

Abstract
Malaria remains one of the most important public health problems, causing significant morbidity and mortality. Malaria is a mosquito borne disease transmitted through an infectious bite from the female Anopheles mosquito. Malaria control will eventually rely on a multitude of approaches, which includes ways to block transmission to, through and from mosquitoes. To study mosquito stages of malaria parasites in the laboratory, we have optimized a protocol to culture highly infectious Plasmodium falciparum gametocytes, a parasite stage required for transmission from the human host to the mosquito vector. P. falciparum gametocytes mature through five morphologically distinct steps, which takes approximately 1–2 weeks. Gametocyte culture described in this protocol is completed in 15 days and are infectious to mosquitoes from days 15–18. These protocols were developed to maintain a continuous cycle of infection competent gametocytes and to maintain uninterrupted supply of mosquito stages of the parasite. Here, we describe the methodology of gametocyte culture and how to infect mosquitoes with these parasites using glass membrane feeders.
Introduction
Malaria is caused by Plasmodium parasites and is transmitted to their vertebrate hosts via infectious bite of female Anopheles mosquitoes. According to the 2019 World Health Organization (WHO) report, there were an estimated 405,000 deaths, from a total of 228 million cases of malaria1. Most of the malaria related deaths were concentrated in the African region, especially among children below five years of age. While the overall incidence rate of malaria has declined globally from 2010, in recent years decline has plateaued and additional control strategies are urgently needed to eliminate the disease.
Cyclic asexual blood stages of malaria parasites cause disease pathogenesis and a small subset of these differentiate into female and male gametocytes. Plasmodium falciparum gametocytes are unique in nature as they take 7–10 days to develop through five morphologically distinct stages. Immature gametocytes from stage I to IV are sequestered in bone marrow parenchyma and largely remain absent from peripheral circulation2, 3, 4, 5. Erythrocytes infected with mature stage V gametocytes are released in the bloodstream and freely circulate to be taken up by mosquitoes. Once inside the mosquito midgut, gametocytes are activated, through a change in temperature and exposure to the midgut environment, transform into female and male gametes and begin development of the mosquito stages, which culminates with the infective stages of sporozoites in the mosquito salivary glands6, 7.
Since Trager and Jenson8 described a standardized method to culture P. falciparum, studies on the asexual blood stages have greatly advanced. However, the lack of a reliable culture system for sexual stages has made it difficult to study P. falciparum gametocytes, transmission biology and mosquito stages. In recent years, several methods have been published which have aided laboratories in establishing gametocyte cultures9, 10, 11, 12. This manuscript describes standardized and reliable protocol to culture P. falciparum gametocytes that can represent a valuable resource for the malaria research community. This method enables the robust production of mature and infectious gametocytes which along with a standardized mosquito feeding protocol, results in highly reliable mosquito infectivity. These methods were established to maintain uninterrupted supply of gametocytes, and mosquito stage parasites. In this manuscript, we describe a thorough gametocyte culture protocol (Figure 1), preparation of glass membrane feeders and infection of mosquitoes using these membrane feeders (Figure 2), dissection of midgut (Figure 3) and salivary gland of mosquitoes (Figure 4), and quantification of infection in mosquito after midgut and salivary gland dissection.
Figure 1: Workflow of P. falciparum gametocyte culture and membrane feeding protocol.
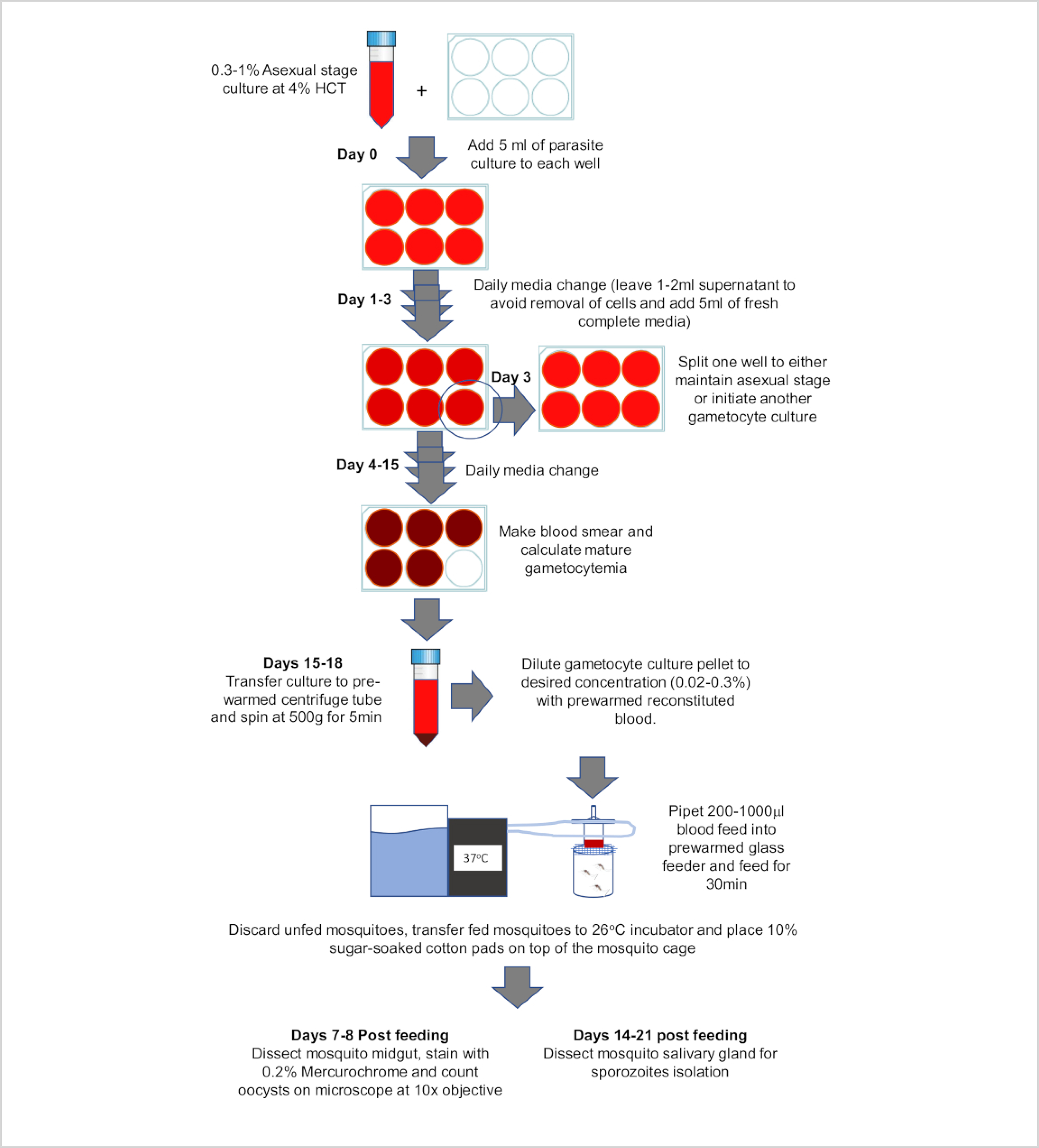
Figure 2: Mosquito blood feeding set-up.

(A) Glass feeder and rectangular piece of paraffin film (B) Two glass feeders displayed before and after parafilm membrane attachment (C) Glass feeder on top of mosquito cup and connected with circulation water bath. (D,E) Pipetting of blood feed into the glass feeder (F) Bottom view of glass feeder showing homogenous distribution of blood feed (G) Several mosquitoes feeding through parafilm membrane. (H) Top view of mosquito cups after feeding, showing drops of blood excreted by the feeding mosquitoes at the bottom of cup. Scale bar = 10 mm.
Figure 3: Graphical representation showing steps of mosquito midgut dissection.
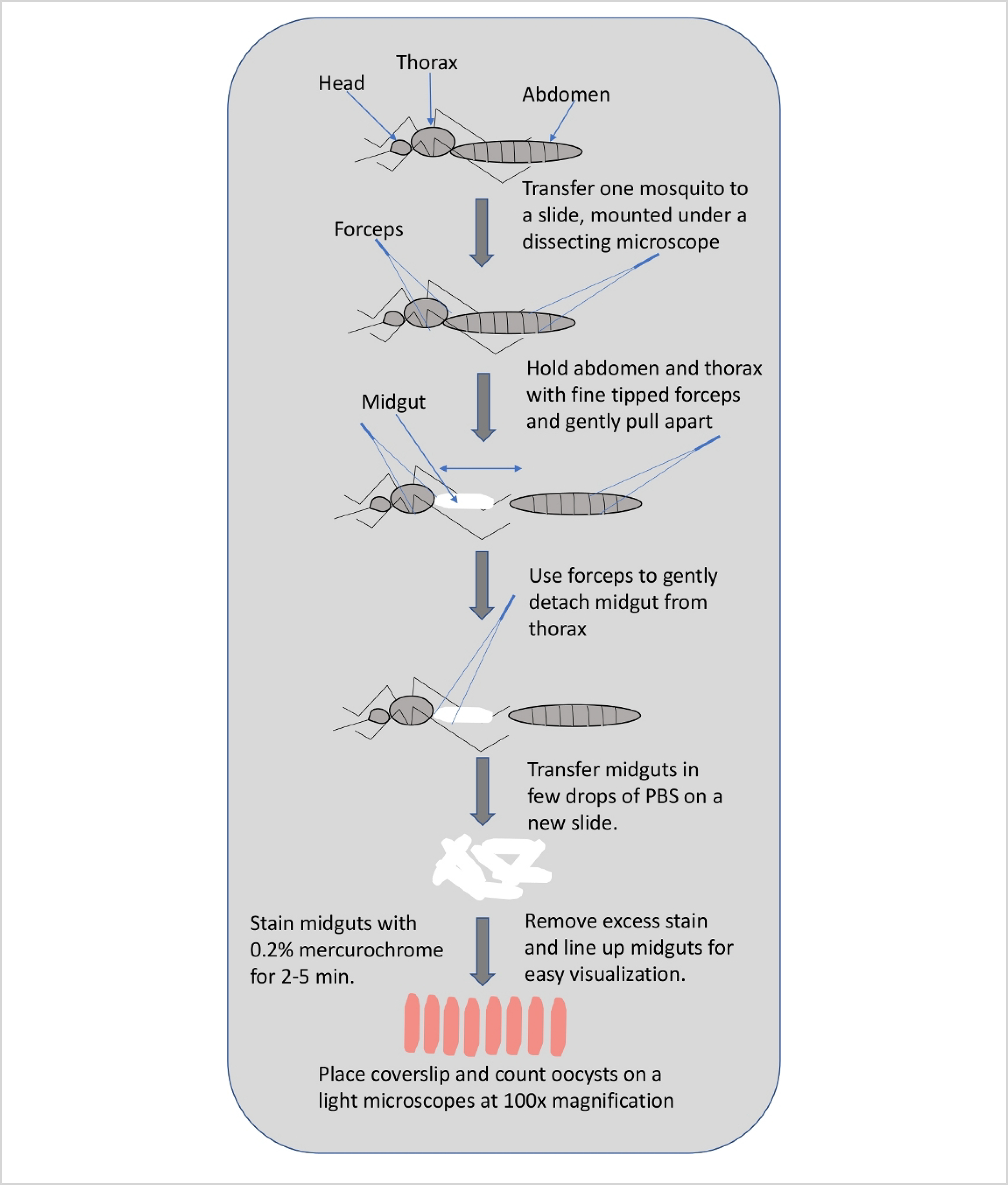
Figure 4: Graphical representation showing steps of mosquito salivary gland dissection.

Protocol
Blood collections described below have been approved by the Institutional Review Board of Johns Hopkins University. P. falciparum is cultured in fresh RBCs under sterile conditions in a biosafety level 2 (BSL2) facility and caution is used to handle biological materials. After each step involving blood or blood products, every plasticware or glassware is rinsed with 10% bleach within the hood prior to proper disposal.
1. Reagents and preparation
-
Parasite isolate P. falciparum NF54 (see Table of Materials) was used, which can produce infectious gametocytes for up to two months while in continuous culture.
NOTE: Not all culture adapted parasite lines produce gametocytes, and low passage NF54 isolates are most consistent.
-
O+ Erythrocytes: Dilute whole blood by adding equal volume of RPMI 1640 media and centrifuge at 500 x g for 5 min at room temperature in a swing out rotor with deacceleration set at 0. Carefully remove the supernatant and buffy coat (a white band between plasma and packed erythrocytes, which contains most of white blood cells and platelets) and add equal volume of RPMI. Repeat the washing step twice and after the final wash add one pellet volume of RPMI to get 50% hematocrit for storage at 4 °C.
NOTE: Gametocytes mature over a period of two weeks which makes it critical to begin cultures with fresh erythrocytes, typically erythrocytes drawn within a week works well.
-
O+ Human Serum: Pool at-least 6 units of serum together to minimize the effect of normal variation in serum from different individuals. Sterilize pooled human serum using 0.2 μm filtration flasks. Aliquot in portions of 50 mL or less based on the need and store at −20 °C.
NOTE: Erythrocytes and serum needs to be from a compatible blood type for P. falciparum cultures.
500x Hypoxanthine (100 mL) solution: Dissolve 0.5 g of hypoxanthine in 100 mL of 1 M NaOH. Filter sterilize and make 5–10 mL aliquots for storage. Stocks can be stored up to a year at −20 °C and up to 2 weeks at 4 °C.
-
Sodium bicarbonate (Optional): Dissolve 7.5 g sodium bicarbonate in 100 mL of deionized, tissue culture grade water and filter sterilize with 0.2 μm filter.
NOTE: This protocol uses the candle jar method to provide microaerophilic conditions for P. falciparum in vitro culture and does not require sodium bicarbonate. However, if parasites are cultured using a malaria gas mix (5% O2, 5% CO2 and 90% N2), it is important to supplement culture media with 0.2% sodium bicarbonate.
Complete media: To prepare 500 mL of complete media, add 1 mL of hypoxanthine solution and 50 mL of pooled human serum to 500 mL of RPMI 1640. Add 15 mL of 7.5% sodium bicarbonate if using a malaria gas mixture. Store complete media at 4 °C and discard if the color of complete media changes from orange to pink. To avoid waste, make enough complete media to be used within three days.
-
Exflagellation media (Optional): Make exflagellation or ookinete media by dissolving 200 mg NaHCO3, 5 mg Hypoxanthine and 100 μL of xanthurenic acid (from 100 mM stock in water) to 100 mL of incomplete media (RPMI 1640 with glutamine and HEPES).
NOTE: Exflagellation is the process of male gamete formation inside the midgut of female Anopheles mosquito few minutes after it takes blood meal infected with gametocytes. Male gametocytes of Plasmodium give rise to 8 male gametes after an exflagellation event. In vitro, this process occurs spontaneously when culture temperature is lowered to room temperature (RT) and can be observed in cultured mature gametocytes.
N-Acetylglucosamine (Optional): Make 500 mM stock solution of N-acetylglucosamine in water or incomplete media, aliquot and store at −20 °C.
NaCl solutions: Dissolve 0.9 g, 1.6 g and 12 g NaCl in 100 mL of tissue culture grade deionized water to make 0.9%, 1.6% and 12% NaCl solutions. Filter sterilize with 0.22 μm filter and store at 4 °C.
Materials.
| Name | Company | Catalog Number | Comments |
|---|---|---|---|
| 10% Sugar solution | |||
| 10ml serological pipet | Falcon | 357551 | |
| 15 ml conical tube | Falcon | 352096 | |
| 1ml serological pipet | Falcon | 357521 | |
| 25 ml serological pipet | Falcon | 357535 | |
| 37°C Incubator | |||
| 50 ml conical tube | Falcon | 352070 | |
| 5ml serological pipet | Falcon | 357543 | |
| 6 well tissue culture plates | Falcon | 353046 | |
| 70% Ethanol | |||
| 9″ glass pipet | Fisherbrand | 13-678-6B | |
| Anopheles Mosquitoes | JHMRI, Insectary core | We use A. stephensi or A. gambiae (keele) | |
| cell counter | |||
| Circulating water bath | |||
| fine tip forceps | Fisherbrand | 12-000-122 | |
| Geimsa stain | Sigma | GS1L | |
| Glass desiccator | |||
| Glass membrane feeder | Chemglass Life Sciences | CG183570 | |
| Glass slides | Fisherbrand | 12-552-3 | |
| HBSS | Sigma | H6648 | |
| Human Blood O+ | JHU | Wash RBCs three times with RPMI and refrigerate at 50% heamatocrit | |
| Human Serum O+ | Interstate blood bank | Pool at-least 6 units of serum from different donors and freeze down aliquots at −20°C. | |
| Hypoxanthine | Sigma | H9337 | Make 500x stock in 1M NaOH |
| Mercurochrome | Sigma | M7011 | Prepare 1% stock solution in PBS that can be diluted to 0.1% when needed |
| Micro Pipette | |||
| Microscope | Olympus | Any microscope with 10x, 40x and 100x objective will work. | |
| Mosquito cups | Neptune cups | ||
| N-acetylglucosamine | Sigma | A3286 | Optional and needed only when pure gametocytes are required. |
| Netting | Make sure it can contain mosquitoes and allow blood feeding | ||
| Parafilm | |||
| PBS | |||
| Petri dish | Thermo Scientific | 249964 | |
| Pipet tips | |||
| Pipetman | |||
| Plasmodium falciparum NF54 | BEI Resources | MRA-1000 | Freeze down large numbers of early passage culture to make sure you have a constant supply |
| RPMI 1640 | Corning | CV-041-CV | Media contains glutamine and HEPES |
| Slide warmer | |||
| Sodium bicarbonate | Sigma | S6297 | Optional for media, add only when using malaria gas mix during culture incubation |
| water bath | |||
| Xanthurenic Acid | Sigma | D120804 | For flagellation media |
2. P. falciparum asexual stage culture
To begin parasite culture, remove a low passage frozen vial of P. falciparum NF54, from the liquid nitrogen tank and quickly thaw in water bath set at 37 °C.
Transfer contents (~1 mL) to a 50 mL sterile centrifuge tube and dropwise add 0.2 volume of prewarmed 12% NaCl, while gently shaking the tube to ensure even mixing. Incubate for 5 min at RT with intermittent gentle shaking.
Add 9 volumes of 1.6% NaCl dropwise, while continuing gentle mixing. Centrifuge the contents at 500 x g for 5 min at RT, carefully discard the supernatant. Add 9 volumes of 0.9% NaCl dropwise, while making sure to consistently mix the parasite pellet. Centrifuge again at 500 x g for 5 min at RT. Remove the supernatant and resuspend parasites into 5 mL of complete media.
Transfer contents to one well of a 6 well tissue culture plate and add 100–200 μL of packed RBCs.
Incubate parasite culture at 37 °C in a candle jar or in a modular incubator chamber purged with special gas mix of 5% O2, 5% CO2 and 90% N2.
Replace media every day and monitor the growth by making a thin blood smear by drawing a few microliters from settled RBC layer after the culture supernatant has been aspirated during regular media change.
Use sterile glass Pasteur pipette to draw and then tap in the center of the glass slide to transfer a drop of culture to the slide. Place another glass slide in front of the of the drop and draw it back to contact the RBCs, quickly pushing it forward in one motion to make thin smear of RBC monolayer.
Place the slide horizontally on a drying rack and let it air dry. Fix the blood smear by dropping absolute methanol onto the smear. Allow the fixed smear to dry completely and then carefully pour 10% Geimsa-stain freshly diluted in water, until blood smear is completely covered. Allow cells to stain for approximately 15 min.
Wash off the excess stain by rinsing the slide under clean tap water and allow the slides to dry in vertical position.
Determine parasitemia by viewing thin blood smear on a microscope using oil immersion 100x objective. Count the number of infected erythrocytes among a total of at-least 500 RBCs to determine percent parasitemia.
3. P. falciparum gametocyte culture
NOTE: Gametocyte cultures take two weeks to produce mature gametocytes infectious to mosquitoes. The steps of the gametocyte culture are outlined in Figure 1. P. falciparum isolates usually lose the ability to produce gametocyte after long term in vitro culture13. To ensure quality of gametocytes, culture should be initiated from low passage feeder culture, not more than 2 months old since thawing. Pre-warm media to 37 °C and perform all the procedures on a slide warmer set at 38 °C. Make sure gametocyte cultures are not out of incubator for extended period to minimize temperature fluctuations.
Seed the gametocyte culture (day 0) using mixed asexual stage feeder culture at 0.3–1% parasitemia at 4% hematocrit.
To set up a six well gametocyte culture, centrifuge 5 mL of feeder culture at 500 x g for 5 min at RT. Discard the supernatant and resuspend the pellet in 30 mL of complete media.
-
Add 1.2 mL of packed RBCs, mix, and dispense 5 mL to each well of the 6 well plate and incubate in a candle jar at 37 °C.
NOTE: Gametocyte culture set up described above is based on a mixed asexual stage feeder culture at 5% parasitemia.
Change media daily for 15–18 days, without the addition of fresh blood, by carefully aspirating about 70–80% culture supernatant to avoid removing blood cells.
-
Add 5 mL of fresh complete media to each well. While changing media, slowly add media using a serological pipette against the wall of the well to avoid disturbing the settled RBC layer.
NOTE: Because 1–2 mL of the culture medium will be left during media change, total volume of culture will be between 6–7 mL, after day 1 of gametocyte culture. While adapting this protocol, it is advisable to make blood smears every alternate day to make sure parasites are healthy. Making a blood smear can often deplete the number of cells in culture. To avoid this, draw a very small volume.
-
To quantify mature gametocytemia, make blood smear between days 15–18 and count the number of mature gametocytes among total number of cells.
NOTE: Mature gametocytes can easily be identified with their classic crescent shape with smooth rounded ends (Figure 5B).
Count a minimum of 1,000 RBCs and calculate the percentage of RBCs infected with mature gametocytes.
To quantify exflagellation events, take 200 μL of gametocyte culture and centrifuge at 500 x g for 5 min at RT in a pre-warmed tube. Resuspend the pellet in 20 μL of exflagellation media and transfer to a glass slide with cover slip.
-
After 15 min incubation at room temperature, begin counting exflagellation centers in phase contrast mode using 10x objective. Count exflagellation centers in at-least four fields to calculate exflagellation events.
NOTE: A detailed methodology of exflagellation assay has been described earlier by Delves et al10. More than 20 exflagellation events per field is considered suitable for membrane feeding assay.
Gametocyte cultures typically have low levels of residual asexual stages. If the experiment requires pure gametocytes, treat the culture with 50 mM N-acetylglucosamine.
-
Add 0.5 mL of N-acetylglucosamine from 10x stock solution per well (5 mL) for a minimum of 3 days while changing media to clear residual asexual stages.
NOTE: N-acetylglucosamine can block invasion of sexually committed merozoites too, treatment should be initiated only after day 7 of gametocyte culture.
Figure 5: P. falciparum gametocyte culture and oocyst visualization on infected mosquito midgut.

(A) Time course of 15 day gametocyte culture, showing steep multiplication of asexual stages to peak parasitemia within first 4 days, followed by gametocytogenesis and maturation over time. (B) Geimsa-stained thin blood smear showing various stages of gametocyte culture, day 1 early asexual stage, day 4 peak asexual parasitemia, day 6 stressed culture due to high parasite load, days 9 & 12 early gametocytes and day 15 showing mature male and female gametocytes. Morphologically early stage gametocytes were indistinguishable from asexual stages, but late stage II showed the crescent shape with pointed ends and elongated as parasite developed into stage III and IV. Mature stage V gametocytes, however, was characterized by classic crescent shape with rounded ends and minimal host cell visibility14. Scale bar = 10 mm
4. Mosquito infection using standard membrane feeding assay (SMFA)
NOTE: Gametocytes grown in vitro can be fed to mosquitos using glass membrane feeders. Setting up of the blood feeding apparatus is shown in Figure 2. As described above, always maintain gametocytes at 37 °C to avoid activation before they are ingested by mosquitoes. Prewarm plasticware, reagents and equipment’s used with gametocyte culture to 37 °C.
Starve 3–7-day old female Anopheles mosquitoes by removing their sugar water for 6.5 h or overnight before membrane feeding.
-
Transfer them using motorized or mouth operated aspirators to paper cups covered with double layer of fine mesh fabric. Alternatively, knock down mosquitoes in a cold room or refrigerator to transfer them to the paper cups.
NOTE: One-pint size cup can be used to feed up to 100 mosquitoes.
Centrifuge washed blood at 500 x g for 5 min at RT and discard supernatant. Add an equal volume of freshly thawed human serum to reconstitute the whole blood. Transfer the blood and serum mix to the water bath set at 37 °C for 30 min.
Transfer gametocyte culture to pre-warmed 15 mL plastic tube and centrifuge at 600 x g for 5 min at 37 °C. Carefully aspirate the culture supernatant and make thin blood smear to determine mature gametocytemia as described above.
-
To make the final blood feed, dilute gametocyte pellet to a desired concentration using reconstituted whole blood. Keep the blood feed at 37 °C until mosquitoes and glass feeders are ready.
NOTE: Mature gametocyte concentrations between 0.02 to 0.3% will provide consistent mosquito infectivity. However, it is advisable to optimize gametocytemia by feeding various concentrations to different cups of mosquitoes.
Ensure that the glass membrane feeders have two openings, a narrow top opening to pipette the infected blood and a bottom opening for membrane attachment. Cut a paraffin film into squares, stretch to an even thickness and stick on to the opening of feeder to create sealed compartment for the blood feed.
-
Attach membrane feeders to a circulatory water-bath by connecting tubing on each side to allow for the passage of warm water through the jacket around membrane feeders.
NOTE: Several glass feeders can be connected in series to accommodate for multiple feeding conditions.
After all the feeders are connected to the water bath, turn it on and inspect for any leaks.
Place feeders in the center of netting on mosquito cups with membrane-side down and secure feeders using clamps or tape. Make sure membrane feeders are not tilted to any side to allow even distribution of the blood feed and close contact of the entire feeder with the netting on cups, to allow mosquitoes access from inside the cup.
-
Pipette about 200–1000 μL of blood feed into membrane feeders.
NOTE: Volume of the blood feed will vary based on the number of mosquitoes and size of the membrane feeder. For 14 mm glass feeder and 50 mosquitoes use 200 μL of blood feed.
Make sure loading was done properly and blood feed was overlaid on the paraffin film.
-
Allow mosquitoes to feed for about 30 min with intermittent monitoring.
NOTE: Gently blow mosquitoes with mouth to provide CO2 which will induce improved blood feeding.
Remove unfed mosquitoes by knocking them down in a cold room. Visibly inspect mosquitoes for bulge and redness in the abdomen as a sign of fresh blood meal. Alternatively, use a mouth operated aspirator to selectively remove unfed mosquitoes.
Discard unfed mosquitoes, after soaking them in 70% ethanol and put fed mosquitoes back into the mosquito cup.
Double cage mosquito cups and transfer them to high containment incubator specified for mosquitoes infected with human malaria parasite P. falciparum.
Place cotton pads soaked in 10% sucrose on the mosquito cups to provide them with sugar meal. Replace cotton pads every other day, until mosquitoes are dissected.
5. Mosquito mid-gut dissection and oocyst load quantification
NOTE: A schematic of midgut dissection is shown in Figure 3.
7–8 days post blood feeding, transfer mosquito cups to 4 °C for 10 min to knock down mosquitoes.
Transfer mosquitoes to be dissected to a Petri dish on ice using fine-tipped forceps.
-
Soak mosquitoes in 70% ethanol for 1–2 min to euthanize them. Add PBS to the Petri dish to wash off the ethanol after all the mosquitoes are euthanized.
NOTE: Before adding PBS make sure all the mosquitoes are dead.
Mount a glass slide on dissecting microscope and pipette 100 μL of PBS in the center. Carefully transfer one mosquito using forceps into the PBS and leave the rest of the mosquitoes in the Petri dish on ice.
Using fine-tipped forceps, hold the third segment of the abdomen from the posterior end of the mosquito and with second forceps hold the junction between the thorax and abdomen. Gently pull the abdomen until the midgut is fully exposed.
Discard the rest of the mosquito tissue and transfer the midgut to a clean slide containing a few drops of PBS.
When the desired number of mosquitoes have been dissected, carefully remove the PBS using a pipette and stain the midgut with 0.2% mercurochrome for 2–5 min.
Remove excess mercurochrome and line-up midguts on the slide so that they can be easily visualized under the light microscope.
Place a cover slip and count oocysts on each midgut under 10x objective. Oocysts stain pink and are circular in shape (Figure 6C).
Figure 6: P. falciparum oocysts counts per mosquito midgut.
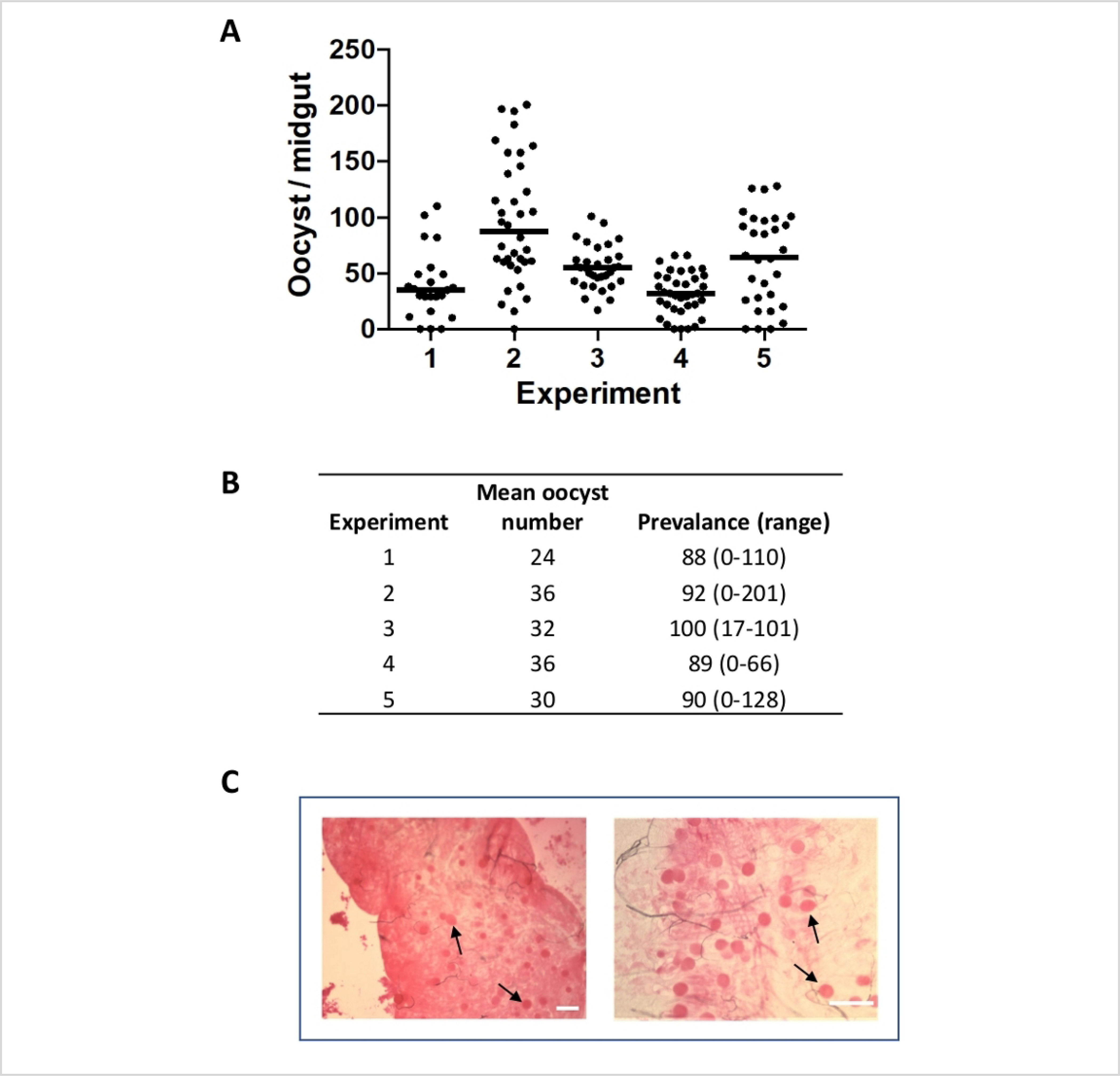
(A) Graph shows oocyst counts over the years, for each experiment, mosquitoes were fed with 0.3% gametocytes and midguts were dissected on day 8 post blood feed. Each dot represents the number of oocysts from individual midguts and the horizontal line represent median value. (B) Table shows mean oocyst count, prevalence of infected mosquitoes and range of infection. (C) Pictures showing oocysts on mosquito midgut from two separate blood feeding experiments at different magnifications. Scale bar = 150 μm.
6. Mosquito salivary gland dissection and sporozoite load quantification
NOTE: A schematic of salivary gland dissection is shown in Figure 4.
-
14–18 days post blood feeding, place mosquito cups at 4 °C for 10 min.
NOTE: It is important to wait till mosquitoes stop moving.
While waiting for mosquitoes to stop moving, prepare tools for dissection.
Place a glass plate on a dissection microscope stage. Prepare 2 sets of 25 G needles on 1 mL syringes and a 9” Pasteur pipette and rubber bulb. Place 70% EtOH and dissection medium (HBSS, L-15 or PBS) in a 6 well plate.
In the cold room, transfer anesthetized mosquitoes in a 6 well plate containing 70% EtOH. Gently move mosquitoes with forceps to make sure all mosquitoes are soaked, then transfer mosquitoes to the dissection medium by using forceps to wash off 70% EtOH.
Move to the dissection room and place mosquitoes on a dissection microscope stage.
Remove excess medium from mosquitoes, add new medium as necessary during dissection. Avoid mosquitoes from drying out yet too much liquid makes dissection more difficult.
Using 2 syringes with 25 G needles, hold mosquito thorax and head, and gently pull the head upward to pull salivary glands from thorax.
Disconnect salivary glands from head and thorax with a needle.
Temporary place salivary glands in a separate droplet of medium on the glass plate.
-
After dissecting 15–20 salivary glands, collect them in a low retention tube using the Pasteur pipette.
NOTE: Once salivary glands enter the wide part of the Pasteur pipette, they will stick to the glass and it is difficult to get them out.
Pellet salivary glands by short pulse spin in a table-top centrifuge. Remove the dissection medium without disrupting the pellet and Add 100 μL of new dissection medium.
Grind salivary glands with a small homogenizer for 1 min to obtain sporozoites.
-
Place 10 μL of dissection medium containing sporozoites on a hemocytometer.
NOTE: Depending on the number of salivary glands, one may need to dilute the sporozoite solution to 1:10 to 1:50 with medium before counting.
- Count the number of sporozoites in two of the four quadrants and calculate number of sporozoites/mosquito:
Representative Results
Here we present results from a series of membrane feeds using P. falciparum NF54 gametocyte cultures generated using the protocol above (see (Figure 5). Gametocyte culture was initiated with approximately 0.5% mixed stage asexual culture on Day 0, which grew to a peak parasitemia of approximately 15% by Day 4 and Day 5. As shown in Figure 5A at this high parasitemia, the parasites are stressed and the asexual stage culture crashes. However, this stress concomitantly results in the induction of gametocytogenesis. Early gametocytes appeared after Day 6 and Day 7 and asexual parasitemia slowly declined but remained at a low level. Presence of asexual stage parasites did not affect mosquito feeding experiments. However, if gametocytes are to be used in experiments which require pure cultures, such as drug sensitivity assays and proteomic or transcriptomic studies, residual asexual stages can be removed by treatment with 50 mM N-acetylglucosamine. The majority of gametocytes mature to stage V by Day 15 at which time they become infectious to mosquitoes and were ready to be fed. Representative images of Giemsa-stained blood smears at different time points after initiation of gametocyte culture are shown in Figure 5B.
Between 8 to 10 days after blood feeding, mosquitoes were dissected to determine the prevalence and intensity of infection. Prevalence is the percentage of fed mosquitoes that have oocysts while intensity is the number of oocysts found in each mosquito. Both are important indicators of the success of the feed. Data from 5 independent mosquito feeds are shown in Figure 6. Feeds were chosen to show the normal variation in the levels of infectivity after feeding with 0.3% mature gametocytes from day 16 culture of P. falciparum NF54. Oocyst intensities provides quantitative data to determine mosquito infectivity of gametocytes and prevalence shows the percentage of fed mosquitoes that became infected. These data can be used to evaluate transmission blocking agents and to identify and characterize targets for transmission blocking vaccines and drugs. Number of oocysts varied both within and between experiments and required 25–50 mosquitoes per cohort to determine the effect of various experimental conditions.
Oocyst intensities are considered as end point of most transmission blocking assays and strategies, however number of sporozoites are usually important for sporozoite biology and for liver stage studies. Table 1 shows the average number sporozoites obtained per mosquito from 12 independent blood feeds. As shown in the table, average number of sporozoites were consistent, however, there was one experiment where we obtained zero sporozoites, representing occasional failure of the assay.
Table 1: Average number of salivary glands sporozoites per mosquito for 12 independent cycles over a 2-year period.
A. stephensi mosquitoes were fed with 0.3% P. falciparum gametocytes and 15–20 mosquitoes were dissected between days 14–17 post blood feeding. Average sporozoite counts per mosquito are shown.
| Experiment | Day of dissection (post blood feed) | Mean sporozoite/mosquito |
|---|---|---|
| 1 | 14 | 42, 000 |
| 2 | 14 | 40, 000 |
| 3 | 14 | 42, 750 |
| 4 | 15 | 25, 411 |
| 5 | 15 | 33, 750 |
| 6 | 14 | 15, 750 |
| 7 | 16 | 0 |
| 8 | 15 | 33, 200 |
| 9 | 14 | 56, 333 |
| 10 | 14 | 45, 333 |
| 11 | 17 | 43, 750 |
| 12 | 16 | 56, 000 |
Discussion
Methods described here have been successfully used at the Johns Hopkins Malaria Research Institute for more than 10 years15, 16, 17, 18, 19, 20, 21, 22. Gametocytes produced using this protocol have been used for high throughput gametocytocidal assays22, for proteomic15, as well as for transcriptomic23 studies. However, a major reason to develop these methods is to use the mature gametocytes to infect mosquitoes for studies on mosquito stages and sporozoites23, 24, 25, 26. This manuscript describes a detailed protocol of generating mature P. falciparum gametocytes and infection of mosquitoes using glass membrane feeders. These methods are critical for any laboratory working on transmission blocking strategies, sporozoite stages and pre-erythrocytic liver stages of P. falciparum.
Ifediba and Vanderberg in 1981 described long term culture of P. falciparum, in the presence of 50 μg/mL hypoxanthine which produced highly infectious mature gametocytes27. Since then there have been numerous publications describing methods to produce gametocytes for different applications9, 10, 11, 12. Most of these publications utilize previously described gametocytogenesis-inducing conditions to increase yields. Using conditioned media, stressing the culture by sudden parasitemia increase, drop in hematocrit to mimic anemia, red blood cell lysis and log phase repression by bulk up, can be used to induce gametocytogenesis. The method described here is simple and time tested. Initiate gametocyte culture with 0.3–1% asexual stage parasitemia at 4% hematocrit and change media every day until day 15–18. To achieve consistent results, it is critical to begin gametocyte culture with low passage asexual stage parasites, use fresh RBCs (<1 week) and making sure that the culture temperature does not fluctuate during media change. Since falciparum gametocyte development process occurs sequestered in static conditions of extravascular spaces in bone marrow4, 5, it’s important to not disturb settled RBC layers throughout the culture period.
Culturing P. falciparum gametocytes with consistency is demanding but getting them to infect mosquitoes presents another level of complexity. Membrane feeding is subject to several variables other than gametocytes, such as age and fitness of mosquito, midgut microbiota, and feeding behavior28, 29. Usually SMFA data show a high degree of variability and require large number of mosquitoes to identify effects of different experimental conditions11, 28. Using low passage culture for gametocytes, healthy 3 – 6-day old mosquitoes and optimized membrane feeding protocol can help with variations in oocyst counts.
The protocol described here for both gametocyte cultivation and membrane feeding, has been optimized over many years. These methods provide a detailed description for obtaining mature transmission competent gametocytes, standard membrane feeding assay, mosquito midgut dissection and oocyst quantification as well as salivary gland dissection and sporozoite quantification. These protocols are consistent in terms of the number of gametocytes needed to provide reliable mosquito infectivity and robust oocyst counts and sporozoite yields.
Acknowledgments
Authors thank Bloomberg Philanthropies for financial support to Johns Hopkins Malaria Research Institute (JHMRI). This work would not have been possible without the expertise provided by JHMRI insect and parasitology core facilities.
Footnotes
Disclosures
The Authors have nothing to disclose.
References
- 1.World Health Organization, World Malaria Report. WHO. (2018). [Google Scholar]
- 2.Sinden RE, Smalley ME Gametocytogenesis of Plasmodium falciparum in vitro: The cell-cycle. Parasitology. 79, (2), 277–296 (1979). [DOI] [PubMed] [Google Scholar]
- 3.Sinden RE Sexual Development of Malarial Parasites. Advances in Parasitology. 22, 153–216 (1983). [DOI] [PubMed] [Google Scholar]
- 4.Joice R et al. Plasmodium falciparum transmission stages accumulate in the human bone marrow. Science Translational Medicine. 6, (244), 244re5 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Abdulsalam AH, Sabeeh N, Bain BJ Immature Plasmodium falciparum gametocytes in bone marrow. American Journal of Hematology. 85, (12), 943 (2010). [DOI] [PubMed] [Google Scholar]
- 6.Ghosh AK, Jacobs-Lorena M Plasmodium sporozoite invasion of the mosquito salivary gland. Current Opinion in Microbiology. 12 (4), 394–400 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bennink S, Kiesow MJ, Pradel G The development of malaria parasites in the mosquito midgut. Cellular Microbiology. 18, (7), 905–918 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Trager W, Jenson JB Cultivation of malarial parasites. Nature. 273, (5664), 621–622 (1978). [DOI] [PubMed] [Google Scholar]
- 9.Duffy S, Loganathan S, Holleran JP, Avery VM Large-scale production of Plasmodium falciparum gametocytes for malaria drug discovery. Nature Protocols. 11, (5), 976–992 (2016). [DOI] [PubMed] [Google Scholar]
- 10.Delves MJ et al. Routine in vitro culture of P. Falciparum gametocytes to evaluate novel transmission-blocking interventions. Nature Protocols. 11, (9), 1668–1680 (2016). [DOI] [PubMed] [Google Scholar]
- 11.Habtewold T et al. Streamlined SMFA and mosquito dark-feeding regime significantly improve malaria transmission-blocking assay robustness and sensitivity. Malaria Journal. 18, (1), 24 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Demanga CG et al. The development of sexual stage malaria gametocytes in a Wave Bioreactor. Parasites and Vectors. 10, (1), 216 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Brockelman CR Conditions favoring gametocytogenesis in the continuous culture of Plasmodium falciparum. Journal of Eukaryotic Microbiology. 29, 454–458 (1982). [DOI] [PubMed] [Google Scholar]
- 14.Meibalan E, Marti M Biology of malaria transmission. Cold Spring Harbor Perspectives in Medicine. 7, (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Essuman E et al. A novel gametocyte biomarker for superior molecular detection of the plasmodium falciparum infectious reservoirs. Journal of Infectious Diseases. 216, (10), 1264–1272 (2017). [DOI] [PubMed] [Google Scholar]
- 16.Simões ML, Mlambo G, Tripathi A, Dong Y, Dimopoulos G Immune regulation of plasmodium is anopheles species specific and infection intensity dependent. mBio. 8, (5), e01631–e01617 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Oakley MS et al. Transcriptome analysis based detection of Plasmodium falciparum development in Anopheles stephensi mosquitoes. Scientific Reports. 8, 11568 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Saraiva RG et al. Chromobacterium spp. mediate their anti-Plasmodium activity through secretion of the histone deacetylase inhibitor romidepsin. Scientific Reports. 8, 6176 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tao D et al. Sex-partitioning of the Plasmodium falciparum stage V gametocyte proteome provides insight into falciparum-specific cell biology. Molecular and Cellular Proteomics. 13, (10), 2705–2724 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Grabias B, Zheng H, Mlambo G, Tripathi AK, Kumar S A sensitive enhanced chemiluminescent-ELISA for the detection of Plasmodium falciparum circumsporozoite antigen in midguts of Anopheles stephensi mosquitoes. Journal of Microbiological Methods. 108, 19–24 (2015). [DOI] [PubMed] [Google Scholar]
- 21.Ferrer P, Vega-Rodriguez J, Tripathi AK, Jacobs-Lorena M, Sullivan DJ Antimalarial iron chelator FBS0701 blocks transmission by Plasmodium falciparum gametocyte activation inhibition. Antimicrobial Agents and Chemotherapy. 59, (3), 1418–1426 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sanders NG, Sullivan DJ, Mlambo G, Dimopoulos G, Tripathi AK Gametocytocidal screen identifies novel chemical classes with Plasmodium falciparum transmission blocking activity. PLoS One. 9, (8), e105817 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lindner SE et al. Transcriptomics and proteomics reveal two waves of translational repression during the maturation of malaria parasite sporozoites. Nature Communications. 10, 4964 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.McLean KJ et al. Generation of Transmission-Competent Human Malaria Parasites with Chromosomally-Integrated Fluorescent Reporters. Scientific Reports. 9, 13131 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Espinosa DA et al. Proteolytic Cleavage of the Plasmodium falciparum Circumsporozoite Protein Is a Target of Protective Antibodies. Journal of Infectious Diseases. 212, (7), 1111–1119 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Swearingen KE et al. Interrogating the Plasmodium Sporozoite Surface: Identification of Surface-Exposed Proteins and Demonstration of Glycosylation on CSP and TRAP by Mass Spectrometry-Based Proteomics. PLoS Pathogens. 12, (4), e1005606 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ifediba T, Vanderberg JP Complete in vitro maturation of Plasmodium falciparum gametocytes. Nature. 294, (5839), 364–366 (1981). [DOI] [PubMed] [Google Scholar]
- 28.Miura K et al. An inter-laboratory comparison of standard membrane-feeding assays for evaluation of malaria transmission-blocking vaccines. Malaria Journal. 15, 463 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Miura K et al. Qualification of Standard Membrane-Feeding Assay with Plasmodium falciparum Malaria and Potential Improvements for Future Assays. PLoS One. 8, (3), e57909 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]