

Abstract
Results of several recent translational studies have suggested that correlating contextual or discriminative stimuli with the delivery and withholding of reinforcement for the functional communication response (FCR) may mitigate resurgence of destructive behavior, but few, if any, have isolated the effects of those stimuli. In the present study, we first trained the FCR, brought it under stimulus control of a multiple schedule, and thinned its reinforcement schedule in one stimulus context. Next, we conducted resurgence evaluations (i.e., baseline, functional communication training [FCT], extinction challenge) in two novel contexts to test the effects of the discriminative stimuli on resurgence. We programmed one context to include the (a) SD during the FCT phase to signal the availability of reinforcement for the FCR and (b) SΔ during a subsequent extinction challenge to signal the unavailability of reinforcement for the FCR. The other context did not include the SD during the FCT phase, nor the SΔ during the extinction challenge. We expected to see greater persistence of the FCR in the context that included the SD during FCT and less persistence of the FCR and destructive behavior in the context that included the SΔ during the extinction challenge. Obtained results confirmed this latter prediction but we observed no reliable difference when the SD was present or absent during the FCT phase. Our results have relevance for practitioners in that they provide further empirical support for the use of discriminative stimuli when treating destructive behavior.
Keywords: autism spectrum disorder, behavioral momentum theory, destructive behavior, functional communication training, resurgence, stimulus control
Functional communication training (FCT) is a commonly prescribed function-based treatment for destructive behavior (Fisher, Greer, & Bouxsein, in press). During FCT, a functional communication response (FCR) produces access to the reinforcer for destructive behavior based on the results of a functional analysis (i.e., functional reinforcement), and destructive behavior is placed on extinction. Reinforcement-schedule thinning is a common practice that behavior analysts use to improve the practicality of FCT in natural settings (e.g., home and school; Saini, Miller, & Fisher, 2016). Although FCT typically reduces destructive behavior by more than 95% post reinforcement-schedule thinning (e.g., Greer, Fisher, Saini, Owen, & Jones, 2016), treatment relapse in the form of resurgence often occurs during the thinning process (Briggs, Fisher, Greer, & Kimball, 2018). Behavior analysts describe relapse of destructive behavior as resurgence when destructive behavior increases following elimination, or a marked reduction in rate, of reinforcement for the FCR. Resurgence may be likely when reinforcement density for the FCR decreases during reinforcement-schedule thinning or when competing responsibilities prevent the caregiver from providing reinforcement for the FCR.
Researchers have conducted extinction challenges to simulate conditions under which resurgence occurs during FCT. The contingences in a resurgence progression typically involve delivery of functional reinforcement for destructive behavior in Phase 1, delivery of functional reinforcement for the FCR and extinction of destructive behavior in Phase 2, and extinction for both responses in Phase 3. Briggs et al. (2018) found that resurgence is common when using FCT—at least one episode of resurgence in over 75% (19/25) of FCT applications during reinforcement-schedule thinning. In addition, Mitteer, Greer, Fisher, Briggs, and Wacker (2018) trained caregivers to implement FCT with high integrity and examined the conditions under which resurgence of undesirable caregiver behavior occurred, with undesirable caregiver behavior including reinforcement of destructive behavior. Mitteer et al. found that caregiver reinforcement of destructive behavior increased when desirable caregiver behavior (i.e., implementing treatment) had no effect on destructive behavior during a context change. This finding is important because resurgence of undesirable caregiver behavior in response to child destructive behavior will negatively impact the long-term effectiveness of FCT.
Given the prevalence of resurgence and its negative impact, translational researchers have evaluated procedures for its mitigation. Previous translational research has shown that reinforcing multiple alternative-response topographies (Falcomata et al., 2018; Lambert, Bloom, Samaha, Dayton, & Rodewald, 2015) or changing from differential reinforcement of alternative behavior to a fixed-time reinforcement schedule (Lieving & Lattal, 2003; Marsteller & St. Peter, 2014) can mitigate resurgence of destructive behavior. Results of several recent studies have suggested that using contextual or discriminative stimuli during FCT also can mitigate resurgence. Betz, Fisher, Roane, Mintz, and Owen (2013) used discriminative stimuli within a multiple schedule and rapidly leaned the per-session reinforcement rate by lengthening the extinction component from 1 to 4 min. They did not observe resurgence of destructive behavior for any of the four children during 4- to 8-min extinction periods. However, the multiple schedule alternated between periods of reinforcement and extinction rather than several, successive extinction sessions typical of an extinction challenge. Whether low levels of destructive behavior (i.e., no resurgence) would have maintained had they conducted a more typical extinction challenge remains untested.
Similarly, Mace et al. (2010) used a multielement design to evaluate the effects of FCT in the presence of unique contextual stimuli, which were the room and the color of the experimenter’s hospital gown. The experimenter delivered (a) no differential consequence for FCRs and functional reinforcement for disruption in the Context 1 baseline; (b) functional reinforcement for FCRs and disruption, similar to FCT without extinction, in the Context 2 baseline; and (c) functional reinforcement for FCRs alone, similar to FCT with extinction, in the Context 3 baseline. The experimenters then introduced extinction challenges in Context 1, Context 2, and a composite of Contexts 1 and 3, which they conducted in the Context 1 room with the experimenter wearing the Context 3 gown color. Rates of disruption were lower in the extinction challenge in the Composite Context relative to the extinction challenge in Context 2 and similarly low in the Composite Context and Context 1. However, a limitation of the study is that the amount of exposure of extinction in Context 1 was twice as much as that in Context 2.
Mace et al. (2010) hypothesized that resurgence was lower in the Composite Context because the experimenters first trained the FCR in Context 3, where the participants did not have a history of reinforcement for disruption, and they correlated therapist gown color with the training (i.e., gown color signaled reinforcement for the FCR and extinction of destructive behavior). The experimenters then conducted the extinction challenge in the Composite Context, which included the room associated with Context 1, where the participants had a history of reinforcement for disruption, but the gown color signaled differential reinforcement of the FCR. However, it remains unclear whether the low rates of resurgence observed in the Composite Context resulted from the stimulus features of Context 1, the color of the gown worn by the therapist to signal differential reinforcement of the FCR, or a combination of the two. That is, the extent to which gown color functioned as a discriminative stimulus that mitigated resurgence in the Mace et al. study remains unknown.
Fuhrman, Fisher, and Greer (2016) compared the effects of a multiple schedule (mult FCT) with discriminative stimuli to traditional FCT (trad FCT) without discriminative stimuli. The discriminative stimuli from mult FCT signaled when the FCR produced reinforcement and when it did not. Fuhrman et al. conducted conditions in different contexts identified by the room and the experimenter’s clothing color. One participant experienced baseline, trad FCT, extinction challenge followed by baseline, mult FCT, extinction challenge. The other participant experienced baseline, mult FCT, extinction challenge followed by baseline, trad FCT, extinction challenge. The SΔ was present during mult FCT and its subsequent extinction challenge but not during trad FCT and its subsequent extinction challenge. The experimenter delivered time-based reinforcement on an FT 200-s schedule during the extinction challenges. They observed lower levels of resurgence during the extinction challenges that followed mult FCT and included the SΔ relative to the extinction challenges that followed trad FCT and did not include the SΔ.
Fuhrman et al. (2016) hypothesized that the SΔ from mult FCT exerted stimulus control over destructive behavior, which resulted in the reduced levels of resurgence during the extinction challenges that followed mult FCT. This conclusion remains tentative given procedural limitations in the study. First, reinforcement-schedule thinning during mult FCT produced different lengths of exposure to and different reinforcement rates during mult FCT and trad FCT. Second, time-based reinforcement delivery on an FT 200-s schedule during the extinction challenges may have increased the rate of destructive behavior through reinstatement or decreased it because an alternative source of reinforcement was available.
In the present investigation, we examined the efficacy of discriminative stimuli in mitigating resurgence of destructive behavior in a clinical setting by addressing the limitations of Fuhrman et al. (2016), using a multielement design and separate stimulus contexts to control for prior exposure to experimental procedures, contextual stimuli, and reinforcement rates. We first conducted pretraining in one context (Context A), in which we trained the FCR, brought it under the control of an SD and SΔ during mult FCT, and thinned the reinforcement schedule. The SD and SΔ in mult FCT in Context A signaled reinforcement availability and unavailability for the FCR, respectively. Next, we conducted a resurgence evaluation consisting of baseline, FCT, and extinction in two novel contexts, Contexts B and C. Table 1 provides an overview of the contexts, presence or absence of discriminative stimuli, and reinforcement schedules across the two contexts for all participants (described in more detail in the Method). We randomly selected one of the two novel contexts (Context C) to include the (a) SD to signal the availability of reinforcement for the FCR during the FCT phase and (b) SΔ to signal the unavailability of reinforcement for the FCR during a subsequent extinction challenge. The other context (Context B) did not include the SD during the FCT phase, nor the SΔ during the extinction challenge. Because we controlled for prior exposure to experiemental procedures, contextual stimuli (i.e., room and therapist gown color), and reinforcement rates, rates of destructive behavior during the extinction challenge reflected the effects of the presence or absence of discriminative stimuli. We expected to see greater persistence of the FCR in the context that included the SD during FCT and less persistence of the FCR and destructive behavior in the context that included the SΔ during the extinction challenge, results that would provide further empirical support for the use of discriminative stimuli when treating destructive behavior.
Table 1.
Reinforcement Schedules During the Resurgence Evaluation in Contexts B and C
| Resurgence Evaluation |
||||||
|---|---|---|---|---|---|---|
| Context B |
Context C |
|||||
| Child |
Baseline |
FCT (No SD) |
Extinction (No SΔ) |
Baseline |
FCT (SD) |
Extinction (SΔ) |
| Owen | VI 40 | VI 20 | - | VI 40 | VI 20 | - |
| Kendrick | VI 4 | VI 2 | - | VI 4 | VI 2 | - |
| Afton | VI 40 | VI 20 | - | VI 40 | VI 20 | - |
| Teddy | VI 8 | VI 4 | - | VI 8 | VI 4 | - |
Note: VI = variable-interval reinforcement schedule.
Method
Subjects and Setting
Participants were four children referred to a university-based clinic for the assessment and treatment of destructive behavior. Afton was a 9-year-old girl diagnosed with attention-deficit/hyperactivity disorder, oppositional defiant disorder, and mixed receptive-expressive language disorder. Kendrick was a 13-year-old boy diagnosed with disruptive mood dysregulation disorder. Owen was a 16-year-old boy diagnosed with autism spectrum disorder. Teddy was a 4-year-old boy diagnosed with autism spectrum disorder. Afton and Kendrick communicated using full sentences and conversational speech. Owen and Teddy communicated with gestures or one- to two-word phrases. Owen participated in Fisher, Saini, et al. (2019) before the present study. Teddy participated in Greer, Mitteer, Briggs, Fisher, and Sodawasser (2019) before the present study and in Greer, Fisher, et al. (2019) after the present study. For the present study, a functional analysis identified the following functional reinforcers for destructive behavior for Afton, Kendrick, Owen, and Teddy—attention, a tangible item and attention, social avoidance, and a tangible item, respectively.
We conducted sessions in a 3-m by 3-m therapy room adjacent to an observation room equipped with a one-way observation window and a two-way intercom. The therapy room had padding on the walls and floors to minimize the risk of self-injurious behavior.
Response Measurement and Reliability
Trained observers sat in the observation room and used BDataPro (Bullock, Fisher, & Hagopian, 2017) to collect and analyze data on laptop computers. Data collectors recorded the frequency of destructive behavior and FCRs, and BDataPro converted these frequencies to responses per minute. Data collectors recorded the duration of reinforcement in seconds, and BDataPro reported the total frequency of reinforcement intervals. We defined destructive behavior as hitting, kicking, scratching, pushing, pinching, throwing objects, self-hitting, self-biting, self-scratching, head banging, body slamming, hitting or kicking furniture or the walls or floor of the therapy room, swiping materials, turning over furniture, or tearing clothing. Functional communication responses consisted of the child handing the therapist an FCR card that contained a picture of him or her consuming his or her functional reinforcer.
A second observer collected data simultaneously with, but independently of, the primary data collector for a minimum of 23% of sessions across experimental conditions to obtain interobserver agreement data. During 39% of sessions for which we assessed interobserver agreement, the second observer was naïve to the experimental question and hypotheses for blinding purposes. BDataPro divided each session into 10-s intervals to calculate interobserver agreement. For destructive behavior and FCRs, BDataPro summed the number of intervals in which both observers scored the same number of responses, divided that sum by the total number of intervals, and converted the quotient to a percentage. For duration of reinforcement, BDataPro divided the smaller duration of reinforcement by the larger duration in each interval, summed the ratios, divided that number by the total number of intervals in the session, and converted the quotient to a percentage. BDataPro substituted a ratio of 1.0 for each interval in which both observers recorded zero seconds of reinforcement. We assessed interobserver agreement on 33%, 33%, 83%, and 23% of sessions for Afton, Kendrick, Owen, and Teddy, respectively. Mean interobserver agreement for destructive behavior and FCRs was 99% (range, 87% to 100%) for Afton, 98% (range, 63% to 100%) for Kendrick, 99% (range, 75% to 100%) for Owen, and 98% (range, 70% to 100%) for Teddy. Mean interobserver agreement for duration of reinforcement was 97% (range, 90% to 100%) for Afton, 98% (range, 89% to 100%) for Kendrick, 96% (range, 85% to 100%) for Owen, and 93% (range, 70% to 100%) for Teddy.
Experimental Design
Pretraining in Context A.
We conducted pretraining in Context A. The therapist provided functional reinforcement for destructive behavior during baseline and then implemented FCT using a multiple schedule. The purpose of pretraining in Context A was twofold. First, we sought to establish the efficacy of mult FCT prior to our resurgence evaluation. Second, we wished to provide a history of mult FCT in a context distinct from those used during the resurgence evaluation.
Resurgence evaluation in Contexts B and C.
We used a multielement design to evaluate resurgence in Context B relative to Context C. Table 1 provides an overview of the contexts, stimuli, and reinforcement schedules across the two contexts for all participants. We used a three-phase resurgence evaluation in which Phase 1 was baseline, Phase 2 was FCT, and Phase 3 was the extinction challenge. The therapist delivered reinforcement for destructive behavior in baseline, placed destructive behavior on extinction and provided functional reinforcement for the FCR during FCT, and placed destructive behavior and the FCR on extinction during the extinction challenge. In Context C alone, the SD from pretraining was present during FCT, and the SΔ from pretraining was present during the extinction challenge. Context B included no discriminative stimuli from pretraining during FCT or during the extinction challenge. We compared levels of destructive behavior and FCRs in Context C with the discriminative stimuli present with Context B, which did not include the discriminative stimuli, to evaluate whether and to what extent the discriminative stimuli exerted stimulus control over responding in both FCT and during the extinction challenge.
General Procedure
Initial treatment evaluation.
Prior to implementing the procedures described below, we evaluated the effects of FCT in a reversal design (data not shown). During this initial treatment evaluation, we delivered the functional reinforcer on a fixed-ratio (FR) 1 schedule only for destructive behavior in baseline and, after teaching the FCR, delivered the functional reinforcer on an FR 1 schedule only for the FCR in treatment. To teach the FCR, we conducted sessions that consisted of 10 trials each, and we used a progressive-prompt delay (Charlop, Schreibman, & Thibodeau, 1985) to transfer stimulus control from a physical prompt to the presentation of the establishing operation. We increased the prompt delay (0 s, 2 s, 5 s, 10 s) following every two sessions with zero instances of destructive behavior and terminated teaching when the participant emitted the FCR independently for 80% of trials with low to zero rates of destructive behavior for two consecutive sessions. All participants showed at least an 85% reduction in destructive behavior during treatment relative to baseline.
Contextual stimuli.
We randomly assigned a color for the stimuli in each of the three contexts to facilitate discrimination between the contexts (Conners et al., 2000). This resulted in three unique contextual stimuli for each participant. The stimuli included light filters on the overhead lights of the therapy room, the FCR card, and the therapists’ scrub top. Stimulus color remained the same in the assigned contexts throughout the experiment. For example, we included blue stimuli in Context A, red in Context B, and yellow in Context C. The therapist asked the child to vocally label the color of his or her scrub top before each session. The therapist provided a vocal prompt (e.g., “My top is red”) if the child did not accurately label the color within 5 s of the therapist’s initial vocal prompt.
Changeover delays and reinforcement intervals.
The therapist used a changeover delay (Herrnstein, 1961) to prevent adventitious reinforcement of destructive behavior during all FCT sessions. The therapist did not deliver reinforcement for FCRs that occurred within 3 s of destructive behavior during FCT and delayed the change from the SΔ to the SD until no FCRs or destructive behavior occurred for at least 3 s during mult FCT. When programmed, the reinforcement interval lasted 20 s. Sessions lasted 10 min.
Pretraining in Context A
The purpose of pretraining was to bring the FCR under the discriminative control of the SD and SΔ with the use of a multiple schedule in Context A. The functional reinforcer was not available when the session started. Thus, the therapist did not provide attention to Afton, did not provide attention or a tangible item to Kendrick, spoke to Owen, and did not provide a tangible item to Teddy.
Baseline.
The therapist began the session by restricting access to the functional reinforcer. For example, for Owen, whose functional reinforcer consisted of escape from social interaction, the therapist began speaking to him. Contingent upon destructive behavior, the therapist provided the participant with access to the functional reinforcer according to an FR 1 schedule of reinforcement. For example, for Owen, the therapist terminated social interaction for 20 s.
Mult FCT.
The therapist used discriminative stimuli to signal the availability (SD) and unavailability (SΔ) of reinforcement for the FCR. Discriminative stimuli were uniquely colored, 7.6-cm by 12.7-cm index cards for Afton, Kendrick, and Owen or uniquely colored 56-cm x 71-cm poster boards for Teddy. Therapists randomly selected the colors for the discriminative stimuli and ensured they were different than the colors programmed for contextual stimuli. The therapist described the contingencies before each session, “When the (color of SD) card is out (while showing the SD to the participant), that means you can hand me the card like this (while the therapist models the FCR), and you can have the (functional reinforcer), but when the (color of SΔ) card is out (while showing the SΔ to the participant), you cannot have the (functional reinforcer), even if you try to hand me the card. I will not switch it back to (color of SD) until you have safe hands and feet.”
During the SD component of mult FCT, the therapist delivered functional reinforcement for the FCR on an FR 1 schedule and placed destructive behavior on extinction. During the SΔ component, the therapist placed FCRs and destructive behavior on extinction. The initial multiple schedule consisted of a 60-s SD component and a 30-s SΔ component. After two consecutive sessions with mean rates of destructive behavior below an 85% reduction from baseline, the therapist lengthened the duration of the SΔ component to 240 s. Each session began with the SD component followed by the SΔ component, and the therapist presented the remaining components quasirandomly, with no more than two of the same component occurring consecutively.
Teddy rarely attended to the discriminative stimuli and did not display discriminative responding during initial mult-FCT sessions. Therefore, therapists conducted additional discrimination training with Teddy (data not displayed) outside of the study context. The procedure included blocking FCRs during the SΔ component, prompting FCRs during the SD component, changing the discriminative stimuli from index cards to poster boards, and affixing the FCR card to the discriminative stimuli so that the FCR card and the discriminative stimulus remained in his line of sight. The therapist gradually faded and then removed the response blocking and prompting procedures.
Resurgence Evaluation in Contexts B and C
We conducted the three-phase resurgence evaluation in two contexts. We randomly selected one context (i.e., Context C) as our condition with discriminative stimuli, and the other context (i.e., Context B) as our condition without discriminative stimuli. Baseline, FCT, and the extinction challenge occurred in both contexts according to a multielement design within each phase of the three-phase resurgence evaluation.
Baseline.
The contingencies were similar to those of the baseline in Context A, except the therapist delivered functional reinforcement for destructive behavior according to a variable-interval (VI) schedule to better control and equate reinforcement rates in the two contexts. We programmed a VI 40-s reinforcement schedule for Owen and Afton and used a progressive-interval assessment (Fisher, Greer, Fuhrman, et al., 2018; Fisher, Saini, et al., 2019) to prescribe a VI 4-s reinforcement schedule with Kendrick and a VI 8-s reinforcement schedule with Teddy.
FCT.
The therapist described the contingencies before each session, “This card [showing FCR] works just like the card in the other room, but I won’t give you the (functional reinforcer) until you have safe hands and safe feet.” The therapist placed destructive behavior on extinction and delivered functional reinforcement for FCRs. We doubled the rate of reinforcement during FCT relative to baseline by halving the VI-schedule value from baseline. For example, if we used a VI 40-s schedule in baseline, we used a VI 20-s schedule during FCT. Therapists delivered functional reinforcement for the FCR on a VI 20-s schedule for Afton and Owen, a VI 2-s schedule for Kendrick, and a VI 4-s schedule for Teddy. Neither the SD nor the SΔ from mult-FCT pretraining were present during FCT in Context B. By contrast, the SD from mult-FCT pretraining that signaled the availability of functional reinforcement for the FCR was present continuously in FCT in Context C. The continuous presence of the SD during FCT in Context C signaled the availability of functional reinforcement for the FCR.
Extinction challenge.
The therapist placed destructive behavior and FCRs on extinction. That is, the therapist did not provide attention to Afton, did not provide attention or a tangible item to Kendrick, spoke to Owen continuously, and did not provide a tangible item to Teddy throughout the session. Neither the SD nor the SΔ from mult-FCT pretraining were present during the extinction challenge in Context B. By contrast, the SΔ from mult-FCT pretraining that signaled the unavailability of functional reinforcement for the FCR was present continuously during the extinction challenge in Context C. The continuous presence of the SΔ during the extinction challenge in Context C signaled that functional reinforcement for the FCR was unavailable throughout the session.
Results
Afton (Figure 1) engaged in high rates of destructive behavior according to the FR 1 reinforcement schedule during baseline in Context A. Following a high rate of destructive behavior during the first mult-FCT session of Context A, Afton engaged in low rates of destructive behavior and high rates of FCRs, and we rapidly thinned the reinforcement schedule for the FCR by increasing the SΔ duration from 30 s to 240 s. Reinforcement-schedule thinning reduced Afton’s overall rate of FCRs, and most of the FCRs occurred during the SD component exclusively, indicating highly discriminated FCRs.
Figure 1.
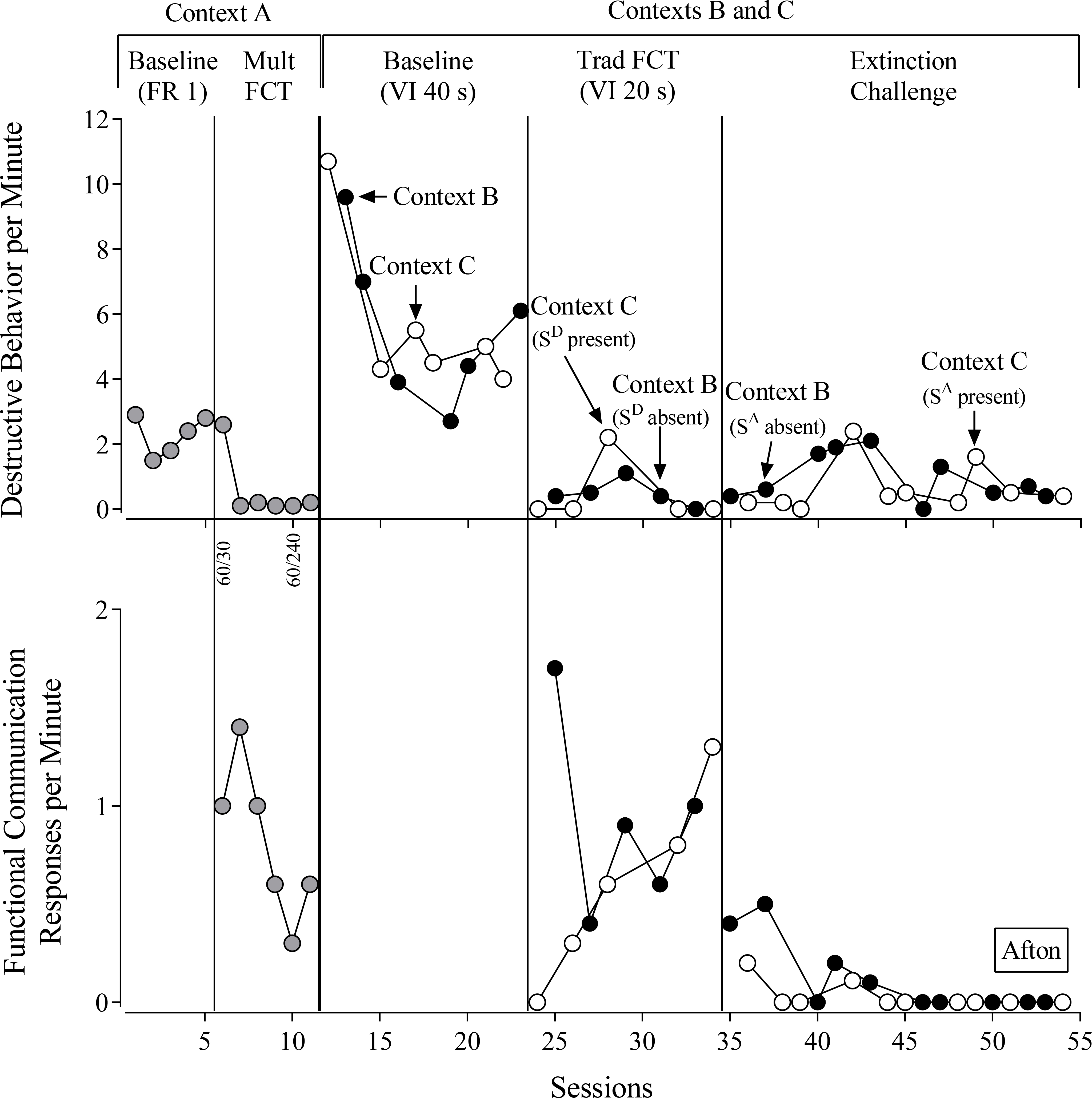
Destructive behavior per minute (top) and functional communication responses per minute (bottom) for Afton. FR = fixed-ratio reinforcement schedule. FCT = functional communication training. VI = variable-interval reinforcement schedule. Numbers below the x-axis for the top panel denote the duration of the SD and SΔ components of the multiple schedule.
During the baselines conducted in Contexts B and C, Afton engaged in high rates of destructive behavior according to the VI 40-s schedule, and obtained similar numbers of reinforcers (see Table 2 for obtained reinforcers). During FCT, Afton engaged in low rates of destructive behavior in Contexts B and C. However, we observed slightly higher rates of destructive behavior in Context B (SD absent) for three of the five sessions with Afton. During the final four FCT sessions, Afton engaged in similar rates of FCRs across the contexts, and she obtained comparable numbers of reinforcers, despite having obtained fewer reinforcers than programmed (i.e., Afton could obtain a maximum of 15 reinforcers per session according to the VI 20-s reinforcement schedule). During the extinction challenge, Afton engaged in variable rates of destructive behavior in both contexts, with slightly higher rates in Context B (SΔ absent) than in Context C (SΔ present). Afton emitted higher rates of FCRs in Context B than in Context C.
Table 2.
Mean and Range of Obtained Reinforcers per Session
| Condition |
||||
|---|---|---|---|---|
| Baseline (VI) |
Trad FCT |
|||
| Child |
Context B |
Context C |
Context B (SD absent) |
Context C (SD present) |
| Owen | 8.6 (7–11) | 9.6 (7–12) | 11.3 (9–14) | 12 (9–14) |
| Kendrick | 20.3 (19–22) | 20.5 (19–22) | 22.5 (22–23) | 21 (20–22) |
| Afton | 8.3 (7–10) | 7.8 (7–10) | 5.8 (4–9) | 3.6 (0–7) |
| Teddy | 16.4 (15–18) | 16.4 (15–18) | 18.3 (16–20) | 17.7 (14–20) |
Note: VI = variable-interval reinforcement schedule.
We defined an episode of resurgence as an increase in responding during the extinction challenge above that observed during any of the last three sessions of the previous FCT phase. Based on this definition, we observed resurgence of destructive behavior in both contexts with Afton. However, we observed greater persistence of FCRs in Context B (SΔ absent) relative to Context C (SΔ present).
Kendrick (Figure 2) displayed high rates of destructive behavior during the baseline in Context A under the FR 1 reinforcement schedule. During mult FCT in Context A, Kendrick engaged in no destructive behavior, even when rapidly thinning the reinforcement schedule by increasing the SΔ duration from 30 s to 240 s. Reinforcement-schedule thinning resulted in a decrease in Kendrick’s FCRs per min from a mean of 1.6 during the first two sessions to 0.4 in the last two sessions of this phase. Kendrick emitted FCRs exclusively during the SD component of mult FCT in Context A. Across mult FCT in Context A, Kendrick emitted 100% of FCRs during the SD component, demonstrating highly discriminated FCRs.
Figure 2.
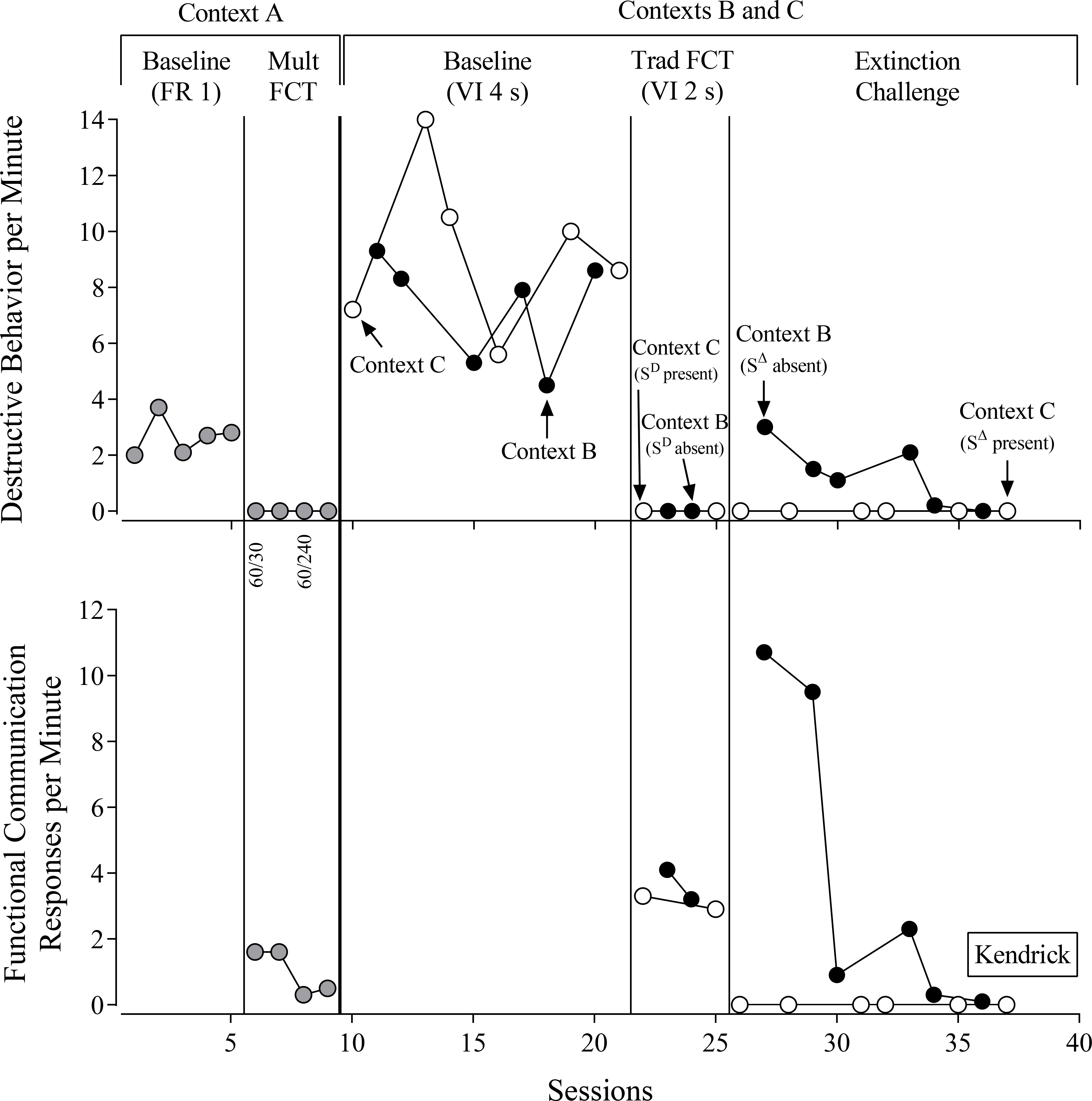
Kendrick’s evaluation. FR = fixed-ratio reinforcement schedule. FCT = functional communication training. VI = variable-interval reinforcement schedule. Numbers below the x-axis for the top panel denote the duration of the SD and SΔ components of the multiple schedule.
During the baselines in Contexts B and C, Kendrick engaged in high rates of destructive behavior under the VI 40-s schedule. Despite Kendrick emitting higher rates of destructive behavior in Context C, he obtained equivalent numbers of reinforcers in each context (see Table 2). During FCT, destructive behavior decreased rapidly to zero rates in both Contexts B and C, and we observed equivalent rates of FCRs in Contexts B and comparable numbers of reinforcers delivered.
During the extinction challenge, Kendrick displayed no destructive behavior in Context C and moderate rates of destructive behavior in Context B. Thus, we observed resurgence of destructive behavior only in Context B (SΔ absent). Presenting the SΔ during sessions in Context C resulted in complete suppression of FCRs throughout the extinction challenge, whereas we observed a burst of FCRs in Context B with the SΔ absent.
Owen (Figure 3) displayed high rates of destructive behavior during the baseline in Context A under the FR 1 reinforcement schedule. During mult FCT in Context A, Owen engaged in no destructive behavior, even when rapidly thinning the reinforcement schedule by increasing the SΔ duration from 30 s to 240 s. Reinforcement-schedule thinning resulted in a decrease in Owen’s FCRs per min from a mean of 1.8 during the first two sessions to 0.5 in the last two sessions of this phase. Owen emitted FCRs exclusively during the SD component of mult FCT in Context A, indicating highly discriminated FCRs.
Figure 3.
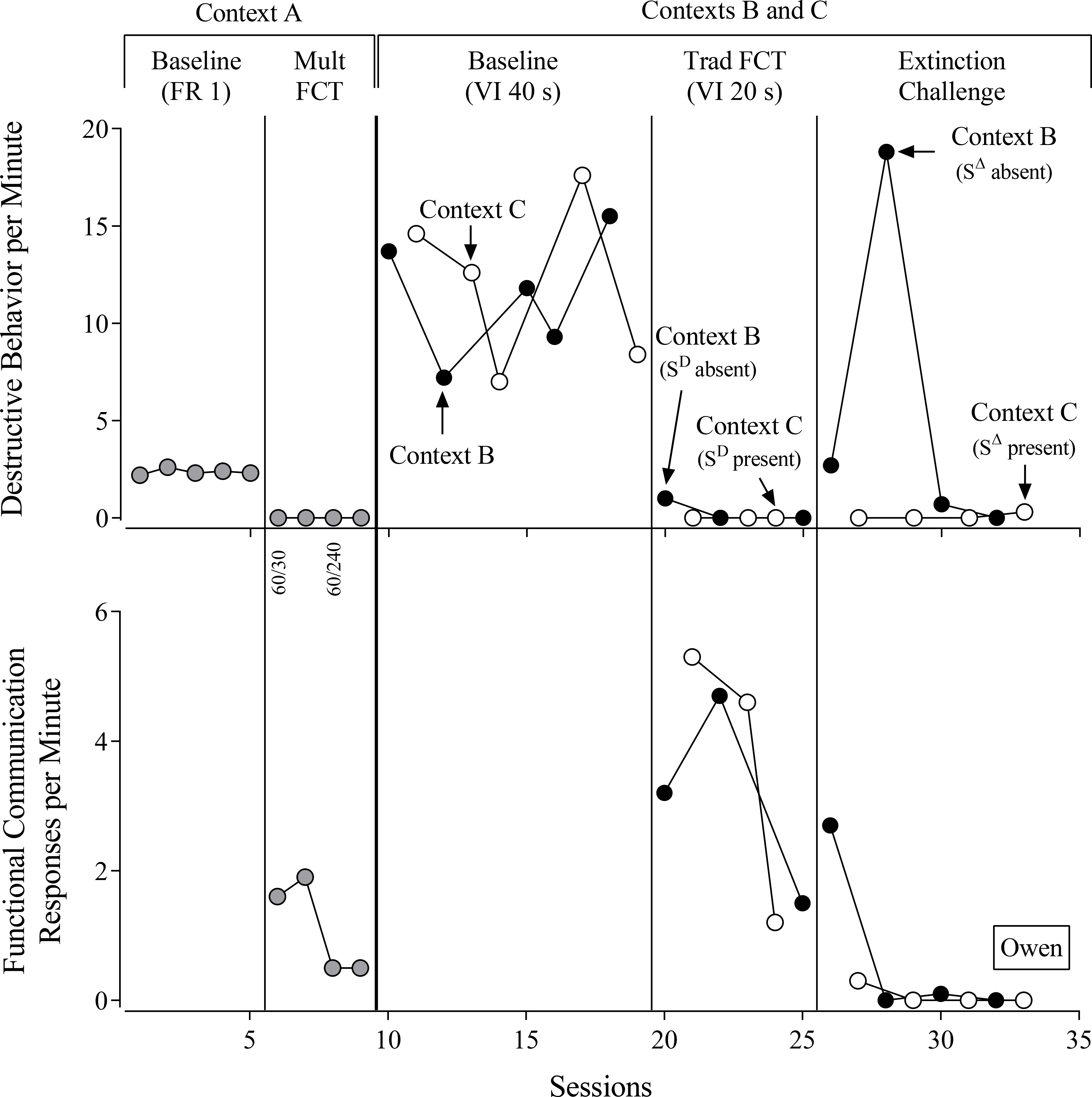
Owen’s evaluation. FR = fixed-ratio reinforcement schedule. FCT = functional communication training. VI = variable-interval reinforcement schedule. Numbers below the x-axis for the top panel denote the duration of the SD and SΔ components of the multiple schedule.
During the baselines in Contexts B and C, Owen engaged in high rates of destructive behavior under the VI 40-s schedule and obtained similar numbers of reinforcers in each context (see Table 2). During FCT, destructive behavior decreased rapidly to near-zero rates in both Contexts B and C, and we observed equivalent rates of FCRs in Contexts B and C and comparable numbers of reinforcers delivered.
During the extinction challenge, Owen engaged in near-zero rates of destructive behavior in Context C (SΔ present) but considerably higher rates of destructive behavior in Context B (SΔ absent). Thus, we observed high levels of resurgence of destructive behavior only in Context B (SΔ absent). We observed roughly equivalent rates of FCRs in Contexts B and C.
Teddy (Figure 4) engaged in high rates of destructive behavior during baseline in Context A under the FR 1 reinforcement schedule. Because Teddy’s FCRs were not discriminated between the multiple-schedule components during the initial mult-FCT session, we conducted multiple-schedule discrimination training. Following these teaching procedures, we returned to mult FCT within Context A, and Teddy engaged in low-to-zero rates of destructive behavior and high rates of FCRs. We then rapidly thinned the reinforcement schedule for the FCR by increasing the SΔ duration from 30 s to 240 s, and Teddy’s destructive behavior remained at near-zero rates. Across mult-FCT sessions in Context A, Teddy emitted 72% of FCRs during the SD, thus showing less discriminated responding than the other three participants.
Figure 4.
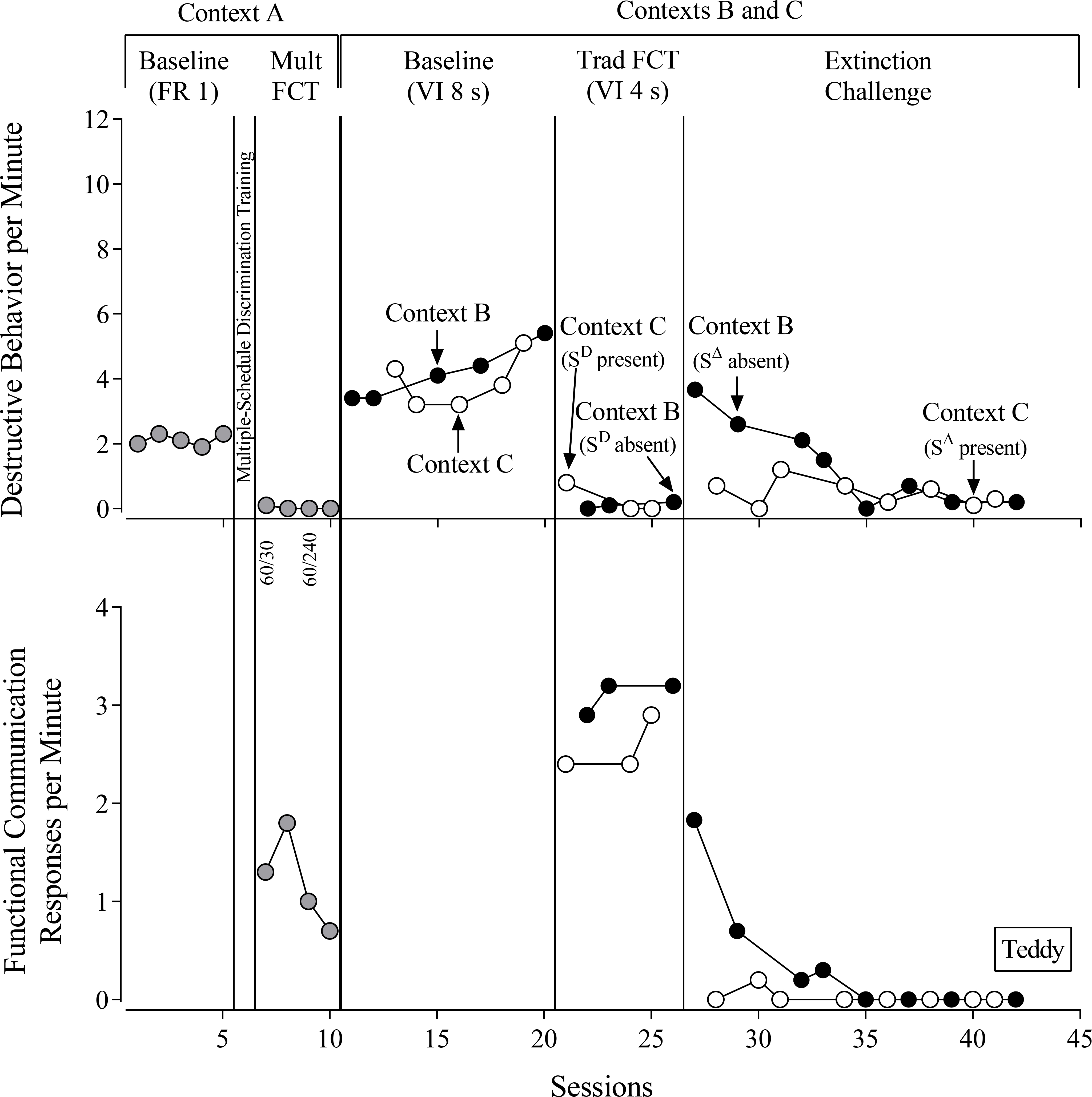
Teddy’s evaluation. FR = fixed-ratio reinforcement schedule. FCT = functional communication training. VI = variable-interval reinforcement schedule. Numbers below the x-axis for the top panel denote the duration of the SD and SΔ components of the multiple schedule. The double phase lines preceding mult FCT indicate direct teaching of the multiple-schedule discrimination for functional communication responses.
During baseline in Contexts B and C, Teddy engaged in similarly high rates of destructive behavior according to the VI 8-s schedule and obtained equal numbers of reinforcers (see Table 2). During FCT, destructive behavior decreased to near-zero rates in both Context B (SD absent) and Context C (SD present). Teddy emitted high rates of FCRs in Context C (SD present) and slightly higher rates in Context B (SD absent). Despite different rates of FCRs across the two contexts, Teddy obtained similar numbers of reinforcers.
During the extinction challenge, Teddy engaged in moderate rates of destructive behavior in Context B (SΔ absent) and much lower rates of destructive behavior in Context C (SΔ present). Thus, we observed higher levels of resurgence of destructive behavior in Context B (SΔ absent). We also observed greater persistence of FCRs in Context B (SΔ absent) relative to Context C (SΔ present).
Figure 5 shows the mean proportion of baseline responding for destructive behavior during the first five (Afton, Kendrick, and Teddy) or all four (Owen) sessions of the extinction challenge conducted in Context B (SΔ absent) and Context C (SΔ present) for each participant. As can be seen, all four participants showed more destructive behavior measured as a proportion of baseline responding in the context without the SΔ (Context B) relative to the context with the SΔ (Context C), with 52%, 100%, 99%, and 72% reductions in destructive behavior in Context C relative to Context B for Afton, Kendrick, Owen, and Teddy, respectively.
Figure 5.
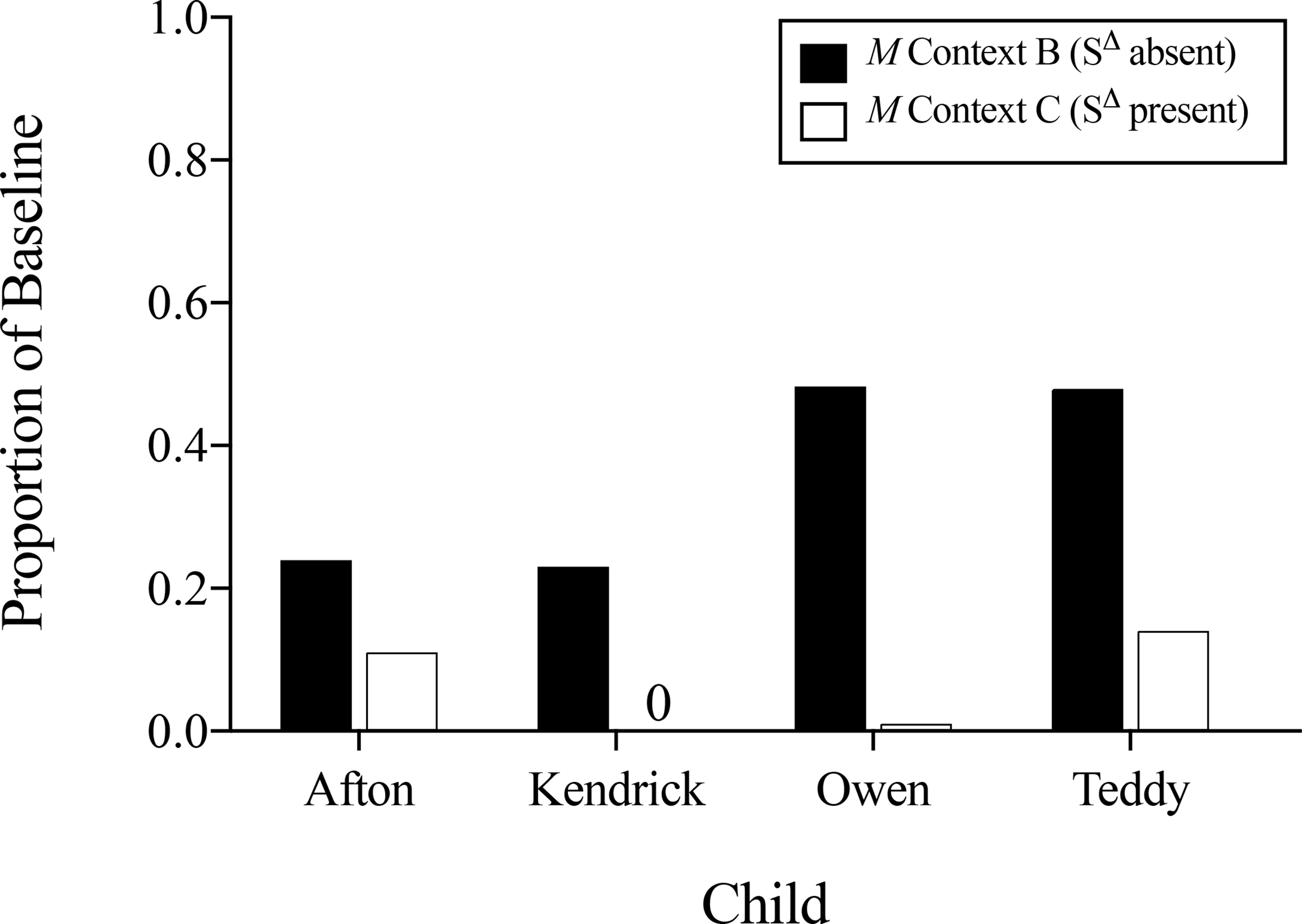
Proportion of baseline responding during all extinction-challenge sessions for each child in Context B (SΔ absent) and Context C (SΔ present).
Statistical analysis.
We also conducted a randomization test (Craig & Fisher, 2019; Edgington, 1995) across all four participants to determine the probability of obtaining differences between the levels of resurgence as large or larger than those we observed between the context with the SΔ absent (Context B) and the context with the SΔ present (Context C). Randomization tests provide more flexible statistical analyses relative to conventional parametric and non-parametric statistical techniques because (a) they make no assumptions about the underlying distribution of outcome variables, (b) show relatively robust results when applied to small-n data sets, and (c) are applicable to between-groups and single-case research designs (Craig & Fisher, 2019). For this analysis, we calculated the mean difference between the levels of resurgence for all sessions of the extinction challenges conducted in Contexts B (SΔ absent) and C (SΔ present) with all participants (total n = 56 sessions, 28 each from Contexts B and C). We then randomly reassigned the data points to the SΔ-absent and SΔ-present conditions and recalculated the mean difference 100,000 times and calculated the proportion of times we obtained a mean difference as larger or larger than the original mean difference, which produced a p-value of less than .001, indicating that it is highly unlikely that the observed differences between the levels of resurgence in Contexts B and C occurred by chance.
Discussion
We taught four participants to emit an FCR that served the same function as their destructive behavior, and we placed destructive behavior on extinction. We then brought the FCR under the discriminative control during mult FCT in Context A using a multiple schedule that signaled quasirandomly alternating periods of extinction (lasting 30 s) and FR-1 reinforcement of the FCR (lasting 60 s) while destructive behavior remained on extinction. Next, we thinned the reinforcement schedule to improve the practicality of the treatment by rapidly lengthening the extinction component of the multiple schedule to 240 s. We then tested whether the discriminative stimuli from mult FCT mitigated resurgence of destructive behavior during an extinction challenge by comparing two equivalent resurgence sequences (baseline → FCT → extinction) in two new contexts, Contexts B and C, and including the SD and SΔ from mult FCT during FCT and the extinction phase, respectively, in Context C but not Context B. Results showed that including the SΔ during the extinction challenge in Context C reduced resurgence by an average of 81% (range, 52% to 100%) and generally decreased persistence of the FCR relative to that we observed in Context B, which did not include the SΔ. We observed no reliable difference when the SD was present or absent during the FCT phase. A randomization test showed that it is highly unlikely that observed differences in resurgence occurred by chance (p < .001). It is also important to note that the function of participants’ destructive behavior (i.e., positive or negative reinforcement) did not seem to impact the results.
The results of the present translational study provide both clinical and theoretical implications. From a clinical standpoint, the present findings replicate and extend the results obtained by Fuhrman et al. (2016) by showing robust effects of including the SΔ from mult FCT during a subsequent extinction challenge relative to an extinction challenge following FCT that did not include the SΔ. Fuhrman et al. observed a mean reduction in resurgence of 82% with two participants, and we observed a mean reduction in resurgence of 81% with four participants. We believe that these data represent the largest reductions in resurgence of destructive behavior for an application of a single mitigation procedure reported in the literature to date. It is worth mentioning that, as in Fuhrman et al., we delivered rules that described the contingencies associated with the discriminative stimuli prior to mult-FCT sessions. Thus, rule-governed behavior may have had a role on the outcomes. For example, Kendrick emitted FCRs only during the SD components of mult FCT, suggesting rule governance of the behavior rather than contingency control. Researchers should investigate how these results hold for participants who do not respond to rules. It is also important to mention that in our terminal treatment packages, we ensure that we present an SD component following extended periods of an SΔ component so individuals have an opportunity to request the reinforcer.
The present findings extend those of Fuhrman et al. by (a) equating the lengths of treatment exposure across contexts; (b) equating the rates of reinforcement across contexts; and (c) eliminating the delivery of time-based reinforcement during the extinction challenge, which may have increased the reoccurrence of destructive behavior through reinstatement or decreased it by providing another source of alternative reinforcement. These changes in the experimental design allowed us to better isolate the effects of the discriminative stimuli of mult FCT.
It is worth noting that we made manipulations similar to those used in studies evaluating extinction cues to decrease operant renewal, or the relapse of previously eliminated responding following a change in context (Podlesnik, Kelley, Jimenez-Gomez, & Bouton, 2017). For example, Nieto, Uengoer, and Bernal-Gamboa (2017) found that the presence of a cue (i.e., tone) from a phase of extinction for lever pressing decreased relapse during a subsequent renewal test. Similarly, Craig, Browning, and Shahan (2017) showed that delivering food-correlated stimuli (i.e., audible pellet-dispenser click and illumination of the food aperture) from a phase of alternative reinforcement for nose pokes decreased relapse of lever pressing during a subsequent resurgence test. Together with the findings of the present study, these results suggest that researchers should continue to investigate how clinicians can use intervention-correlated stimuli to decrease the likelihood of treatment relapse.
Regarding the theoretical implications of the present findings, behavioral momentum theory relies on the generalization-decrement hypothesis to account for the effects of discriminative and contextual stimuli on the persistence and reemergence of destructive behavior following successful initial FCT treatment (Nevin et al., 2017). The generalization-decrement hypothesis states that when we reinforce a response in the presence of a specific stimulus, an SD, the effects of reinforcement tend to generalize to other, untrained stimuli in relation to the degree of physical similarity between the SD and those other stimuli (sometimes called an excitatory generalization gradient; Guttman & Kalish, 1956; Hanson, 1959; Honig & Urcuioli, 1981). Correspondingly, when we program extinction for a specific stimulus, an SΔ, the effects of extinction tend to generalize to other, untrained stimuli in relation to the degree of physical similarity between the SΔ and those other stimuli (sometimes called an inhibitory generalization gradient), although inhibitory stimulus control tends to be less robust than excitatory generalization (Honig, Thomas, & Guttman, 1959; Terrace, 1966).
We hypothesize that mult FCT mitigated the effects of contextual stimuli through processes related to the generalization-decrement hypothesis. First, from a stimulus-control perspective, resurgence can be viewed as generalization of the effects of reinforcement from the baseline and treatment phases to the extinction phase where no reinforcement is delivered. In accordance with the generalization-decrement hypothesis, when the extinction phase is more similar to the baseline phase than the treatment phase, we should see relatively greater resurgence and less persistence of the FCR. Conversely, when the extinction phase is more similar to the treatment phase, we should see relatively less resurgence and greater persistence of the FCR. We hypothesize that the inclusion of the SD and SΔ in the FCT and extinction phases of Context C, respectively, increased the similarity between these two phases and the prior mult-FCT treatment in Context A, where participants experience alternating and signaled periods of reinforcement and extinction for the FCR and extinction of destructive behavior. Including the SD and SΔ from mult FCT in Context C also decreased the similarity between baseline and the extinction challenge in Context C. In particular, presenting the SΔ during the extinction challenge in Context C rendered this condition highly similar to the SΔ components of mult FCT, and we observed relatively low levels of destructive behavior and FCRs, presumably due to the participant’s prior experience with the SΔ signaling extinction for both responses. By contrast, during the resurgence sequence conducted in Context B (baseline → FCT → extinction), with no SD and SΔ present, the stimuli that differentiated these three phases consisted solely of the schedules in effect (e.g., For Owen, VI 40 s for destructive behavior in baseline, VI 20 s for the FCR during FCT, and extinction for both responses during extinction).
It is also worth noting that by correlating the delivery and nondelivery of reinforcement for the FCR with the SD and SΔ, respectively, and programming extinction for destructive behavior in the presence of both the SD and SΔ, we altered the reinforcement and stimulus-control histories for the target and alternative response in Context C. That is, during mult FCT in Context A and during the FCT and extinction phases in Context C, we correlated the SD and SΔ with the delivery and nondelivery of reinforcement, respectively, whereas no other contextual stimuli correlated so fully with the presence or absence of reinforcement (e.g., room color remained constant during periods of reinforcement and nonreinforcement). This difference in the degree of correlation between the discriminative stimuli from mult FCT and the delivery and nondelivery of reinforcement relative to other contextual stimuli probably increased the discriminative effects of the discriminative stimuli from mult FCT during the extinction challenge in Context C relative to the other contextual stimuli in Context C (e.g., room color). That is, the discriminative stimuli from mult FCT had greater discriminative relevance than the contextual stimuli of Context C because the stimuli better predicted the delivery and nondelivery of reinforcement (see Honig & Urcuioli, 1981, for a relevant discussion).
Relatedly, because the SD and SΔ of mult FCT correlated with delivery and nondelivery of reinforcement, they may have heightened the participants’ attention to the SD and SΔ from mult FCT relative to the contextual stimuli of Context C. It is well established that discrimination-training contingencies similar to those involved in mult FCT function to sharpen generalization gradients (i.e., increase stimulus control), thereby weakening the influence of other stimuli not directly involved in the reinforcement contingency (Jenkins & Harrison, 1960; Mackintosh & Honig, 1970; Honig & Urcuioli, 1981). Future translational research should more directly examine whether the relapse-mitigating effects of mult FCT are in part due to stimulus-control blocking. This seems particularly relevant when attempting to mitigate renewal using discrimination-training contingencies similar to those used in the present study.
In summary, our results replicate and extend those obtained by Fuhrman et al. (2016) and demonstrate robust effects of including the SΔ from mult FCT in mitigating resurgence during an extinction challenge. Together, these findings suggest that this single mitigation procedure can produce significant reductions in resurgence, which holds important relevance for practitioners. That is, these findings provide additional empirical support for the use of discriminative stimuli when treating destructive behavior.
Acknowledgments
Grants #5R01HD079113 and #5R01HD083214 from The National Institute of Child Health and Human Development provided partial support for this work. Ashley M. Fuhrman and Daniel R. Mitteer are now at Children’s Specialized Hospital–Rutgers University Center for Autism Research, Education, and Services (CSH–RUCARES). Wayne W. Fisher and Brian D. Greer are now at CSH–RUCARES and Rutgers Robert Wood Johnson Medical School. Cathleen C. Piazza is now at CSH and Rutgers Graduate School of Applied and Professional Psychology.
References
- Betz AM, Fisher WW, Roane HS, Mintz JC, & Owen TM (2013). A component analysis of schedule thinning during functional communication training. Journal of Applied Behavior Analysis, 46, 219–241. 10.1002/jaba.23 [DOI] [PubMed] [Google Scholar]
- Bullock CE, Fisher WW, & Hagopian LP (2017). Description and validation of a computerized behavioral data program: “BDataPro.” The Behavior Analyst, 40, 275–285. 10.1007/s40614-016-0079-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnham K, & Anderson DR (2002). Model selection and multimodel inference: A practical information-theoretic approach. New York: Springer. [Google Scholar]
- Briggs AM, Fisher WW, Greer BD, & Kimball RT (2018). Prevalence of resurgence of destructive behavior when thinning reinforcement schedules during functional communication training. Journal of Applied Behavior Analysis, 51, 620–633. 10.1002/jaba.472 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conners J, Iwata BA, Kahng S, Hanley GP, Worsdell AS, & Thompson RH (2000). Differential responding in the presence and absence of discriminative stimuli during multielement functional analyses. Journal of Applied Behavior Analysis, 33, 299–308. 10.1901/jaba.2000.33-299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charlop MH, Schreibman L, & Thibodeau MG (1985). Increasing spontaneous verbal responding in autistic children using a time delay procedure. Journal of Applied Behavior Analysis, 18, 155–166. 10.1901/jaba.1985.18-155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craig AR, Browning KO, & Shahan TA (2017). Stimuli previously associated with reinforcement mitigate resurgence. Journal of the Experimental Analysis of Behavior, 108, 139–150. 10.1002/jeab.278 [DOI] [PubMed] [Google Scholar]
- Craig AR & Fisher W, W. (2019). Randomization tests as alternative analysis methods for behavior-analytic data. Journal of the Experimental Analysis of Behavior, 111, 309–328. 10.1002/jeab.500 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edgington ES, (1995). Randomization tests (3rd ed.). New York, NY: Marcel Dekker. [Google Scholar]
- Falcomata TS, Muething CS, Silbaugh BC, Adami S, Hoffman K, Shpall C, & Ringdahl JE (2018). Lag schedules and functional communication training: Persistence of mands and relapse of problem behavior. Behavior Modification, 42, 314–334. 10.1177/0145445517741475 [DOI] [PubMed] [Google Scholar]
- Fisher WW, Greer BD, & Bouxsein KJ (in press). Developing function-based reinforcement procedures for problem behavior. In Fisher WW, Piazza CC, & Roane HS (Eds.), Handbook of applied behavior analysis. (2nd ed.). New York: Guilford Publishing. [Google Scholar]
- Fisher WW, Greer BD, Fuhrman AM, Saini V, & Simmons CA (2018). Minimizing resurgence of destructive behavior using behavioral momentum theory. Journal of Applied Behavior Analysis, 51, 831–853. 10.1002/jaba.499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Saini V, Greer BD, Sullivan WE, Roane HS, Fuhrman AM, … Kimball RT (2019). Baseline reinforcement rate and resurgence of destructive behavior. Journal of the Experimental Analysis of Behavior, 111, 75–93. 10.1002/jeab.488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuhrman AM, Fisher WW, & Greer BD (2016). A preliminary investigation on improving functional communication training by mitigating resurgence of destructive behavior. Journal of Applied Behavior Analysis, 49, 884–899. 10.1002/jaba.338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer BD, Fisher WW, Briggs AM, Lichtblau KR, Phillips L, & Mitteer DR (2019). Using discriminative stimuli during functional communication training to promote rapid transfer of treatment effects. Behavioral Development. Advance online publication. 10.1037/bdb0000085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer BD, Mitteer DR, Briggs AM, Fisher WW, & Sodawasser AJ (2019). Comparisons of standardized and interview-informed synthesized reinforcement contingencies relative to traditional functional analysis. Journal of Applied Behavior Analysis. Advance online publication. 10.1002/jaba.601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer BD, Fisher WW, Saini V, Owen TM, & Jones JK (2016). Functional communication training during reinforcement schedule thinning: An analysis of 25 applications. Journal of Applied Behavior Analysis, 49, 105–121. 10.1002/jaba.265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guttman N, & Kalish HI (1956). Discriminability and stimulus generalization. Journal of Experimental Psychology, 51, 79–88. 10.1037/h0046219 [DOI] [PubMed] [Google Scholar]
- Hanson HM (1959). Effects of discrimination training on stimulus generalization. Journal of Experimental Psychology, 58, 321–334. 10.1037/h0042606 [DOI] [PubMed] [Google Scholar]
- Herrnstein RJ (1961). Relative and absolute strength of response as a function of frequency of reinforcement. Journal of the Experimental Analysis of Behavior, 4, 267–272. 10.1901/jeab.1961.4-267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honig WK, Thomas DR, & Guttman N (1959). Differential effects of continuous extinction and discrimination training on the generalization gradient. Journal of Experimental Psychology, 8, 145–152. 10.1037/h0048484 [DOI] [PubMed] [Google Scholar]
- Honig WK & Urcuioli PJ (1981). The legacy of Guttman and Kalish (1956): 25 years of research on stimulus generalization. Journal of the Experimental Analysis of Behavior, 36, 405–445. https://doi.org/jeab.1981.36-405 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenkins HM, & Harrison RH (1960). Effect of discrimination training on auditory generalization. Journal of Experimental Psychology, 59, 246–253. 10.1037/h0041661 [DOI] [PubMed] [Google Scholar]
- Lambert JM, Bloom SE, Samaha AL, Dayton E, & Rodewald A (2015). Serial alternative response training as intervention for target response resurgence. Journal of Applied Behavior Analysis, 48, 765–780. 10.1002/jaba.253 [DOI] [PubMed] [Google Scholar]
- Lieving GA, & Lattal KA (2003). Recency, repeatability, and reinforcer retrenchment: An experimental analysis of resurgence. Journal of the Experimental Analysis of Behavior, 80, 217–233. 10.190/jeab.2003.80-217 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mace FC, McComas JJ, Mauro BC, Progar PR, Taylor B, Ervin R, & Zangrillo AN (2010). Differential reinforcement of alternative behavior increases resistance to extinction: Clinical demonstration, animal modeling, and clinical test of one solution. Journal of the Experimental Analysis of Behavior, 93, 349–367. 10.1901/jeab.2010.93-349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackintosh NJ, & Honig WK (1970). Blocking and enhancement of stimulus control in pigeons. Journal of Comparative and Physiological Psychology, 73, 78–85. 10.1037/h0030021 [DOI] [Google Scholar]
- Marsteller TM, & St. Peter CC (2014). Effects of fixed-time reinforcement schedules on resurgence of problem behavior. Journal of Applied Behavior Analysis, 47, 455–469. 10.1002/jaba.134 [DOI] [PubMed] [Google Scholar]
- Mitteer DR, Greer BD, Fisher WW, Briggs AM, & Wacker DP (2018). A laboratory model for evaluating relapse of undesirable caregiver behavior. Journal of the Experimental Analysis of Behavior, 110, 252–266. 10.1002/jeab.462 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nevin JA, Craig AR, Cunningham PJ, Podlesnik CA, Shahan TA, Sweney MM (2017). Quantitative models of persistence and relapse from the perspective of behavioral momentum theory: Fits and misfits. Behavioural Processes, 141, 92–99. 10.1016/j.beproc.2017.04.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieto J, Uengoer M, & Bernal-Bamboa R (2017). A reminder of extinction reduces relapse in an animal model of voluntary behavior. Learning & Memory, 24, 76–80. 10.11.01/lm.044495.116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podlesnik CA, Kelley ME, Jimenez-Gomez C, & Bouton ME (2017). Renewed behavior produced by context change and its implications for treatment maintenance: A review. Journal of Applied Behavior Analysis, 50, 675–697. 10.1002/jaba.400 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saini V, Miller SA, & Fisher WW (2016). Multiple schedules in practical application: Research trends and implications for future investigation. Journal of Applied Behavior Analysis, 49, 421–444. 10.1002/jaba.300 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terrace HS (1966). Discrimination learning and inhibition. Science, 154, 1677–1680. 10.1126/science.154.3757.1677 [DOI] [PubMed] [Google Scholar]