

Abstract
The Himalayan region is one of the global biodiversity hotspots. However, its biodiversity and ecosystems are threatened due to abiotic and biotic drivers. One of the major biotic threats to biodiversity in this region is the rapid spread of Invasive Alien Species (IAS). Natural forests and grasslands are increasingly getting infested by IAS affecting regeneration of native species and decline in availability of bio-resources. Assessing the current status of IAS and prediction of their future spread would be vital for evolving specific species management interventions. Keeping this in view, we conducted an in-depth study on two IASs, viz., Ageratina adenophora and Lantana camara in the Indian part of Kailash Sacred Landscape (KSL), Western Himalaya. Intensive field surveys were conducted to collect the presence of A. adenophora (n = 567) and L. camara (n = 120) along an altitudinal gradient between 300 and 3000 m a.s.l. We performed Principal Component Analysis to nullify the multi-colinearity effects of the environmental predictors following MaxEnt species distribution model in the current and future climatic scenarios for both the species. All current and future model precision (i.e., Area Under the Curve; AUC) for both species was higher than 0.81. It is predicted that under the current rate of climate change and higher emission (i.e., RCP 8.5 pathway), A. adenophora will spread 45.3% more than its current distribution and is likely to reach up to 3029 m a.s.l., whereas, L. camara will spread 29.8% more than its current distribution range and likely to reach up to 3018 m a.s.l. Our results will help in future conservation planning and participatory management of forests and grasslands in the Kailash Sacred Landscape–India.
Introduction
Invasive alien species (IAS) rank among the top three threats to global biodiversity, the other two being unsustainable harvest of various species from the wild and habitat degradation and loss [1]. Invasive species, coupled with unsustainable resource use and climate change have seriously affected the livelihoods of millions of people in south and south-east Asia [2]. The rapid spread of invasive alien species affects natural habitats, regeneration potential of native species and affects the productivity of forests and grassland habitats [3]. The spread of alien species is particularly challenging in terrestrial ecosystems across the world [4–6]. Changes in climatic conditions and land use practices favour the introduction and spread of IAS in most parts of the world [7–9]. Therefore, it is vital to assess the current extent and future scenarios of invasion for various IAS [10–12].
The Himalayan region, one of the global biodiversity hotspots, is quite vulnerable due to climate change and other abiotic and biotic drivers of change including the rapid spread of IAS. Some of the most obnoxious IAS in this region includes Lantana camara, Ageratina adenophora, Parthenium hysterophorus, and Ageratum conyzoides [13]. During the past few decades increasing anthropogenic pressures such as unabated linear infrastructure development, uncontrolled tourism, livestock grazing, agricultural expansion and extraction of non-timber forest products (NTFPs) have led to the degradation of wildlife habitats and the spread of a large number of invasive species [14]. It is estimated that total 297 IAS species belong to 65 families were reported in the IHR [13]. One of the major causes of the spread of IAS in the Himalayan region is climate change [15]. It has been observed that plant species of higher elevation are projected to shift higher, due to which a few IAS previously limited to the lower elevations are now shifting towards the higher altitudes in the Himalaya [15,16].
As a signatory to Convention on Biological Diversity, India has set its National Target in alignment with Aichi Biodiversity Targets. Accordingly, India’s Biodiversity Strategy and Action Plan aim to identify the priority species of IAS as well as their invasion pathways so that appropriate management strategies could be formulated. It is realized that very few baseline studies have been conducted on the current extent and spread of even common species in the Himalayan region. Anthropogenic pressures coupled with climate change make complex the future prediction of species spread. Several authors [e.g., 15,17–21]; have opined that the IAS which were earlier limited to the lower areas are likely to shift towards the higher elevations in the Himalaya. This calls for the validation of earlier models as well as multi-location studies. In this paper, we present the findings of a case study on two IAS, viz., A. adenophora and L. camara at a landscape level within the Indian part of Kailash Sacred Landscape–India, hereafter referred as KSL–India. The major objectives of the study were to predict the potential current spread of these species in KSL–India and predict their invasion in future climate change scenarios.
Materials and methods
Study area
The study was carried out in KSL–India that is located between 29°26’35" to 30°35’13" N and 80°01’24” to 81°02’44” E. It is characterized by heterogeneous landscape, wide altitudinal range (ca. 800 to over 7000 m a.s.l.), diverse topography and rich biodiversity [17]. This area is contiguous with far-western Nepal and Tibet Autonomous Region (TAR) of China. The study area is spread over 7,120 km2 (Fig 1).
Fig 1. Geographical location of the study area with sampling locations of Lantana camara and Ageratina adenophora.

KSL–India has the predominance of diverse natural ecosystems from grasslands to moist sub-tropical broadleaved to temperate oak forests, sub-alpine conifers, high altitude birch forests, while extensive alpine pastures occur in areas >3000 to 4000 m a.s.l. beside agro-ecosystems in lower reaches [22,23]. The diverse habitats cater to numerous indigenous flora and fauna. The forests in KSL–India falls into two categories—Protected Areas (PAs) and Non-Protected Areas. KSL–India includes only one legally designated PA (i.e. Askot Wildlife Sanctuary) which provides important ecosystem services for the region. However, the non-PA category of forests includes reserve forests and protected forests under the control of the State Forest Department of Uttarakhand, managed for an extended period for the production of timber and non-timber forest products (NTFPs). KSL–India also includes highly interspersed small forest fragments falling under two broad categories, i.e., civil/soyam forests and community forests are under the control of the Revenue Department of the State Government. The latter category of community forests, for nearly nine decades or so, has been managed by the local communities and is referred to as ‘Community Forests’ or ‘Van Panchayat Forests’. This community forest serves as the primary source of livelihood for locals. The KSL–India has experienced a considerable change during recent decades in terms of land use and land cover, along with the development of infrastructure and patterns of natural resources used by the local communities [23,24].
Species presence
After the extensive vegetation survey and consultation with experts, the two most widely spread IAS, i.e., Ageratina adenophora (Asteraceae) and Lantana camara (Verbenaceae) were selected for the present study. A. adenophora is native of Central America [25] and distributed worldwide [26]. L. camara, native to Tropical America [27] is considered to be one of the most troublesome invasive species worldwide [28]. A. Adenophora, a perennial, herb produces a large number of tiny seeds which are dispersed by wind and human activities [29]. Its ubiquitous property to invade diverse habitats, such as roadsides, riverbanks, forest edges, crop fields, wastelands, and rubbish dump edges makes it fit to invade various ecosystems. L. camara is one of the most widely distributed IAS in India [30], resistant to fire and can regrow if burnt [31]. It reproduces primarily by seeds as well as through coppice and prefers to grow in degraded habitats.
Though, the species selected for the study are known to negatively affect the wildlife habitats and native vegetation, they are also known to have a few minor uses to local communities. Leaves of A. adenophora are used for cattle bed [32], or its leaf paste is applied to cuts and wounds and leaves are also have the properties of antimicrobial activity, antipyretic activity, wound healing, antioxidant activity, analgesic activity, anti-tumor activity, insecticidal activity, antiviral activity, anti-inflammatory activity, and larvicidal activity [33,34]. The wood of L. camara is used as fuel [35,36], a source of food for birds [37–39] and nectar for butterflies and moths [40,41] as flowering and fruiting happens throughout the year.
Presence record
We recorded A. adenophora and L. camara presence between 300 and 2500 m a.s.l. covering a wide range of habitats including agricultural lands, grasslands, fallow lands and forests, proximity to major road networks and naturally occurring drainages in different parts of the landscape during 2014 to 2017. A total of 567 and 120 presence locations for A. adenophora and L. camara, respectively were recorded using handheld Garmin N72 GPS (Supplementary Information). Of these, 80% locations were used for probability distribution modeling and the rest 20% were used to evaluate the model performance. The study was conducted outside the protected area and no Schedule-I or -II species were used in the study.
Selection of environmental and bioclimatic variables
We used two-time periods (“current” and “2050”) environmental data to model the A. adenophora and L. camara current and future distribution in KSL–India. A total of 22 environmental variables were used for probability distribution modeling in each period. We retrieved standard 19 bioclimatic variables for WorldClim version 2 [42]. These are the average for the years 1970–2000 with a spatial resolution of 30 seconds (~1 km2). These variables are considered as current bioclimatic variables. We also retrieved 19 bioclimatic variables with a spatial resolution of 30 seconds (~1 km2) for the year 2050 (average for 2041–2060). These are the climate projections as per the Intergovernmental Panel on Climate Change (IPCC) 5th assessment report from Global Climate Models (GCMs) for one Representative Concentration Pathways (RCPs). However, we used RCP 8.5. Because, out of several IPCC climate change scenarios, RCP 8.5, generally taken as the basis for worst-case climate change scenario among all (viz., RCP 2.6, RCP 4.5, RCP 6.0 and RCP 8.5.) [43–45]. The data were downscaled and calibrated using WorldClim 2 as baseline ’current’ climate [42,46]. These bioclimatic data are also the most recent GCM climate projections that have already been considered in the IPCC fifth assessment report.
We also considered four major static topographic factors for both the “current” and “2050” time frame. These are elevation (ASTER, Digital elevation data from Advanced Space-borne Thermal Emission and Reflection Radiometer), slope, aspect, Euclidian distance from water channels and Euclidian distance from major roads. These variables were assumed to be constant across the entire study period. The details of all the variables used in this study are provided in Appendix 1 in S1 Appendix.
Pair-wise Pearson correlations and principal component analysis
We checked for multi co-linearity with pair-wise correlations for both “current” and “2050” environmental data sets to avoid spurious model calibrations [47]. It is common practice to retain only predictors with pair-wise correlation < |0.7| [48]. To nullify the correlation structure of the variables and inter-relationship among them, we implemented the Principal Component Analysis [PCA; 49,50]. The PCA reduces the number of orthogonal predictors each of which is the linear combination of 24 original environmental variables. All the exploratory variables were subjected to PCA to extract the factors of significant contribution using prcomp function and predicting the function of R version 3.3.3 [51]. Furthermore, we obtained 24 Principal Components (PCs) in raster format. For both “current” and “2050” data sets, the first six PCs accounted for 99% of the variability and were chosen for further multivariate species distribution modeling (Table 1).
Table 1. Representation of the first seven principal components derived from all 24 original environmental variables along with the proportion of variance in environmental variables explained by each principal component and their cumulative proportion of variance.
| Principal components | The “current” year | Year 2050 | ||
|---|---|---|---|---|
| Proportion of Variance | Cumulative Proportion | Proportion of Variance | Cumulative Proportion | |
| PC1 | 0.98890 | 0.98890 | 0.98880 | 0.98880 |
| PC2 | 0.00810 | 0.99700 | 0.00810 | 0.99700 |
| PC3 | 0.00276 | 0.99979 | 0.00277 | 0.99973 |
| PC4 | 0.00013 | 0.99992 | 0.00015 | 0.99988 |
| PC5 | 0.00007 | 0.99998 | 0.00008 | 0.99996 |
| PC6 | 0.00001 | 0.99999 | 0.00003 | 0.99999 |
| PC7 | 0.00000 | 1.00000 | 0.00000 | 1.00000 |
MaxEnt species distribution modeling
Maximum Entropy (MaxEnt) distribution modeling was implied to identify the current and future invasion for both, A. adenophora and L. camara in KSL–India. MaxEnt is a machine learning algorithm that is widely used for species distribution modeling that uses presence-only data [52,53] to predict the potential distribution of a particular species in the current as well as future climatic scenarios [54,55].
The occurrence data from the field was collected for both A. adenophora (n = 567) and L. camara (n = 120). One thousand background points were drawn randomly across the KSL–India by considering the pseudo-absence of both species. We generated four main MaxEnt models (two for each species) using the six PCA derived explanatory variables for both the ‘current’ and ‘2050’ year.
We implemented k-fold cross-validation [56] to build each main MaxEnt model. Firstly, we used standard k-fold cross-validation in our randomly partitioned approach. In our k-fold cross-validation approach, the occurrence localities were divided randomly into 10 bins, where each bin constitutes an equal proportion of sample size [57,58]. Then, each model was developed iteratively, using (k − 1) bins for model calibration in each iteration, with the remaining fold retained for evaluation. This is repeated until all the folds were utilized at least once for model evaluation. MaxEnt produces a model based on a series of ‘features’, such as linear, hinge, quadratic, product, threshold and discrete; we used all except for discrete, as all of our explanatory variables were continuous [59]. In the ‘feature’ functions, we set the beta multiplier as 0.5 (medium) that affects the smoothness of the model output. We have opted for the clamping function in each model as the use of the function is strongly suggested for projecting future species distributions [60,61].
As we set the models via k-fold cross-validation (k = 10), for each dataset, we used the respective evaluation localities to calculate Area Under the Curve (AUC) of the Receiver Operation Characteristic (ROC), which is a measure of the overall discriminatory ability of the model or to precisely evaluate model performance by plotting its model sensitivity graph [62]. We averaged those values from each sub-models of each main model and represented the averages for each of the four main MaxEnt models. The AUC score closer to 1 indicates that the model had accurately predicted the habitat, while a value ≤0.5 indicates the low accuracy of the model [63]. We also calculate other model evaluation matrices such as kappa max (kappa), true positive rate (TPR), true negative rate (TNR), and True skill statistics (TSS) using the ‘dismo’ package in R [64].
Binary habitat invasion maps for the ‘current’ and future ‘2050’ were prepared by using the threshold of maximum training sensitivity plus the specificity logistic threshold. The threshold approach of ‘Maximum training sensitivity + specificity’ is a more reliable, restrictive and conservative approach to understand the potential distribution of a species. This particular threshold approach is one of the popular methods either for presence/absence or presence-only data [65]. We used ‘dismo’ package in R [64] along with a source file developed to build all MaxEnt models [66] as well as to evaluate their performance.
Invasion across the elevation gradient
To check the future spread of A. adenophora and L. camara, we extracted all elevational range values from a 30 m resolution digital elevation model (ASTER) using the threshold polygons generated from MaxEnt main models. Elevation gradients that fall within the distribution ranges were re-classified into multiple classes considering 100 m a.s.l. intervals. Area invaded by both species in each elevation class was compared between current and 2050 year using paired sample t—test. The distributions of elevation class vs. invaded areas of both A. adenophora and L. camara were plotted separately for the current year and 2050 to visually assess their spread pattern across the elevation classes.
Results
The pair-wise Pearson correlations were generally higher (r >|0.70|) for most of the variables (Fig 2). We were not able to use these variables directly in MaxEnt as they showed high correlation, and this avoids the overprediction of suitability in MaxEnt models. However, PCA transformed the 24 environmental variables (each for ‘current’ and ‘2050’) into a composite set of 24 components that are orthogonal and un-correlated to each other. Six out of 24 components demonstrated the cumulative proportion of variance > 0.99 for both the ‘current’ and the year ‘2050’ (Table 1). The first six PCs each from the ‘current’ and ‘2050’ were represented in Appendices 2–3 in S1 Appendix.
Fig 2. Representation of Pearson correlation matrix for environmental data of the “current” year (A) and year 2050 (B).

Both training and test area under ROC curve (AUC) values were > 0.81 for both species under the ‘current’ and future ‘2050’ climatic conditions (Fig 3, Table 2). Other model evaluation statistics were provided in Appendix 4 in S1 Appendix. The current occurrence locations of A. adenophora and L. camara were 95.1% and 92.5%, respectively falls within their most climatically suitable current distribution ranges. The relative percent contribution of all PCs from both the ‘current’ and the year ‘2050’ are presented in Table 3, and all response curves associated with the above MaxEnt model predictions were given in Appendix 5 in S1 Appendix. The current and future distributions of the A. adenophora and L. camara along the elevation gradient of KSL–India are determined by the RCP 8.5 pathway projections for 2050 (Fig 4A and 4B).
Fig 3. Receiver operating characteristic (ROC) curve for the predicted distribution MaxEnt models averaged over the ten replicate runs: Ageratina adenophora (the “current” year–A; year 2050 –B) and Lantana camara (the “current” year–C; year 2050 –D).
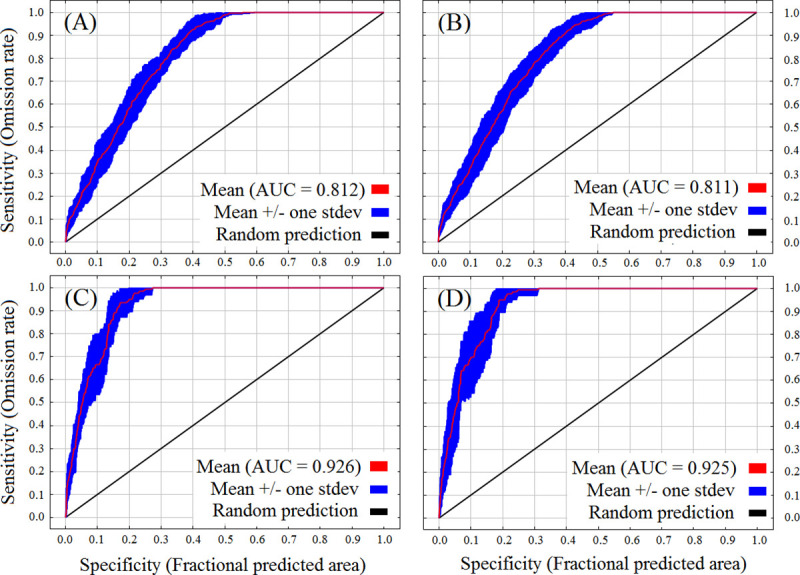
Table 2. The area under the receiver operating curve (AUC) score of MaxEnt models for both Ageratina adenophora and Lantana camara.
| Species | The “current” year | Year 2050 | ||
|---|---|---|---|---|
| Test AUC | Training AUC | Test AUC | Training AUC | |
| Ageratina adenophora | 0.81 | 0.82 | 0.81 | 0.82 |
| Lantana camara | 0.93 | 0.93 | 0.92 | 0.93 |
Table 3. Relative contributions of each principal component (PC) are used in all the MaxEnt models.
| PC | Contribution of Ageratina adenophora (%) | Contribution of Lantana camara (%) | ||
|---|---|---|---|---|
| The “current” year | Year 2050 | The “current” year | Year 2050 | |
| PC1 | 83.00 | 86.10 | 62.8 | 65.2 |
| PC2 | 00.00 | 00.10 | 0.1 | 0.2 |
| PC3 | 01.10 | 02.40 | 9.1 | 15.6 |
| PC4 | 04.60 | 04.60 | 2.1 | 1.1 |
| PC5 | 07.30 | 05.40 | 16.7 | 15.8 |
| PC6 | 04.00 | 01.50 | 9.1 | 2.2 |
Fig 4. Predicted distribution MaxEnt models averaged over the ten replicate runs.

Orange and Red polygons represent maximum training sensitivity + specificity logistic threshold for the distribution of (A) Ageratina adenophora and (B) Lantana camara in the “current” and year 2050.
The current distribution of A. adenophora ranges between 212–2616 m a.s.l., while L. camara distributed mostly below 2045 m a.s.l. The potential area that is currently vulnerable to invasion by A. adenophora and L. camara extends to 1403.7 km2 and 1053.1 km2, respectively. If the climate change continues to follow the higher emission RCP 8.5 pathway, by 2050, invasion of A. adenophora would reach up to 3029 m a.s.l., whereas, L. camara will invade up to 3018 m a.s.l. Both A. adenophora and L. camara will spread across the landscape occupying an area of 2039.7 km2 and 1366.5 km2, respectively. The area invasion by A. adenophora (t = -7.61, df = 28, p ≤ 0.05) and L. camara (t = -2.70, df = 28, p ≤ 0.05) will significantly increase by year 2050 across all designated elevation class (Fig 5).
Fig 5. The predicted spread (in Km2) of Ageratina adenophora (A) and Lantana camara (B) in different elevation classes of KSL–India during the current year and the year 2050.

Discussion
Plant invasion is the emerging area of science that impacts substantial economic and ecological imbalance by altering native plant community composition [67] and depleting native plant species diversity [68] thus affecting ecosystem processes [69–71]. The lower and mid ecosystems in the mountains are more vulnerable to invasion by exotic plant species due to the altering climate change and increasing anthropogenic pressure [72]. These plant species also exceeded their ranges and many of them have become abundant at higher elevations [73,74]. The increasing incidence of invasion in the high-altitude ecosystems [9] poses a significant threat to the native biological diversity of the region and it is expected to expand in the future [21]. Using principal components as input variables in MaxEnt enhances the prediction of good habitat suitability [50,75]. Alien species that seek to establish themselves in mountains easily proliferate and become difficult to sustain and manage due to the rugged and steppe mountain terrains [76]. It is important to detect potential invaders and take preventative action, since this is the most cost-effective way to restrict the spread of IAPS to new locations. This demonstrated an increasing (both vertical and horizontal) trend of plant invasion in KSL–India. Altering climatic condition and mostly increased temperature has been providing favourable conditions for the invasion and spread of these species in the KSL–India.
The presence of L. camara was recorded widely in forest and scrublands [77,78]. Looking forward, L. camara might also affect the regeneration of native biodiversity in the area. It appeared that in the future both of these IAS would colonize more vigorously in the landscape where currently they are not present. This invasion of both species is likely to happen if climate change continues to follow the higher emission RCP 8.5 pathway. Furthermore, L. camara will spread within a similar altitudinal range where it is currently absent. These species have already spread in mountains and along rivers and roadside [79]. A. adenophora presence between 2800 to 3000 m a.s.l. [13], which suggest that the spread of this species in higher regions of western Himalaya can cause a potential threat to the native biodiversity in the alpine and subalpine zone.
Our study also showed a similar trend, as this species will reach 968 m a.s.l. higher than its current distribution range. Our predictive models also depicted high infestation and expansion in the present climatic scenario by A. adenophora and L. camara in the lower parts of KSL–India in western Himalaya. The MaxEnt model also predicted that the landscape provides more suitable conditions for the infestation of A. adenophora as compared to L. camara even at its current climatic condition. Both the species will expand their habitats with the increase in temperature and human activity in the landscape. Our study demonstrated similar results as compared to previous studies [80–82], but in a more robust and spatially explicit manner.
Our study has come up with a current pattern of distribution and established future scenarios of invasion by two highly obnoxious IAS within KSL–India. The distribution maps generated in this exercise would be of much help in further conservation planning and participatory management of forest/grassland management. The findings of our study can be used in the identification of vulnerable areas and important localities which are likely to be infested by these species. Habitat restoration and community-based monitoring at a local scale involving Biodiversity Management Committee (BMC) would be one of the logical steps forward.
Supporting information
(DOCX)
(RAR)
Acknowledgments
The authors are thankful to the Director and Dean, Wildlife Institute of India for their support, and encouragement. Thanks are also due to Uttarakhand Forest Department for granting permission to work in the landscape. We would like to thank Dr. Arti Kala, Mr. Sumit Arya and Dr. Ajaz Hussain for their support during the survey and Dr. Naveen Joshi, Mr. Shahid Dar and Mr. Anukul Nath for providing valuable suggestions during data analysis and interpretation. We would also like to thank Mr. Balwant and Late Hari Shankar Pandey for assisting in the field as well as our gratitude to the local community for help, support and cooperation.
Data Availability
Data for this paper is provided in the Supporting Information files.
Funding Statement
This study was funded by the National Mission on Himalayan Studies to AC (WII/RES/PERS/ALKA CHAUDHARY; 464) and also by International Centre for Integrated Mountain Development (ICIMOD), Nepal under Kailash Sacred Landscape Conservation and Development Initiative - India project to GSR & BSA (WII/KSLCDI-India/2013). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Early R, Bradley BA, Dukes JS, Lawler JJ, Olden JD, Blumenthal DM, et al. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat Commun. 2016; 7: 1–9. 10.1038/ncomms12485 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.IPBES. Summary for policy makers of the regional assessment report on biodiversity and ecosystem services for Asia and the Pacific of the Intergovernmental Science–Policy Platform on Biodiversity and Ecosystem Services. IPBES Secretariat. Bonn, Germany; 2018. pp. 41. [Google Scholar]
- 3.Rai PK, Singh JS. Invasive alien plant species: Their impact on environment, ecosystem services and human health. Ecol Indic. 2020; 111: p.106020. 10.1016/j.ecolind.2019.106020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dukes JS, Mooney HA. Does global change increase the success of biological invaders? Trends Ecol Evol. 1999; 14: 135–139. 10.1016/s0169-5347(98)01554-7 [DOI] [PubMed] [Google Scholar]
- 5.Mack RN, Simberloff D, Mark Lonsdale W, Evans H, Clout M, Bazzaz FA. Biotic invasions: causes, epidemiology, global consequences, and control. Ecol Appl. 2000; 10: 689–710. https://doi.org/10.1890/1051-0761(2000)010[0689: BICEGC]2.0.CO;2. [Google Scholar]
- 6.Lin W, Zhou G, Cheng X, Xu R. Fast economic development accelerates biological invasions in China. PLoS One. 2007; 2: e1208. 10.1371/journal.pone.0001208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Vila M, Weiner J. Are invasive plant species better competitors than native plant species?–evidence from pair‐wise experiments. Oikos. 2004; 105: 229–238. 10.1111/j.0030-1299.2004.12682.x. [DOI] [Google Scholar]
- 8.Hellmann JJ, Byers JE, Bierwagen BG, Dukes JS. Five potential consequences of climate change for invasive species. Conserv biol. 2008; 22: 534–543. 10.1111/j.1523-1739.2008.00951.x [DOI] [PubMed] [Google Scholar]
- 9.Thapa S, Chitale V, Rijal SJ, Bisht N, Shrestha BB. Understanding the dynamics in distribution of invasive alien plant species under predicted climate change in Western Himalaya. PloS One. 2018; 13: e0195752. 10.1371/journal.pone.0195752 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kohli RK, Dogra KS, Batish DR, Singh HP. Impact of Invasive Plants on the Structure and Composition of Natural Vegetation of Northwestern Indian Himalayas. Weed Technol. 2004; 18: 1296–1301. https://doi.org/10.1614/0890-037X(2004)018[1296: IOIPOT]2.0.CO;2. [Google Scholar]
- 11.Tylianakis JM, Didham RK, Bascompte J, Wardle DA. Global change and species interactions in terrestrial ecosystems. Ecol Lett. 2008; 11: 1351–1363. 10.1111/j.1461-0248.2008.01250.x [DOI] [PubMed] [Google Scholar]
- 12.Sekar KC. Invasive alien plants of Indian Himalayan region—diversity and implication Am J Plant Sci. 2012; 3: 177–184. 10.4236/ajps.2012.32021. [DOI] [Google Scholar]
- 13.Pathak R, Negi VS, Rawal RS, Bhatt ID. Alien plant invasion in the Indian Himalayan Region: state of knowledge and research priorities. Biodivers Conserv. 2019; 28: 3073–3102. 10.1007/s10531-019-01829-1. [DOI] [Google Scholar]
- 14.Babu S, Love A, Babu CR. Ecological restoration of lantana-invaded landscapes in Corbett Tiger Reserve, India. Ecol Restor. 2009; 27: 467–477. 10.3368/er.27.4.467. [Google Scholar]
- 15.Telwala Y, Brook BW, Manish K, Pandit MK. Climate-induced elevational range shifts and increase in plant species richness in a Himalayan biodiversity epicenter. PloS One. 2013; 8: e57103. 10.1371/journal.pone.0057103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shrestha BB, Joshi S, Bisht N, Yi S, Kotru R, Chaudhary RP et al. Inventory and impact assessment of invasive alien plant species in Kailash Sacred Landscape: ICIMOD working paper 2018/2. ICIMOD Working Paper id: 12674, e Social Sciences; 2018. http://lib.icimod.org/record/33744.
- 17.Oli KP, Zomer R; (eds.) Kailash sacred landscape conservation initiative–Feasibility assessment report. Kathmandu: ICIMOD, Nepal; 2011. p. 1–92. [Google Scholar]
- 18.Taylor S, Kumar L, Reid N. Impacts of climate change and land‐use on the potential distribution of an invasive weed: a case study of Lantana camara in Australia. Weed Research. 2012; 52: 391–401. 10.1111/j.1365-3180.2012.00930.x. [DOI] [Google Scholar]
- 19.Goncalves E, Herrera I, Duarte M, Bustamante RO, Lampo M, Velasquez G et al. Global invasion of Lantana camara: has the climatic niche been conserved across continents? PLoS One. 2014; 9: e111468. 10.1371/journal.pone.0111468 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Uddin K, Chaudhary S, Chettri N, Kotru R, Murthy M, Chaudhary RP et al. The changing land cover and fragmenting forest on the Roof of the World: A case study in Nepal’s Kailash Sacred Landscape. Landsc Urban Plan. 2015; 141: 1–10. 10.1016/j.landurbplan.2015.04.003. [DOI] [Google Scholar]
- 21.Mungi NA, Coops NC, Ramesh K, Rawat GS. How global climate change and regional disturbance can expand the invasion risk? Case study of Lantana camara invasion in the Himalaya. Biol Invasions. 2018; 20: 1849–1863. 10.1007/s10530-018-1666-7. [DOI] [Google Scholar]
- 22.Singh SP, Bassignana-Khadka I, Singh Karky B, Sharma E. Climate change in the Hindu Kush-Himalayas: the state of current knowledge International Centre for Integrated Mountain Development (ICIMOD), Nepal; 2011. pp. 1–88. [Google Scholar]
- 23.Singh G, Sarkar MS, Pandey A, Lingwal S, Rai ID, Adhikari BS, et al. Quantifying Four Decades of Changes in Land Use and Land Cover in India’s Kailash Sacred Landscape: Suggested Option for Priority Based Patch Level Future Forest Conservation. Journal of the J Indian Soc Remote Sens. 2018; 46: 1625–1635. 10.1007/s12524-018-0817-8. [DOI] [Google Scholar]
- 24.Negi VS, Pathak R, Sekar KC, Rawal RS, Bhatt ID, Nandi SK et al. Traditional knowledge and biodiversity conservation: a case study from Byans Valley in Kailash Sacred Landscape, India. J Environ Plan Manag. 2018; 61: 1722–1743. 10.1080/09640568.2017.1371006. [DOI] [Google Scholar]
- 25.Datta A, Kühn I, Ahmad M, Michalski S, Auge H. Processes affecting altitudinal distribution of invasive Ageratina adenophora in western Himalaya: The role of local adaptation and the importance of different life-cycle stages. PloS One. 2017; 12: e0187708. 10.1371/journal.pone.0187708 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Cronk QC, Fuller JL. Plant invaders: the threat to natural ecosystems. London: Chapman and Hall; 1995. pp. 241. [Google Scholar]
- 27.Holm LG, Plucknett DL, Pancho JV, Herberger JP. The world’s worst weeds: Distribution and biology. University press of Hawaii; 1977. p. 609. [Google Scholar]
- 28.Kohli RK, Batish DR, Singh HP, Dogra KS. Status, invasiveness and environmental threats of three tropical American invasive weeds (Parthenium hysterophorus L, Ageratum conyzoides L, Lantana camara L.) in India. Biol Invasions. 2006; 8: 1501–1510. 10.1007/s10530-005-5842-1. [DOI] [Google Scholar]
- 29.Sang W, Zhu L, Axmacher JC. Invasion pattern of Eupatorium adenophorum Spreng in southern China. Biol Invasions. 2010; 12: 1721–1730. 10.1007/s10530-009-9584-3. [DOI] [Google Scholar]
- 30.Sundaram B, Krishnan S, Hiremath AJ, Joseph G. Ecology and impacts of the invasive species, Lantana camara, in a social-ecological system in South India: perspectives from local knowledge. Hum Ecol. 2012; 40: 931–942. 10.1007/s10745-012-9532-1. [DOI] [Google Scholar]
- 31.Hiremath AJ, Sundaram B. The fire-lantana cycle hypothesis in Indian forests. Conserv Soc. 2005; 3: 26–42. http://eprints.atree.org/id/eprint/22. [Google Scholar]
- 32.Shrestha TB. Development ecology of the Arun river basin in Nepal Country report on biological diversity, ICIMOD, Kathmandu, Nepal; 1989. pp. 1–121. [Google Scholar]
- 33.Joseph J, Kharkongor P. A preliminary ethno-botanical survey in the Khasi and Jaintia hills, Meghalaya. In: Jain S.K (Ed.): Glimpses of Indian Ethnobotany. New Delhi: Oxford IBH Publishing Co; 1981. pp. 115–123. [Google Scholar]
- 34.Poudel R, Neupane NP, Mukeri IH, Alok S, Verma A. An updated review on invasive nature, phytochemical evaluation, & pharmacological activity of Ageratina adenophora. Int J Pharm Sci Res. 2020; 11: 2510–2520. [Google Scholar]
- 35.Kumar R, Chandrashekar N, Pandey KK. 2009. Fuel properties and combustion characteristics of Lantana camara and Eupatorium spp. Curr Sci. 97: 930–935. [Google Scholar]
- 36.Patel S. 2011. A weed with multiple utility: Lantana camara. Rev Environ Sci Biotechnol. 10: 341–351. [Google Scholar]
- 37.Corlett RT. Frugivory and seed dispersal by birds in Hong Kong shrub-land. Forktail 1998; 13: 23–27. [Google Scholar]
- 38.Bhatt D, Kumar A. Foraging ecology of Red-vented bulbul Pycnonotus cafer in Haridwar, India. Forktail. 2001; 17: 109–109. [Google Scholar]
- 39.Aruna R, Balasubramanian P. Fruiting phenology and avian frugivory on Lantana camara in a mixed dry deciduous forest, Western Ghats, Tamilnadu, India. Int J Fauna Biol. 2017; 4: 179–182. [Google Scholar]
- 40.Ram HM, Mathur G. Flower color changes in Lantana camara. Journal of experimental botany. 1984; 35: 1656–1662. [Google Scholar]
- 41.Day MD, Wiley CJ, Playford J, Zalucki MP. Lantana: Current Management, Status and Future Prospects. Australian Centre for International Agricultural Research. 2003; 5: 1–20. [Google Scholar]
- 42.Fick SE, Hijmans RJ. WorldClim 2: new 1‐km spatial resolution climate surfaces for global land areas. Int J Climatol. 2017; 37: 4302–4315. 10.1002/joc.5086. [DOI] [Google Scholar]
- 43.Hausfather Z, Peters GP. Emissions–the ‘business as usual’ story is misleading Nature. 2020; 577: 618–20. 10.1038/d41586-020-00177-3 . [DOI] [PubMed] [Google Scholar]
- 44.Schwalm CR, Glendon S, Duffy PB. RCP8. 5 tracks cumulative CO2 emissions. Proceedings of the National Academy of Sciences. 2020; 117: 19656–19657. ISSN 0027-8424. 10.1073/pnas.2007117117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Collins M. Knutti R, Arblaster J, Dufresne JL, Fichefet T, Friedlingstein P, et al. Long-term climate change: projections, commitments and irreversibility. In Climate Change 2013-The Physical Science Basis: Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press. 2013; pp. 1029–1136. [Google Scholar]
- 46.Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A. Very high-resolution interpolated climate surfaces for global land areas. Int J Climatol. 2005; 25: 1965–1978. 10.1002/joc.1276. [DOI] [Google Scholar]
- 47.Guisan A, Thuiller W. Predicting species distribution: offering more than simple habitat models. Ecol Lett. 2005; 8: 993–1009. 10.1111/j.1461-0248.2005.00792.x. [DOI] [PubMed] [Google Scholar]
- 48.Aguirre-Gutiérrez J, Carvalheiro LG, Polce C, van Loon EE, Raes N, Reemer M, Biesmeijer JC. Fit-for-purpose: species distribution model performance depends on evaluation criteria–Dutch hoverflies as a case study. PloS one. 2013; 8, p.e63708. 10.1371/journal.pone.0063708 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Jolliffe I.T (Ed.) Principal Component Analysis (2nd Eds.) Aberdeen, UK: Springer; 2002. pp. 488. [Google Scholar]
- 50.Cruz-Cárdenas G, López-Mata L, Villaseñor JL, Ortiz E. Potential species distribution modeling and the use of principal component analysis as predictor variables. Revista Mexicana de Biodiversidad. 2014; 85:189–199. [Google Scholar]
- 51.Team R Core. R: A Language and Environment for Statistical Computing. Vienna, Austria. 2018 Jan 25. Available from: https://www.R-project.org/.
- 52.Phillips SJ, Anderson RP, Schapire RE. Maximum entropy modeling of species geographic distributions. Ecol Modell. 2006; 190: 231–259. 10.1016/j.ecolmodel.2005.03.026. [DOI] [Google Scholar]
- 53.Elith J, Phillips SJ, Hastie T, Dudík M, Chee YE, Yates CJ. A statistical explanation of MaxEnt for ecologists. Divers Distrib. 2011; 17: 43–57. 10.1111/j.1472-4642.2010.00725.x. [DOI] [Google Scholar]
- 54.Loarie SR, Carter BE, Hayhoe K, et al. Climate change and the future of California’s endemic flora. PloS One. 2008; 3: e2502. 10.1371/journal.pone.0002502 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Remya K, Ramachandran A, Jayakumar S. Predicting the current and future suitable habitat distribution of Myristica dactyloides Gaertn using MaxEnt model in the Eastern Ghats, India. Ecol Eng. 2015; 82: 184–188. 10.1016/j.ecoleng.2015.04.053. [DOI] [Google Scholar]
- 56.Peterson AT, Soberón J, Pearson RG, Anderson RP, Martínez-Meyer E, Nakamura M, et al. Ecological niches and geographic distributions. Princeton, UK: Princeton University Press. 2011. pp. 328. [Google Scholar]
- 57.Boyce MS, Vernier PR, Nielsen SE, Schmiegelow FK. Evaluating resource selection functions. Ecol Modell. 2002; 157: 281–300. 10.1016/S0304-3800(02)00200-4. [DOI] [Google Scholar]
- 58.Lehmann A, Overton JM, Leathwick JR. GRASP: generalized regression analysis and spatial prediction. Ecol Modell. 2002; 157: 189–207. 10.1016/S0304-3800(02)00195-3. [DOI] [Google Scholar]
- 59.Phillips SJ, Dudík M. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography. 2008; 31: 161–175. 10.1111/j.0906-7590.2008.5203.x. [DOI] [Google Scholar]
- 60.Elith J, Kearney M, Phillips S. The art of modelling range‐shifting species. Methods Ecol. Evol. 2010; 1: 330–342. 10.1111/j.2041-210X.2010.00036.x. [DOI] [Google Scholar]
- 61.Webber BL, Yates CJ, Le Maitre DC, Scott JK, Kriticos DJ, Ota N, et al. Modelling horses for novel climate courses: insights from projecting potential distributions of native and alien Australian acacias with correlative and mechanistic models. Divers Distrib. 2011; 17: 978–1000. 10.1111/j.1472-4642.2011.00811.x. [DOI] [Google Scholar]
- 62.Peterson AT, Papeş M, Soberón J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol modell. 2008; 213: 63–72. 10.1016/j.ecolmodel.2007.11.008. [DOI] [Google Scholar]
- 63.Hanley JA, McNeil BJ. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology. 1982; 143: 29–36. 10.1148/radiology.143.1.7063747 [DOI] [PubMed] [Google Scholar]
- 64.Hijmans RJ, Phillips S, Leathwick J, Elith J, Hijmans MRJ. Package ‘dismo’ Circles. 2017; 9: 1–68. [Google Scholar]
- 65.Liu C, Berry PM, Dawson TP, et al. Selecting thresholds of occurrence in the prediction of species distributions. Ecography. 2005; 28: 385–393. 10.1111/j.0906-7590.2005.03957.x. [DOI] [Google Scholar]
- 66.Feng X, Gebresenbet F, Walker C. Shifting from closed-source graphical-interface to open-source programming environment: a brief tutorial on running Maxent in R (No e3346v1). 2017; PeerJ Preprints. 10.7287/peerj.preprints.3346v1. [DOI] [Google Scholar]
- 67.Gordon DR. Effects of invasive, non‐indigenous plant species on ecosystem processes: lessons from Florida. Ecol Appl. 1998; 8: 975–989. 10.1890/1051-0761(1998)008[0975:EOINIP]2.0.CO;2. [DOI] [Google Scholar]
- 68.Hejda M, Pyšek P Jarošík V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J Ecol. 2009; 97: 393–403. 10.1111/j.1365-2745.2009.01480.x. [DOI] [Google Scholar]
- 69.Weidenhamer JD, Callaway RM. Direct and indirect effects of invasive plants on soil chemistry and ecosystem function. J Chem Ecol. 2010; 36: 59–69. 10.1007/s10886-009-9735-0 [DOI] [PubMed] [Google Scholar]
- 70.Crooks JA. Characterizing ecosystem‐level consequences of biological invasions: the role of ecosystem engineers. Oikos. 2002; 97(2): 153–166. 10.1034/j.1600-0706.2002.970201.x. [DOI] [Google Scholar]
- 71.Dukes JS, Mooney HA. Disruption of ecosystem processes in western North America by invasive species. Revista Chilena de Historia Natural. 2004; 77: 411–437. 10.4067/S0716-078X2004000300003. [DOI] [Google Scholar]
- 72.Priyanka N, Joshi PK. A review of Lantana camara studies in India. IJSRP. 2013; 3: 1–11. [Google Scholar]
- 73.Alexander JM, Lembrechts JJ, Cavieres LA, Daehler C, Haider S, Kueffer C, et al. Plant invasions into mountains and alpine ecosystems: current status and future challenges. Alp. Bot. 2016; 126: 89–103. 10.1007/s00035-016-0172-8. [DOI] [Google Scholar]
- 74.Pauchard A, Kueffer C, Dietz H, Daehler CC, Alexander J, Edwards PJ, et al. Ain’t no mountain high enough: plant invasions reaching new elevations. Front. Ecol. Environ. 2009; 7: 479–486. 10.1890/080072. [DOI] [Google Scholar]
- 75.Moya W, Jacome G, and Yoo C. Past, current, and future trends of red spiny lobster based on PCA with MaxEnt model in Galapagos Islands, Ecuador. Ecol Evol. 2017; 7: 4881–4890. 10.1002/ece3.3054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.McDougall KL, Alexander JM, Haider S, Pauchard A, Walsh NG, Kueffer C. Alien flora of mountains: global comparisons for the development of local preventive measures against plant invasions. Divers Distrib. 2011; 17:103–111. 10.1111/j.1472-4642.2010.00713.x. [DOI] [Google Scholar]
- 77.Bisht N, Joshi S, Shrestha BB, Yi S, Chaudhary RP, Kotru R, et al. Manual on invasive alien plant species in Kailash Sacred Landscape-Nepal. Manual on invasive alien plant species in Kailash Sacred Landscape-Nepal. 2016; pp.44. [Google Scholar]
- 78.Shrestha BB, Joshi S, Bisht N, Yi S, Kotru R, Chaudhary RP, et al. Inventory and impact assessment of invasive plant species in Kailash Sacred Landscape ICIMOD Working Paper 2018 (in press).
- 79.Singh G, Padalia H, Rai ID, Bharti RR, Rawat GS. Spatial extent and conservation status of Banj oak (Quercus leucotrichophora A Camus) forests in Uttarakhand, Western Himalaya. Trop Ecol. 2016; 57: 255–262. [Google Scholar]
- 80.Reese GC, Wilson KR, Hoeting JA, Flather CH. Factors affecting species distribution predictions: a simulation modeling experiment. Ecol Appl. 2005; 15: 554–564. 10.1890/03-5374. [DOI] [Google Scholar]
- 81.Hernandez PA, Graham CH, Master LL, Albert DL. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography. 2006; 29: 773–785. 10.1111/j.0906-7590.2006.04700.x. [DOI] [Google Scholar]
- 82.Wisz MS, Hijmans RJ, Li J, Peterson AT, Graham CH, et al. Predicting Species Distributions Working Group: Effects of sample size on the performance of species distribution models. Divers Distrib. 2008; 14: 763–773. 10.1111/j.1472-4642.2008.00482.x. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(RAR)
Data Availability Statement
Data for this paper is provided in the Supporting Information files.