

Abstract
Tyrophagus putrescentiae (Schrank), commonly known as the cereal mite, cheese mite, or ham mite, is a cosmopolitan species reported from various environments in the wild, including soil, plant material and vertebrate nests. It has also been recognized as a common pest of food storages, mycological collections as well as plant and invertebrate laboratory cultures. Laboratory observations indicate that T. putrescentiae feeds on a large range of dermatophytes, yeasts and molds. We have observed the interspecific relation between this mite and several species of true slime molds (Mycetozoa) under laboratory conditions, which confirms the very broad spectrum of feeding habits of T. putrescentiae. Mycetozoans were grown in semi-sterile in vitro cultures and fed with oat flour or oat flakes. Tyrophagus putrescentiae displayed affinity to all macroscopically identifiable stages of the life cycle of Fuligo septica (L.) F.H. Wigg, Physarum polycephalum Schwein and the Didymium sp. complex [Didymium iridis (Ditmar) Fr., Didymium nigripes (Link) Fr. and Didymium bahiense Gottsb.]: live, decaying or dead plasmodia, sporangia, aethalia, spores and sclerotia. The relation carrying symptoms of various types of interspecific interaction, is hypothesized to form an evolutionarily young phenomenon, which not only identifies a new aspect of mycetozoal biology, but also presents the cereal mite as a species of high adaptive potential.
Supplementary Information
The online version contains supplementary material available at 10.1007/s10493-021-00608-4.
Keywords: Storage mite, Interspecific interaction, Myxomycetes, Fuligo septica, Physarum polycephalum, Didymium
Introduction
Slime molds (Mycetozoa, called also Myxomycetes, Myxogastria, Myxomycota, Eumycetozoa) are simple but unusual eukaryotes. Their plasmodial structure and ameboid-type migration both justify their classification as gigantic microorganisms. Slime molds reveal some features of fungi (lack of autotrophy, production of sporangia in fruit bodies); however, many more characteristics of slime molds are shared with protozoans (Hamilton 2001; Espinosa and Paz-Y-Miño-C 2014). Because of displaying some intermediate traits, in addition to several primitive features, such as lack of histological organization of the body, large size and relatively easy cultivation under laboratory conditions, they have become macroscopic models of the eukaryotic cell and interesting alternate organisms in biological and biomedical research. Their popularity in interdisciplinary studies, such as biocomputing (Adamatzky 2010) or astrobiology (Díez et al. 2020), has increased rapidly.
A detailed description of the slime mold life cycle may be found in numerous handbooks and papers (e.g., Stephenson and Stempen 1994; Díez et al. 2020). Typically, their life cycle starts in sporangia with haploid spores surrounded by a cell wall. After germination, the haploid flagellated forms conjugate and, after a disposition of the flagella, the diploidal ameboid grows to the main vegetative form of the life cycle, the plasmodium. The spores may also be generated by apogamy and remain diploid, which makes the taxonomy of slime molds particularly challenging (Clark and Mires 1999; ElHage et al. 2000; Clark et al. 2001, 2004). The main plasmodial type, the so called phaneroplasmodium, is often pigmented and may reach the macroscopic size scale (up to tens or hundreds cm2; Stephenson and Stempen 1994; Płonka and Rakoczy 1997; Adamatzky 2010). The plasmodium preserves its ability to migrate actively. Under influence of starvation, drought, light, and other stimuli, the plasmodium creates a fruiting body, which may form a simple stalked or sessile sporangium, or a complex aethalium. Under adverse circumstances, the plasmodium may transform into a dormant state—usually the sclerotium that preserves its ability to regenerate into the active plasmodium under normal conditions (Krzywda et al. 2008).
Slime molds are heterotrophic, saprophytic organisms. In the laboratory they may grow on organic substrates (agar, gelatin, lignin, fluid media) absorbing nutrients from the feeder, but in their natural environment slime molds have developed various types of interactions with other organisms. At large, they feed on bacteria and other microorganisms, including fragments of other slime molds which they can absorb and digest (Madelin 1984; Stephenson 1989; Stephenson and Stempen 1994; Adamatzky 2010; Stephenson and Schnittler 2016). Some slime molds are parasitic. A known example, Licea parasitica (Zukal) G.W. Martin, associated with several species of lichens, seems to be a facultative lychenicolous mycetozoan of still unidentified ecological status (Kocourková 1999). Stephenson and Schnittler (2016) report some slime molds, including Fuligo septica (L.) F.H. Wigg, as capable of enzyme excretion to the substrate and extracellular digestion of fruiting bodies (and likely mycelia) of fungi. As commensals, slime molds can also compete with other organisms, e.g., strawberries (Arthaud and Lafon 1972), plant sprouts in hydroponic cultures (Michalczyk et al. 2011) or with cultivated mushrooms (Rakoczy and Szymański 1985; Chung et al. 1998).
Because their plasmodium (large amount of cytoplasm) represents a valuable source of nutrients, slime molds may fall prey to other organisms. It has been documented that slugs may feed on live plasmodia (Keller and Snell 2002; Stephenson and Schnittler 2016), and there is also quite a substantial literature on myxomyceticolous insects: beetles and flies (Stephenson and Stempen 1994; Stephenson and Schnittler 2016; Sugiura et al. 2019). These animals should be therefore called ‘insidious predators’ (Zelmer 1998). As production of spores may be attributed to primitive plants, algae or fungi, this type of interaction may also be called ‘grazing’ (Weiner 2003). However, reports on slime molds as the hosts for parasites are still infrequent, similar to those on the parasitic slime molds.
In our laboratory, on certain occasions, we have observed massive infestations of Tyrophagus putrescentiae in slime mold cultures. Tyrophagus putrescentiae is commonly referred to as the mold mite, cheese mite, or ham mite (Hughes 1961; Amoah 2016). It is a common pest of stored products, especially products with high fat and protein contents, such as whole wheat flour, soy flour, cheese, rye bread, white bread, herring meal, bacon, dry milk and various seeds (Hughes 1961; Duek et al. 2001). It has been reported to be associated with over 140 commodities (Hagstrum et al. 2013), including dried fruits, spices, cultured cheese, and other high-value foods (Rentfrow et al. 2006; Amoah 2016). Indeed, Keller and Smith (1978) and Smith and Keller (1978) reported an acarid mite, T. putrescentiae, feeding on a phaneroplasmodium. Observations of the interaction between the mite and an unidentified slime mold were made on live tree bark under laboratory conditions. They were followed by monitoring of this interaction in the in vitro agar cultures of slime molds of Stemonitis flavogenita Jahn., and Didymium sp. The authors acknowledged Guilford S. Ide (Curator, Acarology Laboratory, Ohio State University, USA), who identified the mite; however, in their short article, they did not include a list of character states that formed the basis for species identification.
Keller and Smith (1978) also observed that mites can positively interact with Didymium sp. by spreading its spores through the digestive system. In the present study, we expand these observations by: (1) confirming that the mite species which lives together with the slime molds is T. putrescentiae—we provide the set of character states which constituted the background for species identification; (2) proving that the mite can be found in cultures of various species of unrelated Myxomycetes, F. septica, various members of Didymium sp. and Physarum polycephalum Schwein, famous for its application in the field of biocomputing (Adamatzky 2010); and (3) revealing that the mites exhibit affinity to various stages of the slime mold life cycle (plasmodium, spores, sporangia and sclerotia). Finally, we discuss the nature of the ecological interaction between the mites and the slime molds.
Material and methods
Slime molds were cultured as described previously (Krzywda et al. 2008; Adamatzky 2010; Michalczyk et al. 2011) at room temperature (19–21 °C) in the dark, in glass Petri dishes (9 and 11 cm diameter), or in 5.5 cm diameter plastic tissue culture dishes (TPP Techno Plastic Products, Trasadingen, Switzerland). Humidity in the laboratory was low, ca. 30–35%, but within the covered Petri dishes it reached up to 84 ± 9% (mean ± SD; DM-9213 thermo-hygrometer, ATM, Hongkong, China). As F. septica is a unique agarolytic eukaryote (Murugan et al. 1996) which causes difficulties in maintaining it on solid agar (Haskins and Wrigley de Basanta 2008), the slime molds were grown on medium density Whatman® filter paper (Sigma-Aldrich, St Louis, MO, USA). They were fed with oat flour or oat flakes (some cultures of P. polycephalum) stored at −80 °C, and watered ad libitum with sterile water (Fig. 1). The cultures were serially passaged ca. 2–4 weeks under sterile conditions; however, we did not sterilize the flour/flakes. After passaging, the old cultures were maintained longer, and sometimes passaged a second or even third time. The mites were found in the old slime mold cultures, wherein they appeared most probably as contaminants of the oat flour/flakes.
Fig. 1.

In vitro cultures of Fuligo septica (a), Didymium sp. (b) and Physarum polycephalum (c). All cultures on Petri dishes with filter paper and oat flour/flakes. Scale bar: 2 cm
The mites were preserved in 90% ethyl alcohol, mounted on microscopic slides in Swan’s fluid (Swan 1936), examined under Nikon Eclipse 50i microscope (Nikon Instruments, Tokyo, Japan) and identified based on the key provided by Solarz (2012). Photographs of T. putrescentiae were taken using the Nikon Eclipse 50i microscope coupled with a Toshiba 1080i camera system. The vouchers are deposited at the Department of Invertebrate Systematics and Ecology, Wrocław University of Environmental and Life Sciences, Poland.
Photographs of slime mold cultures were taken using a Nikon D7000 camera equipped with Nikon Nikkor Micro AF 60 mm 1:2.8 lenses (Nikon, Tokyo, Japan), and the video sequences were recorded using the same camera assembled to a Möller-Weder operating microscope (Möller-Wedel, Wedel, Germany). The sequences were edited using freeware OpenShot Video Editor v.2.4.4. (OpenShot Studios, Rockwall, TX, USA).
Density of mites in the culture dishes was estimated on the base of photographs and magnification (expressed as scale bars), and was only roughly determined, because of a very changeable occurrence of the mites (see, e.g., Smrž and Čatská 1987).
Results and discussion
Identity of the myxomyceticolous mite
The mites interfering with the slime molds represent T. putrescentiae (Sarcoptiformes: Acaridae). According to the data provided by Solarz (2012), the species can be distinguished based on the following characters: anterior margins of propodosomal shield with pigmented spots (corneae); seta d 1 more than twice as long as c 1; seta d 2 only slightly longer than c 1; seta e 2 very long, similar in length to h 2; supracoxal seta (sc x) expanded at base, with 5–6 fairly long pectinations; distal 2/3 of solenidion ω 1 widened, terminated in a distinctly pointed tip. The diagnostic characters used to identify the species and to distinguish it from relatives are presented in Fig. 2—these are: seta sc x relatively long, either slightly or markedly widened in basal half, with long pectinations; tarsi I and II with solenidion ω 1 terminating in a distinctly expanded tip (distal 2/3 of solenidion ω 1 on tarsus I widened); S-shaped aedeagus in males, with two deep curves—one at the base, the other one in the apical third; tarsal (copulatory) suckers on Ta IV.
Fig. 2.

Tyrophagus putrescentiae. a Macrophotography of specimens in a culture, not to scale. b Male, ventral view. c Gnathosoma and leg I (part), dorsal view, ω 1 = solenidion 1 on tarsus I. d Female—hysterosoma, ventral view. e Gnathosoma (part) and legs I, II (part), dorsal view, ω 1 = solenidion 1 on tarsus II. f Male—opisthosoma and hind legs, ventral view. p. p., sex suckers; p. k., copulatory suction cups on tarsus IV. Scale bars: 45 µm (b), 13 µm (c, e), 28 µm (d, f)
Affinity to various species of Mycetozoa in all stages of their life cycle
The mites have been observed to feed on all macroscopic forms which develop in the life cycle of F. septica, P. polycephalum, and of several members of Didymium sp., earlier reported by Michalczyk et al. (2011) as a species complex, which reveals features of Didymium iridis (Ditmar) Fr., Didymium nigripes (Link) Fr. and Didymium bahiense Gottsb. (Stephenson 2003). It is important to note that these mycetozoans represent different family-level taxa. Whereas all belong to the order Physarales, P. polycephalum and F. septica represent family Physaraceae, and Didymium sp. is a member of Didymiaceae (Stephenson and Stempen 1994; Drozdowicz et al. 2003). The mites not only fed on plasmodia, but they also inhabited them (Online Resources 1) and make use of every identified stage of the slime mold life cycle. This included: live plasmodium (Figs. 3a–d, 4a, 5a–c, Online Resources 1), dead and decomposed plasmodium (Figs. 3e–g, 4b, 5d, e, Online Resources 2), aethalium (Fig. 3h–m, Online Resources 3) and sporangium (Figs. 4c, d, 5f–h), spores (Figs. 3i–m, 4d, 5f–h), and sclerotium (Fig. 3n–p) (Krzywda et al. 2008). We observed the same affinity as to F. septica (Fig. 3), also to Didymium sp. and P. polycephalum (Figs. 4, 5) in relation to the stages which we were able to follow under the in vitro conditions.
Fig. 3.

Tyrophagus putrescentiae (white arrows, when necessary) on various forms of Fuligo septica: on live (a–d) and dead (e–g) plasmodium (e—on the slimy track left by a plasmodium) of F. septica. Subsequent stages of two examples of the F. septica aethalium destruction after 8 days (h, i) and after 10 (k), 13 (l) and 17 (m) days following the onset of observation (j). Tyrophagus putrescentiae on a forming sclerotium of F. septica (n) and example of destruction of F. septica sclerotium within 24 h (o, p). White empty arrowheads indicate mites with their digestive tracts filled with various food items: yellow—live plasmodium (b, d), orange—dead plasmodium (f, g, g-inset), black—spores (m-inset). Scale bars: 0.5 cm (a, e, h–p), 1 mm (b–d, f, g)
Fig. 4.

Tyrophagus putrescentiae on live (a) and dead (b) plasmodium of Didymium sp. Sporangium (black arrows) of Didymium sp. before (c) and after invasion of T. putrescentiae (d). White arrows and white empty arrowheads—as in Fig. 3. Scale bars: 1 mm
Fig. 5.

Tyrophagus putrescentiae on various forms of Physarum polycephalum: on live (white arrows) (a–c) and dead (d, e) plasmodium. Subsequent stages of the P. polycephalum sporangium destruction after 5 (g) and 8 (h) days after the initial observation (f). White and black arrows and white empty arrowheads—as in Figs. 3 and 4. Scale bars: 1 mm
According to our observations the mites differed in their preferences to particular form of the slime mold material. Such preferences could be particularly well discernible when there were two or three forms of slime molds in a Petri dish. The mites concentrated around a particular form and then, after ingesting it, they moved to the other one. For example, if there was a dead fragment of plasmodium, besides a live one, the mites concentrated on the dead part first (e.g., Fig. 3e–g), and then moved to the live one. Thus, the following sequence of the preferences was established: dead or decaying plasmodium > spores/sporangia/aethalia > live plasmodium > sclerotia.
Features of various types of ecological interactions
Acarine parasites occur in most of the recognized acarine lineages, except for Opilioacarida (Krantz and Walter 2009; Walter and Proctor 2013). Most taxa among parasitic mites are parasites of vertebrates, invertebrates or plants (phytophages). However, at least T. putrescentiae displays an evident affinity to slime molds as well. Based on our observations, the interaction reported here appears closer to and stronger than what is called ‘insidious predation’ (Zelmer 1998) and seems much more than the food competition reported for T. putrescentiae (Xuan et al. 1993). Because live plasmodia and sclerotia were the least favorized stages of all appearing in the slime mold life cycle, it is disputable whether the mite is a parasite of slime molds. The relation should fulfil at least some criteria of the parasitism suggested by various authors, and/or at least some key features of this phenomenon should be depicted. It must be emphasized here that not all authors accept the full spectrum of the criteria listed below as typical of (and only of) parasitism. However, we can find among them the criteria fulfilled by the analyzed interaction:
1. The habitat relationship (Dogel’ 1966; Zelmer 1998; Weiner 2003). Judging by the ephemeral appearance of the mites in the slime mold cultures, the interaction revealed here is of a facultative or transient (periodic) nature, typical for an evolutionarily young relationship (Weiner 2003). What makes this relationship unique is that it is the microorganismal partner in this relationship, the slime mold, that is the host, and not the metazoan parasite, the mite.
2. The size relationship (Goff 1982; Weiner 2003). The maximal length and width of the body of a mature T. putrescentiae does not exceed 500–750 µm (Liu et al. 2006), whereas the size of a big plasmodium reared as a single individual may exceed hundreds of cm2 in area (Stephenson and Stempen 1994; Płonka and Rakoczy 1997; Adamatzky 2010). In our cultures, the area of a Petri dish (sometimes fully covered with the plasmodium) was ca. 60 cm2. In heavily infested cultures we found parts of the plasmodium ‘covered’ with mites side by side (Figs. 3a, g, 4b, 5d, Online Resources 1–3), and we estimated that the density of mites in such places could reach up to 250 per cm2, i.e., 20,000 mites per dish. It is comparable to the results by Smrž and Čatská (1987) who found up to 10,000 mites per dish of fungal cultures.
3. Immunity (Sprent 1962). The existence of naked plasmodia in the environment densely inhabited by microorganisms proves that the slime molds must possess some effective mechanisms of defense, like the evolutionarily ancient production of nitric oxide (Radomski et al. 1991; Płonka and Rakoczy 2000). Similarly, F. septica secretes fungicidal enzymes to the medium (Stephenson and Schnittler 2016). At present, we cannot identify any mechanism of the mycetozoal defense against the mites, but this may constitute a rationale for further research.
4. Evolution (decrease in host fitness, co-evolution, trophic selectivity and monophagy) (Crofton 1971a; Ewald 1995; Zelmer 1998; Weiner 2003). The overdispersed distribution of mites within the slime mold population, killing the heavily infected hosts (all stages of the slime mold life cycle, except spores), higher reproductive potential of the mite than the slime mold (the latter only exceptionally produces aethalia in vitro), and physiological dependence of the mite on the slime molds (only in mature cultures)—they all strongly point to a decrease in the host fitness in the cultures. However, some spores of Didymium sp. may germinate after passing through the digestive tract of a mite (Keller and Smith 1978), so the interaction sometimes bears the symptoms of protocooperation (facultative mutualism; Weiner 2003). A similar interaction between beetles and slime molds has been classified recently as mutualism (Sugiura et al. 2019). Similarly, mites of genus Tyrophagus sp. spread spores of ciliates, thus interacting positively, which was determined as mutualism and phoresy (Bharti et al. 2020).
5. The trophic relationship (Dogel’ 1966; Weiner 2003) or ‘dependence of the parasite on at least one gene of the host or on its product’ (MacInnis 1976; Zelmer 1998). This has been manifested strongly for at least several species of slime molds investigated here, and for different stages of their life cycle. We not only observed a quick disposal of a given type of slime mold material from the culture, but also a gradual filling of the digestive tract of the mites with the material of a particular coloration (Figs. 3, 4, 5, 6—white empty arrowheads)—white for the flour (Fig. 6), yellow for the plasmodium or the sclerotium (Figs. 3d, 5b), orange for the dead or decaying plasmodium (Fig. 3g), and brown/black for the spores (Figs. 3m, 4d, 5f; see also Keller and Smith 1978). The latter constitutes an evidence that the mites indeed feed on the mycetozoal material. Accordingly, the mites trophically depend on the slime mold, which not only provides the habitat, but also constitutes the source of food for the parasite.
Fig. 6.
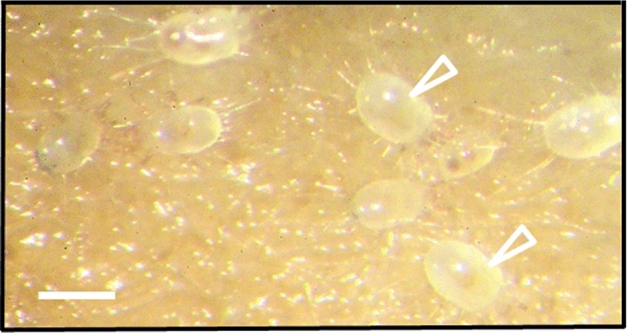
Tyrophagus putrescentiae in control cultures in Petri dishes with filter paper and oat flour but without slime molds. White empty arrowheads indicate mites with their digestive tracts filled with flour. Scale bar: 0.5 mm
It is a good opportunity to consider another type of relationship between the mite and the slime mold. In the in vitro cultures, oat flour is the main source of nutrients for the slime mold and for T. putrescentiae (Fig. 6, Online Resources 4), which may imply food competition (Xuan et al. 1993). In most cases T. putrescentiae has been regarded as synanthropic mite, whereas its occurrence in the soil has been commonly recorded (Smrž and Čatská 1987; Smrž et al. 2016). The major food tested for laboratory populations of this species have been stored products, yeasts or at least synanthropic fungi (Smrž and Čatská 1987; da Silvia et al. 2019; EPPO 2000). The food preference of T. putrescentiae towards fungal taxa indicates some specificity, but also plasticity in food requirements. The latter is also reflected in the array of vernacular names known for this mite (EPPO 2000).
Laboratory observations indicate that T. putrescentiae feeds on spores as well as on hyphae of a large range of dermatophytes, yeasts and molds, but does not feed on bacteria (Duek et al. 2001). The mites often migrate among cultures and thus may act as vectors. Hence, they can be considered as a potential pest of mycology stock cultures that may cause serious damage in microbiological laboratories (Duek et al. 2001). Sustaining of this mite on a variety of food with high protein and fat content, is likely to go beyond the feeding strategies typical of saprophagous organisms. Tyrophagus putrescentiae only partially reveals the characteristics of a true parasite, i.e., monophagy or a tendency towards trophic specialization (Weiner 2003). As a co-habitant of various species of slime molds, it cannot be called specific, nor is it found to co-evolve with its hosts. Importantly, to initiate a co-evolutionary relationship, a size relationship is necessary, often leading to trophic specialization of the co-evolving parasite (Weiner 2003).
Two more types of interactions should still be considered: parasitoidism and pseudo-parasitism. (1) With parasitoidism, the mite infestation can be a direct reason for the plasmodial death; however, the mites often feed on dead or decomposing plasmodia or sclerotia, thus becoming saprobionts or, better, necrophages (Weiner 2003). If feeding on live plasmodium or sclerotium really leads to necrosis of the material, the mite should be regarded here as a ‘facultative’ parasitoid (Croll 1973; Ewald 1995; Weiner 2003). Facultative, because transition from live to dead host body during mite feeding is not obligatory. (2) Pseudo-parasitism—defined as a transient, accidental presence and survival of the invading, normally free-living organism in the organism of the pseudo-host (Złotorzycka et al. 1998)—cannot be excluded, as the mites are free-living and lead a saprobiont-like life under natural conditions. However, in presence of the slime mold, the mites reveal strong affinity to the mycetozoan, and this situation should not be regarded as ‘accidental’ (Online Resources 1). It seems crucial to find out whether such a relation occurs also in the natural environment.
Judging by the time that elapsed between the publication by Keller and Smith (1978) and our observations (2013–2020), the appearance of T. putrescentiae in cultures of slime molds is not just a coincidence but an established affinity. It is evident that depending on the stage of life cycle of the slime mold, the type of interaction between the slime mold and the mite keeps changing. This is another feature of an evolutionarily young interaction (Ewald 1995; Weiner 2003), so it is particularly important to find examples of relationships between other mite species (and other organisms) and/or other types of slime molds. One should recall that Tyrophagus sp. enter also sophisticated and peculiar interactions with parasitic fungi (Xuan et al. 1993), and with ciliates (Bharti et al. 2020). The existing interactions may be a good starting point for further transformations, defining this group of mites as organisms of high evolutionary potential. Follow-up (mathematical) model studies (Crofton 1971a, b; Weiner 2003) will help paint a clear picture of such interactions in their various guises. The mycetozoan laboratory cultures in vitro, as vastly controllable, seem to constitute a particularly suitable system to explore such interactions.
Conclusions
Tyrophagus putrescentiae and the slime molds F. septica, P. polycephalum, and Didymium sp. kept under in vitro conditions create an interesting system of mutual interactions bearing symptoms of several known ecological interactions (i.e., parasitism, pseudo-parasitism, parasitoidism, insidious predation, grazing, trophic competition, necrophagy, mutualism and phoresy) at various stages of their life cycle. Given the growing interest in slime molds, this plethora of relationships create a convenient model of study for further research, both under laboratory conditions and in the natural environment.
Supplementary Information
Below is the link to the electronic supplementary material.
Acknowledgements
The authors are grateful to Prof. Andrew Adamatzky (University of the West of England, Bristol, UK) for the cultures of Physarum polycephalum, and to Prof. Anna Drozdowicz (Institute of Botany, Faculty of Biology, Jagiellonian University in Kraków, Poland) for her help in identification of Fuligo septica. We thank the anonymous reviewers whose comments largely improved the manuscript.
Author contributions
Mycetozoa (65%): DMW, MJ, PMP. Mites (35%): MF, DS, JM. DMW performed the majority of observations, prepared the majority of photos, pictures (except Fig. 2b–f) and movies, contributed to the writing of the text, 25%. MJ contributed to cultivation of the slime molds, photo documented some specimens, and revised the text 15%. PMP authored the concept and design of the paper, participated in observations, substantially participated in the writing of the manuscript 25%. MF participated in identification of mites, in preparing Fig. 2b–f, contributed to the preparation of the text 10%. DS co-authored the idea, participated in the text preparation, 5%. JM participated in the identification of the mite, substantially contributed to the final shape of the text 20%.
Funding
The Faculty of Biochemistry, Biophysics, and Biotechnology of the Jagiellonian University was a partner of the Leading National Research Center (KNOW) supported by the Polish Ministry of Science and Higher Education.
Data availability
The vouchers of the mite are deposited at the Department of Invertebrate Systematics and Ecology, Wrocław University of Environmental and Life Sciences, Poland. All the data and photo documentation are available from the authors on request.
Declarations
Conflict of interest
The authors do not declare any conflict of interest, related to this paper.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Joanna Mąkol, Email: joanna.makol@upwr.edu.pl.
Przemysław M. Płonka, Email: przemyslaw.plonka@uj.edu.pl
References
- Adamatzky A. Physarum machines: computers from slime mould. Hackensack: World Scientific; 2010. [Google Scholar]
- Amoah BA (2016) Monitoring populations of the ham mite, Tyrophagus putrescentiae (Schrank) (Acari: Acaridae): research on traps, orientation behavior, and sampling techniques. PhD dissertation, Department of Entomology, College of Agriculture, Kansas State University. http://hdl.handle.net/2097/32489
- Arthaud J, Lafon R. Un Myxomycète sur nos Fraisiers. Diachea leucopoda Rost. Rev Zool Agric Pathol Veg. 1972;4:145–147. [Google Scholar]
- Bharti D, Kumar S, La Terza A, Chandra K. Dispersal of ciliated protist cysts: mutualism and phoresy on mites. Ecology. 2020;101(9):e03075. doi: 10.1002/ecy.3075. [DOI] [PubMed] [Google Scholar]
- Chung C-H, Liu C-H, Tzean SS. Slime molds in edible mushroom cultivation sites. Plant Pathol Bull. 1998;7:141–146. [Google Scholar]
- Clark J, Mires A. Biosystematics of Didymium: the noncalcareous, long-stalked species. Mycotaxon. 1999;71:369–382. [Google Scholar]
- Clark J, Stephenson SL, Landolt JC. Biosystematics of the Didymium iridis super species complex: additional isolates. Mycotaxon. 2001;79:447–454. [Google Scholar]
- Clark J, Haskins EF, Stephenson SL. Culture and reproductive systems of 11 species of Mycetozoans. Mycologia. 2004;96:36–40. doi: 10.1080/15572536.2005.11832994. [DOI] [PubMed] [Google Scholar]
- Crofton HD. A quantitative approach to parasitism. Parasitology. 1971;62:179–193. doi: 10.1017/S0031182000071420. [DOI] [Google Scholar]
- Crofton HD. A model of host-parasite relationships. Parasitology. 1971;63:343–364. doi: 10.1017/S0031182000079890. [DOI] [PubMed] [Google Scholar]
- Croll NA. Parasitism and other associations. London: Pitman Medical; 1973. [Google Scholar]
- Díez J, Moreno G, Del Peral L, Adams JH, Jr, Rodríguez Frías MD, Manjón JL. Fuligo septica spores onboard of a stratospheric NASA balloon and its complete in vitro life cycle. Astrobiology. 2020;20:394–404. doi: 10.1089/ast.2019.2097. [DOI] [PubMed] [Google Scholar]
- Dogel’ VA (1966) General parasitology, [Revised and enlarged by Polyansky EI, Kheisin E M, transl. from Russian by Kabata Z]. Academic Press, New York
- Drozdowicz A, Ronikier A, Stojanowska W, Panek E. Mycetozoas of Poland. A checklist. Kraków: W Szafer Institute of Botany Polish Academy of Sciences; 2003. [Google Scholar]
- Duek L, Kaufman G, Palevsky E, Berdicevsky I. Mites in fungal cultures. Mycoses. 2001;44:390–394. doi: 10.1046/j.1439-0507.2001.00684.x. [DOI] [PubMed] [Google Scholar]
- ElHage N, Little C, Clark J, Stephenson SL. Biosystematics of the Didymium squamulosum complex. Mycologia. 2000;92:54–64. doi: 10.1080/00275514.2000.12061130. [DOI] [Google Scholar]
- EPPO Global Database (2000) Tyrophagus putrescentiae (TYROPU). European and Mediterranean Plant Protection Organization (EPPO). https://gd.eppo.int/taxon/TYROPU. Accessed 08 Apr 2021
- Espinosa A, Paz-Y-Miño-C G. Evidence of taxa-, clone-, and kin-discrimination in protists: ecological and evolutionary implications. Evol Ecol. 2014;28:019–1029. doi: 10.1007/s10682-014-9721-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ewald PW. The evolution of virulence: a unifying link between parasitology and ecology. J Parasitol. 1995;81:659–669. doi: 10.2307/3283951. [DOI] [PubMed] [Google Scholar]
- Goff LJ. Symbiosis and parasitism: another viewpoint. Bioscience. 1982;32:255–256. doi: 10.2307/1308531. [DOI] [Google Scholar]
- Hagstrum DW, Klejdysz T, Subramanyam B, Nawrot J. Atlas of stored-product insects and mites. St Paul: AACC International; 2013. [Google Scholar]
- Hamilton WD. Narrow roads of gene land. Evolution of sex. New York: Oxford University Press Inc; 2001. [Google Scholar]
- Haskins EF, Wrigley de Basanta D. Methods of agar culture of Myxomycetes: an overview. Rev Mex Mic. 2008;27:1–7. [Google Scholar]
- Hughes AM (1961) The mites of stored food. Technical Bulletin of the Ministry of Agriculture, Fisheries and Food. Tech Bull Min Agric Fish Food 9:1–274
- Keller HW, Smith DM. Dissemination of myxomycete spores through the feeding activities (ingestion–defecation) of an acarid mite. Mycologia. 1978;70:1239–1241. doi: 10.1080/00275514.1978.12020341. [DOI] [Google Scholar]
- Keller HW, Snell KL. Feeding activities of slugs on Myxomycetes and macrofungi. Mycologia. 2002;94:757–760. doi: 10.1080/15572536.2003.11833169. [DOI] [PubMed] [Google Scholar]
- Kocourková J. Lichenicolous fungi of the Czech Republic (the first commented checklist) Acta Mus Nat Pragae Ser B Hist Nat. 1999;55:59–169. [Google Scholar]
- Krantz GW, Walter DE. A manual of acarology. 3. Lubbock: Texas Tech University Press; 2009. [Google Scholar]
- Krzywda A, Petelenz E, Michalczyk D, Płonka PM. Sclerotia of the acellular (true) slime mould Fuligo septica as a model to study melanization and anabiosis. Cell Mol Biol Lett. 2008;13:130–143. doi: 10.2478/s11658-007-0047-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu T, Jin D-C, Guo JJ, Li L. Development and growth of Tyrophagus putrescentiae (Schrank) (Acarina: Acaridae) bred under different temperatures with different nutriments. Acta Entomol Sinica. 2006;49:714–718. [Google Scholar]
- MacInnis AJ. How parasites find their hosts: some thoughts on the inception of host-parasite integration. In: Kennedy CR, editor. Ecological aspects of parasitology. Amsterdam: North-Holland; 1976. pp. 3–20. [Google Scholar]
- Madelin MF. Presidential address—Myxomycete data of ecological significance. Trans Br Mycol Soc. 1984;83:1–19. doi: 10.1016/S0007-1536(84)80240-5. [DOI] [Google Scholar]
- Michalczyk D, Drozdowicz A, Pintscher S, Płonka PM. Mycetozoal bloom in a hydroponic culture of garden cress (Lepidium sativum L.) Int J Food Microbiol. 2011;145:336–341. doi: 10.1016/j.ijfoodmicro.2010.11.042. [DOI] [PubMed] [Google Scholar]
- Murugan M, Mubarak Ali N, Suju Skaria C, Kalyanasundaram I. Agarolytic activity of the myxomycete Fuligo septica. Mycologist. 1996;10:21–22. doi: 10.1016/S0269-915X(96)80045-0. [DOI] [Google Scholar]
- Płonka PM, Rakoczy L. The electron paramagnetic resonance signals of the acellular slime mould Physarum nudum plasmodia irradiated with white light. Curr Top Biophys. 1997;21:83–86. [Google Scholar]
- Płonka PM, Rakoczy L. Heme and non-heme iron complexes of nitric oxide in the plasmodia of acellular slime moulds cultured in vitro. Zesz Probl Post N Roln. 2000;473:249–259. [Google Scholar]
- Radomski M, Martin JF, Moncada S. Synthesis of nitric oxide by the hemocyte of the American horseshoe crab Limulus polyphemus. Philos Trans R Soc Lond B. 1991;334:129–133. doi: 10.1098/rstb.1991.0102. [DOI] [Google Scholar]
- Rakoczy L, Szymański J. Slime moulds in mushroom growings. Owoce, Warzywa. Kwiaty. 1985;13:9–10. [Google Scholar]
- Rentfrow G, Hanson DJ, Schilling MW, Mikel WB (2006) Methyl bromide use to combat mite infestation in dry-cured ham during production. In: Proceedings, annual international research conference on methyl bromide alternatives and emission reduction, Orlando, FL, 5–9 November 2006
- da Silva GL, Esswein IZ, Heidrich D, Dresch F, Maciel MJ, Pagani DM, Valente P, Scroferneker ML, Johann L, Ferla NJ, da Silva OS. Population growth of the stored product pest Tyrophagus putrescentiae (Acari: Acaridae) on environmentally and medically important fungi. Exp Appl Acarol. 2019;78:49–64. doi: 10.1007/s10493-019-00370-8. [DOI] [PubMed] [Google Scholar]
- Smith DM, Keller HW. Spore to spore cultivation of a new species of Didymium mycetozoas in association with an acarid mite Tyrophagus putrescentiae. ASB Bull. 1978;25:67. [Google Scholar]
- Smrž J, Čatská V. Food selection of the field population of Tyrophagus putrescentiae (Schrank) (Acari, Acaridida) J Appl Entomol. 1987;104:329–335. doi: 10.1111/j.1439-0418.1987.tb00533.x. [DOI] [Google Scholar]
- Smrž J, Soukalová H, Čatská V, Hubert J. Feeding patterns of Tyrophagus putrescentiae (Sarcoptiformes: Acaridae) indicate that mycophagy is not a single and homogeneous category of nutritional biology. J Insect Sci. 2016;16(94):1–8. doi: 10.1093/jisesa/iew070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solarz K. House dust mites and storage mites (Acari: Oribatida: Astigmatina). Identification keys. Kraków: Institute of Systematics and Evolution of Animals Polish Academy of Sciences; 2012. [Google Scholar]
- Sprent JFA. Parasitism, immunity, and evolution. In: Leeper GW, editor. The evolution of living organisms: a symposium to mark the centenary of Darwin's origin of species and of the Royal Society of Victoria held in Melbourne, December 1959. Melbourne: Melbourne University Press; 1962. [Google Scholar]
- Stephenson SL. Distribution and ecology of mycetozoans in temperate forests. II. Patterns of occurrence on bark surface of living trees, leaf litter, and dung. Mycologia. 1989;81:608–621. doi: 10.1080/00275514.1989.12025792. [DOI] [Google Scholar]
- Stephenson SL, Stempen H. Myxomycetes: a handbook of slime molds. Portland: Timber Press; 1994. [Google Scholar]
- Stephenson SL. Mycetozoas of New Zealand. Fungi of New Zealand, Vol. 3. Fungal Divers Res Ser. 2003;11:1–238. [Google Scholar]
- Stephenson SL, Schnittler M. Mycetozoas. In: Archibald JM, Simpson AGB, Stamovitz CH, editors. Handbook of the protists. Cham: Springer; 2016. [Google Scholar]
- Sugiura S, Fukasawa Y, Ogawa R, Kawakami S-I, Yamazaki K. Cross-kingdom interactions between slime molds and arthropods: a spore dispersal mutualism hypothesis. Ecology. 2019;100(8):e02702. doi: 10.1002/ecy.2702. [DOI] [PubMed] [Google Scholar]
- Swan DC. Berlese’s fluid: remarks upon its preparation and use as a mounting medium. Bull Entomol Res. 1936;27:389–391. doi: 10.1017/S0007485300058259. [DOI] [Google Scholar]
- Walter DE, Proctor H. Mites: ecology, evolution & behaviour. Dordrecht: Springer; 2013. [Google Scholar]
- Weiner J (2003) Życie i ewolucja biosfery. Podręcznik ekologii ogólnej [Life and evolution of biosphere. A handbook on general ecology]. PWN [State Scientific Editors PWN SA], Warszawa (in Polish)
- Xuan JY, Wang ZS, Wang CB, Yu SD. Preliminary research of relationship between Tyrophagus putrescentiae (Schrank) and two parasitic fungi. GRAIN Storage. 1993;22:21–26. [Google Scholar]
- Zelmer DA. An evolutionary definition of parasitism. Int J Parasitol. 1998;28:531–533. doi: 10.1016/S0020-7519(97)00199-9. [DOI] [PubMed] [Google Scholar]
- Złotorzycka J (ed), Lonc E, Majewska AC, Okulewicz A, Pojmańska T, Wędrychowicz H (1998) Słownik parazytologiczny [The Parasitological Dictionary]. Polskie Towarzystwo Parazytologiczne [Polish Parasitological Society], Warszawa (in Polish)
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The vouchers of the mite are deposited at the Department of Invertebrate Systematics and Ecology, Wrocław University of Environmental and Life Sciences, Poland. All the data and photo documentation are available from the authors on request.