

Abstract
Challenges have been recognized in healthcare of patients with Alzheimer’s disease (AD) in the COVID-19 pandemic, given a high infection and mortality rate of COVID-19 in these patients. This situation urges the identification of underlying risks and preferably biomarkers for evidence-based, more effective healthcare. Towards this goal, current literature review and network analysis synthesize available information on the AD-related gene APOE into four lines of mechanistic evidence. At a cellular level, the risk isoform APOE4 confers high infectivity by the underlying coronavirus SARS-CoV-2; at a genetic level, APOE4 is associated with severe COVID-19; at a pathway level, networking connects APOE with COVID-19 risk factors such as ACE2, TMPRSS2, NRP1, and LZTFL1; at a behavioral level, APOE4-associated dementia may increase the exposure to coronavirus infection which causes COVID-19. Thus, APOE4 could exert multiple actions for high infection and mortality rates of the patients, or generally, with COVID-19.
Keywords: APOE4, Biomarker, Coronavirus, Comorbidity, Peripheral mechanisms, COVID-19
Background
In the midst of the COVID-19 pandemic, patients with Alzheimer’s disease (AD), once infected by the underlying coronavirus SARS-CoV-2, are 5 times likely to die of this infectious disease [1]. In the absence of effective treatment, a mechanistic understanding of how the patients with AD become a vulnerable target of COVID-19 may guide evidence-based healthcare management and targeted therapeutics development. In the earlier literature, it was postulated that APOE4 (italic for gene), a genetic risk factor for AD [2], would be a biomarker for severe COVID-19 [3]; others considered psychological and behavioral contributions [4]. This review aims to capitalize on the evolving literature and database resources and seek a fundamental or molecular understanding of the high vulnerability in patients with AD.
Genetic evidence for APOE4 involvement in the vulnerability for COVID-19
There is evidence for the genetic contribution to comorbidity of other brain disorders with COVID-19 [5] so that genetics may explain the high vulnerability of patients with AD as well. APOE4 is the most established genetic risk factor for late-onset AD. An in vitro study has suggested that cells expressing APOE4 are more vulnerable to SARS-CoV-2 infection than those expressing the nonpathogenic isoform APOE3 [6]. Via induced pluripotent stem cells (iPSC)-based in vitro technologies, cells with APOE4 allowed more significant SARS-CoV-2 infection of artificially differentiated either neurons or astrocytes than those with APOE3. Furthermore, APOE4 astrocytes infected with SARS-CoV-2 presented a more severe cytopathogenic effect than APOE3 astrocytes, which could facilitate the progression and severity of COVID-19. This study indeed provided the first insight to a possible APOE-mediated mechanism for COVID-19 severity. It remained unknown how this genetic vulnerability was achieved and more importantly what this finding meant for a high infection rate in APOE4 carriers and increased COVID-19 mortality in the comorbid patients.
The difference between APOE4 and APOE2/3 is caused by the single nucleotide polymorphism (SNP) rs429358. This SNP carries two alleles, T and C, where T encodes a cystine (APOE3) and C encodes an arginine (APOE4) at residue position 130. In APOE4 carriers, reduced expression levels of APOE in both brain and peripheral systems suggested that this variant causes an increased risk not only for AD [7, 8], but also for systemic susceptibility to coronavirus infection. APOE4 has a worldwide average frequency of 15%, according to the 1000 Genomes Project [9], meaning that approximately 2% of the worldwide population are homozygotes, equal to the current 2% of the world population that have been diagnosed with COVID-19. In fact, APOE is abundantly expressed in cells of various peripheral systems including macrophages and epithelial cells of the lung [10, 11] which confers the most severe impact on coronavirus pathology. As an explanation, patients with APOE4-associated AD may carry higher vulnerability in their peripheral organs such as the lung than those carrying APOE3, possibly via enhancing the receptors’ activities and facilitating the coronavirus entry.
Pathway support for a role of APOE4 in the severity of COVID-19
To explore this possibility, we have used a pathway analysis approach in MetaCore as previously described [12]. Results from this pathway analysis indeed support that possibility by identifying a plausible ten-member network where all ten members have been implicated in AD: APOE [13], ACE2 [14], CTNNB1 [15], NOTCH1 [16, 17], LZTFL1 [18], MMP1 [19], NRP1 [20], RELA [21], SIRT1 [22], and MMP14 [19] (left panel in Fig. 1). Among them, ACE2 and NRP1 are utilized by SARS-CoV-2 in order to enter cells and cause COVID-19 [23, 24]. ACE2 is almost undetectable and NRP1 has low expression levels in the brain. For a better understanding of the epidemiological finding, these functional genetic and pathway findings may encourage and re-direct our attention from the brain with AD to comorbid patients’ peripheral systems, where both receptors are well expressed and the coronavirus has an easy access, does extensive damages during the progression, and causes multiorgan damage-triggered mortality of COVID-19. Of note, it has been experimentally shown that the coronavirus (SARS-CoV) enters cells by binding to ACE2 while ACE2 may recycle subsequently back to cell surface after unloading of the virus [25]. Specifically, ACE2-related endocytosis, besides direct membrane fusion, has been proposed as an entry mechanism [26], consistent with the recent identification of the endosomal protein TMEM106B as another risk factor for the coronavirus infection [27].
Fig. 1.

Possible mechanisms of APOE4-mediated AD and COVID-19 comorbidity. Left panel: A ten-member network shared by both AD (all members) and COVID-19 (*), generated by using MetaCore. In the case of APOE4, reduced/altered APOE has three potential actions in this network alone: (1) disinhibition of ACE2, (2) transcriptional reduction of the protective LZTFL1, and (3) more indirect disinhibition of NRP1 via LZTF1, in exacerbation of COVID-19. Asterisks are for genetic association with severe COVID-19: APOE: rs429358 (APOE2/3 vs 4) p = 0.0026, OR = 1.31; ACE2: chr23:15564667 p = 0.0056, OR = 1.12; CTNNB1: chr3:41204313 p = 0.016, OR = 0.74; LZTFL1: chr3:45834967 p = 1.15 × 10−10, OR = 0.56; NOTCH1: chr9:136510909 p = 0.0092, OR = 0.87; MMP1: rs11621460 p = 0.010, OR = 0.84; NRP1: chr10:33292184 p = 0.00072, OR = 1.47; RelA: rs1049728 p = 0.0063, OR = 0.64 (II); SIRT1: rs12783242 p = 0.0019, OR = 0.78; where chromosome positions are based on HG38 in the absence of rs numbers; OR, odds ratio; II, adjusted with gender and age; all association signals are provided by the GWAS meta-analysis by Ellinghaus et al. Not shown here is the additional APOE-LRP1-PARP1-TMPRSS2 pathway (see text): LRP1: rs4759044 p = 0.023, OR = 0.89; PARP1: chr1:226405149 p = 0.0065, OR = 0.51 (II). Right panel: APOE4-based vulnerability for patients with COVID-19 and AD at double risk: attenuated protective behavior for exposure risk and APOE4-associated infection risk
Further pathway analyses found that SIRT1, the direct target of APOE, might regulate TMEM106 through binding to several AD-related transcription factors (e.g., FOXP3 [28], STAT1 [29], BMAL1 [30, 31], SIRT6 [32], and E2F1 [33]) but the specifics on these regulations and on how TMEM106B regulates coronavirus’ intracellular activity remain to be uncovered. Interestingly, FOXP3 also bound to TMPRSS2 which encoded another cell surface risk factor for SARS-CoV-2 infection [23]. Also, reduced APOE could reduce LRP1 inhibition of the TMPRSS2 activator PARP1, which in turn promotes the coronavirus infection as well (detailed pathway not shown) [34, 35]. Therefore, APOE might regulate the infectivity of SARS-CoV-2 in multiple ways.
Among the ten members, LZTFL1 represents the most significant genetic risk factor for severe COVID-19 as per findings from two genome-wide association studies (GWAS) of severe COVID-19 [18, 36]. Specifically, a minor allele of a SNP (G/GA, without a “rs” number yet) at chr3:45834967 in LZTFL1 was protective against progression to severe forms of COVID-19. Together, APOE4 may have tetrad action: it enhances ACE2 activity by disinhibiting SIRT1 (Sirtuin 1, a generic enzyme), activates TMPRSS2 by the LRP1-PARP1 pathway, decreases the LZTFL1 expression by inhibiting NOTCH1, and activates NRP1 via LZTFL1 indirectly, satisfying the protective roles of both APOE and LZTFL1. That is, reduced APOE levels, which have demonstrated to be associated with the increased risk of AD, may disinhibit ACE2, TMPRSS2, and NRP1 and consistently increase the vulnerability to the coronavirus infection in the patients with AD. In fact, indirect activation of APOE by LZTFL1 via CTNNB1 (generic binding protein) and MPPs (metalloprotease) fits with their protections against the fatal comorbidity.
More interestingly, nine of the ten members in this network, along with LRP1 and PARP1 in the TMPRSS2 pathway, had nominal significance for genetic associations indeed with the severity of COVID-19 (Fig. 1 legend for left panel), as revealed by the meta-analysis of GWAS [18]. For APOE, it was rs429358 that encodes APOE4 (pmeta = 0.0026), but not another nearby (only 138 bp away) SNP rs7412 C/T that differentiates the nonpathogenic APOE2 vs 3 (Cys176Arg) (pmeta = 0.73), that showed an association with severe COVID-19, selectively supporting the underlying risk of APOE4 and the in vitro experimental finding that there is an association between APOE4 and COVID-19 infectivity. The APOE networking had a 17.6-fold enrichment for associations with severe COVID-19 based on pmeta-values, comparing to the whole GWAS, pointing to a shared molecular etiology.
It remains unknown how significant this pathway information contributes to ACE2/TMPRSS2/NRP1-related infection itself. Such information however encourages modeling analysis of human peripheral (epithelial and immune) cells that bear the brunt of the coronavirus infection for further clarification of the APOE4 mechanism in COVID-19 development and progression. Even worse for AD, the molecular vulnerability can be furthered by exposure-related behavioral disadvantage in the patients (right panel in Fig. 1).
Conclusion
Comparing to two other isoforms, APOE4 is genetically associated with reduced APOE levels for increased coronavirus infection and disease progression risks, and consistently with severe COVID-19 as well. As summarized in Fig. 2, the association between APOE4 and coronavirus infectivity supports the hypothesis that APOE4 is an important risk marker for the severity of COVID-19 pathology in patients with AD. If further verified, APOE genotyping may help guide evidence-based healthcare of the comorbid patients.
Fig. 2.
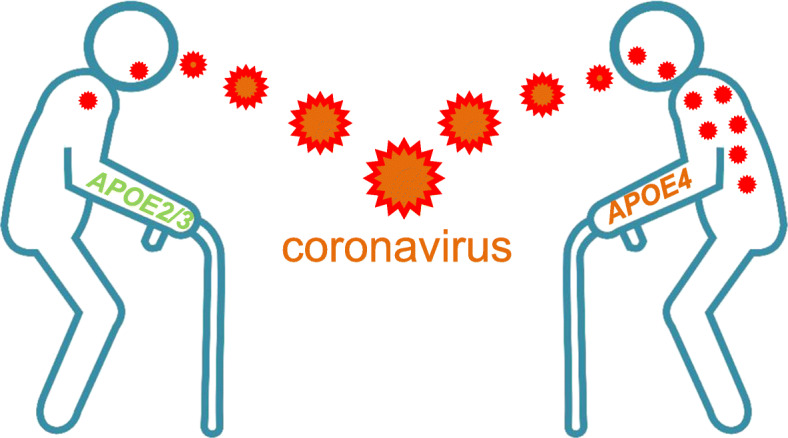
Summary: coronavirus targets AD patients carrying APOE4
Acknowledgements
We thank Dr. Liming Chen for proof-reading and discussing this work.
Authors’ contributions
ZL conceived the work, generated the pathway, collected the data, and drafted the manuscript; NX, JL, and XC contributed to writing and editing; ZL and MRS contributed to revision and finalization. The authors read and approved the final manuscript.
Authors’ information
NX is the President of Wuhan Red Cross Hospital and a neurologist; JL is a Ph.D. student; XC is the head of the Neurology Department and a clinician; MRS is the Director of the Nevada Institute of Personalized Medicine at UNLV; ZL is an associate neurobiologist.
Funding
Not applicable.
Availability of data and materials
The corresponding author has the data available.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Liu N, Sun J, Wang X, Zhao M, Huang Q, Li H. The impact of dementia on the clinical outcome of COVID-19: a systematic review and meta-analysis. J Alzheimers Dis. 2020;78(4):1775–1782. doi: 10.3233/JAD-201016. [DOI] [PubMed] [Google Scholar]
- 2.Emrani S, Arain HA, DeMarshall C, Nuriel T. APOE4 is associated with cognitive and pathological heterogeneity in patients with Alzheimer’s disease: a systematic review. Alzheimers Res Ther. 2020;12(1):141. doi: 10.1186/s13195-020-00712-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kuo CL, Pilling LC, Atkins JL, Masoli JAH, Delgado J, Kuchel GA, Melzer D. APOE e4 genotype predicts severe COVID-19 in the UK Biobank community cohort. J Gerontol A Biol Sci Med Sci. 2020;75(11):2231–2232. doi: 10.1093/gerona/glaa131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Keng A, Brown EE, Rostas A, Rajji TK, Pollock BG, Mulsant BH, Kumar S. Effectively caring for individuals with behavioral and psychological symptoms of dementia during the COVID-19 pandemic. Front Psychiatry. 2020;11:573367. doi: 10.3389/fpsyt.2020.573367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lin, Z., Mechanisms for substance use disorders in COVID-19. Mol Psychiatry. 2021. p. 1-2. 10.1038/s41380-021-01041-0. [DOI] [PMC free article] [PubMed]
- 6.Wang C, et al. ApoE-isoform-dependent SARS-CoV-2 neurotropism and cellular response. Cell Stem Cell. 2021;28(2):331–342.e5. doi: 10.1016/j.stem.2020.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.de Chaves EP, Narayanaswami V. Apolipoprotein E and cholesterol in aging and disease in the brain. Future Lipidol. 2008;3(5):505–530. doi: 10.2217/17460875.3.5.505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Rasmussen KL, Tybjærg-Hansen A, Nordestgaard BG, Frikke-Schmidt R. APOE and dementia - resequencing and genotyping in 105,597 individuals. Alzheimers Dement. 2020;16(12):1624–1637. doi: 10.1002/alz.12165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Abecasis GR, et al. A map of human genome variation from population-scale sequencing. Nature. 2010;467(7319):1061–1073. doi: 10.1038/nature09534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yao X, Gordon EM, Figueroa DM, Barochia AV, Levine SJ. Emerging roles of apolipoprotein E and apolipoprotein A-I in the pathogenesis and treatment of lung disease. Am J Respir Cell Mol Biol. 2016;55(2):159–169. doi: 10.1165/rcmb.2016-0060TR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Martínez-Martínez AB, Torres-Perez E, Devanney N, del Moral R, Johnson LA, Arbones-Mainar JM. Beyond the CNS: the many peripheral roles of APOE. Neurobiol Dis. 2020;138:104809. doi: 10.1016/j.nbd.2020.104809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Liu K, Zhao J, Chen C, Xu J, Bell RL, Hall FS, Koob GF, Volkow ND, Qing H, Lin Z. Epistatic evidence for gender-dependant slow neurotransmission signalling in substance use disorders: PPP1R12B versus PPP1R1B. EBioMedicine. 2020;61:103066. doi: 10.1016/j.ebiom.2020.103066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Michaelson DM. APOE ε4: the most prevalent yet understudied risk factor for Alzheimer’s disease. Alzheimers Dement. 2014;10(6):861–868. doi: 10.1016/j.jalz.2014.06.015. [DOI] [PubMed] [Google Scholar]
- 14.Ding, Q., et al., Protein expression of angiotensin-converting enzyme 2 (ACE2) is upregulated in brains with Alzheimer’s disease. Int J Mol Sci. 2021;22(4):1687. [DOI] [PMC free article] [PubMed]
- 15.Conejero-Goldberg C, Hyde TM, Chen S, Dreses-Werringloer U, Herman MM, Kleinman JE, Davies P, Goldberg TE. Molecular signatures in post-mortem brain tissue of younger individuals at high risk for Alzheimer’s disease as based on APOE genotype. Mol Psychiatry. 2011;16(8):836–847. doi: 10.1038/mp.2010.57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cho SJ, Yun SM, Jo C, Jeong J, Park MH, Han C, Koh YH. Altered expression of Notch1 in Alzheimer’s disease. Plos One. 2019;14(11):e0224941. doi: 10.1371/journal.pone.0224941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Brai E, Alina Raio N, Alberi L. Notch1 hallmarks fibrillary depositions in sporadic Alzheimer’s disease. Acta Neuropathol Commun. 2016;4(1):64. doi: 10.1186/s40478-016-0327-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ellinghaus D, et al. Genomewide association study of severe Covid-19 with respiratory failure. N Engl J Med. 2020;383(16):1522–1534. doi: 10.1056/NEJMoa2020283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bellucci C, Lilli C, Baroni T, Parnetti L, Sorbi S, Emiliani C, Lumare E, Calabresi P, Balloni S, Bodo M. Differences in extracellular matrix production and basic fibroblast growth factor response in skin fibroblasts from sporadic and familial Alzheimer’s disease. Mol Med. 2007;13(9-10):542–550. doi: 10.2119/2007-00034.Bellucci. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Key-Hwan Lim, S.Y., Sung-Hyun Kim, Jae-Yeol Joo, Identifying new COVID-19 receptor Neuropilin-1 in severe Alzheimer’s diseases patients group brain using genome-wide association study approach. Preprint, 2021. [DOI] [PMC free article] [PubMed]
- 21.Chowdhury A, Rajkumar AP. Systematic review of gene expression studies in people with Lewy body dementia. Acta Neuropsychiatr. 2020;32(6):281–292. doi: 10.1017/neu.2020.13. [DOI] [PubMed] [Google Scholar]
- 22.Camporez D, Belcavello L, Almeida JFF, Silva-Sena GG, Pimassoni LHS, Morelato RL, do Carmo Pimentel Batitucci M, de Paula F. Positive association of a Sirt1 variant and parameters of oxidative stress on Alzheimer’s disease. Neurol Sci. 2021;42(5):1843–1851. doi: 10.1007/s10072-020-04704-y. [DOI] [PubMed] [Google Scholar]
- 23.Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH, Nitsche A, Müller MA, Drosten C, Pöhlmann S. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. 2020;181(2):271–280.e8. doi: 10.1016/j.cell.2020.02.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cantuti-Castelvetri L, Ojha R, Pedro LD, Djannatian M, Franz J, Kuivanen S, van der Meer F, Kallio K, Kaya T, Anastasina M, Smura T, Levanov L, Szirovicza L, Tobi A, Kallio-Kokko H, Österlund P, Joensuu M, Meunier FA, Butcher SJ, Winkler MS, Mollenhauer B, Helenius A, Gokce O, Teesalu T, Hepojoki J, Vapalahti O, Stadelmann C, Balistreri G, Simons M. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science. 2020;370(6518):856–860. doi: 10.1126/science.abd2985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wang H, Yang P, Liu K, Guo F, Zhang Y, Zhang G, Jiang C. SARS coronavirus entry into host cells through a novel clathrin- and caveolae-independent endocytic pathway. Cell Res. 2008;18(2):290–301. doi: 10.1038/cr.2008.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Murgolo N, Therien AG, Howell B, Klein D, Koeplinger K, Lieberman LA, Adam GC, Flynn J, McKenna P, Swaminathan G, Hazuda DJ, Olsen DB. SARS-CoV-2 tropism, entry, replication, and propagation: considerations for drug discovery and development. Plos Pathog. 2021;17(2):e1009225. doi: 10.1371/journal.ppat.1009225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Baggen J, Persoons L, Vanstreels E, Jansen S, van Looveren D, Boeckx B, Geudens V, de Man J, Jochmans D, Wauters J, Wauters E, Vanaudenaerde BM, Lambrechts D, Neyts J, Dallmeier K, Thibaut HJ, Jacquemyn M, Maes P, Daelemans D. Genome-wide CRISPR screening identifies TMEM106B as a proviral host factor for SARS-CoV-2. Nat Genet. 2021;53(4):435–444. doi: 10.1038/s41588-021-00805-2. [DOI] [PubMed] [Google Scholar]
- 28.Wu Y, Zhao Y, Xu T, You LW, Zhang H, Liu F. Alzheimer’s disease affects severity of asthma through methylation control of Foxp3 promoter. J Alzheimers Dis. 2019;70(1):121–129. doi: 10.3233/JAD-190315. [DOI] [PubMed] [Google Scholar]
- 29.Zhang Z, Li XG, Wang ZH, Song M, Yu SP, Kang SS, Liu X, Zhang Z, Xie M, Liu GP, Wang JZ, Ye K. δ-Secretase-cleaved Tau stimulates Aβ production via upregulating STAT1-BACE1 signaling in Alzheimer’s disease. Mol Psychiatry. 2021;26(2):586–603. doi: 10.1038/s41380-018-0286-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cronin P, McCarthy MJ, Lim ASP, Salmon DP, Galasko D, Masliah E, de Jager PL, Bennett DA, Desplats P. Circadian alterations during early stages of Alzheimer’s disease are associated with aberrant cycles of DNA methylation in BMAL1. Alzheimers Dement. 2017;13(6):689–700. doi: 10.1016/j.jalz.2016.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yoo, I.D., et al., Elevated CLOCK and BMAL1 contribute to the impairment of aerobic glycolysis from astrocytes in Alzheimer’s disease. Int J Mol Sci. 2020;21(21):7862. [DOI] [PMC free article] [PubMed]
- 32.Akter, R., et al., A comprehensive analysis into the therapeutic application of natural products as SIRT6 modulators in Alzheimer’s disease, aging, cancer, inflammation, and diabetes. Int J Mol Sci. 2021;22(8):4180. [DOI] [PMC free article] [PubMed]
- 33.Rahman MR, Islam T, Turanli B, Zaman T, Faruquee HM, Rahman MM, Mollah MNH, Nanda RK, Arga KY, Gov E, Moni MA. Network-based approach to identify molecular signatures and therapeutic agents in Alzheimer’s disease. Comput Biol Chem. 2019;78:431–439. doi: 10.1016/j.compbiolchem.2018.12.011. [DOI] [PubMed] [Google Scholar]
- 34.Van Gool B, et al. LRP1 has a predominant role in production over clearance of Aβ in a mouse model of Alzheimer’s disease. Mol Neurobiol. 2019;56(10):7234–7245. doi: 10.1007/s12035-019-1594-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Salech F, Ponce DP, Paula-Lima AC, SanMartin CD, Behrens MI. Nicotinamide, a poly [ADP-ribose] polymerase 1 (PARP-1) inhibitor, as an adjunctive therapy for the treatment of Alzheimer’s disease. Front Aging Neurosci. 2020;12:255. doi: 10.3389/fnagi.2020.00255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Pairo-Castineira E, et al. Genetic mechanisms of critical illness in COVID-19. Nature. 2021;591(7848):92–98. doi: 10.1038/s41586-020-03065-y. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The corresponding author has the data available.