

Abstract

Phycobiliproteins (PBPs) are a group of brilliant pigment proteins found in cyanobacteria and red algae; their synthesis and accumulation depend on several factors such as the type of strain employed, nutrient concentration, light intensity, light regimes, and others. This study evaluates the effect of macronutrients (citrate buffer, NaNO3, K2HPO4, MgSO4, CaCl2, Na2CO3, and EDTA) and the concentration of trace metals in BG-11 media on the accumulation of PBPs in a thermotolerant strain of Oscillatoria sp. The strain was grown in BG-11 media at 28 °C with a light:dark cycle of 12:12 h at 100 μmol m–2 s–1 for 15 days, and the effect of nutrients was evaluated using a Plackett–Burman Design followed by optimization using a response surface methodology. Results from the concentration of trace metals show that it can be reduced up to half-strength in its initial concentration without affecting both biomass and PBPs. Results from the Plackett–Burman Design revealed that only NaNO3, Na2CO3, and K2HPO4 show a significant increase in PBP production. Optimization employed a central Non-Factorial Response Surface Design with three levels and four factors (34) using NaNO3, Na2CO3, K2HPO4, and trace metals as variables, while the other components of BG-11 media (citrate buffer, MgSO4, CaCl2, and EDTA) were used in half of their initial concentration. Results from the optimization show that interaction between Na2CO3 and K2HPO4 highly increased PBPs’ concentration, with values of 15.21, 3.95, and 1.89 (% w/w), respectively. These results demonstrate that identifying and adjusting the concentration of critical nutrients can increase the concentration of PBPs up to two times for phycocyanin and allophycocyanin while four times for phycoerythrin. Finally, the reduction in non-key nutrients’ concentration will reduce the production costs of colorants at an industrial scale and increase the sustainability of the process.
1. Introduction
Cyanobacteria are a group of photosynthetic prokaryotes that originated about 3.5 billion years ago. It can be found in different aquatic environments such as lakes, rivers,1 and hot springs.2 Cyanobacteria are considered one of the new sustainable biotechnological sources of various raw materials for the global pharmaceutical, food, clinical, and energy industries; among these metabolites, phycobiliproteins (PBPs) stand out for their full demand,3 especially for pharmaceutical industries.
PBPs are a family of hydrophilic brilliant pigment proteins that play a crucial role in harvesting photonic energy from solar radiation3 in cyanobacteria, red algae, cryptomonads, and cyanelles.4 Among phycobiliproteins, three categories stand out (Figure 1): phycocyanin (C-PC), allophycocyanin (APC), and phycoerythrin (PE).5 Each of these categories possesses a specific spectrum referred to as blue (610–620 nm), blue-green (650–655 nm), and pink (540–570 nm), respectively.6
Figure 1.

Structure of chromophores found in phycocyanin (a), allophycocyanin (b), and phycoerythrin (c) of cyanobacteria. Extract from Oscillatoria sp. OSCI_UFPS001 (d).
Because of their nature, unique color, fluorescence, and antioxidant properties, PBPs are exploited in a wide range of applications such as dyes for the food industry (desserts, gums, jellies, and ice creams) and pharmaceuticals (eyeliners, lipsticks, and makeup) and even in the development of anticancer drugs.7−12
The production, accumulation, and purity of phycobiliproteins in cyanobacteria are strongly regulated by various factors such as the strain to be used, light intensity and its wavelength,13 temperature, pH, and media composition.14−16 Generally, algae and cyanobacteria have been isolated and produced on specific culture media composed of inorganic salts.17 Therefore, it is possible to modulate (or artificially adjust) the final concentration of PBPs.
Media composition plays an essential role in microalgal and cyanobacterial growth and byproduct (lipids, carotenes, etc.) synthesis, affecting the total biomass productivity.18 Generally, growth media such as BG-11,19 Bold’s Basal medium,20 Chu-10 medium,21 and Zarrouk medium22 were initially designed to produce as much biomass as possible from the strains and not for the overproduction of a specific metabolite.
Carbon and nitrogen are the most crucial nutrients for growth of microalgal and cyanobacterial biomass.23 In most algae, nitrogen is an important nutrient that regulates not only biomass production but also the accumulation of lipids and carotenoids.24 However, in cyanobacteria, nitrogen plays a key role in cell viability since it regulates the synthesis of PBPs, which in turn work as the main storage of nitrogen within the cell.25 Other nutrients such as calcium chloride, trace metal mix, and citric acid have proven to modulate the accumulation of PBPs;26 however, the effect on the synthesis and accumulation of PBPs may depend on the unique biology of the selected strain. Therefore, the tuning of nutrient concentration seems to be a critical step to enhance the production of specific metabolites such as PBPs. The conventional method used for improving the overproduction of a specific metabolite by one variable at a time26 is an expensive and time-consuming process.18 A response surface methodology (RSM) is a simple and precise method that allows the user to optimize operational conditions for the system evaluated without affecting production cost.27,28 However, limited studies using the RSM to enhance growth and PBP production have been done. Therefore, the current study investigates the effect of macronutrients (citrate buffer, NaNO3, K2HPO4, MgSO4, CaCl2, Na2CO3, and EDTA) and trace metal concentration from BG-11 media on the accumulation of phycobiliproteins in a thermotolerant strain of Oscillatoria sp. (OSCI_UFPS001), as it shows a high concentration of PBPs, especially C-PC.
2. Results
2.1. Selection of Relevant Media Components
Cyanobacteria constitute a group of photosynthetic microorganisms with morphological and physiological characteristics capable of responding to extreme changes by irradiation, nutrient limitation, salinity, and pH. Therefore, they are considered biotechnological sources to produce phycobiliproteins, carotenoids, and proteins of economic interest.16
According to the Pareto charts (Figure 22a–d), NaNO3, Na2CO3, and K2HPO4 affect the production of biomass and PBPs (C-PC, APC, and PE). On the other hand, only the biomass was affected by other variables such as CaCl2, EDTA, and MgSO4. The only factor that affects neither biomass nor PBPs was citrate buffer.
Figure 2.

Pareto charts for the effects of different nutrients on biomass (a), C-PC (b), APC (c), and PE (d) in Oscillatoria sp.
2.2. Selection of Trace Metal Concentration
According to the results, by reducing the concentration of micronutrients by 50%, the biomass (Figure 3a) remained relatively constant (0.6–0.58 g/L); however, using 25% less of concentration, the biomass was considerably reduced up to 0.48 g/L. In contrast to biomass, PBP concentration and purity were not affected by reducing the concentration of micronutrients (Figure 3b,c).
Figure 3.

Production of biomass (a) and PBPs (b) and its purity (c) under the reduced concentration of trace metals from BG-11 media.
2.3. Optimization by the Response Surface Methodology
Based on the results from the previous stage, four factors (NaNO3, Na2CO3, K2HPO4, and trace metals) were used in the next step of the process for the optimization of PBP production. The other components of BG-11 media (citrate buffer, MgSO4, CaCl2, and EDTA) were used in half concentration. Evaluation was done using a Non-Factorial Response Surface Design with four factors, three levels, and two central points. The resolved design can be found in Table 1.
Table 1. Resolved Design of Four Factors and Three Levels with Two Center Points.
| experiment | NaNO3 (g/L) | Na2CO3 (mL/L) | K2HPO4 (mL/L) | trace metals (mL/L) |
|---|---|---|---|---|
| 8 | 1.0 | 6 | 0.7 | 0.3 |
| 2 | 0.4 | 2 | 0.7 | 0.3 |
| 26 | 0.7 | 4 | 0.23 | 0.9 |
| 4 | 0.4 | 6 | 0.7 | 0.7 |
| 9 (C) | 0.7 | 4 | 0.23 | 0.5 |
| 23 | 0.7 | 4 | 0 | 0.5 |
| 21 | 0.7 | 0 | 0.23 | 0.5 |
| 11 | 0.4 | 2 | 0.7 | 0.7 |
| 20 | 1.3 | 4 | 0.23 | 0.5 |
| 10 | 0.4 | 2 | 0.08 | 0.3 |
| 13 | 0.4 | 6 | 0.7 | 0.3 |
| 18 (C) | 0.7 | 4 | 0.23 | 0.5 |
| 24 | 0.7 | 4 | 0.85 | 0.5 |
| 6 | 1 | 2 | 0.7 | 0.7 |
| 12 | 0.4 | 6 | 0.08 | 0.7 |
| 1 | 0.4 | 2 | 0.08 | 0.7 |
| 7 | 1 | 6 | 0.08 | 0.7 |
| 22 | 0.7 | 8 | 0.23 | 0.5 |
| 17 | 1 | 6 | 0.7 | 0.7 |
| 3 | 0.4 | 6 | 0.08 | 0.3 |
| 25 | 0.7 | 4 | 0.23 | 0.1 |
| 15 | 1 | 2 | 0.7 | 0.3 |
| 14 | 1 | 2 | 0.08 | 0.7 |
| 19 | 0.1 | 4 | 0.23 | 0.5 |
| 5 | 1 | 2 | 0.08 | 0.3 |
| 16 | 1 | 6 | 0.08 | 0.3 |
The experimental data concerning the effect of concentration of NaNO3, Na2CO3, K2HPO4, and trace metals on the production of C-PC, APC, and PE were fitted on two models: linear (L) and quadratic (Q). ANOVA illustrates that not all the factors studied affect the synthesis of PBPs. Table 2 summarizes the most significant factors (p = 0.05). In the case of C-CP, trace metals, NaNO3, and K2HPO4 affect its synthesis. APC is only affected by trace metals and Na2CO3. Finally, PE is affected by trace metals, Na2CO3, and NaNO3. Similarly, it was found that the interaction between the four variables (NaNO3/Na2CO3, NaNO3/K2HPO4, NaNO3/trace metals, Na2CO3/K2HPO4, Na2CO3/trace metals, and K2HPO4/trace metals) affects the production of PBPs; however, the interaction Na2CO3/K2HPO4 substantially increases the final concentration of C-PC, APC, and PE.
Table 2. Analysis of Variance of Quadratic Model for C-PC, APC, and PE Production.
| sum of squares | df | mean square | F value | p-value | ||
|---|---|---|---|---|---|---|
| C-PC (% w/w) R2 = 0.963 | ||||||
| (A) NaNO3 (g/L) | 14.8568 | 1 | 14.8568 | 8.58584 | 0.004262 | significant |
| (B) Na2CO3 (mL/L) | 0.0187 | 1 | 0.0187 | 0.01078 | 0.917518 | |
| (C) K2HPO4 (mL/L) | 10.4787 | 1 | 10.4787 | 6.05572 | 0.015707 | significant |
| (D) micronutrients (mL/L) | 106.0142 | 1 | 106.0142 | 61.26643 | <0.000001 | significant |
| BC | 28.2680 | 1 | 28.2680 | 16.33630 | 0.000109 | significant |
| APC (% w/w) R2 = 0.964 | ||||||
| (A) NaNO3 (g/L) | 0.20976 | 1 | 0.209755 | 0.96771 | 0.327804 | |
| (B) Na2CO3 (mL/L) | 3.48559 | 1 | 3.485587 | 16.08083 | 0.000122 | significant |
| (C) K2HPO4 (mL/L) | 0.03979 | 1 | 0.039794 | 0.18359 | 0.669296 | |
| (D) micronutrients (mL/L) | 6.29052 | 1 | 6.290518 | 29.02143 | 0.000001 | significant |
| BC | 2.49803 | 1 | 2.498033 | 11.52472 | 0.001011 | significant |
| PE (% w/w) R2 = 0.926 | ||||||
| (A) NaNO3 (g/L) | 0.143036 | 1 | 0.143036 | 4.81829 | 0.030649 | significant |
| (B) Na2CO3 (mL/L) | 0.174440 | 1 | 0.174440 | 5.87615 | 0.017281 | significant |
| (C) K2HPO4 (mL/L) | 0.087924 | 1 | 0.087924 | 2.96178 | 0.088580 | significant |
| (D) micronutrients (mL/L) | 1.448332 | 1 | 1.448332 | 48.78810 | <0.000001 | significant |
| BC | 0.185073 | 1 | 0.185073 | 6.23430 | 0.014289 | significant |
Figure 4a–c presents the response surface plots for the effect of Na2CO3/K2HPO4 on the accumulation of C-PC, APC, and PE. According to the results, relatively larger concentrations of Na2CO3 (>4 mL/L) and normal concentrations of K2HPO4 (1 mL/L) substantially increase the accumulation of the three phycobiliproteins studied.
Figure 4.

Surface response and Pareto charts for C-PC (a), APC (b), and PE (c).
Table 3 presents the values for X (Na2CO3 in mL/L) and Y (K2HPO4 in mL/L) used for the validation of the proposed concentrations. In this scenario, NaNO3 and trace metals were used at 1.5 g/L and 0.5 mL/L, respectively, while the other components of BG-11 media (citrate buffer, MgSO4, CaCl2, and EDTA) were used in half concentration.
Table 3. Variables for Optimal PBP Concentration on Oscillatoria sp.
| label | variable | value |
|---|---|---|
| X | Na2CO3 (mL/L) | 6 |
| Y | K2HPO4 (mL/L) | 1 |
| ZC-CP | concentration (% w/w) | 14.3 |
| ZACP | concentration (% w/w) | 3.5 |
| ZPE | concentration (% w/w) | 1.5 |
The contents of PBPs of Oscillatoria sp. grown on BG-11 media without any modification were 7.02, 2.23, and 0.45 (% w/w), respectively (Figure 5a). However, under the optimized conditions, the concentration of PBPs was significantly higher than the control, with values of 15.21, 3.95, and 1.89 (% w/w), respectively, which is 2.12-, 1.77-, and 4.17-fold higher for the three PBPs studied. Another critical result to highlight is the increase in the purity index of PBPs (Figure 5b). In the case of C-PC, purity increased from 7.0 to 8.7; in APC, it increased from 0.28 to 0.38, and the purity of PE increased from 0.45 to 0.6. It should be noted that in this work, any of the extracts were not subjected to any purification process.
Figure 5.
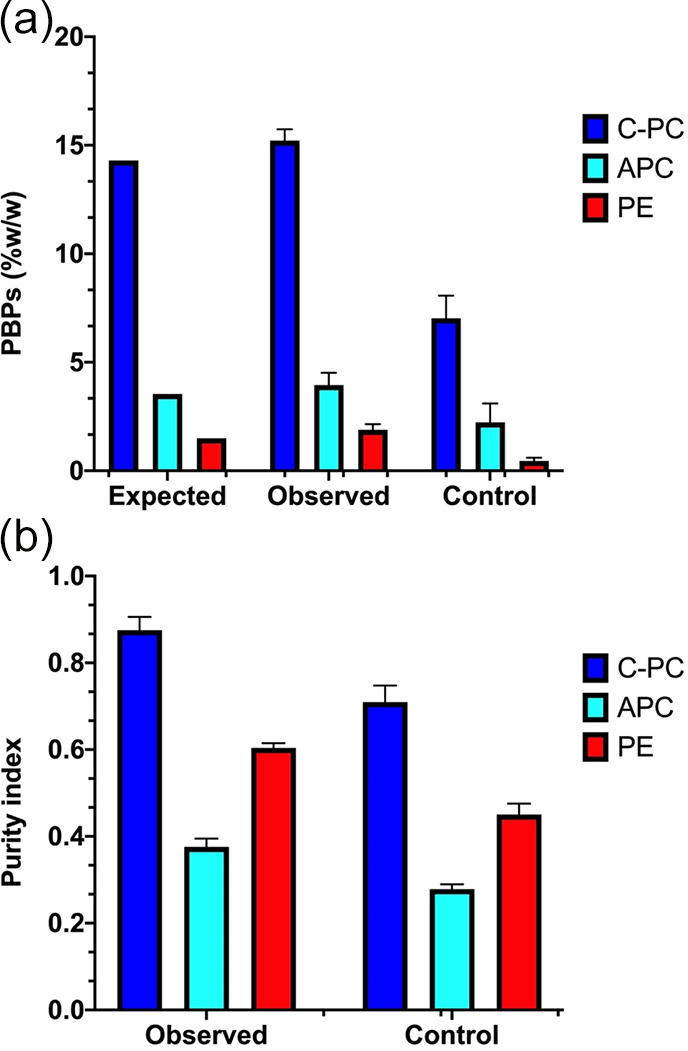
Expected vs observed concentration of PBPs (a) and observed purity of PBPs (b) after optimization of culture media.
3. Discussion
Oscillatoria sp. is a genus of cyanobacteria that presents an unbranched filamentous growth with mucilaginous sheaths (Figure 6). It can be identified by its characteristic movement of oscillating trichomes, which are composed of short cells (usually several times shorter than its width).29 In recent years, different strains of Oscillatoria sp. have been studied for their capacity on PBP production. Soni et al.30 isolated a strain of Oscillatoria quadripunctulata with a purity index of 0.85 in the crude extract. More recently, Chittapun et al.31 proved the capacity of a strain of Oscillatoria okeni to obtain high-purity extracts (0.68–1.65) using diferent extraction solutions. Other studies focused on the inhibitory capacity of C-PC from Oscillatoria sp. over pro-inflammatory enzymes such as lipoxygenase32 and their antioxidant capacity and antiproliferative activity against human cancer cells through apoptosis.33,34 Therefore, this study could allow exploring the potential of developed processes at an industrial scale to produce nutraceutical and pharmaceutical compounds for human health.
Figure 6.

Oscillatoria sp. OSCI_UFPS_001, filamentous cyanobacteria isolated from a thermal spring in the city of Cúcuta (Colombia).
3.1. Relevant Media Components
The adjustment in the concentration of the nutrients of the culture medium (especially N, P, K, and C) is a critical step to improve the production of specific metabolites. The carbon source is a critical element since the cyanobacterial biomass can be up to 50% (w/w) of carbon.35 Therefore, the correct carbon source added into the media, plus an optimal concentration of carbon, allows the accumulation of metabolites of interest.36 Sodium carbonate is the preferred carbon source on different culture media for cyanobacterial production, not only because it is used for pH adjustment, but it also assists in the maintenance and adaption of the microorganism to the autotrophic culture condition.37 In their work Johnson et al.(37) found that a reduction in inorganic carbon (0.008 g/L Na2CO3) promotes the synthesis of accessory pigments in a strain of Nostoc sp. This statement contrasts with the results obtained in this work since 0.16 g/L Na2CO3 (20 times more) was needed to improve PBP production. A possible explanation for this phenomenon is the origin of the two strains, while Johnson et al.37 employed a strain from a lake, the strain used in this study was originally isolated from a hot spring with high carbonate contents (data not shown).
Nitrogen is the second most required nutrient for the biomass and metabolite production, just after carbon,4 and the second most abundant nutrient on algae and cyanobacteria.35 It plays a critical role in the overproduction of PBPs since nitrogen is stored within PBPs. Therefore, under nitrogen shortage, cells selectively degrade their PBP storage.38,39 Typically, cyanobacteria and algae prefer NH4+ rather than NO3– sources; however, due to its high toxicity, nitrate is preferred over ammonia for PBP production.40 Several studies concerning the optimization of PBP production using different nitrogen sources on a wide range of cyanobacteria (including nitrogen-fixing strains) report high concentrations of NaNO3 in their culture media (0.75–4.3 g/L) (Table 4). In the present work, 1.5 g/L NaNO3 increases the production of PBPs. Phosphorous is one of the most important macronutrients for cyanobacterial growth, and its content varies from 0.05% up to 3.3% (w/w).41 P is used for the formation of several organic molecules, such as RNA and DNA, ATP, and membrane phospholipids.42 Unlike carbon, nitrogen and phosphorous are expensive nutrients derived from fossil phosphate rocks.36 On the production of PBPs, the concentration of K2HPO4 varies from species, from 0.04 to 0.75 g/L. Most species evaluated for the overproduction of PBPs use the concentration from BG-11 (0.04 g/L). Other components such as glycine (carbon source and precursor of δ-aminolevulinate) and sodium glutamate (nitrogen source) have improved the maximum yield of cell biomass and PBP production on Oscillatoria sp. BTA170.43
Table 4. Comparison of NaNO3, K2HPO4, and Carbon Source Concentrations Used to Increase PBP Concentration.
| strain | NaNO3 (g/L) | carbon source (g/L) | K2HPO4 (g/L) | biomass (g/L) | PBPs | reference | |
|---|---|---|---|---|---|---|---|
| Anabaena sp. PCC 6803 | 1.5 | Na2CO3 | 0.02 | 0.04 | C-PC: 10% (w/w) | (73) | |
| A. variabilis CCC421 | 0 | Na2CO3 | 0.02 | 0.2 | C-PC: 408.5 (mg/L) | (74) | |
| A. platensis EGEMACC 30 | 2.5 | NaHCO3 | 16.8 | 0.04 | 1.56 | PBPs: 22.3% (w/w) | (66) |
| Euhalothece sp. | 1.67 | Na2CO3 | 0.02 | 0.04 | C-PC: 0.045 (g/g) | (72) | |
| Nostoc sp. | 1.5 | Na2CO3 | 0.008 | 0.04 | 0.625 | C-PC: 0.13 (g/g) | (37) |
| Phormidium sp. EGEMACC 72 | 1.5 | Na2CO3 | 0.02 | 0.04 | 1.18 | PBPs: 18.53% (w/w) | (66) |
| Phormidium ceylanicum | 4.5 | Na2CO3 | 0.02 | 0.04 | 0.97 | C-PC: 0.73 (g/L) | (26) |
| Phormidium rubidum A09DM | 0.75 | Na2CO3 | 0.02 | 0.75 | (75) | ||
| Pseudoscillatoria sp. EGEMACC 74 | 1.5 | 0.04 | 0.87 | PBPs: 18.9% (w/w) | (66) | ||
| Synechocystis sp. PCC 7120 | 1.5 | Na2CO3 | 0.02 | 0.04 | C-PC: 6.5% (w/w) | (74) | |
| Spirulina maxima | 2.5 | CO2 | 0.03% v/v | 0.5 | 3.75 | C-PC: 2.51% (w/w) | (76) |
| APC: 4.11% (w/w) | |||||||
| 2.5 | NaHCO3 | 16.8 | 0.5 | C-PC: 0.08 (g/L) | (77) | ||
| APC: 0.06 (g/L) | |||||||
| PE: 0.02 (g/L) | |||||||
| 2.5 | NaHCO3 | 16.8 | 0.5 | 4.82 | C-PC: 0.53 (g/L) | (78) | |
| Trichromus sp. IMU26 | 0 | Na2CO3 | 0.02 | 0 | CPC: 4% (w/w) | (79) | |
| APC: 3% (w/w) | |||||||
| PE: 5% (w/w) | |||||||
| Oscillatoria sp. OSCI_UFPS001 | 1.5 | Na2CO3 | 0.16 | 0.04 | 0.59 | CPC: 15.2% (w/w) | this study |
| APC: 3.9% (w/w) | |||||||
| PE: 1.8% (w/w) | |||||||
3.2. Trace Metal Concentration
Trace elements or micronutrients greatly influence the growth and production of metabolites in different species of microalgae and cyanobacteria. These are required in minimal amounts from micro-, nano-, or picograms per liter. The most important are iron, cobalt, zinc, nickel, manganese, copper, boron, vanadium, and molybdenum;44 in general, these elements are supplied as a mix of nutrients. According to the literature, different investigations have been carried out to find out the effect of some micronutrients on the growth and content of pigments in some species of cyanobacteria; for example, the importance of four microelements (Fe III, Cu II, Zn II, and Mo VI) was investigated in Arthrospira platensis, where it was concluded that they greatly influence pigment accumulation and biomass production.45 Isao et al.(46) found that iron limitation caused a reduction in cellular chlorophyll and carotenoid concentration. On the other hand, Rueter and Petersen47 reported that high Fe contents promote growth of cyanobacteria in lakes.
3.3. Optimization by the Response Surface Methodology
The traditional method for optimizing medium involves one factor at a time in which each component is changed one by one, keeping other factors constant. This traditional approach is very tedious and time-consuming. Moreover, a biological system is a time-consuming process that is affected by several factors simultaneously.47
The RSM is an interesting approach for the optimization of biomass and specific metabolites. Over the years, this method has been proven to be an interesting statistical tool for better culture media in strains such as Ankistrodesmus falcatus KJ671624,48Auxenochlorella protothecoides,49Botryococcus braunii,24B. braunii LB572,50Chlorella sp.,51Chlorellaprotothecoides,52Chlorella sorokiniana Mb-1,53Chlorella vulgaris,54C. vulgaris 1803,55Chlorococcum oleofaciens,17Desmodesmus armtus,56Dunaliella salina,57C. pyrenoidosa,58Scenedesmus sp. ASK22,59Scenedesmus obliquus,60,61 and Spirulina sp.62 In PBP research, the application of RSM has been focused on their extraction and purification.53−71 Until now, there are few reports such as the works of Singh et al.26 and Mogany et al.(72) that have focused on the effect of multiple nutrients on biomass production and PBP content.
Typically, N and C are the most common factors evaluated because in photosynthetic organisms, up to 50% of the fixed carbon (in the form of CO2) is destined for nitrogen fixation24 while high concentrations of nitrogen promote the synthesis of PBPs; however, factors such as phosphate, sodium, magnesium, and trace metals are less obvious choices. Kumar et al.(73) reported that K2HPO4 (0.2 g/L) and trace metals (0.5 mL/L) enhance PBP production in Anabaena variabilis CCC421. Singh et al.26 evaluated the interaction of trace metals with NaNO3 and citric acid and found that the optimal concentration is between 0.9 and 1 mL/L. On the other hand, Mogany et al.(72) found that the interaction between trace metals, MgSO4, and NaNO3 positively affects the production of PBPs, with concentrations of 10 mL/L. This extreme difference (10 times higher than BG-11) can be due to the unique strain used in the research.
4. Conclusions
The results of this study indicate that the Plackett–Burman Design coupled with the RSM is a robust methodology for identifying critical nutrients for PBP overproduction in Oscillatoria sp. Culture media are designed for cyanobacteria to grow without any possible starvation; therefore, in order to increase the production of specific metabolites, most salts are not required in larger quantities. The development of an optimized concentration of the nutrients from BG-11, especially NaNO3, Na2CO3, K2HPO4, and trace metals (with half-concentration of citrate buffer, MgSO4, CaCl2, and EDTA), resulted in an increase of 2.12-, 1.77-, and 4.17-fold for C-PC, APC, and PE. These results allow reducing the nutrient loss on PBPs’ industrial-scale production. Future studies should be directed at providing further insights into the possible interaction of nutrient availability and abiotic factors such as light regimes (light:dark cycle, intensity, and light quality). Finally, the optimized conditions can allow the industrial-scale production of nutraceutical and pharmaceutical compounds for human health.
5. Materials and Methods
5.1. Strain
Oscillatoria sp. OSCI_UFPS001 was isolated from a thermal spring in Cúcuta (Colombia) and kept at INNOValgae collection (UFPS, Colombia). The strain was grown in a tubular glass flask with a culture volume of 2 L containing BG-11 media.19 The strain was mixed through the injection of filtered air with 1% (v/v) CO2 at a flow rate of 0.78 L min–1 and a light:dark cycle of 12:12 h at 100 μmol m–2 s–1 for 15 days.
5.2. Selection of Relevant Media Components
The effects of seven nutrient stock (citrate buffer, NaNO3, K2HPO4, MgSO4, CaCl2, Na2CO3, and EDTA) from BG-11 media were determined using a Plackett–Burman design with two center points and three replicates (30 runs)80 (Table 5). The concentrations of biomass (expressed as g/L) and C-PC, APC, and PE (expressed as % w/w) were selected as the response variable and subjected to analysis of variance (ANOVA). Pareto charts were employed for the selection of influencing variables.
Table 5. Plackett–Burman Design for the Seven Nutrient Stock from BG-11 Media.
| level | citrate buffer (mL/L) | NaNO3 (g/L) | K2HPO4 (g/L) | MgSO4 (g/L) | CaCl2 (g/L) | Na2CO3 (g/L) | MgNa2EDTA (g/L) |
|---|---|---|---|---|---|---|---|
| low (−) | 0.5 | 0.5 | 0.02 | 0.0375 | 0.018 | 0.01 | 0.0005 |
| high (+) | 1.0 | 1.0 | 0.04 | 0.075 | 0.036 | 0.02 | 0.001 |
5.3. Selection of Trace Metal Concentration
To prove the possible effect of the micronutrient concentration on biomass and PBPs’ final production, several experiments were carried out using reduced quantities of micronutrient mix of BG-11 media (75%, 50%, 25%, and 12.5%).
5.4. Culture Conditions
Each experiment was done in 500 mL flasks with a working volume of 250 mL of culture media. Each flask was mixed through the injection of filtered air with 1% (v/v) CO2 at a flow rate of 0.78 L min–1 and a light:dark cycle of 12:12 h at 100 μmol m–2 s–1 for 15 days.
5.5. Biomass and PBP Quantification
After 15 days, 50 mL samples were removed from each flask (by triplicate) and centrifuged at 3400 rpm for 20 min and the supernatant was withdrawn. The pellet was resuspended in 20 mL of distilled water, filtered on precombusted CF/C glass fiber filters, and dried overnight at 60 °C in an oven containing a bed of silica gel. The dried filters were stored in a desiccator until constant weight. The mass of cyanobacteria was recorded using a digital balance. The filtered biomass was suspended in 10 mL of cold phosphate buffer solution (0.05 M, pH 6.8) and approximately 1 g of glass beads (0.5 mm diameter), and the solution was vortexed at maximum speed for 10 min. The mixture was stored in a refrigerator to promote the solubilization of the phycobiliproteins (4 °C, 24 h). PBPs were separated from cell debris by centrifugation (3400 rpm, 30 min, 20 °C). The supernatant (deep blue) was collected and measured using a spectrophotometer at different wavelengths, i.e., 620, 652, 562, and 280 nm. The concentration of C-PC, APC, and PE was calculated using eqs 1–3 described by Bennett and Bogorad:81
| 1 |
| 2 |
| 3 |
The purities of C-PC, APC, and PE were determined using eqs 4–6 proposed by Patil et al.82 and Antelo et al.:7
| 4 |
| 5 |
| 6 |
5.6. Optimization by the Response Surface Methodology
The optimization of biomass and PBPs production was further refined by using the central Non-Factorial Response Surface Design with two central points on software STATISTICA 7.0 (Statsoft).
Acknowledgments
We would like to express our sincere gratitude to Newton Fund Institutional Links, Universidad de Santander UDES, and Universidad Francisco de Paula Santander for providing the equipment for this research and the Colombian Ministry of Science Technology and Innovation MINCIENCIAS for the support to national Ph.D. Doctorates through the Francisco José de Caldas scholarship program and young researchers “Jovenes Investigadores e Innovadores” scholarship program.
S.O.M.-L. received funding from the “Jovenes Investigadores e Innovadores” scholarship program no. 770-2016. J.B.G.-M. received funding from UFPS Internal Research funding: FINU 27-2019. A.F.B.-S. received funding from the Gen Foundation with the project “Isolation of Thermo-tolerant Algae as a Novel Source of Food Colorants” and Newton Fund Institutional Links (ID 527624805).
The authors declare no competing financial interest.
References
- Kannaujiya V. K.; Kumar D.; Pathak R. J.; Sonker A. S.; Rajneesh.; Singh V.; Sundaram S., Sinha R. P.. Recent advances in production and the biotechnological significance of phycobiliproteins. In New Approaches in Biological Research ;Sinha R.P., Richa., Eds.; Nova Science Publisher: New York, 2017, pp. 1–34. [Google Scholar]
- Santiago-Santos M. C.; Ponce-Noyola T.; Olvera-Ramírez R.; Ortega-López J.; Cañizares-Villanueva R. O. Extraction and Purification of Phycocyanin from Calothrix sp. Process Biochem. 2004, 39, 2047–2052. 10.1016/j.procbio.2003.10.007. [DOI] [Google Scholar]
- Kannaujiya V. K.; Kumar D.; Pathak J.; Sinha R. P.. Phycobiliproteins and Their Commercial Significance. In: Cyanobacteria, From Basic Science to Application ;Mishra A.K., Tiwari D.N., Rai A.N. Eds.; Elsevier Inc: London, 2019, pp. 207–2016 [Google Scholar]
- Pagels F.; Guedes A. C.; Amaro H. M.; Kijjoa A.; Vasconcelos V. Phycobiliproteins from Cyanobacteria: Chemistry and Biotechnological Applications. Biotechnol. Adv. 2019, 37, 422–443. 10.1016/j.biotechadv.2019.02.010. [DOI] [PubMed] [Google Scholar]
- Dasgupta C. N.Algae as a source of phycocyanin and other industrially important pigments. In Algal biorefinery: An integrated approach ;Das D., Ed; Springer International Publishing: New Delhi, 2015. pp. 253–276. [Google Scholar]
- İlter I.; Akyıl S.; Demirel Z.; Koç M.; Conk-Dalay M.; Kaymak-Ertekin F. Optimization of Phycocyanin Extraction from Spirulina platensis Using Different Techniques. J. Food Compos. Anal. 2018, 70, 78–88. 10.1016/j.jfca.2018.04.007. [DOI] [Google Scholar]
- Antelo F. S.; Anschau A.; Costa J. A. V.; Kalil S. J. Extraction and purification of C-phycocyanin from Spirulina platensis in conventional and integrated aqueous two-phase systems. J. Braz. Chem. Soc. 2010, 21, 921–926. 10.1590/S0103-50532010000500022. [DOI] [Google Scholar]
- Fernández-Rojas B.; Hernández-Juárez J.; Pedraza-Chaverri J. Nutraceutical Properties of Phycocyanin. J. Funct. Foods. 2014, 11, 375–392. 10.1016/j.jff.2014.10.011. [DOI] [Google Scholar]
- Manirafasha E.; Ndikubwimana T.; Zeng X.; Lu Y.; Jing K. Phycobiliprotein: Potential Microalgae Derived Pharmaceutical and Biological Reagent. Biochem. Eng. J. 2016, 109, 282–296. 10.1016/j.bej.2016.01.025. [DOI] [Google Scholar]
- Martelli G.; Folli C.; Visai L.; Daglia M.; Ferrari D. Thermal Stability Improvement of Blue Colorant C-Phycocyanin from Spirulina platensis for Food Industry Applications. Process Biochem. 2014, 49, 154–159. 10.1016/j.procbio.2013.10.008. [DOI] [Google Scholar]
- Kumar J.; Singh D.; Tyagi M. B.; Kumar A.. Cyanobacteria: Applications in Biotechnology. In Cyanobacteria, From Basic Science to Application ;Mishra A. K., Tiwari D. N., Rai A. N. Eds.; Elsevier Inc: London, 2019, pp. 327–346. [Google Scholar]
- Wu H.-L.; Wang G.-H.; Xiang W.-Z.; Li T.; He H. Stability and antioxidant activity of food grade phycocyanin isolated from Spirulina platensis. Int. J. Food Prop. 2016, 19, 2349–2362. 10.1080/10942912.2015.1038564. [DOI] [Google Scholar]
- Babu T. S.; Kumar A.; Varma A. K. Effect of Light Quality on Phycobilisome Components of the Cyanobacterium Spirulina platensis. Plant Physiol. 1991, 95, 492–497. 10.1104/pp.95.2.492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee N. K.; Oh H.-M.; Kim H.-S.; Ahn C.-Y. Higher Production of C-Phycocyanin by Nitrogen-Free (Diazotrophic) Cultivation of Nostoc sp. NK and Simplified Extraction by Dark-Cold Shock. Bioresour. Technol. 2017, 227, 164–170. 10.1016/j.biortech.2016.12.053. [DOI] [PubMed] [Google Scholar]
- Boussiba S.; Richmond A. E. Isolation and characterization of phycocyanins from the blue-green alga Spirulina platensis. Arch. Microbiol. 1979, 120, 155–159. 10.1007/BF00409102. [DOI] [Google Scholar]
- Sarada R.; Pillai M. G.; Ravishankar G. A. Phycocyanin from Spirulina Sp.: Influence of Processing of Biomass on Phycocyanin Yield, Analysis of Efficacy of Extraction Methods and Stability Studies on Phycocyanin. Process Biochem. 1999, 34, 795–801. 10.1016/S0032-9592(98)00153-8. [DOI] [Google Scholar]
- Pauline J. M. N.; Achary A. Novel media for lipid production of Chlorococcum oleofaciens: A RSM approach. Acta Protozool. 2019, 58, 31–41. 10.4467/16890027AP.19.003.10834. [DOI] [Google Scholar]
- Bunkaew P.; Kongruang S. Statistical Approach of Nutrient Optimization for Microalgae Cultivation. E3S Web Conf. 2020, 141, 03009 10.1051/e3sconf/202014103009. [DOI] [Google Scholar]
- Allen M. M.; Stanier R. Y. Growth and division of some unicellular blue-green algae. J. Gen. Microbiol. 1968, 199–202. 10.1099/00221287-51-2-199. [DOI] [PubMed] [Google Scholar]
- Bischoff H. W.; Bold H. C.. Phycological Studies IV. Some Soil Algae From Enchanted Rock and Related Algal Species; University of Texas: Austin, 1963, 6318:1–95. [Google Scholar]
- Stein J.Handbook of Phycological methods. Culture methods and growth measurements; Cambridge University Press, Cambridge, 1973. 448 pp. [Google Scholar]
- Zarrouk C.Contribution a l’étude du cyanophycée. Influence de divers facteurs physiques et chimiques sur la croissance et la photosynthèse de Spirulina maxima (setch et gardner) geitl.Thesis Faculte des Sciences, Universite de Paris: paris: 1966. [Google Scholar]
- Quintero-Dallos V.; García-Martínez J. B.; Contreras-Ropero J. E.; Barajas-Solano A. F.; Barajas-Ferrerira C.; Lavecchia R.; Zuorro A. Vinasse as a Sustainable Medium for the Production of Chlorella vulgaris UTEX 1803. Water. 2019, 11, 1526. 10.3390/w11081526. [DOI] [Google Scholar]
- Barajas-Solano A. F.; Guzmán-Monsalve A.; Kafarov V. Effect of Carbon-Nitrogen Ratio for the Biomass Production, Hydrocarbons and Lipids on Botryoccocus Braunii UIS 003. Chem. Eng. Trans. 2016, 49, 247–252. 10.3303/CET1649042. [DOI] [Google Scholar]
- Lamela T.; Márquez-Rocha F. J. Phycocyanin Production in Seawater Culture of Arthrospira maxima. Cienc. Mar. 2000, 26, 607–619. 10.7773/cm.v26i4.619. [DOI] [Google Scholar]
- Singh N. K.; Parmar A.; Madamwar D. Optimization of Medium Components for Increased Production of C-Phycocyanin from Phormidium ceylanicum and Its Purification by Single Step Process. Bioresour. Technol. 2009, 100, 1663–1669. 10.1016/j.biortech.2008.09.021. [DOI] [PubMed] [Google Scholar]
- Ravikumar K.; Ramalingam S.; Krishnan S.; Balu K. Application of Response Surface Methodology to Optimize the Process Variables for Reactive Red and Acid Brown Dye Removal Using a Novel Adsorbent. Dyes Pigm. 2006, 70, 18–26. 10.1016/j.dyepig.2005.02.004. [DOI] [Google Scholar]
- Liu G.-Q.; Wang X.-L. Optimization of critical medium components using response surface methodology for biomass and extracellular polysaccharide production by Agaricus blazei. Appl. Microbiol. Biotechnol. 2007, 74, 78–83. 10.1007/s00253-006-0661-6. [DOI] [PubMed] [Google Scholar]
- Thu N. K.; Tanabe Y.; Matsuura H.; Watanabe M. M. biochemical, and molecular characterization of Oscillatoria kawamurae (Oscillatoriales, Cyanobacteria) isolated from different geographical regions. Phycol Res. 2020, 68, 216–226. 10.1111/pre.12419. [DOI] [Google Scholar]
- Soni B.; Kalavadia B.; Trivedi U.; Madamwar D. Extraction, Purification and Characterization of Phycocyanin from Oscillatoria Quadripunctulata—Isolated from the Rocky Shores of Bet-Dwarka, Gujarat, India. Process Biochem. (Oxford, U. K.) 2006, 41, 2017–2023. 10.1016/j.procbio.2006.04.018. [DOI] [Google Scholar]
- Chittapun S.; Jonjaroen V.; Khumrangsee K.; Charoenrat T. C-Phycocyanin Extraction from Two Freshwater Cyanobacteria by Freeze Thaw and Pulsed Electric Field Techniques to Improve Extraction Efficiency and Purity. Algal Res. 2020, 46, 101789. 10.1016/j.algal.2020.101789. [DOI] [Google Scholar]
- Prasanth S.; Kumar Arun G.; Haridas M.; Sabu A. Phycocyanin of Marine Oscillatoria Sp. Inhibits Lipoxygenase by Protein-Protein Interaction-Induced Change of Active Site Entry Apace: A Model for Non-Specific Biofunctions of Phycocyanins. Int. J. Biol. Macromol. 2020, 165, 1111–1118. 10.1016/j.ijbiomac.2020.09.238. [DOI] [PubMed] [Google Scholar]
- Thangam R.; Suresh V.; Asenath Princy W.; Rajkumar M.; SenthilKumar N.; Gunasekaran P.; Rengasamy R.; Anbazhagan C.; Kaveri K.; Kannan S. C-Phycocyanin from Oscillatoria Tenuis Exhibited an Antioxidant and in Vitro Antiproliferative Activity through Induction of Apoptosis and G0/G1 Cell Cycle Arrest. Food Chem. 2013, 140, 262–272. 10.1016/j.foodchem.2013.02.060. [DOI] [PubMed] [Google Scholar]
- Nainangu P.; Antonyraj A. P. M.; Subramanian K.; Kaliyaperumal S.; Gopal S.; Sampath Renuka P.; A W. A. In Vitro Screening of Antimicrobial, Antioxidant, Cytotoxic Activities, and Characterization of Bioactive Substances from Freshwater Cyanobacteria Oscillatoria Sp. SSCM01 and Phormidium Sp. SSCM02. Biocatal. Agric. Biotechnol. 2020, 29, 101772. 10.1016/j.bcab.2020.101772. [DOI] [Google Scholar]
- Markou G.; Vandamme D.; Muylaert K. Microalgal and Cyanobacterial Cultivation: The Supply of Nutrients. Water Res. 2014, 65, 186–202. 10.1016/j.watres.2014.07.025. [DOI] [PubMed] [Google Scholar]
- Hsieh-Lo M.; Castillo G.; Ochoa-Becerra M. A.; Mojica L. Phycocyanin and Phycoerythrin: Strategies to Improve Production Yield and Chemical Stability. Algal Res. 2019, 42, 101600. 10.1016/j.algal.2019.101600. [DOI] [Google Scholar]
- Johnson E. M.; Kumar K.; Das D. Physicochemical parameters optimization, and purification of phycobiliproteins from the isolated Nostoc sp. Bioresour. Technol. 2014, 166, 541–547. 10.1016/j.biortech.2014.05.097. [DOI] [PubMed] [Google Scholar]
- Gilbert S. M.; Allison G. G.; Rogers L. J.; Smith A. J. Expression of Genes Involved in Phycocyanin Biosynthesis Following Recovery of Synechococcus PCC 6301 from Nitrogen Starvation, and the Effect of Gabaculine on CpcBa Transcript Levels. FEMS Microbiol. Lett. 1996, 140, 93–98. 10.1111/j.1574-6968.1996.tb08320.x. [DOI] [PubMed] [Google Scholar]
- Boussiba S. Nitrogen Fixing Cyanobacteria Potential Uses. Plant Soil 1991, 137, 177–180. 10.1007/BF02187452. [DOI] [Google Scholar]
- Markou G.; Georgakakis D. Cultivation of Filamentous Cyanobacteria (Blue-Green Algae) in Agro-Industrial Wastes and Wastewaters: A Review. Appl. Energy. 2011, 88, 3389–3401. 10.1016/j.apenergy.2010.12.042. [DOI] [Google Scholar]
- Grobbelaar J. U.Algal Nutrition – Mineral Nutrition. In: Handbook of Microalgal Culture: Biotechnology and Applied Phycology, Richmond A. Ed. Blackwell Publishing Ltd., Oxford, 2004, pp. 97–115. [Google Scholar]
- Geider R.; La Roche J. Redfield Revisited: Variability of C:N:P in Marine Microalgae and Its Biochemical Basis. Eur. J. Phycol. 2002, 37, 1–17. 10.1017/S0967026201003456. [DOI] [Google Scholar]
- Tiwari O. N.; Bhunia B.; Chakraborty S.; Goswami S.; Devi I. Strategies for Improved Production of Phycobiliproteins (PBPs) by Oscillatoria Sp. BTA170 and Evaluation of Its Thermodynamic and Kinetic Stability. Biochem. Eng. J. 2019, 145, 153–161. 10.1016/j.bej.2019.02.016. [DOI] [Google Scholar]
- Molnár S.; Kiss A.; Virág D.; Forgó P.. Comparative Studies on Accumulation of Selected Microelements by Spirulina platensis and Chlorella vulgaris with the Prospects of Functional Food Development. J. Chem. Eng. Process Technol. 2013, 04, 10.4172/2157-7048.1000172. [DOI] [Google Scholar]
- Akbarnezhad M.; Mehrgan M. S.; Kamali A.; Baboli M. J. Bioaccumulation of Fe+2 and Its Effects on Growth and Pigment Content of Spirulina (Arthrospira Platensis). AACL Bioflux 2016, 9, 227–238. [Google Scholar]
- Kudo I.; Miyamoto M.; Noiri Y.; Maita Y. Combined effects of temperature and iron on the growth and physiology of the marine diatom phaeodactylum tricornutum (bacillariophyceae). J. Phycol. 2008, 36, 1096–1102. 10.1046/j.1529-8817.2000.99042.x. [DOI] [Google Scholar]
- Rueter J. G.; Petersen R. R. Micronutrient effects on cyanobacterial growth and physiology. N. Z. J. Mar. Freshwater Res. 1987, 21, 435–445. 10.1080/00288330.1987.9516239. [DOI] [Google Scholar]
- Singh P.; Guldhe A.; Kumari S.; Rawat I.; Bux F. Investigation of Combined Effect of Nitrogen, Phosphorus and Iron on Lipid Productivity of Microalgae Ankistrodesmus falcatus KJ671624 Using Response Surface Methodology. Biochem. Eng. J. 2015, 94, 22–29. 10.1016/j.bej.2014.10.019. [DOI] [Google Scholar]
- Polat E.; Yüksel E.; Altınbaş M. Mutual Effect of Sodium and Magnesium on the Cultivation of Microalgae Auxenochlorella protothecoides. Biomass Bioenergy 2020, 132, 105441. 10.1016/j.biombioe.2019.105441. [DOI] [Google Scholar]
- Tran H.-L.; Kwon J.-S.; Kim Z.-H.; Oh Y.; Lee C.-G. Statistical Optimization of Culture Media for Growth and Lipid Production of Botryococcus braunii LB572. Biotechnol. Bioprocess Eng. 2010, 15, 277–284. 10.1007/s12257-009-0127-7. [DOI] [Google Scholar]
- Vishwakarma R.; Dhar D. W.; Pabbi S. Formulation of a Minimal Nutritional Medium for Enhanced Lipid Productivity in Chlorella sp. and Botryococcus sp. Using Response Surface Methodology. Water Sci. Technol. 2018, 77, 1660–1672. 10.2166/wst.2018.044. [DOI] [PubMed] [Google Scholar]
- Cheng K.-C.; Ren M.; Ogden K. L. Statistical Optimization of Culture Media for Growth and Lipid Production of Chlorella protothecoides UTEX 250. Bioresour. Technol. 2013, 128, 44–48. 10.1016/j.biortech.2012.09.085. [DOI] [PubMed] [Google Scholar]
- Chen C.-Y.; Ho S.-H.; Liu C.-C.; Chang J.-S. Enhancing Lutein Production with Chlorella sorokiniana Mb-1 by Optimizing Acetate and Nitrate Concentrations under Mixotrophic Growth. J. Taiwan Inst. Chem. Eng. 2017, 79, 88–96. 10.1016/j.jtice.2017.04.020. [DOI] [Google Scholar]
- Tandon P.; Jin Q.; Huang L.; Song R.; Shan A. Effects of Tryptophan Along with Sodium Pyruvate and Sodium Thiosulfate on Chlorella vulgaris Growth. Waste Biomass Valoriz. 2020, 11, 967–982. 10.1007/s12649-018-00577-7. [DOI] [Google Scholar]
- González-Delgado A. D.; Barajas-Solano A. F.; Ardila-Álvarez A. M. Producción de biomasa y proteínas de Chlorella vulgaris Beyerinck (Chlorellales: Chlorellaceae) a través del diseño de medios de cultivo selectivos. Cienc. Tecnol. Agropecuaria 2017, 18, 451. 10.21930/rcta.vol18_num3_art:736. [DOI] [Google Scholar]
- Wang S.; Cao M.; Wang B.; Deng R.; Gao Y.; Liu P. Optimization of growth requirements and scale-up cultivation of freshwater algae Desmodesmus armatus using response surface methodology. Aquacult. Fish. Manage. 2019, 50, 3313–3325. 10.1111/are.14290. [DOI] [Google Scholar]
- Kadkhodaei S.; Abbasiliasi S.; Shun T. J.; Fard Masoumi H. R.; Mohamed M. S.; Movahedi A.; Rahim R.; Ariff A. B. Enhancement of Protein Production by Microalgae Dunaliella salina under Mixotrophic Conditions Using Response Surface Methodology. RSC Adv. 2015, 5, 38141–38151. 10.1039/C5RA04546K. [DOI] [Google Scholar]
- Mubarak M.; Shaija A.; Suchithra T. V. Cost effective approach for production of Chlorella pyrenoidosa: a RSM based study. Waste Biomass Valoriz. 2019, 10, 3307–3319. 10.1007/s12649-018-0330-x. [DOI] [Google Scholar]
- Pandey A.; Gupta A.; Sunny A.; Kumar S.; Srivastava S. Multi-Objective Optimization of Media Components for Improved Algae Biomass, Fatty Acid and Starch Biosynthesis from Scenedesmus sp. ASK22 Using Desirability Function Approach. Renewable Energy 2020, 150, 476–486. 10.1016/j.renene.2019.12.095. [DOI] [Google Scholar]
- Suastes-Rivas J. K.; Hernández-Altamirano R.; Mena-Cervantes V. Y.; Chairez I. Simultaneous Optimization of Biomass and Metabolite Production by a Microalgae-Yeast Co-culture Under Inorganic Micronutrients. BioEnergy Res. 2020, 13, 974. 10.1007/s12155-020-10116-9. [DOI] [Google Scholar]
- Cuéllar-García D. J.; Rangel-Basto Y. A.; Urbina-Suarez N. A.; Barajas-Solano A. F.; Muñoz-Peñaloza Y. A. Lipids production from Scenedesmus obliquus through carbon/nitrogen ratio optimization. J. Phys.: Conf. Ser. 2019, 1388, 012043 10.1088/1742-6596/1388/1/012043. [DOI] [Google Scholar]
- Tourang M.; Baghdadi M.; Torang A.; Sarkhosh S. Optimization of Carbohydrate Productivity of Spirulina Microalgae as a Potential Feedstock for Bioethanol Production. Int. J. Environ. Sci. Technol. 2019, 16, 1303–1318. 10.1007/s13762-017-1592-8. [DOI] [Google Scholar]
- Fekrat F.; Nami B.; Ghanavati H.; Ghaffari A.; Shahbazi M. Optimization of Chitosan/Activated Charcoal-Based Purification of Arthrospira platensis Phycocyanin Using Response Surface Methodology. J. Appl. Phycol. 2019, 31, 1095–1105. 10.1007/s10811-018-1626-8. [DOI] [Google Scholar]
- Gammoudi S.; Athmouni K.; Nasri A.; Diwani N.; Grati I.; Belhaj D.; Bouaziz-Ketata H.; Fki L.; El Feki A.; Ayadi H. \Optimization, Isolation, Characterization and Hepatoprotective Effect of a Novel Pigment-Protein Complex (Phycocyanin) Producing Microalga: Phormidium versicolor NCC-466 Using Response Surface Methodology. Int. J. Biol. Macromol. 2019, 137, 647–656. 10.1016/j.ijbiomac.2019.06.237. [DOI] [PubMed] [Google Scholar]
- Hadiyanto H.; Suttrisnorhadi S. Response surface optimization of ultrasound assisted extraction (UAE) of phycocyanin from microalgae Spirulina platensis. Emir. J. Food Agric. 2016, 28, 227–234. 10.9755/ejfa.2015-05-193. [DOI] [Google Scholar]
- Khazi M. I.; Demirel Z.; Dalay M. C. Evaluation of Growth and Phycobiliprotein Composition of Cyanobacteria Isolates Cultivated in Different Nitrogen Sources. J. Appl. Phycol. 2018, 30, 1513–1523. 10.1007/s10811-018-1398-1. [DOI] [Google Scholar]
- Martínez J. M.; Luengo E.; Saldaña G.; Álvarez I.; Raso J. C-Phycocyanin Extraction Assisted by Pulsed Electric Field from Artrosphira platensis. Food Res. Int. 2017, 99, 1042–1047. 10.1016/j.foodres.2016.09.029. [DOI] [PubMed] [Google Scholar]
- Nur M. M. A.; Garcia G. M.; Boelen P.; Buma A. G. J. Enhancement of C-Phycocyanin Productivity by Arthrospira platensis When Growing on Palm Oil Mill Effluent in a Two-Stage Semi-Continuous Cultivation Mode. J. Appl. Phycol. 2019, 31, 2855–2867. 10.1007/s10811-019-01806-9. [DOI] [Google Scholar]
- Pereira T.; Barroso S.; Mendes S.; Amaral R. A.; Dias J. R.; Baptista T.; Saraiva J. A.; Alves N. M.; Gil M. M. Optimization of Phycobiliprotein Pigments Extraction from Red Algae Gracilaria gracilis for Substitution of Synthetic Food Colorants. Food Chem. 2020, 126688. 10.1016/j.foodchem.2020.126688. [DOI] [PubMed] [Google Scholar]
- Rodrigues R. D. P.; de Castro F. C.; de Santiago-Aguiar R. S.; Rocha M. V. P. Ultrasound-Assisted Extraction of Phycobiliproteins from Spirulina (Arthrospira) platensis Using Protic Ionic Liquids as Solvent. Algal Res. 2018, 31, 454–462. 10.1016/j.algal.2018.02.021. [DOI] [Google Scholar]
- Ruiz-Domínguez M. C.; Jáuregui M.; Medina E.; Jaime C.; Cerezal P. Rapid Green Extractions of C-Phycocyanin from Arthrospira maxima for Functional Applications. Appl. Sci. 2019, 9, 1987. 10.3390/app9101987. [DOI] [Google Scholar]
- Mogany T.; Swalaha F. M.; Kumari S.; Bux F. Elucidating the Role of Nutrients in C-Phycocyanin Production by the Halophilic Cyanobacterium Euhalothece sp. J. Appl. Phycol. 2018, 30, 2259–2271. 10.1007/s10811-018-1477-3. [DOI] [Google Scholar]
- Kumar Saini D.; Yadav D.; Pabbi S.; Chhabra D.; Shukla P. Phycobiliproteins from Anabaena variabilis CCC421 and Its Production Enhancement Strategies Using Combinatory Evolutionary Algorithm Approach. Bioresour. Technol. 2020, 309, 123347. 10.1016/j.biortech.2020.123347. [DOI] [PubMed] [Google Scholar]
- Xing W.; Lusan L. Effects of the different nitrogen, phosphorus and carbon source on the growth and glycogen reserves in Synechocystis and Anabaena. Afr. J. Microbiol. Res. 2013, 7, 2820–2827. 10.5897/AJMR2013.5787. [DOI] [Google Scholar]
- Rastogi R. P.; Sonani R. R.; Madamwar D. Physico-Chemical Factors Affecting the in Vitro Stability of Phycobiliproteins from Phormidium rubidum A09DM. Bioresour. Technol. 2015, 190, 219–226. 10.1016/j.biortech.2015.04.090. [DOI] [PubMed] [Google Scholar]
- Abd El-Baky H. H.; El-Baroty G. S. Characterization and Bioactivity of Phycocyanin Isolated from Spirulina maxima Grown under Salt Stress. Food Funct. 2012, 3, 381–388. 10.1039/c2fo10194g. [DOI] [PubMed] [Google Scholar]
- Maza L. D. L. Á. R.; Guevara M. Á.; Gómez B. J.; Arredondo-Vega B.; Cortez R.; Licet B. Producción de pigmentos procedentes de Arthrospira maxima cultivada en fotobiorreactores. Rev. Colomb. Biotecnol. 2017, 19, 108–114. 10.15446/rev.colomb.biote.v19n1.59671. [DOI] [Google Scholar]
- Xie Y.; Jin Y.; Zeng X.; Chen J.; Lu Y.; Jing K. Fed-Batch Strategy for Enhancing Cell Growth and C-Phycocyanin Production of Arthrospira (Spirulina) platensis under Phototrophic Cultivation. Bioresour. Technol. 2015, 180, 281–287. 10.1016/j.biortech.2014.12.073. [DOI] [PubMed] [Google Scholar]
- Haddad M. F.; Dayioglu T.; Yaman M.; Nalbantoglu B.; Cakmak T. Long-Term Diazotrophic Cultivation of Trichormus sp. IMU26: Evaluation of Physiological Changes Related to Elevated Phycobiliprotein Content. J. Appl. Phycol. 2020, 32, 881–888. 10.1007/s10811-019-02012-3. [DOI] [Google Scholar]
- Plackett R. L.; Burman J. P. The design of optimum multifactorial experiments. Biometrika 1946, 33, 305–325. 10.1093/biomet/33.4.305. [DOI] [Google Scholar]
- Bennett A.; Bogorad L. Complementary Chromatic Adaptation in a Filamentous Blue-Green Alga. J. Cell Biol. 1973, 58, 419–435. 10.1083/jcb.58.2.419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patil G.; Chethana S.; Sridevi A. S.; Raghavarao K. S. M. S. Method to Obtain C-Phycocyanin of High Purity. J. Chromatogr. A 2006, 1127, 76–81. 10.1016/j.chroma.2006.05.073. [DOI] [PubMed] [Google Scholar]