

Abstract
Intervention studies indicate that children’s early child-care experiences can be leveraged to foster their development of effective self-regulation skills. It is less clear whether typical child-care experiences play a similar role. In addition, evidence suggests that children with a common variant of the DRD4 gene (48-bp VNTR, 7-repeat) may be more sensitive to their experiences than those without this variant. Using data from the NICHD Study of Early Child Care and Youth Development, we considered the degree to which children’s early child-care experiences—quantity, quality, and type—were associated with their attention and self-regulation abilities in prekindergarten, and, in particular, whether these relations were conditional on DRD4 genotype. G × E interactions were evident across multiple neuropsychological and observational measures of children’s attention and self-regulation abilities. Across most outcome measures, DRD4 7+ children spending fewer hours in child care showed more effective attention/self-regulation abilities. For those without a copy of the DRD4 7-repeat allele, such associations were typically null. The results for child-care quality and type indicated no interactions with genotype; the main-effect associations were somewhat inconsistent.
Keywords: attention, early experience, parental care, child care
INTRODUCTION
Developing effective self-regulation skills in early childhood plays an important role in children’s successful entry into formal schooling (Blair, 2002; Ladd, Herald, & Kochel, 2006). Growing evidence suggests that children’s experiences at home may support their emerging capacities to control their emotions, thoughts, and attention (Belsky, Fearon, & Bell, 2007; Bernier, Carlson, & Whipple, 2010; Kochanska, Murray, & Harlan, 2000; NICHD Early Child Care Research Network [ECCRN], 2003a). By the age of five, 63% of children are in regular child-care arrangements (U.S. Census Bureau, 2010). Less is understood about the degree to children’s experiences in childcare support or undermine their self-regulation development.
Although there is good evidence that high-quality and center-cased child-care experiences may support children’s academic skills (Belsky et al., 2007; Loeb, Bridges, Bassok, Fuller, & Rumberger, 2007; NICHD ECCRN & Duncan, 2003)—potentially reflecting better self-regulation abilities (Blair & Razza, 2007; McClelland et al., 2007). There is also, however, mixed evidence linking heightened child-care exposure with children’s development of behavioral problems (Belsky et al., 2007; McCartney et al., 2010; NICHD ECCRN, 2003b), which may reflect self-regulation deficits (Séguin & Zelazo, 2005). Some have proposed that these mixed findings may be explained, in part, by meaningful individual differences in the way children respond to their child-care experiences (Crockenberg & Leerkes, 2005; Phillips, Fox, & Gunnar, 2011; Pluess & Belsky, 2009). In particular, increasing theoretical and empirical work supports the proposition that genetic and experiential differences may lead some children to be more strongly impacted by—or differentially susceptible to—their experiences than others (Belsky & Pluess, 2009; Boyce & Ellis, 2005).
Indeed, evidence suggests that gene-by-environment interactions involving a common variant of the dopamine receptor D4 (DRD4) gene are largely consistent with the differential susceptibility framework (see Bakermans-Kranenburg & van IJzendoorn, 2011). In the present study, we consider the degree to which children’s experiences in nonmaternal child care—quantity, quality and type—are associated with their development of attention and self-regulation skills and, in particular, whether these relations are moderated by DRD4 genotype.
Self-Regulation and Development
The construct of self-regulation is discussed across several literatures. Cognitive perspectives tend to highlight a core set of executive functions—working memory, inhibitory control, and attention shifting—that allow one to hold and manipulate information in mind, inhibit well-learned prepotent responses in order to enact subdominant responses, and volitionally modulate attention, respectively (Miyake et al., 2000). Temperament-based perspectives tend to discuss the related concept of effortful control, in which children employ executive attentional processes to downregulate desires, emotions, and well-learned behaviors, in order to adopt alternative behaviors (Rothbart, Ahadi, Hershey, & Fisher, 2001). Building on commonalties across these areas, we adopt a broad definition of self-regulation, as the largely (but not exclusively) volitional act of managing attention and arousal in a manner that facilitates goal-directed behavior (Blair & Ursache, 2011).
Accumulating evidence suggests that children’s early experiences may play an important role in their development of self-regulation skills. In multiple studies and diverse samples, children who experience emotionally close, sensitive, and responsive caregiving have been found to show more effective self-regulation skills. These statistical associations have been illustrated using neuropsychological measures of attention and executive functioning (Blair et al., 2011; Belsky, Fearon, & Bell, 2007; NICHD ECCRN, 2003a), as well as behavioral tasks (Kochanska et al., 2000; Spinrad et al., 2007) and questionnaire measures tapping effortful control (Spinrad et al., 2007).
Child Care and Self-Regulation
Notably, children spend a substantial proportion of their time in developmental contexts apart from their interactions with their parents. On one hand, it is increasingly clear that contexts outside of the home can be leveraged to deliver early experiences that foster the development of self-regulation skills in early childhood. For example, curricula and child-care interventions designed explicitly to enhance self-regulation skills have shown positive impacts on children’s executive functioning abilities and social behavior in early childhood (Barnett et al., 2008; Bierman, Nix, Greenberg, Blair, & Domitrovich, 2008; Raver et al., 2012). Yet, it is largely unclear whether child-care experiences that are available typically to families play a role in children’s emerging self-regulation abilities.
To date, few studies have considered links between normative early child-care experiences and children’s development of self-regulation skills. The limited available data have been somewhat inconsistent. For example, in prior work using the same sample as the present study, the NICHD ECCRN 1998) found that higher quality care was associated with lower emotional negativity and better abilities to delay gratification at the age of three. Yet, the findings were somewhat inconsistent across child age, and no other child-care indicators (i.e., quantity, stability, type, peer exposure) were associated with children’s self-regulation abilities.
There is, however, strong evidence that high-quality child care may support children’s development of early academic skills—an outcome thought to reflect effective self-regulation skills (Blair, 2002; Blair & Razza, 2007). Several studies, including but not limited to those using the present sample, have shown that children attending higher quality care tend to show better reading and math scores than do their peers attending lower quality care. These relations are evident in early childhood and have been linked with better cognitive-academic outcomes in early elementary school, as well as adolescence (Belsky et al., 2007; Vandell et al., 2010). Experimental studies of high-quality child-care interventions show positive effects on outcomes like earnings that extend into adulthood (Campbell, Ramey, Pungello, Sparling, & Miller-Johnson, 2002; Schweinhart et al., 2005). Similarly, exposure to center-based care, in particular, which tends provide more predictable schedules, better trained caregivers, and richer learning environments (Fuller, Kagan, Loeb, & Chang, 2004), has been linked to better academic achievement (Loeb et al., 2007; Magnuson, Ruhm, & Waldfogel, 2007; NICHD ECCRN & Duncan,2003).
Although associations between child-care quantity and academic achievement have been more mixed (see Burchinal & Clark-Stewart, 2007), there is some indication that—when quality is high—greater child-care exposure may be beneficial for children, at least for those from less affluent families (Dearing, McCartney, & Taylor, 2009).
Notably, there has also historically been some concern that extensive hours in care may have negative developmental impacts on children’s socio-emotional development, which is also thought to reflect children’s attention and self-regulation skills (Hughes & Ensor, 2011; Séguin & Zelazo, 2005). For example, some have proposed that early and extensive exposure to child care may undermine the positive socializing effects of parenting on children’s social development (Belsky, 1999; NICHD ECCRN 1998). In addition, children’s child-care experiences may serve as real-time physiological stressors, known to undermine self-regulatory resources. Increasing evidence suggests that child-care exposure is associated with heightened levels of salivary cortisol, an indicator of hypothalamic–pituitary–adrenal (HPA) axis stress response (see Vermeer & Van IJzendoorn, 2006). In turn, a notable body of work with animals and human adults indicates that very high or very low levels of cortisol tend to be associated with less effective executive self-regulation abilities (deKloet, Oitzl, & Joels, 1999; Lupien, Maheu, Tu, Fiocco, & Schramek, 2007).
The degree to which the greater hours in child care cause children’s behavior problems or whether children’s behavior problems are simply a correlate remains somewhat unclear (McCartney et al., 2010). Positive associations between early child care and more pronounced behavior problems are often robust after accounting for other aspects of care, such as quality and stability and have been illustrated longitudinally to related behaviors like impulsivity and risk-taking that extend to adolescence (Vandell et al., 2010). Notably, work with primarily low-income samples has shown inversed effects, such that greater exposure to high-quality child care may support better socio-emotional outcomes (Votruba-Drzal, Coley, & Chase-Lansdale, 2004).
Individual Differences and Differential Susceptibility
Several research groups have theorized that the inconsistent nature and invariably modest size of child-care quantity effects on children’s behavior problems may be explained in part by predictable individual differences in the way children are affected by their early experiences (Crockenberg & Leerkes, 2005; Ellis, Boyce, Belsky, Bakermans-Kranenburg, & van IJzendoorn, 2011; Phillips et al., 2011; Pluess & Belsky, 2009). In particular, some have proposed that genetic, environmental, and early gene–environment processes may lead some children to be more malleable or susceptible to their early experiences than others. Differential susceptibility is posited to function in a “for better and for worse” manner (Belsky & Pluess, 2009). Highly susceptible children are theorized to reap the largest benefits from environmental supports, yet also be most negatively impacted by deleterious environmental contexts.
Adopting temperament as a potential indicator of environmental susceptibility, some evidence suggests that associations between children’s child-care experiences and their development of behavior problems may be particularly pronounced for temperamentally reactive children. Crockenberg and Leerkes (2005) found that, for highly reactive children attending longer hours of child care, attending non-center-based care was a protective factor against the development of externalizing problems. Using the same sample as the present study, the NICHD SECCYD, Pluess and Belsky (2009) reported that the association between higher quality care—but not quantity—and more optimal social behavior in prekindergarten and kindergarten was particularly pronounced for infants showing high levels of emotional negativity.
To our knowledge, no work to date has considered similar interactive effects with respect to potential genetic indicators of differential susceptibility. Yet, accumulating evidence from studies of G × E processes in the development of substantively related outcomes suggest that genetic factors may lead some children to be more sensitive to their experiences than others. In particular, a common variant of the DRD4 gene—a 48-bp variable nucleotide tandem repeat (VNTR) polymorphism—may be especially relevant to the development of self-regulation skills.
Dopamine is a primary neuromodulator in frontal cortical regions associated with executive functions and self-regulation (Arnsten & Li, 2005). As well, dopamine has well-established associations with other endocrine mediators of the stress response, glucocorticoids and norepinephrine (Butts, Weinberg, Young, & Phillips, 2011). Notably, the D4 dopamine receptor appears to be a highly relevant to behavior regulation in that norepinephrine has high affinity for this receptor, such that the D4 receptor functions more as a general catecholamine receptor than dopamine receptor, exclusively (Arnsten & Li, 2005).
The DRD4 48-bp VNTR is highly variable (2–11 repeats); the 4-repeat allele is the most common, followed by the 7- and 2-repeat alleles (Chang, Kidd, Livak, Pakstis, & Kidd, 1996). The 7-repeat variant has been implicated in reduced D4 receptor expression in vitro (Schoots & Van Tol, 2003) and, via reduced inhibition of cyclic adenosine monophosphate (cAMP), is inferred to lead to “noisier” network transmission in regions such as the frontal cortex where D4 receptors are highly expressed (Meador-Woodruff et al., 1996; Oak, Oldenhof, Hubert, & Van Tol, 2000).
Studies considering the main effects of the DRD4 7-repeat on cognitive and attentional self-regulation have been inconsistent and largely limited to clinical samples of children with attention deficit hyperactivity disorder (ADHD). Within this population, some work has found the 7-repeat to be (albeit, inconsistently across measures) linked with worse executive functioning (Langley et al., 2004; Waldman, 2005). In contrast, other studies have shown inversed (Bellgrove et al., 2005; Johnson et al., 2008) or null relations (Barkley, Smith, Fischer, & Navia, 2006) between the DRD4 7-repeat and measures of executive function and attention.
Beyond genetic main effects, there is increasing evidence that the DRD4 7-repeat variant may moderate the effects of children’s experiences on developmental outcomes thought to partially reflect self-regulation (see Bakermans-Kranenberg & van IJzendoorn, 2011). In recent work with the same data used in the present study, research from our group indicated that the long-term relation between children’s experiences of maternal sensitivity in early childhood and their subsequent development of attention problems across elementary school was moderated by the presence of a DRD4 7-repeat (Berry, Deater-Deckard, McCartney, Wang, & Petrill, in press). Specifically, the relation between early maternal sensitivity and children’s inattention trajectories was evident only for those with a copy of the DRD4 7-repeat. Further, the conditional genetic effects were largely consistent with the “for better and for worse” prediction made by the differential susceptibility framework—the degree to which the DRD4 7-repeat served as a “risk” or “beneficial” factor was conditional on experience. As summarized by Bakermans-Kranenburg and van IJzendoorn (2011), similar G × E interactions are evident for maternal sensitivity and sensation-seeking in toddlerhood (Sheese, Voekler, Rothbart, & Posner, 2007) and externalizing problems in early childhood (Bakermans-Kranenburg & van IJzendoorn, 2006; Bakermans-Kranenburg, van IJzendoorn, Pijlman, Mesman, & Juffer, 2008).
The Present Study
Taken together, experimental evidence indicates that high-quality child-care interventions that are explicitly designed to enhance children’s self-regulation skills can and do impact children’s self-regulation development. Yet, few studies have considered similar relations for the types of child-care experiences that are available typically to families. The aim of the present study is to test the degree to which children’s early child-care experiences—quantity, quality, and type—are predictive of their self-regulation skills in prekindergarten. In particular, a key aim is to test the degree to which such relations may be most substantial for (or perhaps exclusive to) children with a copy of the DRD4 7-repeat allele.
Based on the literature reviewed above, we hypothesized that greater hours in child care and lower quality care would be predictive less effective self-regulation skills, but that these effects would be particularly pronounced for (or limited to) those with a copy of the DRD4 7-repeat. Given that center-based care may simultaneously provide experiences that support (e.g., predictable environments, more extensive teacher training) and challenge (e.g., larger peer groups) we considered these G × E analyses somewhat more exploratory.
METHOD
Participants
In the current study, we examined longitudinal data from Phases I and II of the National Institute of Child Health and Development Study of Early Child Care and Youth Development (NICHD SECCYD), a longitudinal study of 1,364 children (50.1% male) and their families in 10 sites across the U.S. Potential participants were recruited originally from among 8,986 mothers who gave birth within a 24-hr sampling period in 31 hospitals across the sites. Participants were selected such that they represent demographically the catchment area from which they were recruited (see NICHD ECCRN, 1997 for a comprehensive review of the sampling procedure). Eight of the original ten sites (711 children) chose to participate in the genetics protocol of the study. Neither the original sample, nor the representation of those providing genetic data is nationally representative. Yet, the sample is socio-demographically diverse. Approximately, 83% of children are Caucasian, 10% are African-American, 6% are Latino, 2% are other ethnicities. In early infancy, approximately 8% of mothers had less than a high-school education, 20% had a high school education, 57% between 2 and 4 years of college, and 16% had postgraduate educations 1 month after the child was born. Approximately 28% of the children were from low-income families (i.e., income-to-needs ratio ≤ 2, infancy to prekindergarten).
Procedures
When the child was 1-month old, demographic information with regard to maternal education level, age at birth, marital status, and child gestational age were collected, and mothers were administered questionnaires tapping their levels of separation anxiety, their feelings about the developmental benefits of working, and their endorsement of traditional parenting views. Other demographic measures, such as family income-to-needs ratio maternal employment were collected when the child was approximately 1-, 6-, 15-, 24-, 36-, 54-months old (prekindergarten). Mothers were administered a personality questionnaire when the child was approximately 6-months old. Maternal receptive vocabulary was measured when the child was approximately 36-months old. Maternal sensitivity was observed and rated during a mother–child interaction task at five times (6-, 15-, 24-, 36-, 54-months old). All of the attention and self-regulation measures were administered when children were approximately 54-months old. The DNA protocol was conducted in adolescence, when children were approximately 15-years old. Buccal mucosa cells were collected with cotton swabs by the subject. DNA extraction was based on adaptations to the procedure outlined by Freeman et al. (2003).
Measures
Self-Regulation.
Inhibitory control was measured using three tasks: the Continuous Performance Test (CPT; Mirsky, Anthony, Duncan, Ahearn, & Kellam, 1991), a Day-Night Stroop task (Gerstadt, Hong, & Diamond, 1994), and a delay of gratification (DOG) task (Mischel, 1974). The CPT is a computer-delivered task in which the child views target and nontarget, pictorial stimuli on a computer screen and is required to follow two critical rules: (1) to press a response key when the target stimulus is presented on the screen, and (2) to withhold from pressing the response key when nontarget stimuli are presented on the screen. Children viewed illustrations of nontarget objects (e.g., butterfly, fish, flower) and were asked to respond only when the target item (a chair) was presented. Each stimulus appeared on the screen for 500 ms, with a 1,500-ms span between stimuli. Children were presented ten stimuli per block for 22 blocks; the target stimulus was presented randomly, twice per block. Similar to prior work with these data (NICHD ECCRN, 2003a), inhibitory control was operationalized as the number of commission errors (i.e., response to nontarget items) that the child made, scaled on the proportion of delivered trials. Higher scores indicate less effective inhibitory control. Natural logarithm transformations were adopted to adjust for skewness. The CPT shows reasonable test–retest reliability in nonclinical samples (r = .65–.74; Halperin, Sharma, Greenblatt, & Schwartz, 1991), and prior work with these data suggest convergent validity, such that children making more commission or omission errors on the CPT tend to show less effective achievement and language development (NICHD ECCRN, 2003a), as well as higher levels of attention and aggression problems (Berry, 2012; Campbell & von Stauffenberg, 2009).
Based on work by Gerstadt et al. (1994), the Day-Night Stroop task requires the child to downregulate a prepotent response in order to enact a subdominant response. Specifically, the child is presented 18 cards: half of the cards show an illustration of a white moon and stars on a black background, whereas the other half shows an illustration of a yellow sun on a white background. The child is directed to say the word “Day” when the examiner displays an illustration of the moon and to say “Night” when the examiner displays an illustration of the sun. The examiner established that the child understood the rules of the task in the first one or two trials. Inhibitory control was scored as the percentage of postpractice trials that were correct. Higher scores indicate more effective inhibitory control. The task shows reasonable internal consistency reliability (Cronbach’s alpha = .79), and good convergent validity (Montgomery & Koeltzow, 2010).
A DOG task was used to measure more desire-based aspects of inhibitory control. During the 54-month laboratory assessment, DOG was measured as children’s performance during a “waiting game.” The child was first consulted about his/her preferences for three possible food treats (i.e., M&Ms, animal crackers, or pretzels), as well as his/her preferences for small or large quantities of that particular treat. The child was subsequently presented with two plates, one containing a small pile of the child’s chosen treat and the other containing a large pile of the same treat. The examiner told the child that he/she had to leave the room for a while to do some work, but that, if the child waited until the examiner returned, the child would receive the larger pile of treats. If he/she could not wait, the child was instructed to ring a bell—the examiner would return, and the child would then receive only the small pile. Otherwise, the examiner waited 7 min. After 7 min the examiner re-entered the room and rewarded the child with the large pile of treats.
Given the clear bi-modal distribution in waiting time (at each tail), we operationalized DOG in two ways. First, we dummy-coded it as “pass-fail,” with one representing those who waited the entire 7-min span. Second, as there was notable variation in waiting times for those who did not wait the full 7 min, we considered the number of minutes delayed, for those who failed to wait the entire 7-min span. A natural log transformation was adopted to adjust for notable positive skew. Using these data, the DOG task (scored either dichotomously or continuously) shows convergent validity with other known measures of self-regulation (Tab. 1), and minutes waited on this task has shown predictive relations with broader behavioral measures of children’s attention problems (Campbell & von Stauffenberg, 2009).
Table 1.
Zero-Order (Maximum Likelihood) Correlations Between Child-Care Variables and the Attention and Self-Regulation Measures (n = 1,364)
| Inatt/Impuls | Stroop | DOG1 | DOG2 | CPT Comm. | CPT Omiss. | DRD4 7+ | CC Quant. | CC Qual. | CC Center. | |
|---|---|---|---|---|---|---|---|---|---|---|
| Stroop | −.08~ | 1.00 | ||||||||
| DOG 1 | −.20*** | .12*** | 1.00 | |||||||
| DOG 2 | −.07 | .09~ | N/A | 1.00 | ||||||
| CPT Comm. | .36*** | −.21*** | −.33*** | −.15*** | 1.00 | |||||
| CPT Omm. | .24*** | −.08* | −.27*** | −.08 | .38*** | 1.00 | ||||
| DRD4 7+ | −.01 | .08 | .03 | −.02 | −.09* | −.07~ | 1.00 | |||
| CC Quant. | .11** | −.03 | −.02 | .03 | .05 | .01 | −.01 | 1.00 | ||
| CC Qual. | −.17*** | .05 | .10** | .10* | −.22*** | −.13*** | .04 | −.19*** | 1.00 | |
| CC Center. | −.03 | .07* | .11*** | .12* | −.04 | −.08* | −.02 | −.07* | −.07* | 1.00 |
| Mean | — | 74.66 | .53 | −.57 | 1.63 | 1.42 | .25 | 22.95 | .02 | .26 |
| SD | .90 | 20.52 | .50 | 1.66 | 1.58 | 2.17 | .43 | 15.09 | .76 | .44 |
| Min | −1.29 | 12.50 | .00 | −3.62 | −4.61 | −4.61 | .00 | .00 | −2.60 | .00 |
| Max | 3.13 | 100.00 | 1.00 | 1.93 | 5.04 | 3.72 | 1.00 | 57.60 | 1.79 | 1.00 |
| Missingness | 0% | 39% | 29% | 67a% | 26% | 26% | 51% | 0% | 17% | 0% |
p < .10,
p < .05,
p < .01,
p < .001; two-tailed.
Missingness is partially by design; % includes only those failing to delay. Missingness based on overall task participation is 29%. Inatt/Impuls = latent teacher/observer rated inattentive/impulsive classroom behavior; Stroop = % correct on Day-Night Stroop task; DOG1 = fail (0)–pass (1) during a snack delay task; DOG2 = (log) minutes waited during snack delay task, for those failing; CPT Comm = (log) commission errors on Continuous Performance Test; CPT Omiss. = (log) omission errors on Continuous Performance Test; DRD4 7+ = at least one copy of a 7-repeat allele (1), versus other (0); CC Quant. = average weekly hours in child care between 1 and 54 months of age; CC Qual = average child-care quality between 6 and 54 months; CC Center = half or more of child’s time in care is in center-based care (1) versus less than half (0).
Inattention was measured multiple ways. The first task-based measure of cognitive inattention was operationalized as children’s performance on the CPT (see above). Similar to prior work with these data (NICHD ECCRN, 2003a) inattention was scored as the proportion of target items that were missed (i.e., omission errors). Higher scores indicate higher levels of inattention. Natural logarithm transformations were adopted to adjust for skewness.
To measure more broad-scale inattentive and impulsive classroom behavior, we created a latent composite across multiple measures and multiple raters. Specifically, informed by prior work by Kim, Deater-Deckard, Mullineaux, and Allen (2010), we adopted a Confirmatory Factor Analytic (CFA) approach to model children’s inattentive/impulsive behavior as a common latent factor theorized to explain children’s behaviors across 12 observed measures. Based on face validity, we selected nine items tapping inattention and impulsivity/hyperactivity from the (fully administered) attention problem scale of the Teacher Report Form (TRF; Achenbach, 1991). In addition, we included teachers’ reports on an 8-item version of the Attention Focusing subscale of the Child Behavior Questionnaire (CBQ; Rothbart et al., 2001). We also included independent observer ratings of children’s global classroom behaviors indicative of attentiveness to objects and activities in the classroom, as well as children’s overall activity levels, using the Observational Ratings of the Caregiving Environment scale (ORCE; NICHD ECCRN 1996). The questionnaire- and observer-based ratings were each based on Likert-type scales and the latent factor is scaled such that higher ratings represent more inattentive/impulsive behavior. All models were fitted using the robust maximum likelihood estimator available in Mplus (5.21; Muthén & Muthén, 2009). The within-measure residual covariances for the TRF and ORCE indicators, respectively, were freely estimated to account for method covariance.
The common-factor model fit the data well (χ2 = 25.12, df = 17, p = .09; CFI = 1.00: RMSEA = .02). All loadings were statistically significant, with the (absolute) standardized loadings ranging between .19 and .88, and there was a statistically significant variability in the latent factor (φ = .19, p < .001). An alternative two-factor representation of the model to represent inattention and impulsivity as distinct, yet correlated, factors did not improve model fit (Δχ2 = 2.01, df = 1, p = .16). We, thus, adopted the more parsimonious common-factor representation of inattention/impulsivity in our substantive analyses.
Child Care
Hours in Child Care.
Maternal reports of weekly average hours of nonmaternal care were collected by phone interview every 3 months across the first three years of life and every 4 months between 36 and 54 months. Average hours of child care were calculated as the mean hours of care per week between 1 and 54 months of age.
Quality of Child Care.
Quality of nonmaternal care was measured in the context of the caregiver–child interactions when the child was approximately 6, 15, 24, 36, and 54 months of age. Specifically, when the child attended 10 or more hours of nonmaternal care between the ages of 6 and 36 months or 8 or 12 hr of nonmaternal care (i.e., group-based and alone care, respectively) at 54 months of age, caregiver-child interactions were measured by independent observers using the ORCE scale (NICHD ECCRN 1996). At the 6- through 36-month observations observers made two half-day visits to children’s primary nonmaternal child-care setting within a 2-week span. At each visit they rated multiple aspects of caregiver behavior towards the child across two 44-min observation cycles. At the 6- through 36-month observations caregivers were rated on a 4-point Likert-type scale in regard to their sensitivity to child nondistress, stimulation of development, positive regard toward the child, detachment (reflected) and flatness of affect (reflected) and averaged to create a composite. At the 36-month observation two additional scales—fosters child’s development and intrusiveness (reversed)—were added to the composite. At the 54-month observation period, observations were done during one half-day visit. Caregiver behavior was averaged across five scales: sensitivity, stimulation of cognitive development, intrusiveness (reflected), and detachment (reflected). Across all of the composites, higher scores indicate higher quality care. Internal consistency reliability was reasonable at each time point (Cronbach’s alpha = .72–.89), as was inter-rater reliability assessed using either correlation or ANOVA based methods (.86 to .99). Quality was calculated by standardizing each scale averaging across the 6- to 54-month observations.
Type of Child-Care.
Child-care type was operationalized as high and low/no exposure to center-based care—that is, nonmaternal care provided in a formal facility/nursery that is in not the caregiver’s or the child’s residence. Specifically, of the average number of hours in care per week between 1 and 54 months, we calculated the proportion of those hours spent in center-based care. As the distribution showed substantial skew, we created a dummy-variable to represent those attending 50% or more of their time in center-based care (center = 1).
Genotype.
The assay for genotyping the DRD4 VNTR was based on methods developed Sander et al. (1998) and modified by Anchordoquy, McGeary, Liu, Krauter, and Smolen (2003). The Genomics Core Facility at the Pennsylvania State University, Huck Institute of the Life Sciences modified it further as the following: 1× Taq Gold Buffer, 2.25 mM final concentration of MgCl2, 10% DMSO, .2 mM dNTPs, .1 mM deazo GTP, .75 μM primers, 40 ng of DNA, and 1 U of Taq Gold (Applied Biosystems, Foster City, CA) in a volume of 12 μl. The primer sequences are: 5′−6-FAM-GCGACTACGTGGTCTACTCG-3′ and reverse, 5′-AGGACCCT-CATGGCCTTG-3′. The amplification procedure was as described by Anchordoquy et al. One microliter was removed and placed in a 96-well plate and 10 microliters of formamide containing LIZ-500 standard (Applied Biosystems). The plate was run using a Fragment Analysis protocol in the 3730XL DNA Analyzer (Applied Biosystems). Fragments were analyzed using Genemapper software (Applied Biosystems) with PCR products of (in bp): 379, 427, 475, 523, 571, 619, 667, 715, 763, and 811 (i.e., corresponding to the 2–11 repeat alleles, respectively). Reliability analyses were conducted by re-genotyping a second time. Discrepancies were resolved via a third genotyping. In rare cases (n = 27) in which samples became exhausted, such that calls could not be made on the second or third genotyping run, the original genotype was adopted. DRD4 7+ was coded as those who were either homozygous or heterozygous for the 7-repeat allele (1; n = 168) versus those without a single copy of the 7-repeat allele (0; n = 498). Repeat alleles greater than 7 were coded as 7-, based on prior functional work indicating that higher-number repeats (e.g., 10-repeat) show functional properties that are more similar to lower number repeats (e.g., 2-repeat) than to 7-repeats (Jovanovic, Guan, & Van Tol, 1999). There was 83% agreement (κ = .53, p < .001) for the 7+ genotype across genotyping runs, suggesting reasonable reliability.
Exact tests using Markov chains were estimated to test Hardy–Weinberg equilibrium (HWE) for our genotypes of interest (i.e., 7+ vs. 7−; GENEPOP 4.0; Raymond & Rousset, 1995). Given the diverse and geographically stratified nature of the sample, there was a somewhat unsurprising indication that the frequency of the 7+ genotype varied across research site (χ2 = 13.96, df = 7, p = .05). The 7+ genotype ranged from 19% to 35% across sites. Although there was an indication of deviation from HWE, prior to accounting to race and site stratification (p = .002), there was no deviation in HWE for European American children, after adjusting for site (combined across site [Fisher’s method] χ2 = 19.74, df, = 16, p = .23; Raymond & Rousset, 1995). Research site and race were included as controls, along with several other covariates.
Control Covariates.
In an attempt to adjust for the effects of selection bias, we included an extensive array of child (sex, race, gestational age, temperament), maternal (education, personality, separation anxiety, traditional parenting views, benefits of employment), and family (income, employment status, research site) control covariates. Description of each of these variables/measures is provided in Appendix.
Missing Data
Of the total sample, missing data ranged from 0% to 51%, the latter representing the fact that two of the original ten sites did not participate in the genetics protocol. Of those who did participate, 45 children could not be genotyped due to low-quality DNA samples. In an attempt to adjust for biases introduced by missing data, we used full-information maximum likelihood (FIML) estimation methods. FIML estimation helps to adjust for these biases, under the assumption that missing data is “missing at random”—that is, missingness is conditional on observed variables that are included in the model and, after adjusting for these variables, are not conditional on unobserved values of the variables with missing data (Collins, Schafer, & Kam, 2001). We included several control covariates that provide information about the variables of substantive interest, as well as missingness, as predictors in the models (Allison, 2003). As such, all individuals providing any information to the model were included in model estimation. Based on the variables included in our model, all 1,364 children provided some information to the model and were therefore included in the analyses.
Data-Analytic Plan
To address our research questions regarding: (1) the degree to which child-care quantity, quality, and type were predictive of children’s prekindergarten self-regulation and attention skills, and (2) the degree to which these relations were conditional on genotype, we fitted taxonomies of multiple regression models using the robust maximum likelihood estimator in Mplus 5.21 (Muthén & Muthén, 2009). Specifically, we first tested the main effects of child-care quantity, quality and type, and DRD4 genotype on each of the self-regulation outcomes. We then allowed the effects of each of these (nongenetic) predictors to vary as a function of DRD4 genotype by including the respective interaction terms in the model. To adjust for family-wide false discovery rates, Benjamini and Hochberg (1995) corrections were made for each family of tests (i.e., given variable, or interaction term) across the 6 modestly correlated outcomes. The reported p-values marked as “pbh “ reflect these corrections and were calculated using the “multtest” function in the R 2.15.1 statistical program (R Core Team, 2012). We present the most parsimonious “final” models from this taxonomy in Table 2. Model constraints were used to test the simple slopes for all statistically significant interactions. The conditional genetic effects were tested at the 20th (~7 hr) and 80th (~38 hr) percentiles, given its nonnormal distribution.
Table 2.
Fitted Unstandardized Regression Parameters From Six Models Testing the Degree to Which G × E Interactions Between DRD4 7+ Genotype and Child-Care Experiences Are Associated With Children’s Attention and Self-Regulation Skills in Prekindergarten (n = 1,364)
| Stroop | DOG1a (0–1) | DOG2 (#min) | CPT Comm. | CPT Omiss | Inattent/Impuls | |
|---|---|---|---|---|---|---|
| Intercept | 72.76*** | .05 | −.70 | 2.13*** | 2.10*** | — |
| DRD4 7+ | 3.61~ | .17 | .08 | −.25~ | −.15 | −.04 |
| CC Quant | .02 | .01 | .01 | .01 | −.01~ | .00 |
| CC Qual | .00 | −.05 | .19 | −.20*** | −.07 | −.04 |
| CC Center | 1.75 | .41 | .53* | −.09 | −.12 | −.03 |
| Quant × D4 | −.36* | −.04* | .02** | .01** | ||
| Child covariates | ||||||
| Male | −2.14 | −.20 | −.20 | .68*** | .07 | .12*** |
| African American | 5.22* | −1.59** | .30 | .21 | .04 | −.07 |
| Gest. age | .29 | .04 | .02 | −.04 | −.02 | .02* |
| Temperament | 1.68 | .32 | −.17 | .02 | .05 | .08* |
| Birth order | −.88 | .04 | −.08 | .02 | .04 | .02 |
| Maternal/family covariates | ||||||
| Mat. Sens | 1.14 | .71** | .13 | −.14* | −.21*** | −.10** |
| Sep. Anx. | .01 | .01 | −.01~ | .00 | .00 | .00 |
| Income | .43 | .10 | .04 | −.01 | .01 | .00 |
| Education | .51 | −.04 | −.04 | −.04* | −.01 | −.02* |
| Married | 2.70 | .39 | .32 | −.34** | −.09 | .01 |
| Vocab | .05 | .02* | −.00 | .00 | .00 | .00 |
| Neuroticism | −.15 | .00 | .01 | .00 | .00 | .00 |
| Extroversion | −.04 | −.01 | .01 | .01 | −.01 | .00 |
| Agreeableness | −.01 | .01 | .03~ | .00 | .00 | .00 |
| Trad. parent | .00 | .01 | .00 | .01 | .00 | .00 |
| Ben. of work | −.05 | −.02 | .03 | −.04** | .01 | .02** |
| Age | −.07 | .01 | .03 | .00 | .00 | .00 |
| R2 | .08 | — | .15 | .26 | .12 | .19 |
Logistic regression model (n = 498);
p < .10,
p < .05,
p < .01,
p < .001; two-tailed; Inatt/Impuls = latent teacher/observer rated inattentive/impulsive classroom behavior; Stroop = % correct on Day-Night Stroop task; DOG1 = fail (0)–pass (1) during a snack delay task; DOG2 = (log) minutes waited during snack delay task, for those failing; CPT Comm = (log) commission errors on Continuous Performance Test; CPT Omiss. = (log) omission errors on Continuous Performance Test; DRD4 7+ = at least one copy of a 7-repeat allele (1), versus other (0); CC Quant. = average weekly hours in child care between 1 and 54 months of age; CC Qual = average child-care quality between 6 and 54 months; CC Center = half or more of child’s time in care is in center-based care (1) versus less than half (0). Dummy variables representing research site were also included as covariates in all models.
RESULTS
Preliminary Analyses
Across the first 54 months of life, children spent approximately 22.95 hr in child care per week, on average. Just over a quarter of the children (27%) were in ten or fewer hours of care per week, on average, and over a third of the children (37%) were in 30 or more hours of child care per week. Although the average hours per week spent in child care tended to increase toward later toddlerhood and early childhood years, the rank-order stability in hours-of-care was quite strong. For example, the correlation between average hours in care in the first 36 months of life and the average hours per week in child care between 42 and 54 months of age was .69 (p = .001). Approximately a quarter of the children never attended center-based care (24%); this includes the small number of children in exclusively maternal care (e.g., 6% attended 1 or fewer hours of care per week, on average). A slightly larger number of children (26%) spent half or more of their hours in care in center-based care. Children spending more hours in child care and those spending a greater proportion of their time in center-based care tended to experience slightly lower quality care, on average; however these correlations were rather modest, −.19 (p < .001) and −.07 (p = .02), respectively.
As shown in Table 1, the correlations among the outcome variables were typically statistically significant and always in the expected direction, given that they have been discussed collectively as important aspects of self-regulation. However, the modest magnitudes of the relations suggested that there were also be meaningful differences across these measures. Indeed, attempts at data reduction (available from the first author upon request) indicated that each of the behavioral tasks was best represented as an observed variable, rather than as a latent composite across all or a subset of (i.e., inhibitory control) measures/tasks. Zero-order correlations between the child-care variables and the self-regulation outcomes were somewhat inconsistent and modest in magnitude. DRD4 7+ children made significantly fewer commission errors (r = −.09, p = .04), and, at a marginal level of significance, made fewer omission errors on the CPT (r = −.07, p = .09). Notably, genotype type was unassociated with any of the child-care variables.
Child Care, Inhibitory Control, and Inattention
Day-Night Stroop.
Preliminary models indicated that there were no main-effect associations between any of the child-care variables and performance on the Day-Night Stroop task, after adjusting for the other covariates in the model. Subsequent models testing interactive effects showed that the respective effects of child-care type and quality did not show statistical interactions with genotype. However, there was evidence of a statistically significant interaction between DRD4 genotype and child-care quantity (Tab. 2; B = −.36, pbh = .02, ΔR2 = .01).
As displayed in Figure 1, DRD4= 7+ children who spent fewer hours in nonmaternal care tended to show more effective performance on the Stroop task than did their DRD4 7+ peers who spent greater hours in child care, on average (solid slope; BDRD4_7+ = −.35, p = .003, β = −.25). The association for those without a copy of the DRD4 7-repeat allele (dashed slope) was not statistically significant (BDRD4_7− = .02, p = .78, β = .01). Although there was a descriptive indication of a “cross-over” pattern, such that the direction of the conditional genetic effect varied at either end of the child-care-hours distribution, the conditional genetic effect only reached statistical significance at the low end of the distribution (20th percentile; Blow_hours = 9.44, p < .001; β = .46). That is, the “beneficial” effect of the DRD4 7+ was manifest in the context of low hours of child care, yet this same genotype did not appear to be a pronounced risk factor in the context of more extensive hours of child care (e.g., 80th percentile Bhigh_hours = −2.01, p = .53; β = −.10). Collectively, the final model explained approximately 8% of the variation in children’s Stroop performance. Approximately 24% of this explained variation was due to individual differences in the children’s child-care experiences and genotypes.
FIGURE 1.
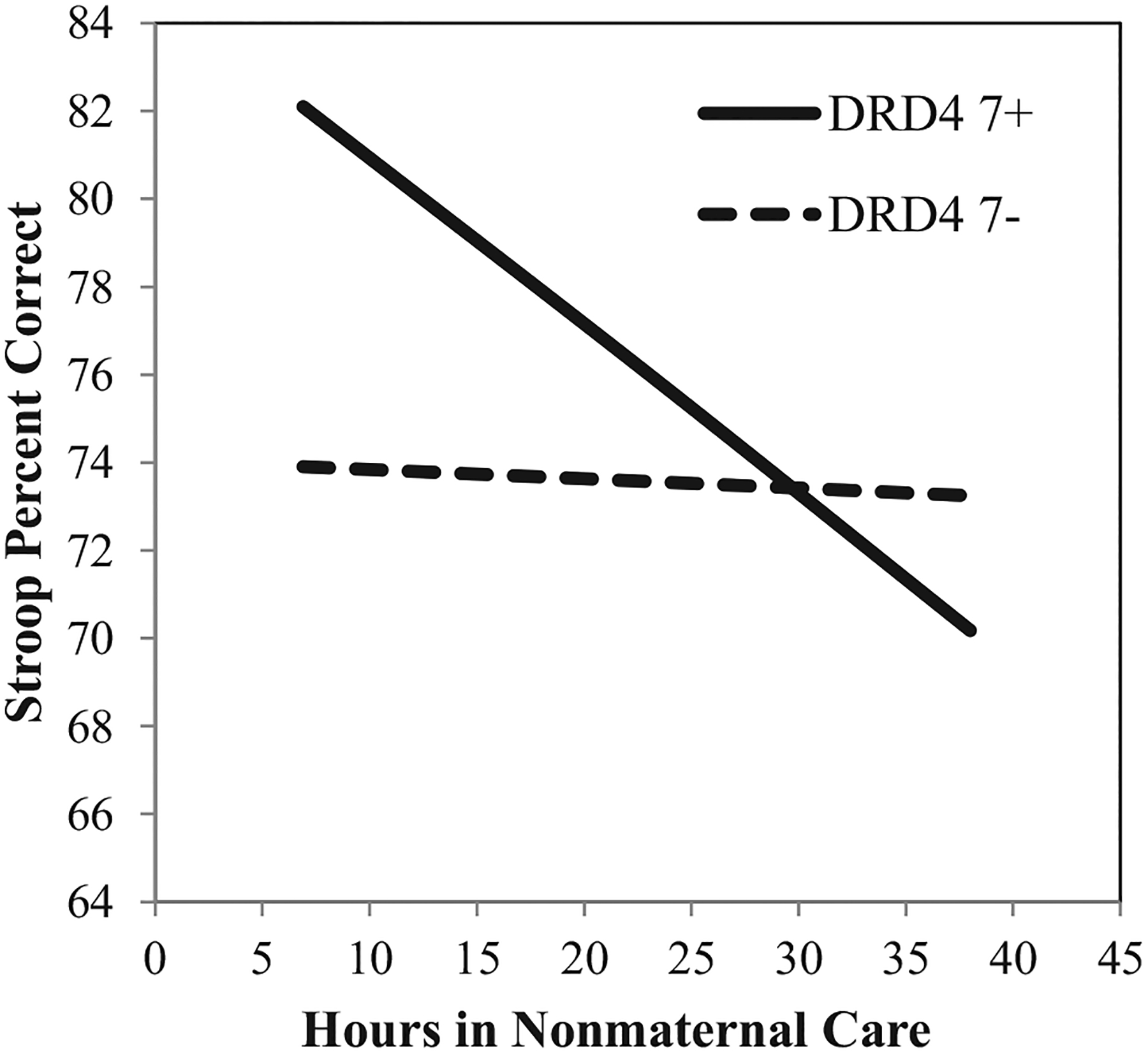
Estimated association between average hours in nonmaternal care between the ages of 1 and 54 months and performance on the Day-Night Stroop taks in prekindergarten, conditional on DRD4 genotype. All other covariates are held at their respective means. Note: Y-axis equals 1 standard deviation.
Delay of Gratification.
Logistic regression analyses using the pass-fail scoring of the DOG task indicated that there were no main-effect associations between any of the child-care variables or genotype with successfully passing the delay task (Tab. 2), nor were the respective effects of child-care quality, quantity, or type moderated by genotype.
When DOG was operationalized as the amount of time waited (for those not passing), the estimates from preliminary regression models suggested that attending center-based care was associated longer delays (i.e., better inhibitory control), after adjusting for the control covariates. There were no main-effect relations for child-care quality or quantity. Subsequent models testing the respective interactive effects showed that neither the effects of child-care type nor quality interacted with genotype. There was, however, evidence of a statistically significant interaction between DRD4 genotype and child-care quantity (Tab. 2; B = −.04, pbh < .05, ΔR2 = .01).
As displayed in Figure 2, for DRD4 7+ children, fewer hours in child care were associated with longer delay times—that is, better inhibitory control (BDRD4_7+ = −.03, p = .02). This corresponded to a Standardized simple slope of approximately −.28. For those without a copy of the DRD4 7-repeat allele, there was no relation between child-care hours and delay (B = .005, p = .43, β = .05). Although there was a descriptive indication of a cross-over interaction, probes of the conditional genetic effects indicated that—at marginal levels of statistical significance—the DRD4 7+ was associated with delay times only for those attending few hours in child care (20th percentile; Blow_hours = .64, p = .07; β = .39). Genotype was not statistically associated with delay times for those attending high hours of care (80th percentile; Bhigh_hours = −.47, p = .15; β = −.28).
FIGURE 2.
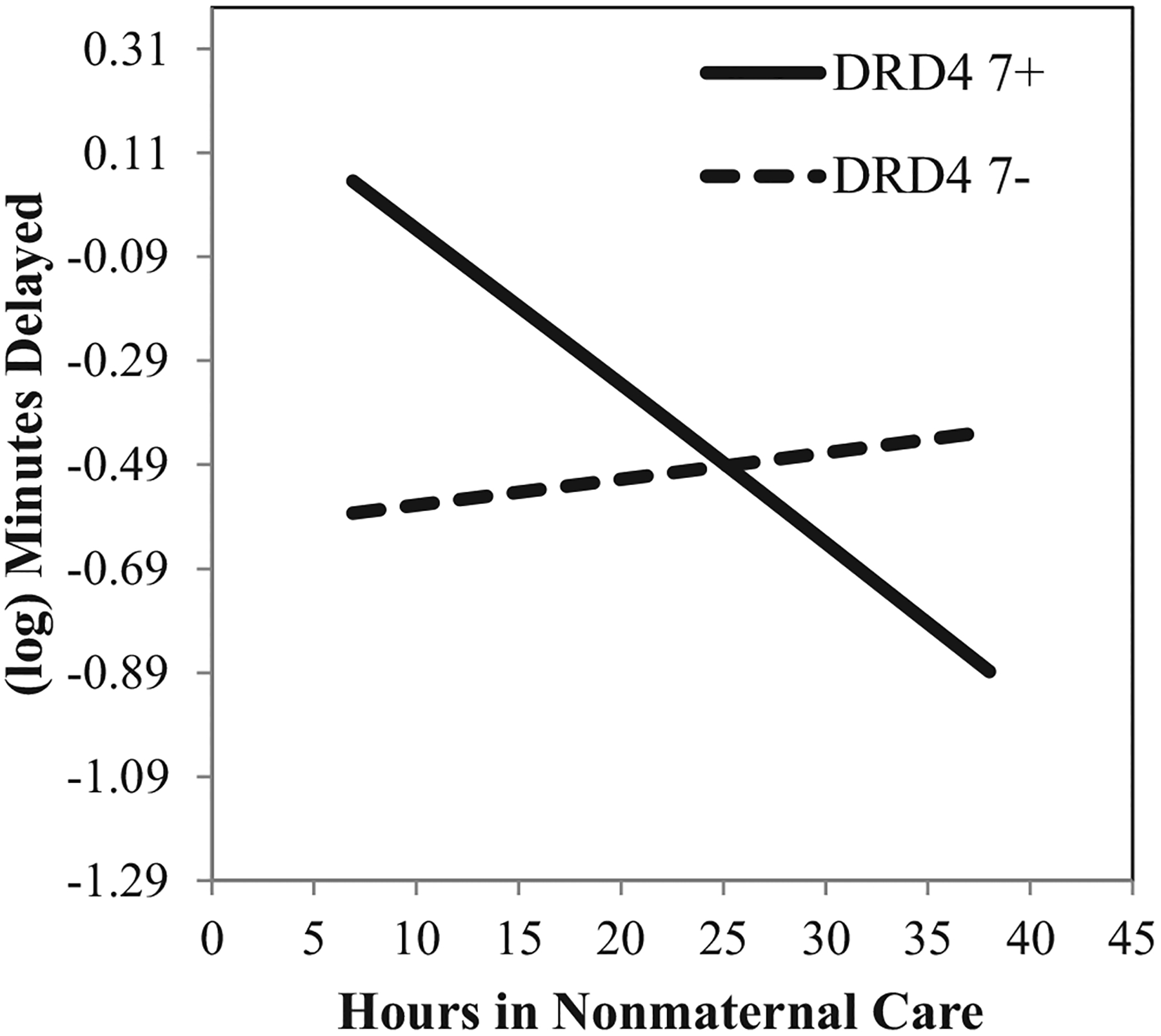
Estimated association between average hours in nonmaternal care between the ages of 1 and 54 months and (log) minutes delayed during a Delay of Gratification task (nonpassers only) in prekindergarten, conditional on DRD4 genotype. All other covariates are held at their respective means. Note: Y-axis equals 1 standard deviation.
After adjusting for the other covariates in the model, attending greater or equal to half of ones time in child-care in center-based care was associated with longer delays (B = .53, pbh .01; β = .32). There was also some indication that higher quality care was predictive of longer delays; however, this statistically marginal association (B = .19, p = .06; β = .10) dropped to statistical nonsignificance after adjusting family-wide false-discovery rate (pbh = .14). Collectively, the final model explained approximately 15% of the variation in children’s delay times. Approximately 26% of this explained variation was due to individual differences in child-care experiences and genotypes.
Continuous Performance Test—Commission Errors
Tests of the main effects indicated a statistically significant association for child-care quality (B = −.20, pbh < .01; β = .13) and a statistically marginal association for genotype (B = −.25, pbh = .06; β = .29), after adjusting for the control covariates and false-discovery rate. Higher quality care was associated with fewer commission errors on the CPT (i.e., better inhibitory control). Child-care type and quantity were unassociated with CPT commission errors. There was no evidence that the respective effects of child-care quantity, quality, or type were moderated by genotype. Collectively, the model explained 26% of the variation in children’s commission errors on the CPT. Approximately 11% of this explained variation was due to individual differences in the children’s child-care experiences and genotypes.
Continuous Performance Test—Omission Errors
Preliminary models indicated that center-based and higher quality child care were associated with fewer omission errors (i.e., better attention) after adjusting for the control covariates. Subsequent models showed that neither relation was moderated by genotype. Although preliminary models indicated that there was no main-effect relation between child-care quantity and omission errors on the CPT, subsequent tests of the interactions showed a statistically significant G × E interaction between DRD4 genotype and child-care quantity (B = .02, pbh = .009, ΔR2 = .01).
As displayed in Figure 3, fewer hours in child care were associated with fewer omission errors, for DRD4 7+ children (solid slope; BDRD4_7+ = .01, p = .01, β = .19). In contrast, there was some indication of an inversed relation for children without a copy of the DRD4 7-repeat allele. This negative simple slope, however, only approached statistical significance (B = −.005, p = .05, β = .08). The parallel interpretation of the interaction indicated that the DRD4 7+ genotype was associated with more effective attention skills when these children spent low hours in care (20th percentile; Blow_hours = −.39, p < .001, β = −.47). In the context of more extensive hours of care (80th percentile), however, DRD4 7+ children were statistically indistinguishable from their peers without a copy of the DRD4 7-repeat allele (Bhigh_hours = .09, p = .35, β = .09).
FIGURE 3.
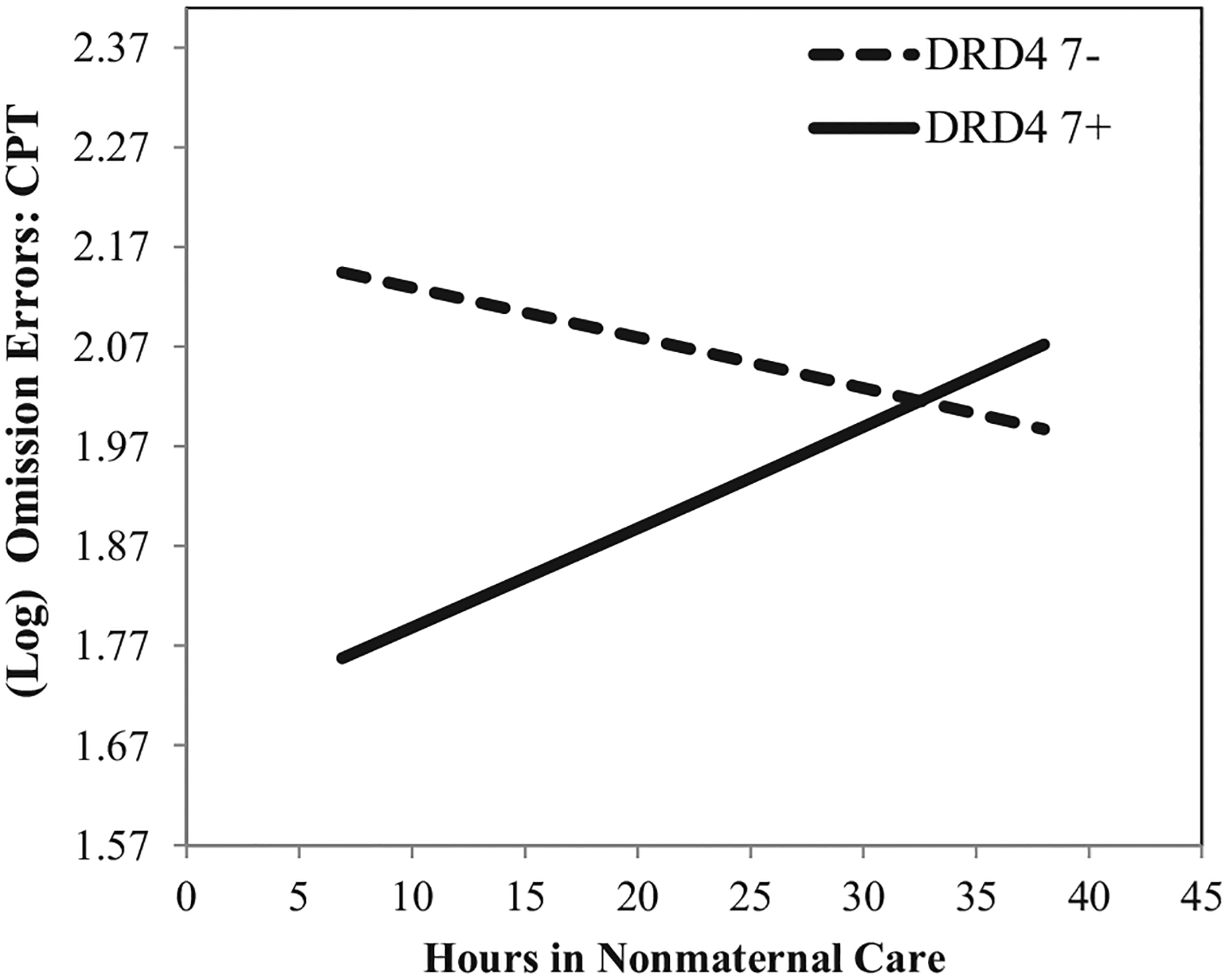
Estimated association between average hours in nonmaternal care between the ages of 1 and 54 months and (log) number of omission errors on the Continuous Performance Test (CPT) in prekindergarten, conditional on DRD4 genotype. All other covariates are held at their respective means. Note: Y-axis equals 1 standard deviation.
After adjusting for this G × E interaction and the other covariates in the model, there was some indication that children spending half or more of their time in center-based care (B = −.12, p = .04, β = −.15) and higher quality care (B = −.07, p = .07, β = −.09) tended to show fewer CPT omission errors, on average. Notably, both relations were statistically nonsignificant, after adjusting for false-discovery rate (pbh = .11 and pbh = .14, respectively). Collectively, the final model explained approximately 12% of the variation in children’s omission errors on the CPT. Approximately 21% of this explained variation was due to individual differences in the children’s child-care experiences and genotype.
Inattention and Impulsivity.
Preliminary models indicated that there were no main-effect associations between child-care quality or type with latent Inattention/Impulsivity. Subsequent models showed that the respective effects of child-care quality and type were also not moderated by genotype. Although preliminary models suggested a positive main-effect association between child-care quantity and higher levels inattentiveness/impulsivity, subsequent models indicated that this effect was moderated by genotype (Tab. 2; B = .01, pbh = .009, ΔR2 = .01).
As displayed in Figure 4, the positive relation between child-care hours and heightened inattentive/impulsive behavior was particularly pronounced for DRD4 7+ children (BDRD4_7+ = .01, p < .001, β = .33). The same relation for those without a copy of the DRD4 7-repeat was not statistically significant (BDRD4_7− = .001, p = .62, β = .03). At a descriptive level, the interaction appeared to show a cross-over effect. However, in statistical terms, the true cross-over effect only approached traditional levels of statistical significance. Similar to several of the other outcomes, the DRD4 7+ genotype was associated with significantly lower levels of inattention/impulsivity, for children attending low hours of nonmaternal care (Blow_hours = −.18, p = .004, β = −.40). In contrast, in the context of high hours of nonmaternal care, the DRD4 7+ genotype was only descriptively (but not statistically) associated with the higher levels of inattention/impulsivity (Bhigh_hours = .10, p = .14, β = .22). Collectively, the final model explained approximately 19% of the variation in children’s omission errors on the CPT. Approximately 13% of this explained variation was due to individual differences in the children’s child-care experiences and genotypes.
FIGURE 4.
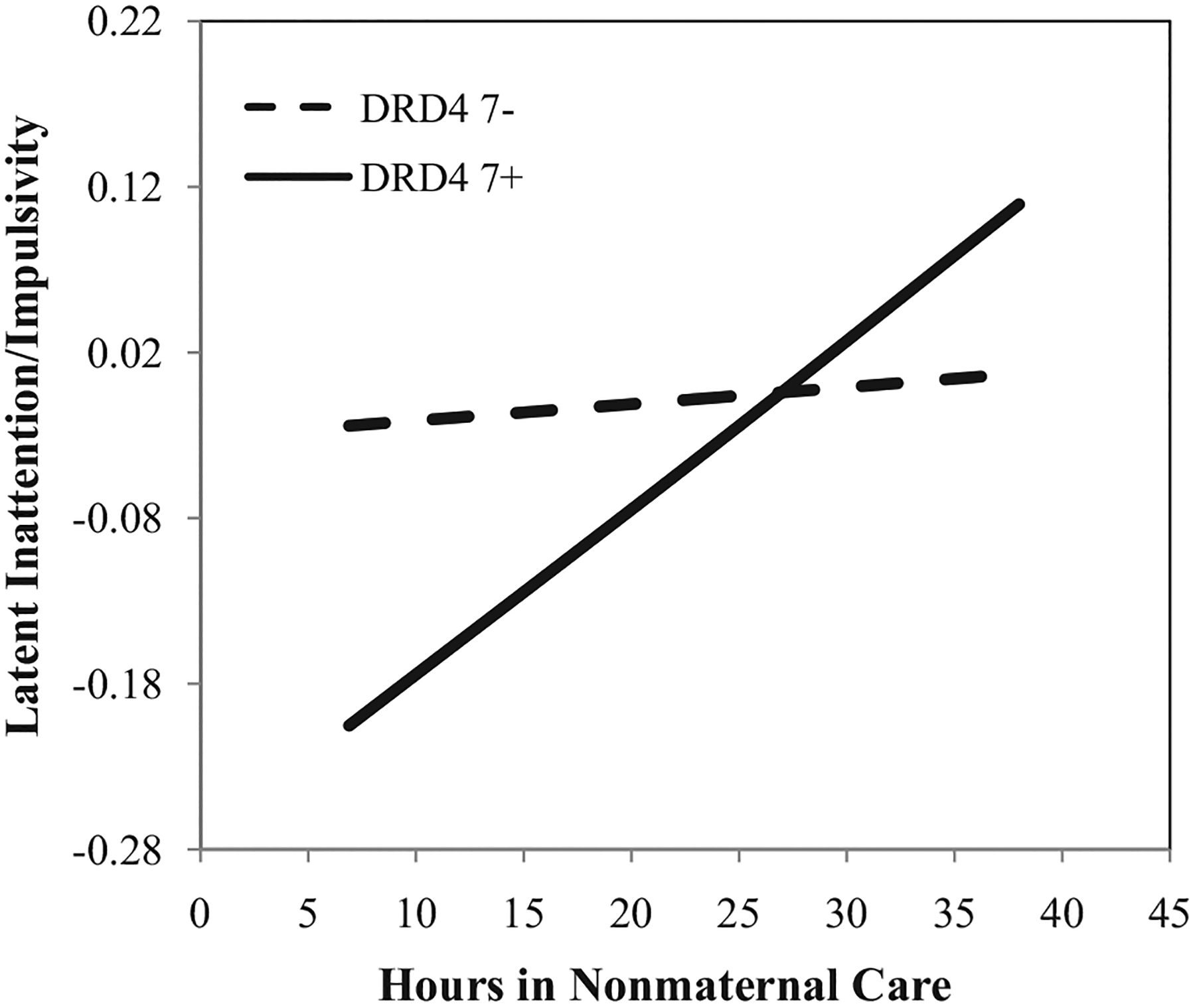
Estimated association between average hours in nonmaternal care between the ages of 1 and 54 months and latent inattention/impulsivity in prekindergarten, conditional on DRD4 genotype. All other covariates are held at their respective means. Note: Y-axis equals 1.5 standard deviations.
In a series of follow-up analyses for each outcome, we subsequently tested the extent to which the respective relations between child-care quantity and type and the given self-regulation outcome were conditional on “dose” (i.e., two-way interaction with child-care quantity) and whether such two-way interactions were, perhaps, particularly pronounced for those with a DRD4 7+ genotypes. No such higher-order interaction emerged at traditional or adjusted levels alpha levels. Also, given the (albeit, mixed) evidence linking the DRD4 7+ allele to aspects and temperament (see Saudino & Wang, 2012), we tested the impact of the temperament control covariate on the G × E effects, as well as potential higher-order interactions between temperament, DRD4 7+, and children’s child-care experiences. Temperament was unassociated with the DRD4 7+ genotype, showed typically null relations with the outcomes in the full models, had no meaningful effect on any G × E finding, and showed no higher-order interactions with genotype and children’s child-care experiences at traditional or adjusted alpha levels.
DISCUSSION
The dual aims of the present study were to model the degree to which children’s experiences in child care were associated with their development of attention and self-regulation skills in prekindergarten, and to test whether these respective relations were conditional on DRD4 genotype. Parallel with prior work considering substantively related outcomes (e.g., behavior problems), we hypothesized that less extensive child-care exposure and higher quality care would be associated with more effective attention and self-regulation skills. Informed by a growing G × E literature, we predicted also that these relations would be particularly pronounced for children with the DRD4 7+ genotype. Our findings provided partial support for these hypotheses. The G × E effects were most consistent with regard to child-care quantity, showing quite similar interactions with respect to performance on the Stroop, DOG (minutes waited), CPT (omission errors) tasks, as well as teacher/observer ratings of inattention/impulsivity in the classroom. In contrast, the respective associations between child-care quality and type with children’s self-regulation abilities were not moderated by genotype, and the main-effect associations were somewhat inconsistent across the outcome measures.
G × E: Child-Care Quantity, Attention, and Self-Regulation
The G × E interactions between child-care quantity and DRD4 genotype were fairly consistent across multiple neuropsychological and behavioral measures of attention and self-regulation. For DRD4 7+ children, fewer hours in nonmaternal care were associated with more effective attention skills in prekindergarten, as measured by omission errors on the CPT, as well as teacher/observer ratings of children’s inattentive and impulsive behavior in the classroom. Similarly, for DRD4 7+ children, less extensive hours in child care were also associated with better inhibitory control, as measured by a Stroop and a delay-of-gratification task. Across the outcome measures, child-care quantity was unassociated with children’s attention and self-regulation abilities, for children without a copy of the DRD4 7-repeat allele.
Although the effects of child-care quantity extended only to DRD4 7+ children, these associations are somewhat consistent with prior work considering associations between child-care quantity and developmental outcomes associated with self-regulation. The findings from multiple studies using this sample (Belsky et al., 2007; McCartney et al., 2010; NICHD ECCRN, 2003b) as well as others (Loeb et al., 2007; Magnuson et al., 2007) have suggested that heightened exposure to child care (or attendance versus nonattendance) in infancy and early childhood may be predictive of problematic social behavior.
Broad behavioral patterns of externalizing behaviors (Hughes & Ensor, 2011; Séguin & Zelazo, 2005) and particularly inattention and hyperactivity (Barkley, 1997; Willcutt, Doyle, Nigg, Faraone, & Pennington, 2005) may reflect, in part, difficulties with executive aspects of self-regulation. The present findings with respect to the neuropsychological measures of attention and cognitive self-regulation are consistent with this possibility. That is, executive self-regulatory processes are perhaps one mechanism through which heightened exposure to child care may affect broad behavioral patterns of inattention, hyperactivity, and aggression. Although the findings for child-care quantity (for DRD4 7+ children) are consistent with prior work, the experiential mechanisms underlying these conditional associations remain unclear. Indeed, like prior work with substantively related outcomes (NICHD ECCRN, 2003b), links between child-care quantity and children’s attention and self-regulation abilities emerged after adjusting for quality and type.
There are potential mechanisms other than child-care quality and type that were not measured in the present study that may explain the conditional relation between child-care quantity and these developmental outcomes. For instance, children in fewer hours of nonmaternal care may have greater exposure to experiences, such as one-on-one scaffolding and mind-minded dialogue, which may help to support more effective attention and self-regulation skills (Bernier et al., 2010; Carlson, 2003). Such highly individualized caregiver–child exchanges may be comparatively less available in child-care contexts in which caregivers’ social resources must be divided across many children.
G × E: Child-Care Quality and Child-Care Type
Links between the quality of children’s child-care experiences and their development of attention and self-regulation skills were rather weak and somewhat inconsistent across outcome measures. A statistically significant association emerged only for children’s commission errors on the CPT. Inconsistent findings were evident also for the effects of child-care type. Spending half or more of one’s time in child-care in center-based care was associated with longer delays of gratification for those not passing the delay task. Yet, attending more center-based care was unassociated with the other measures of attention and self-regulation.
Given a growing body of empirical work suggesting that children’s attention and executive self-regulation abilities play an important role in their development of early academic skills (Blair, 2002; Blair & Razza, 2007; McClelland et al., 2007), and prior work indicating that high quality (Barnett, 1995; Belsky et al., 2007; Campbell et al., 2002; NICHD ECCRN & Duncan, 2003) and center-based (Loeb et al., 2007; Magnuson et al., 2007; NICHD ECCRN & Duncan, 2003) care experiences are associated with higher levels of academic achievement, these null results were somewhat unexpected.
This may be explained, in part, by the nature of the sample. For instance, some evidence suggest that the effects of center-based care experiences may be particularly pronounced for less affluent children, compared to their peers from more affluent families (Loeb et al., 2007). Although socio-economically diverse, the present sample is comparatively more affluent, on average, than are the populations considered in many high-quality child-care interventions (e.g., Campbell et al., 2002; Raver et al., 2012; Schweinhart et al., 2005). Understanding such nonlinear effects across populations and absolute levels of quality have important policy implications and remain critical questions for future work.
G × E: Differential Susceptibility
The overall finding that the magnitude of these relations was much stronger for children with the DRD4 7+ genotype is somewhat consistent with a growing G × E literature. As reviewed in Bakermans-Kranen-burg and van IJzendoorn’s (2011) meta-analysis, several studies have shown that the association between children’s early experiences and developmental outcomes related to attention and self-regulation are often limited to DRD4 7+ children.
Notably—in addition to the pronounced environmental effect predicted for those with the DRD4 7+ genotype—the differential-susceptibility model also posits that the direction of the DRD4 7+ effect is conditional on experience. In terms of the present study, the DRD4 7+ is predicted to be associated with the most effective attention and self-regulation skills in the context of low child-care hours, yet this same DRD4 7+ marker is predicted to be associated with the least effective attention and self-regulation skills, in the context of long child-care hours.
Although there were some descriptive indications of such cross-over interactions in the present findings, in statistical terms, the direction of the conditional genetic effect did not change for those in low (e.g., ~7) compared to high (e.g., ~38) hours of nonmaternal care. Instead, the general trend of the interactions was such that the “beneficial” relation between DRD4 7+ genotype and more effective attention and self-regulation skills extended only to those in lower hours of child care. The magnitude of this conditional relation became increasingly weak as the number of child-care hours increased, statistically reaching zero toward the high end of the child-care-hours distribution. As such, in the present data, the DRD4 7+ marker was not a “risk” factor in the context of high hours of child care, as expected by differential susceptibility. This may be explained in part by the possibility that, for DRD4 7+ children, heightened exposure to child care may limit their exposure to experiences that support self-regulation (e.g., one-to-one scaffolding), without increasing their exposure to environments that are particular deleterious to self-regulation development. Ultimately, however, this remains an unanswered question for future work.
Interestingly, though the present data may fail to align completely with the prediction of the differential susceptibility model, per se, they also fail to align with idea of diathesis-stress—an alternative theoretical model typically pitted against differential susceptibility (Belsky & Pluess, 2009). Specifically, genetic differences are theorized to function as diatheses—or risk factors—that become expressed in the context environmental stressors. Notably, in the present findings, the potential genetic diathesis (i.e., DRD4 7+) did not function as a risk factor that became manifest in the context of an environmental stressor. Rather, the DRD4 7+ seemingly placed carriers at an attentional/self-regulatory advantage that became systematically less advantageous in the context of greater hours of child care.
Irrespective of degree to which the present findings align completely with the differential susceptibility framework, they, along with the findings from prior studies, suggest that modeling G × E processes may be critical to understanding of the roles of genes or experience in children’s development. Across the multiple self-regulation measures considered, the respective conditional associations between child-care quantity and DRD4 7+ genotype with children’s self-regulation abilities would have typically been missed, had the G × E effects not been modeled explicitly.
Limitations and Future Directions
An obvious limitation of the present study is the potential endogeneity of the key question predictors, including passive and evocative gene–environment correlation. We controlled statistically for a wide array of factors associated with selection into child-care quantity, quality level, as well as type and genetic stratification across site and race. Although maternal genotype was unavailable, we also attempted to control for passive gene–environment correlation by including measures of maternal personality that may partially reflect DRD4 genotype in all models (Schinka, Letsch, & Crawford, 2002). There may, however, be unobserved or unobservable variables that could have biased the present estimates. Randomized experimental designs are certainly warranted. Interestingly, emerging work suggests that even intervention studies adopting experimental designs may benefit from considering G × E effects (Bakermans-Kranenburg et al., 2008; Brody, Beach, Philibert, Chen, & Murry, 2009).
The mechanisms underlying the G × E findings also remain unclear at multiple levels. Firstly, the biological pathways linking the DRD4 7+ repeat with heightened sensitivity—across the G × E literature, as well as those specific to this study—are unclear and remain pressing areas of future research. Secondly, as noted above, the ways in which heightened exposure to child care “gets under the skin” to affect children’s attention and self-regulation development remain unknown. Although the present findings suggest that child-care quality and type may play a partial role for some aspects of attention and self-regulation, clarifying the processes through which heighted exposure to nonmaternal care impacts self-regulation has important implications for practice and policy.
Despite these limitations, the present findings suggest that G × E processes between the DRD4 gene and children’s child-care experiences may explain meaningful individual differences in children’s developing attention and self-regulation skills—measured at both the neuropsychological and broad-behavioral levels—and add to a growing literature suggesting that understanding the roles of genes or experience in development may rest on the explicit characterization of G × E processes.
Acknowledgments
This work was supported in part by Grant HD25451 from the National Institute of Child Health and Human Development (NICHD) Study of Early Child Care and Youth Development (to Kathleen McCartney). It was also supported by a Julius B. Richmond Fellowship (Harvard University Center on the Developing Child) and a Graduate Student Research Grant (Harvard University Mind/Brain/Behavior Initiative; to Daniel Berry), as well as by a Research Board grant from the University of Illinois at Urbana-Champaign (to Philip C. Rodkin and Glenn I. Roisman).
DNA extraction and genotyping was performed at the Genome Core Facility in the Huck Institutes for Life Sciences at Pennsylvania State University under the direction of Dr. Deborah S. Grove, Director for genetic analysis. Finally, this research would not have been possible without the inspired work of the NICHD Early Child Care Research Network and research staff, who designed and conducted the overall study, or without the dedicated children, families, and teachers who participated.
Appendix: DESCRIPTIONS OF CONTROL COVARIATES
Child Covariates
Race: We included child race as a control covariates. Because the within-race group sizes were too small for the Latino, Asian, and Other designations, we created a dummy variable to represent ethnicity as African-American versus all other races.
Gender: Boys were coded as one.
Birth Order: Birth order was coded as “1” is first child, “2” is second child, etc.
Gestational Age: Gestational age is scaled in weeks and based on actual birth date and maternal reported due date.
Temperament: Infant temperament scores were based on maternal reports on an adaptation of the Infant Temperament Questionnaire (Carey & McDevitt, 1978), when the child was approximately 6-months old. Higher scores indicate more “difficult” temperaments. The global scale used in the present study shows good internal consistency reliability (alpha = .81).
Maternal/Family Covariates:
Family Income: Annual family income was reported across the five time-points of interest in the present study (i.e., 6-, 15-, 24-, 36-, and 54-months old). At each time-point, family income-to-needs ratio was calculated by dividing family income by the U.S. Census Bureau poverty threshold for the appropriate family size. We average income across the five assessment points, and adopted a natural log transformation to reduce positive skew.
Maternal Age at Birth: Maternal age at the target child’s birth is based on maternal reports and is scaled in years.
Maternal Education: Maternal education level is based on self-reported number of years that she had attended school when the child was 1-month old.
Maternal Employment Beliefs: Mothers’ beliefs about the developmental benefits of maternal employment on children was measured when the target child was approximately 1-month old, using five items from the Attitude Toward Maternal Employment Questionnaire (Greenberger, Goldberg, Crawford, & Granger, 1988). Higher scores reflect greater benefits or working. Internal consistency reliability for this scale was acceptable (alpha = .80).
Maternal Separation Anxiety: Maternal feelings of anxiety, concern, guilt, and sadness when separated from her child were measured when the target child was approximately 1-month old, based on their ratings on 21 items from the Parental Care Questionnaire (Hock, DeMeis, & McBride, 1987). Higher score indicate higher levels of separation anxiety. Internal consistency reliability for this scale was acceptable (alpha = .93).
Traditional Parenting Views: The degree to which mothers endorsed more traditional/authoritarian parenting views was based on maternal reports on 22 items from the Parental Modernity Scale (Schaefer & Edgerton, 1985), when the target child was approximately 1-month old. Higher scores indicate more authoritarian, less progressive parenting beliefs. Internal consistency reliability for this scale was acceptable (alpha = .90).
Maternal Personality: When the target child was approximately 6-months old, maternal personality traits were measured using the Agreeableness, Extra-version, and Neuroticism subscales of the NEO Five-Factor Inventory (NEO-FFI; Costa & McCrae, 1985) and NEO Personality Inventory scales (NEO-PI; Costa & McCrae, 1985). The Agreeableness subscale was taken from the NEO-FFI; mothers rated the degree to which items tapping trust, modesty, compliance, altruism, straightforwardness, and tender-mindedness were reflective of their personalities. Higher scores indicate higher levels of compassion and cooperation. The remaining subscales were taken from the NEO-PI. On the extraversion subscale, mothers rated the degree to which items measuring the quantity and intensity of interpersonal interactions, activity levels, need for stimulation, and capacity for joy reflected their personalities. Higher scores indicate more sociable, active, and person-oriented personalities. The neuroticism subscale comprises items reflecting adjustment and emotional instability, proneness to psychological distress, unrealistic ideas, and/or maladaptive coping responses. Higher scores on this subscale indicate higher levels of worrying and anxiety. Internal-consistency reliability estimates were acceptable (alpha = .76–.92).
Maternal Vocabulary: Maternal receptive vocabulary was measured when the child was approximately 36-month old, using the Peabody Picture Vocabulary Test-R (Dunn & Dunn, 1981). Higher scores are indicative of broader maternal vocabulary. This commonly used measure reports reasonable split-half reliability (.80–.83).
Maternal Sensitivity: Observational measures of maternal sensitivity were conducted when children were approximately 6-, 15-, 24-, 36-, and 54-months old (see NICHD ECCRN, 1999 for review). At all time-points mother–child interactions were videotaped during semi-structured play procedures. At the 6-month in-home assessment mothers were asked to play with their children as they would typically, for 7 min. In the remaining time, mothers and children were provided with a standardized set of toys/objects to play with (or not). At the 15- (home), 24- (laboratory), and 36-month (laboratory) assessment, mothers and children were presented with three boxes, each containing a set of developmentally appropriate books/toys and were asked to play as they would typically. No other directions were given than to go through the boxes in a particular order. Blind raters coded the videotaped interactions for maternal sensitivity. The 6-, 15-, and 24-month sensitivity scores were based on a composite score of maternal sensitivity to nondistress, positive regard, and intrusiveness (reversed). Each was rated on a 1–4 scale and then summed to create the composite. Higher ratings indicate higher levels of maternal sensitivity. The 36- and 54-month composites were adjusted for developmental appropriateness and included ratings of maternal supportive presence, respect for autonomy, and hostility (reversed). Each was rated on a 1–7 scale and summed to create the composite. Approximately, 20% of the videotapes were coded by two raters, at each assessment period. Inter-rater reliability estimates based upon repeated-measures ANOVA (Winer, 1971) ranged from .83 to .88. Internal-consistency reliability for the maternal sensitivity scales ranged from .70 to .84. In the present study, we adopted a composite representing average maternal sensitivity across infancy and early childhood by standardizing each scale and averaging the standardized scores across the five measures.
Site: Site dummy variables were included to adjust for biases due to research site.
Descriptive Statistics for the Control Covariates Included in All Multiple Regression Models
| Mean | SD | Min | Max | Missing (%) | |
|---|---|---|---|---|---|
| Male | .52 | .50 | .00 | 1.00 | .00 |
| African American | .13 | .34 | .00 | 1.00 | .00 |
| Gest. age | 39.25 | 1.44 | 33.00 | 43.00 | .01 |
| Temperament | 3.18 | .40 | 1.54 | 4.72 | .06 |
| Birth order | 1.83 | .95 | 1.00 | 7.00 | .00 |
| Maternal sensitivity | −.02 | .75 | −3.07 | 1.57 | .04 |
| Separation anxiety | 70.26 | 13.25 | 35.00 | 105.00 | .01 |
| Income | 3.51 | 2.96 | .00 | 22.47 | .05 |
| Education | 14.23 | 2.51 | 7.00 | 21.00 | .00 |
| Married | .76 | .42 | .00 | 1.00 | .00 |
| Vocab | 99.01 | 18.35 | 40.00 | 159.00 | .14 |
| Neuroticism | 29.77 | 7.16 | 13.00 | 59.00 | .07 |
| Extroversion | 42.49 | 5.83 | 23.00 | 60.00 | .07 |
| Agreeableness | 46.28 | 5.29 | 28.00 | 60.00 | .07 |
| Trad. parent | 60.34 | 15.21 | 23.00 | 104.00 | .00 |
| Ben. of work | 19.19 | 3.17 | 5.00 | 30.00 | .00 |
| Age | 28.11 | 5.63 | 18.00 | 46.00 | .00 |
REFERENCES
- Achenbach TM (1991). Integrative guide for the 1991 CBCL/4–18, YSR, and TRF profiles. Burlington: University of Vermont, Department of Psychiatry. [Google Scholar]
- Allison PD (2003). Missing data techniques for structural equation modeling. Journal of Abnormal Psychology, 112, 545–557. [DOI] [PubMed] [Google Scholar]
- Anchordoquy HC, McGeary C, Liu L, Krauter KS, & Smolen A (2003). Genotyping of three candidate genes following whole genome preamplification of DNA collected from buccal cells. Behavior Genetics, 33, 73–78. [DOI] [PubMed] [Google Scholar]
- Arnsten AF, & Li BM (2005). Neurobiology of executive functions: Catecholamineinfluences on prefrontal cortical functions. Biological Psychiatry, 57, 1377–1384. [DOI] [PubMed] [Google Scholar]
- Bakermans-Kranenburg MJ, & van IJzendoorn MH (2006). Gene–environment interaction of the dopamine D4 receptor (DRD4) and observed maternal insensitivity predicting externalizing behavior in preschoolers. Developmental Psychobiology, 48, 406–409. [DOI] [PubMed] [Google Scholar]
- Bakermans-Kranenburg MJ, & van IJzendoorn MH (2011). Differential susceptibility to rearing environment depending on dopamine-related genes: New evidence and a meta-analysis. Development and Psychopathology, 23, 39–52. [DOI] [PubMed] [Google Scholar]
- Bakermans-Kranenburg MJ, van IJzendoorn MH, Pijlman FT, Mesman J, & Juffer F (2008). Experimental evidence for differential susceptibility: Dopamine D4 receptor polymorphism (DRD4 VNTR) moderates intervention effects on toddlers’ externalizing behavior in a randomized controlled trial. Developmental Psychology, 44, 293–300. [DOI] [PubMed] [Google Scholar]
- Barkley RA (1997). Behavioral inhibition, sustained attention, and executive functions: Constructing a unifying theory of ADHD. Psychological Bulletin, 121, 65–94. [DOI] [PubMed] [Google Scholar]
- Barkley RA, Smith KM, Fischer M, & Navia B (2006). An examination of the behavioral and neuropsychological correlates of three ADHD candidate gene polymorphisms (DRD4 7+, DBH TaqI A2, and DAT1 40 bp VNTR) in hyperactive and normal children followed to adulthood. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 141, 487–498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnett WS (1995). Long-term effects of early childhood programs on cognitive and school outcomes. The Future of Children, 5, 25–50. [PubMed] [Google Scholar]
- Barnett WS, Jung K, Yarosz DJ, Thomas J, Hornbeck A, Stechuk R, & Burns S (2008). Educational effects of the Tools of the Mind curriculum: A randomized trial. Early Childhood Research Quarterly, 23(3), 299–313. [Google Scholar]
- Bellgrove MA, Hawi Z, Lowe N, Kirley A, Robertson IH, & Gill M (2005). DRD4 gene variants and sustained attention in attention deficit hyperactivity disorder (ADHD): Effects of associated alleles at the VNTR and − 521 SNP. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 136B, 81–86. [DOI] [PubMed] [Google Scholar]
- Belsky J (1999). Quantity of nonmaternal care and boys’ problem behavior/adjustment at 3 and 5: Exploring the meditating role of parenting. Psychiatry: Interpersonal and Biological Processes, 62, 1–21. [DOI] [PubMed] [Google Scholar]
- Belsky J, & Pluess M (2009). The nature (and nurture?) of plasticity in early human development. Perspectives in Psychological Science, 4, 345–351. [DOI] [PubMed] [Google Scholar]
- Belsky J, Fearon RMP, & Bell B (2007). Parenting, attention and externalizing problems: Testing mediation longitudinally, repeatedly and reciprocally. Journal of Child Psychology and Psychiatry and Allied Disciplines, 48, 1233–1242. [DOI] [PubMed] [Google Scholar]
- Belsky J, Vandell DL, Burchinal M, Clarke-Stewart KA, McCartney K, Owen MT, & the NICHD Early Child Care Research Network. (2007). Are there long-term effects of early child care? Child Development, 78, 681–701. [DOI] [PubMed] [Google Scholar]
- Benjamini Y, & Hochberg Y (1995). Controlling the false discovery rate: A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society, 57(Series B), 289–300. [Google Scholar]
- Bernier A, Carlson SM, & Whipple N (2010). From external regulation to self-regulation: Early parenting precursors of young children’s executive functioning. Child Development, 81, 326–339. [DOI] [PubMed] [Google Scholar]
- Berry D (2012). Inhibitory control and teacher–child conflict: Reciprocal associations across the elementary-school years. Journal of Applied Developmental Psychology, 33, 66–76. [Google Scholar]
- Berry D, Deater-Deckard K, McCartney K, Wang Z, & Petrill S (in press). Gene–environment interaction between DRD4 7-repeat polymorphism and early maternal sensitivity predicts inattention trajectories across middle childhood. Development and Psychopathology. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bierman KL, Nix RL, Greenberg MT, Blair C, & Domitrovich CE (2008). Executive functions and school readiness intervention: Impact, moderation, and mediation in the Head Start REDI program. Developmental Psychopathology, 20, 821–843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blair C (2002). School readiness: Integrating cognition and emotion in a neurobiological conceptualization of child functioning at school entry. American Psychologist, 57, 111–127. [DOI] [PubMed] [Google Scholar]
- Blair C, Granger D, Willoughby M, Mills-Koonce R, Cox M, Greenberg MT, … the FLP Investigators. (2011). Salivary cortisol mediates effects of poverty and parenting on executive functions in early childhood. Child Development, 82, 1970–1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blair C, & Razza RP (2007). Relating effortful control, executive function, and false-belief understanding to emerging math and literacy ability in kindergarten. Child Development, 78, 647–663. [DOI] [PubMed] [Google Scholar]
- Blair C, & Ursache A (2011). A bidirectional model of executive functions and self-regulation. In Baumeister RF & Vohs KD (), Handbook of self-regulation (2nd ed., pp. 300–320). New York: Guilford. [Google Scholar]
- Boyce WT, & Ellis BJ (2005). Biological sensitivity to context: I. An evolutionary developmental theory of the origins and functions of stress reactivity. Development and Psychopathology, 17, 271–301. [DOI] [PubMed] [Google Scholar]
- Brody GH, Beach SRH, Philibert RA, Chen Y-F, & Murry VM (2009). Prevention effects moderate the association of 5-HTTLPR and youth risk behavior initiation: Gene × environment hypotheses tested via a randomized prevention design. Child Development, 80, 645–661. [DOI] [PubMed] [Google Scholar]
- Burchinal MR, & Clarke-Stewart AK (2007). Maternal employment and child cognitive outcomes: The importance of analytic approach. Developmental Psychology, 43, 1140–1155. [DOI] [PubMed] [Google Scholar]
- Butts KA, Weinberg J, Young AH, & Phillips AG (2011). Glucocorticoid receptors in the prefrontal cortex regulate stress-evoked dopamine efflux and aspects of executive function. Proceedings of the National Academy of Sciences of the United States of America, 108, 18459–18464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campbell FA, Ramey CT, Pungello E, Sparling J, & Miller-Johnson S (2002). Early childhood education: Young adult outcomes from the Abecedarian Project. Applied Developmental Science, 6, 42–57. [Google Scholar]
- Campbell SB, & von Stauffenberg C (2009). Delay and inhibition as early predictors of ADHD symptoms in third grade. Journal of Abnormal Child Psychology, 37, 1–15. [DOI] [PubMed] [Google Scholar]
- Carey WB, & McDevitt SC, (1978). Revision of the infant temperament questionnaire. Pediatrics, 61, 735–739. [PubMed] [Google Scholar]
- Chang FM, Kidd JR, Livak KJ, Pakstis AJ, & Kidd KK (1996). The world-wide distribution of allele frequencies at the human dopamine D4 receptor locus. Human Genetics, 98, 91–101. [DOI] [PubMed] [Google Scholar]
- Carlson SM (2003). Executive function in context: Development, measurement, theory, and experience. Mono-graphs of the Society for Research in Child Development, Serial No. 274, 68, 138–151. [PubMed] [Google Scholar]
- Collins LM, Schafer JL, & Kam CM (2001). A comparison of inclusive and restrictive strategies in modern missing data procedures. Psychological Methods, 6, 330–351. [PubMed] [Google Scholar]
- Costa PT, & McCrae RR (1985). The NEO personality inventory manual. Odessa FL: Psychological Assessment Resources. [Google Scholar]
- Crockenberg SC, & Leerkes EM (2005). Infant temperament moderates associations between childcare type and quantity and externalizing and internalizing behaviors at 2½ years. Infant Behavior and Development, 28, 20–35. [Google Scholar]
- Dearing E, McCartney K, & Taylor BA (2009). Does higher quality early child care promote low-income children’s math and reading achievement in middle childhood? Child Development, 80, 1329–1349. [DOI] [PubMed] [Google Scholar]
- deKloet ER, Oitzl MS, & Joëls M (1999). Stress and cognition: Are corticosteroids good or bad guys? Trends in Neuroscience, 2210, 422–426. [DOI] [PubMed] [Google Scholar]
- Dunn LM, & Dunn LM (1981). Peabody Picture Vocabulary Test – Revised. Circle Pines, MN: American Guidance Service. [Google Scholar]
- Ellis BJ, Boyce WT, Belsky J, Bakermans-Kranenburg MJ, & van IJzendoorn MH (2011). Differential susceptibility to the environment: An evolutionary-neurodevelopmental theory. Development and Psychopathology, 23, 7–28. [DOI] [PubMed] [Google Scholar]
- Freeman B, Smith N, Curtis C, Huckett L, Mill J, & Craig IW (2003). DNA from buccal swabs recruited by mail: Evaluation of storage effects on long-term stability and suitability for multiplex polymerase chain reaction genotyping. Behavior Genetics, 33, 67–72. [DOI] [PubMed] [Google Scholar]
- Fuller B, Kagan SL, Loeb S, & Chang Y (2004). Child care quality: Centers and home-based settings that serve poor families. Early Childhood Research Quarterly, 19, 505–527. [Google Scholar]
- Gerstadt CL, Hong YJ, & Diamond A (1994). The relationship between cognition and action: Performance of children 3.5–7 years old on a Stroop-like day–night test. Cognition, 53, 129–153. [DOI] [PubMed] [Google Scholar]
- Greenberger E, Goldberg WA, Crawford TJ, & Granger J (1988). Beliefs about the consequences of maternal employment for children. Psychology of Women Quarterly, 12, 35–59. [Google Scholar]
- Halperin JM, Sharma V, Greenblatt E, & Schwartz ST (1991). Assessment of the Continuous Performance Test: Reliability and validity in a nonreferred sample. Psychological Assessment, 3, 603–608. [Google Scholar]
- Hock E, DeMeis D, & McBride S (1987). Maternal separation anxiety: Its role in the balance of employment and motherhood in mothers of infants. In Gottfried AE, & Gottfried AW, (Eds.), Maternal employment and children’s development: Longitudinal research (pp. 191–227). New York: Plenum Press. [Google Scholar]
- Hughes C, & Ensor R (2011). Individual differences in growth in executive function across the transition to school predict externalizing and internalizing behaviors and children’s self-perceived academic success at age 6. Journal of Experimental Child Psychology, 108, 663–676. [DOI] [PubMed] [Google Scholar]
- Johnson KA, Kelly SP, Robertson IH, Barry E, Mulligan A, Daly M, … Bellgrove MA (2008). Absence of the 7-repeat variant of the DRD4 VNTR is associated with drifting sustained attention in children with ADHD but not in controls. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 147B, 927–937. [DOI] [PubMed] [Google Scholar]
- Jovanovic V, Guan HC, & Van Tol HH (1999). Comparative pharmacological and functional analysis of the human dopamine D4.2 and D4.10 receptor variants. Pharmacogenetics, 9, 561–568. [PubMed] [Google Scholar]
- Kim J, Deater-Deckard K, Mullineaux PY, & Allen B (2010). Longitudinal studies of anger and attention span: Context and informant effects. Journal of Personality, 78, 419–440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kochanska G, Murray KT, & Harlan ET (2000). Effortful control in early childhood: Continuity and change, antecedents, and implications for social development. Developmental Psychology, 36, 220–232. [PubMed] [Google Scholar]
- Ladd GW, Herald SL, & Kochel KP (2006). School readiness: Are there social prerequisites? Early Education and Development, 17, 115–150. [Google Scholar]
- Langley K, Marshall L, van den Bree M, Thomas H, Owen M, O’Donovan M, & Thapar A (2004). Association of the dopamine D4 receptor gene 7-repeat allele with neuropsychological test performance of children with ADHD. American Journal of Psychiatry, 161, 133–138. [DOI] [PubMed] [Google Scholar]
- Loeb S, Bridges M, Bassok D, Fuller B, & Rumberger RW (2007). How much is too much? The influence of preschool centers on children’s social and cognitive development. Economics of Education Review, 26, 52–66. [Google Scholar]
- Lupien SJ, Maheu F, Tu M, Fiocco A, & Schramek TE (2007). The effects of stress and stress hormones on human cognition: Implications for the field of brain and cognition. Brain and Cognition, 653, 209–237. [DOI] [PubMed] [Google Scholar]
- Magnuson K, Ruhm C, & Waldfogel J (2007). Does prekindergarten improve school preparation and performance? Economics of Education Review, 26, 33–51. [Google Scholar]
- McCartney K, Burchinal M, Clarke-Stewart A, Bub K, Owen M, Belsky J, & NICHD Early Child Care Research Network. (2010). Testing a series of causal propositions relating time in child care to children’s externalizing behavior. Developmental Psychology, 46, 1–17. [DOI] [PubMed] [Google Scholar]
- McClelland MM, Cameron CE, Connor CM, Farris CL, Jewkes AM, & Morrison FJ (2007). Links between behavioral regulation and preschoolers’ literacy, vocabulary, and math skills. Developmental Psychology, 43, 947–959. [DOI] [PubMed] [Google Scholar]
- Meador-Woodruff JH, Damask SP, Wang J, Haroutunian V, Davis KL, & Watson SJ (1996). Dopamine receptor mRNA expression in human striatum and neocortex. Neuropsychopharmacology, 15, 17–29. [DOI] [PubMed] [Google Scholar]
- Mirsky AF, Anthony BJ, Duncan CC, Ahearn MB, & Kellam SG (1991). Analysis of the elements of attention: A neuropsychological approach. Neuropsychology Review, 2, 109–145. [DOI] [PubMed] [Google Scholar]
- Mischel W (1974). Processes in delay of gratification. In Berkowitz L (Ed.), Advances in experimental social psychology (Vol. 7). New York: Academic Press. [Google Scholar]
- Miyake A, Friedman NP, Emerson MJ, Witzki AH, Howerter A, & Wager TD (2000). The unity and diversity of executive functions and their contributions to complex “frontal lobe” tasks: A latent variable analysis. Cognitive Psychology, 41, 49–100. [DOI] [PubMed] [Google Scholar]
- Montgomery DE, & Koeltzow TE (2010). A review of the day–night task: The Stroop paradigm and interference control in young children. Developmental Review, 30, 308–330. [Google Scholar]
- Muthén LK, & Muthén BO (2009). Mplus. Statistical analysis with latent variables: User’s guide. Los Angeles: Muthén & Muthén. [Google Scholar]
- National Institute of Child Health and Human Development Early Child Care Research Network. (1996). Characteristics of infant child care: Factors contributing to positive caregiving. Early Childhood Research Quarterly, 11, 296–306. [Google Scholar]
- NICHD Early Child Care Research Network (1997). The effects of infant child care on infant–mother attachment security: Results of the NICHD Study of Early Child Care. Child Development, 68, 860–879. [DOI] [PubMed] [Google Scholar]
- National Institute of Child Health and Human Development Early Child Care Research Network. (1998). Child care and self control, compliance, and problem behavior at 24 and 36 months. Child Development, 69, 1145–1170. [PubMed] [Google Scholar]
- NICHD Early Child Care Research Network. (1999). Child care and mother-child interaction in the first three years of life. Developmental Psychology, 35, 1399–1413. [PubMed] [Google Scholar]
- National Institute of Child Health and Human Development Early Child Care Research Network. (2003a). Do children’s attention processes mediate the link between family predictors and school readiness? Developmental Psychology, 39, 581–593. [DOI] [PubMed] [Google Scholar]
- National Institute of Child Health and Human Development Early Child Care Research Network. (2003b). Does amount of time spent in child care predict socioemotional adjustment during the transition to kindergarten? Child Development, 74, 976–1005. [DOI] [PubMed] [Google Scholar]
- National Institute of Child Health and Human Development Early Child Care Research Network & Duncan GJ (2003). Modeling the impacts of child care quality on children’s preschool cognitive development. Child Development, 74, 1454–1475. [DOI] [PubMed] [Google Scholar]
- Oak JN, Oldenhof J, & Van Tol HHM (2000). The dopamine D4 receptor: One decade of research. European Journal of Pharmacology, 405, 303–327. [DOI] [PubMed] [Google Scholar]
- Phillips DA, Fox NA, & Gunnar MR (2011). Same place, different experiences: Bringing individual differences to research in child care. Child Development Perspectives, 5, 44–49. [Google Scholar]
- Pluess M, & Belsky J (2009). Differential susceptibility to rearing experience: The case of childcare. Journal of Child Psychology and Psychiatry and Allied Disciplines, 50, 396–404. [DOI] [PubMed] [Google Scholar]
- R Development Core Team. (2012). R: A Language and Environment for Statistical Computing (http://www.r-projet.org).
- Raver CC, Jones SM, Li-Grining C, Zhai F, Bub K, & Pressler E (2011). CSRP’s impact on low-income preschoolers’ preacademic skills: Self-regulation as a mediating mechanism. Child Development, 821, 362–378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raymond M, & Rousset F (1995). GENEPOP (version 1.2): A population genetics software for exact test and ecumenicism. Journal of Heredity, 86, 248–249. [Google Scholar]
- Rothbart MK, Ahadi SA, Hershey KL, & Fisher P (2001). Investigations of temperament at 3–7 years: The Children’s Behavior Questionnaire. Child Development, 72, 1394–1408. [DOI] [PubMed] [Google Scholar]
- Sander T, Harms H, Dufeu P, Kuhn S, Rommelspacher H, & Schmidt LG (1998). Dopamine D4 receptor exon III alleles and variation of novelty seeking in alcoholics. American journal of medical genetics, 74(5), 483–487. [DOI] [PubMed] [Google Scholar]
- Saudino KJ, & Wang M (2012). Quantitative and molecular genetic studies of temperament. In Zentner M & Shiner RL (Eds.), Handbook of temperament. New York: Guilford. [Google Scholar]
- Schaefer E, & Edgerton M (1985). Parental and child correlates of parental modernity. In Sigel IE, (Ed). Parental belief systems: The psychological consequences for children. Hillsdale, NJ: Erlbaum. [Google Scholar]
- Schinka JA, Letsch EA, & Crawford FC (2002). DRD4 and novelty seeking: Results of meta-analyses. American Journal of Medical Genetics, 114, 643–648. [DOI] [PubMed] [Google Scholar]
- Schoots O, & Van Tol HHM (2003). The human dopamine D4 receptor repeat sequences modulate expression. Pharmacogenomics Journal, 3, 343–348. [DOI] [PubMed] [Google Scholar]
- Schweinhart LJ, Montie J, Xiang Z, Barnett WS, Belfield CR, & Nores M (2005). Lifetime effects: The High/Scope Perry Preschool study through age 40. Ypsilanti, MI: High/Scope Educational Research Foundation. [Google Scholar]
- Séguin JR, & Zelazo PD (2005). Executive function in early physical aggression. In Tremblay RE, Hartup WW, & Archer J (Eds.), Developmental origins of aggression (pp. 307–329). New York: Guilford. [Google Scholar]
- Sims M, Guilfoyle A, & Parry TS (2006). Children’s cortisol levels and quality of child care provision. Child: Care, Health and Development, 32(4), 453–466. [DOI] [PubMed] [Google Scholar]
- Sheese BE, Voelker PM, Rothbart MK, & Posner MI (2007). Parenting quality interacts with genetic variation in dopamine receptor D4 to influence temperament in early childhood. Development and Psychopathology, 19, 1039–1046. [DOI] [PubMed] [Google Scholar]
- Spinrad TL, Eisenberg N, Gaertner B, Popp T, Smith CL, Kupfer A, Greving K et al. (2007). Relations of maternal socialization and toddlers’ effortful control to children’s adjustment and social competence. Developmental Psychology, 43, 1170–1186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- U.S. Census Bureau. (2010). Who’s minding the kids? Child care arrangements Spring 2010. Washington, DC: U.S. Census Bureau. [Google Scholar]
- Vandell DL, Belsky J, Burchinal M, Steinberg L, Vandergrift N, & NICHD Early Child Care Research Network. (2010). Do effects of early child care extend to age 15 years? Results from the NICHD Study of Early Child Care and Youth Development. Child Development, 81, 737–756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vermeer HJ, & Van IJzendoorn MH (2006). Children’s elevated cortisol levels at daycare: A review and meta-analysis. Early Childhood Research Quarterly, 21, 390–401. [Google Scholar]
- Votruba-Drzal E, Coley RL, & Chase-Lansdale PL (2004). Child care and low-income children’s development: Direct and moderated effects. Child Development, 75, 296–312. [DOI] [PubMed] [Google Scholar]
- Waldman ID (2005). Statistical approaches to complex phenotypes: Evaluating neuropsychological endophenotypes for attention-deficit/hyperactivity disorder. Biological Psychiatry, 57, 1347–1356. [DOI] [PubMed] [Google Scholar]
- Willcutt EG, Doyle AE, Nigg JT, Faraone SV, & Pennington BF (2005). Validity of the executive function theory of Attention Deficit/Hyperactivity Disorder: A meta-analytic review. Biological Psychiatry, 57, 1336–1346. [DOI] [PubMed] [Google Scholar]
- Winer BJ (1971). Statistical principles in experimental design. Second edition. New York: McGraw-Hill. [Google Scholar]