

Introduction
“Nothing in biology makes sense except in the light of evolution”[1] and this is no less true for viral pathogens than it is for their host species. To appreciate the full extent of the challenges posed by current and future emerging infectious diseases (EIDs), I believe it is insightful to take a coevolutionary perspective. Coevolution comprises the reciprocal, adaptive genetic changes that occur when two species interact. During coevolution, the direction and intensity of natural selection is constantly changing because adaptations in one species provoke counter-adaptations in the coevolving species. Despite continuous adaptations, none of the interacting species gain a sustained fitness advance, making antagonistic coevolution a zero-sum game [2–4]. This is also why coevolution has been likened to the character of the Red Queen in Lewis Carroll’s “Through the Looking Glass”, who said: “ … it takes all the running you can do, to keep in the same place”. But what would happen if one of the parties (our livestock and crops) lacked the coevolutionary means to run, whilst the other party (their parasites) had just received a significant fitness performance-enhancing boost? Here, I will discuss how recent changes in interactions between the evolutionary forces have fundamentally altered the Red Queen dynamics between parasites and their hosts. I believe we have created the perfect coevolutionary storm, and the question is: what can we do about it now?
Evolutionary forces
There are five evolutionary forces; three that create genetic variation (mutation, recombination and gene flow), one that changes the distribution of genetic variation randomly (genetic drift), and one force that molds the genetic variation (selection).
Mutation
Ultimately, the origin of all genetic variation stems from mutations[5]. The pathogens with the highest mutation rate are the RNA viruses (Figure 1a). Although this group represents less than 25% of the pathogens that account for EIDs, they are responsible for the most devastating outbreaks in the past decades: HIV, influenza H1N1 and H5H1, severe acute respiratory syndrome coronavirus, Lassa virus, Ebola virus, the Middle East respiratory syndrome coronavirus, and SARS-CoV-2 [6–8]. The diversity generated by mutation helps in the evasion of host responses, and these so-called “mutant clouds” also enable RNA viruses to replicate in multiple host species[9]. Given their high mutation rate[10], rapid rate of replication, and vast population size [11] (Figure 1b), RNA viruses were the first out of the evolutionary starting blocks, able to take best advantage of the new opportunities in our rapidly changing world. With the vast population of well-interconnected human hosts and livestock (Figure 1c), the size of the gene pool of many zoonotic pathogens has significantly increased, thereby augmenting their evolutionary potential (cf. Fundamental Theorem of Natural Selection [12]). Indeed, zoonotic virus richness in domesticated mammalian species is highly correlated with the global abundance of humans and domesticated species[13]. The high mutation rate and abundance of virions makes the development of durable treatments more challenging[9]. This has implications also for the current COVID-19 pandemic, which in all likelihood will require annual booster vaccinations against SARS-CoV-2 to ensure vaccine efficiency and avoid the outbreak of new variants in the future. A vaccination program will work most effectively when hand in glove with public health surveillance and contact tracing, to identify people who have been in close contact those infected with variants of concern to minimize their spread.
Figure 1.
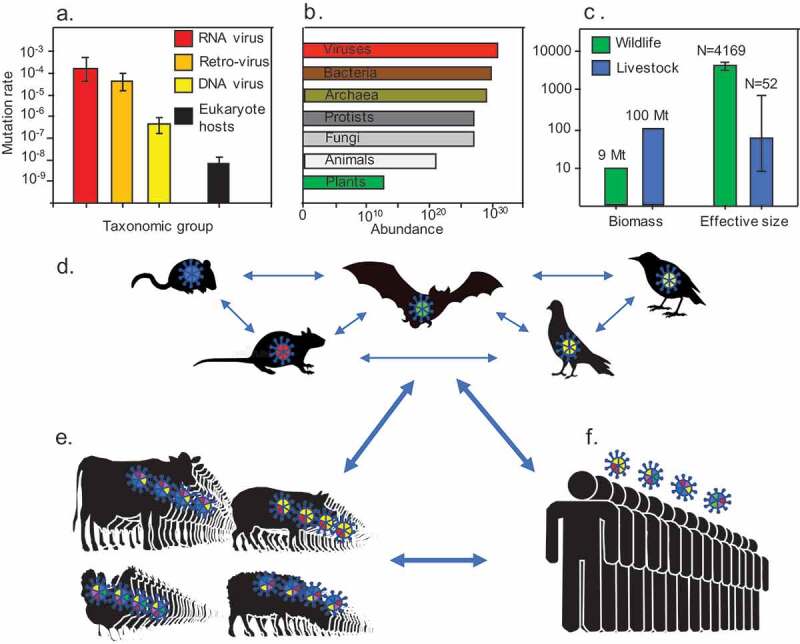
Coevolutionary disequilibrium between livestock, humans and pathogens. (a) Viruses have an extremely high mutation rate.[10], and (b) a high abundance[11]. In combination with their rapid rate of replication, these properties give viruses a high evolutionary potential. (c) The biomass of livestock is over an order of magnitude higher than that of all wildlife combined[11], yet their genetic effective size [30] is 80 times lower than the minimum viable population size of free-living species[31], which makes livestock vulnerable to infectious disease. (d) Habitat destruction, illegal wildlife trade and human activities have increased gene flow and brought many species into contact, which is facilitating spillover and hybridization of pathogens[16]. (e) The highly abundant and genetically depleted livestock act as “mixing vessels” where different pathogens can hybridize[7]. (f) Close contact between livestock and humans can lead to spillovers, which make zoonotic hybrid pathogens a significant threat to human health now and in the future[40]
Recombination
By mixing the already available genetic variation into new combinations, recombination can also generate evolutionary novelty. Unlike mutation, recombination is not a strictly blind process, given that the segregating variation has already been “tried and tested” by natural selection in another genomic background. Furthermore, recombination acts fast and it can have profound effects by swapping multiple nucleotides in a single event. In sexually reproducing organisms, meiotic recombination mixes the genetic contributions from both parents in the offspring, which is the principle evolutionary advantage of sex[14]. Although many pathogens reproduce asexually, some can still recombine their genome by template switching[15], i.e. the exchange between closely related (homologous) genes from other viruses. Several studies have documented evidence of genetic recombination in coronaviruses, both in animals, such as MHV, TGEV, and feline and canine coronaviruses, as well as in human-infecting coronaviruses, including MERS-CoV, SARS-CoV-1 (reviewed in Su et al [6].), and SARS-CoV-2[8]. Recombination can rapidly increase the level of variation, but its potential to generate evolutionary novelty is considerably enhanced when combined with another evolutionary force – gene flow.
Gene flow
This evolutionary force is perhaps most under our control, as it involves the physical movement of plants, animals, humans, and their associated pathogens. Such migration results in gene flow when it leads to genetic exchange between otherwise isolated gene pools. However, moderating gene flow has significant social-economic consequences as decision makers need to trade-off the benefits of economic growth, and people’s well-being and education, against the costs to human health and the environment. In addition, climate change can further elevate the rate of gene flow of the pathogens associated with humans, livestock and wildlife. Gene flow of pathogens can also be mediated by natural vectors, and it can be an indirect result of habitat destruction, environmental change, illegal wildlife trade, and other human activities [16,17]. Migration and the gene flow of SARS-CoV-2 during the COVID-19 pandemic need to be minimized, which requires difficult political decisions and broad public support for a vaccine passport (or “immunity passport”, “immunity-based license”), such as the UK’s “COVID status certification” and the EU’s “Digital Green Certificate”. Vaccine passports provide proof that a person has been vaccinated against COVID-19, received a negative test result, or recently recovered from COVID-19, and they offer three important benefits: (1) helping to open up the economy and society more rapidly; (2) protecting lives by reducing the risk of people contracting the virus; and (3) shortening the transmission chains, thereby reducing the opportunity for the virus to evolve increased virulence. Implementation of such vaccine passports will have significant implications across a wide range of legal and ethical issues affecting individuals and societies globally (see below).
Genetic introgression
When gene flow brings two previously isolated populations into contact so that they can hybridize and recombine their genomes, this may result in genetic introgression. Genetic introgression can release vast levels of evolutionary novelty, and it is thought to have been responsible for one of the largest known speciation events of vertebrates; the radiation of cichlid fishes in the Great African Lakes [18]. Introgression between diverged lineages has also led to the evolution of human-specialized pathogens[19]. The potential for hybridization between previously isolated species is at a record high (Figure 1), and pathogens are exceptionally well poised to exploit the new opportunities offered by genetic exchanges[20]. The genetic introgression that results from hybridization can produce mosaic-like genomes[21], and these novel combinations of genes occasionally enable the hybrids to infect new host species[22], or increase virulence[23]. According to the “mixing vessel” theory[24], some species of domesticated animals provide potential contact zones for multiple pathogens to exchange their genes (Figure 1). Pigs, for example, are susceptible to both avian and human influenza viruses, and these multiple infections can result in the evolution of novel hybrid strains such as H2N3[24]. Given the zoonotic origin of SARS-CoV-2, and the close contact between its most common host (humans) and domesticated animals, the vaccination of some domesticated species may be required in the future[25].
Selection
Natural selection molds the genetic variation introduced by mutation, recombination, and gene flow, resulting in adaptive evolution and increased fitness. Due to their rapid rate of replication and vast population size, pathogens are able to evolve rapidly, adaptively responding to changes in their environment. The discipline of evolutionary epidemiology studies how ecology, human behavior, and (medical) interventions shape the evolution of virulence in pathogens[26]. Although we are still unable to predict the exact trajectory of virulence evolution, we do know that adaptive evolutionary changes in pathogens increase their momentary fitness. This means that decisions made presently on how to deal with COVID-19 globally are likely to have long-term consequences for the virulence evolution of this pathogen. As vaccination programmes are being executed globally, we will have to assess and minimize the risks posed by SARS-CoV-2 reservoirs in parts of the human population that cannot, or do not wish to, be vaccinated. Furthermore, we need to be aware of other reservoir host species and their spillback potential, which could lead to the reemergence of (localized) COVID-19 outbreaks in the future. Hence, the continued monitoring of the disease, the global rollout of vaccines, vaccination of susceptible host species, and vaccine passports are measures that will need to be employed to successfully combat this disease.
The Red Queen in a globalized world
So, how did we get here? To better understand what has shifted the status quo between (human) hosts and our zoonotic pathogens, it is useful to examine the theory of coevolution, and in particular the Red Queen’s hypothesis. This hypothesis was originally proposed to explain the constant rate of extinction specific to a given group of species[2]. According to this hypothesis, the amount of resources controlled by a species determines its fitness, and hence, its population size. Furthermore, the amount of resources that one species gains results in an equivalent loss in its competing species, which could result in their extinction[3]. If we are to relate this evolutionary law to humans, we should acknowledge that our fitness gain has not been accomplished by evolving superior physical traits to control more resources, but rather, by adapting our environment. Many people have access to near limitless amounts of animal protein and carbohydrates produced in agriculture by our livestock and crops. However, the production of these resources has been driven by economic forces rather than evolutionary forces, and I believe this has tipped the coevolutionary balance in favor of our pathogens.
In the textbook example of coevolution, the frequencies of two antagonistic species fluctuate over time. Parasites tend to disproportionately infect the most common host genotype, whilst the rare genotypes escape becoming parasitized. The rare genotypes can thereby increase in frequency until one of them becomes the most common genotype. In turn, this eases the selection pressure against the once-common genotype, saving it from being pushed to extinction. In this sense, “a genotype is its own worst enemy, its fitness will decrease as it becomes more common”[27]. This process leads to Red Queen dynamics, and it results in a large amount of genetic variation that can be maintained over long periods of time[4]. This genetic variation is important because it allows recombination to generate novel genotypes. Recombination might be the most important evolutionary advantage of sexual reproduction [14,28], shuffling the genetic variation of the parents into new offspring genotypes, enabling slowly evolving hosts to coexist with much faster evolving parasites. Importantly, however, recombination can only generate novel variation when the parents possess genetic differences.
Adaptations in the host and parasite can also proceed in a “tit-for-tat” mode of coevolution, whereby both parties are under selection to “exceed” the antagonistic trait of the other species. This second type of coevolution is the other face of the Red Queen[4], and it is commonly known as the coevolutionary arms race. In this case, the antagonistic coevolution plays out between the virulence genes of the parasite and the immune genes (or resistance genes) of the host, and such coevolution is typified by a series of recurrent selective sweeps [4]. The host and parasite genotypes are rapidly replacing each other, and this process can drive one of the species to extinction[29]. In an arms race scenario, there are no second prizes; once the resistance of a gene or genotype has been broken, it can only be mend by evolving yet another resistant variant. The same rationale holds true for vaccines; once a pathogen like SARS-CoV-2 has evolved resistance to the vaccine, new vaccines need to be developed and applied to continue protecting the population. This need for variation is fundamental to both Red Queen processes. However, in the first process (Red Queen dynamics), both the host and parasite have genetic variation “in credit”, whereas during an arms race scenario, new variation (or vaccines) need to be constantly generated as the system is “in debit”.
Due to centuries of selective breeding, many of our livestock have become severely inbred; for example, the genetic variation present in 150 breeds of sheep and cattle is equivalent to a gene pool with a median effective population size of 52 individuals [30] (Figure 1c). This is 80 times lower than the minimum viable population size calculated for the 212 free-living species reviewed by Traill et al [31]. With little genetic variation, this livestock lacks the “coevolutionary fuel” to adaptively respond to the challenges posed by new pathogens, forcing coevolution into an arms race scenario. Consequently, our livestock is continually losing fitness with each novel pathogen it contracts. For our livestock – and perhaps also for us humans – the fitness ratchet clicks with every new EID that evolves and establishes itself. This is very pertinent to the original concept of the Red Queen hypothesis[2], which was originally formulated to understand the biotic conflicts over energy distribution among species. According to Van Valen: “ … an increase in momentary fitness by one species causes an equal total decline in momentary fitness among ecologically interacting species”[3]. Given the extraordinarily high biomass of our livestock (and us humans), the momentary fitness gains that parasites could accomplish by exploiting this plentiful resource are truly astronomical. Parasites and pathogens will continue to adapt to exploit these resources, and it is high time we recognize this evolutionary inevitability.
Pathogens have shaped societies over the past 13,000 years, playing a major role in wars, the colonization of continents, and the demise of civilizations[32]. True, infectious pathogens are not our only threat, but the risks and their impacts are closely interlinked with various other challenges [33,34]. In 2015, the UN set out 17 Sustainable Development Goals and 169 targets for 2030, which address economic, social and environmental dimensions of sustainable development[35]. Two-thirds of these targets may not be reached as a result of the COVID-19 pandemic[36]. In particular, Goal 3 aims to ensure healthy lives and promote well-being, and Target 3.3 aims to end or combat many infectious diseases. Similarly, the Global Health Security Agenda vision statement reads: “A world safe and secure from global health threats posed by infectious diseases, whether natural, deliberate, or accidental”[37]. These visions, goals, and targets are ambitious and transformational, but they do not recognize the danger posed by the evolution of novel zoonotic EIDs, and they do not prepare the public for the concessions we all will need to make as individuals to safeguard the future well-being of our species.
Toward a coevolutionary sustainable world
Control and reduce gene flow
To reduce the risks of future outbreaks of novel pathogens, we must control the gene flow of pathogens. Firstly, we can dramatically reduce the gene flow of human pathogens by altering some of our behaviors. Social distancing has already altered how we work and interact, which has markedly increased the uptake of online communication, such as Skype, Zoom, and FaceTime. Synchronous virtual communication platforms are giving us a unique advantage over other social animals that are threatened by EIDs. [38] COVID-19 is likely to accelerate the development of virtual communication, and such technologies will not only be key to reducing the impact of EIDs, but they may also be part of the solution to other environmental challenges. In addition, the introduction of “vaccine passports” could be instrumental in facilitating safe, free movement during the COVID-19 pandemic, enabling resumption of many social and economic activities. This will help to break transmission chains, which is not only important to prevent deaths now, but also halt the evolution of SARS-CoV-2 and to save lives in the future. There is significant political and public resistance against such passports, as some fear that these could be used discriminate against those refusing a COVID-19 vaccine, thereby applying pressure to raise vaccination rates. Some of the already most disadvantaged and discriminated against minorities have the lowest uptake rate of COVID-19 vaccine[39], putting these communities at increased risk, and potentially exacerbating inequalities. If we accept to need for a high level of vaccination to ameliorate potential inequalities arising from noncompliance, it may be necessary to consider compelling the uptake of vaccines. Sadly, this might be the price we have to pay for trailing in the Red Queen arms race. Similar to the momentary fitness loss of species at the receiving end of a coevolutionary arms race[3], we may have to accept the need for such measures until we have managed to reset the coevolutionary imbalance. Importantly, implementing these urgent changes requires us to carefully negotiate a wide range of legal and ethical issues that could inadvertently discriminate or exacerbate existing inequalities. How best to adjust to this new status quo is being considered in various reports and committees, including the United Nations, UNESCO, EQUINET (the European Network of Equality Bodies), ENNHRI (the European Network of National Human Rights Institutions), the SET-C (Science in Emergencies Tasking – COVID-19) and many others.
Secondly, we have to better understand our enemy. Brooks et al. spelled it out in an acronym, DAMA – Document, Assess, Monitor, Act[40]. Zoonotic EIDs pose the most significant threat to our society, but the pandemic potential differs markedly between pathogens[7]. We need to dedicate more resources to sequencing the virome of wildlife and our livestock, enabling us to better track gene flow, and assess spillover events and hybridization. Various programmes are already in place. For example, the Global Virome Project [41] conducts viral discovery and provide timely data for public health interventions against future pandemics. An estimated 1.67 million viral species have yet to be discovered from mammal and bird hosts, and the costs of sequencing viruses with the highest zoonotic potential may be great (~1.2 to 7 billion US dollars)[41], but they are dwarfed by the cost of another pandemic. The PREventing ZOonotic Disease Emergence (PREZODE) [42] is a particularly promising initiative that was announced recently at the One Planet Summit in January 2021. It aims to bring together the scientific and medical communities, promoting an internationally coordinated strategy to tackle emerging risks, focussing on prevention and sustainable solutions, as well as preparedness for the outbreak of EIDs.
Thirdly, we will need to better assess the EID risks posed by other host species. Hosts vary in their suitability as “mixing vessels and their ability to spread and transmit zoonotic pathogens to humans”[7]. Although we have known this for over a century[26], we have failed to act[39]. We now have the tools to identify hosts that pose the greatest zoonotic threat. For example, Sun et al [43]. examined whether SARS-CoV-2 could infect other species and found that one of the receptors in human ACE2 is identical to that of the receptor in macaques and chimpanzees. Such comparative genomics could help to assess the host-specific risks of spillovers and spillbacks, enabling us to identify potential reservoir host species.
Fourthly, to minimize highest-risk gene flow, we will need to bring a halt to the loss of natural habitats in biodiversity hotspots, thereby reducing human-wildlife conflict and spillover events[17]. These actions will complement the SDG targets, of which 30 (18%) are predicted to help reduce the risk of another global pandemic[36]. In this coevolutionary One Health approach, the protection of natural habitats and the environment is not just for the conservation of biodiversity, but also for the prevention of the next pandemic.
Restore genetic variation in livestock
In order to limit the reservoir of susceptible hosts, we will need to restore genetic diversity of farmed and domesticated animals (as well as crops), going beyond the goals and targets set out by the Strategic Plan for Biodiversity 2011–2020[44], and the UN 2030 Agenda for Sustainable Development [35]. Our livestock is at an overwhelming coevolutionary disadvantage, and it has become a “sitting duck” in the middle of a rapidly heating arms race with EIDs. However, the genetic variation that is lost in our livestock and crops is still largely present, in wild varieties and wild relatives, and dispersed among different breeds. Understandably, breeding programmes have been focussing on a few, highly productive breeds to maximize yield. Unfortunately, however, this has resulted in the extinction of 3.5% and 10% of domesticated breeds of birds and mammals, respectively[45]. In the first global assessment of biodiversity for food and agriculture, the FAO found that the genetic resources of many production systems are in serious decline[46]. The report also shows that biodiversity loss increases disease risk, but sadly, it fails to anticipate the extreme coevolutionary vulnerability of many production systems. Unless we act now, massive losses of crops and livestock are an evolutionary inevitability.
Interestingly, however, some wild vertebrate species such as the mangrove killifish (Kryptolebias marmoratus) appear to have evolved a genomic architecture that enables them to survive without novel, recombinant genetic variation. This fish can reproduce through self-fertilization, resulting in genetically near-uniform individuals. Remarkably, to offset its coevolutionary disadvantage, it has duplicated many of its MHC immune genes[47]. Emulating this genomic solution, transgenic, cisgenic and intragenic approaches, gene editing, site-specific mutagenesis (genome editing), and gene pyramiding are applied or considered for more durable resistance in crops [48–50]. However, without genetic variation between individuals, there can be no coevolutionary balance with pathogens. Frankel [51] already wrote this in his landmark paper: “Variation – the essence of life.”. We are now half a century later, and it is time we act.
Reduce consumption of animal protein
Mass food production has underpinned our “ecological success”, but it is entirely unsustainable based on environmental, ecological, and coevolutionary grounds. We urgently need to reduce our reliance on animal protein, in particular the consumption of other mammals. Emission of potent greenhouse gases by ruminant livestock is contributing significantly to anthropogenic “radiative forcing” and global warming[52]. The vast amount of antibiotics used to enhance growth and control infections has led to the emergence of new, more virulent, and more resistant microorganisms[53]. Given that mammals are phylogenetically closely related to humans, they also pose the most significant threat for the evolution and transmission of zoonotic EIDs. Producing animal protein from aquaculture and aquaponics has the potential to become an environmentally and coevolutionary more sustainable way of feeding part of the world’s population[46]. In addition, we will also need to develop a larger variety of palatable non-meat-based alternatives, which will have direct and indirect health benefits[54]. Reducing the number of livestock is imperative for human health, as well as for the health of the planet. To implement this and other changes, we must embrace a coevolutionary robust One Health approach and focus on what really matters: One World – One Health.
Concluding remarks
Emerging infectious diseases (EIDs) are possibly the greatest existential threat to humanity, but fortunately, we have never been in a stronger position to defend ourselves against these threats. COVID-19 has elicited a huge pan-global response among our medical and scientific communities, and as a species, we have shown remarkable behavioral plasticity to cope with the new conditions. Nevertheless, COVID-19 has revealed the tension between our rights as individuals and the well-being of society as a whole. However, tackling this pandemic requires us to act as an eusocial species rather than as selfish individuals. It calls for increased social responsibility, in particular in the uptake of the vaccine and adherence to rules designed to control the COVID-19 pandemic. Misinformation on the web and “playing politics” can significantly undermine the much-needed concerted response. Going forward, we need to recognize that our health, the environment, and our global economy are all closely interlinked, and that the world’s natural resources must be governed in a way that attains a coevolutionary balance. The “One World – One Health” concept was created in 2004, inspired by the “One Medicine” approach [55] that integrates human and veterinary medicine to respond to zoonoses[56]. By considering the health of humans, wildlife and ecosystems, the holistic One Health approach has recruited multisector expertise in dealing with humanity’s greatest challenge. New initiatives are emerging that envision innovative prevention strategies, such as PREZODE[42]. I advocate that these initiatives also address the coevolutionary imbalance, integrating this into the One Health framework. Importantly, all these changes require broad public support underpinned by an understanding of the significant threat our society is facing, and what we can do at an individual and societal level to improve our long-term (coevolutionary) prospects as a species. I believe that this is essential because with increased globalization, we have created a perfect storm for the evolution and outbreak of EIDs. Rather than just combating the existing EIDs and minimizing their social-economic impacts, we must also curtail the evolution of novel EIDs, thereby reducing the risks of future outbreaks, and the possible transition from the “Anthropocene” into a “Pathocene”.
Funding Statement
This work was supported by the Earth and Life Systems Alliance.
Data availability statement
Data sharing is not applicable to this article as no new data were generated in this study. Figure 1 is based on data published in, or associated with: Holmes (2009) (doi.org/10.1146/annurev.ecolsys.110308.120248), Traill et al. (2007) (doi.org/10.1016/j.biocon.2007.06.011), Bar-On et al. (2018) (doi.org/10.1073/pnas.1711842115), and Zhang et al. (2018) (doi.org/10.1038/s41598-018-30061-0).
References
- [1].Dobzhansky T. Nothing in biology makes sense except in the light of evolution. Am Biol Teacher. 1973;35(3):125–129. [Google Scholar]
- [2].Van Valen L. A new evolutionary law. Evol Theory. 1973;1:1–30. [Google Scholar]
- [3].Van Valen L. Molecular evolution as predicted by natural selection. J Mol Evol. 1974;3(2):89–101. [DOI] [PubMed] [Google Scholar]
- [4].Lighten J, Papadopulos AS, Mohammed RS, et al. Evolutionary genetics of immunological supertypes reveals two faces of the Red Queen. Nat Commun. 2017;8:1294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Crow JF, Kimura M. An introduction to population genetics theory. New York: Harper and Row; 1970. [Google Scholar]
- [6].Su S, Wong G, Shi W, et al. Epidemiology, genetic recombination, and pathogenesis of coronaviruses. Trends Microbiol. 2016;24(6):490–502. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Olival K, Hosseini P, Zambrana-Torrelio C, et al. Host and viral traits predict zoonotic spillover from mammals. Nature. 2017;546:646–650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Zhou H, Chen X, Hu T, et al. A novel bat coronavirus closely related to SARS-CoV-2 contains natural insertions at the S1/S2 cleavage site of the spike protein. Curr Biol. 2020;30(11):2196–2203.e3. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Domingo E, Perales C. Viral quasispecies. PLoS Genet. 2019;15(10):e1008271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Holmes EC. The evolutionary genetics of emerging viruses. Annu Rev Ecol Evol Syst. 2009;40:353–372. [Google Scholar]
- [11].Bar-On YM, Phillips R, Milo R. The biomass distribution on Earth. Proc Nat Acad Sci. 2018;115(25):6506–6511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Fisher RA. The genetical theory of natural selection. In: Variorum, Bennett JH, editors. Oxford: Oxford University Press; 1999. [Google Scholar]
- [13].Johnson CK, Hitchens PL, Pandit PS, et al. Global shifts in mammalian population trends reveal key predictors of virus spillover risk. Proc R Soc B. 2020;287(1924):20192736. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Felsenstein J. The evolutionary advantage of recombination. Genetics. 1974;78:737–756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Cromer D, Grimm AJ, Schlub TE, et al. Estimating the in-vivo HIV template switching and recombination rate. Aids. 2016;30(2):185–192. [DOI] [PubMed] [Google Scholar]
- [16].Wilkinson DA, Marshall JC, French NP, et al. Habitat fragmentation, biodiversity loss and the risk of novel infectious disease emergence. J R Soc Interface. 2018;15:20180403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Dobson AP, Pimm SL, Hannah L, et al. Ecology and economics for pandemic prevention. Science. 2020;369(6502):379–381. . [DOI] [PubMed] [Google Scholar]
- [18].Nichols P, Genner MJ, Van Oosterhout C, et al. Secondary contact seeds phenotypic novelty in cichlid fishes. Proc R Soc B. 2015;282(1798):20142272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Nader JL, Mathers TC, Ward BJ, et al. Evolutionary genomics of anthroponosis in Cryptosporidium. Nat Microbiol. 2019;4(5):826–836. . [DOI] [PubMed] [Google Scholar]
- [20].Jouet A, Saunders DG, McMullan M, et al. Albugo candida race diversity, ploidy and host‐associated microbes revealed using DNA sequence capture on diseased plants in the field. New Phytol. 2019;221(3):1529–1543. . [DOI] [PubMed] [Google Scholar]
- [21].Ward BJ, Van Oosterhout C. HYBRIDCHECK: software for the rapid detection, visualization and dating of recombinant regions in genome sequence data. Mol Ecol Resour. 2016;16(2):534–539. [DOI] [PubMed] [Google Scholar]
- [22].McMullan M, Gardiner A, Bailey K, et al. Evidence for suppression of immunity as a driver for genomic introgressions and host range expansion in races of Albugo candida, a generalist parasite. Elife. 2015;4:e04550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Schelkle B, Faria PJ, Johnson MB, et al. Mixed infections and hybridisation in monogenean parasites. PLoS One. 2012;7(7):e39506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Ma W, Kahn RE, Richt JA. The pig as a mixing vessel for influenza viruses: human and veterinary implications. J Mol Genetic Med. 2009;3(1):158–166. [PMC free article] [PubMed] [Google Scholar]
- [25].Van Oosterhout C, Hall N, Ly H, et al. COVID-19 evolution during the pandemic – implications of new SARS-CoV-2 variants on disease control and public health policies. Virulence. 2021;12(1):507–508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Restif O. Evolutionary epidemiology 20 years on: challenges and prospects. Infect Genet Evol. 2009;9(1):108–123. [DOI] [PubMed] [Google Scholar]
- [27].Lewontin RC. The genetic basis of evolutionary change. New York: Columbia University Press; 1974. [Google Scholar]
- [28].Lively CM. Host-parasite coevolution and sex. Bioscience. 1996;46(2):107–114. [Google Scholar]
- [29].Dawkins R, Krebs JR. Arms races between and within species. Proc R Soc Lond B. 1979;205:489–511. [DOI] [PubMed] [Google Scholar]
- [30].Zhang M, Peng WF, Hu XJ, et al. Global genomic diversity and conservation priorities for domestic animals are associated with the economies of their regions of origin. Sci Rep. 2018;8(1):1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Traill LW, Bradshaw CJ, Brook BW. Minimum viable population size: a meta-analysis of 30 years of published estimates. Biol Conserv. 2007;139(1–2):159–166. [Google Scholar]
- [32].Diamond J. Guns, germs and steel: a short history of everybody for the last 13,000 years. London: Random House; 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Wu T. The socioeconomic and environmental drivers of the COVID-19 pandemic: a review. Ambio. 2021;50:822–833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Di Marco M, Baker ML, Daszak P, et al. Opinion: sustainable development must account for pandemic risk. Proc Nat Acad Sci. 2020;117(8):3888–3892. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].United Nations . The sustainable development goals report 2016 [internet]. 2016. [cited 2020 September07]. Available from: http://www.un.org.lb/Library/Assets/The-Sustainable-Development-Goals-Report-2016-Global.pdf
- [36].Naidoo R, Fisher B. Reset sustainable development goals for a pandemic world. Nature. 2020;583:198–201. [DOI] [PubMed] [Google Scholar]
- [37].GHSA . Global health security agenda. [internet] 2019. [cited 2020 September07]. Available from: https://ghsagenda.org/
- [38].Townsend AK, Hawley DM, Stephenson JF, et al. Emerging infectious disease and the challenges of social distancing in human and non-human animals. Proc R Soc B. 2020;287(1932):20201039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Razai MS, Osama T, McKechnie DGJ, et al. Covid-19 vaccine hesitancy among ethnic minority groups. BMJ. 2021;372:n513. [DOI] [PubMed] [Google Scholar]
- [40].Brooks DR, Hoberg EP, Boeger WA. The Stockholm paradigm: climate change and emerging disease. Chicago: University of Chicago Press; 2019. [Google Scholar]
- [41].Carroll D, Daszak P, Wolfe ND, et al. The global virome project. Science. 2018;359(6378):872–874. . [DOI] [PubMed] [Google Scholar]
- [42].Peyre M, Vourc’h G, Lefrançois T, et al. PREZODE: preventing zoonotic disease emergence. Lancet. 2021;397(10276):792–793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Sun J, He WT, Wang L. COVID-19: epidemiology, evolution, and cross-disciplinary perspectives. Trends Mol Med. 2020;26(5):483–495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].The strategic plan for biodiversity 2011–2020 and the aichi biodiversity targets. Nagoya, Japan: Convention on Biological Diversity; 2010. October 29. Available from: https://go.nature.com/2Vuh0Ck [Google Scholar]
- [45].Brondizio ES, Settele J, Díaz S, et al.editors. IPBES: global assessment report on biodiversity and ecosystem services of the intergovernmental science-policy platform on biodiversity and ecosystem services [internet]. Bonn, Germany: IPBES Secretariat. 2019. cited 2020 September07. Available from: https://ipbes.net/global-assessment [Google Scholar]
- [46].Bélanger J, Pilling D editors. FAO: the state of the world’s biodiversity for food and agriculture [internet]. Rome: FAO Commission on Genetic Resources for Food and Agriculture Assessments. 2019. [cited 2020 September7]. Available from: http://www.fao.org/documents/card/en/c/ca3129en/ [Google Scholar]
- [47].Van Oosterhout C. Maintenance of major histocompatibility supertype variation in selfing vertebrate is no evidence for overdominant selection. Proc R Soc B. 2013;280(1754):20122501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Borrelli GM, Mazzucotelli E, Marone D, et al. Regulation and evolution of NLR genes: a close interconnection for plant immunity. Int J Mol Sci. 2018;19(6):1662. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Witek K, Lin X, Karki HS, et al. A complex resistance locus in Solanum americanum recognizes a conserved Phytophthora effector. BioRxiv. 2020. Available from. DOI: 10.1101/2020.05.15.095497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Witek K, Lin X, Karki HS, et al. A complex resistance locus in Solanum americanum recognizes a conserved Phytophthora effector. Nat Plants. 2021;7:198–208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Frankel OH. Variation - the essence of life. Proc Linnean Soc New South Wales. 1970;95:158–169. [Google Scholar]
- [52].Lassey KR. Livestock methane emission: from the individual grazing animal through national inventories to the global methane cycle. Agric Forest Meteorol. 2007;142:120–132. [Google Scholar]
- [53].Gilchrist MJ, Greko C, Wallinga DB, et al. The potential role of concentrated animal feeding operations in infectious disease epidemics and antibiotic resistance. Environ Health Perspect. 2007;115(2):313–316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Sabaté J. The contribution of vegetarian diets to health and disease: a paradigm shift? Am J Clin Nutr. 2003;78(3):502S–507S. [DOI] [PubMed] [Google Scholar]
- [55].Schwabe CW. Veterinary medicine and human health. 3rd ed. Baltimore, MD: The Williams and Wilkins Company; 1984. [Google Scholar]
- [56].Zinsstag J, Schelling E, Waltner-Toews D, et al. From “one medicine” to “one health” and systemic approaches to health and well-being. Prev Vet Med. 2011;101:148–156. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data sharing is not applicable to this article as no new data were generated in this study. Figure 1 is based on data published in, or associated with: Holmes (2009) (doi.org/10.1146/annurev.ecolsys.110308.120248), Traill et al. (2007) (doi.org/10.1016/j.biocon.2007.06.011), Bar-On et al. (2018) (doi.org/10.1073/pnas.1711842115), and Zhang et al. (2018) (doi.org/10.1038/s41598-018-30061-0).