

Abstract
Lateral organization in the plane of the plasma membrane is an important driver of biological processes. The past dozen years have seen increasing experimental support for the notion that lipid organization plays an important role in modulating this heterogeneity. Various biophysical mechanisms rooted in the concept of liquid–liquid phase separation have been proposed to explain diverse experimental observations of heterogeneity in model and cell membranes with distinct but overlapping applicability. In this review, we focus on the evidence for and the consequences of the hypothesis that the plasma membrane is poised near an equilibrium miscibility critical point. Critical phenomena explain certain features of the heterogeneity observed in cells and model systems but also go beyond heterogeneity to predict other interesting phenomena, including responses to perturbations in membrane composition.
Keywords: critical composition fluctuations, thermodynamics, cell membrane, membrane microdomains, lipid rafts
1. OVERVIEW
The spatial organization of the plasma membrane on 10–100-nm length scales has been a topic of interest in biology for decades. Heterogeneity has been hypothesized to play important roles in many membrane-associated biological processes, from the coordination of signal transduction machinery, to endo- and exocytosis, to polarization, as iconically described in the lipid raft hypothesis (1). Various biophysical notions have been marshaled to provide explanations of membrane heterogeneity, especially from equilibrium thermodynamics and phase transitions, as coexisting liquid phases are readily observed in both purified membranes and membranes isolated from plasma membranes (2, 3). One important thread of this ongoing conversation explains nanoscale membrane structure in terms of critical phenomena. In contrast to a classical phase separation picture of plasma membrane domains, which implies stable, discrete regions of defined composition, critical phenomena are subtle, dynamic, and malleable and inhabit the relevant nanoscopic length scales.
In this review, we conduct a brief historical survey of membrane domains in both model and biological membranes and describe the consensus that has been reached regarding the macroscopic miscibility phase behavior of model membranes as well as the remaining controversies regarding the microscopic heterogeneity reported in other regions of phase space. We introduce membrane criticality and the accumulated evidence that eukaryotic plasma membranes are near-critical. We discuss how the concept of criticality fits into conventional descriptions of raft phenomenology and describe some unique areas in which criticality could play roles in biological function that go beyond the simple organization of components.
2. AN EARLY HISTORY OF DOMAINS IN MODEL AND CELL MEMBRANES
There is a long history of detecting heterogeneity in bilayer membranes containing cholesterol. Some of the earliest studies that interrogated purified membranes in the late 1960s and early 1970s found evidence that liquid membranes containing a single phospholipid species and cholesterol contained structure on the 1–100-nm length scales accessible to the spectroscopic methods available at the time (4–9). Over the decades, evidence for these microscopic domains in binary mixtures of phospholipids and cholesterol continued to accumulate through additional spectroscopic studies; calorimetry; the application of fluorescence techniques such as fluorescence quenching, anisotropy, and Förster resonance energy transfer (FRET); and imaging methodologies such as freeze-fracture electron microscopy (10–21).
Largely in parallel with this physical chemistry characterization of membranes, cell biologists also postulated that membranes within cells might contain lipid-mediated substructures that could be important for cell functions. Early evidence of this came from the observation that cells could polarize their membrane lipid composition via the differential trafficking of lipid species (22). In addition, cholesterol or similar sterols have long been known to be vital to the proper functioning of most eukaryotic plasma membranes, as manipulation of membrane cholesterol content has been shown to interfere with functions including endocytosis, several receptor-mediated signaling cascades, and cell cycle control (23–26). In the early 1990s it was discovered that some detergents only partially solubilize cellular membranes, especially the plasma membrane (27, 28), and it was postulated that the insoluble fractions represented distinct membrane domains (1, 20). Researchers found that artificially clustering some membrane components individually could drive macroscopic colocalization, a process termed copatching, which was detectable by conventional fluorescence microscopy (29). Subsequent spectroscopic and fluorescence measurements supported the hypothesis that cell membranes were heterogeneous in their lipid and protein content (30–32), and many studies correlated biological function with presumed membrane heterogeneity that had been probed and perturbed via these and related assays (33–36). Each of these methods had well-documented flaws (37), but the remarkable consistency of the conclusions drawn from different methodologies provided convincing evidence that lipid-driven heterogeneity is relevant to the plasma membrane.
3. LIQUID–LIQUID PHASE SEPARATION IN MODEL MEMBRANES
3.1. Purified Membranes
In 2001, the first observations of macroscopically phase-separated fluid domains were reported in bilayer membranes reconstituted from cellular extracts (38, 39). These initial observations emphasized similarities between the composition of the coexisting phases in model membranes and the heterogeneity measured in cells by detergent solubilization methods. Soon after, minimal systems of at least three lipid components were shown to form macroscopically phase-separated domains in bilayers: a high melting temperature (Tm) lipid, a low Tm lipid, and a sterol such as cholesterol (40). Phase diagrams describing this macroscopic phase transition have now been mapped by numerous groups using a range of methods and are in good qualitative agreement (41–49). An introduction to reading and interpreting three component phase diagrams is given in Figure 1, and several experimental phase diagrams are presented in Figure 2.
Figure 1.
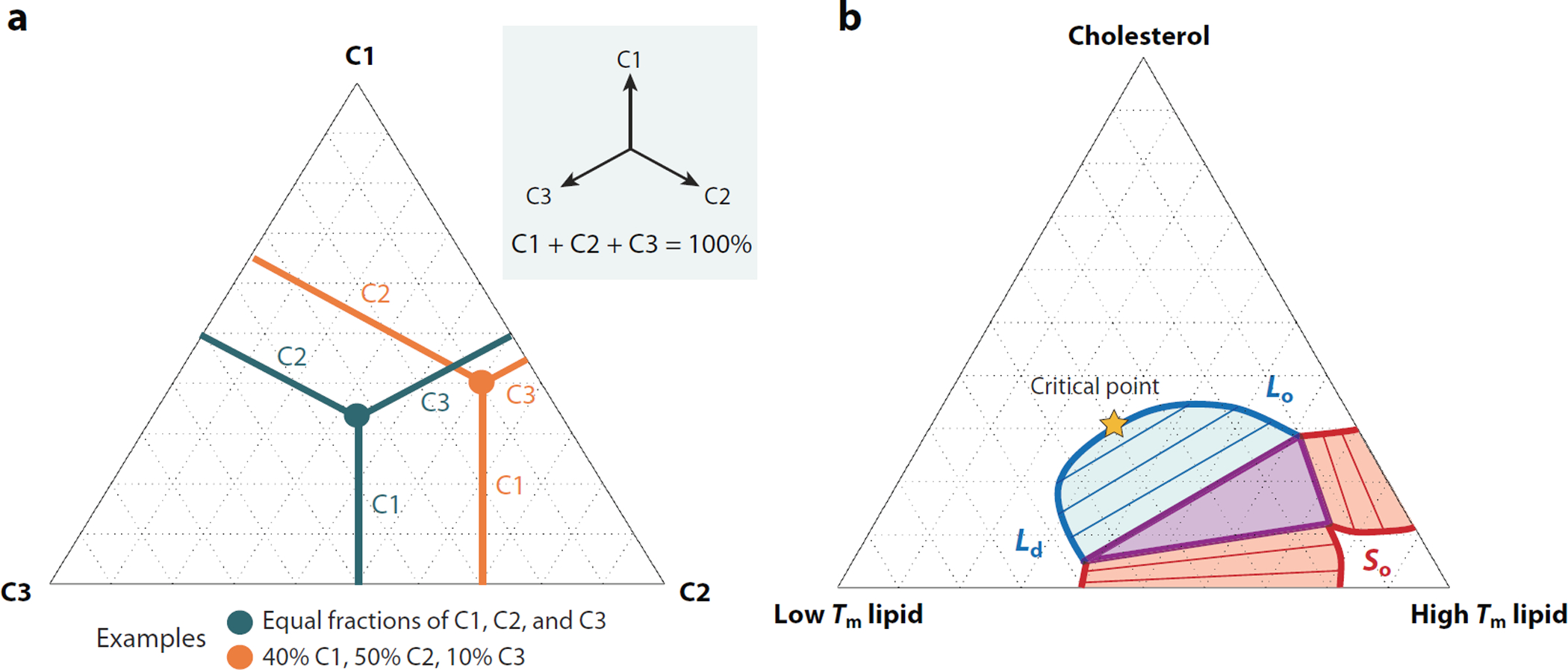
Phase diagram of lipid mixtures. (a) Phase diagrams of three component mixtures (C1, C2, and C3) are conventionally drawn on an equilateral triangle. The three vertices are pure mixtures of each lipid component, points along the edges are binary mixtures, and points within the triangle contain all three components. Compositions can be read by measuring the perpendicular distance to each edge and adding the resulting percentages, which always sum to 100%. Two examples are shown. (b) A qualitative phase diagram for ternary lipid mixtures of high melting temperature (Tm) lipids, low Tm lipids, and cholesterol. Thick red lines indicate the boundaries of liquid–solid (So) coexistence, and the thick blue line represents the boundary of liquid-liquid coexistence of the liquid-disordered (Ld) and liquid-ordered (Lo) phases. Points along this boundary also indicate the composition of coexisting phases, and the specific compositions in coexistence are indicated by blue and red shaded areas within binary coexistence regions. The purple triangle represents compositions that exhibit all three phases in coexistence. The compositions of the three phases are indicated by the three vertices of the triangle. The Ld–Lo coexistence region terminates at a miscibility critical point along the high cholesterol edge that is indicated by an orange star.
Figure 2.
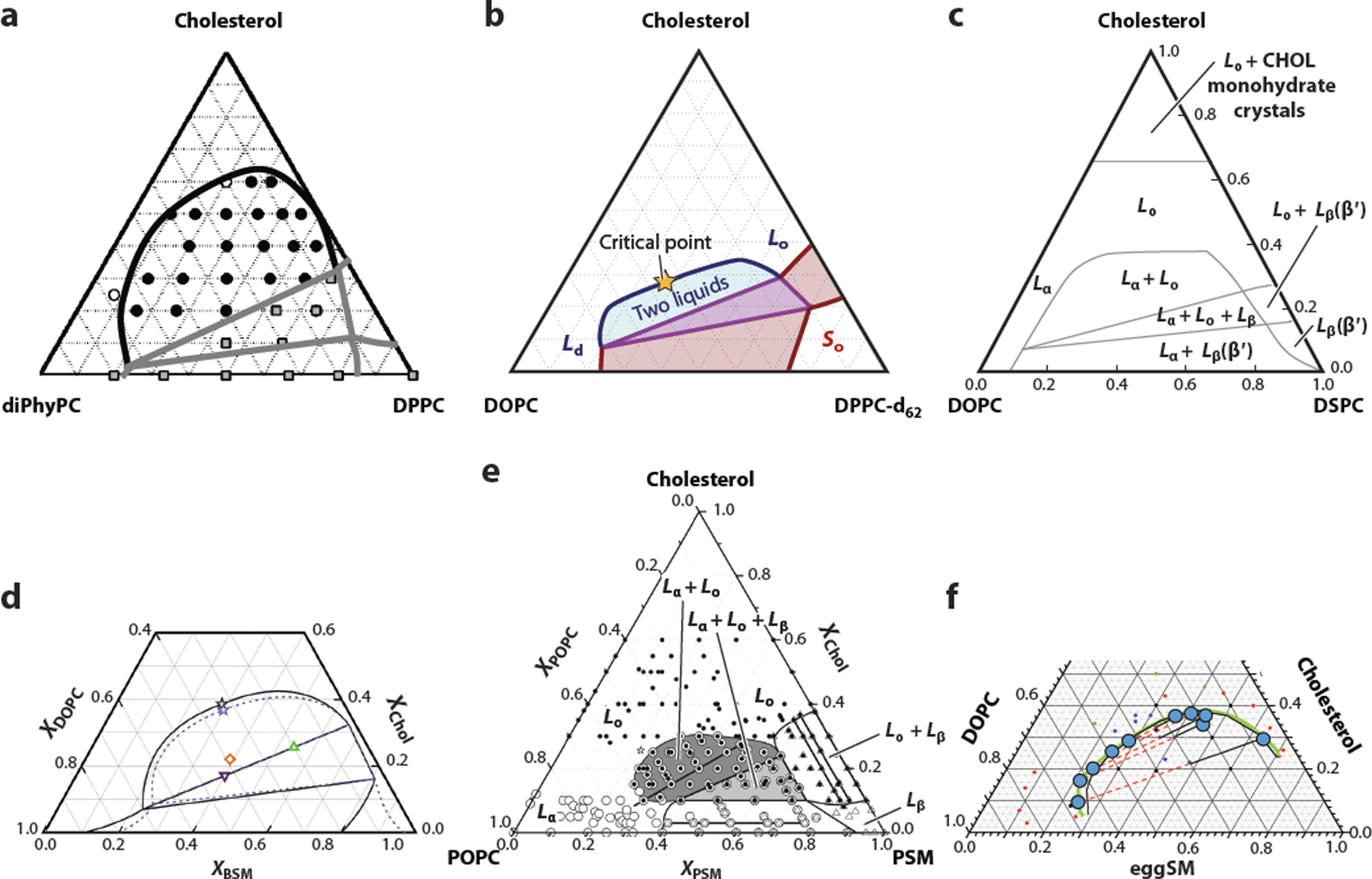
Phase diagrams of ternary lipid mixtures exhibit the same overall topology. Curves on the phase diagrams indicate phase boundaries. In particular, the high cholesterol phase boundary defines the boundary of the liquid-liquid miscibility gap in each case. Sample points and/or deduced tie-line endpoints are indicated in some cases, and some diagrams label single phases as Lα or Ld, Lo, and Lβ or So with abbreviations given below. (a) DPhPC/ DPPC/Chol by fluorescence microscopy at 16°C. Panel a adapted from Reference 41 with permission. (b) DOPC/DPPC/Chol by deuterium NMR spectroscopy. Panel b adapted from Reference 45 with permission. (c) DOPC/DSPC/Chol by fluorescence microscopy and FRET. Panel c adapted from Reference 46 with permission. (d) DPC/PSM/Chol by FRET, neutron scattering, and DSC. Panel d adapted from Reference 47 with permission. (e) POPC/PSM/Chol by EPR spectroscopy. Panel e adapted from Reference 43 with permission. (f) DOPC/eggSM/Chol by atomic force microscopy at 28°C. Panel f adapted from Reference 48 with permission. Abbreviations: BSM, brain sphingomyelin; Chol, cholesterol; DOPC, 1,2-dioleoyl-sn-glycero-3-phosphocholine; DPC, dodecylphosphocholine; DPhPC, diphytanoyl phosphatidylcholine; DPPC, dipalmitoylphosphatidylcholine; DSC, differential scanning calorimetry; DSPC, 1,2-distearoyl-sn-glycero-3-phosphocholine; eggSM, egg sphingomyelin; EPR, electron paramagnetic resonance; FRET, Förster resonance energy transfer; Lα, lamellar liquid crystalline phase; Lβ, a lamellar gen phase; Ld, liquid-disordered phase; Lo, liquid-ordered phase; NMR, nuclear magnetic resonance; POPC,1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine; PSM, palmitoylsphingomyelin; So, solid or gel phase.
A range of experimental (50–56) and simulation (57, 58) approaches using different lipid combinations have produced results consistent with phase diagrams topologically similar to those shown in Figure 1b, and their detailed characteristics have been described in several comprehensive review articles (3, 59, 60). Liquid immiscibility is most often observed at temperatures below the Tm of the high Tm lipid (42, 44–46), although there are exceptions (41). In the typical case, reported phase diagrams have a region of liquid–liquid coexistence, a region of three-phase coexistence (two liquids and a solid), and two regions of liquid–solid coexistence. One of the liquid phases is called the liquid-disordered (Ld) phase, and it resembles the liquid crystalline phase (Lα) of pure phospholipids (61). The second liquid phase, which is called the liquid-ordered (Lo) phase (15), was first characterized in mixtures of saturated phospholipids and cholesterol (8, 16, 18). Most authors refer to both Lo and Ld as Lα phases. The solid phase (So) is often called gel and there are several distinct gel phases observed in purified membranes, including the solid lamellar phases designated Lβ and Lβ′ (62).
The high cholesterol edge of the three-phase triangle is sloped such that the Lo phase contains a higher cholesterol mole fraction than does the Ld phase. This edge of this triangle is also the first tie-line in the liquid–liquid coexistence region. As cholesterol is increased further, the tie-lines run roughly parallel to one another, meaning that the cholesterol concentration increases roughly linearly in both phases. The Lo–Ld coexistence region terminates in a miscibility critical point at which in principle the tie-lines merge to a single point. In practice, this region of the phase diagram is surprisingly flat, meaning that the tie-lines remain long and shorten over a very small range of compositions. As the temperature is lowered, the Lo–Ld immiscibility gap extends to higher concentrations of cholesterol and low Tm lipid, as does the concentration of components at the critical point (41, 45, 48). At a constant temperature, the miscibility gap expands when the Tm of the high Tm component is increased or when the Tm of the low Tm lipid is decreased (41, 42, 44). No macroscopic miscibility gap is observed for some combinations of low and high Tm lipids (42, 44, 63). A closed loop miscibility gap is found when the extremely low Tm lipid diphytanoyl phosphatidylcholine (DPhPC) is used (41), meaning that Lo–Ld coexistence occurs at temperatures above the Tm of the saturated component and that there are two critical points. The phase behavior of the mixed system can depend on more than just the Tm of its components. For example, sphingomyelin (SM) lipids are more effective at establishing coexisting phases than are glycerol-phospholipids with phosphatidylcholine (PC) headgroups (44, 58) even when the main chain transition occurs at similar temperatures for the SM and PC lipids used.
Although the equilibrium thermodynamics description of the macroscopic miscibility transition is now largely accepted, questions remain regarding the thermodynamic basis of the microscopic heterogeneity also routinely observed in membranes containing cholesterol. These structures are frequently reported at temperatures and compositions at which membranes remain uniform on a macroscopic scale using methods that are sensitive to molecular-scale organization such as FRET, fluorescence quenching, or electron spin resonance (ESR) spectroscopy. Membranes can be tuned from a state with macroscopic phase separation to one with microscopic heterogeneity by raising the temperature (64, 65), by titrating in an additional component that disrupts the macroscopic phase transition (see, e.g., References 50, 52, and 66), or by probing different ratios of the same lipid species at a fixed temperature (65). Submicrometer structure is also reported in binary mixtures of saturated lipids and cholesterol (7, 67) and in some ternary membranes that do not exhibit macroscopic Lo–Ld immiscibility at any temperature or lipid ratio (21, 47, 63, 68). Recent experimental developments in purified membranes have begun to probe how leaflet asymmetry impacts phase separation and the presence of submicrometer structure in purified membranes (69, 70).
Experimental observations have motivated numerous theories to explain the presence of microstructure at thermodynamic equilibrium. Critical phenomena provide a possible mechanism to bridge between the macro- and microscales in the form of dynamic fluctuations (45, 71). Other theories enable static, finite-sized domains by including some repulsive mechanism to oppose the formation of macroscopic domains (72–74). These theories too predict domains that span macro- to microscales. Recent experimental work (75) has begun to directly test some of these theories.
3.2. Isolated Membranes
The first observations of macroscopic liquid immiscibility in vesicles blebbed directly from living cells came in 2007 (2), and these vesicles were named giant plasma membrane vesicles (GPMVs) due to their close resemblance to the giant unilamellar vesicles (GUVs) used widely in fluorescence microscopy investigations of purified lipid mixtures. Earlier work using a similar vesicle preparation had characterized their lipid and protein content by mass spectrometry (76) and had observed heterogeneity using ESR (30), a spectroscopic method that can detect heterogeneity on the molecular scale. Baumgart et al. (2) detected the sorting of fluorescent lipid analogs and fluorescently tagged proteins with respect to phases in GPMVs at temperatures well below those at which cells were grown, leading the researchers to conclude that this phase transition was not relevant for cells under normal growth conditions.
Soon after, other methods emerged to isolate plasma membranes from cells, all of which could yield coexisting liquid phases (77, 78), although the conditions needed to achieve phase separation differed depending on the method used. A subsequent study found correlations between biochemically defined detergent-resistant membranes and the Lo phase detected in GPMVs (79) and that the surface fraction of Lo phase at low temperature was altered by acute treatments to manipulate cholesterol levels in vesicles (79, 80). In all cases, miscibility transition temperatures (Tmix) remained well below growth temperatures in isolated cells, emphasizing that such macroscopic domains were not likely to form under physiological conditions. A possible explanation came in 2008 when it was shown that freshly isolated GPMVs exhibited hallmarks of criticality, placing them close to a room temperature miscibility critical point (81). Over time, additional studies have documented how GPMV phase behavior is impacted by growth conditions in cells (82–84) and differentiation into other cell types (85, 86), and lipidomics analysis has begun to characterize the vast compositional complexity of these membranes (82, 84, 87).
While there are many similarities between the phase behavior observed in GPMVs and purified model membranes, there are also key differences (88). The coexisting phases detected in GPMVs differ in their physical properties from their purified membrane counterparts. The viscosity and hydration of phases is more similar in GPMVs compared to that in GUVs, as measured through the diffusion of membrane components or using order-sensing fluorophores that report on local hydration within the hydrophobic region of the membrane (89). These different physical properties can be sensed by incorporated proteins. Some transmembrane proteins, particularly those with palmitoylated cysteines, are observed to partition into the Lo phase in GPMVs, whereas few are reported to partition into the Lo phase in purified membranes (90).
GPMVs are model membranes that differ from intact plasma membranes in important ways. Plasma membranes exist in close association to the actin cytoskeletal cortex, while GPMVs are missing polymerized cytoskeletal components and tend to be depleted in proteins that associate with actin (91). Notably, the cell plasma membrane does not macroscopically phase separate even under conditions that cause GPMVs to phase separate, or in fact under any known conditions, even when phase-separated GPMVs remain attached to an intact cell membrane (92). GPMVs are depleted of phosphatidylinositol 4,5-bisphosphate [PI(4,5)P2] (93), which typically makes up several mole percent of the inner plasma membrane leaflet in intact cells (94). A recent report demonstrates that isolated GPMVs are frequently permeable to large hydrophilic markers (95), indicating that their membranes contain long-lived defects. While cell membranes are asymmetric in their lipid and protein composition, at least some of this asymmetry is lost in the GPMV generation and isolation process (2, 93). The interpretation of GPMV experiments is also complicated by their sensitivity to methodological choices. For example, the most common method used to prepare GPMVs involves incubating cells with a low concentration of formaldehyde and a reducing agent, and the choice of the reducing agent can greatly impact the Tmix and physical properties of phases of the resulting GPMVs (78, 96). This is due, at least in part, to the ability of some reducing agents to modify membrane proteins and lipids.
4. PHASE-SEPARATED DOMAINS ARE RELATED TO BUT DIFFERENT FROM RAFT HETEROGENEITY IN INTACT CELLS
Phases are robust macroscopic entities with well-defined compositions and, with the exception of in the yeast vacuole (97, 98), structures resembling liquid–liquid phase separation are not observed in intact cells. Instead, the vast majority of membrane domains in cells are microscopic and require significant perturbations to be visualized or isolated. In line with this, several recent studies have directly concluded that there is no evidence of a miscibility phase transition in intact cells when cells are examined through the lens of several different experimental observables (99, 100). Nonetheless, a large number of experimental studies provide strong evidence that the heterogeneity reported in intact cells is closely related to the macroscopic phase separation observed in GPMVs. Two excellent recent reviews (101, 102) discuss many of these findings, as well as some exceptions. We highlight several lines of evidence below.
In many cases, proteins that are associated with live-cell heterogeneity or with detergent-resistant membranes are also found to partition into the Lo -like phase of GPMVs. In particular, single-pass transmembrane and peripheral proteins containing palmitoylations are more likely to be found in detergent-resistant membranes and to partition into the Lo phase in GPMVs, while proteins containing branched and unsaturated geranylgeranyl or prenyl groups tend to be solubilized by detergents and to partition into the Ld phase in GPMVs. Recent work has begun to extend this to multipass proteins that can accommodate more complex protein–lipid interactions (103–105). Protein partitioning is likely dictated by the identity of lipids that solvate transmembrane proteins in complex membranes (106, 107)
Cells actively tune their plasma membrane Tmix in response to changes in growth conditions, and conditions that change Tmix lead to different phenotypes. For example, experiments indicate that cells in culture actively tune the Tmix of their membrane to be a fixed temperature below their growth temperature (82) and that the Tmix decreases in cells under conditions that inhibit cell growth (83). Other studies have documented that acute treatments with lipophilic small molecules or dietary lipids that alter Tmix correlate with changes in signaling outcomes, cellular differentiation, and even the general anesthetic response (85, 108, 109). While these correlations with Tmix may be a result of a mutual correlation with an unrelated quantity, the accumulated evidence suggests that maintaining Tmix plays important roles in cellular processes.
In model membranes, Tmix predicts structure observed in single-phase membranes. While model membranes appear homogeneous at the macroscopic scale above Tmix, the value of Tmix can predict the presence of structure at smaller scales or under conditions in which membranes are perturbed by clustering one component. A recent report (96) documents the temperature dependence of both macroscopic phase behavior and microscopic heterogeneity in GPMVs under a range of perturbation conditions. This work found that the microscopic heterogeneity in GPMVs at elevated temperature was highly correlated with their macroscopic transition temperature Tmix, suggesting that the same could be true in intact cells. An older study found that macroscopic membrane domains enriched in phase-marking probes could be stabilized even well above Tmix (80). These and other studies have led to the idea that the value of Tmix is a measure of the stability of raft-like domains under physiological conditions, even though cells do not appear to experience phase separation directly.
Theoretical arguments support that coupling to an intact cytoskeletal network disrupts the macroscopic phase transition. It is well established that quenched disorder abolishes first-order phase transitions in two-dimensional (2D) systems such as membranes (110). The cortical actin cytoskeleton, which is linked to the plasma membrane through a system of adapter proteins, is likely to play the role of quenched disorder in the intact plasma membrane (92, 111). Experimental studies in model membranes support the main conclusion of this theory, which is that a broadly distributed cytoskeletal network disrupts macroscopic domains while stabilizing small-scale structure (112). Direct tests in intact cells have yet to be reported, but broad evidence exists for important connections between cortical actin and raft heterogeneity (113–117).
The evidence for phase-driven partitioning within intact cells is increasing. Recent work using single-molecule fluorescence methods is beginning to draw more direct connections between phases in vesicles and domains in cells (118, 119). These studies monitor the recruitment and exclusion of probes with respect to clustered proteins in intact cells and have found that probe concentration in clusters mirrors partitioning with respect to phase-separated domains in model membranes, although typically with a smaller magnitude.
To explain these experimental developments, several theoretical avenues have been explored, with a goal of explaining the lack of a macroscopic phase separation in intact cells. Any complete biophysical model of membrane heterogeneity must first account for this fact, and therefore must be more complex than the simple phase-separation picture. These explanations include coupling to quenched disorder in the form of the cytoskeleton (92, 111); coupling to curvature in a way that produces a microemulsion (72, 74, 120); and nonequilibrium suppression of domain growth, for example, by active lipid transport or the remodeling of membrane-coupled actin structures (121, 122). Many of these models, including the critical phenomena that are discussed in the next section, are closely related to the canonical phase-separation picture (123).
5. CRITICALITY AND ITS CONNECTION TO THE PLASMA MEMBRANE
Equilibrium critical phenomena are special relationships between the thermodynamic properties of a system that emerge due to a diverging correlation length of the system in the vicinity of a critical point. Key physical quantities such as the heat capacity, susceptibility, and interfacial energy between domains also exhibit specific behaviors as the critical point is approached. These properties of critical systems are universal, meaning that the dominant behavior is governed by a small number of effective parameters regardless of the complicated details of microscopic interactions (Figure 3). The physics of criticality has been covered in detail in many textbooks (124–127). A useful introduction for biophysicists is given in Reference 128.
Figure 3.
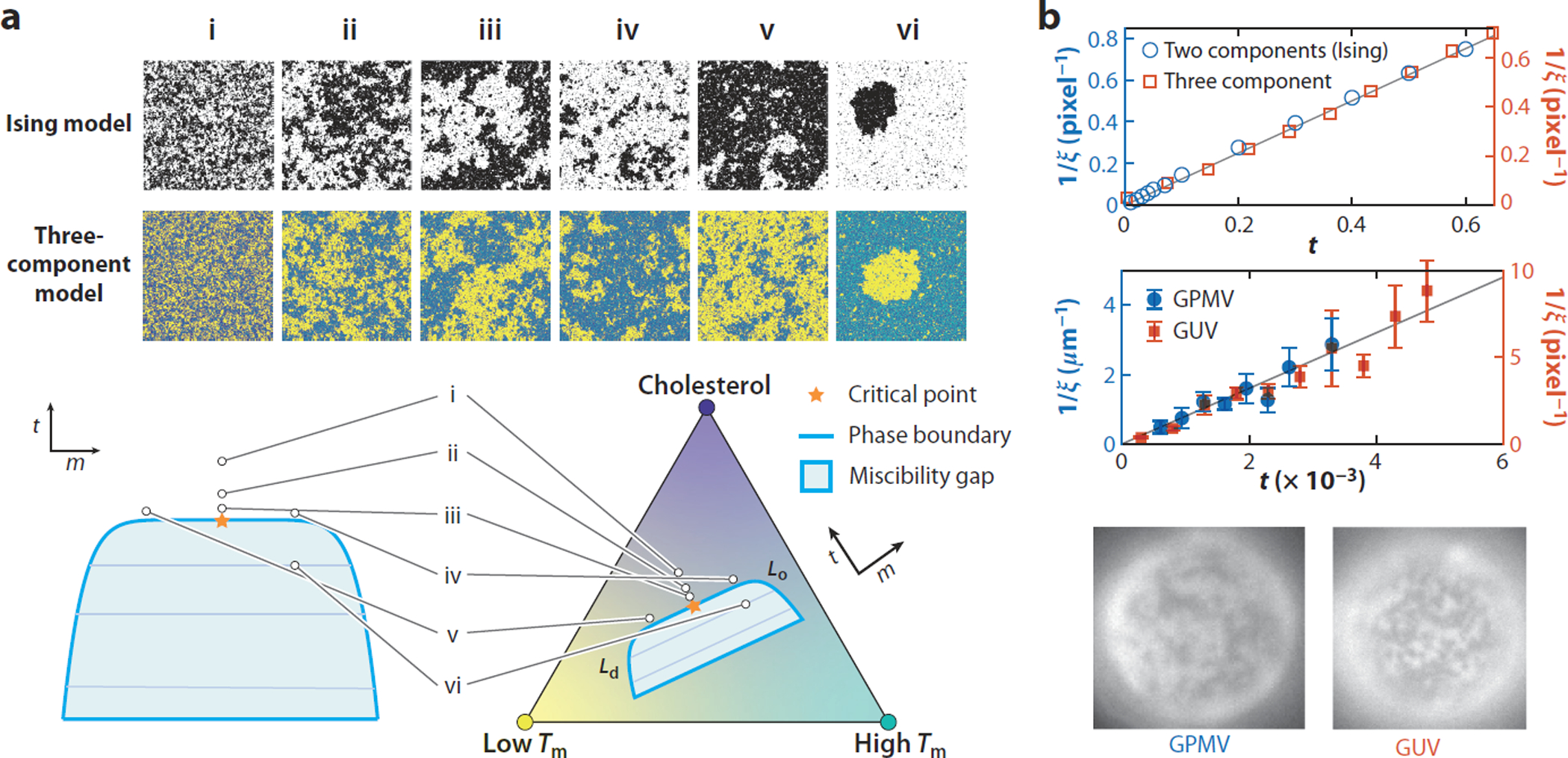
Critical systems exhibit universal features that extend beyond the two-phase region. (a) Simulation snapshots of two lattice models. (Top) An Ising model at various reduced temperatures t and compositions m, and (middle) a three-component model with interactions designed to reproduce typical phase diagrams of ternary mixtures of Cholesterol (dark blue), a low melting temperature (Tm) lipid (yellow) and a high Tm lipid (cyan) at fixed temperature and various compositions. (Bottom) Model parameters used for (i)-(vi) in each model are indicated on the respective phase diagram. Compositions of the three-component model are chosen so that the effective reduced temperature t and effective magnetization m of the model match the t and m of the corresponding Ising model. Miscibility gaps, tie-lines, and the directions of t and m are shown on each schematic phase diagram. The regions that correspond to liquid-ordered (Lo) and liquid-disordered (Ld) mixtures in ternary lipid mixtures are also indicated. (b) Measurements of the divergence of correlation length ξ versus t as the critical point is approached from the one-phase region. (Top) In the two models from panel a. For the three-component model, t is the cholesterol (Chol) content normalized by the Chol content at the critical point. (Middle) In GUVs and giant plasma membrane vesicles (GPMVs) with varying temperature. Data from Reference 128. In each case, the expected 2D Ising power law ξ ∝ t−1 is observed although the proportionality constant differs, resulting in each data set being plotted with a different y-axis scale. (Bottom) Representative GUV and GPMV images near the critical point.
5.1. Historical Foundations of Critical Phenomena
The physics of criticality has developed over the last 200 years, beginning with the observation of a critical temperature for several liquids above which there is no liquid–gas transition (129). The van der Waals equation of state first published in 1873 was the first model that featured a critical point with this property (130). In 1918, Ornstein & Zernicke (131) formalized the study of spatial fluctuations in liquid–gas systems, a key development. In the first half of the twentieth century, precise measurements of near-critical liquid–gas systems (132) as well as magnetic systems (133) indicated that classical equations of state were inadequate to explain the near-critical region of phase space, for example, that the shape of the liquid–gas coexistence curve is qualitatively different than that predicted by the van der Waals equation. In 1944, Lars Onsager (134) exactly solved the 2D Ising model, which allowed for the study of critical exponents in that system. In 1952, Lee & Yang (135) observed that phase transitions correspond to nonanalyticities of the partition function in the thermodynamic limit. Soon after, Widom (136) and others proposed power-law scaling for various quantities at critical points and derived relationships between different critical exponents from that hypothesis. Real-space [Kadanoff (137)] and momentum-space [Wilson (138)] renormalization group methods explained the emergence of these power laws and provided avenues for computing approximate values for the critical exponents in general systems,. They also set a uniform framework for other conceptual advances. A broader class of nonequilibrium critical points can also be defined, generalizing this equilibrium concept. Biological nonequilibrium critical points have received considerable attention in recent years; for a review see, e.g., Mora & Bialek (139).
5.2. Unique Properties of Critical Systems
Coexistence regions generically terminate in a critical point except in special circumstances, and the Lo–Ld miscibility of purified and isolated membranes is no exception. On the phase triangle shown in Figure 1b, the critical point occurs on the high cholesterol edge of the Lo–Ld miscibility gap. Initial evidence of critical behavior in membranes came from nuclear magnetic resonance (NMR) studies of multilamellar vesicles, which detected enhanced line broadening in the vicinity of known critical points (45). This was attributed to the diffusion-mediated exchange of lipids between fluctuations with submicrometer dimensions. Later work directly visualized micron-sized critical fluctuations (above the critical temperature Tc) and fluctuating phases (below Tc) in GUVs of purified lipids, providing evidence that membranes belong to the 2D Ising model universality class, meaning that fluctuations exhibited a temperature dependence that is universal to 2D systems with a 1D order parameter (71). Subsequent measurements confirmed this observation in supported membranes by atomic force microscopy (48) and probed the dynamics of critical membranes (140). At the same time, isolated GPMVs were also discovered to exhibit critical behaviors in the vicinity of a room temperature critical point (81). Again, fluctuations were consistent with the 2D Ising model universality class. These observations and their implications have been previously reviewed in greater detail (128).
One of the key features of a critical point is that its fingerprints extend well beyond the phase transition itself (Figure 3). An important parameter is t, the difference between the temperature of the system and the Tc, normalized by Tc in units of Kelvin. The correlation length ξ, or characteristic size of critical composition fluctuations, is predicted to vary as ξ(t) = ξ0/t, where ξ0 is a parameter with dimensions close to the size of molecules in the system and in membranes was measured to be roughly 1 nm. Note that as T → Tc, t → 0, so that the correlation length becomes infinite. If extrapolating this relationship using a room temperature critical point (Tc = 22°C = 295 K), then 20-nm-sized fluctuations are expected at 37°C, which corresponds to t = 0.05. This prediction is in good agreement with recent experimental work probing heterogeneity in GPMVs by FRET, which detects evidence for larger than 10-nm structures in GPMVs over this same temperature range (96). Similar observations have also been made in purified model membranes (141). Note that t need not be a physical temperature. Instead, it is any trajectory in the phase diagram that runs perpendicular to tie-lines close to the critical point. Thus, while some Ising model images of Figure 3a are obtained by varying temperature in the model, the corresponding three-component lattice model images are obtained by varying composition at fixed temperature, as indicated in the phase diagram.
Another physical property that can extend well beyond the phase transition itself is the susceptibility (χ). The susceptibility measures how large a local composition difference arises from a local force applied to components of one of the phases, e.g., by clustering components that prefer Lo lipids. In other words, in a highly susceptible membrane, a domain of distinct local composition can be stabilized by clustering only a small subset of components or by weakly biasing the concentration of many components that have the same order preference. In the Ising universality class, χ varies with t as χ ∝ t−7/4. This too has experimental support in vesicles, where robust domains are stabilized well above Tmix by organizing a small subset of components via an actin network or a streptavidin crystal that partially decorates a vesicle surface (114, 142) or by adhesion to a supported membrane (80).
Direct theoretical predictions of critical phenomena such as the scaling of the correlation length and the magnitude of the susceptibility are quite useful for predicting the consequences of perturbations to membrane heterogeneity, but the theory quickly becomes intractable when coupled to more complex biological phenomena. Statistical mechanical lattice models based on the Ising model can be useful in this situation. These models typically contain only two components (often called up and down spins because of the Ising model’s origin as a model of magnetic systems) positioned on a lattice, where the components at the lattice sites are either allowed to change identity (such that the composition or magnetization can vary) or are allowed to exchange with other sites on the lattice (such that the composition remains fixed). Universality guarantees that, as long as the system is close to the critical point, the Ising model captures the relevant mesoscopic heterogeneity of the membrane for appropriate choices of the Ising reduced temperature t and magnetization m (Figure 3). That is, the Ising model accurately recapitulates the thermodynamics of the effective Lo order parameter at length scales beyond a few lipid diameters, despite the extreme simplicity of the microscopic interaction in the model, which is a simple nearest-neighbor interaction potential. As a result, when a biological system is coupled to the Lo order parameter, an Ising model modified to include this coupling is expected to reflect the relevant biophysical phenomena. Past work has used this approach to model the coupling of fluctuations to cortical actin (92), to explain changes in phosphorylation steady states upon clustering of a component for various values of t and m (118, 143), and to predict how proximity to the critical point affects conformational state equilibria of proteins whose boundaries are sensitive to lipid order (144).
5.3. Evidence for Criticality Playing a Role in Cells
If intact plasma membranes exhibit heterogeneity similar to that observed in GPMVs, this could easily be relevant to the biological function of membrane proteins. An important line of evidence that criticality plays a role in biological function has come from the tuning of the GPMV critical point. For a system to be near a critical point, two parameters must be tuned, those corresponding to t (temperature) and m (composition) of the (fixed-composition) Ising model. In the extremely large space of lipid mixtures of varying composition, there are many critical points—an n − 2-dimensional manifold in the n-dimensional space. However, there is no generic reason that tuning the concentration of any given lipid corresponds to tuning just t, or just m, or neither; general perturbations affect both t and m. Thus, it is somewhat surprising that the cell arrives near a critical point if it constructs its membranes without explicitly or implicitly tuning to the critical point, given that lipid composition is modulated by a wide variety of perturbations. In other words, the fact that plasma membrane composition is near-critical is unlikely to simply be a coincidence. Furthermore, at least in certain cases, eukaryotic cells adapt to perturbations in ways that preserve the distance to the critical point and corresponding physical properties. Zebrafish cells cultured at a range of temperatures from 20°C to 32°C produce GPMVs with correspondingly altered Tc values (82).
The concept of high susceptibility near a critical point is useful in interpreting recent single-molecule and super-resolution studies documenting the partitioning of phase-marking probes to protein clusters in intact cells (118, 119, 143). In these studies, antibodies are used to cross-link a membrane component that prefers either the Lo or Ld phase, then the differential partitioning of probes is monitored with respect to these domains. When proteins that prefer the Lo phase are clustered, probes that also prefer Lo tend to be recruited, and those that prefer Ld tend to be excluded. In contrast, when proteins that prefer the Ld phase are clustered, probes that prefer Lo are excluded, and probes that prefer Ld are recruited. Similar to experiments with vesicles adhered to supported membranes (80), the act of clustering a protein or peptide biases the concentration of many components in ways that can be detected when membranes have high susceptibility. In some cases, the extent of probe partitioning approaches that observed in phase-separated vesicles (143), while in others the sorting of components is much weaker (118). These differences could arise from differences in either the coupling of protein clusters to membranes or the susceptibility of the membrane in different experimental systems.
6. CRITICALITY AS IT RELATES TO BIOLOGICAL FUNCTION
Since the inception of the raft hypothesis, the study of the functional relevance of membrane domains has focused on their ability to compartmentalize protein and lipid components so that they can optimally function within biochemical networks (1). Critical phenomena are in many ways consistent with this framework. A supercritical membrane contains domains resembling ordered and disordered phases, and components that partition with the same phase colocalize within these domains. The fluctuations are small and dynamic, consistent with evolving descriptions of rafts over the decades (145–148), but fluctuations alone are not an effective means to strongly colocalize or confine membrane components. This new reality requires us to move beyond the simple mechanisms proposed in the early raft literature to propose and test mechanisms that exploit the unique material properties of critical systems. Several proposals are highlighted in Figure 4 and described in this section.
Figure 4.
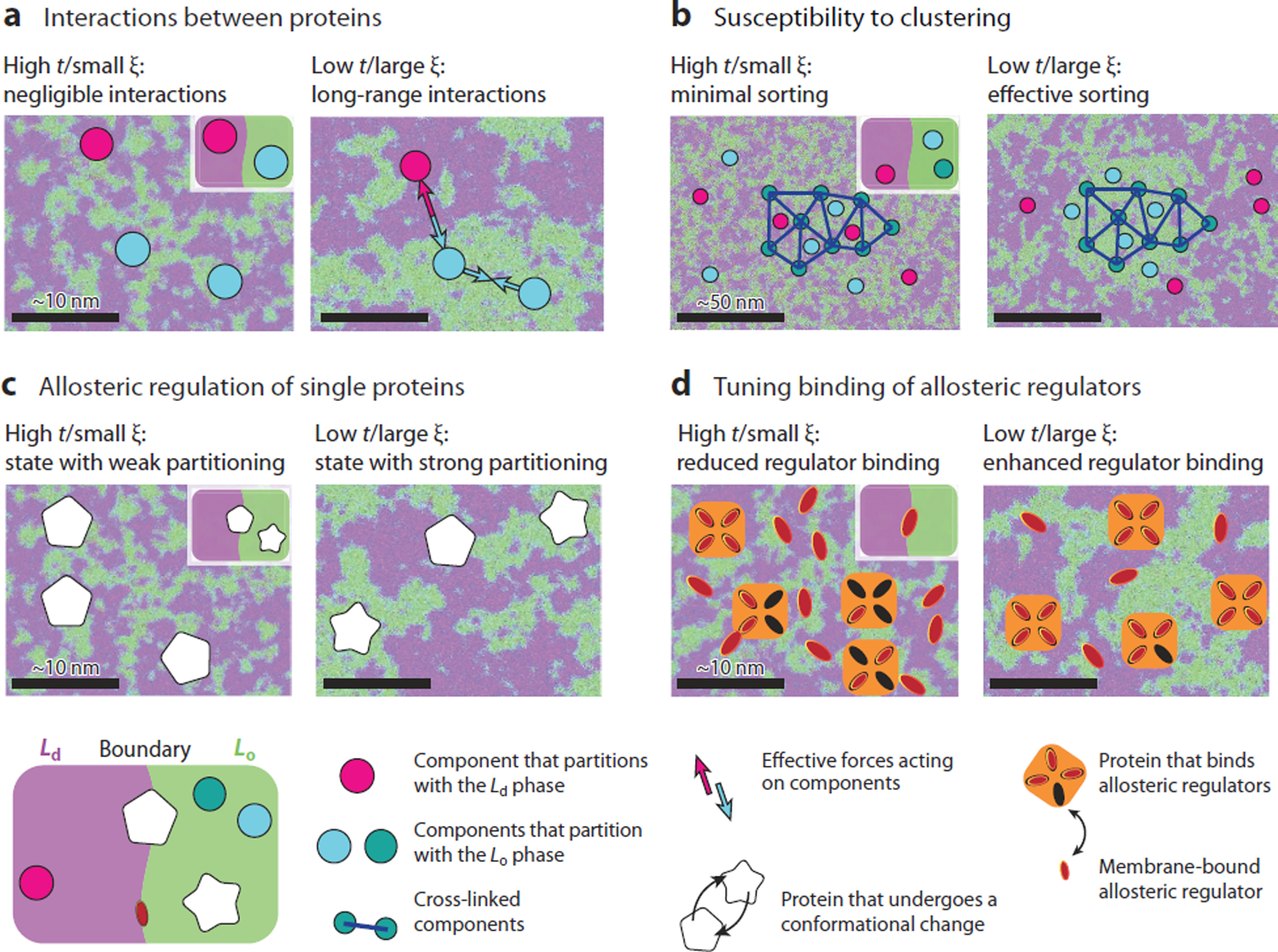
Four functional mechanisms primarily driven by liquid-ordered (Lo)–liquid-disordered (Ld) phase partitioning in a near-critical membrane. Membrane proteins and other components are characterized by how they partition into Lo–Ld domains, as shown here schematically. The combination of the partitioning of various components confers to the systems particular properties that can be used to drive biological function. a) Critical Casimir forces yield an effective attraction between components with like order preference (blue-blue circles), and an effective repulsion between components that prefer opposite lipid order (blue-magenta circles). The strength of these interactions increases rapidly when t is reduced andξ becomes large. (b) Clustering a protein that prefers Lo lipids (green circle) induces a distinct Lo domain due to the high susceptibility of the membrane near the critical point. As a result, other Lo-preferring proteins (blue circles) are recruited to the cluster, and Ld -preferring proteins (magenta circles) are excluded. Similarly, an Ld domain can be stabilized by clustering an Ld-preferring component. (c) The white shape is a large protein with two conformations, pentagon or star. One conformation (pentagon) has no order preference, and the other (star) prefers an Lo environment. The star conformation becomes more likely as t is decreased, because there are large patches of Lo membrane that satisfy its preferred boundary condition. (d) Changes in t also result in changes in the chemical potentials of some membrane components, including components (red ovals) that can bind to a membrane protein (orange squares) as an allosteric modulator. Therefore, a change in t can induce differences in binding-site occupancy and the resulting distribution of protein states.
6.1. Interactions Between Proteins
Composition fluctuations can mediate forces between membrane proteins via a process termed critical Casimir forces that was first described between conducting plates in vacuum (149). In essence, proteins sense an effective attractive potential if they partition into the same phase because coming into close proximity allows them to share the same local lipids, as shown in Figure 4a (150, 151). In contrast, proteins that prefer different environments sense an effective repulsion, because there is an energetic cost to mixing their local environments. These potentials are weak (on the order of the thermal energy kBT) but have a range given by the correlation length and the size of the protein or protein cluster. This range is long compared to other interaction modes experienced by membrane proteins such as curvature, electrostatics, and van der Waals potentials. Note that repulsion of components that prefer different phases has a larger magnitude than does attraction between components that prefer the same phase. Casimir forces may contribute to the stability of protein assemblies, including phase-separated polymer droplets that assemble on membranes. Casimir forces are also expected to alter biochemistry occurring at the membrane by increasing or decreasing the rates at which the proteins encounter one another. It is tempting to speculate that one functional role of palmitoylation, the posttranslational modification that places a saturated acyl chain on proteins, is to tune the magnitude of these Casimir forces for specific protein species.
6.2. Susceptibility to Receptor Clustering
The high susceptibility of a critical membrane provides a means for the cell to sense the redistribution of a subset of membrane constituents by an external force (Figure 4b). Clustering a membrane protein that prefers one phase biases the local lipid composition in proportion to the heightened susceptibility of the system. This effect can impact biochemical reactions that take place within these clusters, drastically altering the chemical steady state of the system. We have studied this effect in the context of B cell–receptor (BCR) signaling (118, 143). Here, the act of clustering the BCR or another ordered membrane component by an extracellular ligand stabilizes an ordered domain that contains a higher local concentration of kinase and a lower concentration of phosphatase than the membrane as a whole. This establishes a local environment that favors receptor phosphorylation and activation. In principle, this class of activation mechanism could contribute to a wide range of signaling pathways that are initiated by receptor clustering at the cell surface. This type of mechanism could also play a role in establishing biochemical environments in membrane regions where components are organized by processes occurring at the inner plasma membrane leaflet, such as at junctions between the endoplasmic reticulum and plasma membrane (152), or at sites where scaffolding adaptor proteins are anchored to membranes, such as in neuronal synapses (153).
6.3. Allosteric Regulation of Single Proteins
Beyond contributing to the organization of proteins, the size and stability of fluctuations in the membrane can also impact the functioning of single proteins. One mechanism that has been proposed requires that two conformational states of the protein in question have different boundary lipid preferences for Lo or Ld lipids (144). If that is the case, then a change in Tc differentially affects the free energies of the two conformational states. Roughly, a spatially extended lipid preference carries a free energy cost that decreases near Tc, so that a conformational state with a strong order preference becomes more probable when fluctuations are large compared to the protein diameter. This model was proposed to explain the striking correlations between Tc-altering effects and the anesthetic or anesthetic-reversing potencies of a wide range of treatments, including short- and long-chain n-alcohols and hydrostatic pressure (108, 109).
6.4. Tuning Binding of Allosteric Regulators
The chemical potential μ of a component is the thermodynamic parameter that controls the proclivity of that component to enter or exit a system, for example, its availability to bind in a binding pocket. Formally, the chemical potential is the increment of free energy to move one particle into the system from a particle bath. Equivalently, the chemical activity α ∝ eμ/kT can be used. In an ideal gas or ideal dilute solution, the chemical potential has a simple logarithmic relationship to concentration and is linear in temperature (so that activity is proportional to concentration) and insensitive to the concentrations of other components (154). However, a near-critical mixture is far from ideal: The critical point is precisely where weak cooperative interactions between the many components lead to strong effects (124). Therefore, we expect strong relationships between the chemical potentials of different components, especially when those components modulate Tc.
Many transmembrane proteins have been shown to be modulated by binding to membrane components, prominently including cholesterol (154, 155) and phosphatidylinositol lipids (156) as well as many other signaling lipids (157). If the chemical potential of some of these components is strongly modulated by the concentration changes of other components, binding-site occupancy also varies, and we expect to see changes in protein functions that depend on the binding of those components. Recent work by Ayuyan & Cohen (154) has resulted in the development of sensitive methods for measuring and controlling the chemical potential of cholesterol in the plasma membrane and found that cholesterol chemical potential varies by ~2 kBT in different physiologically relevant cellular conditions. That amount is certainly adequate to induce substantial changes in binding-site occupancy. Work remains to be done to explore if these differences can be attributed in any way to the critical phase transition, but we can speculate that perturbations that change membrane criticality may act indirectly by impacting the activity of membrane components.
6.5. Criticality Coupled to Other Processes Describes a Broad Array of Raft Phenomena
The Ising universal critical phenomena are a good start for understanding membrane heterogeneity, but they are also clearly insufficient to explain all phenomena. As stated in Section 5, critical phenomena alone are not expected to give rise to regions of tight clustering or confinement of proteins and lipids, as is sometimes attributed to membrane domains. This said, the local membrane environment could impact the conformational states sampled by membrane proteins in ways that facilitate binding through stronger protein-binding sites. This type of synergistic effect could underlie a range of cholesterol-dependent processes observed at the plasma membrane, including, for example, the transient pinning observed in studies of membrane protein and lipid dynamics (158).
Another example in which there is potential for synergy between criticality and other organizing principles relates to the active composite model proposed by Rao & Mayor (159). This model posits that the plasma membrane interacts with a heterogeneous cortical actin network composed of both active and passive components. The passive components largely resemble quenched disorder, as described in Section 3. The active component is composed of motor-driven short actin filaments that can actively drive certain membrane proteins and lipids into close proximity, coupling across membrane leaflets. Considering this model in a critical membrane provides a simple means to correlate domain structure across leaflets without requiring strong interactions such as interdigitation, because the cooperativity inherent in a phase transition can amplify weak couplings that may be present. Moreover, the high susceptibility of a critical membrane allows it to robustly remodel when external forces are applied, including those originating from the actin cortex.
More broadly, we envision that plasma membrane criticality is only one of several organizing principles that contribute to plasma membrane functions. Interactions mediated by curvature and electrostatics may be superimposed over those mediated via criticality to define the plasma membrane interactome. There also may be interesting cross talk between these various interaction modes. For example, studies have shown that the sorting of lipids into curved membranes can be mediated by the binding of curvature-sensing proteins that have preferences for one membrane phase (160). Proteins and peptides can also organize lipids through electrostatics, which in turn can stabilize domains impacted by the fluctuations of Lo and Ld lipids. The broader implications of this potential cross talk are largely unexplored and could give rise to qualitatively new phenomena accessible to cell membranes (161).
7. CONCLUDING REMARKS
While it has long been appreciated that plasma membrane lipids are capable of intriguing, nonideal behaviors, much of the past literature is clouded by imperfect methods and an incomplete conceptual framework with which to conceptualize experimental observations. This backdrop led to controversial and often unphysical descriptions of lipid rafts. The past decade or so has brought key advances, including membrane isolations that largely preserve plasma membrane protein and lipid content and super-resolution imaging methods that do not suffer from the same pitfalls that plagued early raft research. Alongside these advances, the membrane community has begun to appreciate the rich phenomena that naturally occur near miscibility critical points, many of which exhibit strong parallels with long-standing observations in both the membrane biophysics and membrane biology literatures. Moving forward, the challenges will be to isolate these effects to enable a definitive measurement of the role of criticality in cell membranes and to explore how these immiscibility-mediated interactions work alongside other physical and biochemical organizing principles to contribute to the rich array of biological functions at the cell surface.
ACKNOWLEDGEMENTS:
We thank Sarah Shelby and Ben Machta for helpful discussions, and Aurelia Honerkamp-Smith for supplying the raw data used to generate parts of Figure 3. The authors’ work is funded by grants from the National Science Foundation (MCB1552439 and DMR1905600) and the National Institutes of Health (R01GM129347 and R01GM110052).
Footnotes
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings that might be perceived as affecting the objectivity of this review.
LITERATURE CITED
- 1.Simons K, Ikonen E. 1997. Functional rafts in cell membranes. Nature 387(6633):569–72 [DOI] [PubMed] [Google Scholar]
- 2.Baumgart T, Hammond AT, Sengupta P, Hess ST, Holowka DA, et al. 2007. Large-scale fluid/fluid phase separation of proteins and lipids in giant plasma membrane vesicles. PNAS 104(9):3165–70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Veatch SL, Keller SL. 2005. Seeing spots: complex phase behavior in simple membranes. Biochim. Biophys. Acta Mol. Cell Res 1746(3):172–85 [DOI] [PubMed] [Google Scholar]
- 4.Oldfield E, Chapman D. 1971. Effects of cholesterol and cholesterol derivatives on hydrocarbon chain mobility in lipids. Biochem. Biophys. Res. Commun 43(3):610–16 [DOI] [PubMed] [Google Scholar]
- 5.Oldfield E, Chapman D. 1972. Dynamics of lipids in membranes: heterogeneity and the role of cholesterol. FEBS Lett 23(3):285–97 [DOI] [PubMed] [Google Scholar]
- 6.Engelman DM, Rothman JE. 1972. The planar organization of lecithin-cholesterol bilayers. J. Biol. Chem 247(11):3694–97 [PubMed] [Google Scholar]
- 7.Shimshick EJ, McConnell HM. 1973. Lateral phase separations in binary mixtures of cholesterol and phospholipids. Biochem. Biophys. Res. Commun 53(2):446–51 [DOI] [PubMed] [Google Scholar]
- 8.Chapman D, Penkett SA. 1966. Nuclear magnetic resonance spectroscopic studies of the interaction of phospholipids with cholesterol. Nature 211(5055):1304–5 [DOI] [PubMed] [Google Scholar]
- 9.Ladbrooke BD, Williams RM, Chapman D. 1968. Studies on lecithin-cholesterol-water interactions by differential scanning calorimetry and X-ray diffraction. Biochim. Biophys. Acta Biomembr 150(3):333–40 [DOI] [PubMed] [Google Scholar]
- 10.Opella SJ, Yesinowski JP, Waugh JS. 1976. Nuclear magnetic resonance description of molecular motion and phase separations of cholesterol in lecithin dispersions. PNAS 73(11):3812–15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lentz BR, Barrow DA, Hoechli M. 1980. Cholesterol-phosphatidylcholine interactions in multilamellar vesicles. Biochemistry 19(9):1943–54 [DOI] [PubMed] [Google Scholar]
- 12.Rubenstein JLR, Owicki JC, McConnell HM. 1980. Dynamic properties of binary mixtures of phosphatidylcholines and cholesterol. Biochemistry 19(3):569–73 [DOI] [PubMed] [Google Scholar]
- 13.Recktenwald DJ, McConnell HM. 1981. Phase equilibriums in binary mixtures of phosphatidylcholine and cholesterol. Biochemistry 20(15):4505–10 [DOI] [PubMed] [Google Scholar]
- 14.Alecio MR, Golan DE, Veatch WR, Rando RR. 1982. Use of a fluorescent cholesterol derivative to measure lateral mobility of cholesterol in membranes. PNAS 79(17):5171–74. Corrigendum. 1982. PNAS 79(24):7949 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hjort Ipsen J, Karlström G, Mourtisen OG, Wennerström H, Zuckermann MJ. 1987. Phase equilibria in the phosphatidylcholine-cholesterol system. Biochim. Biophys. Acta Biomembr 905(1):162–72 [DOI] [PubMed] [Google Scholar]
- 16.Vist MR, Davis JH. 1990. Phase equilibria of cholesterol/dipalmitoylphosphatidylcholine mixtures: deuterium nuclear magnetic resonance and differential scanning calorimetry. Biochemistry 29(2):451–64 [DOI] [PubMed] [Google Scholar]
- 17.Almeida PF, Vaz WL, Thompson TE. 1992. Lateral diffusion in the liquid phases of dimyristoylphosphatidylcholine/cholesterol lipid bilayers: a free volume analysis. Biochemistry 31(29):6739–47 [DOI] [PubMed] [Google Scholar]
- 18.Huang TH, Lee CW, Das Gupta SK, Blume A, Griffin RG. 1993. A 13C and 2H nuclear magnetic resonance study of phosphatidylcholine/cholesterol interactions: characterization of liquid–gel phases. Biochemistry 32(48):13277–87 [DOI] [PubMed] [Google Scholar]
- 19.McMullen TPW, McElhaney RN. 1995. New aspects of the interaction of cholesterol with dipalmitoylphosphatidylcholine bilayers as revealed by high-sensitivity differential scanning calorimetry. Biochim. Biophys. Acta Biomembr 1234(1):90–98 [DOI] [PubMed] [Google Scholar]
- 20.Ahmed SN, Brown DA, London E. 1997. On the origin of sphingolipid/cholesterol-rich detergent-insoluble cell membranes: physiological concentrations of cholesterol and sphingolipid induce formation of a detergent-insoluble, liquid-ordered lipid phase in model membranes. Biochemistry 36(36):10944–53 [DOI] [PubMed] [Google Scholar]
- 21.Feigenson GW, Buboltz JT. 2001. Ternary phase diagram of dipalmitoyl-PC/dilauroyl-PC/cholesterol: nanoscopic domain formation driven by cholesterol. Biophys. J 80(6):2775–88 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.van Meer G, Simons K. 1982. Viruses budding from either the apical or the basolateral plasma membrane domain of MDCK cells have unique phospholipid compositions. EMBO J 1(7):847–52 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Heiniger H-J, Kandutsch AA, Chen HW. 1976. Depletion of L-cell sterol depresses endocytosis. Nature 263(5577):515–17 [DOI] [PubMed] [Google Scholar]
- 24.Alderson JCE, Green C. 1975. Enrichment of lymphocytes with cholesterol and its effect on lymphocyte activation. FEBS Lett 52(2):208–11 [DOI] [PubMed] [Google Scholar]
- 25.Lohr KM, Snyderman R. 1982. Amphotericin B alters the affinity and functional activity of the oligopeptide chemotactic factor receptor on human polymorphonuclear leukocytes. J. Immunol 129(4):1594–99 [PubMed] [Google Scholar]
- 26.Dahl C, Biemann HP, Dahl J. 1987. A protein kinase antigenically related to pp60v-src possibly involved in yeast cell cycle control: positive in vivo regulation by sterol. PNAS 84(12):4012–16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Brown DA, Rose JK. 1992. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell 68(3):533–44 [DOI] [PubMed] [Google Scholar]
- 28.Fiedler K, Kobayashi T, Kurzchalia TV, Simons K. 1993. Glycosphingolipid-enriched, detergent-insoluble complexes in protein sorting in epithelial cells. Biochemistry 32(25):6365–73 [DOI] [PubMed] [Google Scholar]
- 29.Harder T, Scheiffele P, Verkade P, Simons K. 1998. Lipid domain structure of the plasma membrane revealed by patching of membrane components. J. Cell Biol 141(4):929–42 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ge M, Gidwani A, Brown HA, Holowka D, Baird B, Freed JH. 2003. Ordered and disordered phases coexist in plasma membrane vesicles of RBL-2H3 mast cells. An ESR study. Biophys. J 85(2):1278–88 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gidwani A, Holowka D, Baird B. 2001. Fluorescence anisotropy measurements of lipid order in plasma membranes and lipid rafts from RBL-2H3 mast cells. Biochemistry 40(41):12422–29 [DOI] [PubMed] [Google Scholar]
- 32.Kenworthy AK, Edidin M. 1998. Distribution of a glycosylphosphatidylinositol-anchored protein at the apical surface of MDCK cells examined at a resolution of <100 Å using imaging fluorescence resonance energy transfer. J. Cell Biol 142(1):69–84 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sheets ED, Holowka D, Baird B. 1999. Critical role for cholesterol in Lyn-mediated tyrosine phosphorylation of FcεRI and their association with detergent-resistant membranes. J. Cell Biol 145(4):877–87 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Pierce SK. 2002. Lipid rafts and B-cell activation. Nat. Rev. Immunol 2(2):nri726. [DOI] [PubMed] [Google Scholar]
- 35.Plowman SJ, Muncke C, Parton RG, Hancock JF. 2005. H-ras, K-ras, and inner plasma membrane raft proteins operate in nanoclusters with differential dependence on the actin cytoskeleton. PNAS 102(43):15500–5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Brown DA, London E. 1998. Functions of lipid rafts in biological membranes. Annu. Rev. Cell Dev. Biol 14:111–36 [DOI] [PubMed] [Google Scholar]
- 37.Munro S 2003. Lipid rafts: elusive or illusive? Cell 115(4):377–88 [DOI] [PubMed] [Google Scholar]
- 38.Dietrich C, Bagatolli LA, Volovyk ZN, Thompson NL, Levi M, et al. 2001. Lipid rafts reconstituted in model membranes. Biophys. J 80(3):1417–28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Samsonov AV, Mihalyov I, Cohen FS. 2001. Characterization of cholesterol-sphingomyelin domains and their dynamics in bilayer membranes. Biophys. J 81(3):1486–500 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Veatch SL, Keller SL. 2002. Organization in lipid membranes containing cholesterol. Phys. Rev. Lett 89(26):268101. [DOI] [PubMed] [Google Scholar]
- 41.Veatch SL, Gawrisch K, Keller SL. 2006. Closed-loop miscibility gap and quantitative tie-lines in ternary membranes containing diphytanoyl PC. Biophys. J 90(12):4428–36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Veatch SL, Keller SL. 2003. Separation of liquid phases in giant vesicles of ternary mixtures of phospholipids and cholesterol. Biophys. J 85(5):3074–83 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ionova IV, Livshits VA, Marsh D. 2012. Phase diagram of ternary cholesterol/palmitoylsphingomyelin/palmitoyloleoyl-phosphatidylcholine mixtures: spin-label EPR study of lipid-raft formation. Biophys. J 102(8):1856–65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Veatch SL, Keller SL. 2005. Miscibility phase diagrams of giant vesicles containing sphingomyelin. Phys. Rev. Lett 94(14):148101. [DOI] [PubMed] [Google Scholar]
- 45.Veatch SL, Soubias O, Keller SL, Gawrisch K. 2007. Critical fluctuations in domain-forming lipid mixtures. PNAS 104(45):17650–55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Zhao J, Wu J, Heberle FA, Mills TT, Klawitter P, et al. 2007. Phase studies of model biomembranes: complex behavior of DSPC/DOPC/cholesterol. Biochim. Biophys. Acta Biomembr 1768(11):2764–76 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Petruzielo RS, Heberle FA, Drazba P, Katsaras J, Feigenson GW. 2013. Phase behavior and domain size in sphingomyelin-containing lipid bilayers. Biochim. Biophys. Acta Biomembr 1828(4):1302–13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Connell SD, Heath G, Olmsted PD, Kisil A. 2013. Critical point fluctuations in supported lipid membranes. Faraday Discuss 161:91–111 [DOI] [PubMed] [Google Scholar]
- 49.Khadka NK, Ho CS, Pan J. 2015. Macroscopic and nanoscopic heterogeneous structures in a three-component lipid bilayer mixtures determined by atomic force microscopy. Langmuir 31(45):12417–25 [DOI] [PubMed] [Google Scholar]
- 50.Konyakhina TM, Wu J, Mastroianni JD, Heberle FA, Feigenson GW. 2013. Phase diagram of a 4-component lipid mixture: DSPC/DOPC/POPC/chol. Biochim. Biophys. Acta Biomembr 1828(9):2204–14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mills TT, Tristram-Nagle S, Heberle FA, Morales NF, Zhao J, et al. 2008. Liquid-liquid domains in bilayers detected by wide angle X-ray scattering. Biophys. J 95(2):682–90 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Konyakhina TM, Feigenson GW. 2016. Phase diagram of a polyunsaturated lipid mixture: brain sphingomyelin/1-stearoyl-2-docosahexaenoyl-sn-glycero-3-phosphocholine/ cholesterol. Biochim. Biophys. Acta Biomembr 1858(1):153–61 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.de Almeida RFM, Borst J, Fedorov A, Prieto M, Visser AJWG. 2007. Complexity of lipid domains and rafts in giant unilamellar vesicles revealed by combining imaging and microscopic and macroscopic time-resolved fluorescence. Biophys. J 93(2):539–53 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Sibold J, Tewaag VE, Vagedes T, Mey I, Steinem C. 2020. Phase separation in pore-spanning membranes induced by differences in surface adhesion. Phys. Chem. Chem. Phys 22(17):9308–15 [DOI] [PubMed] [Google Scholar]
- 55.Kahya N, Scherfeld D, Bacia K, Poolman B, Schwille P. 2003. Probing lipid mobility of raft-exhibiting model membranes by fluorescence correlation spectroscopy. J. Biol. Chem 278(30):28109–15 [DOI] [PubMed] [Google Scholar]
- 56.Aufderhorst-Roberts A, Chandra U, Connell SD. 2017. Three-phase coexistence in lipid membranes. Biophys. J 112(2):313–24 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sodt AJ, Sandar ML, Gawrisch K, Pastor RW, Lyman E. 2014. The molecular structure of the liquid-ordered phase of lipid bilayers. J. Am. Chem. Soc 136(2):725–32 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Sodt AJ, Pastor RW, Lyman E. 2015. Hexagonal substructure and hydrogen bonding in liquid-ordered phases containing palmitoyl sphingomyelin. Biophys. J 109(5):948–55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Goñi FM, Alonso A, Bagatolli LA, Brown RE, Marsh D, et al. 2008. Phase diagrams of lipid mixtures relevant to the study of membrane rafts. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1781(11):665–84 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Heberle FA, Feigenson GW. 2011. Phase separation in lipid membranes. Cold Spring Harb. Perspect. Biol 3(4):a004630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ladbrooke BD, Chapman D. 1969. Thermal analysis of lipids, proteins and biological membranes a review and summary of some recent studies. Chem. Phys. Lipids 3(4):304–56 [DOI] [PubMed] [Google Scholar]
- 62.Janiak MJ, Small DM, Shipley GG. 1979. Temperature and compositional dependence of the structure of hydrated dimyristoyl lecithin. J. Biol. Chem 254(13):6068–78 [PubMed] [Google Scholar]
- 63.Heberle FA, Wu J, Goh SL, Petruzielo RS, Feigenson GW. 2010. Comparison of three ternary lipid bilayer mixtures: FRET and ESR reveal nanodomains. Biophys. J 99(10):3309–18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Pathak P, London E. 2011. Measurement of lipid nanodomain (raft) formation and size in sphingomyelin/POPC/cholesterol vesicles shows TX-100 and transmembrane helices increase domain size by coalescing preexisting nanodomains but do not induce domain formation. Biophys. J 101(10):2417–25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Pathak P, London E. 2015. The effect of membrane lipid composition on the formation of lipid ultrananodomains. Biophys. J 109(8):1630–38 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Konyakhina TM, Goh SL, Amazon J, Heberle FA, Wu J, Feigenson GW. 2011. Control of a nanoscopic-to-macroscopic transition: modulated phases in four-component DSPC/DOPC/POPC/chol giant unilamellar vesicles. Biophys. J 101(2):L8–10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Collado MI, Goñi FM, Alonso A, Marsh D. 2005. Domain formation in sphingomyelin/cholesterol mixed membranes studied by spin-label electron spin resonance spectroscopy. Biochemistry 44(12):4911–18 [DOI] [PubMed] [Google Scholar]
- 68.de Almeida RFM, Fedorov A, Prieto M. 2003. Sphingomyelin/phosphatidylcholine/cholesterol phase diagram: boundaries and composition of lipid rafts. Biophys. J 85(4):2406–16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Doktorova M, Heberle FA, Eicher B, Standaert RF, Katsaras J, et al. 2018. Preparation of asymmetric phospholipid vesicles for use as cell membrane models. Nat. Protoc 13(9):2086–101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Lin Q, London E. 2015. Ordered raft domains induced by outer leaflet sphingomyelin in cholesterol-rich asymmetric vesicles. Biophys. J 108(9):2212–22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Honerkamp-Smith AR, Cicuta P, Collins MD, Veatch SL, den Nijs M, et al. 2008. Line tensions, correlation lengths, and critical exponents in lipid membranes near critical points. Biophys. J 95(1):236–46 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Leibler S, Andelman D. 1987. Ordered and curved meso-structures in membranes and amphiphilic films. J. Phys 48(11):2013–18 [Google Scholar]
- 73.Palmieri B, Yamamoto T, Brewster RC, Safran SA. 2014. Line active molecules promote inhomogeneous structures in membranes: theory, simulations and experiments. Adv. Colloid Interface Sci 208:58–65 [DOI] [PubMed] [Google Scholar]
- 74.Brodbek L, Schmid F. 2016. Interplay of curvature-induced micro- and nanodomain structures in multicomponent lipid bilayers. Int. J. Adv. Eng. Sci. Appl. Math 8(2):111–20 [Google Scholar]
- 75.Cornell CE, Skinkle AD, He S, Levental I, Levental KR, Keller SL. 2018. Tuning length scales of small domains in cell-derived membranes and synthetic model membranes. Biophys. J 115(4):690–701 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Fridriksson EK, Shipkova PA, Sheets ED, Holowka D, Baird B, McLafferty FW. 1999. Quantitative analysis of phospholipids in functionally important membrane domains from RBL-2H3 mast cells using tandem high-resolution mass spectrometry. Biochemistry 38(25):8056–63 [DOI] [PubMed] [Google Scholar]
- 77.Lingwood D, Ries J, Schwille P, Simons K. 2008. Plasma membranes are poised for activation of raft phase coalescence at physiological temperature. PNAS 105(29):10005–10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Levental I, Grzybek M, Simons K. 2011. Raft domains of variable properties and compositions in plasma membrane vesicles. PNAS 108(28):11411–16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Levental I, Byfield FJ, Chowdhury P, Gai F, Baumgart T, Janmey PA. 2009. Cholesterol-dependent phase separation in cell-derived giant plasma-membrane vesicles. Biochem. J 424(2):163–67 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Zhao J, Wu J, Veatch SL. 2013. Adhesion stabilizes robust lipid heterogeneity in supercritical membranes at physiological temperature. Biophys. J 104(4):825–34 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Veatch SL, Cicuta P, Sengupta P, Honerkamp-Smith A, Holowka D, Baird B. 2008. Critical fluctuations in plasma membrane vesicles. ACS Chem. Biol 3(5):287–93 [DOI] [PubMed] [Google Scholar]
- 82.Burns M, Wisser K, Wu J, Levental I, Veatch SL. 2017. Miscibility transition temperature scales with growth temperature in a zebrafish cell line. Biophys. J 113(6):1212–22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Gray EM, Díaz-Vázquez G, Veatch SL. 2015. Growth conditions and cell cycle phase modulate phase transition temperatures in RBL-2H3 derived plasma membrane vesicles. PLOS ONE 10(9):e0137741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Levental KR, Lorent JH, Lin X, Skinkle AD, Surma MA, et al. 2016. Polyunsaturated lipids regulate membrane domain stability by tuning membrane order. Biophys. J 110(8):1800–10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Levental KR, Surma MA, Skinkle AD, Lorent JH, Zhou Y, et al. 2017. ω−3 polyunsaturated fatty acids direct differentiation of the membrane phenotype in mesenchymal stem cells to potentiate osteogenesis. Sci. Adv 3(11):eaao1193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Cammarota E, Soriani C, Taub R, Morgan F, Sakai J, et al. 2020. Criticality of plasma membrane lipids reflects activation state of macrophage cells. J. R. Soc. Interface 17(163):20190803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Symons JL, Cho K-J, Chang JT, Du G, Waxham MN, et al. 2020. Lipidomic atlas of mammalian cell membranes reveals hierarchical variation induced by culture conditions, subcellular membranes, and cell lineages. Soft Matter In press. 10.1039/D0SM00404A [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Sezgin E, Levental I, Grzybek M, Schwarzmann G, Mueller V, et al. 2012. Partitioning, diffusion, and ligand binding of raft lipid analogs in model and cellular plasma membranes. Biochim. Biophys. Acta Biomembr 1818(7):1777–84 [DOI] [PubMed] [Google Scholar]
- 89.Kaiser H-J, Lingwood D, Levental I, Sampaio JL, Kalvodova L, et al. 2009. Order of lipid phases in model and plasma membranes. PNAS 106(39):16645–50 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Lorent JH, Diaz-Rohrer B, Lin X, Spring K, Gorfe AA, et al. 2017. Structural determinants and functional consequences of protein affinity for membrane rafts. Nat. Commun 8:1219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Menon AK, Holowka D, Webb WW, Baird B. 1986. Cross-linking of receptor-bound IgE to aggregates larger than dimers leads to rapid immobilization. J. Cell Biol 102(2):541–50 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Machta BB, Papanikolaou S, Sethna JP, Veatch SL. 2011. Minimal model of plasma membrane heterogeneity requires coupling cortical actin to criticality. Biophys. J 100(7):1668–77 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Keller H, Lorizate M, Schwille P. 2009. PI(4,5)P2 degradation promotes the formation of cytoskeleton-free model membrane systems. ChemPhysChem 10(16):2805–12 [DOI] [PubMed] [Google Scholar]
- 94.McLaughlin S, Wang J, Gambhir A, Murray D. 2002. PIP2 and proteins: interactions, organization, and information flow. Annu. Rev. Biophys. Biomol. Struct 31:151–75 [DOI] [PubMed] [Google Scholar]
- 95.Skinkle AD, Levental KR, Levental I. 2020. Cell-derived plasma membrane vesicles are permeable to hydrophilic macromolecules. Biophys. J 118(6):1292–300 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Li G, Wang Q, Kakuda S, London E. 2020. Nanodomains can persist at physiologic temperature in plasma membrane vesicles and be modulated by altering cell lipids. J. Lipid Res 61:758–66 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Toulmay A, Prinz WA. 2013. Direct imaging reveals stable, micrometer-scale lipid domains that segregate proteins in live cells. J. Cell Biol 202(1):35–44 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Rayermann SP, Rayermann GE, Cornell CE, Merz AJ, Keller SL. 2017. Hallmarks of reversible separation of living, unperturbed cell membranes into two liquid phases. Biophys. J 113(11):2425–32 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Lee I-H, Saha S, Polley A, Huang H, Mayor S, et al. 2015. Live cell plasma membranes do not exhibit a miscibility phase transition over a wide range of temperatures. J. Phys. Chem. B 119(12):4450–59 [DOI] [PubMed] [Google Scholar]
- 100.Sevcsik E, Brameshuber M, Fölser M, Weghuber J, Honigmann A, Schütz GJ. 2015. GPI-anchored proteins do not reside in ordered domains in the live cell plasma membrane. Nat. Commun 6:6969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Levental I, Levental KR, Heberle FA. 2020. Lipid rafts: controversies resolved, mysteries remain. Trends Cell Biol 30(5):341–53 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kusumi A, Fujiwara TK, Tsunoyama TA, Kasai RS, Liu A-A, et al. 2020. Defining raft domains in the plasma membrane. Traffic 21(1):106–37 [DOI] [PubMed] [Google Scholar]
- 103.Castello-Serrano I, Lorent JH, Ippolito R, Levental KR, Levental I. 2020. Myelin-associated MAL and PLP are unusual among multipass transmembrane proteins in preferring ordered membrane domains. J. Phys. Chem. B 124(28):5930–39 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Marinko JT, Kenworthy AK, Sanders CR. 2020. Peripheral myelin protein 22 preferentially partitions into ordered phase membrane domains. PNAS 117(25):14168–77 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Yang S-T, Kreutzberger AJB, Kiessling V, Ganser-Pornillos BK, White JM, Tamm LK. 2017. HIV virions sense plasma membrane heterogeneity for cell entry. Sci. Adv 3(6):e1700338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Anderson RGW, Jacobson K. 2002. A role for lipid shells in targeting proteins to caveolae, rafts, and other lipid domains. Science 296(5574):1821–25 [DOI] [PubMed] [Google Scholar]
- 107.Corradi V, Mendez-Villuendas E, Ingólfsson HI, Gu R-X, Siuda I, et al. 2018. Lipid–protein interactions are unique fingerprints for membrane proteins. ACS Cent. Sci 4(6):709–17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Gray E, Karslake J, Machta BB, Veatch SL. 2013. Liquid general anesthetics lower critical temperatures in plasma membrane vesicles. Biophys. J 105(12):2751–59 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Machta BB, Gray E, Nouri M, McCarthy NLC, Gray EM, et al. 2016. Conditions that stabilize membrane domains also antagonize n-alcohol anesthesia. Biophys. J 111(3):537–45 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Grinstein G, Ma S. 1982. Roughening and lower critical dimension in the random-field Ising model. Phys. Rev. Lett 49(9):685–88 [Google Scholar]
- 111.Yethiraj A, Weisshaar JC. 2007. Why are lipid rafts not observed in vivo? Biophys. J 93(9):3113–19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Honigmann A, Sadeghi S, Keller J, Hell SW, Eggeling C, Vink R. 2014. A lipid bound actin meshwork organizes liquid phase separation in model membranes. eLife 3:e01671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Kwik J, Boyle S, Fooksman D, Margolis L, Sheetz MP, Edidin M. 2003. Membrane cholesterol, lateral mobility, and the phosphatidylinositol 4,5-bisphosphate-dependent organization of cell actin. PNAS 100(24):13964–69 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Liu AP, Fletcher DA. 2006. Actin polymerization serves as a membrane domain switch in model lipid bilayers. Biophys. J 91(11):4064–70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Shelby SA, Veatch SL, Holowka DA, Baird BA. 2016. Functional nanoscale coupling of Lyn kinase with IgE-FcεRI is restricted by the actin cytoskeleton in early antigen-stimulated signaling. Mol. Biol. Cell 27(22):3645–58 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Gudheti MV, Curthoys NM, Gould TJ, Kim D, Gunewardene MS, et al. 2013. Actin mediates the nanoscale membrane organization of the clustered membrane protein influenza hemagglutinin. Biophys. J 104(10):2182–92 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Chichili GR, Rodgers W. 2009. Cytoskeleton-membrane interactions in membrane raft structure. Cell. Mol. Life Sci 66(14):2319–28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Stone MB, Shelby SA, Núñez MF, Wisser K, Veatch SL. 2017. Protein sorting by lipid phase-like domains supports emergent signaling function in B lymphocyte plasma membranes. eLife 6:e19891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Kinoshita M, Suzuki KGN, Matsumori N, Takada M, Ano H, et al. 2017. Raft-based sphingomyelin interactions revealed by new fluorescent sphingomyelin analogs. J. Cell Biol 216(4):1183–204 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Allender DW, Giang H, Schick M. 2020. Model plasma membrane exhibits a microemulsion in both leaves providing a foundation for “rafts.” Biophys. J 118(5):1019–31 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Foret L 2005. A simple mechanism of raft formation in two-component fluid membranes. Europhys. Lett 71(3):508–14 [Google Scholar]
- 122.Gowrishankar K, Ghosh S, Saha S, Rumamol C, Mayor S, Rao M. 2012. Active remodeling of cortical actin regulates spatiotemporal organization of cell surface molecules. Cell 149(6):1353–67 [DOI] [PubMed] [Google Scholar]
- 123.Schmid F 2017. Physical mechanisms of micro- and nanodomain formation in multicomponent lipid membranes. Biochim. Biophys. Acta Biomembr 1859(4):509–28 [DOI] [PubMed] [Google Scholar]
- 124.Goldenfeld N 2018. Lectures on Phase Transitions and the Renormalization Group Boca Raton, FL:CRC Press [Google Scholar]
- 125.Cardy J 1996. Scaling and Renormalization in Statistical Physics Cambridge, UK: Cambridge Univ. Press [Google Scholar]
- 126.Kardar M 2007. Statistical Physics of Fields Cambridge, UK: Cambridge Univ. Press [Google Scholar]
- 127.Chaikin PM, Lubensky TC. 2010. Principles of Condensed Matter Physics Cambridge, UK: Cambridge Univ. Press [Google Scholar]
- 128.Honerkamp-Smith AR, Veatch SL, Keller SL. 2009. An introduction to critical points for biophysicists; observations of compositional heterogeneity in lipid membranes. Biochim. Biophys. Acta Biomembr 1788(1):53–63 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Cagniard de la Tour C. 1822. Exposé de quelques résultats obtenu par l’action combinée de la chaleur et de la compression sur certains liquides, tels que l’eau, l’alcool, l’éther sulfurique et l’essence de pétrole rectifiée. Ann. Chim. Phys 21:127–32 [Google Scholar]
- 130.van der Waals JD, Rowlinson JS. 2004. On the Continuity of the Gaseous and Liquid States Mineola, NY: Dover [Google Scholar]
- 131.Ornstein L, Zernike F. 1918. Die linearen Dimensionen der Dichteschwankungen. Phys. Zeit 19:134–37 [Google Scholar]
- 132.Guggenheim EA. 1945. The principle of corresponding states. J. Chem. Phys 13(7):253–61 [Google Scholar]
- 133.Fisher ME. 1967. The theory of equilibrium critical phenomena. Rep. Prog. Phys 30(2):615–730. Corrigendum. 1968. Rep. Prog. Phys. 31:418–20 [Google Scholar]
- 134.Onsager L 1944. Crystal statistics. I. A two-dimensional model with an order-disorder transition. Phys. Rev 65(3–4):117–49 [Google Scholar]
- 135.Lee TD, Yang CN. 1952. Statistical theory of equations of state and phase transitions. II. Lattice gas and Ising model. Phys. Rev 87(3):410–19 [Google Scholar]
- 136.Widom B 1965. Equation of state in the neighborhood of the critical point. J. Chem. Phys 43(11):3898–905 [Google Scholar]
- 137.Kadanoff LP. 1966. Scaling laws for Ising models near Tc. Phys. Phys. Fiz 2(6):263–72 [Google Scholar]
- 138.Wilson KG. 1971. Renormalization group and critical phenomena. I. Renormalization group and the Kadanoff scaling picture. Phys. Rev. B 4(9):3174–83 [Google Scholar]
- 139.Mora T, Bialek W. 2011. Are biological systems poised at criticality? J. Stat. Phys 144(2):268–302 [Google Scholar]
- 140.Honerkamp-Smith AR, Machta BB, Keller SL. 2012. Experimental observations of dynamic critical phenomena in a lipid membrane. Phys. Rev. Lett 108(26):265702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Frazier ML, Wright JR, Pokorny A, Almeida PFF. 2007. Investigation of domain formation in sphingomyelin/cholesterol/POPC mixtures by fluorescence resonance energy transfer and Monte Carlo simulations. Biophys. J 92(7):2422–33 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Manley S, Horton MR, Lecszynski S, Gast AP. 2008. Sorting of streptavidin protein coats on phase-separating model membranes. Biophys. J 95(5):2301–7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Núñez MF, Wisser K, Veatch SL. 2019. Synergistic factors control kinase-phosphatase organization in B-cells engaged with supported bilayers. Mol. Biol. Cell 31(7):667–82 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Kimchi O, Veatch SL, Machta BB. 2018. Ion channels can be allosterically regulated by membrane domains near a de-mixing critical point. J. Gen. Physiol 150(12):1769–77 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Lingwood D, Simons K. 2010. Lipid rafts as a membrane-organizing principle. Science 327:46–50 [DOI] [PubMed] [Google Scholar]
- 146.Pike LJ. 2006. Rafts defined: a report on the Keystone symposium on lipid rafts and cell function. J. Lipid Res 47:1597–98 [DOI] [PubMed] [Google Scholar]
- 147.Levental I, Veatch SL. 2016. The continuing mystery of lipid rafts. J. Mol. Biol 428(24, Part A):4749–64 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Sezgin E, Levental I, Mayor S, Eggeling C. 2017. The mystery of membrane organization: composition, regulation and roles of lipid rafts. Nat. Rev. Mol. Cell Biol 18(6):361–74 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Casimir HBG. 1948. On the attraction between two perfectly conducting plates. Indag. Math 10:261–63 [Google Scholar]
- 150.Machta BB, Veatch SL, Sethna JP. 2012. Critical Casimir forces in cellular membranes. Phys. Rev. Lett 109(13):138101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Reynwar BJ, Deserno M. 2008. Membrane composition-mediated protein-protein interactions. Biointerphases 3(2):FA117–24 [DOI] [PubMed] [Google Scholar]
- 152.Saheki Y, De Camilli P. 2017. Endoplasmic reticulum-plasma membrane contact sites. Annu. Rev. Biochem 86:659–84 [DOI] [PubMed] [Google Scholar]
- 153.Zeng M, Chen X, Guan D, Xu J, Wu H, et al. 2018. Reconstituted postsynaptic density as a molecular platform for understanding synapse formation and plasticity. Cell 174(5):1172–87. e16 [DOI] [PubMed] [Google Scholar]
- 154.Ayuyan AG, Cohen FS. 2018. The chemical potential of plasma membrane cholesterol: implications for cell biology. Biophys. J 114(4):904–18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Fantini J, Epand RM, Barrantes FJ. 2019. Cholesterol-recognition motifs in membrane proteins. Adv. Exp. Med. Biol 1135:3–25 [DOI] [PubMed] [Google Scholar]
- 156.De Craene J-O, Bertazzi DL, Bär S, Friant S. 2017. Phosphoinositides, major actors in membrane trafficking and lipid signaling pathways. Int. J. Mol. Sci 18(3):634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Shimizu T 2009. Lipid mediators in health and disease: enzymes and receptors as therapeutic targets for the regulation of immunity and inflammation. Annu. Rev. Pharmacol. Toxicol 49:123–50 [DOI] [PubMed] [Google Scholar]
- 158.Eggeling C, Ringemann C, Medda R, Schwarzmann G, Sandhoff K, et al. 2009. Direct observation of the nanoscale dynamics of membrane lipids in a living cell. Nature 457(7233):1159–62 [DOI] [PubMed] [Google Scholar]
- 159.Rao M, Mayor S. 2014. Active organization of membrane constituents in living cells. Curr. Opin. Cell Biol 29:126–32 [DOI] [PubMed] [Google Scholar]
- 160.Sorre B, Callan-Jones A, Manneville J-B, Nassoy P, Joanny J-F, et al. 2009. Curvature-driven lipid sorting needs proximity to a demixing point and is aided by proteins. PNAS 106(14):5622–26 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Mitra ED, Whitehead SC, Holowka D, Baird B, Sethna JP. 2018. Computation of a theoretical membrane phase diagram and the role of phase in lipid-raft-mediated protein organization. J. Phys. Chem. B 122(13):3500–13 [DOI] [PMC free article] [PubMed] [Google Scholar]