

Abstract
In this paper, we discuss the basic reproduction number of stochastic epidemic models with random perturbations. We define the basic reproduction number in epidemic models by using the integral of a function or survival function. We study the systems of stochastic differential equations for SIR, SIS, and SEIR models and their stability analysis. Some results on deterministic epidemic models are also obtained. We give the numerical conditions for which the disease-free equilibrium point is asymptotically stable.
Keywords: Basic reproduction number, Random perturbations, Brownian motion, Stability analysis
Introduction
Pandemics can cause sudden and drastic increases in mortality and morbidity rates as well as social, political, and economic disruptions. Humanity can defend itself against these types of problems with advances in science and with professionals in medicine, immunology, genetics, epidemiology, and statisticians. Finding the necessary measures to guarantee people’s access to medical centers is a topic of great interest; controlling the sources and vectors of contagion is the most efficient way to slow down a pandemic. Reducing infection rates guarantees not only well-being but also a reduction in mortality rates. Knowing the mechanisms of spread, infection, and death, modeling them mathematically, and making predictions of populations at risk are the most advantageous state tools to guarantee the right to life. Epidemic models are widely used to analyze the dynamics of populations under infectious diseases. They are crucial for studying the epidemic development and transmission dynamics of a disease. Mathematical models play an important role in predicting, assessing, and controlling potential outbreaks. One of the first epidemic models developed was the SIR model proposed in 1927 by Kermack and McKendrick (see [14]) based on the ordinary differential system given by equation (1.1). The SIR model is a compartmental model where the population is divided into different types of individuals: the susceptible (), the infected (), and the recovered () individuals, respectively, at time t. The transmission-dynamic epidemic models help us understand that the risk of infection among susceptible individuals depends on the prevalence of infectious individuals. An infected individual becomes recovered after receiving treatment. We now give the system of differential equations:
| 1.1 |
where β represents the rate of infection, the infection recovery rate is γ, and N is the total population size such that . However, these previous models do not assume the possibility of immigrants and emigrants. We consider a model with demography, for which μ is considered as the emigration rate and η is the immigration rate. Sometimes the rate μ is considered as the mortality rate and η is the birth rate in standard branching processes.
| 1.2 |
We note that if then the population will be constant. In the above model, we assume that the disease for which infection does not confer immunity is called the population of type SIS (susceptible(S)–infection(I)–susceptible(S)) model since individuals return to the susceptible class when they recover from the infections. Such infections do not have a recovered state and individuals become susceptible again after recovery from infection. Now we describe the population of type SEIR (susceptible(S)–exposed(E)–infection(I)–recovered(R)), and the system of differential equations for the SEIR model (with demography) is given as follows:
| 1.3 |
where the average incubation time is the time for which the infectious agent takes a time to convert an exposed individual into an infected individual. Note that during incubation time the exposed individual cannot transmit the disease. The above models are deterministic. However, the epidemics tend to occur in cycles of outbreaks due to variations in the infection rate mainly related to certain external factors such as people’s social activities and climatic fluctuations (see [24]). In fact, the climatic variations can affect the infection rate . The epidemic models with random perturbation have been widely studied to accommodate randomness in the model, see for example [3, 7, 13, 20, 27]. More recently the evidence of the mechanism by which climate change could have played a direct role in the emergence of COVID-19 has been reported [2].
In this paper, we study the basic reproduction number in epidemic models with random perturbations. We define the basic reproduction number in epidemic models by using the survival function and demonstrate the numerical conditions under which the disease-free equilibrium point is asymptotically stable. The paper is organized as follows: In Sect. 2, we introduce the framework and basic concepts of the stochastic models with random perturbation and establish the stability conditions of the SIS, SIR, and SEIR epidemic models. Section 3 is devoted to the main results illustrated with simulation results for the basic reproduction number for the SIR, SIS, and SEIR models. Section 4 discusses the basic reproduction variable with double perturbation terms for the transmission rate; and finally, Sect. 5 concludes the paper with the future work.
Stochastic model
In this section, we introduce the stochastic modeling of epidemics with random perturbations. In our model, we consider environmental variations and social behaviors in the infection rate [9]. In this paper, we assume to be a complete probability space with a filtration satisfying the usual conditions. We define
| 2.1 |
where β and σ are positive constants, and is the standard Brownian motion with . We note that the constant β is the deterministic mean infection rate, and σ is the perturbation parameter which describes changes in the infection rate changes over time with respect to β. We now introduce the stochastic perturbations (1.1) in the system of stochastic differential equations(SDE) for the SIR model. The resulting SDE is given by
| 2.2 |
Reasoning analogously as in (2.2), we now propose the following system of stochastic differential equations for the SEIR model with random perturbations:
| 2.3 |
The basic reproduction number is defined as the expected number of secondary cases produced by a single infection in a completely susceptible population [4, 6, 10]. In many definitions of basic reproduction number that have been proposed, the basic conceptual framework is similar. This is also called the basic reproduction ratio, which is an epidemiological metric used to describe the transmission of an infectious disease. Mathematically, the basic reproduction number is defined as follows [11].
The basic reproduction number of an epidemic model is given by
| 2.4 |
where is the average number of new infected individuals (in a completely susceptible population) by an infected individual if it is infectious during all the time between 0 and a. is the probability of a new infected individual continuous infecting during the time interval between 0 and a. This is also called the underlying survival probability (or function). Note that in the case of the SEIR model and . For SIR model, and . In this way, the basic reproduction numbers for SIR and SEIR models are, respectively,
| 2.5 |
See the example in Appendix A.1. The basic reproduction number is built for the SEIR model with demography. We now give some basic definitions and preliminary results for the benefit of the readers in the following subsection.
Preliminaries and basic definitions
In this section, we introduce the basic notions and the theoretical framework that we need in this paper. The following definition of equilibrium point is given [12].
Definition 2.1
Let an ordinary differential system be given by
with the matrix notation
| 2.6 |
where is a locally Lipschitz function for all . is called an equilibrium point , where is a matrix with size .
Let be an equilibrium point of the ordinary differential system . If is different to , it is possible to consider the substitution obtaining . In this case, the stability with respect to the point [12] and the reason why the stability and the asymptotic stability are defined for the point have been studied.
Definition 2.2
The point of system (2.6) is called
(i) Stable if and only if, for all , there exists such that
(ii) Asymptotically stable if and only if it is stable and can be chosen such that
Intuitively, is stable if the solutions which start near enough to the path which starts in () remain near enough to the path for every (), that is, if a solution starts near to , then it will never move away enough from the path . The point is asymptotically stable if the solutions which start near to the path with origin in converge to that path (see [12]).
The disease-free equilibrium point results to be locally asymptotically stable if the reproduction number is less than unity, while the endemic equilibrium point is locally asymptotically stable if such a number exceeds unity. In the deterministic epidemic models, the disease-free equilibrium points are locally asymptotically stable if the reproduction number is less than unity. In contrast, the endemic equilibrium point is locally asymptotically stable if the reproduction number exceeds unity (see [23]). For the SEIR model, assume and for any t, and for the models SIS and SIR, . For the deterministic case, the disease-free equilibrium points of the SIR and SEIR models with demography are and , respectively. Under the SIR model we have that if , then is asymptotically stable. Note that a numerical condition on the basic reproduction number holds for the stability of the SIR model. Hence, we establish numerical conditions for which some deterministic epidemic models are asymptotically stable on the disease-free equilibrium points (for more details, see Appendix B). We now briefly discuss the stability analysis for stochastic differential equations on epidemic models with random perturbations. For more details, we recommend readers to refer to [15] and [18].
Definition 2.3
Let the system of stochastic differential equations be as follows:
| 2.7 |
where are locally Lipschitz functions from to . We say that for some is an equilibrium point of the system if it holds .
If is an equilibrium point, and substituting , we have the system
is an equilibrium point. Using this, the stability and the asymptotic stability are defined as follows.
Definition 2.4
Let be a system defined by (2.7), for which is an equilibrium point. We say that is
(i) Stable (in probability) if and only if, for all , there exists such that if , then
(ii) Asymptotically stable if it is stable in probability, and there exists such that if then
Definition 2.5
Let be an Itô process and . We define the differential operator for h as follows:
| 2.8 |
For observing the stability in SIS and SEIR models with random perturbations, using adequate Lyapunov functions, we state now the following theorem given in [22] without proof.
Theorem 2.1
Let defined on be a Lyapunov function.
(i) If for all , then is stable in probability.
(ii) If V satisfies (i) and , then is asymptotically stable.
We prove the following theorem by constructing a Lyapunov function and give the sufficient conditions at which the point is asymptotically stable in the SEIR model with random perturbations. In [17] the author used a similar approach for constructing a Lyapunov function to prove that the endemic equilibrium state is globally asymptotically stable.
Theorem 2.2
If the parameters of the SEIR model with random perturbations satisfy the following:
| 2.9 |
and , then the point is asymptotically stable.
Proof
Let the function be given by
where are adequately chosen. As for all and . In addition, the partial derivatives of V are continuous, therefore V is a Lyapunov function.
We rewrite in the matrix form , with , and f, g given by
For calculating , we have
where
On the other hand, when we have
therefore
such that
See (i) of the proof for Theorem B.2, it is clear that .
On the other hand, as , then
therefore . If , the proof is analogous to Theorem B.2, having for which for any .
Choosing adequately , , and , for any case, it has that
for all , showing that the point is asymptotically stable. □
Theorem 2.3
If the parameters of the SIS model with random perturbation satisfy that
| 2.10 |
then the point is asymptotically stable.
Proof
The proof is similar to the previous theorem. Take V defined by
where are positive constants adequately chosen. □
Theoretically, by inequality (2.10) it is shown that (Theorem 2.3) if
| 2.11 |
then the point is asymptotically stable.
According to Theorem 2.2, that in the SEIR model with random perturbations is asymptotically stable, and it is necessary that and inequality (2.9) hold and can be written as
| 2.12 |
Simulation results for the stability of the stochastic models
In this section, we discuss simulation results of the reproduction numbers , and respectively for SIR, SIS, and SEIR models with random perturbations. Our objective is to find the smallest value of such that and for which the SIS model with random perturbation is asymptotically stable on (according to Theorem 2.3). Similarly, we search for the smallest value of such that and is asymptotically stable on the SEIR model with random perturbations (according to Theorem 2.2). We now observe through simulations the smallest values of and for which the asymptotic stability holds.
We now apply the Euler–Maruyama method for simulating the SIS and SEIR models with random perturbations [21]. The approximation equations of the models are given by
| 3.1 |
| 3.2 |
The numeric conditions for which the disease-free equilibrium for the simulations presented at the point ) on the SIS model with random perturbation is asymptotically stable. Note that when (see Fig. 1, upper left) the asymptotic stability is clear since the functions remain “near” to the constant functions and , varying these functions +0.0005 and −0.0005. Similarly, the asymptotic stability is observed when and (Fig. 1, upper right). When (Fig. 1, lower left), the stability is not so clear, while it is clear when (Fig. 1, lower right). We observe that as guarantees the asymptotic stability for the disease-free equilibrium, based on the simulation results, we propose the following conjecture.
Figure 1.

Stability modeled using the parameters , , , , , and (a) (upper right), (b) (upper left), (c) (lower left) and (d) (lower right). The initial condition is for all of them
Conjecture 3.1
If
| 3.3 |
then is asymptotically stable on the SIS model with random perturbation.
Now, we focus our attention on the simulations of the stability for the SEIR model with random perturbations which are shown for determining the numeric conditions under which the point is asymptotically stable on the SEIR model with random perturbations, for example, the values of ) are verified numerically.
In all of the previous simulations, we assume that . Note that when (see Fig. 2, upper left) the asymptotic stability is clear since the functions remain “near” to the constant functions and , varying these functions +0.0005 and −0.0005. Similarly, the asymptotic stability is observed when and (Fig. 2, upper right). When (Fig. 2, lower left), the instability is not so clear, while the instability is clear when (Fig. 2, lower right) since it is observed that the varied solutions move away from the disease-free equilibrium. As and guarantee the asymptotic stability for the disease-free equilibrium (according to the simulations), we now propose the conjecture.
Figure 2.

Stability modeled using the parameters , , , , , and (a) and (upper right), (b) and (upper left), (c) and (lower left) and (d) and (lower right). The initial condition is for all of them
Conjecture 3.2
If and
| 3.4 |
then is asymptotically stable on the SEIR model with random perturbations.
As the basic reproduction number of the SEIR model with random perturbations (with , ) is the lower number for which is asymptotically stable. In the Fig. 3, we show that the condition is not satisfied.
Figure 3.

Stability modeled using the parameters , , , , , and (a) and (right), (b) and (left). The initial condition is for all of them
It is clear that despite of being , if , the stability is not so clear. Similarly, if and , according to the simulation, the point (in this case ) is unstable. But it is important to have the condition for retaining the asymptotic stability on the SEIR model with random perturbations.
We wish to note that, for the SIR model with random perturbation, the following inequality holds for having the asymptotic stability in for the model proposed in [25] and [28]
| 3.5 |
Basic reproduction variable and their statistical tests
We now study the basic reproduction number as a normally distributed random variable. For the deterministic model, is defined in integral (2.4). Consider the SIR model with random perturbation, the survival integral is given by
| 3.6 |
where is a normally distributed random variable. We refer the reader to consult (A.1) for the SEIR deterministic model. Set , from the above equation, is given by
using the integration-by-parts rule [19], we have an expression which involves given by
where is a Brownian motion, thus
The above integral is well defined, we get (see [16, p. 393])
| 3.7 |
By the law of the iterated logarithm [1, p. 66], we get
we have
| 3.8 |
On the other hand, we have
and by applying the L’Hôpital’s rule
thus,
then inequality (3.8) can be written as
which means that a.s. We see that
Then is the random basic reproduction variable on the SIR model with random perturbation and is given by
| 3.9 |
Similarly, we assume that random basic reproduction variables on the SIS and SEIR models with random perturbations are normally distributed and are given as follows.
Definition 1
| 3.10 |
| 3.11 |
From definition (1) and inequalities (2.10) and (2.9), the following inequalities hold:
| 3.12 |
| 3.13 |
Note that
where is the distribution function of Z such that . The probability p satisfies
that is,
this inequality holds if and only if .
On the other hand, if μ tends to 0, then1, therefore,
which means . This means that when the emigration rate is lower, the random variable is closer to the number . If , then2
thus, if (except for ), then since
Analogously, for the SIS model with random perturbation the following holds:
and
We now discuss the confidence intervals and hypothesis tests from the basic reproduction. Let be the average number of cases of infected people for , respectively. According to the previously mentioned, we assume that , all independent. Note that
to determinate a confidence set under a confidence level , knowing μ, β, γ, υ, and σ, observe that
| 3.14 |
therefore,
Thus,
Then
Similarly, the confidence set is given by , where . For calculating the size of sample with an error e, see that
therefore,
The statistic test Z is given by (3.14) and the critical sets are , , and for the alternative test , , and .
Basic reproduction variable with double stochastic component
In this section, we determine the basic reproduction variable for the model based on the stochastic differential equations with two kinds of perturbation terms. We consider the SEIRS epidemic model with stochastic transmission proposed by Witbooi [26] to include two stochastic perturbation terms in the disease model. It is given by
| 4.1 |
Analogously, the deterministic version of the SEIR model with demography is given by
| 4.2 |
Using the approach of the next generation matrix method(see [5]) for the deterministic model, the matrix T (transmissions) and the matrix Σ (transitions), respectively, are given by
and
The eigenvalues of correspond to
with . It is clear that the greatest eigenvalue is , which is the basic reproduction number for system (4.2).
For system (4.2), we assume that , and as in example (A.1) with function
For system (4.1), take and ([8] and [9]). Based on the construction of integral (3.6), we define the basic reproduction variable for the system:
| 4.3 |
where . Observe that
-
(i)
-
(ii)
, with
-
(iii)Note that ; where , and . The roots of are given by
thereforeIt is easy to observe that, for all ,
with . Note by equation (3.7) that
due to (reasoning similarly to inequality (3.8)), note that .On the other hand, , therefore .
Taking the random variables
we have and are normally distributed with variance and means and , respectively. In addition, for all , it is clear thatWriting , we have that . The distance between and corresponds toObserve that
that is, . The Fig. 4 shows that the function is decreasing for all with . Therefore, when , then , then . This happens when σ, p, q, γ, μ, or υ tends to ∞.On the other hand, note that if , then , which lets us conclude that . This happens when , or . However, if then , thus . If and (at the same time), then , thus . In case that , then , therefore the mean of does not have sense.
Figure 4.
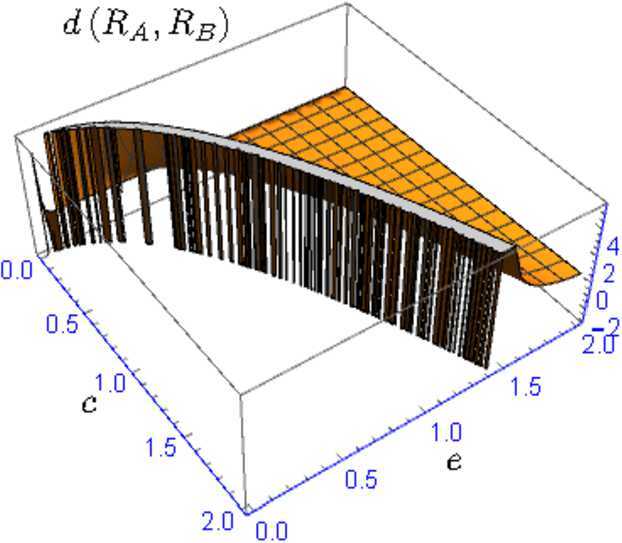
Graphic of the function with restricted to . Our case considers , a function which has similar behavior to
By the procedures done in items (i), (ii), and (iii) of this section, it is possible to see that the basic reproduction number of system (4.1), , is a random variable whose expectation holds
| 4.4 |
with and . If , then .
Conclusions
In this paper, we have studied the basic reproduction number in stochastic epidemic models to include random perturbations in the infection rate as the contributing factor for the spread of the epidemics. We have established stability conditions for the SIS, SIR, and SEIR epidemic models. As in the case of the deterministic SEIR model, the condition is not enough for the disease-free equilibrium point to be asymptotically stable. We showed that it is also necessary that . Also, in some deterministic models, the basic reproduction number is defined as the survival probability, which coincides with the value . If , then the disease-free equilibrium point is asymptotically stable. However, epidemic models with random perturbations need not be the same. In this paper, we considered the basic reproduction number as a random variable. Under stability conditions (Theorems 2.3 and 2.2), we proved that the basic reproduction number depends on the perturbation parameter σ, which means that the variations can affect the epidemic spread. We also presented simulation results that the value of for which the disease-free equilibrium point is asymptotically stable is less than the value found in the proofs of Theorems 2.3 and 2.2. Finally, we presented conjectures (3.1) and (3.2) to conclude that the transmission velocity of an epidemic is lower than the variation fluctuations, and for the values of proved in Theorems 2.3 and 2.2. The limitation of the proposed model is that populations that make transitions to the compartment are assumed to interact homogeneously and death rates are equal. The future work in this direction comprises considering a more realistic scenario using data from the recent COVID-19 outbreak in the city of Bogotá to include the lockdown restrictions and social mobility in the spread of infections that would allow us to address the issue of dependence control measures and epidemics mitigation.
Acknowledgments
Acknowledgements
The excellent comments of the anonymous reviewers are greatly acknowledged and have helped a lot in improving the quality of the paper. This research work is supported by Directorate-Bogotá campus (DIB), Universidad Nacional de Colombia.
Authors’ information
AR, a doctoral student of Statistics Program at Universidad Nacional de Colombia; has Master’s degree in Statistics. ST, full professor and senior researcher at Universidad de Valparaiso, Valparaiso, Chile; published numerous research papers in international journals including Stochastic Analysis and Applications, Statistics & Probability Letters, Journal of Theoretical Probability, Nonlinear Analysis: Real World Applications etc. VA, associate professor at Universidad Nacional de Colombia; published research papers in several journals such as Mathematical Biosciences, Stochastic Analysis and Applications, Journal of Risk, Journal Biological Systems, Annals of Operations Research, Computers and Mathematics with Applications, among others. He is a co-author of a text book entitled “Introduction to Probability and Stochastic Processes with Applications” in John Wiley & Sons 2012.
Appendix A: Construction of the basic reproduction number on deterministic epidemic models
Example A.1
(Basic reproduction number in a deterministic SEIR model with demography)
Let be “number of exposed population which become infected individuals and remain infected from the time 0 to a”. Note that the number of individuals per unit of time which avoid being exposed people during the period are those that died or who became infected individuals, that is, individuals per unit of time. The individuals who recovered from the disease or died are those who do not remain infected during the period , namely individuals per unit of time. The others continue being exposed people or they are infected individuals which remain infected during .
solving the differential equation, we get
Initially it needs to have at least an infected individual or an exposed individual which becomes infected, for when the epidemic occurs, then is the number of initial infected people. is the number of infected individuals which remain infected during the period . Note that corresponds to multiplied by the probability that an infected individual continues to be infected during all the interval . Therefore, the probability previously described. In this way,
is the survival function.
On the other hand, if an infected individual, , arrives at a place where the population is completely susceptible, , then it is expected to have βN exposed individuals in total. From the βN expected exposed individuals, corresponds to the total infected population, therefore
as long as the mortality rate is the same as the birth rate. In another case, note that
and the solution of the above equation is given by
when , then tends to . Therefore, if an infected individual arrives in a completely susceptible population, then it will have new infected on an enough big period of time. Thus, the function is given by
then the basic reproduction number for the model SEIR with demography is
| A.1 |
Appendix B: Stability on deterministic epidemic models
We give the following theorem which gives sufficient conditions for a point to be asymptotically stable using the appropriate Lyapunov functions ([12] and [25]).
Theorem B.1
Let be an equilibrium point of system (2.6) (in the other case, it is possible to do the substitution , where is an equilibrium point) defined for all , and is a Lyapunov function. Then, for some ,
(i) If V satisfies that
| B.1 |
then is stable.
(ii) If V satisfies (i) and furthermore
| B.2 |
then is asymptotically stable.
To prove the stability of an ordinary equation system, we use construction of the Lyapunov functions. The definition is given in the following definition given in [12].
Definition B.1
Let be an ordinary differential equation system defined for all , and let be a continuous function with continuous derivatives.
(i) The rate of V with respect to is defined as
| B.3 |
(ii) If V satisfies that and for all , then V is called a Lyapunov function.
We give the theorems which relate with the basic reproduction number and disease-free equilibrium point for the deterministic models. The proofs are based on [18] and [27]. Now we give the following theorem for the SEIR model with the demography.
Theorem B.2
If and , then is asymptotically stable in the SEIR model with demography.
Proof
Assume . Define the function W given by
where are positive constants adequately chosen. Clearly for all and . Given , , , and are continuous functions and
are continuous too, then V is a Lyapunov function. Notice that
where3
Now, it is an objective to show that for all with . If
-
(i)As for all (by ), then
when then . -
(ii)Notice that for all and as
then , therefore
so by (i) and (ii) we have that . In this way it is possible to chose the values for that hold on
Then by Theorem B.1 it is concluded that for the SEIR model with demography is asymptotically stable.
If , and also for all for which , it is clear that . If there exists for which , analogously like it was made for the proof of Theorem B.3 and following that
it is shown that and for all for some .
For any case, we have that , which is why it is possible to choose adequate values for , , and such that
In consequence, the point is asymptotically stable in the SEIR model with demography. □
Theorem B.3
If and , then and are asymptotically stable in (i) SIR y (ii) SIS models with demography, respectively.
Proof
The proof is similar to the previous theorem, taking V defined by
where are appropriately chosen positive constants. □
Authors’ contributions
AR, ST, and VA performed the stochastic analysis, and AR analyzed simulations. All authors wrote and revised the final version of the manuscript.
Funding
This work was financially supported by Directorate-Bogotá campus (DIB), Universidad Nacional de Colombia under project No. 41097 and project No. 50803.
Availability of data and materials
Not applicable. All data generated or analysed during this study are simulated and included in this manuscript.
Competing interests
The authors declare that they have no competing interests.
Footnotes
by the L’Hôpital’s rule.
by the L’Hôpital’s rule.
Later, it is shown that is negative.
Contributor Information
Andrés Ríos-Gutiérrez, Email: asriosg@unal.edu.co.
Soledad Torres, Email: soledad.torres@uv.cl.
Viswanathan Arunachalam, Email: varunachalam@unal.edu.co.
References
- 1.Bernt Ø. Stochastic Differential Equations: An Introduction with Applications. Berlin: Springer; 2013. [Google Scholar]
- 2.Beyer R.M., Manica A., Mora C. Shifts in global bat diversity suggest a possible role of climate change in the emergence of SARS-CoV-1 and SARS-CoV-2. Sci. Total Environ. 2021;767:145413. doi: 10.1016/j.scitotenv.2021.145413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cai Y., Jiao J., Gui Z., Liu Y., Wang W. Environmental variability in a stochastic epidemic model. Appl. Math. Comput. 2018;329:210–226. [Google Scholar]
- 4.Diekmann O., Heesterbeek J.A.P., Metz J.A. On the definition and the computation of the basic reproduction ratio in models for infectious diseases in heterogeneous populations. J. Math. Biol. 1990;28(4):365–382. doi: 10.1007/BF00178324. [DOI] [PubMed] [Google Scholar]
- 5.Diekmann O., Heesterbeek J.A.P., Roberts M.G. The construction of next-generation matrices for compartmental epidemic models. J. R. Soc. Interface. 2009;7(47):873–885. doi: 10.1098/rsif.2009.0386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dietz K. The estimation of the basic reproduction number for infectious diseases. Stat. Methods Med. Res. 1993;2(1):23–41. doi: 10.1177/096228029300200103. [DOI] [PubMed] [Google Scholar]
- 7.Divine W. Complete global analysis of a two-scale network SIRS epidemic dynamic model with distributed delay and random perturbations. Appl. Math. Comput. 2017;294:49–76. [Google Scholar]
- 8.El Fatini M., Laaribi A., Pettersson R., Taki R. Lévy noise perturbation for an epidemic model with impact of media coverage. Stochastics. 2019;91:998–1019. doi: 10.1080/17442508.2019.1595622. [DOI] [Google Scholar]
- 9.Gray A., Greenhalgh D., Hu L., Mao X., Pan J. A stochastic differential equation SIS epidemic model. SIAM J. Appl. Math. 2011;71(3):876–902. doi: 10.1137/10081856X. [DOI] [Google Scholar]
- 10.Heesterbeek J.A.P. A brief history of and a recipe for its calculation. Acta Biotheor. 2002;50(3):189–204. doi: 10.1023/A:1016599411804. [DOI] [PubMed] [Google Scholar]
- 11.Heffernan J.M., Smith R.J., Wahl L.M. Perspectives on the basic reproductive ratio. J. R. Soc. Interface. 2005;2(4):281–293. doi: 10.1098/rsif.2005.0042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Hokayem, P.A., Gallestey, E.: Lyapunov Stability Theory. Nonlinear Syst. Control, Spring (2015)
- 13.Ji C., Jiang D. The threshold of a non-autonomous SIRS epidemic model with stochastic perturbations. Math. Methods Appl. Sci. 2017;40(5):1773–1782. doi: 10.1002/mma.4096. [DOI] [Google Scholar]
- 14.Kermack W.O., McKendrick A.G. Proceedings of the Royal Society of London. Series A, Containing Papers of a Mathematical and Physical Character. 1927. Contribution to the mathematical theory of epidemics; pp. 700–721. [Google Scholar]
- 15.Khasminskii R. Stochastic Stability of Differential Equations. Berlin: Springer; 2011. [Google Scholar]
- 16.Klebaner F.C. Introduction to Stochastic Calculus with Applications. Singapore: World Scientific; 2012. [Google Scholar]
- 17.Lahrouz A., Omari L., Kiouach D. Global analysis of a deterministic and stochastic nonlinear SIRS epidemic model. Nonlinear Anal., Model. Control. 2011;16(1):59–76. doi: 10.15388/NA.16.1.14115. [DOI] [Google Scholar]
- 18.Lahrouz A., Settati A. Necessary and sufficient condition for extinction and persistence of SIRS system with random perturbation. Appl. Math. Comput. 2014;233:10–19. [Google Scholar]
- 19.Lai Chung K., Williams R.J., Williams R.J. Introduction to Stochastic Integration. Berlin: Springer; 1990. [Google Scholar]
- 20.Lin Y., Jiang D., Xia P. Long-time behavior of a stochastic SIR model. Appl. Math. Comput. 2014;236:1–9. [Google Scholar]
- 21.Liu W., Mao X. Strong convergence of the stopped Euler–Maruyama method for nonlinear stochastic differential equations. Appl. Math. Comput. 2013;223:389–400. [Google Scholar]
- 22.Mao X. Stochastic Differential Equations and Applications. 2. Cambridge: Woodhead Publishing; 2008. [Google Scholar]
- 23.Morris Q. Analysis of a co-epidemic model. SIAM Undergrad. Res. Online. 2010;4:121–133. doi: 10.1137/11S010852. [DOI] [Google Scholar]
- 24.Sturrock R.N., Frankel S.J., Brown A.V., Hennon P.E., Kliejunas J.T., Lewis K.J., Worrall J.J., Woods A.J. Climate change and forest diseases. Plant Pathol. 2011;60(1):133–149. doi: 10.1111/j.1365-3059.2010.02406.x. [DOI] [Google Scholar]
- 25.Tornatore E., Buccellato S.M., Vetro P. Stability of a stochastic SIR system. Phys. A, Stat. Mech. Appl. 2005;354:111–126. doi: 10.1016/j.physa.2005.02.057. [DOI] [Google Scholar]
- 26.Witbooi P.J. An SEIRS epidemic model with stochastic transmission. Adv. Differ. Equ. 2017;2017(1):109. doi: 10.1186/s13662-017-1166-6. [DOI] [Google Scholar]
- 27.Yang Q., Jiang D., Shi N., Ji C. The ergodicity and extinction of stochastically perturbed SIR and SEIR epidemic models with saturated incidence. J. Math. Anal. Appl. 2012;388(1):248–271. doi: 10.1016/j.jmaa.2011.11.072. [DOI] [Google Scholar]
- 28.Zhou Y., Zhang W., Yuan S. Survival and stationary distribution of a SIR epidemic model with stochastic perturbations. Appl. Math. Comput. 2014;244:118–131. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable. All data generated or analysed during this study are simulated and included in this manuscript.