to clinical diagnosis and treatment is still a challenge for us. More validation studies with larger numbers of cases are warranted to confirm the findings and exploring the potential overlap between different biofluids. Moreover, metabolomics studies on other glaucoma subtypes should be encouraged to make a precise signature for glaucoma detection and treatment.
Supplementary Material
Acknowledgments
Supported by the National Natural Science Foundation of China [Grant Number: 81973061], the Training Program for Medical Leaders of Yunnan Health Commission [Grant Number: D-2018016] and the Tang Scholar of Soochow University.
Disclosure: Y. Wang, None; X.-W. Hou, None; G. Liang, None; C.-W. Pan, None
References
- 1. Gungor K, Hotez PJ, Ozdemir V, Aynacioglu S.. Glaucomics: a call for systems diagnostics for 21(st) century ophthalmology and personalized visual health. OMICS. 2014; 18: 275–279. [DOI] [PubMed] [Google Scholar]
- 2. Tham YC, Li X, Wong TY, Quigley HA, Aung T, Cheng CY.. Global prevalence of glaucoma and projections of glaucoma burden through 2040: a systematic review and meta-analysis. Ophthalmology. 2014; 121: 2081–2090. [DOI] [PubMed] [Google Scholar]
- 3. Schuster AK, Erb C, Hoffmann EM, Dietlein T, Pfeiffer N.. The Diagnosis and Treatment of Glaucoma. Dtsch Arztebl Int. 2020; 117: 225–234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Evangelho K, Mogilevskaya M, Losada-Barragan M, Vargas-Sanchez JK.. Pathophysiology of primary open-angle glaucoma from a neuroinflammatory and neurotoxicity perspective: a review of the literature. Int Ophthalmol. 2019; 39: 259–271. [DOI] [PubMed] [Google Scholar]
- 5. Nath M, Halder N, Velpandian T.. Circulating biomarkers in glaucoma, age-related macular degeneration, and diabetic retinopathy. Indian J Ophthalmol. 2017; 65: 191–197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Weinreb RN, Aung T, Medeiros FA.. The pathophysiology and treatment of glaucoma: a review. JAMA. 2014; 311: 1901–1911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Trivedi DK, Hollywood KA, Goodacre R.. Metabolomics for the masses: The future of metabolomics in a personalized world. New Horiz Transl Med. 2017; 3: 294–305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Barbosa-Breda J, Himmelreich U, Ghesquiere B, Rocha-Sousa A, Stalmans I.. Clinical Metabolomics and Glaucoma. Ophthalmic Res. 2018; 59: 1–6. [DOI] [PubMed] [Google Scholar]
- 9. Kelly RS, Vander Heiden MG, Giovannucci E, Mucci LA.. Metabolomic Biomarkers of Prostate Cancer: Prediction, Diagnosis, Progression, Prognosis, and Recurrence. Cancer Epidemiol Biomarkers Prev. 2016; 25: 887–906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Rodrigues D, Jeronimo C, Henrique R, et al.. Biomarkers in bladder cancer: A metabolomic approach using in vitro and ex vivo model systems. Int J Cancer. 2016; 139: 256–268. [DOI] [PubMed] [Google Scholar]
- 11. Qureshi MI, Vorkas PA, Coupland AP, Jenkins IH, Holmes E, Davies AH.. Lessons from Metabonomics on the Neurobiology of Stroke. Neuroscientist. 2017; 23: 374–382. [DOI] [PubMed] [Google Scholar]
- 12. Sun Y, Gao HY, Fan ZY, He Y, Yan YX.. Metabolomics Signatures in Type 2 Diabetes: A Systematic Review and Integrative Analysis. J Clin Endocrinol Metab. 2020; 105: dgz240. [DOI] [PubMed] [Google Scholar]
- 13. Dai L, Yang W, Qin X, et al.. Serum metabolomics profiling and potential biomarkers of myopia using LC-QTOF/MS. Exp Eye Res. 2019; 186: 107737. [DOI] [PubMed] [Google Scholar]
- 14. Liu K, Fang J, Jin J, et al.. Serum Metabolomics Reveals Personalized Metabolic Patterns for Macular Neovascular Disease Patient Stratification. J Proteome Res. 2020; 19: 699–707. [DOI] [PubMed] [Google Scholar]
- 15. Hou XW, Wang Y, Pan CW.. Metabolomics in Age-Related Macular Degeneration: A Systematic Review. Invest Ophthalmol Vis Sci. 2020; 61: 13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Hysi PG, Khawaja AP, Menni C, et al.. Ascorbic acid metabolites are involved in intraocular pressure control in the general population. Redox Biol. 2019; 20: 349–353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Umeno A, Tanito M, Kaidzu S, Takai Y, Horie M, Yoshida Y.. Comprehensive measurements of hydroxylinoleate and hydroxyarachidonate isomers in blood samples from primary open-angle glaucoma patients and controls. Sci Rep. 2019; 9: 2171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Doganay S, Cankaya C, Alkan A.. Evaluation of corpus geniculatum laterale and vitreous fluid by magnetic resonance spectroscopy in patients with glaucoma; a preliminary study. Eye (Lond). 2012; 26: 1044–1051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Lumbreras B, Porta M, Marquez S, Pollan M, Parker LA, Hernandez-Aguado I.. QUADOMICS: an adaptation of the Quality Assessment of Diagnostic Accuracy Assessment (QUADAS) for the evaluation of the methodological quality of studies on the diagnostic accuracy of '-omics'-based technologies. Clin Biochem. 2008; 41: 1316–1325. [DOI] [PubMed] [Google Scholar]
- 20. Cabrerizo J, Urcola JA, Vecino E.. Changes in the lipidomic profile of aqueous humor in open-angle glaucoma. J Glaucoma. 2017; 26: 349–355. [DOI] [PubMed] [Google Scholar]
- 21. Buisset A, Gohier P, Leruez S, et al.. Metabolomic profiling of aqueous humor in glaucoma points to taurine and spermine deficiency: findings from the Eye-D Study. J Proteome Res. 2019; 18: 1307–1315. [DOI] [PubMed] [Google Scholar]
- 22. Myer C, Perez J, Abdelrahman L, et al.. Differentiation of soluble aqueous humor metabolites in primary open angle glaucoma and controls. Exp Eye Res. 2020; 194: 108024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Pan CW, Ke C, Chen Q, et al.. Differential metabolic markers associated with primary open-angle glaucoma and cataract in human aqueous humor. BMC Ophthalmol. 2020; 20: 183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Burgess LG, Uppal K, Walker DI, et al.. Metabolome-wide association study of primary open angle glaucoma. Invest Ophthalmol Vis Sci. 2015; 56: 5020–5028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Leruez S, Marill A, Bresson T, et al.. A metabolomics profiling of glaucoma points to mitochondrial dysfunction, senescence, and polyamines deficiency. Invest Ophthalmol Vis Sci. 2018; 59: 4355–4361. [DOI] [PubMed] [Google Scholar]
- 26. Kouassi Nzoughet J, Guehlouz K, Leruez S, et al.. A data mining metabolomics exploration of glaucoma. Metabolites. 2020; 10(2): 49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Javadiyan S, Burdon KP, Whiting MJ, et al.. Elevation of serum asymmetrical and symmetrical dimethylarginine in patients with advanced glaucoma. Invest Ophthalmol Vis Sci. 2012; 53: 1923–1927. [DOI] [PubMed] [Google Scholar]
- 28. Gong H, Zhang S, Li Q, et al.. Gut microbiota compositional profile and serum metabolic phenotype in patients with primary open-angle glaucoma. Exp Eye Res. 2020; 191: 107921. [DOI] [PubMed] [Google Scholar]
- 29. Rossi C, Cicalini I, Cufaro MC, et al.. Multi-Omics Approach for Studying Tears in Treatment-Naive Glaucoma Patients. Int J Mol Sci. 2019; 20: 4029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Boucard CC, Hoogduin JM, van der Grond J, Cornelissen FW.. Occipital proton magnetic resonance spectroscopy (1H-MRS) reveals normal metabolite concentrations in retinal visual field defects. PLoS One. 2007; 2: e222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Sidek S, Ramli N, Rahmat K, Ramli NM, Abdulrahman F, Kuo TL.. In vivo proton magnetic resonance spectroscopy (1H-MRS) evaluation of the metabolite concentration of optic radiation in primary open angle glaucoma. Eur Radiol. 2016; 26: 4404–4412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Rong S, Li Y, Guan Y, et al.. Long-chain unsaturated fatty acids as possible important metabolites for primary angle-closure glaucoma based on targeted metabolomic analysis [published online ahead of print March 26, 2017]. Biomed Chromatogr, 10.1002/bmc.3963. [DOI] [PubMed] [Google Scholar]
- 33. Chen X, Chen Y, Wang L, Sun X.. Metabolomics of the aqueous humor in patients with primary congenital glaucoma. Mol Vis. 2019; 25: 489–501. [PMC free article] [PubMed] [Google Scholar]
- 34. Myer C, Abdelrahman L, Banerjee S, et al.. Aqueous humor metabolite profile of pseudoexfoliation glaucoma is distinctive. Mol Omics. 2020; 16: 425–435. [DOI] [PubMed] [Google Scholar]
- 35. Barbosa Breda J, Croitor Sava A, Himmelreich U, et al.. Metabolomic profiling of aqueous humor from glaucoma patients - The metabolomics in surgical ophthalmological patients (MISO) study. Exp Eye Res. 2020; 201: 108268. [DOI] [PubMed] [Google Scholar]
- 36. Becquet F, Courtois Y, Goureau O.. Nitric oxide in the eye: multifaceted roles and diverse outcomes. Surv Ophthalmol. 1997; 42: 71–82. [DOI] [PubMed] [Google Scholar]
- 37. Koss MC. Functional role of nitric oxide in regulation of ocular blood flow. Eur J Pharmacol. 1999; 374: 161–174. [DOI] [PubMed] [Google Scholar]
- 38. Minois N, Carmona-Gutierrez D, Madeo F.. Polyamines in aging and disease. Aging (Albany NY). 2011; 3: 716–732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Coull BM, Cutler RW.. Light-evoked release of endogenous glycine into the perfused vitreous of the intact rat eye. Invest Ophthalmol Vis Sci. 1978; 17: 682–684. [PubMed] [Google Scholar]
- 40. Pourcho RG. Neurotransmitters in the retina. Curr Eye Res. 1996; 15: 797–803. [DOI] [PubMed] [Google Scholar]
- 41. Moura AP, Grings M, Marcowich GF, et al.. Evidence that glycine induces lipid peroxidation and decreases glutathione concentrations in rat cerebellum. Mol Cell Biochem. 2014; 395: 125–134. [DOI] [PubMed] [Google Scholar]
- 42. Said HM. Biotin: biochemical, physiological and clinical aspects. Subcell Biochem. 2012; 56: 1–19. [DOI] [PubMed] [Google Scholar]
- 43. Ochoa-Ruiz E, Diaz-Ruiz R, Hernandez-Vazquez Ade J, et al.. Biotin deprivation impairs mitochondrial structure and function and has implications for inherited metabolic disorders. Mol Genet Metab. 2015; 116: 204–214. [DOI] [PubMed] [Google Scholar]
- 44. Mohlin C, Sandholm K, Kvanta A, Ekdahl KN, Johansson K.. A model to study complement involvement in experimental retinal degeneration. Ups J Med Sci. 2018; 123: 28–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Boczonadi V, Horvath R.. Mitochondria: impaired mitochondrial translation in human disease. Int J Biochem Cell Biol. 2014; 48: 77–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Csosz E, Deak E, Toth N, Traverso CE, Csutak A, Tozser J.. Comparative analysis of cytokine profiles of glaucomatous tears and aqueous humour reveals potential biomarkers for trabeculectomy complications. FEBS Open Bio. 2019; 9: 1020–1028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Teahan O, Gamble S, Holmes E, et al.. Impact of analytical bias in metabonomic studies of human blood serum and plasma. Anal Chem. 2006; 78: 4307–4318. [DOI] [PubMed] [Google Scholar]
- 48. Hirayama A, Sugimoto M, Suzuki A, et al.. Effects of processing and storage conditions on charged metabolomic profiles in blood. Electrophoresis. 2015; 36: 2148–2155. [DOI] [PubMed] [Google Scholar]
- 49. Kaluarachchi M, Boulange CL, Karaman I, et al.. A comparison of human serum and plasma metabolites using untargeted (1)H NMR spectroscopy and UPLC-MS. Metabolomics. 2018; 14: 32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Budenz DL, Feuer WJ, Barton K, et al.. Postoperative Complications in the Ahmed Baerveldt Comparison Study During Five Years of Follow-up. Am J Ophthalmol. 2016; 163: 75–82.e73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Yu Z, Kastenmuller G, He Y, et al.. Differences between human plasma and serum metabolite profiles. PLoS One. 2011; 6: e21230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Wu J, Yu X, Ping X, et al.. Socioeconomic disparities in the global burden of glaucoma: an analysis of trends from 1990 to 2016. Graefes Arch Clin Exp Ophthalmol. 2020; 258: 587–594. [DOI] [PubMed] [Google Scholar]
- 53. Shaikh Y, Yu F, Coleman AL.. Burden of undetected and untreated glaucoma in the United States. Am J Ophthalmol. 2014; 158: 1121–1129.e1121. [DOI] [PubMed] [Google Scholar]
- 54. Tang J, Liang Y, O'Neill C, Kee F, Jiang J, Congdon N.. Cost-effectiveness and cost-utility of population-based glaucoma screening in China: a decision-analytic Markov model. Lancet Glob Health. 2019; 7: e968–e978. [DOI] [PubMed] [Google Scholar]
- 55. Heijl A, Leske MC, Bengtsson B, et al.. Reduction of intraocular pressure and glaucoma progression: results from the Early Manifest Glaucoma Trial. Arch Ophthalmol. 2002; 120: 1268–1279. [DOI] [PubMed] [Google Scholar]
- 56. Sacca SC, Corazza P, Gandolfi S, et al.. Substances of Interest That Support Glaucoma Therapy. Nutrients. 2019; 11: 239. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.