

Abstract
In the clinic, differential reinforcement of alternative behavior (DRA) often involves programming extinction for destructive behavior while reinforcing an alternative form of communication (e.g., a functional communication response); however, implementing extinction can be unsafe or impractical under some circumstances. Quantitative theories of resurgence (i.e., Behavioral Momentum Theory and Resurgence as Choice) predict differences in the efficacy of treatments that do and do not involve extinction of target responding when reinforcement conditions maintaining alternative responding worsen. We tested these predictions by examining resurgence following two DRA conditions in which we equated rates of reinforcement. In DRA without extinction, target and alternative behavior produced reinforcement. In DRA with extinction plus noncontingent reinforcement, only alternative behavior produced reinforcement. We conducted this study in a reverse-translation sequence, first with participants who engaged in destructive behavior (Experiment 1) and then in a laboratory setting with rats (Experiment 2). Across both experiments, we observed proportionally lower levels of target responding during and following the DRA condition that arranged extinction for the target response. However, levels of resurgence were similar following both arrangements.
Keywords: behavioral momentum theory, destructive behavior, extinction, differential reinforcement of alternative behavior, resurgence, resurgence as choice, reverse-translational
Differential reinforcement of alternative behavior (DRA) is an effective strategy for treating socially mediated destructive behavior (Greer, Fisher, Saini et al., 2016; Hagopian et al., 1998; Kurtz et al., 2011; Rooker et al., 2013). During DRA, the therapist delivers the functional reinforcer following an alternative response (e.g., a functional communication response [FCR]) often while placing destructive behavior on extinction. Despite several studies showing the extinction component of DRA to be important in reducing destructive behavior (Fisher et al., 2000; Hagopian et al., 1998; Petscher et al., 2009; Shirley et al., 1997), arranging extinction may be unsafe or impractical under some conditions. For example, extinction may be unsafe if it entails ignoring risky (e.g., furniture climbing) or dangerous (e.g., self-injurious) behavior. Likewise, extinction may be impractical if it requires physically guiding an individual of a large physical stature (Athens & Vollmer, 2010) or if destructive behavior (e.g., aggression) is sensitive to small changes in the therapist’s behavior (Fisher, Ninness et al., 1996; Kodak et al., 2007; Piazza et al., 1999). For these reasons, applied researchers have investigated DRA treatments that do not rely on extinction, yet still produce clinically significant reductions in destructive behavior (Athens & Vollmer, 2010; Lalli & Casey, 1996; Lalli et al., 1999; Parrish et al., 1986; Piazza, Fisher et al., 1997; Russo et al., 1981; for a related review, see Payne & Dozier, 2013).
Despite the widespread use and effectiveness of DRA-based interventions (Petscher et al., 2009; Tiger, Hanley, & Bruzek, 2008), questions remain regarding the durability of these interventions over time. Recent translational research on DRA-based interventions has shown that destructive behavior is likely to reemerge under certain conditions, such as when the alternative response fails to produce the functional reinforcer (Berg et al., 2015; Fisher et al., 2020; Fisher, Greer, Fuhrman et al., 2018; Fisher, Saini et al., 2018; Fuhrman et al., 2016; Greer et al., 2020; Hoffman & Falcomata, 2014; Lieving et al., 2004; Mace et al., 2010; Marsteller & St. Peter, 2014; Volkert et al., 2009; Wacker et al., 2011; Wacker et al., 2013) and following other decrements in the rate of alternative reinforcement (Briggs et al., 2018; Volkert et al., 2009). This reemergence of destructive behavior following successful treatment is known as treatment relapse (Pritchard et al., 2014), and researchers are increasingly concerned with how different forms of relapse (e.g., resurgence, renewal, reinstatement) affect clinical care (e.g., Briggs et al., 2018; Craig, Sullivan, Browning et al., 2019; Kimball et al., 2020; Kimball et al., 2018; Podlesnik & DeLeon, 2015). Translational researchers have placed considerable focus on resurgence, or the reemergence of previous behavior resulting from a general worsening of reinforcement conditions (Epstein, 1985; Lattal et al., 2017), as such conditions occur frequently in and outside the clinic.
Quantitative theories of resurgence (i.e., Behavioral Momentum Theory [BMT; Shahan & Sweeney, 2011] and Resurgence as Choice [RaC; Shahan & Craig, 2017]) offer a framework for understanding the behavioral processes responsible for treatment relapse by offering unique insights on resurgence. As such, these theories have been integral to recent translational research on the resurgence of destructive behavior following DRA-based treatments (see Fisher, Greer, Craig et al., 2018 for a summary of four such studies). It is worth noting that the findings from these recent translational studies have often suggested refinements to standards of care for treating destructive behavior, and in some cases the findings directly oppose those standards of care. For example, the recent study by Fisher, Saini et al. (2018) showed that programming a lean schedule of reinforcement for destructive behavior in baseline, as opposed to the widely adopted practice of programming a dense schedule of reinforcement for destructive behavior, better suppressed resurgence when alternative reinforcement was later suspended during an extinction-challenge condition. Such basic-to-applied translations have the potential to inform new standards of care for the treatment of destructive behavior, new standards not possible without the predictions afforded by quantitative theories of relapse.
Despite recent extensions of laboratory findings on resurgence to the clinic, both BMT and RaC make explicit predictions about DRA-based treatments for destructive behavior that have yet to receive empirical scrutiny, yet also have the potential to inform new standards of care. For example, both BMT and RaC predict better suppression of target responding during a resurgence test (Phase 3) when target responding results in extinction during Phase 2 of a three-phase resurgence evaluation than when target responding continues to produce reinforcement. According to BMT, arranging extinction of target responding in Phase 2 severs the response–reinforcer relation established in Phase 1, and reinforcer omission in Phase 2 affects generalization decrement (1985 Nevin et al., 2001; see Greer, Fisher, Romani et al., 2016, for a description within the context of functional communication training). RaC makes a similar prediction but does so based on a different set of assumptions. According to RaC, discontinuing reinforcement for the target response in Phase 2 lessens the relative value of that response in relation to the value of the alternative response, and this lessened relative value has important implications when all reinforcers are later discontinued in Phase 3 (Greer & Shahan, 2019). These quantitative predictions have important implications for practitioners who rely on DRA-based interventions to treat destructive behavior. To our knowledge, no study, basic or applied, has compared resurgence following DRA implemented with and without extinction.
In the present study, we examined this confluence of predictions from BMT and RaC by evaluating whether and to what extent target behavior reemerged following DRA treatments that did and did not arrange extinction. Because both BMT and RaC predict differing levels of resurgence following DRA treatments that deliver different rates of alternative reinforcement, we yoked the rates of alternative reinforcement across the two DRA conditions to isolate the effects of extinction on resurgence. Additionally, we conducted a similar number of sessions of each DRA condition to minimize the likelihood that differences in resurgence were due to different durations of each treatment type, as also suggested by both BMT and RaC. We examined this question in a reverse-translational preparation in which we first conducted Experiment 1 in an applied setting with two children referred for the assessment and treatment of destructive behavior. To better isolate extinction and expand upon the findings of Experiment 1, we conducted Experiment 2 in a laboratory setting with rats.
Experiment 1
Method
Participants and Setting
Two children1 referred for the assessment and treatment of destructive behavior participated. Charlie, an 8-year-old boy, carried the diagnoses of autism spectrum disorder and attention deficit hyperactivity disorder. Charlie displayed aggression, self-injurious behavior, and property destruction. We targeted Charlie’s aggression and self-injurious behavior in the present study and subsequently treated his property destruction. Charlie communicated primarily by guiding adults to preferred materials. Charlie also participated in a study on functional analysis conducted by Fisher et al. (2016). Charlie attended our clinic 2 days per week (Tuesdays and Thursdays) for 3-hr appointments. Dan, an 8-year-old boy, carried the diagnoses autism spectrum disorder and attention deficit and hyperactivity disorder. He displayed aggression, self-injurious behavior, and property destruction. Dan communicated in full sentences. He attended our clinic 3 days per week (Mondays, Wednesdays, and Fridays) for 2-hr appointments. Both participants were on stable drug regimens throughout the study and had no comorbid health conditions.
We conducted all study procedures under the oversight of a pediatrics institutional review board, and we used the session-termination criteria recommended by Betz and Fisher (2011). No session terminated prematurely due to meeting these criteria. We conducted all sessions in 3-m by 3-m padded therapy rooms equipped with a two-way intercom and a one-way observation window. Dan’s therapy room contained chairs, but therapists removed all furniture from Charlie’s therapy room to prevent self-injury directed at hard surfaces. The therapy rooms for both participants included materials relevant for the upcoming session (e.g., preferred tangible items, FCR cards). Following completion of the study, therapists implemented treatment procedures similar to those used within the study and observed a 100% and a 90% reduction in baseline rates of destructive behavior during the final five treatment sessions for Charlie and Dan, respectively.
Response Measurement
Data collectors used laptop computers to collect frequency data for each participant. Aggression (Charlie and Dan) included hitting, kicking, biting, scratching, head-butting, pinching, grabbing, pushing, and throwing objects at the therapist. Property destruction (Dan only) included overturning, hitting, kicking, swiping, and throwing furniture or objects (not directed at the therapist). Self-injurious behavior consisted of head banging, head hitting, and body hitting for Charlie but consisted of self-biting for Dan. Alternative responses (Charlie and Dan) consisted of handing the therapist a card with a picture of the participant consuming the reinforcer that maintained destructive behavior. Data collectors also measured reinforcer frequency, duration, and type (i.e., delivered contingently or noncontingently) during the resurgence evaluation.
Interobserver Agreement and Treatment Integrity
A second independent observer collected data simultaneously with the primary data collector on 47% and 31% of sessions for Charlie and Dan, respectively. We scored an agreement when both observers recorded the same number of responses or seconds of the response within each 10-s interval (i.e., exact agreement within the interval). We calculated interobserver agreement for each session by summing the number of agreement intervals, dividing by the total number of intervals, and converting the resulting quotient into a percentage. For Charlie, agreements averaged 94.1% for aggression (range, 78.3–100%), 94.1% for self-injury (range, 70–100%), and 94.7% for FCRs (range, 81–100%). For Dan, agreements averaged 86.7% for aggression (range, 63.3–100%), 98.7% for self-injury (range, 93.3–100%), 93.3% for property destruction (range, 85–100%), and 99.1% for FCRs (range, 66.6–100%).
We also assessed whether therapists delivered reinforcers according to their programmed duration of 20 s for 36% and 38% of sessions during the resurgence evaluation for Charlie and Dan, respectively. We did this by scoring a reinforcer delivery as correctly implemented if it lasted between 15 s and 25 s (i.e., a 5-s window above and below the target duration). Observers scored reinforcer deliveries lasting fewer than 15 s or greater than 25 s as incorrect. For Charlie, treatment integrity averaged 95% (range, 75–100%), and for Dan, treatment integrity averaged 96% (range, 62–100%).
Procedure
Analyses Informing the Resurgence Evaluation
Functional Analysis.
We first conducted a functional analysis of each participant’s destructive behavior using procedures similar to those described by Iwata et al. (1982/1994), which incorporated (a) screening procedures for automatic reinforcement described by Querim et al. (2013); (b) the fixed sequence of multielement conditions described by Hammond et al. (2013); (c) reinforcer-duration modifications described by Fisher, Piazza and Chiang (1996); and (d) condition-specific colored clothing worn by the therapists described by Conners et al. (2000). We selected items for the tangible and attention conditions of the functional analysis based on the results of a paired-stimulus preference assessment (Fisher et al., 1992), informed by caregiver report (Fisher, Piazza, Bowman & Amari, 1996). Both participants selected an iPad as their most preferred item. The results of the functional analysis demonstrated Charlie and Dan reliably displayed destructive behavior in the tangible condition, suggesting that access to preferred tangible items (i.e., the iPad) reinforced their destructive behavior.
Alternative Response Training.
Following the functional analysis, we taught each participant the alternative response, which consisted of handing the therapist a picture the size of an index card showing the participant playing with the iPad. The therapist guided the alternative response after increasing periods of time without access to the reinforcer for destructive behavior until alternative responding occurred independently. Alternative responses resulted in 20-s access to the iPad. Therapists ignored destructive behavior but implemented a changeover delay (COD; Herrnstein, 1961) to prevent adventitious reinforcement if the FCR occurred within 3 s of destructive behavior. This 3-s COD remained in place across all subsequent DRA conditions.
Reinforcement-Schedule Assessments.
We employed two methods to determine appropriate schedules of intermittent reinforcement for each participant’s alternative response and destructive behavior for the purpose of evaluating resurgence following each of the two DRA arrangements. Because we aimed to isolate the effects of the extinction component of DRA, an ideal schedule of intermittent reinforcement for our purposes would produce high rates of the alternative response and lower, but not zero, rates of destructive behavior. We hoped to ensure that destructive behavior would contact the presence or absence of its reinforcement contingency across the two DRA conditions. Thus, our resurgence evaluation differed somewhat from those reported elsewhere in the literature in that we did not expect destructive behavior to extinguish in one of the two DRA conditions (i.e., DRA without extinction), and as such, we initiated the extinction challenge after a fixed number of DRA sessions rather than awaiting complete suppression of destructive behavior.
We had three additional requirements for the reinforcement schedules programmed in the resurgence evaluation. First, the schedule of reinforcement used in baseline for destructive behavior across the progressions had to be the same as that programmed for destructive behavior in the DRA condition that did not include extinction, thereby minimizing the discriminability that a contingency change had occurred for destructive behavior upon transitioning from baseline to DRA without extinction. Second, both DRA conditions had to arrange the same reinforcement schedule for the alternative response. Third, the anticipated lower rates of alternative reinforcement obtained in the DRA condition that included extinction needed to be offset by delivering reinforcers on a time-based schedule in that condition, thereby yoking the obtained rates of reinforcement to the DRA condition that did not include extinction. We took measures to satisfy these requirements in order to ensure that the two DRA conditions differed only in terms of the presence or absence of the extinction component. However, doing so required us to conduct two reinforcer assessments prior to conducting the resurgence evaluation.
Progressive Interval Assessment.
For the resurgence evaluation, we wanted to identity the leanest schedule of intermittent reinforcement that did not produce untoward side effects (e.g., a burst in the rate of destructive behavior). One way to accomplish this is to thin the schedule of reinforcement maintaining the alternative response until the point at which destructive behavior occurs and to then select a schedule of reinforcement just denser than the schedule associated with destructive behavior. Recent translational studies (e.g., Fisher, Greer, Fuhrman et al., 2018; Fisher, Greer, Mitteer et al., 2018) have used this general premise to select an appropriately lean schedule of reinforcement for destructive behavior by arranging a series of progressively thinning intervals of reinforcement (Findley, 1958). Our progressive interval assessment began with the therapist presenting the establishing operation for destructive behavior (i.e., withholding the iPad) and delivering 20-s access to the functional reinforcer for the FCR according to the following progression: fixed-interval [FI] 2 s, FI 5 s, FI 10 s, FI 20 s, FI 40 s, FI 60 s, FI 120 s, FI 180 s, and FI 240 s. The therapist conducted two trials at each FI schedule before progressing to the subsequent FI schedule, yielding a maximum of 18 possible trials per assessment. The progressive interval assessment ended following either (a) the first instance of destructive behavior or (b) completion of the second FI 240-s trial, whichever came first. If destructive behavior occurred, the therapist withheld the functional reinforcer and terminated the session. We selected the longest interval that did not produce destructive behavior as the initial variable interval (VI) schedule for destructive behavior in the subsequent concurrent VI VI assessment, the results of which then informed the schedules of reinforcement used in the resurgence evaluation for each participant. For example, if destructive behavior occurred during the first FI 40-s trial, but not during the preceding two FI 20-s trials, we programmed a VI 20-s schedule of reinforcement for destructive behavior in the concurrent VI VI assessment. Results of the progressive interval assessments suggested VI 20-s and VI 10-s reinforcement schedules for Charlie and Dan’s destructive behavior, respectively. We used the constant-probability algorithm described by Fleshler and Hoffman (1962) to construct all VI schedules of reinforcement. When a VI schedule was in effect for responding, the first response of each session produced the reinforcer, after which time, subsequent responses produced the reinforcer according to the programmed interval currently in effect.
Concurrent VI VI Assessment.
After identifying an initial VI schedule for each participant’s destructive behavior via the progressive interval assessment, we next evaluated how each participant allocated responding between concurrently available VI VI schedules arranged for destructive behavior and for the alternative response. Recall that we aimed to generate high rates of the alternative response and lower, but not zero, rates of destructive behavior when we programmed reinforcement for both responses. Thus, we began by programming a relatively dense schedule of reinforcement for each child’s alternative response in 10-min sessions and by then adjusting the relative density of the reinforcement schedules for destructive behavior and for the alternative response across sessions until we achieved the desired response pattern. After manipulating the concurrent VI VI schedules for Charlie and Dan, our desired response pattern was achieved under a VI 10-s schedule for destructive behavior and a VI 5-s schedule for the alternative response for both participants. When reinforcement was available, we used these reinforcement schedules throughout the remainder of the experiment, unless otherwise noted. The final VI 10-s schedule consisted of 20 intervals (range, 0.3 s–40.2 s), and the final VI 5-s schedule consisted of 24 intervals (range, 0.1 s–20.9 s). All reinforcer deliveries lasted 20 s.
Resurgence Evaluation
We used a three-phase progression common to resurgence evaluations. In Phase 1 (i.-e., baseline), the therapist delivered the functional reinforcer maintaining destructive behavior only for the destructive response. In Phase 2 (i.e., DRA), the therapist delivered the functional reinforcer for the alternative response; however, the therapist also delivered reinforcers for destructive behavior during DRA without extinction or noncontingently during DRA with extinction plus noncontingent reinforcement (NCR). In Phase 3 (i.e., extinction challenge), therapists no longer reinforced destructive behavior or the alternative response but delivered the functional reinforcer on a variable-time (VT) 200-s schedule, which we included to make the transition from DRA to the extinction challenge less discriminable (cf. Fuhrman et al., 2016; Koegel & Rincover, 1977). Table 1 provides an overview of the reinforcement contingencies in place across conditions and phases.
Table 1.
Reinforcement Contingencies Across Conditions and Phases in Experiment 1
| Condition | Phase 1 (Baseline) |
Phase 2 (DRA) |
Phase 3 (EXT Challenge) |
|---|---|---|---|
| DRA Without EXT | Target – VI 10 s | Target – VI 10 s | Target – EXT |
| Alt. – VI 5 s | Alt. – EXT | ||
| NCR – VT 200 s | |||
| DRA With EXT + NCR | Target – VI 10 s | Target – EXT | Target – EXT |
| Alt. – VI 5s | Alt. – EXT | ||
| NCR – Yoked to DRA without EXT | NCR – VT 200 s |
We repeated this three-phase progression four times with each participant, twice with DRA without extinction and twice with DRA with extinction plus NCR. The sequence of these progressions alternated according to an ABAB reversal design in which we explicitly did not counterbalance the order of each progression. That is, in order to equate the rates of reinforcement across conditions, we always conducted DRA without extinction first and then yoked the number of reinforcer deliveries during DRA with extinction plus NCR to the obtained rates of reinforcement from the preceding session of DRA without extinction. We did this by delivering NCR at those times when a reinforcer delivery occurred in the preceding session of DRA without extinction if that reinforcer had not already been produced by the alternative response in the current session of DRA with extinction plus NCR. We yoked reinforcer deliveries in this manner on a session-bysession basis. Additionally, we yoked the number of sessions of each DRA type across consecutive AB sequences of the ABAB reversal design as well as the number of sessions of the extinction challenge across all sequences (15 sessions of each extinction challenge for Charlie, 10 sessions of each extinction challenge for Dan). All reinforcer deliveries lasted 20 s, and all sessions lasted 10 min. This portion of the study occurred across 14 appointments with an average of seven sessions per appointment (range, 3–11 sessions per appointment) for Charlie and across 17 appointments with an average of five sessions per appointment (range, 1–7 sessions per appointment) for Dan.
We attempted to decrease the likelihood of carryover between the two progressions by programming discriminative stimuli unique to each progression. Baseline, DRA, and the extinction challenge for each progression occurred in a unique stimulus context. Each context was correlated with a both a unique therapist and color (e.g., Therapist A who wore a yellow scrub top and conducted all sessions for one progression in front of a yellow poster board and under yellow light covers and Therapist B who wore a blue scrub top and conducted all sessions for the other progression in front of a blue poster board and under blue light covers). Therapists removed these discriminative stimuli when sessions ended if the participant remained in the therapy room (e.g., over lunch), as was typically the case outside of session time.
Phase 1: Baseline.
Therapists reinforced destructive behavior on a VI 10-s schedule while the alternative response remained unavailable. We conducted at least five baseline sessions and continued baseline until we obtained five consecutive sessions for which (a) the standard deviation of the rates of destructive behavior did not exceed 50% of its mean and (b) the trend in the rates of destructive behavior was either flat or increasing. Additionally, we never transitioned from baseline to either DRA condition at the start of an appointment (i.e., at least one baseline session immediately preceded the start of DRA).
Phase 2: DRA Without Extinction.
Therapists reinforced destructive behavior on the same VI 10-s schedule used in baseline but also reinforced the alternative response on a VI 5-s schedule. Reinforcing either response paused the VI timer for the other response such that reinforcement was not guaranteed to be available for the other response immediately upon terminating reinforcement for the first response. Therapists conducted at least five sessions in this condition and continued until rates of destructive behavior met the first portion of the stability criteria (related to response variability) used in baseline for at least two consecutive sessions. Additionally, we never transitioned from DRA without extinction to the extinction challenge at the start of an appointment (i.e., at least one session of DRA without extinction immediately preceded the start of the extinction challenge).
Phase 2: DRA With Extinction Plus NCR.
Therapists no longer reinforced destructive behavior but reinforced the alternative response on the same VI 5-s schedule used in DRA without extinction. To determine the NCR schedule for each session, data collectors reviewed the corresponding session from the immediately preceding phase of DRA without extinction. We yoked the first session of DRA with extinction plus NCR to the first session of DRA without extinction, and we yoked each subsequent session in the same manner. By yoking the number of reinforcers delivered in this condition to those delivered in the immediately preceding phase of DRA without extinction, we closely equated the obtained rates of reinforcement across the DRA conditions. To do so, therapists reviewed the timestamps at which they delivered reinforcers in the corresponding DRA-without-extinction session prior to the start of the upcoming session of DRA with extinction plus NCR. Observers recorded these times, and as the yoked session progressed, they instructed the therapist via earbud to deliver the time-based reinforcer at the programmed times. Therapists withheld scheduled, time-based reinforcers during DRA-with-extinction-plus-NCR sessions if the number of reinforcer deliveries at that point in the session equaled or exceeded the number delivered at the same point in time in the corresponding DRA-without-extinction session. We subtracted future time-based reinforcer deliveries from upcoming sessions if needed to ensure that obtained reinforcement rates across both DRA conditions were equal.
In addition to the 3-s COD in place to minimize adventitious reinforcement of destructive behavior following reinforcer deliveries for the alternative response, we implemented another 3-s COD for all time-based reinforcer deliveries. That is, therapists withheld time-based reinforcers for 3 s following any instance of destructive behavior or alternative response. We used the same termination criteria for ending DRA with extinction plus NCR as those used for ending DRA without extinction. Additionally, we never transitioned from DRA with extinction plus NCR to the extinction challenge at the start of an appointment (i.e., at least one session of DRA with extinction plus NCR immediately preceded the start of the extinction challenge). Furthermore, we conducted additional sessions of DRA with extinction plus NCR whenever the preceding phase of DRA without extinction required more than five sessions to meet progression criteria.
Phase 3: Extinction Challenge.
Therapists no longer reinforced destructive behavior or the alternative response but delivered the functional reinforcer on a VT 200-s schedule. We continued to implement the 3-s COD for all time-based reinforcer deliveries. Each extinction challenge ended after 15 sessions for Charlie and after 10 sessions for Dan.
Results and Discussion
Figure 1 depicts results of the resurgence evaluation for Charlie with rates of destructive behavior in the top panel, rates of alternative responding in the middle panel, and the frequency of reinforcer deliveries in the bottom panel. Charlie displayed high rates of destructive behavior across the four baseline phases, and Charlie’s therapist delivered reinforcers at similar rates across these phases. Both DRA conditions reduced Charlie’s baseline rates of destructive behavior, with greater reductions during DRA with extinction plus NCR relative to DRA without extinction. Charlie displayed high rates of the alternative response in both DRA conditions, and his therapist delivered reinforcers at similar rates across DRA phases and conditions. Finally, we observed lower rates of destructive behavior during extinction-challenge phases that followed DRA with extinction plus NCR relative to extinction-challenge phases that followed DRA without extinction, despite identical reinforcement rates produced by the VT 200-s schedule.
Fig. 1. Charlie’s resurgence-evaluation results across sequences of phases in which differential reinforcement of alternative behavior (DRA) was implemented either without extinction (DRA without EXT) or with extinction plus noncontingent reinforcement (DRA with EXT + NCR).

Note. The three panels depict rates of destructive behavior (top panel), rates of the alternative response (middle panel), and frequencies of noncontingent (NCR) or contingent (SR) reinforcer deliveries, depicted as stacked open and closed bars, respectively (bottom panel).
Figure 2 depicts results of the resurgence evaluation for Dan. Dan displayed high and increasing rates of destructive behavior across the four baseline phases. Despite these differences in response rates, we observed similar reinforcement rates across baseline phases. Similar to Charlie, both DRA conditions reduced Dan’s rates of destructive behavior relative to baseline, with slightly greater overall reductions during DRA with extinction plus NCR relative to DRA without extinction. Dan displayed high and often increasing rates of the alternative response in both DRA conditions, and his therapist delivered similar rates of reinforcement across DRA phases and conditions. Unlike Charlie, rates of destructive behavior for Dan across the extinction-challenge phases did not differ markedly, despite similar reinforcement rates produced by the VT 200-s schedule. We present data on proportion of baseline responding for both Charlie and Dan in the context of the findings of Experiment 2 in the sections below.
Fig. 2. Dan’s resurgence-evaluation results across sequences of phases in which differential reinforcement of alternative behavior (DRA) was implemented either without extinction (DRA without EXT) or with extinction plus noncontingent reinforcement (DRA with EXT + NCR).

Note. The three panels depict rates of destructive behavior (top panel), rates of the alternative response (middle panel), and frequencies of noncontingent (NCR) or contingent (SR) reinforcer deliveries, depicted as stacked open and closed bars, respectively (bottom panel).
A few limitations of Experiment 1 led to the design of a better controlled experiment, which we conducted in a nonclinical (i.e., laboratory) setting using an experimental design that minimized potential order effects. First, highly variable rates of destructive behavior for both Charlie and Dan clouded our ability to evaluate the question at hand in a conclusive manner. Second, this effect was magnified somewhat for Dan because rates of destructive behavior increased across successive baseline phases, making it difficult to relate performance in prior phases to that in latter phases when testing under the same experimental conditions. Third, the high rates of destructive behavior we sometimes observed with Charlie and Dan, paired with the extended nature of the successive design of Experiment 1, suggested to us that further study of the question at hand would be better suited for evaluation in a laboratory setting and with nonclinical subjects and response alternatives. Fourth, we delivered time-based reinforcers during the extinction challenge, which is common when testing for reinstatement (e.g., Podlesnik & Shahan, 2009). Because relapse can be greater when combining tests for resurgence and reinstatement than when testing either in isolation (Liggett et al., 2018), our delivery of time-based reinforcers during the extinction challenge may have confounded resurgence and reinstatement models of relapse.
Experiment 2
Method
Subjects
Ten male Long-Evans rats (Charles River Laboratories, Stone Ridge, NY; IDs R1-R10) served as subjects. Rats were approximately 12 months old at the beginning of the experiment and had no previous experience with resurgence preparations. They did, however, have a previous history with reinforcement and extinction across various contextual manipulations (see Craig, Sullivan, Browning et al., 2019; Craig, Sullivan, & Roane, 2019). Rats were pair-housed in a temperature- and humidity-controlled colony room with a 12:12 hr light/dark cycle. All sessions occurred daily at approximately the same time during the light cycle. Rats were maintained at 80% of their free-feeding weights and received postsession feedings in individual feeding cages (approximately 2 hr), after which they were returned to their home cages. Rats had free access to water in both their home and feeding cages. Animal care and housing were in accordance with the guidelines set forth by SUNY Upstate Medical University’s Institutional Animal Care and Use Committee.
Apparatus
Four identical Med-Associates (St. Albans, VT) operant chambers, housed in sound- and light-attenuating cubicles, were used. A fan affixed to each cubicle masked extraneous sounds at all times. Each chamber measured 30 cm × 24 cm × 21 cm. The front and back walls were constructed of brushed aluminum work panels. The ceiling, door, and wall opposite the door were constructed of clear Plexiglas. The chamber floor consisted of cylindrical stainless-steel bars (0.47 cm diameter) spaced 1.57 cm from one another and arranged parallel to the front and back walls. A house light was centered at the top of the back wall and provided general illumination. On the front wall, two retractable levers with stimulus lights above them were positioned left and right of a centered feeder aperture located in the wall. On the center of the back wall was a single nose-poke aperture that was internally illuminated by a yellow LED. Reinforcement entailed delivery of a single 45 mg grain-based food pellet (Bio Serv, Flemington, NJ) into the feeder aperture. During reinforcement, all lights, excluding the house light, were turned off and there was a 3-s timeout from sessions. Experimental events and data recording were controlled by Med-PC software running on a computer in the session room.
Statistical Analyses
All statistical tests reported below were deemed significant at α level of .05. We used Greenhouse-Geisser corrections to degrees of freedom when the assumption of sphericity in analyses of variance (ANOVA; tested using Mauchly’s method) was violated.
Procedure
Magazine Training
We first ensured rats would retrieve food pellets from the feeder aperture in a single session. During this session, the chamber was illuminated by the house light, levers were retracted, and food was delivered according to a VT 60-s schedule. All schedules of reinforcement used in the current experiment consisted of 10 interreinforcer intervals derived from the constant-probability algorithm provided by Fleshler and Hoffman (1962). Magazine training, and sessions for the remainder of the experiment, lasted 30 min, excluding time for reinforcer deliveries.
Resurgence Evaluation
We used the same three-phase progression resurgence evaluation described in Experiment 1, with the exception that during Phase 2, we used a multielement design. With this modification, we were able to yoke reinforcement rates across DRA contexts while addressing the previous limitation in Experiment 1 of a potential sequence effect. Table 2 provides an overview of the reinforcement contingencies in place across conditions and phases.
Table 2.
Reinforcement Contingencies Across Conditions and Phases in Experiment 2
| Condition | Phase 1 (Baseline) |
Phase 2 (DRA) |
Phase 3 (EXT Challenge) |
|---|---|---|---|
| DRA Without EXT | Target – VI 60 s | Target – VI 60 s | Target – EXT |
| Alt. – VI 15 s | Alt. – EXT | ||
| DRA With EXT + NCR | Target – VI 60 s | Target – EXT | Target – EXT |
| Alt. – VI 15 s | Alt. – EXT | ||
| NCR – Yoked to DRA without EXT |
Phase 1: Baseline.
During sessions of Phase 1, both response levers were inserted into the chamber, and the stimulus light positioned above one of the levers (either left or right, counterbalanced across rats) was illuminated. In the first session of Phase 1, the initial target-lever press produced food to facilitate acquisition, after which pressing the target lever produced food according to a VI 60-s schedule. Presses to the other inactive lever, or nose pokes into the nose-poke aperture, produced no consequences but were recorded. Phase 1 lasted for 25 sessions.
Phase 2: DRA With and Without Extinction.
Before beginning Phase 2, rats were assigned to either the DRA with extinction plus NCR group (n = 5) or to the DRA without extinction group (n = 5). Given that past research has suggested baseline response rates predict the magnitude of relapse (e.g., Craig, Sullivan, & Roane, 2019; Podlesnik et al., 2017; Sweeney & Shahan, 2013), rats were matched such that the mean rates of target-lever pressing during the final three sessions of Phase 1 did not differ between groups. Sessions of Phase 2 began similarly to Phase 1 in that both levers were inserted into the chamber with the stimulus light positioned above the target lever illuminated. In addition, the nose-poke aperture on the back chamber wall was illuminated. For both groups of rats, during all sessions of Phase 2, the first nose poke into the illuminated nose-poke aperture produced food to initially facilitate acquisition and thereafter to ensure the rats continued to contact alternative reinforcement. After the initial reinforcer delivery, alternative nose pokes produced food reinforcement according to a VI 15-s schedule for the rest of the experimental session.
Contingencies for target-lever pressing differed between groups. For rats in the DRA without extinction group, the first target-lever press produced reinforcement throughout Phase 2, after which a VI 60-s schedule commenced, as in Phase 1. Additionally, a COD was in place for both target-lever presses and alternative nose pokes such that reinforcement was unavailable for either response if they occurred within 3 s of one another. In the DRA with extinction plus NCR group, target-lever pressing was placed on extinction. However, to maintain similar rates of reinforcement across groups, food reinforcement was also delivered according to a VT schedule. During the first session of Phase 2, the VT schedule was yoked to the mean rate of reinforcement from the final session of Phase 1 For the remaining sessions in Phase 2, the VT schedule was yoked to the mean rate of reinforcement produced by target-lever pressing from the DRA without extinction group’s previous session. Furthermore, target-lever presses initiated a COD such that a reinforcer could not be delivered within 3 s of the target-lever press. Presses to the inactive lever produced no consequences for either group but were recorded. Phase 2 lasted for 14 sessions.
Phase 3: Extinction Challenge.
In Phase 3, all stimuli were identical to those of Phase 2. That is, levers were inserted into the chamber, the stimulus light about the target lever was illuminated, the nose-poke aperture was illuminated, and the house light was on. All sources of reinforcement, however, were suspended. Phase 3 lasted for five sessions.
Results and Discussion
The top panels of Figure 3 show target-lever responses per minute across sessions of Experiment 2. Data from individual rats in the DRA with extinction plus NCR and without extinction groups are shown in the top-left and top-center panels of the figure, respectively. Mean target-lever response rates are shown in the top-right panel of the figure. Session 0 in each panel represents mean target-lever response rates from the last three sessions of Phase 1. Rates of target-lever pressing were comparable between groups during these final sessions of the phase, t(8) = 0.01, p = .994, d = 0.01. The inactive-lever press occurred at near-zero rates in both groups (for the DRA with extinction plus NCR and without extinction groups, respectively: M = 1.29, SD = 1.89; and M = 1.03, SD = 0.54 responses per minute) during the final three sessions of Phase 1.
Fig. 3. Rates of target (top panels) and alternative responses (bottom panels) across phases in which differential reinforcement of alternative behavior (DRA) was implemented either with extinction plus noncontingent reinforcement (DRA with EXT + NCR) or without extinction (DRA without EXT).

Note. Aggregated responding for each group is depicted in the right column.
Reinforcer rates for target behavior during the last three sessions of Phase 1 (Session 0) and all sessions of Phase 2 are shown in Figure 4. From the left to the right, the panels depict target-reinforcer rates for individual subjects in the DRA with extinction plus NCR group, individual subjects in the DRA without extinction group, and these rates aggregated across subjects in both groups, respectively. Note that the Phase-2 reinforcer rates shown for rats in the DRA with extinction plus NCR group represent rates of time-based VT reinforcement that were yoked to target-reinforcer rates from the DRA without extinction group. During the final three sessions of Phase 1, target-reinforcer rates did not differ between groups, t(8) = 0.61, p = .561, d = 0.43, and approximated programmed reinforcer rates (i.e., one reinforcer per minute).
Fig. 4. Rates of noncontingent and contingent reinforcer deliveries for target responses (top panels) and alternative responses (bottom panels) across phases in which differential reinforcement of alternative behavior (DRA) was implemented either with extinction plus noncontingent reinforcement (DRA with EXT + NCR) or without extinction (DRA without EXT).

Note. Aggregated rates of reinforcer deliveries for each group are depicted in the right column.
During Phase 2, rates of target-lever pressing decreased across sessions for rats in the DRA with extinction plus NCR group and were less than five responses per minute during Session 5 and all subsequent sessions of the phase. Target-lever response rates for all rats in the DRA without extinction group decreased relative to baseline during the first session of Phase 2, but the relation between baseline response rates and Phase-2 response rates varied between rats and across sessions. For Rat R4, responding occurred at roughly baseline rates throughout the phase. For R8, target-lever response rates decreased initially, after which they varied nonsystematically across sessions of Phase 2. For the remaining rats, target-lever pressing decreased early in Phase 2, but responding gradually recovered across sessions until, by the end of the phase, it occurred at roughly baseline rates. A 2 × 14 (Group × Session) mixed-model ANOVA was used to compare target-lever responding between groups across sessions, and this test returned significant main effects of Group, F (1,8) = 13.90, p = .002, η2p = .635; Session, F (13, 104) = 2.75, p = .002, η2p = .256; and a significant Group × Session interaction, F (13, 104) =4.93, p < .001, η2p = .381. Thus, target-lever responding decreased across sessions of Phase 2 for rats in the DRA with extinction plus NCR group and, on the whole, remained elevated and reasonably stable across sessions for rats in the DRA without extinction group. These findings are generally consistent with those reported in Experiment 1—providing alternative reinforcement with extinction for target behavior more effectively reduced target responding than did providing alternative reinforcement without extinction.
Rates of the alternative nose-poke response across sessions of Phases 2 and 3 are shown in the bottom panels of Figure 3. Data from individual rats in the DRA with and without extinction groups are shown in the bottom-left and bottom-center panels of the figure, respectively. Mean alternative nose-poke response rates are shown in the bottom-right panel of the figure. The alternative nose-poke response occurred at near-zero rates in both groups (for the DRA with extinction plus NCR and without extinction groups, respectively: M = 0.81, SD = 0.69; and M = 0.42, SD = 0.27 responses per minute) during the final three sessions of Phase 1. Alternative nose poking increased across sessions of Phase 2, with some variability, for all rats. A 2 × 14 (Group × Session) mixed-model ANOVA conducted on these data revealed a main effect of Session, F (13, 104) = 17.93, p < .001, η2p = .691, but the main effect of Group and Group × Session interaction were not significant—respectively, F(1, 8) = 0.11, p = .744, η2p = .014; and F (13, 104) = 0.41, p = .963, η2p = .049. Thus, alternative responding increased across sessions of the phase, and the rate at which it increased was the same between groups.
Inactive-lever pressing occurred at near-zero rates (for the DRA with extinction plus NCR and without extinction groups, respectively: M = 0.67, SD = 0.80; and M = 0.40, SD = 0.29 responses per minute) during the final session of Phase 2.
Across sessions of Phase 2, rates of reinforcement for target-lever pressing for rats in the DRA without extinction group and rates of yoked VT reinforcement for rats in the DRA with extinction plus NCR group were reasonably stable and similar between groups (see the top-center and top-left panels of Fig. 4, respectively). A 2 × 14 (Group × Session) mixed-model ANOVA conducted on these data revealed a significant Group × Session interaction, F(13, 104) = 2.12, p = .018, η2p = .210, though neither of the main effects were significant—for Group and Session, respectively: F(1, 8) = 4.73, p = .061, η2p = .372; and F(13, 104) = 0.97, p = .487, η2p = .108. Though visual inspection of data in the top-right panel of Figure 4 does not immediately indicate the source of the significant Group × Session interaction noted above, the ANOVA may have detected the slight decreasing trend in VT reinforcer rates across sessions in the DRA with extinction plus NCR group compared to the reasonably consistent rates of reinforcement for target-lever pressing in the DRA without extinction group. Reinforcer rates for the alternative nose-poke response tended to increase across sessions for both groups (see the bottom panels of Fig. 4), and mean alternative-reinforcer rates tended to be slightly higher in the DRA with extinction plus NCR group than in the DRA without extinction group. A 2 × 14 (Group × Session) mixed-model ANOVA conducted on these data revealed only a significant main effect of Session, F(13, 104) = 22.69, p < .001, η2p = .739 (for the main effect of Group and Group × Session interaction, respectively: F(1, 8) = 1.29, p = .288, η2p = .139; and F(13, 104) = 0.26, p = .996, η2p = .031). Thus, though alternative reinforcer rates were, on average, consistently higher across sessions for the DRA with extinction plus NCR group than for the DRA without extinction group, these differences were small and not statistically reliable.
Relative to the final session of Phase 2, rates of target-lever pressing increased when reinforcement was suspended during the first session of Phase 3 for every animal (see the top panels of Fig. 3). A 2 × 2 (Group × Phase) mixed-model ANOVA conducted on these data revealed a significant main effect of Phase, F(1, 8) = 28.66, p = .001, η2p = .782, and Group, F(1, 8) = 13.68, p = .006, η2p = .631. The Group × Phase interaction, however, was not significant, F(1, 8) = 0.02, p = .905, η2p = .002. Thus, even though target-lever response rates tended to be significantly higher for rats in the DRA without extinction group than for rats in the DRA with extinction plus NCR group during these sessions, the absolute increase in response rates between phases was the same between groups.
Across sessions of Phase 3, target-lever response rates tended to be initially higher for rats in the DRA without extinction group than for rats in the DRA with extinction plus NCR group (see the top panels of Fig. 3). However, rates of target-lever pressing decreased across sessions for rats in both groups such that responding occurred at comparable, low rates in both groups during the final session of the phase. A 2 × 5 (Group × Session) mixed-model ANOVA conducted on these data revealed significant main effects of Group, F(1, 8) = 11.85, p = .009, η2p = .597, and Session, F(1.33, 10.65) = 10.72, p = .005, η2p = .573, and a significant Group × Session interaction, F(1.33, 10.65) = 7.49, p = .015, η2p = .484. These statistical outcomes are consistent with the visual analysis provided above.
Alternative nose poking occurred at similar rates in both groups during Phase 3, and nose poking decreased across sessions similarly for both groups (see the bottom panels of Fig. 3). These findings were supported by a 2 × 5 (Group × Session) mixed-model ANOVA conducted on nose-poke response rates during Phase 3. This test returned only a significant main effect of Session, F(1.82, 14.57) = 51.08, p < .001, η2p = .865—for the main effect of Group and Group × Session interaction, respectively: F(1, 8) = 0.52, p = .493, η2p = .061; and F(1.82, 14.57) = 0.45, p = .626, η2p = .054.
The outcomes from the resurgence evaluation reported above are generally consistent with the resurgence data reported in Experiment 1. That is, during both Phase 2 and the Phase 3 extinction challenge, target behavior for rats in the DRA without extinction group occurred at higher rates than for rats in the DRA with extinction plus NCR group. It is important to note, however, that the dependency between treatment type and the absolute magnitude of the increase in target-response rates between the end of Phase 2 and the beginning of Phase 3 differed between experiments. In Experiment 1, response rates tended to increase more following DRA without extinction than following the DRA with extinction plus NCR, whereas no such between-treatment differences were apparent in Experiment 2.
Any of various between-experiment differences (e.g., the species whose behavior was studied, the reinforcement schedules used) could have been responsible for this discrepancy. One factor that likely contributed to this difference, however, was the extent to which target behavior was reduced by the end of the DRA without extinction treatment. On average, target behavior occurred at nearer baseline rates at the end of Phase 2 in Experiment 2 than in Experiment 1. Thus, a ceiling effect may have masked between-group differences in resurgence in Experiment 2. That is, because target behavior was near baseline rates for most rats in the DRA without extinction group, it could not increase much beyond its terminal Phase 2 rate.
Nevertheless, these findings reinforce those from Experiment 1 while addressing some of the procedural limitations from that experiment. In particular, order effects were avoided in the present experiment by using a between-groups design. Also, because time-based reinforcers were not delivered in Phase 3 of the present experiment, response-rate increases may be more confidently attributed to resurgence rather than reinstatement processes.
General Discussion
We compared the efficacy of two DRA-based treatments for target responding when the overall rate of reinforcement did not differ across the DRA conditions. In one DRA condition, target responding continued to produce reinforcement according to the same schedule from baseline. In the other DRA condition, target responding no longer produced reinforcement, and time-based reinforcers replaced those reinforcers available for target responding in the first DRA condition. Across Experiments 1 and 2, target responding was lower when DRA included extinction plus NCR than when DRA did not include extinction. This same pattern of responding persisted when all reinforcer deliveries ceased (Experiment 2) or were available only on a time-based schedule (Experiment 1).
These findings were furthered clarified when we analyzed responding in Phases 2 and 3 as a proportion of baseline responding, which we calculated by dividing target-response rates in each session of Phases 2 and 3 by the mean rate of target responding from the final five (Experiment 1) or all (Experiment 2) sessions of the preceding baseline phase. The left column of panels in Figure 5 display destructive behavior for Charlie and Dan, expressed as a proportion of baseline responding for the first and second AB comparisons of the ABAB design during DRA with extinction plus NCR (top-left panel) and DRA without extinction (middle-left panel). The bottom-left panel depicts these same data but averaged across AB comparisons and participants for each DRA condition. The right column of panels in Figure 5 display these same data but for Experiment 2. Target-response rates tended to be lower within subject (Experiment 1) and across subjects (Experiments 1 and 2) during and following the DRA condition that included extinction plus NCR than the DRA condition that continued to reinforce target responding. As noted above, Experiment 1 showed greater increases in target responding when transitioning from Phase 2 to Phase 3 with DRA without extinction than DRA with extinction plus NCR, but this finding was not replicated in Experiment 2. It remains possible that better suppression of target responding during DRA without extinction in Experiment 2 would have led to the replication of this finding.
Fig. 5. Proportion of baseline target responding during the last two sessions of Phase 2 and the first five sessions of Phase 3 for Experiment 1 (left panels) and Experiment 2 (right panels) across differential reinforcement of alternative behavior (DRA) implemented with extinction plus noncontingent reinforcement (NCR; top panels) and without extinction (middle panels).
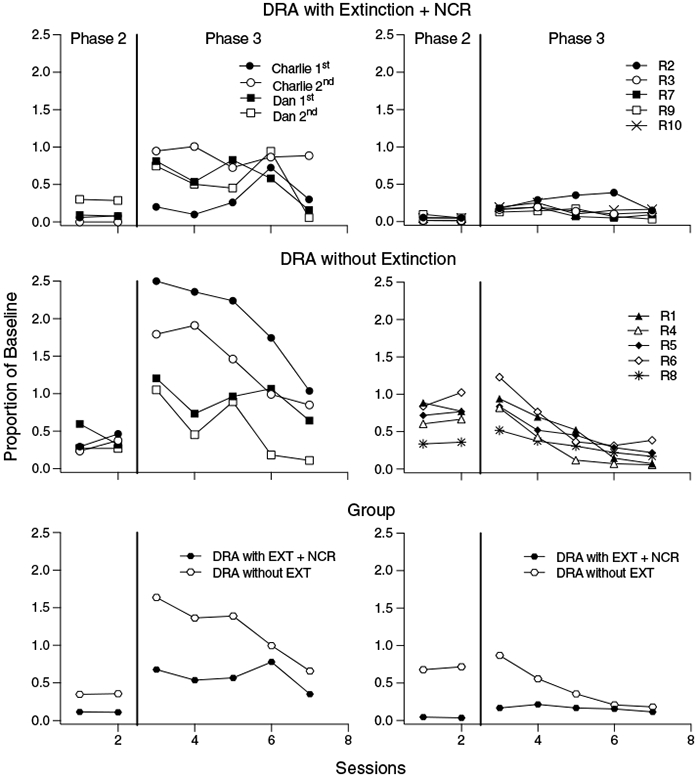
Note. Bottom panels show aggregated target responding.
These collective findings add to the growing literature on variables that promote and mitigate treatment relapse when DRA interventions are temporarily suspended. Prior investigations have shown that DRA interventions often result in resurgence when alternative reinforcement ceases, both when the intervention includes (e.g., Volkert et al., 2009; Wacker et al., 2013) and does not include (e.g., Mace et al., 2010) extinction. The present results extend this literature by suggesting that when target responding is sufficiently suppressed in Phase 2, resurgence may be greater following DRA without extinction relative to resurgence following DRA with extinction plus NCR. If with additional study this finding is confirmed, such a result would conform to the predictions of both BMT and RaC. According to BMT, this effect may happen as a result of severing the response–reinforcer relation maintaining target behavior and altering generalization decrement (Nevin et al., 2001; Greer, Fisher, Romani et al., 2016). According to RaC, DRA treatments that arrange extinction for target behavior lessen the relative value of that response in relation to the value of the alternative response, which is likely to affect resurgence (Greer & Shahan, 2019).
Regardless of whether resurgence is greater following either DRA condition, our finding across both experiments that target-response rates were lower during and following one of the two DRA conditions, has important implications for practice. Whenever feasible, clinicians should arrange extinction for destructive behavior. When it is unsafe or otherwise impractical to do so, quantitative theories of resurgence offer a number of suggestions for mitigating resurgence other than programming extinction for destructive behavior, such as signaling the unavailability of reinforcement for the alternative response (e.g., Fisher et al., 2020; Fuhrman et al., 2016; Nevin et al., 2016) and conducting treatment in a novel context (e.g., Craig et al., 2018). Additionally, RaC suggests a number of procedural modifications for mitigating resurgence that could be adapted to DRA treatments that do not include extinction. For example, RaC suggests that any increment in the relative value of the alternative response over that of destructive behavior will mitigate resurgence. Such value-altering procedures may include decreasing the relative effort of the alternative response (e.g., Horner & Day, 1991) and increasing the relative rate, immediacy, quality, and/or magnitude of the reinforcer for the alternative response (e.g., Athens & Vollmer, 2010). Similar, but opposite, modifications could be made for destructive behavior. In fact, many of the procedural modifications that applied behavior analysts already implement when treating destructive behavior with DRA without extinction are predicted by RaC to mitigate resurgence.
Translational researchers should pay special attention to those procedural modifications that show promise for mitigating resurgence following DRA-based interventions that do not include extinction, as these same modifications should only be enhanced with similar interventions that include extinction. Furthermore, predictions shared by two or more quantitative theories of relapse (e.g., the prediction shared by BMT and RaC regarding differential levels of resurgence following DRA interventions that do and do not include extinction) should be of special interest to translational researchers, for these are likely to be the most robust and fruitful lines of inquiry for improving practice.
Acknowledgments
Experiment 1 fulfilled partial requirements of the first author’s Master of Arts degree from the University of Nebraska Omaha and was supported in part by grants 5R01HD079113 and 5R01HD083214 from The National Institute of Child Health and Human Development. Experiment 2 was conducted at Upstate Medical University. Katherine R. Brown is now at Utah State University. Brian D. Greer and Wayne W. Fisher are now at Children’s Specialized Hospital–Rutgers University Center for Autism Research, Education, and Services (CSH–RUCARES) and Rutgers Robert Wood Johnson Medical School.
Footnotes
All participant names are pseudonyms.
References
- Athens ES, & Vollmer TR (2010). An investigation of differential reinforcement of alternative behavior without extinction. Journal of Applied Behavior Analysis, 43, 569–589. 10.1016/j.ridd.2010.12.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berg WK, Ringdahl JE, Ryan SE, Ing AD, Lustig N, Romani P, Wacker DP, Andersen JK, & Durako E (2015). Resurgence of mands following functional communication training. Mexican Journal of Behavior Analysis, 41, 166–186. [PMC free article] [PubMed] [Google Scholar]
- Betz AM, & Fisher WW (2011). Functional analysis: History and methods. In Fisher WW & Roane HS (Eds.), Handbook of applied behavior analysis (pp. 206–255). Guilford. [Google Scholar]
- Briggs AM, Fisher WW, Greer BD, & Kimball RT (2018). Prevalence of resurgence of destructive behavior during reinforcement schedule thinning during functional communication training. Journal of Applied Behavior Analysis, 51, 620–633. 10.1002/jaba.472 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conners J, Iwata BA, Kahng S, Hanley GP, Worsdell AS, & Thompson RH (2000). Differential responding in the presence and absence of discriminative stimuli during multielement functional analyses. Journal of Applied Behavior Analysis, 33, 299–308. 10.1901/jaba.2000.33-299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craig AR, Cunningham PJ, Sweeney MM, Shahan TA, & Nevin JA (2018). Delivering alternative reinforcement in a distinct context reduces its counter-therapeutic effects on relapse. Journal of the Experimental Analysis of Behavior, 109, 492–505. 10.1002/jeab.431 [DOI] [PubMed] [Google Scholar]
- Craig AR, Sullivan WE, Browning KO, DeRosa NM, & Roane HS (2019). Re-exposure to reinforcement in Context A during treatment in Context B reduces ABC renewal. Journal of the Experimental Analysis of Behavior, 113, 141–152. 10.1002/jeab.569 [DOI] [PubMed] [Google Scholar]
- Craig AR, Sullivan WE, & Roane HS (2019). Further evaluation of a nonsequential approach to studying operant renewal. Journal of the Experimental Analysis of Behavior, 112, 210–223. 10.1002/jeab.546 [DOI] [PubMed] [Google Scholar]
- Epstein R (1985). Extinction-induced resurgence: Preliminary investigations and possible applications. The Psychological Record, 35, 143–153. 10.1007/bf03394918 [DOI] [Google Scholar]
- Findley JD (1958). Preference and switching under concurrent scheduling. Journal of the Experimental Analysis of Behavior, 1, 123–144. 10.1901/jeab.1958.1-123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Fuhrman AM, Greer BD, Mitteer DR, & Piazza CC (2020). Mitigating resurgence of destructive behavior using the discriminative stimuli of a multiple schedule. Journal of the Experimental Analysis of Behavior, 113, 263–277. 10.1002/jeab.552 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Greer BD, Craig AR, Retzlaff BJ, Fuhrman AM, Lichtblau KR, & Saini V (2018a). On the predictive validity of behavioral momentum theory for mitigating resurgence of problem behavior. Journal of the Experimental Analysis of Behavior, 109, 281–290. 10.1002/jeab.303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Greer BD, Fuhrman AM, Saini V, & Simmons CA (2018b). Minimizing resurgence of destructive behavior using behavioral momentum theory. Journal of Applied Behavior Analysis, 51, 831–853. 10.1002/jaba.499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Greer BD, Mitteer DR, Fuhrman AM, Romani PW, & Zangrillo AN (2018c). Further evaluation of differential exposure to establishing operations during functional communication training. Journal of Applied Behavior Analysis, 51, 360–373. 10.1001/jaba.451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Greer BD, Romani PW, Zangrillo AN, & Owen TM (2016). Comparisons of synthesized and individual reinforcement contingencies during functional analysis. Journal of Applied Behavior Analysis, 49, 596–616. 10.1002/jaba.314 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Ninness HAC, Piazza CC, & Owen-DeSchryver JS (1996a). On the reinforcing effects of the content of verbal attention. Journal of Applied Behavior Analysis, 29, 235–238. 10.1901/jaba.1996.29-235 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Piazza CC, Bowman LG, & Amari A (1996b). Integrating caregiver report with systematic choice assessment to enhance reinforcer identification. American Journal of Mental Retardation, 101, 15–25. [PubMed] [Google Scholar]
- Fisher W, Piazza CC, Bowman LG, Hagopian LP, Owens JC, & Slevin I (1992). A comparison of two approaches for identifying reinforcers for persons with severe and profound disabilities. Journal of Applied Behavior Analysis, 25, 491–498. 10.1901/jaba.1992.25-491 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Piazza CC, & Chiang CL (1996c). Effects of equal and unequal reinforcer duration during functional analysis. Journal of Applied Behavior Analysis, 29, 117–120. 10.1901/jaba.1996.29-117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Saini V, Greer BD, Sullivan WE, Roane HS, Fuhrman AM, Craig AR, & Kimball RT (2018d). Baseline reinforcement rate and resurgence of destructive behavior. Journal of the Experimental Analysis of Behavior, 111, 75–93. 10.1002/jeab.488 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Thompson RH, Hagopian LP, Bowman LG, & Krug A (2000). Facilitating tolerance of delayed reinforcement during functional communication training. Behavior Modification, 24, 3–29. 10.1177/0145445500241001 [DOI] [PubMed] [Google Scholar]
- Fleshler M, & Hoffman HS (1962). A progression for generating variable-interval schedules. Journal of the Experimental Analysis of Behavior, 5, 529–530. 10.1901/jeab.1962.5-529 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuhrman AM, Fisher WW, & Greer BD (2016). A preliminary investigation on improving functional communication training by mitigating resurgence of destructive behavior. Journal of Applied Behavior Analysis, 49, 884–899. 10.1002/jaba.338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer BD, Fisher WW, Retzlaff BJ, & Fuhrman AM (2020). A preliminary evaluation of treatment duration on the resurgence of destructive behavior. Journal of the Experimental Analysis of Behavior, 113, 251–262. 10.1002/jeab.567 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer BD, Fisher WW, Romani PW, & Saini V (2016a). Behavioral momentum theory: A tutorial on response persistence. The Behavior Analyst, 39, 269–291. 10.1007/s40614-016-0050-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer BD, Fisher WW, Saini V, Owen TM, & Jones JK (2016b). Functional communication training during reinforcement schedule thinning: An analysis of 25 applications. Journal of Applied Behavior Analysis, 49, 105–121. 10.1002/jaba.265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer BD, & Shahan TA (2019). Resurgence as Choice: Implications for promoting durable behavior change. Journal of Applied Behavior Analysis, 52, 816–846. 10.1002/jaba.573 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagopian LP, Fisher WW, Sullivan MT, Acquisto J, & LeBlanc LA (1998). Effectiveness of functional communication training with and without extinction and punishment: A summary of 21 inpatient cases. Journal of Applied Behavior Analysis, 31, 211–235. 10.1901/jaba.1998.31-211 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond JL, Iwata BA, Rooker GW, Fritz JN, & Bloom SE (2013). Effects of fixed versus random condition sequencing during multielement functional analyses. Journal of Applied Behavior Analysis, 46, 22–30. 10.1002/jaba.7 [DOI] [PubMed] [Google Scholar]
- Herrnstein RJ (1961). Relative and absolute strength of response as a function of frequency of reinforcement. Journal of the Experimental Analysis of Behavior, 4, 267–272. 10.1901/jeab.1961.4-267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman K, & Falcomata TS (2014). An evaluation of resurgence of appropriate communication in individuals with autism who exhibit severe problem behavior. Journal of Applied Behavior Analysis, 47, 651–656. 10.1002/jaba.144 [DOI] [PubMed] [Google Scholar]
- Horner RH, & Day HM (1991). The effects of response efficiency on functionally equivalent competing behaviors. Journal of Applied Behavior Analysis, 24, 719–732. 10.1901/jaba.1991.24-719 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iwata BA, Dorsey MF, Slifer KJ, Bauman KE, & Richman GS (1994). Toward a functional analysis of self-injury. Journal of Applied Behavior Analysis, 27, 197–209. 10.1901/jaba.1994.27-197 (Reprinted from Analysis and Intervention in Developmental Disabilities, 2, 3-20, 1982). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimball RT, Greer BD, Randall KR, & Briggs AM (2020). Investigations of operant ABA renewal during differential reinforcement. Journal of the Experimental Analysis of Behavior, 113, 187–205. 10.1002/jeab.577 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimball RT, Kelley ME, Podlesnik CA, Forton A, & Hinkle B (2018). Resurgence with and without an alternative response. Journal of Applied Behavior Analysis, 51, 854–865. 10.1002/jaba.466 [DOI] [PubMed] [Google Scholar]
- Kodak T, Northup J, & Kelley ME (2007). An evaluation of the types of attention that maintain problem behavior. Journal of Applied Behavior Analysis, 40, 167–171. 10.1901/jaba.2007.43-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koegel RL, & Rincover A (1977). Research on the difference between generalization and maintenance in extra-therapy responding. Journal of Applied Behavior Analysis, 10, 1–12. 10.1901/jaba.1977.10-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurtz PF, Boelter EW, Jarmolowicz DP, Chin MD, & Hagopian LP (2011). An analysis of functional communication training as an empirically supported treatment for problem behavior displayed by individuals with intellectual disabilities. Research in Developmental Disabilities, 32, 2935–2942. 10.1016/j.ridd.2011.05.009 [DOI] [PubMed] [Google Scholar]
- Lalli JS, & Casey SD (1996). Treatment of multiply controlled problem behavior. Journal of Applied Behavior Analysis, 29, 391–395. 10.1901/jaba.1996.29-391 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lalli JS, Vollmer TR, Progar PR, Wright C, Borrero J, Daniel D, Barthold CH, Tocco K, & May W (1999). Competition between positive and negative reinforcement in the treatment of escape behavior. Journal of Applied Behavior Analysis, 32, 285–296. 10.1901/jaba.1999.32-285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lattal KA, Cançado CR, Cook JE, Kincaid SL, Nighbor TD, & Oliver AC (2017). On defining resurgence. Behavioural Processes, 141, 85–91. 10.1016/j.beproc.2017.04.018 [DOI] [PubMed] [Google Scholar]
- Lieving GA, Hagopian LP, Long ES, & O’Conner J (2004). Response-class hierarchies and resurgence of severe problem behavior. The Psychological Record, 54, 621–634. [Google Scholar]
- Liggett AP, Nastri R, & Podlesnik CA (2018). Assessing the combined effects of resurgence and reinstatement in children diagnosed with autism spectrum disorder. Journal of the Experimental Analysis of Behavior, 109, 408–421. 10.1002/jeab.315 [DOI] [PubMed] [Google Scholar]
- Mace FC, McComas JJ, Mauro BC, Progar PR, Taylor B, Ervin R, & Zangrillo AN (2010). Differential reinforcement of alternative behavior increases resistance to extinction: Clinical demonstration, animal modeling, and clinical test of one solution. Journal of the Experimental Analysis of Behavior, 93, 349–367. 10.1901/jeab.2010.93-349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marsteller TM, & St. Peter CC (2014). Effects of fixed-time reinforcement schedules on resurgence of problem behavior. Journal of Applied Behavior Analysis, 47, 455–469. 10.1002/jaba.134 [DOI] [PubMed] [Google Scholar]
- Nevin JA, Mace FC, DeLeon IG, Shahan TA, Shamlian KD, Lit K, Sheehan T, Frank-Crawford MA, Trauschke SL, Sweeney MM, Tarver DR, & Craig AR (2016). Effects of signaled and unsignaled alternative reinforcement on persistence and relapse in children and pigeons. Journal of the Experimental Analysis of Behavior, 106, 34–57. 10.1002/jeab.213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nevin JA, McLean AP, & Grace RC (2001). Resistance to extinction: Contingency termination and generalization decrement. Animal Learning & Behavior, 29, 176–191. 10.3758/bf03192826 [DOI] [Google Scholar]
- Parrish JP, Cataldo MF, Kolko DJ, Neef NA, & Egel AL (1986). Experimental analysis of response covariation among compliant and inappropriate behaviors. Journal of Applied Behavior Analysis, 19, 241–254. 10.1901/jaba.1986.19-241 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Payne SW, & Dozier CL (2013). Positive reinforcement as treatment for problem behavior maintained by negative reinforcement. Journal of Applied Behavior Analysis, 46, 699–703. 10.1002/jaba.54 [DOI] [PubMed] [Google Scholar]
- Petscher ES, Rey C, & Bailey JS (2009). A review of empirical support for differential reinforcement of alternative behavior. Research in Developmental Disabilities, 30, 409–425. 10.1016/j.ridd.2008.08.008 [DOI] [PubMed] [Google Scholar]
- Piazza CC, Bowman LG, Contrucci SA, Delia MD, Adelinis JD, & Goh H (1999). An evaluation of the properties of attention as reinforcement for destructive behavior and appropriate behavior. Journal of Applied Behavior Analysis, 32, 437–449. 10.1901/jaba.1999.32-437 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piazza CC, Fisher WW, Hanley GP, Remick ML, Contrucci SA, & Aitken TL (1997). The use of positive and negative reinforcement in the treatment of escape-maintained destructive behavior. Journal of Applied Behavior Analysis, 30, 279–298. 10.1901/jaba.1997.30-279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podlesnik CA, & DeLeon IG (2015). Behavioral Momentum Theory: Understanding persistence and improving treatment. In DiGennaro Reed F & Reed D (Eds.), Autism Service Delivery. Autism and Child Psychopathology Series (pp. 327–351). Springer. 10.1007/978-1-4939-2656-5_12 [DOI] [Google Scholar]
- Podlesnik CA, Kelley ME, Jimenez-Gomez C, & Bouton ME (2017). Renewed behavior produced by context change and its implications for treatment maintenance: A review. Journal of Applied Behavior Analysis, 50, 675–697. 10.1002/jaba.400 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podlesnik CA, & Shahan TA (2009). Behavioral momentum and relapse of extinguished operant responding. Learning & Behavior, 37, 357–364. 10.3758/lb.37.4.357 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pritchard D, Hoerger M, & Mace FC (2014). Treatment relapse and behavioral momentum theory. Journal of Applied Behavior Analysis, 47, 814–833. 10.1002/jaba.163 [DOI] [PubMed] [Google Scholar]
- Querim AC, Iwata BA, Roscoe EM, Schlichenmeyer KJ, Virues Ortega J, & Hurl KE (2013). Functional analysis screening for destructive behavior maintained by automatic reinforcement. Journal of Applied Behavior Analysis, 46, 47–60. 10.1002/jaba.26 [DOI] [PubMed] [Google Scholar]
- Rooker GW, Jessel J, Kurtz PF, & Hagopian LP (2013). Functional communication training with and without alternative reinforcement and punishment: An analysis of 58 applications. Journal of Applied Behavior Analysis, 46, 708–722. 10.1002/jaba.76 [DOI] [PubMed] [Google Scholar]
- Russo DC, Cataldo MF, & Cushing PJ (1981). Compliance training and behavioral covariation in the treatment of multiple behavior problems. Journal of Applied Behavior Analysis, 14, 209–222. 10.1901/jaba.1981.14-209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahan TA, & Craig AR (2017). Resurgence as choice. Behavioural Processes, 141, 100–127. 10.1016/j.beproc.2016.10.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahan TA, & Sweeney MM (2011). A model of resurgence based on behavioral momentum theory. Journal of the Experimental Analysis of Behavior, 95, 91–108. 10.1901/jeab.2011.95-91 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shirley MJ, Iwata BA, Kahng S, Mazaleski JL, & Lerman DC (1997). Does functional communication training compete with ongoing contingencies of reinforcement? An analysis during response acquisition and maintenance. Journal of Applied Behavior Analysis, 30, 93–104. 10.1901/jaba.1997.30-93 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sweeney MM, & Shahan TA (2013). Effects of high, low, and thinning rates of alternative reinforcement on response elimination and resurgence. Journal of the Experimental Analysis of Behavior, 100, 102–116. 10.1002/jeab.26 [DOI] [PubMed] [Google Scholar]
- Tiger JH, Hanley GP, & Bruzek J (2008). Functional communication training: A review and practical guide. Behavior Analysis in Practice, 1(1), 16–23. 10.1007/bf03391716 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkert VM, Lerman DC, Call NA, & Trosclair-Lasserre N (2009). An evaluation of resurgence during treatment with functional communication training. Journal of Applied Behavior Analysis, 42, 145–160. 10.1901/jaba.2009.42-145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wacker DP, Harding JW, Berg WK, Lee JF, Schieltz KM, Padilla YC, Nevin JA, & Shahan TA (2011). An evaluation of persistence of treatment effects during long-term treatment of destructive behavior. Journal of the Experimental Analysis of Behavior, 96, 261–282. 10.1901/jeab.2011.96-261 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wacker DP, Harding JW, Morgan TA, Berg WK, Schieltz KM, Lee JF, & Padilla YC (2013). An evaluation of resurgence during functional communication training. The Psychological Record, 63, 3–20. 10.11133/j.tpr.2013.63.1.001 [DOI] [PMC free article] [PubMed] [Google Scholar]