

Abstract
Stress resistance correlates with longevity and this pattern has been exploited to help identify genes that can influence lifespan. Reciprocally, genes and pharmacological agents that have been studied primarily in the context of longevity may be an untapped resource for treating acute stresses. Here we summarize the evidence that targeting SIRT1, studied primarily in the context of longevity, can improve outcomes in hemorrhagic shock and resuscitation. Hemorrhagic shock is a potentially fatal condition that occurs when blood loss is so severe that tissues no longer receive adequate oxygen. While stabilizing the blood pressure and reperfusing tissues are necessary, re-introducing oxygen to ischemic tissues generates a burst of reactive oxygen species that can cause secondary tissue damage. Reactive oxygen species not only exacerbate the inflammatory cascade but also can directly damage mitochondria, leading to bioenergetic failure in the affected tissues. Treatments with polyphenol resveratrol and with nicotinamide adenine dinucleotide (NAD) precursors have both shown promising results in rodent models of hemorrhagic shock and resuscitation. Although a number of different mechanisms may be at play in each case, a common theme is that resveratrol and NAD both enhance the activity of SIRT1. Moreover, many of the physiologic improvements observed with resveratrol and NAD precursors are consistent with modulation of known SIRT1 targets. Because small blood vessels and limited blood volume make mice very challenging for the development of hemorrhagic shock models, there is a paucity of direct genetic evidence testing the role of SIRT1. However, the development of more robust methods in mice as well as genetic modifications in rats should allow the study of SIRT1 transgenic and KO rodents in the near future. The potential therapeutic effect of SIRT1 in hemorrhagic shock may serve as an important example supporting the value of considering “longevity” pathways in the mitigation of acute stresses.
Keywords: Hemorrhagic shock, NAD, Sirtuins, Aging, Stress, Riboside, Nicotinamide, Mononucleotide, SIRT1, Resveratrol
Longevity and resilience
It has long been appreciated that there is a correlation between longevity and stress resistance. Human centenarians achieve their longevity not by surviving a series of ailments but by remaining remarkably healthier over time than their counterparts—in fact, compared to the general population, centenarians on average spend less time suffering from disabilities, despite their longer lives [1]. The same is true for behaviors that influence longevity. Factors that shorten life, such as obesity and smoking, simultaneously increase susceptibility to a host of conditions that go well beyond the obvious and well-known associations [2, 3]. For example, a weight gain of 15 kg for men or 13 kg for women is estimated to increase the risks of certain cancers by more than 50% [4], and smoking is associated with impairments in hearing [5], vision [6], and fertility [7, 8]. On the other hand, regular exercise increases longevity and decreases the risk of almost every major cause of death in the developed world [9, 10]. Similarly, calorie restriction (CR), which is the most robust and reproducible intervention known to extend life in model organisms [11, 12], confers resistance to chronic diseases as well as many acute stresses including ischemic injuries [13, 14], radiation [15], and multiple toxins and oxidative stressors [16, 17]. Recently, it has been shown that relatively short-term fasting can confer stress resistance that resembles the effect of longer-term CR. For example, fasted animals tolerate the side effects of chemotherapy drugs better than fed animals and are protected from ischemic injuries during surgical procedures [18]. Although CR has not been proven to extend life in humans, it does improve many indicators of general health and delays age-related diseases in primates, suggesting that many of its benefits are likely to be conserved [11, 19–21].
The general association between longevity and stress resistance has been exploited experimentally to identify longevity genes in model organisms. Survivors of acute stresses are enriched for mutations that also confer increased longevity. Hits from such a screen in yeast led directly to the discovery of sirtuins (homologs of the yeast silent information regulator 2, SIR2) as genes that can influence lifespan in model organisms (albeit with some controversy) [22–28] and are required for some aspects of the response to CR in mammals [29–31]. While new data have continued to support the notion that sirtuins and other longevity genes boost stress resistance in lower organisms, the majority of research in mammals has been focused on their potential to impact aging and chronic diseases [32]. Here we discuss the evidence that targeting sirtuin 1 (SIRT1) may be a viable therapeutic strategy to improve outcomes in hemorrhagic shock, a type of severe acute stress. These data may serve as an example to support exploring the potential of longevity pathways in general to promote short-term resilience.
Hemorrhagic shock
Hemorrhagic shock occurs when severe blood loss leads to inadequate tissue oxygen delivery and is often fatal if not promptly treated. Although acute injuries are often not thought of in the context of aging, outcomes are drastically worse in the elderly [33]. Hemorrhagic shock can occur in a variety of clinical situations (e.g., childbirth, gastrointestinal hemorrhage, and aneurysmal rupture), but traumatic injury remains the most common cause and accounts for an estimated 1.5 million deaths per year worldwide [34]. Clinically, hemorrhagic shock is treated with a rapid control of bleeding in conjunction with intravenous fluid and blood products. Thus, poor access to emergency medical services and limited transfusion resources can negatively impact survival.
Unfortunately, even with prompt resuscitation, patients who survive the initial period of hemorrhagic shock are at risk of developing multi-organ failure (MOF). During acute blood loss, poorly perfused tissues resort to anaerobic metabolism, becoming progressively more dysfunctional with declining ATP levels and lactic acid buildup [35]. Although absolutely essential, restoring perfusion and oxygen can further contribute to organ dysfunction. Reperfusing hypoxic tissues releases a storm of free radical oxygen species, toxic metabolites, and inflammatory cytokines—all of which can lead to mitochondrial dysfunction and activate a systemic inflammatory response that further damages critical organs [36]. In fact, MOF is the leading cause of late death (after 7 days) in trauma [37]. As such, strategies that improve resilience to the acute stress of hemorrhagic shock could significantly decrease the incidence of MOF and improve survival following hemorrhagic shock.
Potential benefits of SIRT1 activation in hemorrhagic shock
SIRT1, in contrast to its perception as a long-term modulator of general health, performs multiple actions that may be acutely beneficial during hemorrhagic shock and reperfusion. These include suppressing inflammation, mitigating oxidative damage, promoting mitochondrial activity, and rescuing vascular endothelial function (Fig. 1). SIRT1 has many downstream effects, including deacetylation and inactivation of the ReIA subunit of NFκB, a central mediator of the immune response that is responsible for the transcription of hundreds of pro-inflammatory cytokines and chemokines [38, 39]. By inhibiting NFκB, SIRT1 inhibits the transcription of pro-inflammatory genes, including IL6, TNF-α, CINC, COX2, and ICAM-1 [39, 40]. In non-hemorrhage models, inhibition of SIRT1 promotes the expression of tissue factor and clot propagation mediated by NFκB [41], indicating that SIRT1 may play a role in post-hemorrhage coagulopathy. However, its effects on coagulation have not yet been studied in a hemorrhagic shock model. During reperfusion following hemorrhagic shock, reactive oxygen species (ROS) are produced by dysfunctional mitochondria and by nicotinamide adenine dinucleotide phosphate oxidases (NOX) [42]. ROS can cause oxidative damage to nucleic acids and proteins leading to cell damage and leukocyte recruitment. In addition, ROS can damage mitochondria, which can further increase ROS production in a cycle that prevents energetic recovery after perfusion [43]. Accordingly, therapies that decrease free radical damage can improve organ function following hemorrhagic shock and reperfusion (HSR) [44–46]. Upregulation of antioxidant enzymes such as superoxide dismutase (SOD2) can be achieved through SIRT1 deacetylating and activating forkhead box O3 (FOXO3) [47]. SIRT1 activates PGC1α, which is known as the master regulator of mitochondrial biogenesis [48]. Along with the induction of genes required to increase mitochondrial abundance, PGC1α promotes transcription of antioxidant genes such as SOD2, HO-1, and glutathione reductase [49]. Additionally, pharmacological activation or genetic overexpression of SIRT1 has been shown to decrease defective autophagy and improve mitochondrial stability in ischemic livers, thus reducing tissue damage [50] (Fig. 2).
Fig. 1.
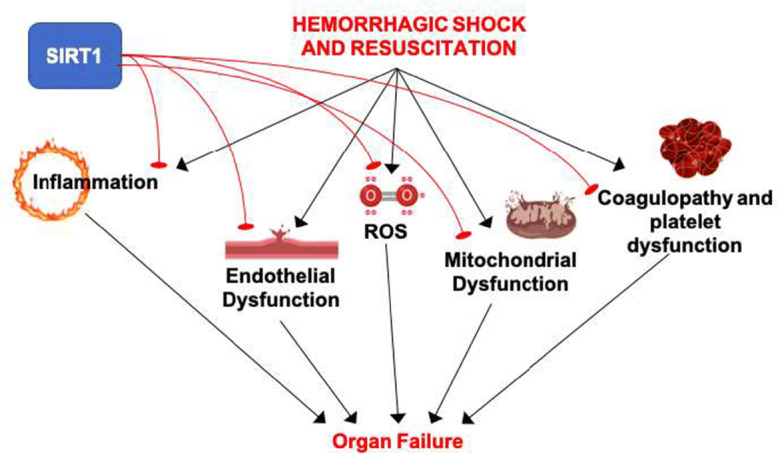
Effects of hemorrhagic shock and reperfusion that may be mitigated by SIRT1. Hemorrhagic shock in trauma results in activation of the inflammatory response, vascular endothelial dysfunction, oxidative stress, mitochondrial dysfunction, and platelet dysfunction and coagulopathy. Together, these effects contribute to organ dysfunction and multi-organ failure. SIRT1 inhibits the inflammatory response, promotes vascular endothelial function, decreases ROS, improves mitochondrial function, and can improve coagulopathy in different settings. Thus, SIRT1 has the potential to mitigate the key features of hemorrhagic shock and resuscitation that lead to organ damage and thereby improve survival
Fig. 2.

SIRT1 activation is a common effect of resveratrol and NAD that may drive beneficial effects in hemorrhagic shock and reperfusion. Resveratrol not only directly activates SIRT1 but may also indirectly activate it by increasing NAD levels via the AMPK pathway. Resveratrol inhibits COX2 leading to reduced platelet aggregation and stimulates the estrogen receptor PI3K/AKT pathway. Activation of this pathway promotes vascular endothelial function, decreases inflammation, and protects against oxidative stress. NAD is required by SIRT1 as well as the other sirtuins (SIRT2–7). NAD also plays essential roles in redox metabolism and is a substrate for CD38 and PARPs. CD38 and PARPs can contribute to inflammation and apoptosis, and by competing for NAD, may reduce SIRT1 activity. SIRT1 acts via multiple downstream pathways that could ameliorate damage in hemorrhagic shock and resuscitation. It inhibits NFκB, in turn inhibiting the expression of multiple downstream pro-inflammatory cytokines. By inhibiting p53, SIRT1 can suppress cellular apoptosis and mitigate mitochondrial dysfunction. SIRT1 activates FOXO3 and PGC1α to increase antioxidant gene expression and increase mitochondrial biogenesis and respiratory activity. SIRT1 also differentially modulates the hypoxia-inducible factors HIF1α and HIF2α, which may favor mitochondrial respiration while promoting erythropoietin expression. Together, these effects are expected to increase cellular ATP levels, preserve vascular endothelial cell function, and ameliorate organ dysfunction
The benefits of SIRT1 activation during periods of physiological stress in vivo have been documented in multiple studies. For instance, the loss of SIRT1 significantly increased the risk of death in hepatocytes following ischemia/reperfusion injury [50]. In contrast, SIRT1 overexpression decreased oxidative stress following cardiac ischemia/reperfusion. When compared with wild-type mice, SIRT1 transgenic hearts not only had increased expression of superoxide dismutase but also had decreased myocardial tissue infarction and improved cardiac contractility [51]. Similarly, SIRT1 has been found to decrease cerebral infarct volume following ischemia in mice via inhibition of NLRP3 inflammasome activation [52]. Thus, there are multiple lines of evidence to support the hypothesis that therapies targeting SIRT1 would be beneficial in HSR. Unfortunately, there is a lack of direct genetic evidence in HSR due mainly to the relatively small blood vessels and blood volume of mice compared to rats, causing difficulty in creating a hemorrhagic shock model in mice.
Therapeutic pathways targeting SIRT1 in hemorrhagic shock
Resveratrol and polydatin
Because of the challenges of obtaining direct genetic evidence, most HSR research has utilized chemical modulators of SIRT1, such as the SIRT1 activator resveratrol. Resveratrol, a naturally occurring molecule found in grapes, has been shown to reduce inflammation and oxidative stress in a variety of conditions. Although described as an allosteric activator of SIRT1 [53, 54], resveratrol may increase SIRT1 activity through multiple different mechanisms including activating AMPK [55, 56] which promotes expression of NAMPT [57]; this in turn increases the amount of available nicotinamide adenine dinucleotide (NAD), an essential co-substrate for SIRT1 [58–60]. Recently, resveratrol and polydatin, a natural resveratrol glucoside, have been investigated as potential treatments for hemorrhagic shock [61]. Although resveratrol does have multiple targets as well as its own antioxidant properties, various studies have suggested that its effects in hemorrhagic shock are at least partially, if not principally, mediated by SIRT1 [62–74].
In addition to increasing SIRT1’s activity, resveratrol increases the levels of SIRT1 mRNA expression [67, 72, 73, 75]. It also increases the expression and activity of PGC1α, a promoter of antioxidant activity and mitochondrial biogenesis that can control its own expression. This is thought to occur via deacetylation and thus direct activation of PGC1α by SIRT1 [48, 54, 67, 76]. Consistently, treatment with resveratrol following HSR leads to increased levels of ATP, increased expression of SOD2 and catalase, and decreased ROS and lipid peroxidation, a marker of oxidative stress. Resveratrol treatment improved mitochondrial respiratory function after HSR, as evidenced by increased complex I, II, and IV activity, and mitigated renal injury [66, 67, 77]. Similar effects were observed in cardiac tissue and arteriolar smooth muscle cells, where resveratrol treatment was able to preserve the levels of PGC1α, increase the expression of key mitochondrial transcription factors, and improve myocardial contractility, cardiac output, mean arterial pressure (MAP), and survival in a SIRT1-dependent manner [63, 64, 73, 77]. Polydatin, which appears to work at least partially via SIRT1-mediated deacetylation of p53, also decreases ROS and ameliorates mitochondrial damage in HSR. Treatment with polydatin decreased mitochondrial swelling, inhibited mitochondrial membrane depolarization, thus decreasing membrane permeability, and inhibited apoptosis in multiple different cell types [78–80]. Various studies have found that these changes were associated with decreased tissue damage in the small intestine [81], improved renal function, increased MAP, and a survival benefit [78, 82]. Mitochondrial dysfunction and reduced cardiac output after hemorrhagic shock are known to cause increased cellular dysfunction and tissue damage, and treatment with resveratrol and polydatin could ameliorate these downstream effects through a SIRT1-dependent mechanism. Improved cardiac output could likewise improve post-shock tissue damage by improving organ perfusion [63, 67, 75, 77–80]. Therefore, SIRT1’s ability to improve mitochondrial activity and oxidative stress not only confers resistance against chronic stresses during aging but also protects organisms from the acute stress of hemorrhage.
While SIRT1-dependent mechanisms of resveratrol do play an important role in its ability to ameliorate tissue damage following hemorrhagic shock, resveratrol also has SIRT1-independent mechanisms of action. Resveratrol has been found to decrease NOX, a major source of vascular ROS [83], mitigate endothelial dysfunction, and improve aortic vasoreactivity after hemorrhagic shock. Resveratrol can also inhibit cyclooxygenase 2 (COX2), thereby inhibiting platelet aggregation and inflammation [84]. Multiple studies have shown decreased levels of cytokines, chemokines, and adhesion molecules leading to decreased neutrophil migration and improved organ function in mice treated with resveratrol after HSR. Upregulation of HO-1 via the estrogen receptor–mediated PI3K/AKT pathway may play a key role in these findings [65, 68–70, 85–87]. While estrogen receptor–dependent activation of HO-1 provides a SIRT1-independent pathway, there is substantial evidence to intimate that preservation of endothelial function after HSR, which improves vasoresponsiveness and MAP, depends on SIRT1 activity [63, 68, 70, 77]. Thus, although resveratrol does have its own antioxidant and anti-inflammatory activity, SIRT1 is likely necessary for many of its key benefits.
NMN, niacin, and other NAD precursors
NAD plays a key role in cellular metabolism and redox reactions. Not only does NAD drive oxidative phosphorylation by accepting reducing equivalents and providing them to complex I, it also serves as an essential co-substrate for a number of enzymes involved in cellular resilience including SIRT1 [88]. Moreover, NAD falls with age in some tissues and chronic treatment with precursor supplements decreases age-related degeneration in mice [89–91]. Although NAD can be synthesized de novo, the majority in most tissues is recycled from nicotinamide. Nicotinamide is converted to nicotinamide mononucleotide (NMN) in a rate-limiting reaction catalyzed by nicotinamide phosphoribosyltransferase (NAMPT) [92]. Depending on its subcellular location, NMN is then converted to NAD by one of the three different NMN adenylyltransferase isoforms (NMNAT1–3) [93] (Fig. 3). NAD is critical for cellular function and its depletion leads to mitochondrial dysfunction and cellular death. In contrast, enhancing NAD levels promotes cellular resilience and improves function [94–98].
Fig. 3.

NAD salvage pathway and hemorrhagic shock. NAD+ is a key reduction-oxidation (redox) cofactor that accepts high-energy electrons in the form of a hydride ion (H−) during glycolysis and the TCA cycle, generating NADH. Electrons are subsequently donated to complex I of the mitochondrial electron transport chain to support ATP generation, or to pyruvate via lactate dehydrogenase in order to recycle NAD+. NAD also serves as a substrate for several classes of enzymes, including SIRT1 and other sirtuins, which can modulate a broad range of stress responses and other behaviors. These reactions consume NAD, which necessitates regeneration of the pool to support cellular metabolism. Some NAD is synthesized from dietary sources including tryptophan, niacin, and NAD itself or synthetic intermediates, but the bulk of synthesis results from recycling of nicotinamide. This begins with an energetically costly reaction catalyzed by the enzyme NAMPT to make nicotinamide mononucleotide (NMN). NMNAT1–3 then convert NMN to NAD to complete the recycling process. Hemorrhagic shock causes damage and inflammation that can activate NAD consumers such as PARPs and CD38, and may further decrease the levels of NAD by causing a redox shift in favor of NADH due to the limited activity of the electron transport chain in hypoxic conditions, as well as by limiting the NAMPT activity due to lack of ATP. In this case, providing exogenous NMN directly during HS may be especially beneficial by bypassing the NAMPT reaction, thus increasing the NAD levels
During hemorrhagic shock, the tissue levels of NAD fall proportionally to the degree and duration of hemorrhagic shock [99]. As such, supplementing NAD either preinjury or during resuscitation could potentially be beneficial. The use of NAD, nicotinamide, and niacin to treat hemorrhagic shock was first described by Chaudry et al. in 1976 [100]. Although this approach did enhance the NAD levels in both the kidney and liver tissues, it did not restore the ATP levels nor improve survival. These negative results, however, may have been a consequence of the technique. The resuscitation protocol used in these early experiments provided minimal restoration of blood volume which could have limited the therapeutic potential of NAD supplementation. Indeed, more recently, high-dose oral niacin has been shown to significantly mitigate lung damage, decrease inflammation, and improve survival when given in conjunction with appropriate volume resuscitation [101]. Concordantly, inhibition of poly (ADP-ribose) polymerase (PARP), an NAD-consuming enzyme that is activated during hemorrhagic shock, is therapeutic in preclinical models [102].
Supplementing NMN may be particularly helpful because it bypasses the rate-limiting NAMPT conversion step and increases tissue NAD levels rapidly [103]. Not only is the NAMPT-catalyzed reaction energetically costly, but also the activity of this critical enzyme may be suppressed in the setting of ischemia/reperfusion [104]. Providing NMN, therefore, directly circumvents this metabolic constraint, and there is increasing data to suggest that NMN can mitigate the damage associated with a wide range of ischemia-reperfusion events in a SIRT1-dependent fashion [105–108].
We have recently demonstrated that the use of exogenous NMN significantly improves both cellular function and physiologic reserve in a rodent model of decompensated hemorrhagic shock [109]. Animals pretreated with NMN not only demonstrated increased tissue NAD levels, but also the severe mitochondrial dysfunction normally observed post-resuscitation in the kidney and liver tissues was entirely absent. Moreover, NMN treatment decreased both systemic and tissue-level inflammation. Importantly, NMN increased the time animals could tolerate severe shock before decompensating by nearly 25% and significantly improved survival from roughly 10 to 55% at 48 h [109]. A more recent study not only replicated the protective effects of NMN but also showed that a lower dose of niacin was effective and that the low-dose niacin effect depends on the niacin receptor (GPR109a) [110]. Low-dose intravenous niacin was sufficient to prolong survival in an otherwise lethal model of hemorrhagic shock even in the absence of fluid resuscitation [110]. Thus, it appears that NAD precursors may have therapeutic effects through two distinct mechanisms, a GPR109a pathway that is unique to niacin and an NAD/SIRT1-dependent pathway that is likely shared among precursors. Although more research is required, there is compelling preclinical data to suggest that NAD precursors including NMN may be one strategy to improve resilience, mitigate inflammation, improve cellular metabolism, and promote survival following hemorrhagic shock.
Further considerations and potential pitfalls
The activation of SIRT1 appears to be largely advantageous during reperfusion; however, there are also risks that should be carefully considered. For example, modulating SIRT1 may interfere with normal homeostatic pathways in acute hypoxia. Hypoxia-inducible factor 1α (HIF1α) and HIF2α are deacetylated by SIRT1, resulting in inhibition and activation, respectively [72, 111]. Acetylated HIF1α can stimulate various adaptive metabolic responses, such as the transition from oxidative phosphorylation to anaerobic metabolism, which could potentially improve cell survival under conditions of hypoperfusion [112]. Additionally, some findings suggest that as deacetylated HIF1α accumulates in cells during hypoxic conditions, the subsequent upregulation of HIF1α target genes may ultimately promote cancer cell invasion [113].
Another example is that while NFκB is generally considered pro-inflammatory, it also acts to regulate cellular stress resistance pathways and its upregulation in response to ROS formation may aid cellular apoptosis resistance during HSR [38, 40]. Thus, while inhibition of inflammation by SIRT1 may be helpful in the setting of chronic stresses or aging, excessive stimulation of SIRT1 during an acute injury has the potential to interfere with managing damage responses and metabolic shifts under conditions of hypoperfusion.
Treatment with resveratrol may present additional risks through SIRT1-independent pathways. The inhibition of cyclooxygenase by resveratrol causes decreased production of thromboxane, which is a potent facilitator of platelet aggregation and vasoconstriction. Its inhibition may be detrimental due to decreased activation of the clotting cascade, potentially promoting bleeding. In the emergency department, it is estimated that nearly 30% of trauma patients have impaired clotting [114]. Multiple studies have evidenced a prolonged clotting time in washed or partially enriched platelet fractions, and a marked reduction in collagen-induced clotting was discovered in the blood from subjects who drank grape juice for 1 week [84, 115, 116]. In contrast, the antiplatelet activity of resveratrol appears to be weakened or masked in circulating whole blood, with concentrations as high as 100 μm having no effect on platelet aggregation [117]. Even after multiple weeks of resveratrol administration in rats, no change in clotting time was observed [118]; hence, the clinical relevance of these findings remains partially unknown. Thus, resuscitation with resveratrol could theoretically be harmful by increasing blood loss in susceptible patients, although the risk remains uncertain.
In addition, there is some evidence that animals treated with resveratrol following HSR had decreased insulin resistance and lower blood glucose levels [66]. While the increase in insulin sensitivity is most likely beneficial, clinician awareness of possible hypoglycemic episodes during treatment will be imperative.
Finally, it is important to note that the effects of SIRT1 in the setting of septic shock have been shown to depend on timing. It has been suggested that SIRT1 activity is advantageous in the early stages of shock, but may be harmful at later stages, due to the body’s transition from a pro-inflammatory to a hypo-inflammatory response state. A significant prolongation in survival was demonstrated in a cecal ligation-puncture model through the inhibition of SIRT1 beginning 24 h after the onset of sepsis [119]. Although SIRT1 activation has proven to be largely beneficial, these findings delineate the necessity to further evaluate delivery timings of SIRT1 activators during the setting of hemorrhagic shock. Similarly, CR has not been universally beneficial. CR has been reported to increase lipid peroxidation and shorten the lifespan in a model of amyotrophic lateral sclerosis [120], and conflicting results have been reported on the effect of CR in sepsis [17, 121]. Thus, some caution is warranted when generalizing the effects of longevity interventions on the responses to specific stresses.
Conclusion
Genes and other factors that confer increased longevity often confer resistance not only to chronic diseases but also to acute stresses. Thus, such factors may represent a largely untapped source of new therapeutic approaches to treat the pathophysiology of acute stresses such as trauma and hemorrhagic shock. Both resveratrol and NAD precursors have been shown to decrease organ damage and improve survival in hemorrhagic shock and interestingly, both converge on activation of SIRT1, the mammalian homolog of a protein that influences longevity in lower organisms. In fact, there is evidence that a combination therapy including both resveratrol and NAD precursors may be more effective than either alone [122]. While there is little direct genetic evidence on the effects of SIRT1 in hemorrhagic shock due to the inherent difficulties in creating a mouse model of hemorrhage, SIRT1 has been shown in other systems to work through multiple mechanisms to inhibit inflammation, oxidative stress, mitochondrial dysfunction, vascular endothelial dysfunction, and coagulopathy. The potential benefit of SIRT1 in mitigating hemorrhagic shock offers a new therapeutic avenue and supports the continued focus on longevity pathways as a source of interventions that can mitigate acute stresses.
Funding
J.A.B. is supported by a grant from the National Institute of Diabetes and Digestive and Kidney Diseases (DK 098656).
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Carrie A. Sims, Email: Carrie.Sims@osumc.edu
Joseph A. Baur, Email: baur@mail.med.upenn.edu
References
- 1.Ismail K, Nussbaum L, Sebastiani P, Andersen S, Perls T, Barzilai N, Milman S. Compression of morbidity is observed across cohorts with exceptional longevity. J Am Geriatr Soc. 2016;64(8):1583–1591. doi: 10.1111/jgs.14222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Koene RJ, Prizment AE, Blaes A, Konety SH. Shared risk factors in cardiovascular disease and cancer. Circulation. 2016;133(11):1104–1114. doi: 10.1161/CIRCULATIONAHA.115.020406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Anstey KJ, Kingston A, Kiely KM, Luszcz MA, Mitchell P, Jagger C. The influence of smoking, sedentary lifestyle and obesity on cognitive impairment-free life expectancy. Int J Epidemiol. 2014;43(6):1874–1883. doi: 10.1093/ije/dyu170. [DOI] [PubMed] [Google Scholar]
- 4.Renehan AG, Tyson M, Egger M, Heller RF, Zwahlen M. Body-mass index and incidence of cancer: a systematic review and meta-analysis of prospective observational studies. Lancet. 2008;371(9612):569–578. doi: 10.1016/S0140-6736(08)60269-X. [DOI] [PubMed] [Google Scholar]
- 5.Rogha M, et al. Cigarette smoking effect on human cochlea responses. Adv Biomed Res. 2015;4:148. doi: 10.4103/2277-9175.161575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Fernandes TMP, Almeida NL, Santos NAD. Effects of smoking and smoking abstinence on spatial vision in chronic heavy smokers. Sci Rep. 2017;7(1):1690. doi: 10.1038/s41598-017-01877-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kovac JR, Khanna A, Lipshultz LI. The effects of cigarette smoking on male fertility. Postgrad Med. 2015;127(3):338–341. doi: 10.1080/00325481.2015.1015928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Merritt T, Mazela J, Merritt A. Tobacco smoking and its consequences on reproductive health: the impact of a lifestyle choices including cigarette smoke exposure on fertility and birth defects. Przegl Lek. 2013;70(10):779–783. [PubMed] [Google Scholar]
- 9.Kramer A. An overview of the beneficial effects of exercise on health and performance. Adv Exp Med Biol. 2020;1228:3–22. doi: 10.1007/978-981-15-1792-1_1. [DOI] [PubMed] [Google Scholar]
- 10.Marshall CH, al-Mallah MH, Dardari Z, Brawner CA, Lamerato LE, Keteyian SJ, Ehrman JK, Visvanathan K, Blaha MJ. Cardiorespiratory fitness and incident lung and colorectal cancer in men and women: results from the Henry Ford Exercise Testing (FIT) cohort. Cancer. 2019;125(15):2594–2601. doi: 10.1002/cncr.32085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mattison JA, Colman RJ, Beasley TM, Allison DB, Kemnitz JW, Roth GS, Ingram DK, Weindruch R, de Cabo R, Anderson RM. Caloric restriction improves health and survival of rhesus monkeys. Nat Commun. 2017;8:14063. doi: 10.1038/ncomms14063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Anderson RM, Weindruch R. The caloric restriction paradigm: implications for healthy human aging. Am J Hum Biol. 2012;24(2):101–106. doi: 10.1002/ajhb.22243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Menezes-Filho SL, Amigo I, Prado FM, Ferreira NC, Koike MK, Pinto IFD, Miyamoto S, Montero EFS, Medeiros MHG, Kowaltowski AJ. Caloric restriction protects livers from ischemia/reperfusion damage by preventing Ca(2+)-induced mitochondrial permeability transition. Free Radic Biol Med. 2017;110:219–227. doi: 10.1016/j.freeradbiomed.2017.06.013. [DOI] [PubMed] [Google Scholar]
- 14.Zhang J, Zhang W, Gao X, Zhao Y, Chen D, Xu N, Pu H, Stetler RA, Gao Y. Preconditioning with partial caloric restriction confers long-term protection against grey and white matter injury after transient focal ischemia. J Cereb Blood Flow Metab. 2019;39(7):1394–1409. doi: 10.1177/0271678X18785480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gross L, Dreyfuss Y. Prevention of spontaneous and radiation-induced tumors in rats by reduction of food intake. Proc Natl Acad Sci U S A. 1990;87(17):6795–6797. doi: 10.1073/pnas.87.17.6795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Harper JM, Salmon AB, Chang Y, Bonkowski M, Bartke A, Miller RA. Stress resistance and aging: influence of genes and nutrition. Mech Ageing Dev. 2006;127(8):687–694. doi: 10.1016/j.mad.2006.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sun D, Muthukumar AR, Lawrence RA, Fernandes G. Effects of calorie restriction on polymicrobial peritonitis induced by cecum ligation and puncture in young C57BL/6 mice. Clin Diagn Lab Immunol. 2001;8(5):1003–1011. doi: 10.1128/CDLI.8.5.1003-1011.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Brandhorst S, Harputlugil E, Mitchell JR, Longo VD. Protective effects of short-term dietary restriction in surgical stress and chemotherapy. Ageing Res Rev. 2017;39:68–77. doi: 10.1016/j.arr.2017.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Most J, Gilmore LA, Smith SR, Han H, Ravussin E, Redman LM. Significant improvement in cardiometabolic health in healthy nonobese individuals during caloric restriction-induced weight loss and weight loss maintenance. Am J Physiol Endocrinol Metab. 2018;314(4):E396–E405. doi: 10.1152/ajpendo.00261.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kraus WE, Bhapkar M, Huffman KM, Pieper CF, Krupa Das S, Redman LM, Villareal DT, Rochon J, Roberts SB, Ravussin E, Holloszy JO, Fontana L, CALERIE Investigators 2 years of calorie restriction and cardiometabolic risk (CALERIE): exploratory outcomes of a multicentre, phase 2, randomised controlled trial. Lancet Diabetes Endocrinol. 2019;7(9):673–683. doi: 10.1016/S2213-8587(19)30151-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Grigolon RB, Brietzke E, Trevizol AP, McIntyre RS, Mansur RB. Caloric restriction, resting metabolic rate and cognitive performance in non-obese adults: a post-hoc analysis from CALERIE study. J Psychiatr Res. 2020;128:16–22. doi: 10.1016/j.jpsychires.2020.05.018. [DOI] [PubMed] [Google Scholar]
- 22.Kennedy BK, Austriaco NR, Jr, Zhang J, Guarente L. Mutation in the silencing gene SIR4 can delay aging in S. cerevisiae. Cell. 1995;80(3):485–496. doi: 10.1016/0092-8674(95)90499-9. [DOI] [PubMed] [Google Scholar]
- 23.Tissenbaum HA, Guarente L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature. 2001;410(6825):227–230. doi: 10.1038/35065638. [DOI] [PubMed] [Google Scholar]
- 24.Kaeberlein M, McVey M, Guarente L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 1999;13(19):2570–2580. doi: 10.1101/gad.13.19.2570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rogina B, Helfand SL. Sir2 mediates longevity in the fly through a pathway related to calorie restriction. Proc Natl Acad Sci U S A. 2004;101(45):15998–16003. doi: 10.1073/pnas.0404184101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Whitaker R, Faulkner S, Miyokawa R, Burhenn L, Henriksen M, Wood JG, Helfand SL. Increased expression of Drosophila Sir2 extends life span in a dose-dependent manner. Aging (Albany NY) 2013;5(9):682–691. doi: 10.18632/aging.100599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Burnett C, Valentini S, Cabreiro F, Goss M, Somogyvári M, Piper MD, Hoddinott M, Sutphin GL, Leko V, McElwee JJ, Vazquez-Manrique RP, Orfila AM, Ackerman D, Au C, Vinti G, Riesen M, Howard K, Neri C, Bedalov A, Kaeberlein M, Sőti C, Partridge L, Gems D. Absence of effects of Sir2 overexpression on lifespan in C. elegans and Drosophila. Nature. 2011;477(7365):482–485. doi: 10.1038/nature10296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Viswanathan M, Guarente L. Regulation of Caenorhabditis elegans lifespan by sir-2.1 transgenes. Nature. 2011;477(7365):E1–E2. doi: 10.1038/nature10440. [DOI] [PubMed] [Google Scholar]
- 29.Chalkiadaki A, Guarente L. Sirtuins mediate mammalian metabolic responses to nutrient availability. Nat Rev Endocrinol. 2012;8(5):287–296. doi: 10.1038/nrendo.2011.225. [DOI] [PubMed] [Google Scholar]
- 30.Chen D, Steele AD, Lindquist S, Guarente L. Increase in activity during calorie restriction requires Sirt1. Science. 2005;310(5754):1641. doi: 10.1126/science.1118357. [DOI] [PubMed] [Google Scholar]
- 31.Someya S, Yu W, Hallows WC, Xu J, Vann JM, Leeuwenburgh C, Tanokura M, Denu JM, Prolla TA. Sirt3 mediates reduction of oxidative damage and prevention of age-related hearing loss under caloric restriction. Cell. 2010;143(5):802–812. doi: 10.1016/j.cell.2010.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Imai SI, Guarente L. It takes two to tango: NAD(+) and sirtuins in aging/longevity control. NPJ Aging Mech Dis. 2016;2:16017. doi: 10.1038/npjamd.2016.17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Vanzant EL, et al. Advanced age is associated with worsened outcomes and a unique genomic response in severely injured patients with hemorrhagic shock. Crit Care. 2015;19:77. doi: 10.1186/s13054-015-0788-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cannon J, et al. Resuscitative endovascular balloon occlusion of the aorta (REBOA) for hemorrhagic shock. Mil Med. 2018;183(suppl_2):55–59. doi: 10.1093/milmed/usy143. [DOI] [PubMed] [Google Scholar]
- 35.Chaudry IH. Cellular mechanisms in shock and ischemia and their correction. Am J Phys. 1983;245(2):R117–R134. doi: 10.1152/ajpregu.1983.245.2.R117. [DOI] [PubMed] [Google Scholar]
- 36.Peitzman AB, Harbrecht BG, Udekwu AO, Billiar TR, Kelly E, Simmons RL. Hemorrhagic shock. Curr Probl Surg. 1995;32(11):925–1002. doi: 10.1016/s0011-3840(05)80008-5. [DOI] [PubMed] [Google Scholar]
- 37.Sauaia A, Moore FA, Moore EE, Moser KS, Brennan R, Read RA, Pons PT. Epidemiology of trauma deaths: a reassessment. J Trauma. 1995;38(2):185–193. doi: 10.1097/00005373-199502000-00006. [DOI] [PubMed] [Google Scholar]
- 38.Shenkar R, et al. Hemorrhage activates NF-kappa B in murine lung mononuclear cells in vivo. Am J Phys. 1996;270(5 Pt 1):L729–L735. doi: 10.1152/ajplung.1996.270.5.L729. [DOI] [PubMed] [Google Scholar]
- 39.Yeung F, Hoberg JE, Ramsey CS, Keller MD, Jones DR, Frye RA, Mayo MW. Modulation of NF-kappaB-dependent transcription and cell survival by the SIRT1 deacetylase. EMBO J. 2004;23(12):2369–2380. doi: 10.1038/sj.emboj.7600244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Pahl HL. Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene. 1999;18(49):6853–6866. doi: 10.1038/sj.onc.1203239. [DOI] [PubMed] [Google Scholar]
- 41.Breitenstein A, Stein S, Holy EW, Camici GG, Lohmann C, Akhmedov A, Spescha R, Elliott PJ, Westphal CH, Matter CM, Lüscher TF, Tanner FC. Sirt1 inhibition promotes in vivo arterial thrombosis and tissue factor expression in stimulated cells. Cardiovasc Res. 2011;89(2):464–472. doi: 10.1093/cvr/cvq339. [DOI] [PubMed] [Google Scholar]
- 42.Liu FC, Tsai HI, Yu HP. Organ-protective effects of red wine extract, resveratrol, in oxidative stress-mediated reperfusion injury. Oxidative Med Cell Longev. 2015;2015:568634. doi: 10.1155/2015/568634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Guo C, Sun L, Chen X, Zhang D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen Res. 2013;8(21):2003–2014. doi: 10.3969/j.issn.1673-5374.2013.21.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Fink MP, Macias CA, Xiao J, Tyurina YY, Delude RL, Greenberger JS, Kagan VE, Wipf P. Hemigramicidin-TEMPO conjugates: novel mitochondria-targeted antioxidants. Crit Care Med. 2007;35(9 Suppl):S461–S467. doi: 10.1097/01.CCM.0000279192.96303.E7. [DOI] [PubMed] [Google Scholar]
- 45.Powell RD, Goodenow DA, Mixer HV, Mckillop IH, Evans SL. Cytochrome c limits oxidative stress and decreases acidosis in a rat model of hemorrhagic shock and reperfusion injury. J Trauma Acute Care Surg. 2017;82(1):35–41. doi: 10.1097/TA.0000000000001257. [DOI] [PubMed] [Google Scholar]
- 46.Bowler RP, et al. Extracellular superoxide dismutase attenuates lung injury after hemorrhage. Am J Respir Crit Care Med. 2001;164(2):290–294. doi: 10.1164/ajrccm.164.2.2011054. [DOI] [PubMed] [Google Scholar]
- 47.Kops GJ, et al. Forkhead transcription factor FOXO3a protects quiescent cells from oxidative stress. Nature. 2002;419(6904):316–321. doi: 10.1038/nature01036. [DOI] [PubMed] [Google Scholar]
- 48.Rodgers JT, Lerin C, Haas W, Gygi SP, Spiegelman BM, Puigserver P. Nutrient control of glucose homeostasis through a complex of PGC-1alpha and SIRT1. Nature. 2005;434(7029):113–118. doi: 10.1038/nature03354. [DOI] [PubMed] [Google Scholar]
- 49.St-Pierre J, Drori S, Uldry M, Silvaggi JM, Rhee J, Jäger S, Handschin C, Zheng K, Lin J, Yang W, Simon DK, Bachoo R, Spiegelman BM. Suppression of reactive oxygen species and neurodegeneration by the PGC-1 transcriptional coactivators. Cell. 2006;127(2):397–408. doi: 10.1016/j.cell.2006.09.024. [DOI] [PubMed] [Google Scholar]
- 50.Biel TG, Lee S, Flores-Toro JA, Dean JW, Go KL, Lee MH, Law BK, Law ME, Dunn WA, Jr, Zendejas I, Behrns KE, Kim JS. Sirtuin 1 suppresses mitochondrial dysfunction of ischemic mouse livers in a mitofusin 2-dependent manner. Cell Death Differ. 2016;23(2):279–290. doi: 10.1038/cdd.2015.96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Hsu CP, Zhai P, Yamamoto T, Maejima Y, Matsushima S, Hariharan N, Shao D, Takagi H, Oka S, Sadoshima J. Silent information regulator 1 protects the heart from ischemia/reperfusion. Circulation. 2010;122(21):2170–2182. doi: 10.1161/CIRCULATIONAHA.110.958033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wang J, Guo M, Ma R, Wu M, Zhang Y. Tetrandrine alleviates cerebral ischemia/reperfusion injury by suppressing NLRP3 inflammasome activation via Sirt-1. PeerJ. 2020;8:e9042. doi: 10.7717/peerj.9042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Howitz KT, Bitterman KJ, Cohen HY, Lamming DW, Lavu S, Wood JG, Zipkin RE, Chung P, Kisielewski A, Zhang LL, Scherer B, Sinclair DA. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature. 2003;425(6954):191–196. doi: 10.1038/nature01960. [DOI] [PubMed] [Google Scholar]
- 54.Hubbard BP, Gomes AP, Dai H, Li J, Case AW, Considine T, Riera TV, Lee JE, E SY, Lamming DW, Pentelute BL, Schuman ER, Stevens LA, Ling AJY, Armour SM, Michan S, Zhao H, Jiang Y, Sweitzer SM, Blum CA, Disch JS, Ng PY, Howitz KT, Rolo AP, Hamuro Y, Moss J, Perni RB, Ellis JL, Vlasuk GP, Sinclair DA. Evidence for a common mechanism of SIRT1 regulation by allosteric activators. Science. 2013;339(6124):1216–1219. doi: 10.1126/science.1231097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Zang M, Xu S, Maitland-Toolan KA, Zuccollo A, Hou X, Jiang B, Wierzbicki M, Verbeuren TJ, Cohen RA. Polyphenols stimulate AMP-activated protein kinase, lower lipids, and inhibit accelerated atherosclerosis in diabetic LDL receptor-deficient mice. Diabetes. 2006;55(8):2180–2191. doi: 10.2337/db05-1188. [DOI] [PubMed] [Google Scholar]
- 56.Baur JA, Pearson KJ, Price NL, Jamieson HA, Lerin C, Kalra A, Prabhu VV, Allard JS, Lopez-Lluch G, Lewis K, Pistell PJ, Poosala S, Becker KG, Boss O, Gwinn D, Wang M, Ramaswamy S, Fishbein KW, Spencer RG, Lakatta EG, le Couteur D, Shaw RJ, Navas P, Puigserver P, Ingram DK, de Cabo R, Sinclair DA. Resveratrol improves health and survival of mice on a high-calorie diet. Nature. 2006;444(7117):337–342. doi: 10.1038/nature05354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Fulco M, Cen Y, Zhao P, Hoffman EP, McBurney MW, Sauve AA, Sartorelli V. Glucose restriction inhibits skeletal myoblast differentiation by activating SIRT1 through AMPK-mediated regulation of Nampt. Dev Cell. 2008;14(5):661–673. doi: 10.1016/j.devcel.2008.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Price NL, Gomes AP, Ling AJY, Duarte FV, Martin-Montalvo A, North BJ, Agarwal B, Ye L, Ramadori G, Teodoro JS, Hubbard BP, Varela AT, Davis JG, Varamini B, Hafner A, Moaddel R, Rolo AP, Coppari R, Palmeira CM, de Cabo R, Baur JA, Sinclair DA. SIRT1 is required for AMPK activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab. 2012;15(5):675–690. doi: 10.1016/j.cmet.2012.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Park SJ, Ahmad F, Philp A, Baar K, Williams T, Luo H, Ke H, Rehmann H, Taussig R, Brown AL, Kim MK, Beaven MA, Burgin AB, Manganiello V, Chung JH. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell. 2012;148(3):421–433. doi: 10.1016/j.cell.2012.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Toiber D, Sebastian C, Mostoslavsky R. Characterization of nuclear sirtuins: molecular mechanisms and physiological relevance. Handb Exp Pharmacol. 2011;206:189–224. doi: 10.1007/978-3-642-21631-2_9. [DOI] [PubMed] [Google Scholar]
- 61.Sims CA, Baur JA. The grapes and wrath: using resveratrol to treat the pathophysiology of hemorrhagic shock. Ann N Y Acad Sci. 2017;1403(1):70–81. doi: 10.1111/nyas.13444. [DOI] [PubMed] [Google Scholar]
- 62.Baur JA, Sinclair DA. Therapeutic potential of resveratrol: the in vivo evidence. Nat Rev Drug Discov. 2006;5(6):493–506. doi: 10.1038/nrd2060. [DOI] [PubMed] [Google Scholar]
- 63.Ayub A, Poulose N, Raju R. Resveratrol improves survival and prolongs life following hemorrhagic shock. Mol Med. 2015;21:305–312. doi: 10.2119/molmed.2015.00013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Jian B, Yang S, Chaudry IH, Raju R. Resveratrol improves cardiac contractility following trauma-hemorrhage by modulating Sirt1. Mol Med. 2012;18:209–214. doi: 10.2119/molmed.2011.00365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Yu HP, Hsu JC, Hwang TL, Yen CH, Lau YT. Resveratrol attenuates hepatic injury after trauma-hemorrhage via estrogen receptor-related pathway. Shock. 2008;30(3):324–328. doi: 10.1097/SHK.0b013e318164f013. [DOI] [PubMed] [Google Scholar]
- 66.Wang H, Guan Y, Widlund AL, Becker LB, Baur JA, Reilly PM, Sims CA. Resveratrol ameliorates mitochondrial dysfunction but increases the risk of hypoglycemia following hemorrhagic shock. J Trauma Acute Care Surg. 2014;77(6):926–933. doi: 10.1097/TA.0000000000000358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Wang H, et al. Resveratrol rescues kidney mitochondrial function following hemorrhagic shock. Shock. 2015;44(2):173–180. doi: 10.1097/SHK.0000000000000390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Yu HP, Hwang TL, Hsieh PW, Lau YT. Role of estrogen receptor-dependent upregulation of P38 MAPK/heme oxygenase 1 in resveratrol-mediated attenuation of intestinal injury after trauma-hemorrhage. Shock. 2011;35(5):517–523. doi: 10.1097/SHK.0b013e318209e931. [DOI] [PubMed] [Google Scholar]
- 69.Yu HP, Yang SC, Lau YT, Hwang TL. Role of Akt-dependent up-regulation of hemeoxygenase-1 in resveratrol-mediated attenuation of hepatic injury after trauma hemorrhage. Surgery. 2010;148(1):103–109. doi: 10.1016/j.surg.2009.12.008. [DOI] [PubMed] [Google Scholar]
- 70.Yu HP, Hwang TL, Hwang TL, Yen CH, Lau YT. Resveratrol prevents endothelial dysfunction and aortic superoxide production after trauma hemorrhage through estrogen receptor-dependent hemeoxygenase-1 pathway. Crit Care Med. 2010;38(4):1147–1154. doi: 10.1097/CCM.0b013e3181cd124e. [DOI] [PubMed] [Google Scholar]
- 71.Halliwell B. Dietary polyphenols: good, bad, or indifferent for your health? Cardiovasc Res. 2007;73(2):341–347. doi: 10.1016/j.cardiores.2006.10.004. [DOI] [PubMed] [Google Scholar]
- 72.Lim JH, Lee YM, Chun YS, Chen J, Kim JE, Park JW. Sirtuin 1 modulates cellular responses to hypoxia by deacetylating hypoxia-inducible factor 1alpha. Mol Cell. 2010;38(6):864–878. doi: 10.1016/j.molcel.2010.05.023. [DOI] [PubMed] [Google Scholar]
- 73.Jian B, Yang S, Chaudry IH, Raju R. Resveratrol restores sirtuin 1 (SIRT1) activity and pyruvate dehydrogenase kinase 1 (PDK1) expression after hemorrhagic injury in a rat model. Mol Med. 2014;20:10–16. doi: 10.2119/molmed.2013.00077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Poolman TM, et al. Inhibition of the respiratory burst by resveratrol in human monocytes: correlation with inhibition of PI3K signaling. Free Radic Biol Med. 2005;39(1):118–132. doi: 10.1016/j.freeradbiomed.2005.02.036. [DOI] [PubMed] [Google Scholar]
- 75.Li P, Wang X, Zhao M, Song R, Zhao KS. Polydatin protects hepatocytes against mitochondrial injury in acute severe hemorrhagic shock via SIRT1-SOD2 pathway. Expert Opin Ther Targets. 2015;19(7):997–1010. doi: 10.1517/14728222.2015.1054806. [DOI] [PubMed] [Google Scholar]
- 76.Scarpulla RC. Metabolic control of mitochondrial biogenesis through the PGC-1 family regulatory network. Biochim Biophys Acta. 2011;1813(7):1269–1278. doi: 10.1016/j.bbamcr.2010.09.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Li P, Meng X, Bian H, Burns N, Zhao KS, Song R. Activation of sirtuin 1/3 improves vascular hyporeactivity in severe hemorrhagic shock by alleviation of mitochondrial damage. Oncotarget. 2015;6(35):36998–37011. doi: 10.18632/oncotarget.6076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Wang X, Song R, Bian HN, Brunk UT, Zhao M, Zhao KS. Polydatin, a natural polyphenol, protects arterial smooth muscle cells against mitochondrial dysfunction and lysosomal destabilization following hemorrhagic shock. Am J Physiol Regul Integr Comp Physiol. 2012;302(7):R805–R814. doi: 10.1152/ajpregu.00350.2011. [DOI] [PubMed] [Google Scholar]
- 79.Wang X, Song R, Chen Y, Zhao M, Zhao KS. Polydatin—a new mitochondria protector for acute severe hemorrhagic shock treatment. Expert Opin Investig Drugs. 2013;22(2):169–179. doi: 10.1517/13543784.2013.748033. [DOI] [PubMed] [Google Scholar]
- 80.Zeng Z, et al. Polydatin protecting kidneys against hemorrhagic shock-induced mitochondrial dysfunction via SIRT1 activation and p53 deacetylation. Oxidative Med Cell Longev. 2016;2016:1737185. doi: 10.1155/2016/1737185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Zeng Z, et al. Polydatin alleviates small intestine injury during hemorrhagic shock as a SIRT1 activator. Oxidative Med Cell Longev. 2015;2015:965961. doi: 10.1155/2015/965961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Xu S, Zeng Z, Zhao M, Huang Q, Gao Y, Dai X, Lu J, Huang W, Zhao K. Evidence for SIRT1 mediated HMGB1 release from kidney cells in the early stages of hemorrhagic shock. Front Physiol. 2019;10:854. doi: 10.3389/fphys.2019.00854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Chow SE, et al. Resveratrol attenuates oxLDL-stimulated NADPH oxidase activity and protects endothelial cells from oxidative functional damages. J Appl Physiol (1985) 2007;102(4):1520–1527. doi: 10.1152/japplphysiol.00881.2006. [DOI] [PubMed] [Google Scholar]
- 84.Olas B, Wachowicz B, Saluk-Juszczak J, Zieliński T. Effect of resveratrol, a natural polyphenolic compound, on platelet activation induced by endotoxin or thrombin. Thromb Res. 2002;107(3–4):141–145. doi: 10.1016/s0049-3848(02)00273-6. [DOI] [PubMed] [Google Scholar]
- 85.Powell RD, Swet JH, Kennedy KL, Huynh TT, Mckillop IH, Evans SL. Resveratrol attenuates hypoxic injury in a primary hepatocyte model of hemorrhagic shock and resuscitation. J Trauma Acute Care Surg. 2014;76(2):409–417. doi: 10.1097/TA.0000000000000096. [DOI] [PubMed] [Google Scholar]
- 86.Wu CT, Yu HP, Chung CY, Lau YT, Liao SK. Attenuation of lung inflammation and pro-inflammatory cytokine production by resveratrol following trauma-hemorrhage. Chin J Physiol. 2008;51(6):363–368. [PubMed] [Google Scholar]
- 87.Tsai YF, Liu FC, Lau YT, Yu HP. Role of Akt-dependent pathway in resveratrol-mediated cardioprotection after trauma-hemorrhage. J Surg Res. 2012;176(1):171–177. doi: 10.1016/j.jss.2011.05.039. [DOI] [PubMed] [Google Scholar]
- 88.Canto C, Menzies KJ, Auwerx J. NAD(+) metabolism and the control of energy homeostasis: a balancing act between mitochondria and the nucleus. Cell Metab. 2015;22(1):31–53. doi: 10.1016/j.cmet.2015.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Zhang H, Ryu D, Wu Y, Gariani K, Wang X, Luan P, DAmico D, Ropelle ER, Lutolf MP, Aebersold R, Schoonjans K, Menzies KJ, Auwerx J. NAD(+) repletion improves mitochondrial and stem cell function and enhances life span in mice. Science. 2016;352(6292):1436–1443. doi: 10.1126/science.aaf2693. [DOI] [PubMed] [Google Scholar]
- 90.Yoshino J, Baur JA, Imai SI. NAD(+) intermediates: the biology and therapeutic potential of NMN and NR. Cell Metab. 2018;27(3):513–528. doi: 10.1016/j.cmet.2017.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Mills KF, Yoshida S, Stein LR, Grozio A, Kubota S, Sasaki Y, Redpath P, Migaud ME, Apte RS, Uchida K, Yoshino J, Imai SI. Long-term administration of nicotinamide mononucleotide mitigates age-associated physiological decline in mice. Cell Metab. 2016;24(6):795–806. doi: 10.1016/j.cmet.2016.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Revollo JR, Grimm AA, Imai S. The NAD biosynthesis pathway mediated by nicotinamide phosphoribosyltransferase regulates Sir2 activity in mammalian cells. J Biol Chem. 2004;279(49):50754–50763. doi: 10.1074/jbc.M408388200. [DOI] [PubMed] [Google Scholar]
- 93.Berger F, Lau C, Dahlmann M, Ziegler M. Subcellular compartmentation and differential catalytic properties of the three human nicotinamide mononucleotide adenylyltransferase isoforms. J Biol Chem. 2005;280(43):36334–36341. doi: 10.1074/jbc.M508660200. [DOI] [PubMed] [Google Scholar]
- 94.van Wijk SJ, Hageman GJ. Poly(ADP-ribose) polymerase-1 mediated caspase-independent cell death after ischemia/reperfusion. Free Radic Biol Med. 2005;39(1):81–90. doi: 10.1016/j.freeradbiomed.2005.03.021. [DOI] [PubMed] [Google Scholar]
- 95.Alano CC, Garnier P, Ying W, Higashi Y, Kauppinen TM, Swanson RA. NAD+ depletion is necessary and sufficient for poly(ADP-ribose) polymerase-1-mediated neuronal death. J Neurosci. 2010;30(8):2967–2978. doi: 10.1523/JNEUROSCI.5552-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Canto C, et al. The NAD(+) precursor nicotinamide riboside enhances oxidative metabolism and protects against high-fat diet-induced obesity. Cell Metab. 2012;15(6):838–847. doi: 10.1016/j.cmet.2012.04.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Gomes AP, Price NL, Ling AJY, Moslehi JJ, Montgomery MK, Rajman L, White JP, Teodoro JS, Wrann CD, Hubbard BP, Mercken EM, Palmeira CM, de Cabo R, Rolo AP, Turner N, Bell EL, Sinclair DA. Declining NAD(+) induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. Cell. 2013;155(7):1624–1638. doi: 10.1016/j.cell.2013.11.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Gero D, Szabo C. Salvage of nicotinamide adenine dinucleotide plays a critical role in the bioenergetic recovery of post-hypoxic cardiomyocytes. Br J Pharmacol. 2015;172(20):4817–4832. doi: 10.1111/bph.13252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Wurth MA, Sayeed MM, Baue AE. Nicotinamide adenine dinucleotide (NAD) content of liver with hemorrhagic shock. Proc Soc Exp Biol Med. 1973;144(2):654–658. doi: 10.3181/00379727-144-37656. [DOI] [PubMed] [Google Scholar]
- 100.Chaudry IH, Zweig S, Sayeed MM, Baue AE. Failure of nicotinamide in the treatment of hemorrhagic shock. J Surg Res. 1976;21(1):27–32. doi: 10.1016/0022-4804(76)90006-8. [DOI] [PubMed] [Google Scholar]
- 101.Jeong KY, Suh GJ, Kwon WY, Kim KS, Jung YS, Kye YC. The therapeutic effect and mechanism of niacin on acute lung injury in a rat model of hemorrhagic shock: down-regulation of the reactive oxygen species-dependent nuclear factor kappaB pathway. J Trauma Acute Care Surg. 2015;79(2):247–255. doi: 10.1097/TA.0000000000000761. [DOI] [PubMed] [Google Scholar]
- 102.Szabo C. Poly (ADP-ribose) polymerase activation and circulatory shock. Novartis Found Symp. 2007;280:92–103. doi: 10.1007/0-387-36005-0_16. [DOI] [PubMed] [Google Scholar]
- 103.Yoshino J, Mills KF, Yoon MJ, Imai SI. Nicotinamide mononucleotide, a key NAD(+) intermediate, treats the pathophysiology of diet- and age-induced diabetes in mice. Cell Metab. 2011;14(4):528–536. doi: 10.1016/j.cmet.2011.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Hsu CP, Oka S, Shao D, Hariharan N, Sadoshima J. Nicotinamide phosphoribosyltransferase regulates cell survival through NAD+ synthesis in cardiac myocytes. Circ Res. 2009;105(5):481–491. doi: 10.1161/CIRCRESAHA.109.203703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Park JH, Long A, Owens K, Kristian T. Nicotinamide mononucleotide inhibits post-ischemic NAD(+) degradation and dramatically ameliorates brain damage following global cerebral ischemia. Neurobiol Dis. 2016;95:102–110. doi: 10.1016/j.nbd.2016.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Yamamoto T, Byun J, Zhai P, Ikeda Y, Oka S, Sadoshima J. Nicotinamide mononucleotide, an intermediate of NAD+ synthesis, protects the heart from ischemia and reperfusion. PLoS One. 2014;9(6):e98972. doi: 10.1371/journal.pone.0098972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Wang P, Xu TY, Guan YF, Tian WW, Viollet B, Rui YC, Zhai QW, Su DF, Miao CY. Nicotinamide phosphoribosyltransferase protects against ischemic stroke through SIRT1-dependent adenosine monophosphate-activated kinase pathway. Ann Neurol. 2011;69(2):360–374. doi: 10.1002/ana.22236. [DOI] [PubMed] [Google Scholar]
- 108.Wei CC, Kong YY, Li GQ, Guan YF, Wang P, Miao CY. Nicotinamide mononucleotide attenuates brain injury after intracerebral hemorrhage by activating Nrf2/HO-1 signaling pathway. Sci Rep. 2017;7(1):717. doi: 10.1038/s41598-017-00851-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Sims CA, et al. Nicotinamide mononucleotide preserves mitochondrial function and increases survival in hemorrhagic shock. JCI Insight. 2018;3(17):e120182. doi: 10.1172/jci.insight.120182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Subramani K, Chu X, Warren M, Lee M, Lu S, Singh N, Raju R. Deficiency of metabolite sensing receptor HCA2 impairs the salutary effect of niacin in hemorrhagic shock. Biochim Biophys Acta Mol basis Dis. 2019;1865(3):688–695. doi: 10.1016/j.bbadis.2019.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Dioum EM, Chen R, Alexander MS, Zhang Q, Hogg RT, Gerard RD, Garcia JA. Regulation of hypoxia-inducible factor 2alpha signaling by the stress-responsive deacetylase sirtuin 1. Science. 2009;324(5932):1289–1293. doi: 10.1126/science.1169956. [DOI] [PubMed] [Google Scholar]
- 112.DeBerardinis RJ, Lum JJ, Hatzivassiliou G, Thompson CB. The biology of cancer: metabolic reprogramming fuels cell growth and proliferation. Cell Metab. 2008;7(1):11–20. doi: 10.1016/j.cmet.2007.10.002. [DOI] [PubMed] [Google Scholar]
- 113.Joo HY, Yun M, Jeong J, Park ER, Shin HJ, Woo SR, Jung JK, Kim YM, Park JJ, Kim J, Lee KH. SIRT1 deacetylates and stabilizes hypoxia-inducible factor-1alpha (HIF-1alpha) via direct interactions during hypoxia. Biochem Biophys Res Commun. 2015;462(4):294–300. doi: 10.1016/j.bbrc.2015.04.119. [DOI] [PubMed] [Google Scholar]
- 114.MacLeod JB, et al. Early coagulopathy predicts mortality in trauma. J Trauma. 2003;55(1):39–44. doi: 10.1097/01.TA.0000075338.21177.EF. [DOI] [PubMed] [Google Scholar]
- 115.Keevil JG, Osman HE, Reed JD, Folts JD. Grape juice, but not orange juice or grapefruit juice, inhibits human platelet aggregation. J Nutr. 2000;130(1):53–56. doi: 10.1093/jn/130.1.53. [DOI] [PubMed] [Google Scholar]
- 116.Stef G, Csiszar A, Lerea K, Ungvari Z, Veress G. Resveratrol inhibits aggregation of platelets from high-risk cardiac patients with aspirin resistance. J Cardiovasc Pharmacol. 2006;48(2):1–5. doi: 10.1097/01.fjc.0000238592.67191.ab. [DOI] [PubMed] [Google Scholar]
- 117.Kirk RI, Deitch JA, Wu JM, Lerea KM. Resveratrol decreases early signaling events in washed platelets but has little effect on platelet in whole blood. Blood Cells Mol Dis. 2000;26(2):144–150. doi: 10.1006/bcmd.2000.0289. [DOI] [PubMed] [Google Scholar]
- 118.Johnson WD, Morrissey RL, Usborne AL, Kapetanovic I, Crowell JA, Muzzio M, McCormick DL. Subchronic oral toxicity and cardiovascular safety pharmacology studies of resveratrol, a naturally occurring polyphenol with cancer preventive activity. Food Chem Toxicol. 2011;49(12):3319–3327. doi: 10.1016/j.fct.2011.08.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Vachharajani VT, Liu T, Brown CM, Wang X, Buechler NL, Wells JD, Yoza BK, McCall CE. SIRT1 inhibition during the hypoinflammatory phenotype of sepsis enhances immunity and improves outcome. J Leukoc Biol. 2014;96(5):785–796. doi: 10.1189/jlb.3MA0114-034RR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Patel BP, Safdar A, Raha S, Tarnopolsky MA, Hamadeh MJ. Caloric restriction shortens lifespan through an increase in lipid peroxidation, inflammation and apoptosis in the G93A mouse, an animal model of ALS. PLoS One. 2010;5(2):e9386. doi: 10.1371/journal.pone.0009386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Starr ME, Steele AM, Cohen DA, Saito H. Short-term dietary restriction rescues mice from lethal abdominal sepsis and endotoxemia and reduces the inflammatory/coagulant potential of adipose tissue. Crit Care Med. 2016;44(7):e509–e519. doi: 10.1097/CCM.0000000000001475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Chu X, Schwartz R, Diamond MP, Raju RP. A combination treatment strategy for hemorrhagic shock in a rat model modulates autophagy. Front Med (Lausanne) 2019;6:281. doi: 10.3389/fmed.2019.00281. [DOI] [PMC free article] [PubMed] [Google Scholar]